










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.44 n.2 Buenos Aires abr./jun. 2007
The first shelly fauna from metamorphic rocks of the Sierras Pampeanas (La Cébila Formation, Sierra de Ambato, Argentina): age and paleogeographic implications
Sebastián O. Verdecchia1, Edgardo G. Baldo2, Juan L. Benedetto3 and Pablo A. Borghi1
1Departamento de Geología Básica, Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba. Av. Vélez Sarsfield 1611, 5016 Córdoba. sverdecchia@efn.uncor.edu, pabloborghi@yahoo.com.ar
2CONICET. Departamento de Geología Básica, Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba. Av. Vélez Sarsfield 1611, 5016 Córdoba. ebaldo@efn.uncor.edu
3CONICET. Centro de Investigaciones Paleontológicas (CIPAL), Facultad de Ciencias Exactas Físicas y Naturales, Universidad Nacional de Córdoba, Av. Vélez Sarsfield 299, 5000 Córdoba. jbenedetto@arnet.com.ar
Introduction
In Argentina, Cambro-Ordovician shelly faunas are confined to the Precordillera, Famatina and Northwestern basins within the present-day Andean belt. Outside the Andes, fossiliferous rocks are known only from the Tandilia System, in the Buenos Aires province. These rocks, however, though rich in Ordovician trace fossils, are barren in body fossils. The Sierras Pampeanas of central Argentina are a series of mountainous ridges bounded by reverse faults, uplifted during the Tertiary Andean Orogeny in response to the flat-subduction of the Nazca plate at this latitude (Jordan et al., 1983). This basement complex is largely composed of low- to high-grade polymetamorphic rocks and extensive granitoid plutons whose Mesoproterozoic to early Paleozoic ages were based essentially on U-Pb isotopic data (Pankhurst and Rapela, 1998 and references therein). Here we provide the first report of a shelly fauna from metamorphic rocks of the Sierras Pampeanas, consisting of numerous, relatively well-preserved rhynchonelliformean brachiopod valves, and a few trilobite and bivalve remains. This fauna allows to better constrain the age of the La Cébila Formation and sheds new light on the lower Paleozoic paleogeography and depositional history of the Gondwana margin. Moreover, this discovery opens new and exciting perspectives for searching fossils in the metamorphic rocks formerly considered of Neoproterozoic-Cambrian age exposed in large areas between 27° S and 33° S, through Tucumán, Catamarca, La Rioja, Córdoba and San Luis provinces.
Specimens illustrated in this paper are housed in the paleontological collection of Centro de Investigaciones Paleobiológicas (CIPAL), Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba, Argentina (prefix CEGH-UNC).
Geological setting
The metasedimentary rocks of the La Cébila Formation crop out along the Quebrada La Cébila, a north-south erosive valley separating the Sierra de Velasco from the Sierra de Ambato, near the limit between La Rioja and Catamarca provinces (figure 1). These metasediments were firstly described by Sgrosso (1949), and subsequently they were referred to the La Cébila Formation by González Bonorino (1951). Espisúa and Caminos (1979) recognized in this formation four main lithologic units: phyllites, schists with andalucite, metaquartzites, and micaschists ("micacitas inyectadas"). Rocks are arranged according to a metamorphic zonation prograding from southeast to northwest, which starts with the biotite zone (in the phyllites), continues with the andalucite zone (in the micaschists and metaquartzites), and culminates with the sillimanite - potassium feldspar zone (in the metaquartzites and cordierite migmatite) (figure 1.d). The low pressure metamorphic event is linked to the intrusion of the Velasco granites.
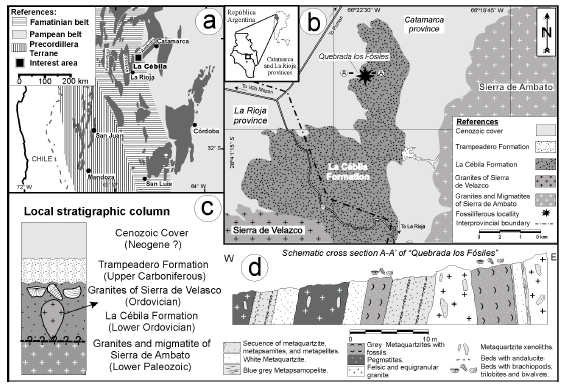
Figure 1. a, Geographic location of the Sierras Pampeanas and the La Cébila area / ubicación geográfica de las Sierra Pampeanas y del área de la Cébila; b, Geological sketch showing the La Cébila Formation outcrops and fossiliferous locality / mapa geológico esquemático donde se muestran los afloramientos de la Formación La Cébila; c, Local stratigraphic column / columna estratigráfica local; d, schematic cross-section of Quebrada Los Fósiles / sección transversal esquemática de la Quebrada Los Fósiles.
The fossils described here were collected from outcrops of the La Cébila Formation located about 6 km north of quebrada La Cébila, in the Catamarca province (figure 1). In this area this unit consists of a succession of white to grey fossiliferous metaquartzites interbedded with metapelites displaying strain deformation (figure 1). These rocks are intruded by tourmalinerich pegmatite and felsic equigranular granite. Metaquartzites are finegrained (0.3 mm in average) and show a subpolygonal granoblastic texture composed by an equigranular setting of quartz, plagioclase, clino-amphibole (pale green), apatite, epidote, titanite, chlorite, zircon, and ore minerals (magnetite and others unidentified oxides). The brachiopod shells are replaced entirely by clino-amphibole and oxides. The metamorphic clino-amphibole in the metaquartzites and the andalucite + fibrolite + biotite assemblage in the metapelites suggest that the metamorphic event reached the hornfels-hornblende facies (~ 600º - 650º C).
Preservation and age
Fossils come from numerous, more or less discrete, densely packed shell concentrations as much as 5 cm thick (figure 2.A), which are irregularly distributed within the metaquartzite beds. All brachiopod shells are disarticulated but dorsal and ventral valves are represented in nearly equal proportion. Trilobites consist of an isolated pygidium (figure 2.J) and a few thoracic segments. Degree of fragmentation and sorting appears to be low. Almost all brachiopod valves are preserved in the convexup position. Such preservational features are suggestive of event concentrations (Kidwell, 1991) in which fossils have been reworked and displaced during high-energy, short-term events, but deposited not too far from their original habitat. Shell beds of this kind are compatible with a nearshore zone above the fair weather wave base, probably a shoreface environment. This is consistent with the fine-grained, well-sorted, quartzrich sandstones enclosing coquinas.

Figure 2. A-I, Ffynnonia sp. A. Slab showing brachiopod shell concentration / laja con una concentración de valvas de braquiópodos, CEGHUNC 22041 (x 0,75); B, Exterior of ventral valve, latex cast / valva ventral, molde de caucho, CEGH-UNC 22397 (x2); C, Exterior of ventral valve, latex cast / valva ventral, molde de caucho, CEGH-UNC 22399 (x2.5); D, Exterior of dorsal valve, latex cast / valva dorsal, molde de caucho, CEGH-UNC 22398 (x2); E, Exterior of ventral valve, latex cast / valva ventral, molde de caucho, CEGH-UNC 22400 (x2); F, Internal mold of ventral valve / molde interno de valva ventral, CEGH-UNC 22401 (x3); G, Exterior of dorsal valve, latex cast / valva dorsal, molde de caucho, CEGH-UNC 22402 (x2); H. Internal mold of dorsal valve / molde interno de valva dorsal, CEGH-UNC 22402 (x2); I, Exterior of dorsal valve / exterior de valva dorsal, CEGH-UNC 22403. J, Pygidium of an undetermined asaphid trilobite / pigidio de un trilobite asáfido indeterminado, CEGH-UNC 22409 (x2).
Despite the metamorphism affecting fossiliferous beds, both shell outline and valve convexity were not substantially modified. Brachiopods are preserved as internal and external molds. In many specimens, details of external surface and internal structures are obscured by overgrowth of clino-amphibole crystals.
The sedimentary protoliths of the La Cébila Formation were considered to be Precambrian in age by Sgrosso (1949) and González Bonorino (1951). Later, Espisúa and Caminos (1979) correlated this metamorphic unit with the Cambrian Mesón Group. Aceñolaza and Aceñolaza (2003, p. 4) mentioned the presence of the Beltanelliformis ichnoassociation in 'the southwards La Cébila-Angulos areas in La Rioja province' and referred these metamorphic rocks to the Upper Proterozoic-Lower Cambrian Punco-viscana Formation of northwestern Argentina. More recently, Finney et al. (2003) showed that detrital zircon ages from the metasedimentary rocks of the La Cébila Formation fall into three populations, the youngest one forming a peak at 480-520 Ma indicating a Lower Ordovician maximum age (around the Tremadoc-Arenig boundary).
The La Cébila shell beds consist of nearly monospecific assemblages of Ffynnonia sp. (figure 2.A). The type material of this genus [F. costata (Bates)] comes from the Treiorwerth Formation of Anglesey, northwest Wales, of late Arenig age (Neuman and Bates, 1978; Neuman, 1984). Outside Wales, Ffynnonia is known only from the Precordillera and Famatina basins of Argentina. The Precordilleran San Juan Formation has yielded three species, of which F. spondyliformis Benedetto comes from the upper Arenig Monorthis cumillangoensis Zone, and F. minuta (Benedetto and Herrera) and F. fasciculata (Benedetto and Herrera) occur in the uppermost Arenig-lower Llanvirn Ahtiella argentina Zone (Benedetto and Herrera, 1987; Benedetto, 2001). In the Famatina Group succession, Ffynnonia famatinensis Benedetto is very common in the Suri Formation (Benedetto, 2003) of well constrained mid-Arenig age (O. evae conodont Zone; Albanesi and Astini, 2000). This is consistent with the 470± 2 Ma U-Pb SHRIMP age of zircons from the Las Planchadas rhyolite exposed at Chaschuil River (Baldo et al., 2003), and the 477 ± 4 Ma rhyolite from Quebrada Potrero Grande (Dahlquist et al., 2005), in the northern Famatina belt. According to the last published ICS International Geologic Time Scale (Ogg, 2004) this volcanic event is of mid-late Arenig age. Cisterna (2001) stated that these rhyolites are partially equivalent to the marine volcanosedimentary unit referred to the 'Suri' Formation by Mángano and Buatois (1996). It should be noted, however, that the rich paleontological content of the Chaschuil beds indicates correlation with the Molles Formation rather than the Suri Formation (Benedetto, 2003).
Although the La Cébila material can be confidently referred to Ffynnonia, new and better preserved specimens are needed for specific identification. However, judging from the known stratigraphic range of the genus, and considering the morphological similitude with the Famatina species, the age of fossiliferous levels is probably Arenig.
Paleogeographic implications
The monospecific assemblage from the La Cébila Formation is inadequate for paleobiogeographical comparisons. However, the record of Ffynnonia is significant as this genus is always associated with key taxa of the Celtic Realm. Ffynnonia has been recorded in the classical Celtic locality of Anglesey (Wales) and in the Famatina Range, whose brachiopods form a statistically significant cluster with other Celtic localities (i.e. the Central Volcanic belt of New-foundland; Neuman, 1984). In the Precordillera basin, species of Ffynnonia are also associated with distinctive Celtic Realm forms [i.e. Inversella (Reinversella), Rugostrophia, Skenidioides]. The Celtic realm is a paleobiogeographic unit developed mainly at mid- to high-latitudes, along the northwest Gondwana continental margins and around Iapetus Ocean-related terranes as Avalonia, Famatina, and Precordillera (Neuman and Harper, 1992; Benedetto, 1998, 2003). The presence of Ffynnonia in the La Cébila Formation suggests a faunal connection between the Sierras Pampeanas seaway and the Famatina basin located to the west.
The shallow-water marine succession of the La Cébila Formation would record deposition near the outermost Gondwana foreland. This unit developed probably along an incipiently eroded mountain belt uplifted during the early to mid Cambrian Pampean compressive deformation (Rapela et al., 1998). Between this eastern (in present coordinates) shore-line and the rapidly subsiding Famatina backarc basin (Astini and Benedetto, 1996) located to the west, an over 150 km wide marine platform system must have developed. Astini et al. (2003), based on detrital zircon data (c. 480 Ma), proposed a tentative correlation between the La Cébila Formation and the low-grade metamorphic La Aguadita Formation exposed along the eastern slope of the Famatina Range. These authors interpreted this immature sandy unit as a relict of a clastic wedge developed in a back-arc setting during the rapid exhumation of the Famatinian granitoids, so that it should be younger than the volcanosedimentary Suri and Molles formations. However, the discovery of Ffynnonia -a typical brachiopod of the Suri Formation -in the La Cébila Formation favours the correlation of these units. The profound eastward flooding of the Gondwana plate may have been associated with a major global sealevel rise, probably the 'Evae Drowning Event' (Nielsen, 2004). In the Precordillera carbonate ramp, a high stand sealevel is well documented in the mid-Arenig O. evae conodont Zone (Cañas, 1999; Carrera et al., 1999), and in the Atacama region of Chile a mid-Arenig transgressive deepening upwards marine succession bearing Monorthis (a genus usually associated with Ffynnonia) overlaps unconformably the Cordón de Lila sedimentary-igneous Complex (Benedetto et al., 2005). The upper part of the Acoite Formation in the Cordillera Oriental of northwestern Argentina, of mid Arenig age, has also been interpreted as deposited during a highstand system tract (Astini et al., 2004).
Certainly, available isotopic and paleontologic data from the metamorphic units exposed to the east of the Famatina Range are by far too fragmentary for accurate paleogeographic and paleotectonic interpretations. The main challenge in the near future will be to decipher sedimentary facies and ages of metamorphic marine strata deposited in the vast epicontinental seaway extending between the Famatina-Puna magmatic arc and the Pampean foreland.
Systematic paleontology (J.L.B)
Order Orthida Schuchert and Cooper, 1932
Suborder Orthidina Schuchert and Cooper, 1932
Superfamily Plectorthoidea Schuchert and LeVene, 1929
Family Platystrophiidae Schuchert and LeVene, 1929
Genus Ffynnonia Neuman and Bates, 1978
Type species. Pleurorthis costatus Bates, 1968.
Ffynnonia sp. Figures 2.A-I
Material. Three internal molds of ventral valves and two of dorsal valves, CEGH-UNC 22397-22408. Four slabs bearing coquinas with numerous molds of fragmentary ventral and dorsal valves.
Occurrence. La Cébila Formation, about 6 km north of quebrada La Cébila (Quebrada Los Fósiles), Catamarca province.
Description. Subrectangular shell, up to 35 mm wide, semielliptical in outline, about 60% as long as wide, with right-angled to acute cardinal extremities. Hingeline long, representing the greatest shell width. Ventral valve gently convex, with a wide, well-defined median sulcus and evenly convex lateral slopes. Interarea planar, moderately low, apsaclie. Dorsal valve moderately and evenly convex, with an incipient narrow median sulcus confined to the posterior half of the valve, being replaced anteriorly by a weak fold. Ornament coarsely fascicostellate formed by subangular costellae arising by intercalation and bifurcation, in number of 4-5 per 5 mm, reaching about 25 at the anterior margin in one nearly complete ventral valve (figure 2.C). A few coarser growth interruptions developed near anterior margin in one specimen.
Ventral interior with large, triangular teeth supported by receding divergent dental plates. Well developed pseudospondylium supporting a large, deeply impressed, subrhomboidal ventral muscle field, bounded laterally by ridge-like extensions of dental plates and anteriorly by an arcuate ridge, extending anteriorly for 45% of valve length. Diductor muscle scars long, subrectangular, narrower than the median adductor field. Dorsal interiors poorly preserved, showing vestiges of a subtriangular notothyrial platform and laterally divergent brachiophores.
Remarks. Assignment of the La Cébila material to Ffynnonia is based mainly upon the fascicostellate ornament, sulcate ventral valve, and ventral muscle field located on a well developed spondylium. In shape, ornament and ventral muscle field morphology, the La Cébila material is rather similar to the type species Ffynnonia costata (Bates), from the Arenig of Wales (Bates, 1968; Neuman and Bates, 1978), from which differs in its larger size, the less numerous costellae and the proportionally larger ventral muscle field. Ffynnonia famatinensis Benedetto (2003), from the Suri Formation of Famatina Range, also resembles the La Cébila specimens but can be differentiated by its finer ornament and its shallower ventral sulcus. Externally, our material is closer to the Precordilleran species F. spondyliformis Benedetto (2001), but the latter differs in having a shorter and very prominent spoon-shaped pseudospondylium. F. minuta (Benedetto and Herrera), from the uppermost San Juan Formation, differs in its more transverse shell outline, its rarely branched costellae and its suboval ventral muscle field, whereas the nearly coeval P. fasciculata (Benedetto and Herrera), formerly referred to Platystrophia by Benedetto and Herrera (1987), differs in having finer fasciculate ornament, strongly biconvex shells and more prominent dorsal fold.
Acknowledgments
We thank V. Ramos (Universidad Nacional de Buenos Aires) and P.R. Racheboeuf (Université de Bratagne Occidentale, Brest) for their useful comments on the manuscript. We also thank CRILAR for providing logistic support. This study was supported by grants from the Consejo Nacional de Investigaciones Científicas y Técnicas, PIP 5599 (to J.L.B), and the Agencia Nacional de Promoción Científica y Tecnológica, PICT 07-10735 (to E.G.B.).
References
1. Aceñolaza, F.G. and Aceñolaza, G. 2003. Trace fossils, microbial mats and sedimentary structures in the Puncoviscana Formation of northwestern Argentina (Neoproterozoic-Lower Cambrian): Their record in a varied spectrum of palaeoenvironmental settings. 3º International Colloquium Vendian-Cambrian of Gondwana, IGCP 478 Project, Abstracts, pp. 4-7. [ Links ]
2. Albanesi, G.L. and Astini, R.A. 2000. New conodont fauna from Suri Formation (Early-middle Ordovician), Famatina System, western Argentina. Ameghiniana 37, Suplemento Resúmenes: 68R. [ Links ]
3. Astini, R.A. and Benedetto, J.L. 1996. Paleoenvironmental features and basin evolution of a complex volcanic arc region in the protoAndean western Gondwana. 3º International Symposium on Andean Geodynamics (St. Malô), pp. 755-75. [ Links ]
4. Astini, R.A., Dávila, F.M., Rapela, C.W., Pankhurst, R.J. and Fanning, C.M. 2003. Ordovician back-arc clastic wedge in the Famatina Ranges: new ages and implications for reconstruction of the protoAndean Gondwana margin. In: G.L Albanesi, M.S. Beresi and S.H. Peralta (eds.), Ordovician from the Andes. INSUGEO, Serie Correlación Geológica 17: 375-380. [ Links ]
5. Astini, R.A., Toro, B.A., Waisfeld, B.G. and Benedetto, J.L. 2004. El Paleozoico inferior y medio de la región de Los Colorados, borde occidental de la Cordillera Oriental (provincia de Jujuy). Revista de la Asociación Geológica Argentina 59: 243-260. [ Links ]
6. Baldo, E.G., Fanning, C.M., Rapela, C.W., Pankhurst, R.J., Casquet, C. and Galindo, C. 2003. U-Pb shrimp dating of rhyolite volcanism in the Famatina belt and K-bentonites in the Precordillera. In: G.L. Albanesi, M.S. Beresi and S.H. Peralta (eds.), Ordovician from the Andes. INSUGEO, Serie Correlación Geológica 17: 185-190. [ Links ]
7. Bates, D.E.B. 1968. The Lower Palaeozoic brachiopod and trilobite faunas from Anglesey. Bulletin of the British Museum (Natural History), Geology 16: 127-199. [ Links ]
8. Benedetto, J.L. 1998. Early Palaeozoic brachiopods and associated shelly faunas from western Gondwana: its bearing on the geodynamic history of the preAndean margin. In: R.J. Pankhurst and C.W. Rapela (eds.), The protoAndean margin of Gondwana. Geological Society, London, Special Publications 142: 57-83. [ Links ]
9. Benedetto, J.L. 2001. Silicified Early Ordovician (Arenig) brachiopods from the San Juan Limestone, Argentine Precordillera. Geologica et Palaeontologica 35: 1-29. [ Links ]
10. Benedetto, J.L. 2003. Early Ordovician (Arenig) brachiopods from volcaniclastic rocks of the Famatina Range, northwest Argentina. Journal of Paleontology 77: 212-242. [ Links ]
11. Benedetto, J.L. and Herrera, Z.A. 1987. El género Platystrophia King (Brachiopoda) en la Formación San Juan de la Precordillera Argentina. Ameghiniana 24: 51-59. [ Links ]
12. Benedetto, J.L., Niemeyer, H., González, J. and Brussa, E.D. 2005. First occurrence of Ordovician brachiopods and graptolites from Cordón de Lila (Salar de Atacama), northern Chile: stratigraphic and paleobiogeographic implications. In: R.J. Pankhurst and G.D. Veiga (eds.), Gondwana 12, Geological and Biological Heritage of Gondwana, Abstracts, Academia Nacional de Ciencias (Córdoba), p. 68. [ Links ]
13. Cañas, F.L. 1999. Facies sequences of Late Cambrian to Early Ordovician carbonates of the Argentine Precordillera: A physical stratigraphic comparison with Laurentian platforms. In:. D. Keppie and V.A. Ramos (eds.), Laurentia-Gondwana connections before Pangea, Geological Society of America, Special Paper 336: 43-62. [ Links ]
14. Carrera, M., Sánchez, T.M. and Benedetto, J.L. 1999. Paleoenvironmental controls on biofacies in the early Ordovician limestones of the Argentine Precordillera. In: P. Kraft and O. Fatka (eds.), Quo vadis Ordovician? Acta Universitatis Carolinae, Geologica 43: 475-478. [ Links ]
15. Cisterna, C. E. 2001. Volcanismo subácueo en el Eopaleozoico del Sistema de Famatina, noroeste de Argentina. Revista de la Asociación Geológica Argentina 56: 16-24. [ Links ]
16. Dahlquist, J.A., Pankhurst, R.J., Rapela, C.W., Galindo, C., Alasino, P., Casquet, C., Fanning, C.M., Saavedra, J., Baldo, E. and González-Casado, J. 2005. New SHRIMP ages in the Sierra de Famatina, NW of Argentina: Implications for the Famatinian Orogen. In: R.J. Pankhurst and G.D. Veiga (eds.), Gondwana 12, Geological and Biological Heritage of Gondwana, Abstracts, Academia Nacional de Ciencias (Córdoba), p. 123. [ Links ]
17. Espisúa, S.M. and Caminos, R. 1979. Las rocas metamórficas de la Formación La Cébila, Sierra de Ambato, Provincias de Catamarca y La Rioja. Boletín de la Academia Nacional de Ciencias (Córdoba), 53: 125-142. [ Links ]
18. Finney, S., Gleason, J., Gehrels, G., Peralta, S.H. and Aceñolaza, G. 2003. Early Gondwanan connection for the Argentine Precordillera terrane. Earth and Planetary Science Letters 205: 349-359. [ Links ]
19. González Bonorino, F. 1951. Una nueva formación precámbrica en el noroeste argentino. Comunicaciones Científicas del Museo de La Plata 5: 4-6. [ Links ]
20. Jordan, T.E. , Isacks, B.I., Allmendinger, R.W., Brewer, J.A., Ramos, V.A. and Ando, C.J. 1983. Andean tectonics related to geometry of subducted Nazca plate. Geological Society of America Bulletin 94: 341-361. [ Links ]
21. Kidwell, S.M. 1991. The stratigraphy of shell concentrations. In: P.A. Allison and D.E.G. Briggs (eds.), Taphonomy: Releasing the Data Locked in the Fossil Record. Plenum Press, New York. pp. 115-208. [ Links ]
22. Mángano, M.G. and Buatois, L.A. 1996. Shallow marine event sedimentation in a volcanic arcrelated setting: The Ordovician Suri Formation, Famatina Range, northwest Argentina. Sedimentary Geology 105: 63-90. [ Links ]
23. Neuman, R.B. 1984. Geology and paleobiology of islands in the Ordovician Iapetus Ocean: review and implications. Geological Society of America Bulletin 95: 188-1201. [ Links ]
24. Neuman, R.B. and Bates, D.E.B. 1978. Reassessment of Arenig and Llanvirn age (Early Ordovician) brachiopods from Anglesey, north-west Wales. Palaeontology 21: 571-613. [ Links ]
25. Neuman, R.B. and Harper, D.A.T. 1992. Paleogeographic significance of Arenig-Llanvirn Toquima Table Head and Celtic brachiopod assemblages. In: B.D. Webby and J. Laurie (eds.), Global perspectives in Ordovician Geology, Balkema, Rotterdam. pp. 241-254. [ Links ]
26. Nielsen. A.T. 2004. Ordovician sea level changes: a Baltoscandian perspective. In: B.D. Webby, F. Paris, M.L. Droser and I.G. Percival (eds.), The Great Ordovician Biodiversification Event, Columbia University Press. pp. 84-93. [ Links ]
27. Ogg, J.G. 2004. Status of divisions of the International Geologic Time Scale. Lethaia 37: 183-199. [ Links ]
28. Rapela, C.W., Pankhurst, R.J., Casquet, C., Baldo, E., Saavedra, J., Galindo, C. and Fanning, C.M. 1998. The Pampean Orogeny of the southern protoAndes: Cambrian continental collision in the Sierras de Córdoba. In: R.J. Pankhurst and C.W. Rapela (eds.), The protoAndean margin of Gondwana. Geological Society, London, Special Publications 142: 181-217. [ Links ]
29. Pankhurst, R.J. and Rapela, C.W. (eds.). 1998. The protoAndean margin of Gondwana. Geological Society, London, Special Publications 142: 1-383 [ Links ]
30. Sgrosso, P. 1949. Nota sobre el yacimiento de Antimonita de la Quebrada de La Cébila, Prov. de La Rioja. Revista del Museo de La Plata (nueva serie), Sección Geología 4: 245. [ Links ]
Recibido: 26 de abril de 2006.
Aceptado: 11 de agosto de 2006.

