










Serviços Personalizados
Journal

Artigo
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAmeghiniana
versão On-line ISSN 1851-8044
Ameghiniana v.44 n.2 Buenos Aires abr./jun. 2007
El género Coleites Plummer (Rotaliina, Foraminiferida) en la Argentina
Andrea Caramés1 y Carolina Náñez2
1CONICET. Departamento de Ciencias Geológicas, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Intendente Güiraldes 2160, Ciudad Universitaria, 1428 Buenos Aires, Argentina. carames@gl.fcen.uba.ar
2CONICET-SEGEMAR. Benjamín Juan Lavaisse 1194, 1107 Buenos Aires, Argentina. cnaniez@fullzero.com.ar
Introducción
Hasta el momento llamó la atención de los investigadores que estudiaron las faunas de foraminíferos paleocenos argentinos, la aparente ausencia de Coleites reticulosus (Plummer), una especie que, aunque siempre infrecuente, se considera guía de la fauna "tipo Midway". La fauna "tipo Midway", caracterizada por lagénidos, polimorfínidos y anomalínidos, fue descripta para ambientes de plataforma media a externa (50-200 m), y aunque tipificada por el Grupo Midway de la Planicie Costera Atlántica, regiones del Caribe y Antillas, posee una amplia distribución global en sedimentos marinos paleocenos y ha sido reconocida para el Daniano de la Argentina (Bertels, 1964). Coleites reticulosus, por su particular morfología, no pudo haber pasado desapercibido ni haber sido confundido con otra especie de la microfauna tipo Midway de la Argentina. Su aparente ausencia se debió probablemente a su escasez, pero el estudio intensivo de los afloramientos de la Formación Roca en Liu Malal (provincia de Mendoza) y de la Formación Salamanca en la hoja geológica Campamento Villegas (provincia del Chubut), ha confirmado su presencia. De esta manera, Coleites reticulosus, que posee las características de un buen fósil guía por su morfología conspicua, restringida distribución temporal y amplia distribución geográfica (figura 1), se agrega a la lista de 234 especies bentónicas danianas reconocidas en Argentina en la revisión taxonómica de Malumián y Caramés (1995).
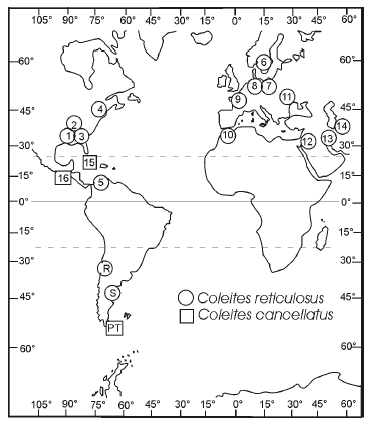
Figura 1. Distribución mundial de Coleites reticulosus (Plummer) y C. cancellatus (Cushman y Bermúdez). Coleites reticulosus (Plummer) en: 1-4, EEUU: 1, Formación Wills Point, Daniano, Texas. 2, Formación Midway, Paleoceno, Arkansas. 3, Formación Porters Creek, Paleoceno, Alabama. 4, Formación Vincentown, Selandiano superior-Thanetiano, New Jersey. 5, Venezuela, Formación Guasare, Paleoceno. 6, Suecia, Daniano medio, Daniano superior y Selandiano. 7, Polonia, Montiano. 8, Bélgica, Holanda y Alemania, Daniano-Montiano, provincia de Limburg ; Alemania Oriental, Montiano. 9, Francia, Montiano, cuenca de París. 10, Algeria y Túnez Paleoceno; Túnez, Paleoceno inferior-superior bajo. 11, Unión Soviética, Crimea, Paleoceno. 12, Líbano y Siria, Paleoceno. 13, Irán, Paleoceno. 14, Unión Soviética, Turkmenia, Paleoceno. R-S, Argentina: R, Formación Roca, Daniano inferior, perfíl Liu Malal, provincia de Mendoza (este trabajo); S, Formación Salamanca, Daniano superior, perfil Sector Linares, provincia del Chubut (este trabajo). Coleites cancellatus (Cushman y Bermúdez) en: 15, Cuba, Eoceno superior. 16, Guatemala, Paleoceno. PT, Argentina, Formación Punta Torcida, Eoceno Inferior, perfil Cabo Campo del Medio, isla de Tierra del Fuego (este trabajo) / geographic distribution of Coleites reticulosus (Plummer) and C. cancellatus (Cushman y Bermúdez). Coleites reticulosus (Plummer) in: 1-4, EEUU. 1, Wills Point Formation, Danian, Texas. 2, Midway Formation, Paleocene, Arkansas. 3, Porters Creek Formation, Paleocene, Alabama. 4, Vincentown Formation, Upper Selandian-Thanetian, New Jersey. 5, Venezuela, Guasare Formation, Paleocene. 6, Sweden, Middle Danian, Upper Danian and Selandian. 7, Poland, Montian. 8, Belgium, The Netherlands and Germany, Danian-Montian, Limburg region; East Germany, Montian. 9, France, Montian, Paris Basin. 10, Algeria and Tunisia, Paleocene; Tunisia, Lower to lower Middle Paleocene. 11, Soviet Union, Crimea, Paleocene. 12, Lebanon and Syria, Paleocene. 13, Iran, Paleocene. 14, Soviet Union, Turkmenia, Paleocene. R and S, Argentina: R, Roca Formation, Lower Danian, Liu Malal section, Mendoza province (this work); S, Salamanca Formation, Upper Danian, Sector Linares section, Chubut province (this work). Coleites cancellatus (Cushman y Bermúdez) in: 15, Cuba, Upper Eocene. 16, Guatemala, Paleocene. PT, Argentina, Punta Torcida Formation, Lower Eocene, Cabo Campo del Medio section, Tierra del Fuego Island (this work).
Coleites cancellatus (Cushman y Bermúdez), otro representante del género Coleites, de morfología conspicua y una distribución restringida al Paleoceno de Guatemala y el Eoceno Superior de Cuba (cf. Cushman y Bermúdez, 1948), fue hallado en el Eoceno Inferior de la Formación Punta Torcida, de la Isla Grande de Tierra del Fuego.
La presente contribución da a conocer el registro del género Coleites Plummer en la Argentina representado por las especies Coleites reticulosus y Coleites cancellatus.
Los ejemplares ilustrados se depositaron en la colección de Micropaleontología de la Dirección Nacional del Servicio Geológico bajo los números DNSG 2500 a 2513.
Paleontología sistemática
Se utiliza la clasificación sistemática propuesta por Loeblich y Tappan (1988).
Suborden Rotaliina Delage y Hérouard, 1896
Superfamilia Chilostomellacea Brady, 1881
Familia Coleitidae Loeblich y Tappan, 1984
Género Coleites Plummer, 1934
Especie tipo. Pulvinulina reticulosa Plummer, 1927.
Coleites reticulosus (Plummer, 1927) Figuras 2.1-2.15
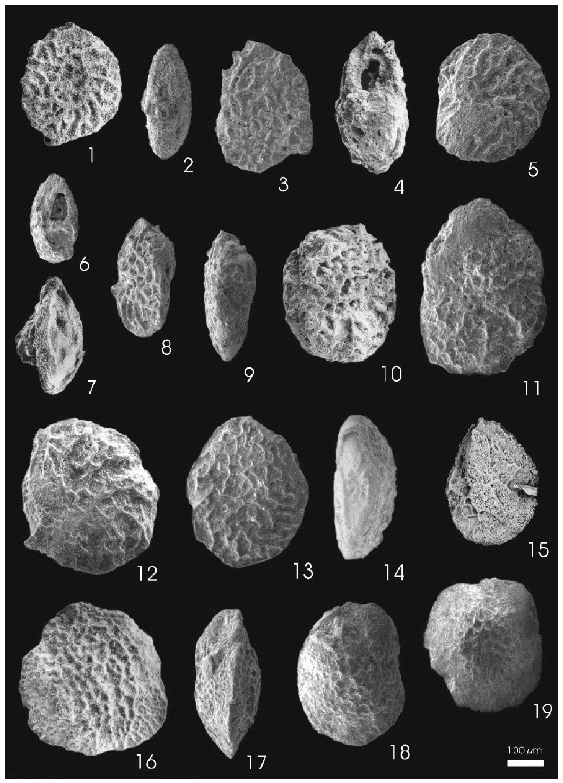
Figura 2.1-2.10, Coleites reticulosus (Plummer), Formación Salamanca / Salamanca Formation. 1, DNSG 2503, lado espiral / spiral side; 2, DNSG 2504, vista periférica / peripheral view; 3-4, DNSG 2505, 3, lado espiral / spiral side, 4, vista periférica / peripheral view; 5, DNSG 2506, lado umbilical / umbilical side; 6, DNSG 2507, vista periférica / peripheral view; 7, DNSG 2508, vista periférica / peripheral view; 8, DNSG 2509, vista periférica / peripheral view; 9-10, DNSG 2510, 9, vista periférica / peripheral view, 10, lado umbilical / umbilical side. 2.11-2.15, Coleites reticulosus (Plummer), Formación Roca / Roca Formation. 11, DNSG 2511, lado umbilical / umbilical side; 12-14, DNSG 2512, 12, lado umbilical / umbilical side, 13, lado espiral / spiral side, 14, vista periférica / peripheral view; 15, DNSG 2513, lado umbilical / umbilical side. 2.16-2.19, Coleites cancellatus (Cushman y Bermúdez), Formación Punta Torcida / Punta Torcida Formation. 16-17, DNSG 2500, 16, lado espiral / spiral side, 17, DNSG 2500, vista periférica / peripheral view; 18, DNSG 2501, lado umbilical / umbilical side; 19, DNSG 2502, lado umbilical / umbilical side.
1927. Pulvinulina reticulosus Plummer, p. 152, lám. 12, fig. 5.
1934. Coleites reticulosus (Plummer); Plummer, p. 605.
Material. Cuenca Neuquina, Formación Roca, Daniano inferior (Concheyro y Villa, 1996; Pires et al., 1999), perfil Liu Malal, muestra M43, facies de areniscas finas basales: 1 ejemplar; muestra M52, facies de calizas suprayacentes: 8 ejemplares. Cuenca del Golfo San Jorge, Formación Salamanca, Daniano superior (Panza et al., 2004), hoja geológica Campamento Villegas, perfil sector Linares, muestra 1235, sección superior carbonáticoclástica de la formación: 72 ejemplares.
Descripción. Conchilla biconvexa asimétrica a plano convexa, periferia aguda con carena lisa y roma. Lado espiral plano o ligeramente convexo, evoluto, ornamentado en su totalidad por un fuerte reticulado que oblitera las suturas; sólo en unos pocos ejemplares son visibles las últimas suturas camerales. Lado umbilical fuertemente convexo, involuto, con 6 a 8 cámaras visibles, zona umbilical variable, al ras (figura 2.10) o ligeramente elevada por engrosamiento de la pared (figura 2.5) en los ejemplares de la Formación Salamanca, o ligeramente deprimida (figuras 2.11, 2.12) en los ejemplares de la Formación Roca; suturas camerales al ras excepto algunos ejemplares con las últimas suturas deprimidas; superficie ornamentada por elevaciones delgadas que componen un diseño reticulado poligonal, en general menos definido que en el lado espiral ya que las elevaciones cambian de dirección o se interrumpen en las suturas camerales. Zona cercana a la abertura lisa. Abertura oval, sobre el lado umbilical, próxima a la periferia.
Observaciones. Según Pozaryska (1965), probablemente Coleites vanbelleni Marie sea sinónimo de C. reticulosus. Coleites reticulosus se diferencia de C. cancellatus de la Formación Punta Torcida, por la conchilla más asimétrica, periferia más redondeada y reticulado del lado espiral más grueso. Ejemplares juveniles (sin estadío adulto rectilíneo) de C. reticulosus se diferencian de la ilustración original de C. danicus Brotzen (1948) del Daniano y Selandiano de Suecia, también correspondiente a un ejemplar juvenil, por poseer una conchilla más asimétrica ornamentada con retículos de tamaño más o menos uniforme sobre la totalidad de ambos lados, en lugar de retículos en general más grandes y de tamaño variable, desarrollados únicamente sobre la zona central del lado espiral, y pequeñas costillas en la zona central del lado umbilical. Coleites laevigatus Toulmin (1941), descripto en Alabama y referido a la parte inferior, paleocena media (P4), del Grupo Wilcox, y del Daniano de su infrayacente, el Grupo Midway superior en Georgia, no posee la ornamentación característica del género, y según la descripción original carecería del tubo interno mencionado por Plummer en la descripción del genotipo, planteando una duda sobre su posición genérica. Cabe mencionar el registro de Coleites (?) sp. en Mabesoone et al. (1967), formando parte de una asociación paleocena de tipo Midway en la Formación María Farinha del nordeste de Brasil.
Distribución (figura 1). Descrito originalmente para la Formación Midway de Texas (Plummer, 1927). Otros registros en América del Norte corresponden a la Formación Wills Point, Daniano cuspidal, Zona P1d y/o P2, Texas (Kellough, 1965); Formación Midway, Paleoceno, Arkansas (Cushman, 1951); Formación Porters Creek, Paleoceno, Alabama (Cushman, 1951); y Formación Vincentown, Paleoceno superior, Zona P4, New Jersey (Hofker, 1955). En América del Sur se registra en Venezuela, Formación Guasare, Paleoceno, Estado de Zulia (Sutton, 1946). En Europa se registra en Suecia, en sedimenti-tas del Daniano medio, Daniano superior y Selandiano (Brotzen, 1948, 1959); en Polonia, en el Montiano (parcialmente equivalente al Selandiano) (Pozaryska, 1965; Pozaryska y Szczechura, 1968); en Bélgica, Holanda y Alemania, en la provincia de Limburg, en el Daniano-Montiano (Pozaryska, 1965) y en Alemania Oriental, en el Montiano (Kiessel, 1970); y en Francia, cuenca de París, en rocas del Montiano (Pozaryska, 1965). En África se conocen registros en el Paleoceno de Argelia y Túnez (Cuvillier et al., 1955); y en Túnez, en la Zona P1 del Daniano (Aubert y Berggren, 1976) y en la parte inferior de la sección inferior de El Kef, Paleoceno temprano y Paleoceno medio basal (Zonas de foraminíferos planctónicos IIa,b,c y III, cf. Salaj et al., 1976). Los registros en Asia, también en sedimentitas paleocenas, se ubican en Unión Soviética, en Crimea (Mjatluk, 1953, y Vasilenko, 1961, según Pozaryska, 1965) y en Turkmenia (Kalugin et al., 1964, y/o Morozova et al., 1967, en Berggren y Aubert, 1975); en Líbano y Siria (Cuvillier et al., 1955); y en Irán (página web del Geological Survey of Irán www. gsi.ir/?Lang=en&p=52).
Coleites cancellatus (Cushman y Bermúdez), 1936 Figuras 2.16-2.19
1936. Pulvinulinella cancellata Cushman y Bermúdez, p. 35, lám. 6, fig. 7.
1948. Coleites cancellatus (Cushman y Bermúdez); Cushman y Bermúdez, p. 83, lám. 13, figs. 10-11.
Material. Cuenca Austral, Formación Punta Torcida, Eoceno inferior (Olivero y Malumián, 1999), perfil Cabo Campo del Medio, muestra 13a, facies de areniscas del Miembro inferior: 5 ejemplares.
Descripción. Conchilla trocoespiral, biconvexa asimétrica, periferia aguda carenada y ligeramente lobulada. Lado espiral evoluto con dos vueltas, superficie ornamentada por elevaciones elongadas que forman un reticulado amplio y profundo en la parte central, y uno más pequeño y menos profundo próximo a la periferia. Lado umbilical con 6-7 cámaras, excepcionalmente 8, suturas deprimidas, y ornamentación reticular restringida a las suturas deprimidas y a la zona umbilical levemente deprimida. Zona próxima a la abertura lisa. Abertura oval, sobre el lado umbilical y próxima a la periferia.
Observaciones. Se diferencia de C. reticulosus por su periferia aguda, el lado espiral con reticulado más pequeño y menos uniforme, y el umbilical más convexo con ornamentación reticular limitada a la zona umbilical y a las suturas más claramente visibles. Coleites abuillotensis Cushman y Bermúdez (1948) del Eoceno inferior de Haití y C. guatemalensis Cushman y Bermúdez (1948) del Eoceno inferior de Guatemala, se diferencian de C. cancellatus por sus conchillas extremadamente comprimidas, además, C. guatemalensis, posee estadío juvenil trocoespiral más finamente reticulado. Coleites pasionensis Cushman y Bermúdez (1948), del Eoceno inferior de Guatemala, se diferencia por la periferia más irregular, reticulado mucho más grueso en la porción inicial y suturas elevadas y anchas sobre el lado dorsal.
Distribución (figura 1). Descrito originalmente para el Eoceno superior de Cuba (Cushman y Bermúdez, 1936). Según Cushman y Bermúdez (1948), la especie es frecuente en el Eoceno de Cuba y rara en el Paleoceno de Guatemala.
Comentarios paleogeográficos y paleoecológicos
El dimorfismo del género Coleites fue mencionado desde su descripción original (Plummer, 1934). Marie (1943) indicó que las conchillas completamente trocoespirales son megalosféricas, y que aquellas con estadío inicial trocoespiral y estadío adulto rectilíneo, son microsféricas. Notablemente, todos los ejemplares hallados en la Argentina carecen del estadío adulto desenroscado, por lo que a priori serían megalosféricos. Sin embargo, al desconocer la existencia de estudios biométricos específicos, y como entre los foraminíferos se conocen algunas especies trimorficas y otras dimórficas con algunos ejemplares megalosféricos con prolóculos más pequeños que los de las formas microsféricas (cf. Boltovskoy, 1965), antes de aceptar una relación directa entre prolóculos megalosféricos y ausencia de estadío adulto rectilíneo, se intentó medir el diámetro de los prolóculos bajo microscopio con luz transmitida y líquido de inmersión, pero la fuerte ornamentación y la recristalización y/o relleno impidió su distinción.
Coleites es un género que se conoce desde el Daniano al Lutetiano, pero si consideramos que, como en la mayoría de los géneros de foraminíferos actuales, las conchillas más pequeñas y con menor número de cámaras son megalosféricas y resultan de la reproducción asexual de la generación microsférica, la ausencia de conchillas microsféricas podría explicarse, como sucede en varias especies actuales, por una disolución selectiva de las mismas durante esta etapa asexual de la reproducción (cf. Boltovskoy, 1965). Además, según Boltovskoy y Wright (1976) y Nigam y Caron (2000), entre otros, la reproducción dimórfica de los foraminíferos requeriría de determinadas condiciones ambientales, incluso del tipo de alimento disponible. Al respecto, Hallock (1985), menciona que en foraminíferos grandes, el predominio de formas megalosféricas refleja el hecho habitual de que entre dos generaciones microsféricas, que resultan de etapas de reproducción sexual, se suceden varias generaciones que son producto de etapas de reproducción asexual, y esto indica una relativa estabilidad ambiental. Determinados parámetros ambientales explicarían el hecho de que en varias especies actuales, las generaciones megalosféricas se suceden una detrás de la otra y solamente de vez en cuando aparece una generación microsférica (cf. Boltovskoy, 1965), y por otra parte, sugieren la posibilidad de que el paleoambiente daniano de las Formaciones Roca y Salamanca no haya sido propicio para la reproducción sexual por lo cual se tienen solamente conchillas microsféricas.
La distribución de C. reticulosus (figura 1) puede explicarse porque su estadío inicial, como el de la mayoría de los foraminíferos bentónicos, debió ser pelágico (Bettenstaedt y Wicher, 1955). Su dispersión, como la de otros foraminíferos pequeños, debió efectuarse en el estadío de gameta, que ayudadas por corrientes oceánicas y de fondo, se establecieron en plataformas, se reprodujeron y continuaron su avance (cf. Berggren y Phillips, 1972). Sin embargo, la morfología discoidal del género Coleites sugiere un hábito de vida epifaunal, probablemente epífito, que explicaría más simplemente su distribución. La extensión de condiciones climáticas equitativas y la casi uniforme estructura térmica vertical durante el Paleoceno, con una circulación más controlada por diferencias de salinidad que de temperatura (cf. Kennett, 1982), explicaría la amplia distribución.
Coleites reticulosus habría encontrado un ambiente propicio en los ambientes que originaron los estratos de la sección superior, carbonáticoclástica, de la Formación Salamanca. Estos estratos afloran en el área de la Hoja Geológica General Villegas a unos 6 m por encima del contacto con las tobas de la Formación Marifil, y corresponden a areniscas medianas a gruesas hasta sabulíticas, cuarzosas y con clastitos verdosos de rocas jurásicas, y concreciones más resistentes que el conjunto. Su depositación se produjo bajo condiciones marinas someras de salinidad normal (cf. Panza et al., 2004). Aunque para la Formación Salamanca es más extendido el desarrollo de microfaunas con baja diversidad correspondientes a biofacies de fondos blandos, arcillosos, como las dominadas por Buliminella isabelleana (cf. Malumián y Caramés, 1995), también se habrían formado sustratos firmes que habrían permitido el desarrollo de fondos vegetados, y C. reticulosus habría vivido adherido a esa vegetación. Otro testimonio de la presencia de sustratos firmes en la misma zona son las icnofacies de Glossifungites que destacan la presencia de discontinuidades erosivas (Rodríguez y Panza, 2003). Coleites reticulosus también habría encontrado un hábitat adecuado en las calizas de la Formación Roca, que según Pires et al. (1999) contienen restos de algas que indican una profundidad dentro de la zona fótica, y corresponden a una facies de moderada energía por debajo del tren de olas, con fauna de aguas relativamente someras (foraminíferos planctónicos y bentónicos, moluscos, briozoos, crustáceos, equinodermos). Siendo C. reticulosus una especie con registro en todo el Daniano e incluso más joven, en el Selandiano y Thanetiano, su aparente desaparición en los estratos más jóvenes de la Formación Salamanca habría sido consecuencia del extendido desarrollo de fondos blandos, testimoniados por el dominio de especies infaunales (cf. Malumián y Caramés, 1995). En la Formación Roca, la desaparición correspondería al desarrollo de facies algo más profundas y más finas.
Coleites reticulosus parece ser una especie limitada a temperaturas templado-cálidas según indican la presencia de cocodrilos, la ausencia de anillos criogénicos en maderas de gimnospermas en la Formación Salamanca (cf. Volkheimer, 1971; Petriella y Archangelsky, 1975; Brea et al., 2005), y algunas especies de nanoplancton calcáreo típicas de aguas someras templadas, elementos fosfatizados que sugieren temperaturas superiores a los 18°C durante su depositación, e indicadores de clima árido como sedimentitas evaporíticas y caliche en la Formación Roca (cf. Ramos, 1981; Concheyro y Villa, 1996; Pires et al., 1999).
Respecto de C. cancellatus, es extremadamente escaso en los niveles arenosos más bajos del miembro inferior de la Formación Punta Torcida (Eoceno inferior), en su perfil tipo en Cabo Campo del Medio, Tierra del Fuego. Olivero y Malumián (1999) y Olivero et al. (2002) interpretan los niveles arenosos como depósitos de sistemas turbidíticos depositados bajo condiciones de fondos disaeróbicos a anóxicos por sus microfósiles calcáreos bien conservados, baja diversidad de la asociación de foraminíferos bentónicos, contenido de materia orgánica y microfósiles piritizados. Sin embargo, según Jannou (comunicación verbal, 2005), la asociación de foraminíferos de los niveles arenosos entre los que se encuentra C. cancellatus tiene una muy baja relación planctónicos/bentónicos y especies bentónicas de ambientes marinos más someros por lo que interpreta fueron transportadas.
Agradecimientos
Las autoras agradecen muy especialmente a N. Malumián la lectura crítica del manuscrito y haber facilitado las muestras de la Formación Punta Torcida y los ejemplares de Coleites reticulosus de la Formación Salamanca; a los licenciados J.L. Panza y M. Pires, las muestras de las Formaciones Salamanca y Roca, respectivamente, y al Lic. G. Jannou los ejemplares de C. cancellatus de la Formación Punta Torcida. Es un aporte al proyecto PICT 2002 N°12257, Cenozoico de la Plataforma Continental Argentina y Patagonia.
Bibliografía
1. Aubert, J. y Berggren, W.A. 1976. Paleocene benthic foraminiferal biostratigraphy and paleoecology of Tunisia. Bulletin du Centre de Recherches Pau-SNPA 10: 379-469. [ Links ]
2. Berggren, W.A. y Aubert, J. 1975. Paleocene benthonic foraminiferal biostratigraphy, paleobiogeography and paleoecology of Atlantic Tethyan regions: Midway-type fauna. Palaeo-geography, Palaeoclimatology, Palaeoecology 18: 73-192. [ Links ]
3. Berggren, W.A. y Phillips, J.D. 1972. The influence of the continental drift on the distribution of Cenozoic Benthonic foraminifera in the Mediterranean and Caribbean regions: Symposium on Geology of Libya (Tripoli, 1969), Catholic Press, Beirut: 263-299. [ Links ]
4. Bertels, A. 1964. Micropaleontología del Paleoceno de General Roca (provincia de Río Negro). Revista del Museo de La Plata (nueva serie), Paleontología 23: 125-184. [ Links ]
5. Bettenstaedt, F. y Wicher, C.A. 1955. Stratigraphic correlation on the Upper Cretaceous and Lower Cretaceous in the Tethys and Boreal by the aid of microfossils. Proceedings of the 4º World Petroleum Congress (Roma), Section 1/D, paper 5: 494-516. [ Links ]
6. Boltovskoy, E. 1965. Los Foraminíferos recientes: biología, método de estudio, aplicación oceanográfica. Manuales de EUDEBA, Ciencias Naturales, EUDEBA, Buenos Aires, 510 pp. [ Links ]
7. Boltovskoy, E. y Wright, W.R. 1976. Recent Foraminifera. Dr. W. Junk bv Publishers, The Hague, 515 pp. [ Links ]
8. Brea, M., Matheos, S., Zamuner, A. y Ganuza, D. 2005. Análisis de los anillos de crecimiento del bosque fósil de Victor Szlápelis, Terciario inferior del Chubut, Argentina. Ameghiniana 42: 407-418. [ Links ]
9. Brotzen, F. 1948. The Swedish Paleocene and its foraminiferal fauna. Sveriges Geologiska Undersökning, serie C, 42: 3-140. [ Links ]
10. Brotzen, F. 1959. On Tylocidaris species (Echinoidea) and the stratigraphy of the Danian of Sweden. Sveriges Geologiska Undersökning 54: 1-81. [ Links ]
11. Concheyro, A. y Villa, G. 1996. Maestrichtian-Danian (K/P) calcareous nannofossils in the Liu Malal section, Northern Patagonia, Argentina. Palaeopelagos 6: 281-297. [ Links ]
12. Cushman, J.A. 1951. Paleocene foraminifera of the Gulf Coastal Region of the United States and adjacent areas. Geological Survey Professional Paper 232: 1-75. [ Links ]
13. Cushman, J.A. y Bermúdez, P.J. 1936. New genera and species of foraminifera from the Eocene of Cuba. Contributions from the Cushman Laboratory for Foraminiferal Research 12: 27-38. [ Links ]
14. Cushman, J.A. y Bermúdez, P.J. 1948. The genus Coleites and its species. Contributions from the Cushman Laboratory for Foraminiferal Research 24: 81-84. [ Links ]
15. Cuvillier, J., Dalbiez, F., Glintzboeckel, C., Lys, M., Magné, J., Perebaskine, V. y Rey, M. 1955. Études micropaléontologiques de la limite Crétacé-Tertiaire dans les mers mésogéennes. Proceedings of the 4º World Petroleum Congress (Roma), Sect. 1/D, Paper 6: 517-544. [ Links ]
16. Hallock, P. 1985. Why are larger foraminifera large? Paleobiology 11: 195-208. [ Links ]
17. Hofker, H. 1955. The foraminifera from the Vincentown Formation. Reports from the McLean Foraminiferal Laboratory 2: 1-21. [ Links ]
18. Kalugin, P.I., Dmitriev, A.V. y Kozhevnikova, G.E. 1964. Stratigrafiya verkhnemelovykh i paleotsenovykh otlozhenii Kopet-Daga I Badkhyza. Ashkhabad, 343 pp. [ Links ]
19. Kellough, G.R. 1965. Paleoecology of the Foraminiferida of the Wills Point Formation (Midway Group) in northeast Texas. Transactions Gulf Coast Associations of Geological Societies 15: 73-153. [ Links ]
20. Kennett, J.P. 1982. Marine Geology. Prentice-Hall. Englewood Cliffs, New Jersey, 813 pp. [ Links ]
21. Kiessel, Y. 1970. Die Foraminifernfauna der paläozänen und eozänen Schichtenfolge der Deutschen Demokratischen Republik. Paläontolgische Abhandlungen, Abteilung A 4: 163-394. [ Links ]
22. Loeblich, A.R. y Tappan, H. 1988. Foraminiferal genera and their classification. Van Nostrand Reinhold, New York, 1: 970 p. 2: 212 p. 847 láms. [ Links ]
23. Mabesoone, J.M., Tinoco, I.M. y Coutinho, P.N. 1967. The Mesozoic-Tertiary boundary in northeastern Brazil. Palaeogeography, Palaeoclimatology, Palaeoecology 4: 161-185. [ Links ]
24. Malumián, N. y Caramés, A. 1995. El Daniano marino de Patagonia (Argentina): paleobiogeografía de los foraminíferos bentónicos. Asociación Paleontológica Argentina, Publicación Especial N°3 Paleogeno de América del Sur: 83-105. [ Links ]
25. Marie, P. 1943. Sur la valeur stratigraphique du genre Coleites Plummer. Compte Rendus Sommaire des Séances, Société Géologique de France 3: 10-20. [ Links ]
26. Morozova, V.G., Kozhevnikova, G.E. y Kuryleva, A.M. 1967. Datsko-Paleotsenovye Raznofatsial'nye otlozheniya Kopel-Data i metody ikh korrelyatsii po foraminiferam. Trudy Geologicheskogo Instituta, Akademiya Nauk SSSR 157, 208 pp. [ Links ]
27. Mjatluk, E.V. 1953. Spirillinidy, Rotalinidy, Epistominidy i Asterigerinidy. Trudy Vsesoyznogo Neftyanogo Nauchno-issledo-vatel'skogo Geologo-razvedochnogo Instituta (VNIGRI), n. ser 71, 273 pp. [ Links ]
28. Nigam, R. y Caron, D.A. 2000. Does temperature affect dimorphic reproduction in benthic foraminifera? A culture experiment on Rosalina leei. Research Communications, Current Science 79: 1105-1106. [ Links ]
29. Olivero, E.B. y Malumián, N. 1999. Eocene stratigraphy of Southeastern Tierra del Fuego Island, Argentina. Bulletin of the American Association of Petroleum Geologists 83: 295-313. [ Links ]
30. Olivero, E.B., Malumián, N. y Jannou, G. 2002. Estratigrafía y facies de sistemas turbidíticos, estuáricos y de plataforma, Eoceno, Andes Fueguinos, Argentina. 9° Reunión Argentina de Sedimentología (Córdoba), Resúmenes: p. 57. [ Links ]
31. Panza, J.L., Sacomani, L.E., Rodríguez, M.F. y Aragón, E. 2004. Hoja Geológica 4366-27 "Campamento Villegas", provincia del Chubut, escala 1: 100.000. Instituto de Geología y Recursos Minerales, Servicio Geológico Minero Argentino, Boletín 330, 156 pp. [ Links ]
32. Petriella, B.T.P. y Archangelsky, S. 1975. Vegetación y ambiente en el Paleoceno de Chubut. 1° Congreso Argentino de Paleontología y Bioestratigrafía (San Miguel de Tucumán), Actas 2: 257-270. [ Links ]
33. Pires, M., Náñez, C., Casadío, S. y Parras, A. 1999. Sedimentología, paleontología y estratigrafía del límite Cretácico-Paleógeno. 14° Congreso Geológico Argentino (Salta), Actas 1: 463-466. [ Links ]
34. Plummer, H.J. 1927. Foraminifera of the Midway Formation in Texas. Bulletin University of Texas Bureau of Economic Geology and Technology 2644, 206 pp. [ Links ]
35. Plummer, H.J. 1934. Epistominoides and Coleites new genera of foraminifera. American Midland Naturalist 15: 601-608. [ Links ]
36. Pozaryska, K. 1965. Foraminifera and biostratigraphy of the Danian and Montian in Poland. Paleontologia Polonica 14: 156 pp. [ Links ]
37. Pozaryska, K. y Szczechura, J. 1968. Foraminifera from the Paleocene of Poland, their ecological and biostratigraphical meaning. Paleontologia Polonica 20: 107 pp. [ Links ]
38. Ramos, V.A. 1981. Descripción Geológica de la Hoja 33c, Los Chihuidos Norte, Provincia del Neuquén. Servicio Geológico Nacional, Boletín 182: 103 pp. [ Links ]
39. Rodríguez, M.F. y Panza, J.L. 2003. La icnofacies de Glossifungites en la Formación Salamanca, Paleoceno, paraje Campamento Villegas, Chubut, Argentina. Asociación Paleontológica Argentina, Publicación Especial N° 9, 4º Reunión Argentina de Icnología y 2º Reunión de Icnología del Mercosur: 157-167. [ Links ]
40. Salaj, J., Pozaryska, K. y Szczechura, J. 1976. Foraminiferida, zonation and subzonation of the Paleocene of Tunisia. Acta Paleontologica Polonica 21: 127-190. [ Links ]
41. Sutton, F.A. 1946. Geology of the Maracaibo Basin, Venezuela. Bulletin of the American Association of Petroleum Geologists 30: 1621-1741. [ Links ]
42. Toulmin, L.D. 1941. Eocene smaller foraminifera from the Salt Mountain Limestone of Alabama. Journal of Paleontology 15: 567-611. [ Links ]
43. Vasilenko, V.P. 1961. Foraminifery verchnego mela poluostrova Mangylakja. Proceedings of the All Union Petroleum Scientific Research Institute for Geological Survey (VNIGRI) 171: 487 pp. [ Links ]
44. Volkheimer, W. 1971. Aspectos paleoclimáticos del Terciario Argentino. Revista del Museo Argentino de Ciencias Naturales Bernardino Rivadavia, Paleontología 1: 243-264. [ Links ]
Recibido: 6 de diciembre de 2005.
Aceptado: 29 de noviembre de 2006.

