










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.44 n.2 Buenos Aires abr./jun. 2007
A new Cretaceous chelid turtle from the northern Neuquén Basin, Argentina
Marcelo S. De La Fuente1, Jorge O. Calvo2 and Bernardo J. González Riga3
1CONICET. Museo de Historia Natural de San Rafael Parque Mariano S/Nº, 5600 San Rafael, Mendoza, Argentina. mdlafu@infovia.com.ar
2Centro Paleontológico Lago Barreales, Universidad Nacional del Comahue. Proyecto Dino, Ruta Prov. 51, km 65, 8300 Neuquén, Argentina. jorgecalvo@digimedia.com.ar
3CONICET. Unidad de Paleovertebrados, Instituto Argentino de Nivología, Glaciología y Ciencias Ambientales, Centro Regional de Investigaciones Científicas y Tecnológicas, Avda. Ruiz Leal s/n, Parque Gral. San Martín, 5500 Mendoza, Argentina. bgonriga@lab.cricyt.edu.ar
Introduction
The Family Chelidae is a group of freshwater aquatic to semi aquatic pleurodiran turtles with extant species distributed in South America and Australasia (Pritchard and Trebbau, 1984; Iverson, 1992). Its fossil record is known as early as the Albian in Patagonia (Lapparent de Broin and de la Fuente, 2001), Paleocene?Eocene in Australia (Lapparent de Broin and Molnar, 2001), and Oligocene in Tasmania (Warren, 1969). This extant disjoint geographic range and the early fossil record of this family might be a result of an old biogeographical and phylogenetic history that was developed in Gondwanan landmasses (Broin, 1987, 1988; de la Fuente, 1988, 1992, 1997; Broin and de la Fuente, 1993a, 1993b). These turtles are characterized by two main authapomorphic conditions: the formulae of articulation of the cervical vertebrae including, after the biconcave atlas, three opisthocoelous (second to fourth), two biconvexes (fifth and eight), one procoelous (sixth), and one biconcave (seventh) [This condition is shown as)1((2((3((4((5))6))7((8), sensu Williams (1950) notation], and the developed lateral cheek emargination with loss of quadratojugal (see Gaffney, 1977; Broin and de la Fuente, 1993b).
The knowledge of Cretaceous chelids begun in Patagonia with Broin´s study of chelonian fauna of Los Alamitos Formation in the context of Bonaparte´s research of the fauna of Los Alamitos (Bonaparte et al., 1984; Bonaparte, 1986, 1987). The preliminary Broin´s study was followed by studies of additional and more complete chelid material recovered from several localities. They were included in synthetic articles about the Argentinean chelonian fauna (Broin and de la Fuente, 1993a, 1993b) and more detailed studies (Gasparini and de la Fuente, 2000; de la Fuente et al., 2001; de la Fuente, 2003; Lapparent de Broin and de la Fuente, 2001). References about them can also be seen in abstracts (Lapparent de Broin et al., 1997; Lapparent de Broin and de la Fuente, 1999; de la Fuente, 1997; de la Fuente et al., 2000) or in synthetic articles about the tetrapod assemblages of Patagonia (Bonaparte, 1996; Gasparini et al., 2001; Gasparini et al., 2002; Goin et al., 2002; Leanza et al., 2004).
Recently, a new partially preserved specimen of a chelid was recovered from an Upper Cretaceous horizon of the Neuquén Group. Although there are chelid specimens represented by complete shells, cervical vertebrae and skulls from this and other basins (de la Fuente et al., 2001; Lapparent de Broin and de la Fuente, 2001; Bona and de la Fuente, 2005), the specimen here described preserves an almost complete forelimb, skeletal elements unknown in Patagonian chelids. The presence of this forelimb associated with a typical plastron and cervical vertebrae remains of chelids allows us to determine the specimen of Loma del Lindero as a new taxon of this family of pleurodiran turtles.
In northwestern Patagonia, the Neuquén Group is the richest reptiles bearing unit. It comprises a thick continental succession (maximum thickness 1300 m) deposited from early Cenomanian (97 Ma) to early Campanian (74 Ma) (Leanza, 1999; Leanza and Hugo, 2001; Leanza et al., 2004). In the last years, extraordinary titanosaurid sauropods were found in Rincón de los Sauces area, Northern Neuquén Province, Patagonia (Calvo et al., 1997). In the Cañadón Arroyo Río Seco site, located 2 km south to Rincón de los Sauces Town, a slender Eutitanosauria named Rinconsaurus caudamirus (Calvo and González Riga, 2003) and Pehuenchesuchus enderi, a basal sebecosuchian crocodyliform (Turner and Calvo, 2005), have also been found.
The main scope of this work is to describe a new species of chelid turtle from the Río Neuquén Subgroup. The fossils were found at Loma del Lindero site (figure 1), 10 km west to Rincón de los Sauces Town (Calvo and González Riga, 1999). This locality is an important paleontological site , that was excavated during four years (1998-2001) under the direction of Jorge O. Calvo, where numerous disarticulated titanosaur remains (nearly 300 bones) and theropods associated with the turtle specimen described herein have been found (González Riga and Calvo, 1999). The fossils were collected in mudstones and fine sandstones of overbank facies related to channel fluvial systems. In this quarry, numerous bones of titanosaur sauropods and other vertebrates are preserved in a taphonomic mode preliminary named "overbank bone assemblage" (González Riga et al., 2003). Turtle specimen is housed at the "Museo Argentino Urquiza-Rincón de los Sauces" (MAU-PV).

Figure 1. Location map of the Loma del Lindero locality / mapa de ubicación de la localidad de Loma del Lindero.
Systematic paleontology
TESTUDINES Batsch, 1788
PLEURODIRA Cope, 1865
CHELIDAE Lindholm, 1929
Linderochelys gen. nov.
Diagnosis. Short-necked chelid turtle having a plastron length of moderate size (290 mm) with an elongated abdominal seam. Differs from the extant taxa assigned to Phrynops Wagler (sensu lato) in the presence of cuneiform mesoplastra. Differs from Palaeophrynops patagonicus Lapparent de Broin and de la Fuente in the bell shape of entoplastron, and the more depressed common process of postzygapophyses. Differs from Bonapartemys bajobarrealis Lapparent de Broin and de la Fuente in a short plastral bridge, absence of ridges at the bridge limit of anterior plastral lobe, and in the different plastral scale formulae. Differs from Prochelidella portezuelae de la Fuente and the cenomanian Prochelidella spp. in a large and rounded anterior plastral lobe and the length of interadominal seam
Etimology. "Lindero" from the site Loma del Lindero; "chelys" from Greek turtle.
Linderochelys rinconensis sp. nov.
Diagnosis. The same as the genus by monotypy.
Etimology. "Rinconensis" species from the town of Rincon de Los Sauces.
Holotype. MAU-PV-LL 69. A fragment of the suprapygal and the eight right costal bone of the carapace, a partially preserved plastron, one cotyle and one condyle probably of a second to fourth opisthocoelus vertebrae or a seventh biconcave vertebrae, as well as fragments of two postzigapophyses of cervical vertebrae, left humerus, radius and proximal carpal elements and right metacarpals and digits, the pelvic girdle, femora and fibula, and right tibia.
Locality and age. The turtle was recovered in mudstones and fine sandstones that crop out at Loma del Lindero site, 10 km West to Rincón de los Sauces City, Neuquén Province, Patagonia, Argentina (figure 1). These rocks belong to the Río Neuquén Subgroup, Neuquén Group, Late Cretaceous late Turonian-Coniacian according to Leanza (1999), Leanza and Hugo (2001) and Leanza et al. (2004).
Description
Shell. The carapace is poorly preserved and is represented by several small indeterminate fragments. Only one piece might be identified as one anterolateral fragment of the suprapygal bone sutured to the medial part of the eight right costal bone. On the visceral surface of this fragment a transversal elliptic section of the proximal ilium attached to the carapace bone is recognized (figures 2.1-2.2).
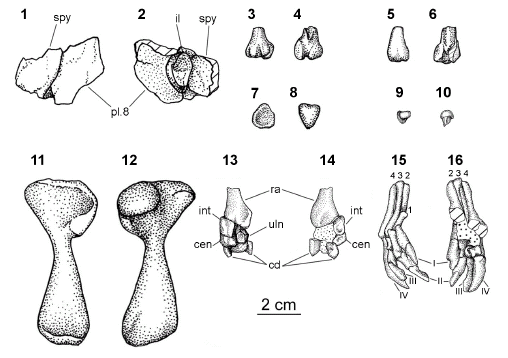
Figure 2. Linderochelys rinconensis gen et sp. nov. 1, dorsal view of the carapace / vista dorsal del caparazón; 2, ventral view of the carapace with remains of the ilium attached / vista visceral del caparazón con el ilion adherido; 3 and 5, dorsal view / vista dorsal; 4 and 6, ventral view of two posterior process of cervical vertebrae bearing postzigapophysis / vista ventral de dos procesos posteriores de las vértebras cervicales portadores de las postzigapófisis; 7-8, posterior and ventral view of a cotyle of a cervical vertebra / vista posterior y ventral de un cótilo de una vértebra cervical; 9-10, anterior and ventral view of a condyle of a cervical vertebra / vista anterior y ventral de un cóndilo de una vértebra cervical; 11-12, ventral and dorsal view of the left humerus / vista ventral y dorsal del húmero izquierdo; 13-14, dorsal and ventral view of the left radius and carpal bones / vista dorsal y ventral del radio izquierdo y huesos carpales; 15-16, dorsal view of right metacarpal bones and digits / vista dorsal de metacarpales y dígitos derechos. References / referencias. 1-2-3-4, metacarpals / metacarpales; I-II-III-IV, digits / dígitos; cen, centrals bones / huesos centrales; cd, distal carpals / carpales distales; int, intermedium; il, ilium, pl 8, pleural (= costal) bone 8 / placa pleural (= costal) 8; ra, radius / radio; spy, suprapygal bone / placa suprapigal; uln, ulnar.
The plastron (figures 3.1-3.2) is better preserved than the carapace and its length is estimated as 290 mm. Although the anterior plastral lobe is not complete and only the posterior part of the right plastral bridge is preserved, it is possible to estimate that the posterior plastral lobe is longer than the anterior one, and the bridge is the shortest plastral element. Likewise, the anterior and posterior plastral lobes are wider than longer. The left epiplastron is only visible in visceral view. The curved lateral margin of the left epiplastron suggests a curved anterior margin of the plastron lobe probably followed by subparallel hyoplastral margins. The epiplastron is longer than wide, and its peculiar shape in visceral view is defined posteriorly by a sinuous epihyoplastral suture. This sinuous suture runs from the entoplastron to the lateral margin of the anterior plastral lobe, thus resulting in an anterior and medial process of the hyoplastron that protrude the epiplastron. Although the entoplastron is partially preserved, a bell shape is recognized. Contrary to the anterior plastral bones (left epiplastron and the fragments of entoplastron), the hyo, hypo and xifhiplastron are partially preserved in ventral and visceral sides. The hyoplastra are the largest plastral bones. The hyoplastron is also longer than the hypoplastron, but it is shorter than xiphiplastron, mea-sured in the middle plastral line. The right mesoplastron is included between hyo and hypoplastron, and is short and wide. The scute pattern of the anterior plastral lobe is poorly preserved and only the humero-pectoral sulcus is preserved. It is strongly curved posteriorly, and is well posterior to the entoplastron. Only the left pectoro-abdominal sulcus is preserved. The pectoral scute (interpectoral seams) is the shortest preserved scute, while the abdominal scute (interabdominal seams) is the longest scute, measured in the middle plastral line. Consecuently, the plastral formulae of Linderochelys rinconensis gen. et sp. nov. is Ab > Fe >An > Pe. The lateral margins of the posterior plastral lobe are slightly convergent distally ending in xiphiplastral points that delimit a U shape anal notch.
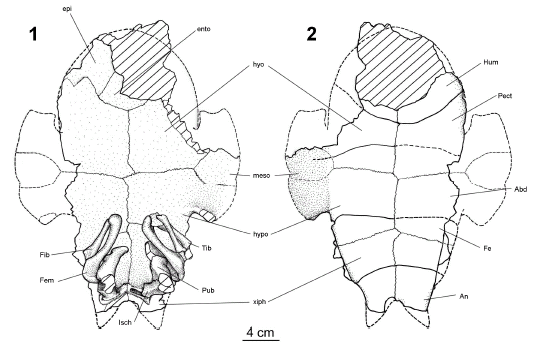
Figure 3. Linderochelys rinconensis gen et sp. nov. 1, visceral view of the plastron with remains of pelvic girdle, both femora, fibula and right tibia / vista visceral del plastrón con restos de la cintura pélvica, ambos fémures, fíbulas y la tibia derecha; 2, ventral view of the plastron / vista ventral del plastrón. References / referencias. Abd, abdominal scute / escudo abdominal; An, anal scute / escudo anal; ento, entoplastron / entoplastrón; epi, epiplastron / epiplastrón; Fe, femoral scute/escudo femoral; Fem, femur/fémur; Fib, fibula / fíbula; hyo, hyoplastron / hioplastrón; hypo, hypoplastron / hipoplastrón; Hum, humeral scute / escudo humeral; isch, ischion / isquión; meso, mesoplastral bone / placa mesoplastral; Pub, pubis / púbis; Pect, pectoral scute / escudo pectoral; xiph, xiphyplastron/ xifiplastrón.
Cervical vertebrae (figures 2.3-2.10). The cervical vertebrae are represented by fragmentary remains of a posterior part of a vertebral centrum with a concave subquadrangular cotyle. It is slightly elongated and narrow and without posterior inferolateral tuberosities like the posterior cotyle of the third or fourth opisthocoelous vertebrae of the extant Phrynops tuberculatus Luederwaldt. Other isolated remains of a centrum belong to an anterior condyle, subtriangular in shape, of the another opistocoelus or biconvex cervical vertebra with a well developed ventral keel, as well as fragments of two postzigapophyses oriented ventrolaterally of two cervical vertebrae.
Humerus (figures 2.11-2.12). The general morphology of left humerus is similar to other chelid species of the Phrynops group (e.g., Prochelidella portezuelae de la Fuente). However, it differs from the extant Phrynops and Acanthochelys Gray species in the robustness of this bone. It has a subtriangular articular head, a wide proximal end with a shallow intertubercular fossa lying between the large medial process and a small lateral process. The humerus narrows to a shaft, subcylindrical in section, that archs dorsally. Distally the shaft flattens dorsoventrally and ends in a broad extremity. The articular surface has a small ventrally facing capitellum where the radio articulates, and a more extensive trochlea for the ulna. An ectepycondilar open foramen and groove are recognized in a extreme distal position as it is typical in turtles.
Forearm and hand (figures 2.13-2.16). The left radius is only preserved by its expanded and flat distal end. This end contacts with displaced carpal elements. Among them it is recognized a trapezoidal inverted intermedium, an almost subpentagonal ulnar, with the long axis to 90 degree to normal horizontal position. These bones are moved from this natural position (articulate with two facets of the distal end of the ulna). Other carpal elements are interpreted as two central and two distal carpals. Some right carpal elements are also preserved. The right I to IV elongate metacarpal bones are recognizable. Only the III and IV are completely preserved. The I and II metacarpal are preserved in their distal and proximal parts. In dorsal view, the preserved distal metacarpal I overlaps the metacarpal II. Four digits (I-IV) are preserved in bad condition. As it is typical in turtles (as in many other tetrapods), the first digit is the shortest while the remaining digits are longer than the first. This difference in length between the first and the other digits suggests the presence of a typical phalangeal formulae of casichelydian turtles 2-3-3-3 for the four preserved digits.
Pelvic girdle (figure 3.1). The pelvic girdle is seen in dorsal view. The ilium, pubis and ischium are united to compose the acetabulum, seen on the left side where the head of the femur is slightly displaced. The left and right pubis and ischium are partially preserved. They are separated each other by a large thyroid fenestra. Pubis and ischium extend ventrally and medially to the acetabulum toward the plastron midline where they almost contact each other. The pubis and the ischium are attached by suture with the xiphiplastron. The left and right ilium preserved only their acetabular portions, but the left one also lacks its dorsal end, while the dorsal end of the right ilium is attached by suture to the eight costal bone (figure 2.2).
Femur (figure 3.1). The right and left femora are preserved in good condition. The femur is slightly longer than the humerus. The left femur is exposed in posterior view, and only the trochanter major and the partial articular head are seen. The right femur is preserved in postero-anterior view, showing most of the oval articular head surface that is set off from the dorsal surface of the femur at an obtuse angle of approximately 120 degree with the longitudinal axis of the femur. The femoral shaft is subcilindric in cross section and arched dorsally as in other chelids. The condyles that bear the fibular articulation are visible in the distal end of both femora.
Tibia (figure 3.1). Only the right tibia is preserved. This bone is only slightly displaced from its natural position. The tibia is a massive bone expanded in both ends and the shaft is slightly bent. Unfortunately, the proximal and posterior ends of the right tibia are damaged and, consequently, they are not available for an accurate description.
Fibula (figure 3.1). The left and right fibula are preserved. The left fibula is partially articulated with the fibular condyle, while the right one is slightly displaced from the articular surface of the right femur, and is exposed in lateral view. As in other chelids the fibula is a slender bone, more slender than the tibia. The proximal articular surface of the right fibula is slightly rounded, while the visible distal end of the left tibia is expanded.
Discussion
Taxonomic implications
The shell and vertebrae morphology of Linderochelys rinconensis gen. et sp. nov. is comparable with that of chelid pleurodiran turtles. The pelvic girdle sutured with the shell is a recognized synapomorphy of Pleurodira (see Gaffney and Meylan, 1988, and references therein). This condition associated with the presence of a short and wide mesoplastra crossed by a humeropectoral sulci, and the presence of opisthocoelous or biconcave cervical vertebrae, and loose carapace-plastron contact (see Lapparent de Broin and de la Fuente , 2001) allow us to assign this species to the family Chelidae.
Although isolated cervical vertebrae with similar primitive characters of the Australasian extant species of the Emydura-Elseya group have been recognized in the Upper Cretaceous horizon of Patagonia (Broin and de la Fuente, 1993b: pl. 1, figs. 2-3), until now all the named chelid taxa recovered from Upper Cretaceous horizons of Patagonia have been considered closely related to the South American short and long-necked chelids (Lapparent de Broin and de la Fuente, 2001; de la Fuente et al., 2001). In this sense, Linderochelys rinconensis gen. nov. et sp. is also tentatively referred to the Phrynops group (sensu Broin and de la Fuente, 1993b) by the morphology of the preserved cervical vertebrae. These cervical vertebrae are similar to those of the extant species of the Phrynops group. These vertebrae are slightly elongated and lower than the cervical vertebrae of the Australasian short necked species, and are characterized by the presence of ventral keels curved and reduced posteriorly in relation to the central length. Linderochelys rinconensis gen nov et sp. is a short-necked chelid characterized by an elongated abdominal seam. It differs clearly from the South American extant species assigned to Phrynops (sensu lato) in the presence of cuneiform mesoplastra crossed by an pectoral-abdominal sulcus. Likewise, this new taxa also differs from the hitherto known Upper Cretaceous chelids referred to the Phrynops group by several characters (Lapparent de Broin and de la Fuente, 2001). Differs from Palaeophrynops patagonicus from the Los Alamitos Formation (Late Campanian-Early Maastrichtian) in the bell shape of entoplastron, and the more depressed common process of postzygapophyses. Differs from the Bajo Barreal Formation (Turonian-Campanian) turtle Bonapartemys bajobarrealis in having a short plastral bridge, absence of ridges at the bridge limit of anterior plastral lobe, and in the different plastral scale formulae, and also differs from the Portezuelo Formation (Late Turonian-Early Coniacian) turtle Prochelidella portezuelae and the Cenomanian Prochelidella spp. (Lapparent de Broin et al., 1997) in having a large and rounded anterior plastral lobe and in the length of interadominal seam.
Comments about the relationships among extant species of Phrynops (sensu lato) and the constraints in the inclusion of the extinct species in a phylogenetic scenario
Broin and de la Fuente (1993b) suggested to use the Phrynops Group (= Phrynops sensu lato) to recognize a South American group of short-necked chelid turtles. Among the extant forms, this group includes: a) Phrynops s.s. with three extant subgenera without a good definition (Phrynops Wagler, Mesoclemmys Gray and Batrachemys Stejenger) follows sensu Bour (1973) and Bour and Pauler (1987); b) Acanthochelys, excluded from Platemys auctoris; and c) Platemys Wagler restricted to P. platycephala (Schneider). Although Broin and de la Fuente (1993b) suggested that the genus Phrynops (s.s.) could be a paraphyletic tax-on in relation to the other two genera, these authors proposed that Acanthochelys and Platemys might be sister groups and could be close relative to Phrynops (s.s). According to Broin and de la Fuente (1993b), Phrynops (s.l.) is characterized by two synapomorphies: the dorsal inflation of the antrum squamosum and the inclination of the columella-fenestra ovalis axis. The species included in Phrynops (s.l.) are characterized by a short neck as the members of the Emydura-Elseya group, but with cervical vertebrae slightly more elongated and lower, and with ventral keels curved and reduced posteriorly in relation to the central length, relative to the cervical vertebrae of Australasian short necked species. The cotyle fragment preserved in Linderochelys rinconensis gen et sp. nov. is similar to that of some species of Phrynops (s.s.)
The Broin and de la Fuente´s concept of Phrynops sensu stricto is only comparable to Phrynops sensu lato of McCord et al. (2001), the last reviewers of Phrynops. Previously to McCord's work, several authors explored the loose relationships among the extant species assigned to the genus Phrynops (s.l.). Gaffney (1977) was unable to found derived cranial characters among some species of Phrynops available in his study [Phyrnops (P.) geoffroanus (Schweigger), P (M.) gibbus (Schweigger) and P (B.) nasutus (Schweigger) complex]. The monophyly of the genus Phrynops (s.l.) seemed to be supported by genetic data of Frair (1980, 1982) and Reed et al. (1991). Likewise, Shaffer et al. (1997) provided a phylogeny where the genus Phrynops (s.l.) resulted as a monophyletic taxon. This conclusion was achieved using mitochondrial DNA sequencing, morphological and paleontological data. Contrary to this proposal, Seddon et al. (1977) strongly supported the paraphyly among the subgenera of Phrynops (s.l.). This conclusion was sustained after Seddon et al. (1977) sequenced 411 mitochondrial 12S rRNA nucleotides of 16 species assigned to 11 chelid genera. Analysis using parsimony and neighbor-joining algorithms supported the generic differentiation among the subgenera of Phrynops (s.l.). In this context Mesoclemmys was proposed as the sister taxon of Batrachemys, and Phrynops was closely related to the genus Chelus. Likewise, the results of Georges et al. (1998) studies on molecular data supported the conclusion of Seddon et al. (1997) about the paraphyly of the genus Phrynops (s.l.) and resolved to elevate once again the subgenera (Phrynops, Mesoclemmys and Batrachemys) to generic level. Finally, McCord et al. (2001) demonstrated full recognition of these taxa as genera based on morphometric analysis, skull osteology, and nuclear and mitochondrial gene secuences. These three distinguished taxa were anteriorly demoted to subgenera, and then synonymized with Phrynops (see Pritchard, 1979; Pritchard and Trebbau, 1984).
Contrary to the almost completeness condition of the long necked chelid species of the genus Yaminuechelys de la Fuente et al., which allows us to include these taxa in a phylogenetic context (de la Fuente et al., 2001; Bona, 2004; Bona and de la Fuente, 2005), the incompleteness nature of most of the short-necked chelid species from the Cretaceous of Patagonia prevents their phylogenetic resolution. Among them the species assigned to Phrynops sensu lato by Lapparent de Broin and de la Fuente (2001), and the new taxon here described, are not easy to include in McCord (2001) phylogenetic scenario. In this sense, 50 per cent of the morphological character (9 on 18) scored by these authors are no comparable in Patagonian fossil turtles and the remainder characters are not useful when the analyses is broadened to include the Patagonian Cretaceous taxa.
Acknowledgements
J. Gonzalez drew the figures 2 and 3 of this article. C. Marsicano and an anonymous reviewer provided constructive comments. The fieldtrips were founded by projects of the National Agency of Science and Technology of Argentina (PICT / project 07-01513 and 08277), the Municipality of Rincon de los Sauces, National University of Comahue and Chevron - Texaco (T/013-021), and Pecom S.A. This paper was also partially supported by PIP -CONICET 02262/00.
References
1. Bona, P. 2004. [Sistemática y biogeografía de las tortugas y de los cocodrilos paleocenos de la Formación Salamanca, Provincia de Chubut, Argentina. Ph Dissertation Thesis. Universidad Nacional de la Plata, La Plata, Buenos Aires, 185 pp. Unpublished.]. [ Links ]
2. Bona, P. and de la Fuente, M.S. 2005. Phylogenetic and palaeobiogeographical implications of Yaminuechelys maior (Staesche, 1929) new comb., a large long-necked chelid turtle from the early Paleocene of Patagonia. Journal of Vertebrate Paleontology 25: 569-582. [ Links ]
3. Bonaparte, J.F. 1986. History of the terrestrial Cretaceous vertebrates of Gondwana. 4º Congreso Argentino de Paleontología y Bioestratigrafía (Mendoza), Actas 2: 63-95. [ Links ]
4. Bonaparte, J.F., 1987. The Late Cretaceous fauna of Los Alamitos, Patagonia, Argentina. Revista del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia". Paleontología 3: 103-178. [ Links ]
5. Bonaparte, J.F., 1996. Cretaceous Tetrapods of Argentina. Münchner Geowissenschaftliche Abhandlungen. Reihe A. Geologie und Paläontologie 30: 73-130. [ Links ]
6. Bonaparte, J.F., Franchi, M.R, Powell, J.E. and Sepúlveda, E.G. 1984. La Formación Los Alamitos (Campaniano-Maastrichtiano) del sudeste de Río Negro, con descripción de Kritosaurus australis n. sp. (Hadrosauridae). Significado paleogeográfico de los vertebrados. Revista de la Asociación Geológica Argentina 39: 284-299. [ Links ]
7. Bour, R. 1973. Contribution á la connaissance de Phrynpos nasutus (Schweigger: 1812) et Phrynops tuberculatus (Luederwaldt: 1926). Description dé una nouvelle sousespece originaire de Paraguay, Phrynops tuberculatus vanderhaegei (Testudinata-Pleurodira-Chelidae). Bullettin de la Société Zoologique de France 98: 175-190. [ Links ]
8. Bour, R. and Pauler, I. 1987. Identité de Phrynops vanderhaegei Bour, 1973, et des especes affines (Reptilia-Chelonii-Chelidae). Mesogée 47: 3-23. [ Links ]
9. Broin, F. de. 1987. The Late Cretaceous Fauna of Los Alamitos, Patagonia, Argentina. Part IV, Chelonia. Revista Museo Argentino de Ciencias Naturales, "Bernardino Rivadavia" Paleontología 3: 131-139. [ Links ]
10. Broin, F. de.1988. Les tortues et le Gondwana. Examen des rapports entre le fractionnement du Gondwana au Crétacé et la dispersion géographique des tortues pleurodires à partir du Crétacé. Studia Geológica Salmanticensia. Studia Palaeocheloniologica 2: 103-142. [ Links ]
11. Broin, F. de and de la Fuente, M.S. 1993a. Les tortues fossiles d'Argentine: premièr synthèse. Table Ronde Européenne Paléontologie et Stratigraphie d'Amerique latine. (Lyon, 1992), Documents des laboratoires de Géologie Lyon 125: 73-84. [ Links ]
12. Broin, F. de. and de la Fuente, M.S. 1993 b. Les tortues fossiles d'Argentine: Synthèse. Annales de Paléontologie 79: 169-232. [ Links ]
13. Calvo, J.O., Coria, R.A. and Salgado, L. 1997. Uno de los más completos titanosáuridos (Dinosauria-Sauropoda) registrados en el mundo. Ameghiniana 34: 534. [ Links ]
14. Calvo, J.O. and González Riga, B. J. 1999. Hallazgos de Theropoda y Titanosauridae, y su paleoambiente en una nueva localidad de Rincón de los Sauces, Neuquén, Argentina. Ameghiniana 36: 97. [ Links ]
15. Calvo, J.O. and González Riga, B.J. 2003. Rinconsaurus caudamirus gen. et sp. nov., a new titanosaurid (Dinosauria, Sauropoda) from the Late Cretaceous of Patagonia, Argentina. Revista Geológica de Chile 30: 333-353. [ Links ]
16. De la Fuente, M.S. 1988. [Las tortugas Chelidae (Pleurodira) y Testudinidae (Cryptodira) del Cenozoico argentino. Ph. Dissertation Thesis. Universidad Nacional de La Plata, 270 pp., Unpublished.]. [ Links ]
17. De la Fuente, M.S. 1992. Las tortugas Chelidae del Terciario superior y Cuaternario del territorio argentino. Ameghiniana 29: 211-299 [ Links ]
18. De la Fuente, M.S. 1997. Las tortugas Chelidae del Cretácico de Patagonia. Aspectos paleobiogeográficos. 3º Congreso Argentino de Herpetología (Corrientes), Resúmenes: p. 22 [ Links ]
19. De la Fuente, M.S. 2003. Two new pleurodiran turtles from the Portezuelo Formation (Upper Cretaceous) of northern Patagonia, Argentina. Journal of Paleontology 77: 559-575. [ Links ]
20. De la Fuente, M.S., Lapparent de Broin, F. de and Manera de Bianco, T. 2000. El más antigüo y completo esqueleto de un quélido de cuello largo del subgrupo Hydromedusa (Chelidae, Pleurodira) del Cretácico Supérior de Patagonia. Reunion Anual de Comunicaciones de la Asociación Paleontológica Argentina (Mar del Plata). Ameghiniana Suplemento Resúmenes 37: 72R. [ Links ]
21. De la Fuente, M.S., Lapparent de Broin, F. de and Manera de Bianco, T. 2001. The oldest and first nearly complete skeleton of a chelid, of the Hydromedusa subgroup (Chelidae, Pleurodira), from the Upper Cretaceous of Patagonia. Bullettin de la Société Géologique de France 172: 237-244. [ Links ]
22. Frair, W. 1980. Serological survey of Pleurodiran turtle. Comparative Biochemistry Physiology 65B: 505-511. [ Links ]
23. Frair, W. 1982. Serological studies of the red turtle, Phrynops rufipes. Herpetological Bulletin 17: 4-9. [ Links ]
24. Gasparini, Z and de la Fuente, M.S. 2000. Tortugas y plesiosaurios de la Formación La Colonia (Cretácico Superior) de la Patagonia. Revista Española de Paleontología 15: 23-35 [ Links ]
25. Gasparini, Z., de la Fuente, M.S., Fernández, M.S. and Bona, P. 2001. Reptiles from coastal environments of the Upper Cretaceous of northern Patagonia. 7º International Symposium on Mesozoic Terrestrial Ecosystems (Buenos Aires, 1999). Asociación Paleontológica Argentina Publicación Especial 7: 101-105. [ Links ]
26. Gasparini, Z., Casadío, S., de la Fuente, M.S, Salgado, L., Fernández, M. and Concheyro, A. 2002. Reptiles acuáticos en sedimentitas lacustres y marinas del Cretácico superior de Patagonia (Río Negro, Argentina) 15º Congreso Geológico Argentino (El Calafate). Actas 1: 473-478. [ Links ]
27. Gaffney, E.S. 1977. The side-necked turtle family Chelidae: A theory of relationships using shared derived characters. American Museum Novitates 2620: 1-28. [ Links ]
28. Gaffney, E.S. 1979. Fossil chelid turtles of Australia. American Museum Novitates 2681: 1-23. [ Links ]
29. Gaffney, E.S. and Meylan, P.A. 1988. A phylogeny of Turtles. En: M. J. Benton (ed.), The Phylogeny and Classification of the Tetrapods 1: Amphibians, Reptiles, Birds. Systematic Association, Special Volume, Clarendon Press, Oxford, 35A: pp. 157-219. [ Links ]
30. Georges, A., Birrell, J., Saint, K.M., McCord, W. and Donnellan, S.C. 1998. A phylogeny for side-necked turtles (Chelonia: Pleurodira) based on mitochondrial and nuclear gene sequence variation. Biological Journal of the Linnean Society 67: 213-246. [ Links ]
31. Goin, F.J., Poire, D., de la Fuente, M.S, Cione, A.L; Novas, F.E., Bellosi, E.S., Ambrosio, A., Ferrer, O., Canessa, N.D., Carloni, A., Ferigolo, J., Ribeiro, A.M., Sales Viana, Reguero, M., Vucetich, M.F, Marenssi, S., de Lima Filho, M.F. and Agostinho, S. 2002. Paleontología y Geología de los sedimentos del Cretácico Supérior aflorantes al sur del río Shehuen (Mata Amarilla, provincia de Santa Crruz, Argentina). 15º Congreso Geológico Argentino (El Calafate), Actas 1: 603-608. [ Links ]
32. González Riga, B.J. and Calvo, J.O. 1999. Unusual caudal series of Titanosauridae of the Late Cretaceous in the Rio Colorado Formation, border between the Neuquen and Mendoza provinces, Argentina. 7º International Symposium on Mesozoic Terrestrial Ecosystems (Buenos Aires), Abstracts: 29-30. [ Links ]
33. González Riga, B.J., Calvo, J.O., Astini, R.A. and Parras, A.M. 2003. Preservación tafonómica de titanosaurios (Dinosauria, Sauropoda) en facies fluviales del sector septentrional de la Cuenca Neuquina. Reunión Anual de Comunicaciones de la Asociación Paleontológica Argentina y Simposio de Tafonomía y Paleoecología (Santa Rosa). Ameghiniana Suplemento Resúmenes 40: 102R. [ Links ]
34. Iverson, J.B. 1992. A Revised Checklist with Distribution Maps of the Turtles of the World. Privately Print, Richmond (Indiana), 363 pp. [ Links ]
35. Lapparent de Broin, F. de and de la Fuente, M.S. 1999. Particularidades de la fauna continental de tortugas del Cretácico de Argentina. 14º Jornadas Argentinas de Paleontología de Vertebrados (Neuquén, 1998), Ameghiniana 36: 104. [ Links ]
36. Lapparent de Broin, F. de and de la Fuente, M.S. 2001. Oldest world Chelidae (Chelonii, Pleurodira), from the Cretaceous of Patagonia. Comptes Rendues de l´Académie des Sciences de Paris 333: 463-470. [ Links ]
37. Lapparent de Broin F. de and Molnar, R. 2001. Eocene chelid turtles from Redbank Plains, southeastern Queensland, Australia. Geodiversitas 23: 41-79. [ Links ]
38. Lapparent de Broin, F. de, de la Fuente, M.S. and Calvo, J. 1997. Presencia de los más antiguos Quélidos (Tortugas Pleurodiras) en el Cretácico Inferior de 'El Chocón', Provincia del Neuquén, Argentina. 13º Jornadas Argentinas de Paleontología de Vertebrados (La Rioja), Ameghiniana 34: 538. [ Links ]
39. Leanza, H.A. 1999. The Jurassic and Cretaceous terrestrial beds from Southern Neuquén Basin, Argentina. Field Guide. Instituto Superior de Correlación Geológica. INSUGEO. Serie Miscelánea 4: 1-30. [ Links ]
40. Leanza, H.A. and Hugo, C.A. 2001. Cretaceous red beds from southern Neuquén Basin (Argentina): age, distribution and stratigraphic discontinuities. Asociación Paleontológica Argentina, Publicación Especial 7: 117-122. [ Links ]
41. Leanza, H.A., Apesteguia, S., Novas, F.E. and de la Fuente, M.S. 2004. Cretaceous terrestrial beds from Neuquén Basin (Argentina) and their tetrapods assemblages. Cretaceous Research 25: 61-87. [ Links ]
42. McCord, W., Joseph-Ouni, M. and Lamar, W.W. 2001. A taxonomic reevaluation of Phrynops (Testudines: Chelidae) with the description of two new genera and a new species of Batrachemys. Revista de Biologia Tropical 49: 715-764. [ Links ]
43. Pritchard, P.C.H. 1979. Encyclopedia of Turtles. T: F: H: Publications Inc., Neptune, New Jersey, 895 pp. [ Links ]
44. Pritchard, P.C.H. and Trebbau, P. 1984. The Turtles of Venezuela. Contribution to Herpetology, Society Study Amphibians, Reptiles Publications 2: 1-403. [ Links ]
45. Reed, K.M, Hanks, B.G., Bickham, J.W, Rhodin, A.G. J., Greenbaum, F., Mittermeier, R.A. and Fedullo, L.P. 1991. Cytogenetic analysis of the pleurodire turtle Phrynops hogei and its taxonomic implications. Amphibia-Reptilia 12: 203-212. [ Links ]
46. Shaffer, H.B., Meylan, P. and Mcknight, M.L. 1997. Test of turtle phylogeny: molecular, morphological, and paleontological approaches. Systematic Biology 46: 235-268. [ Links ]
47. Seddon, J.M., Georges, A., Bavesrstock, P. and McCord, W. 1997. Phylogenetic relationships of chelid turtles (Pleurodira: Chelidae) based on mitochondrial 12S rRNA gene sequence variation. Molecular Phylogenetics and Evolution 7: 55-61. [ Links ]
48. Turner, A.H. and Calvo, J.O. 2005. A new sebecosuchian crocodyliform from the Late Cretaceous of Patagonia. Journal of Vertebrate Paleontology 25: 87-98 [ Links ]
49. Williams, E.E. 1950. Variation and selection in the cervical central articulations of living turtles. Bulletin of the American Museum of Natural History 94: 511-561. [ Links ]
50. Warren, J.W. 1969. Chelid turtles from the Mid-Tertiary of Tasmania. Journal of Paleontology 43: 179-182. [ Links ]

