










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.44 n.2 Buenos Aires abr./jun. 2007
The Triassic insect fauna from the Los Rastros Formation (Bermejo Basin), La Rioja Province (Argentina): its context, taphonomy and paleobiology
Adriana C. Mancuso1, Oscar F. Gallego2 and Rafael G. Martins-Neto3
1Museo de Ciencias Naturales y Antropológicas "Juan Cornelio Moyano" e Instituto Argentino de Nivología, Glaciología y Ciencias Ambientales, CRICYT (Centro Regional de Investigaciones Cientificas y Tecnológicas, Consejo Nacional de Investigaciones Científicas y Técnicas), Adrián Ruiz Leal s/n -Parque Gral. San Martín, 5500 Mendoza, Argentina, C.C. 330. amancu@lab.cricyt.edu.ar
2Paleontología (Facultad de Ciencias Exactas y Naturales y Agrimensura, Universidad Nacional del Nordeste) y Área Paleontología (Centro de Ecología Aplicada del Litoral, CECOAL (Consejo Nacional de Investigaciones Científicas y Técnicas), 3400 Corrientes, Argentina, C.C. 128. ofgallego@hotmail.com
3Programa de Pós-graduação em Ciências Biológicas, Comportamento e Biologia Animal, Universidade Federal de Juiz de Fora - UFJF. Campus Universitário, Martelos. 36036-900 Juiz de Fora, MG, Brasil. martinsneto@terra.com.br
Abstract. In the Bermejo Basin, the Los Rastros Formation bears an abundant insect fauna, mainly of terrestrial winged adult organisms related to the Blattoptera and the Coleoptera orders. The insect remains are found in the black shales of the offshore lacustrine facies and their taphonomic features suggest that the specimens were allochthonous to the lake. The individuals appear to arrived alive to the lake and had a rapid fall trough the water column, thus, they are preserved intact. In contrast, some of them may have suffered fragmentation in air traspontation and/or biological attack during long periods of flotation.
Resumen. La fauna de insectos triásicos de la Formación Los Rastros (cuenca Bermejo), provincia de La Rioja (Argentina): su contexto, tafonomía y paleobiología. En la cuenca Bermejo, la Formación Los Rastros es portadora de una abundante fauna de insectos, principalmente organismos adultos terrestres y alados pertenecientes a los órdenes Blattoptera y Coleoptera. Los restos de insectos son encontrados en las pelitas negras de la facies de lago abierto. Las características tafonómicas de los insectos sugieren que los especímenes son alóctonos al lago. Los individuos pudieron llegar vivos al lago y sufrir una rápida caída a través de la columna de agua, preservándose intactos, o sufrir fragmentación en el transporte aéreo o por ataques biológicos durante largos períodos de flotación.
Key words. Triassic; Insecta; Taphonomy; Los Rastros Formation; Argentina.
Palabras clave. Triásico; Insecta; Tafonomía; Formación Los Rastros; Argentina.
Introduction
Generally, the fossil insect bearing deposits are found in very quiet depositional environments, such as lacustrine systems and transitional environments (Martinell and Martinez-Delclós, 1990). During the biostratinomic stage, there are several processes which can affect the insect remains. In some cases, insects are attracted by physical factors, such as temperature and moisture, and these factors can promote the presence of these organisms in a specific environment. The wind factor is considered completely external to the organism and is one of the most important factors in the generation of insect deposits (Martinell and Martinez-Delclós, 1990).
The Los Rastros succession has an abundant insect fauna. Most of the individuals are adult winged organisms and terrestrial representatives belonging to the Blattoptera and the Coleoptera orders. The remains are only associated with the black shales of the offshore lacustrine facies.
Geological setting
The Bermejo Basin is an extensional basin formed along the western margin of Gondwana during the Late Permian and Early Triassic in the pre-breakup stage of Pangea (Uliana and Biddle, 1988). Basin deposits crop out in the northwest of Argentina along the border between San Juan and La Rioja provinces, and shows a NW-SE orientation. The sedimentary infilling of the Bermejo Basin is dominated by 2.000 to 6.000 m of Triassic alluvial, fluvial and lacustrine deposits (Stipanicic, 2002).
From a lithostratigraphic point of view, the sedimentary succession has been subdivided, in ascending order, into the Talampaya and Tarjados formations (Romer and Jensen, 1966), and the Agua de la Peña Group, which was divided into four units (Mancuso, 2005a, 2005b) which are from the base to the top: the Chañares Formation (Romer and Jensen, 1966), the Los Rastros Formation (Frenguelli, 1944; Ortiz, 1968), the Ischigualasto Formation (Frenguelli, 1944; Stipanicic, 1969) and the Los Colorados Formation (Groeber and Stipanicic, 1953; Stipanicic, 1969) (figures 1, 2).
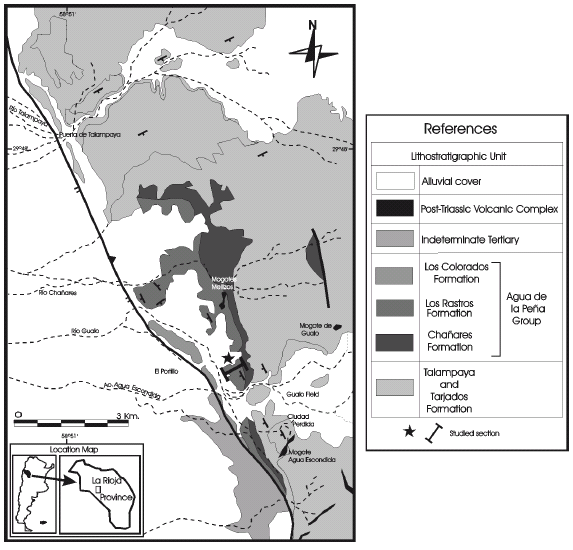
Figure 1. Geologic map of the Río Gualo section in the Bermejo Basin (modified from Mancuso, 1999) / mapa geológico de la sección de Río Gualo en la cuenca del Bermejo (modificado de Mancuso, 1999).
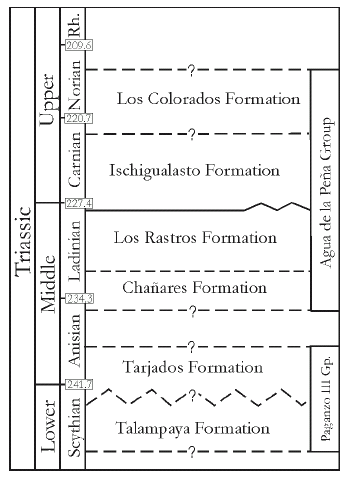
Figure 2. Triassic lithostratigraphy and chronostratigraphy in the Bermejo Basin (modified from Rogers et al., 2001). Limits in my. / litoestratigrafía y cronoestratigrafía triásica en la cuenca Bermejo (modificado de Rogers et al., 2001). Límites en ma.
The Chañares Formation is dominated by tuffaceous sandstones and siltstones of fluvial-lacustrine deposits, and by alternation of polymictic clast- and matrix-supported conglomerates belonging to alluvial fan deposits (Rogers et al., 2001; Mancuso, 2005a, 2005b). This succession passes upward into the lacustrine black shales and deltaic sandstones of the Los Rastros Formation (Rogers et al., 2001; Mancuso, 2005a, 2005b). The sandstones, mudstones and tuffs of the Ischigualasto Formation deposited by a moderate to high-sinuosity river system overlay the Los Rastros succession (Milana and Alcober, 1994). The succession culminates with the red beds of the Los Colorados Formation. Following these, a regional unconformity separates the Triassic succession from the overlaying ?Cretaceous Cerro Rajado Formation (Bossi, 1977; Stipanicic and Bonaparte, 1979; López-Gamundí et al., 1989; Caselli et al., 2001).
Facies analysis of the Los Rastros Formation
Based on the field data recovered at the Río Gualo locality (figure 1), several lacustrine-deltaic cycles were defined, which correspond to a shallowing-upward sequence (Vail et al., 1991). Each cycle is characterized by a coarsening-upward succession, and consists of Facies association A (offshore lacustrine deposits) dominated by claystones with iron mudstones interbedded, and Facies association B (deltaic deposits) represented by coarsening-upward siltstones and sandstones with rare organicrich shales interbedded (figure 3). The individual coarsening-upward cycle ranges, approximately, from 20 to 40 m.
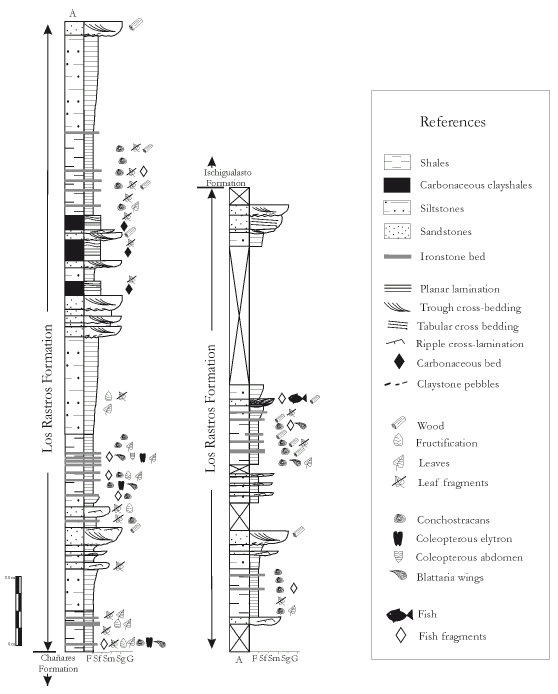
Figure 3. Detailed stratigraphic section of Los Rastros Formation at Río Gualo locality indicating fossiliferous horizons / sección estratigráfica detallada de la Formación Los Rastros en Río Gualo indicando los horizontes fosilíferos.
Facies assemblage A (offshore lacustrine deposits). This facies assemblage is dominated by dark gray to black carbonaceous claystones (Fl) interbedded with iron mudstones and very fine-grained sandstones (SFm) (figures 3, 4). The tabular beds form a laterally persistent succession with thickness that ranges from 2 to 5 m. The claystones are horizontally laminated, and formed by continuous microlamination with different grain sizes (figure 4). The ironstone beds are massive or laminated and rarely gradated; they range in thickness from 0.01 to 0.3 m (figure 4).

Figure 4. Exposures of the Los Rastros Formation in Río Gualo locality, A-B, Facies association A; C-D, Facies association B / fotos de la Formación Los Rastros en la localidad de Río Gualo, A-B, Asociación de facies A, C-D, Asociación de facies B.
The fossil remains preserved in the claystone facies include plant debris, and sometimes well-preserved plant fragments related to the "Dicroidium-type Flora"; conchostracans, insects and, rarely, fish are also recorded (Mancuso, 2003). On the other hand, the ironstone facies contains plant and wood remains, conchostracans, fish, and putative fish locomotion traces (cf. Undichnia) and grazing traces (Mancuso, 2003).
The features observed in the facies assemblage A suggest that it was deposited in a deep off-shore lacustrine depositional environment. The claystones were deposited by suspension in distal-lake conditions, whereas the ironstones by distal turbiditic currents (Farquharson, 1982; Mancuso, 2003; Talbot and Allen, 1996).
Facies assemblage B (delta deposits). This facies assemblage consists of a coarsening and thickening-upward succession with a thickness of 15 to 30 m. It is dominated by green siltstones, grey claystones, fine to coarse-grained sandstones and rare gravelly sandstones deposited by stream load (figures 3, 4). The facies assemblage begins with an alternation of green siltstones and gray claystones (Fm) with very fine sandstones interbedded, which form "rhythmic packages" of massive individual beds with a thickness of approximately 0.1 m (figure 4). The thin layers of small ripple cross-laminated fine to mediumgrained sandstones interbedded are more abundant and thicker upwards (figure 4). These layers preserved plant debris. Upward, medium to coarsegrained sandstones prevail. They begin with horizontal lamination (Sh), which passes upwards to planar cross-bedded (Sp) and culminate with trough cross-bedded (St) in the upper coarser material (figure 4). This coarsening and thickening-upward succession shows more abundant and thicker sandstone layers upwards, occasionally interbedded with scarce thin siltstone levels. The individual bed thickness ranges between 0.05 and 3 m. Fossil fish bodies and woody tissues were preserved in the horizontal laminated sandstones, while only fossil woods were remains found in the planar and trough cross-bedded sandstones (Mancuso, 2003, 2005a).
To the top, the assemblage is dominated by dark gray mudstones and very fine-grained sandstones (FSC) interbedded with planar and trough cross-bedded sandstones (figures 3, 4). They are internally massive, or pass upward from planar to rippled-lamination, and present a significant amount of organic matter, which are represented mainly by carbonized fragments. The important content of organic matter can form coal layers of 0.2 m thickness (figure 4) (Mancuso, 2003, 2005a).
Based on the stratal geometry and the interpretation of depositional mechanisms of the succession, this facies assemblage is interpreted as deltaic mouth-bar deposits (Guirin et al., 1994; Milana, 1998; Rogers et al., 2001; Mancuso, 2003, 2005a). The gradation from suspension deposits through cross-stratified sandstones represents a shallowing upward trend, which evidences the progradation of a mouthbar of the fluvial system into the lake body (see Farquharson, 1982; Talbot and Allen, 1996; Mancuso, 2003, 2005a).
Paleontology of the Los Rastros Formation
At present, the Los Rastros flora record is represented by an abundant macrofloristic assemblage related to the "Dicroidium-type Flora" (Frenguelli, 1948; Stipanicic and Bonetti, 1969; Stipanicic and Bonaparte, 1979; Kokogian et al., 1999; Spalletti et al., 1999) and a rich palynological association related to "Ipswich-type microflora" (Yrigoyen and Stover, 1970; Zavattieri and Melchor, 1999; Mancuso, 2005a; Ottone et al., 2005). Nevertheless the faunal record of the unit has increased during the last years. Invertebrate remains include different groups of conchostracans (clam shrimps) (Gallego, 1999; Mancuso and Gallego, 2000; Mancuso, 2005a), insects (Gallego, 1997; Gallego and Martins-Neto, 1999; Martins-Neto and Gallego, 1999, 2001; Mancuso and Gallego, 2000; Martins-Neto et al., 2003, 2005, 2006; Mancuso, 2005a) and bivalves (Frenguelli, 1945). The vertebrate fauna is represented by skeletal remains and ichnites. The skeletal remains are dominated by actinopterygian fishes (Cabrera, 1944; Forster et al., 1995; Mancuso, 2003), and a single temnospondyl amphibian (Contreras et al., 1997; Mancuso, 2002). The remaining tetrapod fauna is represented by non-mammalian therapsid, archosaur and possible dinosaur footprints (von Huene, 1931; Arcucci et al., 1995; Marsicano et al., 2004).
Preservation and taphonomy
The insect remains include blattopteran and hemipteran wings, and coleopteran elytra and fragmented bodies (figure 5). This material is preserved as hard parts, for example elytra, and/or impressions with partial or total loss of the original material, mainly through recent weathering.

Figure 5. Insect remains exhumed of the Los Rastros Formation, A-B, blattopteran wings, C, nearly complete coleopteran bodies, D, articulated coleopteran elytra, E, coleopteran abdomen, F, coleopteran elytra. / restos de insectos exhumados de la Formación Los Rastros, AB, alas de blattóptero, C, cuerpo de coleóptero casi completo, D, élitros de coleóptero articulados, E, abdomen de coleóptero, F, élitro de coleóptero. Scale bar 3 mm. / Escala gráfica 3 mm.
Most of the insect remains are constituted by disarticulated wings and elytra, and, only in some cases, the articulated elytra and nearly complete coleopteran bodies were found (figures 5.A-F). Thus, the state of articulation of the remains observed in this assemblage is mainly isolated and dispersed, and sometimes articulated.
With regard to postmortem modification, only few fractures were observed in blattopteran wings (figures 5.A, 5.B), mainly related to recent surface exposure and collection. The remaining insect material does not show evidence of postmortem modification (e.g. abrasion, cracking, breakage).
All insect remains have been found in the laminated black shales of the offshore-lacustrine facies, and show a spatial arrangement pattern parallel to the bedding plane (figure 5). The material has been reported to be, in some cases, preserved in several successive levels, and without preferential orientation.
The association shows a moderate to low abundance of remains. Thus, the biofabric is characterized by a moderate to low density of accumulation, a dispersed packing, and a good sorting in size and shape.
In the insect-bearing facies, plant debris, conchostracans and isolated fish scales (figure 6) are also preserved. Only in seldom cases, these remains have been reported to be directly associated with insect material.
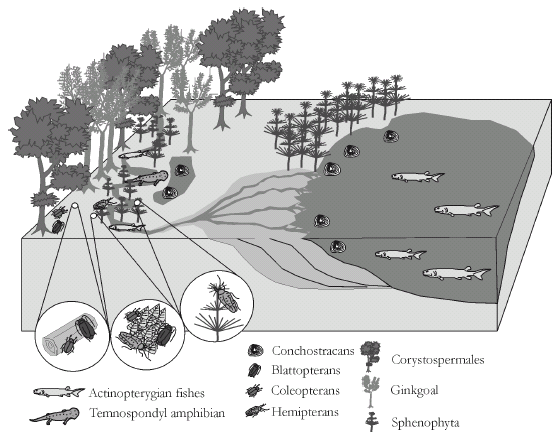
Figure 6. Paleobiological reconstruction of the Los Rastros biota taken from Mancuso, 2005a. It shows the different habitats and diet that the blattopterans, coleopterans and hemipterans may have developed. Organisms and environment are out of scale / reconstrucción paleobiológica de la biota de Los Rastros tomado de Mancuso, 2005a. Se muestran los diferentes hábitats y dietas que los blattópteros, coleópteros y hemípteros pudieron desarrollar. Organismos y ambiente fuera de escala.
All collected insect specimens are winged organisms with terrestrial habit (Gallego, 1997; Gallego and Martins-Neto, 1999; Martins-Neto and Gallego, 1999, 2001; Martins-Neto et al., 2003, 2005, 2006). Thus, they are considered allochthonous to the lacustrine depositional sub-environment where they were found.
Paleobiology and paleoecology
The Los Rastros insect assemblage includes representatives of different orders: Blattoptera (cockroaches), Coleoptera (beetles), Hemiptera (planthoppers), Ensifera (crickets), Glosselytrodea, Odonatoptera (dragonflies), Miomoptera, and Plecoptera (stoneflies) (Martins-Neto et al., 2003, 2005, 2006).
Blattoptera is the most conspicuous group among the described species. The extant nymph and adult of this order are terrestrial omnivores. They are primarily nocturnal, unspecialized scavengers in the leaf litter, feeding largely on decomposing plant material (figure 6). The abundance and diversity of the Los Rastros blattopterans was probably due to the large Dicroidium forests and the Taeniopteris and Baiera thickets around or next to the Los Rastros lake area, which provided many ecotopes where the blattopterans could survive, consuming litter. They had a major role in the mechanical reduction of litter (see also Martins-Neto et al., 2005).
The coleopteran assemblage is mainly composed by members of the suborder Archostemata (Protocoleoptera sensu Crowson, 1975). This primitive group (presence of the crossveins ornamentation) includes the Permosynidae and Cupedidae families, which are closely related phylogenetically, and known from the Lower Permian. The Permosynidae vanished in the Jurassic. The life habit of the fossil and extant forms probably are the same, and the recent cupedids are characterized by living in different habitats such as closed forest, open forest, rotting wood and woodland (figure 6). Many of the cupedid species are endemic, known only in limited areas (Neboiss, 1960, 1984). Like the Ommatidae they can be found in rainforests as well as in arid grazing or open woodland. Adults are poor fliers (Atkins, 1958). The structure of adult mouthparts suggests pollen-feeding habits; this is supported by the presence of pollen grains in the gut of one species and observations of another species frequenting flowers (Crowson, 1962). All known larvae feed in dead wood, which may be firm and dry, but has been attacked by wood-rotting fungi (figure 6). The other coleopterans belong to the Polyphaga suborder and the Elateridae family (represented up today), which are common and abundant throughout the world (Lawrence, 1982). The biology of this family varies considerably with the larvae being found in a variety of habitats. Larval stages can be found under the bark of trees, in rotten wood or in the soil. Larvae of different genera can live adjacent to termite nests and feed upon their inhabitants, the soil, where they prey upon other coleopteran larvae, or partially rotten wood, feeding upon cerambycid coleopteran larvae. However, the natural habitat of the larval stage of numerous elaterid species is either grassland or pasture (Calder, 1996).
The Hemiptera order is represented by Cercopoidea (froghoppers, planthoppers), Dysmorphoptilidae and Scytinopteridae families (Martins-Neto et al., 2003). In general extant forms of the hemipterans have jumping adults and underground scavenger nymphs. They consume leaves and roots of great variety of non-herbaceous plants (figure 6). The other hemipteran represented in the Los Rastros is a small group: the Coleorrhyncha. Today, it is only present in the southern hemisphere, living in caves, leaf-litter of the southern beech-tree forest or feeding on wet mosses (Bryophyta) (figure 6).
The Ensifera (crickets), are one of the less common insect orders represented in the Los Rastros fossil biota, together with the Miomoptera, Plecoptera and Odonatoptera (Martins-Neto et al., 2003). These Triassic ensiferans belong to the Haglidae family, which had a great diversity from the late Permian to the early Cretaceous (Sharov, 1968; Storozhenko, 1997) but only five relict species survive today. Most of the available biological information on the family is from haglines. The recent hagline genera inhabit coniferous taiga zones (Morris and Gwynne, 1978; Storozhenko, 1980), and emerge in spring being only active at night when the insects move into low bushes or coniferous trees to mate and to feed on vegetation (staminate cones and leaf-galls of sagebrush are known food items). During daylight, adults and nymphs retreat into shallow burrows, often under stones. Oviposition behavior is unknown, but as female haglines have lost the sword-like ovipositor characteristic of most Ensifera, they probably lay eggs in burrows; the few ensiferans that show maternal care of eggs or young have a reduced or absent ovipositor (Gwynne, 1995).
Living Tettigoniidae, the closest related group to the Haglidae, are characterized by living in vegetated areas with trees and bush, and they are represented by both carnivorous and omnivorous forms. The species of Notopamphagopsis (Haglidae) was one of the unique sound producers amon the insect fossil biota, and its sound probably was strong if we consider wing span reported from other species of this genus (fore wing length 36 mm in N. bolivari Cabrera from the Potrerillos Formation, Mendoza province).
The aquatic component of the Los Rastros fossil biota is very scarce, but Wilson (1988) suggests that under-representation of aquatic groups is common in the lacustrine fossil record. In the Los Rastros succession the only indirect evidence of this fact, up to date, is the record of adult forms of the orders Miomoptera, Plecoptera and Odonatoptera. Their nymph stages have aquatic habits, and are active predators. Immature stoneflies are aquatic nymphs (naiads) that usually live under stones in fast-moving, well-aerated water. Most species are herbivorous, feeding on algae and other submerged vegetation, but two families are predators on mayfly nymphs (Ephemeroptera) and on other small aquatic insects. Adult stoneflies emerge from their streams and can generally be found on the banks next to their previous habitat. They are not active fliers and usually remain near the ground where they feed on algae or lichens. In many species, the adults are short-lived and do not have functional mouthparts. Stoneflies are most abundant in cool climates with a few exceptions in the southern hemisphere living on damp soil; stonefly nymphs dwell in aquatic habitats. The preferred habitat is in rocky streams with a noticeable current at least. Some species can live in damp sandy areas. The usual habitat in running water that contains rocky, stony, or gravel substrata with more diversity in cooler, swifter water. Some studies have shown a correlation between certain species and certain habitats. For instance, Perlidae and Perlodidae are usually encountered under large stones, while Chloroperlidae tends to appear in gravel and Pteronarcyidae are frequently found in leaf packs. Extant members of the Gripopterigiidae (Plecoptera) have aquatic nymphs, with both predator and herbivorous habits (Richard and Davies, 1984).
Discussion
Biostratinomic processes are very important as control on insect preservation, and are mainly responsible for the disarticulation and fragmentation of organisms. Particularly in the aquatic setting, the floating time of insects in the surface is uneven, and is also an important control on preservation (Martinell and Martinez-Delclós, 1990; Martinez-Delclós and Martinell, 1993; Martinez-Delclós et al., 2004). One of the main variables is whether the insect arrives to the water dead or alive. The insects that die in aerial or terrestrial conditions, and are later introduced into water, float for a longer time. In contrast, live insects fall to the bottom in relatively short time. This is caused by the increase of their weight, due to water penetrating into the body when the individuals try to escape from surface tension (Martinell and Martinez-Delclós, 1990; Martinez-Delclós and Martinell, 1993; Martinez-Delclós et al., 2004).
During floating time on the water surface, insects may undergo several processes. The main process, in the aquatic setting, is the biologic activity produced by possible predators such as other insects, fishes or amphibians. They eat most of the insect and result in an important taphonomic filter (Martinell and Martinez-Delclós, 1990; Martinez-Delclós and Martinell, 1993; Martinez-Delclós et al., 2004). The predators may also produce insect disarticulation and/or fragmentation. The predator attack may be random or selective, thus the soft tissues, as the abdomen, are selected, whereas other tissues, like wings, are despised. These attacks produce fragments with different floating potential (Martinell and Martinez-Delclós, 1990; Martinez-Delclós and Martinell, 1993; Martinez-Delclós et al., 2004). The decomposition is the other process that affects the insect body on the water surface and any biological or physical disturbance promotes fragmentation and rapid decay.
The floating time depends on several factors. The relation between volume and surface of the insect body is one of these factors. For example, the rough outside organisms show void floating time whereas, the organisms with big wing surface can experience complete decomposition processes on the water surface (Martinell and Martinez-Delclós, 1990; Martinez-Delclós and Martinell, 1993; Martinez-Delclós et al., 2004). During the decay process, the water infilled the soft tissue producing the increase of insect weight and the fall through the water column. However, there are external factors that produce the fall of the remains, such as rain, waves produced by wind, and variation of surface tension. Moreover, push from other organisms may produce selective fall, as observed by Martinell and Martinez-Delclós (1990) where the shocks produced by crustaceans cause some body parts to fall and leave the rest of the body afloat.
The fall of an insect carcass, in a quiet environment, is usually slow and vertical; during the fall the same processes as described above may affect them. So, the biological attacks may both occur on water surface or during the fall through the oxygenated zone. Besides, the insects may also suffer the normal degradation of the soft tissues during their fall in the oxygenated zone (Martinell and Martinez-Delclós, 1990; Martinez-Delclós and Martinell, 1993; Martinez-Delclós et al., 2004).
Wilson (1980, 1988) observed that the insect taxa with weak flying ability (e.g. Coleoptera, Hemiptera Fulgoroidea, Hemiptera, Heteroptera) are relatively more common at near-shore facies, whereas insect taxa with strong flying ability (Diptera, Hymenoptera) are more common in offshore associations.
Generally, the insect-bearing sediments show evidence of anoxic conditions such as absence of benthonic organisms, invertebrate trails and bioturbation. Therefore, insect disarticulation and/or fragmentation presumably occurred during the arrival to the water, floating, or sinking through the oxygenated zone. In contrast, the presence of partially or totally articulated insects suggests that they probably arrived alive to the water and suffered a relatively rapid fall. Thus, the insect remains were safeguarded from attacks and degradation on the water surface and in the oxygenated zone. Finally, there are no biologic and/or physical disturbing agents in the bottom.
The Los Rastros insects are mainly adult, terrestrial and winged, and they are considered allochthonous to the lake. Thus, they may stem from the paleo-shoreline of the lake or from more distant sectors into the affluent system. The insect record includes from almost complete individuals to isolated wings, and its taphonomic features allow to characterize several possible pathways. The disarticulated insect remains represented by isolated wings (blattopteran and hemipteran wings and coleopteran elytra) can be produced by three possible pathways. The first pathway is that the insect remains arrived already disarticulated to the lake; the second is that the insects are selectively attacked and only wings are preserved; and the third is that the individuals suffered the normal decay on the water surface and/or during sinking through the oxygenated zone. Another pathway is denoted by the articulated remains represented by almost complete individuals (coleopteran elytra and bodies), which suggests that some living organisms arrived to the lake and suffered a relatively rapid fall, and housed in the anoxic bottom, where they were saved from biological attacks and decay.
Conclusions
The insect assemblage from the Los Rastros Formation bears the same composition as the other known classic Triassic Gondwana deposits, specially constituted by isolated wings of Hemiptera, Coleoptera and Blattoptera. They suggest a global taphonomic pattern and fossilization process, with a complex of anoxic and calm lakes and streams associated to a low diversity pattern, after the Permian-Triassic transition. The whole insect assemblage is essentially allochthonous, and the remarkable amount of isolated wings of different types and thickness, suggests different taphonomic histories. The thick elytra of the coleopteran specimens suffer less fragmentation like (in a minor scale) the fragmentation of the coreiceous wings of the cercopoid-like Dysmorphoptilidae. The presence of articulated elytra indicates that specimens arrived alive to the lake, suffered a relatively rapid fall through the water column, and were housed in the anoxic bottom, which preserved them intact. The blattopteran wings, together with the more fragile wings of other insect groups, are more liable to undergo fragmentation during transport and long periods of flotation on the water surface, as well as during the fall through the water column by the attack of other organisms, like conchostracans and small fishes.
Acknowlegdements
We greatly acknowledge to T. Heredia for the critical reading of the manuscript and his pertinent comments; A. Caselli, C. Marsicano and A. Arcucci for their suggestions and assistance in the field; C. Forster, C. May and R. Rogers from National Geographic Society Grant 5317-94, and M.E. Gonaldi from Museo de Ciencias Naturales (Universidad Nacional de La Rioja, Argentina) for her assistance with the catalogue of the specimens. Field research was supported by PIP CONICET 0535 (A. Arcucci). This research was partly supported by the Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) and by the Secretaría General de Ciencia y Técnica, Universidad Nacional del Nordeste Grant PI-64/04 (O.F. Gallego).
References
1. Atkins, M.D. 1958. Observations on the flight, wing movements and wing structure of male Priacma serrata (Lec.) (Coleoptera:Cupedidae). Canadian Entomology 90: 339-347 [ Links ]
2. Arcucci, A.B., Forster, C., Abdala, F., May, C. and Marsicano, C. 1995. "Theropods" tracks from the Los Rastros Formation (Middle Triassic), La Rioja Province, Argentina. Journal of Vertebrate Paleontology 15, supplement 3: 16A. [ Links ]
3. Bossi, G. 1977. La Formación Cerro Rajado, Provincia de La Rioja. Acta Geológica Lilloana 14: 19-40. [ Links ]
4. Cabrera, A. 1944. Dos nuevos peces ganoideos del Triásico de Argentina. Notas del Museo de La Plata 9, Paleontología 81: 569-576. [ Links ]
5. Calder, A.A. 1996. Click Beetles. Genera of Australian Elateridae (Coleoptera). Monography of Invertebrate Taxonomy 2: i-x, 1-401. [ Links ]
6. Caselli, A.T., Marsicano, C. and Arcucci, A.B. 2001. Sedimentología y paleontología de la Formación Los Colorados, Triásico superior (provincias de La Rioja y San Juan, Argentina). Revista de la Asociación Geológica Argentina 56: 173-188. [ Links ]
7. Contreras, V.H., Bracco, A. and Sill, W. 1997. Primer registro de restos de tetrápodos en la Formación Los Rastros (Triásico Medio), Parque Provincial Ischigualasto, San Juan, Argentina. Ameghiniana 34: 534. [ Links ]
8. Crowson, R.A. 1962. Observations on the beetle family Cupedidae, with descriptions of two new fossil forms and a key to the recent genera. Annual Magazine Natural History 13: 147-157. [ Links ]
9. Crowson, R.A. 1975. The evolutionary history of Coleoptera, as documented by fossil and comparative evidence. 10º Congressi Nazionali. Italiani Entomologi, Atti, pp. 47-90. [ Links ]
10. Farquharson, G.W. 1982. Lacustrine deltas in a Mesozoic alluvial sequence from Camp Hill, Antarctica. Sedimentology 29: 717-725. [ Links ]
11. Forster, C., Arcucci, A., Marsicano, C., Abdala, F. and May, C. 1995. New vertebrate material from the Los Rastros Formation (Middle Triassic), La Rioja Province, Northwestern Argentina. Journal of Vertebrate Paleontology 15, supplement 3: 29A. [ Links ]
12. Frenguelli, J. 1944. La serie del llamado "Rético" en el oeste argentino (nota preliminar). Notas Museo de La Plata 9, Geología 30: 261-269. [ Links ]
13. Frenguelli, J. 1945. Moluscos continentales en el Paleozoico Superior y en Triásico de la Argentina. Notas del Museo de La Plata 10, Paleontología 83: 180-204. [ Links ]
14. Frenguelli, J. 1948. Estratigrafía y edad del llamado Rético en la Argentina. Anales de la Sociedad Argentina de Estudios Geográficos, Sociedad Argentina de Estratigrafía Geológica, GAEA, Buenos Aires, 8: 159-310. [ Links ]
15. Gallego, O.F. 1997. Hallazgos de Insectos triásicos en la Argentina. Ameghiniana 34: 511-516. [ Links ]
16. Gallego, O. F. 1999. [Estudio Sistemático de las Faunas de Conchóstracos Triásicos de la República Argentina. PhD thesis of Universidad Nacional de Córdoba, Córdoba, Argentina, 210 pp. Unpublished.]. [ Links ]
17. Gallego, O.F. and Martins-Neto, R.G. 1999. La entomofauna mesozoica de Argentina. Estado actual del conocimiento. Revista de la Sociedad Entomológica Argentina 58: 86-94. [ Links ]
18. Groeber, P. and Stipanicic, P.N. 1953. Mesozoico: Triásico, Jurásico y Cretácico. Geografía de la República Argentina 2, GAEA. 1º parte, Sociedad Argentina de Estratigrafía Geológica, Buenos Aires pp. 13-141. [ Links ]
19. Guirin, R.A., Limarino, C.O., Caselli, A.T., Net, L. and Page, S.M. 1994. Sedimentología de las facies lacustres de la Formación Lomas Blancas (Triásico Medio, Cuenca Ischigualasto-Villa Unión). 5º Reunión Argentina de Sedimentología (San Miguel de Tucumán), Actas (supl.): 307-312. [ Links ]
20. Gwynne, D.T. 1995. Phylogeny of the Ensifera (Orthoptera): a hypothesis supporting multiple origins of acoustical signalling, complex spermatophores and maternal care in crickets, katydids, and weta. Journal of Orthoptera Research 4: 203-218. [ Links ]
21. Huene, F. von 1931. Die fossilen Fährten im Rhät von Ischigualasto in Nordwest-Argentinien. Palaeobiologica 4: 99-112. [ Links ]
22. Kokogian, D., Spalletti, L., Morel, E., Artabe, A., Martínez, R., Alcober, O., Milana, J., Zavattieri, A. and Papú, O.H. 1999. Los depósitos continentales triasicos. In: R. Caminos and J. Panza (eds.), Geología Regional Argentina, Instituto de Geología y Recursos Minerales, Anales 29: 377- 398. [ Links ]
23. Lawrence, J.F., 1982. Coleoptera. pp. 482-553. In: S.P. Parker (ed.), Synopsis and Classification of Living Organisms. New York, McGraw-Hill Vol. 2, 1232 pp. [ Links ]
24. López Gamundí, O.R., Alvarez, L., Andreis, R., Bossi, G., Espejo, I., Fernandez-Seveso, F., Legarreta, L., Kokogian, D., Limarino, O. and Sesarego, H. 1989. Cuencas Intermontanas. In: G. Chebli, and L. Spalletti (eds.), Cuencas Sedimentarias Argentinas, Serie Correlación Geológica Nº 6, Instituto Superior de Correlación Geológica, Universidad Nacional de Tucumán, pp. 123-167. [ Links ]
25. Mancuso, A.C. 1999. [Análisis tafonómico de una asociación fósil continental del Triásico Medio de la Cuenca de Ischigualasto-Villa Unión (Provincia de La Rioja). Lic. thesis, Universidad de Buenos Aires, Buenos Aires, 71 pp. Unpublished.]. [ Links ]
26. Mancuso, A.C. 2002. Vertebrate taphonomy in a Triassic lake: tetrapods of the Los Rastros Formation (Ischigualasto-Villa Unión Basin, central-western Argentina). 8º Congreso Argentino de Paleontología y Bioestratigrafía, Actas: p. 35. [ Links ]
27. Mancuso, A.C. 2003. Continental fish taphonomy: a case study in the Triassic of Argentina. Journal of South American Earth Sciences 16: 275-286. [ Links ]
28. Mancuso, A.C. 2005a. [Tafonomía en ambientes lacustres: estudio integral de las asociaciones fósiles de las secuencias lacustres del Triásico Medio de la Cuenca de Ischigualasto-Villa Unión (formaciones Chañares, Los Rastros e Ischichuca). PhD thesis of Universidad de Buenos, Aires Buenos Aires, Argentina, 208 pp. Unpublished.]. [ Links ]
29. Mancuso, A.C. 2005b. Revisión y aportes a la estratigrafía de la sección inferior del Grupo Agua de la Peña (Triásico Medio, Argentina). 16º Congreso Geológico Argentino (La Plata), Actas 3: 415-422. [ Links ]
30. Mancuso, A.C. and Gallego O.F. 2000. Primer análisis tafonómico de una asociación fósil de conchóstracos e insectos del Triásico medio (Fm. Los Rastros) de la Argentina. 1º International Meeting on Paleoarthropodology (San Pablo), Actas p. 64 [ Links ]
31. Marsicano, C. A., Arcucci, A., Mancuso, A. C. and Caselli, A. 2004. Early-Middle Triassic tetrapod footprints of southern South America: a phylogenetic approach. Ameghiniana 41: 171-184. [ Links ]
32. Martinell, J. and Martinez-Delclós, X. 1990. Observaciones de laboratorio sobre la flotabilidad de los insectos. Reunión de Tafonomía y Fosilización (Madrid), pp. 201-209. [ Links ]
33. Martínez-Declòs, X. and Martinell, J. 1993. Insect taphonomy experiments: Their application to the Cretaceous outcrops of lithographic limestones from Spain. Kaupia 2: 133-144. [ Links ]
34. Martínez-Declòs, X., Briggs, D.E.G. and Peñalver, E. 2004. Taphonomy of insects in carbonates and amber. Palaeogeography, Palaeoclimatology, Palaeoecology 203: 19-64. [ Links ]
35. Martins-Neto, R.G. and Gallego, O. 1999. The palaeontomofauna from Argentina. I. Auchenorrhyncha, Miomoptera and Ensifera. Revista Española de Paleontología 14: 27- 38. [ Links ]
36. Martins-Neto, R.G. and Gallego, O.F. 2001. The Triassic insect fauna from Argentina. IV. Glosselytrodea and complements on Auchenorhyncha. Acta Geologica Leopoldensia 52: 74-83. [ Links ]
37. Martins-Neto, R.G., Gallego, O.F. and Melchor, R.N. 2003. The Triassic insect fauna from South America (Brazil, Argentina and Chile): a checklist (except Blattoptera and Coleoptera) and descriptions of new taxa. Acta Zoologica Cracoviensia (suppl.-Fossil Insects) 46: 229-256. [ Links ]
38. Martins-Neto, R.G., Mancuso, A.C. and Gallego, O.F. 2005. The Triassic insect fauna from Argentina. Blattoptera from the Los Rastros Formation (Bermejo Basin), La Rioja Province. Ameghiniana 42: 705-723. [ Links ]
39. Martins-Neto, R.G., Gallego, O.F. and Mancuso, A.C. 2006. The Triassic insect fauna from Argentina. Coleoptera from the Los Rastros Formation (Bermejo Basin), La Rioja Province. Ameghiniana 43: 591-609. [ Links ]
40. Milana, J.P. 1998. Anatomía de parasecuencias en un lago de rift y su relación con la generación de hidrocarburos, cuenca triásica de Ischigualasto, San Juan. Revista de la Asociación Geológica Argentina 53: 365-387. [ Links ]
41. Milana, J.P. and Alcober, O. 1994. Modelo tectosedimentario de la cuenca triásica de Ischigualasto (San Juan, Argentina). Revista de la Asociación Geológica Argentina 49: 217-235. [ Links ]
42. Morris, G.K. and Gwynne, D.T. 1978. Geographical distribution and biological observations of Cyphoderris (Orthoptera: Haglidae) with a description of a new species. Psyche 85: 147- 167. [ Links ]
43. Neboiss, A. 1960. On the family Cupedidae, Coleoptera. Proceedings of the Royal Society of Victoria 72: 12-20. [ Links ]
44. Neboiss, A. 1984. Reclassification of Cupes Fabricius (s. lat.), with descriptions of new genera and species (Cupedidae: Coleoptera). Systmatic Entomology 9: 443-477 [ Links ]
45. Ortiz, A. 1968. Los denominados Estratos de Ischichuca como sección media de la Formación Los Rastros. 3º Jornadas Geológicas Argentinas (Comodoro Rivadavia) 1: 333-341. [ Links ]
46. Ottone, E.G., Mancuso, A.C. and Resano, M. 2005. Miospores and Chlorococcales from the Los Rastros Formation, Middle Triassic of central- western Argentina. Ameghiniana 42: 347-362. [ Links ]
47. Richards, O.W. and Davies, R.G. 1984. Tratado de Entomología Imms. Vol. 2: Clasificación y Biología. Ediciones Omega S.A., Barcelona, 998 pp. [ Links ]
48. Rogers, R.R., Arcucci, A.B., Abdala, F., Sereno, P.C., Forster, C.A. and May, C.L. 2001. Paleoenvironment and Taphonomy of the Chañares Formation Tetrapod Assemblage (Middle Triassic), Northwestern Argentina: Spectacular Preservation in Volcanogenic Concretions. Palaios 16: 461-481. [ Links ]
49. Romer, A.S. and Jensen, J.A. 1966. The Chañares (Argentina) triassic reptile fauna. II Sketch of the geology of the Rio Chañares-Rio Gualo region. Breviora 252: 1-20. [ Links ]
50. Sharov, A.G. 1968. Phylogeny of the Orthopteroidea. Trudy Paleontologicheskogo Instituta 118: 1-251. [ Links ]
51. Spalletti, L.A., Artabe, A., Morel, E. and Brea, M. 1999. Biozonación paleoflorística y cronoestratigrafía del Triásico Argentino. Ameghiniana 36: 419- 451. [ Links ]
52. Stipanicic, P.N. 1969. El avance en los conocimientos del Jurásico argentino a partir del esquema de Groeber. Revista de la Asociación Geológica Argentina 24: 367-388. [ Links ]
53. Stipanicic, P.N. 2002. Introducción. In: P.N. Stipanicic and C. Marsicano (eds.), Léxico Estratigráfico de la Argentina: Triásico. Asociación Geológica Argentina, volumen 8, Serie "B" (Didática y Complementaria), pp. 1-24. [ Links ]
54. Stipanicic, P.N. and Bonaparte, J. 1979. Cuenca triásica de Ischigualasto-Villa Unión (Provincia de La Rioja y San Juan). In: Geología Regional Argentina, Academia Nacional de Ciencias (Córdoba) 1: 523- 575. [ Links ]
55. Stipanicic, P.N. and Bonetti, M.I.R. 1969 Consideraciones sobre la cronología de los terrenos triásicos argentinos. In: Gondwana Stratigraphy IUGS Symposium, Mar del Plata, UNESCO, Ciencias de la Tierra 2: 1081-1120. [ Links ]
56. Storozhenko, S. Yu., 1980. Haglidae (Orthoptera)-a new family for the URSS fauna. Revue d'Entomologie de l'URSS 59: 114-117. [ Links ]
57. Storozhenko, S. Yu., 1997. New Triassic genera of the Family Atactophlebiidae (Grylloblattida). Far Easthern Entomologist 40: 8. [ Links ]
58. Talbot, M.R. and Allen, P.A. 1996. Lakes. In: H.G. Reading (ed.), Sedimentary Environments: Processes, Facies and Stratigraphy, Blackwell Science, Oxford, pp. 5-36. [ Links ]
59. Uliana, M.A. and Biddle, K. 1988. Mesozoic-Cenozoic paleogeographic and geodynamic evolution of southern South America. Revista Brasileira de Geociências 18: 172-190. [ Links ]
60. Vail, P.R., Audermard, F., Bowman, S. A., Eisner, P.N. and Perez Cruz, C. 1991. The stratigraphic signatures of tectonics, eustacy and sedimentology- an overview. In: G. Einsele, G. Ricken and A. Seilacher (eds.), Cycles and events in stratigraphy, Springer-Berlag, Berlin, pp. 617-659. [ Links ]
61. Wilson, M.V.H. 1980. Eocene lake environments: depth and distance-from-shore variation in fish, insect, and plant assemblages. Palaeogeography, Palaeoclimatology, Palaeoecology 32: 21-44. [ Links ]
62. Wilson, M.V.H. 1988. Reconstruction of ancient lake environments using both autochthonous and allochthonous fossils. Palaeogeography, Palaeoclimatology, Palaeoecology 62: 609-623. [ Links ]
63. Yrigoyen, M.R. and Stover, L.E. 1970. La palinología como elemento de correlación del Triásico en la Cuenca Cuyana. 4° Jornadas Geológicas Argentinas (Mendoza), Actas 2: 427-447. [ Links ]
64. Zavattieri, A.M. and Melchor, R.N. 1999. Estudio palinológico preliminar de la Formación Ischichuca (Triásico), en su localidad tipo (quebrada de Ischichuca Chica), provincia de La Rioja, Argentina. Asociación Paleontológica Argentina, Publicación Especial 6: 33-38. [ Links ]
Recibido: 19 de agosto de 2005.
Aceptado: 29 de noviembre de 2006.

