










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.44 n.3 Buenos Aires jul./sep. 2007
Nuevos registros para la flora cretácica descripta por Halle (1913) en lago San Martín, Santa Cruz, Argentina
Mauro G. Passalia 1
1CONICET, Grupo de Estudios Ambientales, Centro Regional Universitario Bariloche-Universidad Nacional del Comahue, Quintral 1250, 8400 San Carlos de Bariloche, Río Negro, Argentina. mpassalia@crub.uncoma.edu.ar
Resumen. Se han estudiado dos nuevos sitios portadores de plantas fósiles cretácicas, Bajo Comisión 1 y 2, localizados en proximidades del área prospectada originalmente por Thor Halle, al norte del lago San Martín, provincia de Santa Cruz, Argentina. Sus depósitos corresponden a la Formación Kachaike (Albiano tardío - Cenomaniano). Se han identificado alrededor de veinte especies, pertenecientes en su mayoría a pteridófitas, pteridospermas, coníferas y frondes de afinidad sistemática incierta. Briófitas, bennettitales, ginkgoales y angiospermas están también presentes, aunque como elementos subordinados. Entre ellos, se reconocieron nuevos taxones para esta unidad: Pachypteris auriculata sp. nov., Brachyphyllum kachaikensis sp. nov. y Thorhallenia dentata gen. et sp. nov. A su vez, se proponen reasignaciones sistemáticas para otros taxones (Gleichenites juliensis Herbst en vez de Gleichenites cf. G. micromerus Heer y Phyllopteroides cf. P. laevis Cantrill y Webb en vez de Asplenites lanceolatus Halle). Existe una evidente similitud entre las asociaciones Bajo Comisión 1 y 2 con aquella descripta por Halle, lo cual sugiere que estos niveles son, en efecto, los estudiados originalmente por este autor.
Abstract. New records of the cretaceous flora described by Halle (1913) from San Martín Lake, Santa Cruz, Argentina. Two new sites with Cretaceous fossil plants, Bajo Comisión 1 and 2, have been studied. They are located north of San Martín Lake, Santa Cruz Province, Argentina, and their deposits belong to the Kachaike Formation (Late Albian - Cenomanian). This area was previously studied by Thor Halle who provided the description of a fossil plant assemblage. About twenty species have been recognized at the new sites, mostly belonging to pteridophytes, pteridosperms, conifers as well as morphogenera of uncertain botanical affinity. Bryophytes, bennettites, ginkgophytes and angiosperms are also present as subordinated components. Among them, new taxa are described: Pachypteris auriculata sp. nov., Brachyphyllum kachaikensis sp. nov. and Thorhallenia dentata gen. et sp. nov. In turn, two species, previously described by Halle, have been reassigned to other genera or species: Phyllopteroides cf. P. laevis Cantrill and Webb instead of Asplenites lanceolatus Halle, and Gleichenites juliensis Herbst instead of Gleichenites cf. G. micromerus Heer and Phyllopteroides cf. P. laevis Cantrill y Webb instead of Asplenites lanceolatus Halle. There is a considerable resemblance between Bajo Comisión 1 and 2 assemblages and the flora previously described by Halle, suggesting they belong to the same levels.
Palabras clave. Argentina; Santa Cruz; Formación Kachaike; Cretácico; Pteridófitas; Gimnospermas; Angiospermas.
Key Words. Argentina; Santa Cruz; Kachaike Formation; Cretaceous; Pteridophytes; Gymnosperms; Angiosperms.
Introducción
Durante el Jurásico Superior y todo el Cretácico la cuenca Austral recibió el aporte de abundante sedimentación de origen marino y continental. En la región cordillerana comprendida entre los lagos San Martín, Cardiel y Belgrano (en la provincia de Santa Cruz) hay numerosos afloramientos, en parte correspondientes a las Formaciones Kachaike, Piedra Clavada y Belgrano, que reflejan una sucesión de eventos transgresivo-regresivos, y cuyo contenido fosilífero incluye mega, microflora y restos de invertebrados.
El estudio de las megafloras fósiles contenidas en estas secuencias ha sido llevado a cabo, primero, por Halle (1913a), luego por Frenguelli (1935) y Piátnitzky (1938) y, posteriormente, por Baldoni y Ramos (1981), Ruiz (1984), Longobucco et al. (1985), Llorens y Del Fueyo (2003) y Cúneo y Gandolfo (2005). El yacimiento estudiado por Halle (1913a), abarca una franja de dirección norte-sur, dispuesta al este de bahía de la Lancha en lago San Martín. El mismo comprende cuatro localidades: "a", "b", "c" y una sección denominada "Upper part of the Río Fósiles Valley". Es de estas dos últimas de donde proviene la mayor parte del material fósil descripto por Halle (1913a) y que incluye abundantes restos megaflorísticos, incluyendo briófitas, pteridófitas, gimnospermas y dos posibles fragmentos de angiospermas. La necesidad de un mayor conocimiento de la diversidad mega y microflorística de estas secuencias y la posible existencia de facies heterocrónicas para el Cretácico Inferior de la región, han motivado el emprendimiento de un nuevo estudio de estos depósitos.
En el presente artículo se dan a conocer los resultados del análisis de nuevos restos megaflorísticos, pertenecientes a la Formación Kachaike, obtenidos en las proximidades de los yacimientos estudiados originalmente por Halle (1913a), los que se cotejan con la lista proporcionada por este autor, con el fin de establecer sus eventuales relaciones.
Procedencia, paleoambiente y edad
En un área situada al norte del lago San Martín, en la provincia de Santa Cruz (Argentina), han sido localizados dos nuevos sitios portadores de plantas fósiles, denominados Bajo Comisión 1 (BC1) y Bajo Comisión 2 (BC2), que son el objeto del presente trabajo (figura 1). El primero de ellos (BC1, 48º51'9.9"S, 72º09'9.6"O) presenta un perfil de más de 250 metros de espesor (Cladera y Limarino, inédito) en el que se han reconocido seis niveles portadores de plantas (denominados NF A al NF F) (figura 2). Los fósiles consisten en impresiones y compresiones de fragmentos foliares y estructuras reproductivas. En la base del perfil, correspondiente a un ambiente sedimentario de prodelta, se encuentra el nivel NF A. Los restantes niveles se hallan en los términos superiores de la parte media de la secuencia. El nivel NF B corresponde a un frente deltaico y los niveles NF C y D a una plataforma subácuea (NF B y NF C distantes sólo por escasos metros). Por último, los niveles NF E y NF F fueron depositados en un ambiente de plataforma subaérea (Cladera y Limarino, com. pers., 1994). El marco paleoambiental del perfil estudiado puede resumirse como una secuencia deltaica progradante, sujeta, al menos, a tres eventos transgresivos (Cladera y Limarino, com. pers.) El segundo sitio (BC2, 48º51'00''S, 72º10'00''O), consiste en una pequeña barranca (situada sobre una de las márgenes de un afluente del arroyo Bajo Comisión) compuesta por intercalaciones de areniscas portadoras de restos de plantas fósiles. La relación litoestratigráfica entre las secuencias BC1 y BC2, por el momento, es poco clara; no obstante, ambas forman parte de la Formación Kachaike y su depositación corresponde al Subciclo Regresivo Kachaike-Piedra Clavada (Ciclo lago San Martín), definido por Arbe (2002).
La Formación Kachaike, identificada por Riccardi (1971) para la región del lago San Martín, se dispone transicionalmente sobre los depósitos marinos del Miembro Superior de la Formación Río Mayer (Arbe, 2002). Sobre la base de asociaciones de micro y megasporas, polen y quistes de dinoflagelados, Baldoni et l. (2001) sugirieron una edad Aptiano tardía - Albiano temprana (probablemente Albiano temprana), para los depósitos de la Formación Kachaike en el perfil Arroyo Caballo Muerto. Una asociación de dinoquistes, de la parte media de este mismo perfil, fue asignada al Albiano tardío-Cenomaniano (Guler y Archangelsky, 2002). Asimismo, esta última edad fue también propuesta por Aguirre-Urreta (2002), sobre la base del contenido de amonites de depósitos de la Formación Kachaike en el área de lago San Martín. Asociaciones de dinoflagelados, recuperados de la sección inferior del perfil BC1, sugieren una edad Albiano temprano para estos niveles (Guler y Archangelsky, 2006); mientras que el contenido de polen de angiosperma indica una edad Albiano tardío para las porciones media y superior de este perfil (Barreda y Archangelsky, 2006).

Figura 1. Mapa de ubicación de la localidad fosilífera 'Bajo Comisión' / location map of the 'Bajo Comisión' fossiliferous locality.
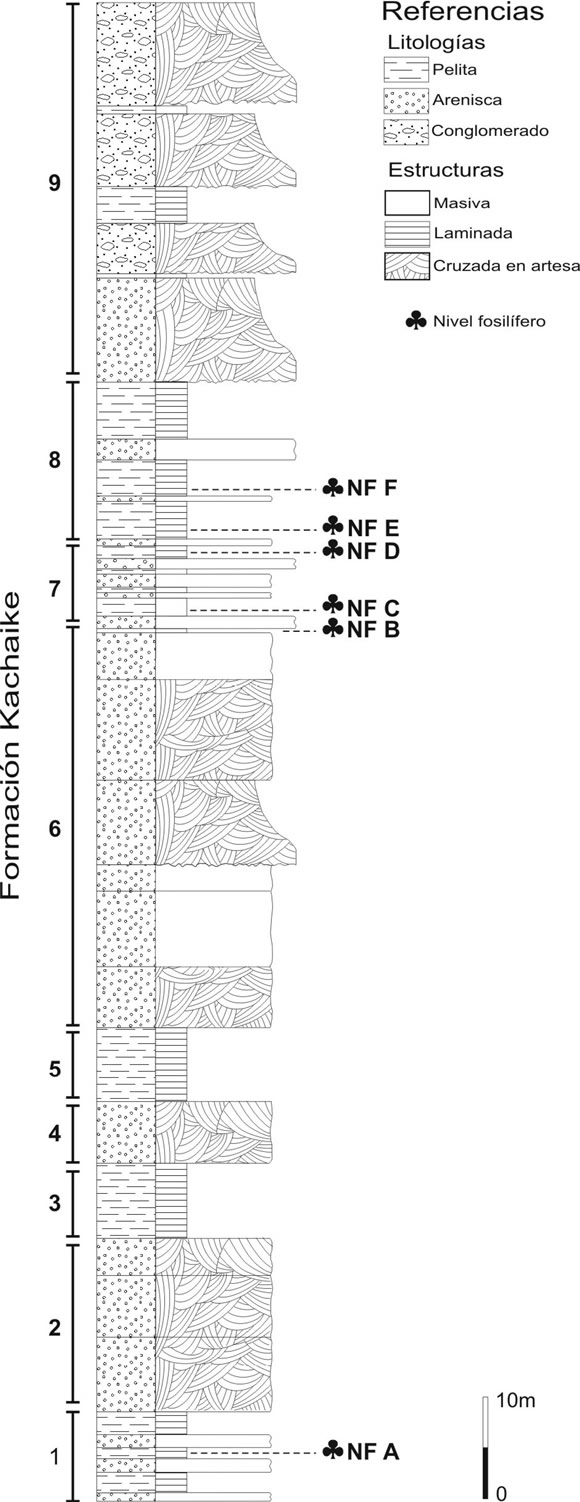
Figura 2. Perfil Bajo Comisión 1 basado en datos inéditos de Cladera y Limarino con niveles fosilíferos (NFA-NFF). 1-9, ambientes sedimentarios; 1, prodelta; 2, frente deltaico; 3, prodelta; 4, frente deltaico; 5, prodelta; 6, frente deltaico hasta plataforma subaérea; 7, plataforma subacuea. 8, plataforma subaérea; 9, fluvial. / stratigraphical section and sedimentary environment of Bajo Comisión 1 based on inedit data of Cladera and Limarino showing megafossil sampling levels (NFA-NFF). 1-9, sedimentary environment; 1, prodelta; 2, delta front; 3, prodelta; 4, delta front; 5, prodelta; 6, delta front to upper delta plain; 7, shelf; 8, upper delta plain; 9, fluvial.
Materiales y métodos
En las descripciones de impresiones foliares de angiospermas se siguió la terminología de Hickey (1973) y, en parte, aquella propuesta por el Leaf Architecture Working Group (1999). Las cutículas fueron maceradas con HNO3 al 40% y HONH3 al 5%, sometidas a ultrasonido y montadas en gelatina/glicerina. Las fotografías fueron tomadas con cámara digital Nikon/Coolpix 990. Para las observaciones con microscopio electrónico de barrido se utilizó un equipo Phillips XL30 del Museo Argentino de Ciencias Naturales Bernardino Rivadavia. Los especímenes fósiles se encuentran depositados en la colección paleobotánica del Museo Argentino de Ciencias Naturales Bernardino Rivadavia (BA Pb y BA Pb Pm)
Paleontología sistemática
División BRYOPHYTA
Clase BRYOPSIDA Rothmaler 1951
Género Muscites Brongniart 1828
Especie tipo. Muscites tournalii Brongniart 1828.
Muscites sp.
Figuras 3.1-3.3
Descripción. Fragmento de talo gametofítico densamente foliado, de 2 cm de largo y ancho máximo de 3 mm; eje ramificado dicotómicamente, al menos, dos veces, con no más de 0,4 mm de grosor (figura 3.1). Filodios elípticos, dispuestos helicoidalmente, insertos con ángulos cercanos a 45º; de 1,4-2,5 mm de largo y 0,8-1,2 mm de ancho máximo (relación largo: ancho 2:1); de margen entero; sésiles; con ápice agudo acuminado. Lámina recorrida por una vena media (figura 3.2). No se observa esporofito.
Comentarios. Si bien se trata de un único fragmento, la suma de características morfológicas reunidas (talo con ramificación dicotómica, forma, tamaño, inserción y vascularización de las hojas) nos permite interpretar este ejemplar como un gametofito de briófita y asignarlo a Muscites Brongniart, morfogénero de amplia definición, que reúne vegetales fósiles con caracteres similares a los de musgos actuales.
Comparaciones.La ausencia de tejidos de resistencia otorga, a las briófitas, un bajo potencial de fosilización por lo cual su registro megascópico es escaso. El único antecedente para el Mesozoico de Patagonia es Marchantites hallei Lundblad (1955), briófita de probable afinidad Hepaticae descripta originalmente (como Marchantites? sp.) por Halle (1913a) como parte de la tafoflora cretácica de Río Fósiles. Muscites antarcticus Cantrill (2000) del Aptiano de Antártida, difiere de nuestro ejemplar por poseer filodios notablemente amplexifolios y recurvados. Drinnan y Chambers (1986) describieron dos gametofitos para el Aptiano de Australia con los cuales los ejemplares de Bajo Comisión podrían compararse. Sin embargo, los caracteres compartidos entre ambos (ángulo de inserción de los filodios, éstos con ápice acuminado y presencia de vena media) son comunes para gametofitos de numerosas especies de musgos.
Procedencia. Bajo Comisión 1 (BC1), nivel fosilífero gNF C.
Material estudiado. BAPb 12481a y 12481c.
División PTERIDOPHYTA
Orden FILICALES Tryon y Tryon 1982
Familia GLEICHENIACEAE (?) (R. Br.) Presl 1825
Género Gleichenites Goeppert 1836
Especie tipo. Gleichenites porsildii Seward 1926.
Gleichenites sanmartini Halle emend. Herbst 1962ª
Figuras 3.4, 9.13
1913a. Gleichenites sanmartini Halle, pág. 22, lám. 1, figs. 14-15
1962a. Gleichenites sanmartini Halle, Herbst, pág. 142, figs. 1-5, 12-13, 16.
Descripción. Segmentos de fronde estéril, al menos, bipinnada; el mayor de 4 cm de largo. Raquis de 1 mm de grosor. Pinnas imparipinadas; lineares; insertas con ángulo cercano a 80º; alternas. Pínnulas pecopterídeas, insertas con ángulos de 70-90º; alternas a subopuestas; de 1-1,5 mm de largo y 1-1,2 mm de ancho máximo (relación largo:ancho 1,5:1 a típicamente 1:1). Vena media recta (levemente decurrente) y dos a tres laterales simples. No se observan pinas fértiles.
Comparaciones. Si bien no se observan pinnas fértiles, por los caracteres vegetativos reunidos, se considera aquí que el material estudiado corresponde a segmentos estériles de Gleichenites sanmartini Halle emend. Herbst.
Esta especie, descripta por Halle (1913a) para Río Fósiles, fue citada, posteriormente, para varias localidades cretácicas de cuenca Austral, presumiblemente coetáneas (Baldoni y Ramos, 1981; Longobucco et al., 1985), o más antiguas (Feruglio, 1951; Herbst, 1962a), como así también para el Jurásico Inferior (Liásico) de cuenca Neuquina (Arrondo y Petriella, 1980 como Gleichenites cf. G. sanmartini). Cabe destacarse, que los ejemplares asignados a esta especie por Baldoni y Ramos (1981) presentan algunas diferencias con los aquí descriptos, como así también con aquellos procedentes del Grupo Baqueró (Feruglio, 1951; Herbst, 1962a) y Río Fósiles (Halle, 1913a). Estas diferencias residen en el ángulo de inserción de las pinnas y pínnulas, y en las dimensiones y venación de estas últimas, con venas laterales bifurcadas. Los ejemplares descriptos por Longobucco et al. (1985) son muy similares a los de Bajo Comisión, salvo, nuevamente, por la presencia de venas laterales bifurcadas.
La especie G. sanmartini fue citada por Césari et al. (1999), para el Aptiano de Antártida, pero luego sinonimizada en la especie Eocyathea remesaliae Césari (2006). Microphyllopterissp. cf. M. gleichenioides (Oldham y Morris) Walkom, descripta por McLoughlin et al. (1995) para el Cenomaniano de Australia, consiste en un segmento estéril al menos bipinnado, con pínnulas cuya forma, tamaño y ángulo de emergencia son semejantes a G. sanmartini Halle, observación ya hecha por los propios autores. La ausencia de venación en los ejemplares australianos no permite una comparación más rigurosa. Procedencia. Bajo Comisión 1 (BC1), niveles fosilíferos NF B y F.
Material estudiado. BAPb 12440 al 12443, 12446, 12448 al 12449 y 12453 al 12457.
Gleichenites juliensis Herbst 1962b
Figuras 3.5-3.9, 9.12
1913a. Gleichenites cf. G. micromerus Heer, Halle, pág. 22, lám. 1, figs. 14-15.
Descripción. Fragmentos de fronde estéril, al menos, bipinnada. Segmento mayor medido de 4,5 cm de largo (figura 3.5). Raquis principal de 1,5 mm a 3 mm de ancho máximo. Pinnas de disposición alterna a subopuestas, insertas al raquis con un ángulo de 70º - 80º; imparipinnadas. Pinnas contiguas separadas por no más de 1-2 mm; ápice no preservado. Pinna mayor medida de aproximadamente 5 cm de largo y 5 mm de ancho máximo. Pínnulas alternas a subalternas, insertas al raquis por toda la base, con ángulo de 60-70º; a razón de 5 pínnulas por cm (en las pinnas mayores); márgenes enteros, paralelos; base catadrómica decurrente; ápice redondeado. Pínnulas contiguas separadas o algo confluentes en la base, pudiendo estar en contacto pero no imbricadas. En algunos ejemplares (BAPb 12465b, 12470) las pínnulas basales se superponen al raquis principal. Las pínnulas presentan dimensiones casi uniformes a lo largo de la pinna, no obstante, próximo al ápice, disminuyen gradualmente su tamaño. Pínnulas más grandes con 2,5 mm de largo y 1,5 mm de ancho en la base. Relación largo-ancho 1,3-2: 1, en pínnulas proximales, a casi 1: 1, en las distales. Pínnulas vascularizadas por una vena media decurrente, bifurcada cerca del ápice, de grosor considerable en la base, el cual disminuye rápidamente. Venas laterales en número máximo de tres a cada lado de la vena media, bifurcándose una vez cerca del origen y alcanzando el margen sin anastomosarse. De la base de la vena media, o algo por encima de este punto, emerge una vena bifurcada que vasculariza la porción catadrómica basal de la pínnula. Estas venas basales catadrómicas pueden presentar una segunda bifurcación de la rama distal (figura 3.9). No se observan pinnas fértiles.
Comentarios. Las pínnulas del NF E son algo más grandes que las del NF C (aunque conservan igual relación largo:ancho) coincidiendo en todos los demás aspectos, razón por la cual son consideradas pertenecientes a la misma especie.
Comparaciones. Coincide en todo aspecto con las frondes de Río Fósiles que Halle (1913a) describió como Gleichenites cf. G. micromerus Heer, por cuanto se trataría de nuevos ejemplares de la misma especie. Halle (1913a: 24-25) encontró cierta similitud entre sus ejemplares de Gleichenites cf. G. micromerus con Gleichenia micromera Heer (1874) de los "Kome bed" (Albiano-Cenomaniano?) de Groenlandia. Sin embargo, tal como señaló Herbst (1962a), los argumentos son inconsistentes, toda vez que esta última posee pínnulas insertas casi perpendicularmente al raquis y venas laterales simples. Más aún, el propio Halle (1913a) arrojó dudas sobre la identidad de la especie de Heer.
Por otro lado, los especímenes aquí descriptos presentan una muy marcada similitud en los caracteres vegetativos con Gleichenites juliensis Herbst (1962b), del Jurásico Medio del Gran Bajo de San Julián. Al respecto, Herbst (1962b) señaló semejanzas y diferencias entre sus ejemplares y los descriptos por Halle (1913a). Una de las diferencias señaladas por Herbst (1962b) consiste en el ángulo de inserción de las pinnas. No obstante, en los ejemplares de Bajo Comisión, dicho ángulo se encuentra muy cercano a los 90º descriptos para G. juliensis. Además, en el ejemplar ilustrado en Herbst (1962b: fig. 4), se observa que las pinnas no son ortogonales al raquis que las porta. Otra diferencia, remarcada por este autor, es respecto a la forma de las pínnulas, argumento que no se comparte en el presente trabajo. La inserción de las pínnulas al raquis es otra diferencia señalada por Herbst (1962b). Al respecto, si bien Halle (1913a) sugirió que podrían estar insertas por un punto, en los ejemplares de Bajo Comisión parecen hacerlo por toda la base tal como ocurre en G. juliensis. Por último, el marcado grosor basal de la vena media en cada pínnula es otro carácter común a los especímenes de ambas localidades. La ausencia de frondes fértiles en los ejemplares de Bajo Comisión como en los descriptos por Halle (1913a) no permite una comparación con los del Gran Bajo de San Julián. Sin embargo, por lo arriba expuesto y habiendo podido revisar el material original descripto por Herbst (1962b) considero que tanto los ejemplares aquí descriptos como los atribuidos por Halle (1913a) a Gleichenites cf. G. micromerus corresponden a Gleichenites juliensis Herbst.
Otra especie afín a G. juliensis, en sus caracteres vegetativos, es Gleichenites argentinica (Berry) emend Herbst (Formación Punta del Barco, Aptiano). No obstante, G. juliensis puede separarse de ésta sobre la base del tamaño de las pínnulas, tal como señaló Herbst (1962a), y por la presencia, en G. argentinica, de venas laterales basales que salen directamente del raquis y se bifurcan casi siempre dos veces (véase Herbst, 1962a: fig. 1a).
Los ejemplares de Bajo Comisión presentan cierta semejanza con segmentos estériles de Gleicheniaceapyllum acutum Nagalingum y Cantrill (2006) y Microphyllopteris unisora Cantrill y Nagalingum (2005), del Cretácico (Albiano tardío) de Antártida. No obstante, G. acutum se distingue por sus pínnulas rómbicas con ápice agudo; en tanto, M. unisora presentaría pínnulas con cierta anastomosis (Cantrill y Nagalingum, 2005), carácter que lo diferenciaría de G. juliensis. Gleichenites gleichenoides (Oldham y Morris) emend. Seward y Sahni (en Surange, 1966: 100) del Jurásico de Rajmahal (India) es otra especie afín a G. juliensis aunque se diferencia por sus venas secundarias generalmente no bifurcadas.
Procedencia. Bajo Comisión 1 (BC1), niveles fosilíferos NF C y E.
Material estudiado. BAPb 12463, 12464, 12465a y b, 12467, 12470 al 12473, 12475, 12490 al 12492 (NF C), BAPb 12429a-d (NF E).

Figura 3. 1-3, Muscites sp. BAPb 12481. 1 y 3, aspecto general del gametófíto / general view of gametophyte. Las flechas indican sitios de ramificación dicotómica / arrows indicate dichotomous branched sites. 2, detalle de hojas vascularizadas por una vena media (flecha) / detail of leaves innerved by a middle vein (arrow). 4, Gleichenites sanmartini Halle emend. Herbst. BAPb 12449; 5-9, Gleichenites juliensis Herbst; 5, BAPb 12473; 6, BAPb 12429; 7, BAPb 12491; 8, detalle pinnas / detail of pinnae. BAPb 12463. 9, detalle de pínnula / detail of pinnule. BAPb 12464. Escala / scale 2 = 0,2 cm; 3, 4, 9 = 0,5 cm; 1, 5-8 = 1 cm.
Familia MATONIACEAE (?) Presl 1848
Género Nathorstia Heer 1880
Especie tipo. Nathorstia angustifolia Heer 1880.
Nathorstia alata Halle 1913a
Figuras 4.1-4.3
1913a. Nathorstia alata Halle, pág. 20, lám.1, figs. 1-9.
1938. Nathorstia alata Halle, Piátnitzky, pág. 81, lám. 10, fig. 40.
Descripción. Segmentos de fronde pinnada. El mayor conservado mide alrededor de 12 cm de largo. Raquis de 5 mm de ancho. Pínnulas opuestas a subopuestas, insertas lateralmente (en ángulos cercanos a los 80-90º), espaciadas, no superpuestas, lineares, de 4-5 mm (a 6 mm en BAPb 259) de ancho y hasta 15 cm de largo. Margen entero suavemente revoluto y textura cartácea a coriácea. Base algo constricta; lámina angostándose muy progresivamente en dirección apical, ápice agudo (figura 4.1). Vena media fuerte, con 0,5-0,6 mm de grosor, venas laterales insertas con ángulo de 80-90º y 0,1 mm de grosor, a razón de 6-7 venas por cm. Segmentos fértiles morfológicamente idénticos a los estériles. Presentan, a cada lado de la vena media, una hilera de soros circulares de 0,7-1,5 mm de diámetro y a razón de 8-9 por cm (figura 4.3); esporangios numerosos en forma de cuña. No se observa evidencia de indusio.
Comentarios. En Matoniaceae actuales, el indusio frecuentemente se desprende de la cara abaxial de las pínnulas en forma total o parcial (Klavins et al., 2004). Por tal razón, la no preservación de un indusio en los fósiles de Bajo Comisión, como en los descriptos por Halle (1913a) y Piátnitzky (1938), no necesariamente es conclusiva respecto a este carácter.
Comparaciones. Esta especie fue citada por Halle (1913a) para Río Fósiles, y por Piátnitzky (1938) para depósitos de la Formación Piedra Clavada en proximidades de lago Cardiel, provincia de Santa Cruz. Krassilov (1964) sinonimizó a N. alata Halle, en una nueva combinación: Nathorstia pectinata (Goeppert) Krassilov. No obstante, N. alata presenta pínnulas más largas y menor número de esporangios por soro. Además, Van Konijnenburg-Van Cittert (1993) encontró pequeñas diferencias entre las esporas de Nathorstia pectinata y N. alata revalidando la identidad específica de los ejemplares patagónicos. Otras especies fósiles semejantes a N. alata son: Phlebopteris sp. (Surange, 1966: 113, fig. 72) del Jurásico de Rajmahal (India); Matonidium brownii var. magnipinnulum Rushforth (1970) del Cretácico Inferior de Estados Unidos; Phlebopteris(?) elongata Bell (1956) del Cretácico Inferior (Neocomiano-Barremiano) de Canadá, aunque todas ellas presentan pínnulas notablemente más cortas. Matonia jeffersonii Nagalingum y Cantrill (2006) del Albiano tardío de Antártida (Formación Triton Point), presenta pínnulas alargadas semejantes a las de N. alata. No obstante, Nagalingum y Cantrill (2006) destacan, en M. jeffersonii, la presencia de indusio y venas secundarias no ramificadas como caracteres distintivos de esta especie.
Procedencia. Bajo Comisión (BC1), nivel fosilífero NF F.
Material estudiado. BAPb 245, 258, 259.
Familia SCHIZAEACEAE (?) Martius 1834
Género Mohriopsis Appert 1973
Especie tipo. Mohriopsis plastica Appert 1973.
cf. Mohriopsis plastica Appert 1973
Figura 4.8
Descripción. Fragmento de fronde, al menos, bipinnada. Segmento mayor medido de 2 cm de largo. Raquis liso, de 1 mm de grosor. Pinnas alternas, insertas en ángulo de 45º, de forma triangular; la más larga medida de 1,3 cm de longitud. Pínnulas alternas, insertas en ángulo de 45º, triangulares, lobuladas, con base constricta y ápice redondeado; de 0,5 cm de largo máximo y 0,3 cm de ancho medido en la base. Pínnulas contiguas en contacto, las que, en dirección al ápice de la pinna, disminuyen rápidamente su tamaño y confluyen. Venación mal preservada, sólo se observa una vena media vascularizando cada pínnula.
Comentarios. El material es escaso y fragmentario. Aparece en niveles ampliamente dominados por segmentos estériles de cf. Sphenopteris sp. "f" Douglas. Comparaciones. Por su forma general y dimensiones presenta similitud con Mohriopsis plastica Appert (Schizaeaceae), del Jurásico Superior de Madagascar (Appert, 1973, lám. 2, figs. 1-2). Sin embargo, por presentar un raquis algo más grueso y, por la ausencia de otros caracteres diagnósticos del género (en particular, de estructuras fértiles), su asignación se hace con reservas. Cabe destacar, por otro lado, cierto parecido, en el aspecto vegetativo, con la fronde fértil de Amanda floribunda Douglas (1973, lám. 39 figs. 1-3). Procedencia. Bajo Comisión (BC1), nivel fosilífero NF E.
Material estudiado. BAPb 13466, BAPb 13467, BAPb 13469, BAPb 13456, BAPb 13441, BAPb 13457, BAPb 13463, BAPb 13432.
Familia OSMUNDACEAE (?) Brown 1810
Género Phyllopteroides Medwell 1954
Especie tipo. Phyllopteroides dentata (Medwell) Cantrill y Webb 1987.
Phyllopteroides cf. P. laevis Cantrill y Webb 1987
Figuras 4.9-4.10
1913a. Asplenites lanceolatus Halle, pág. 33, lám. 4, figs. 11-21.
Descripción. Pínnula de forma oblonga u oval lanceolada; de 3 cm de largo y 1 cm de ancho máximo; carente de extremo apical; base aguda, levemente cuneada; margen finamente serrado (correspondiendo cada diente a la terminación de una vena lateral), algo ondulado. Pecíolo presente, aparentemente corto (conservado hasta 1 mm de su longitud); textura cartácea. Sección transversal en forma de "V" abierta. Vena media de recorrido recto y con 0,5 mm de grosor el cual persiste, al menos, hasta el extremo apical conservado; venas laterales con emergencia decurrente (en ángulos próximos a 30º) y recurvadas; dicotomizadas hasta dos veces con sus ramificaciones paralelas y alcanzando el margen (en ángulos entre 55-65º) sin anastomosarse. Venas laterales en número de 20 venas por cm (medido sobre el margen), con grosor no mayor a 0,1-0,2 mm.
Comparaciones. Aunque carente de su extremo apical, no hay dudas de que se trata de la misma especie descripta por Halle (1913a, lám. 4, figs. 11-21) para Río Fósiles como Asplenites lanceolatus Halle. No encontrando un organogénero dentro del cual incluir sus ejemplares, Halle (1913a) sugirió el uso provisorio de Asplenites Goeppert, no por su parecido con la especie tipo, sino por su similitud con especies del género actual Asplenium L. Medwell (1954) estableció el género Phyllopteroides para hojas del Cretácico Inferior de Victoria (Australia), corrigiendo un antiguo problema de nomenclatura (para detalles véase Cantrill y Webb, 1987). Los ejemplares de Río Fósiles y de Bajo Comisión presentan características morfológicas concordantes con Phyllopteroides. Por ello, se considera que el ejemplar aquí descripto y los de Halle (1913a) (como Asplenites lanceolatus Halle) tienen mejor ubicación en este organogénero.
El género Phyllopteroides Medwell incluye seis especies definidas para el Cretácico (Neocomiano -Cenomaniano) de Australia: P. dentata (Medwell) Cantrill y Webb (1987), P. lanceolata (Walkom) Medwell (1954), P. serrata Cantrill y Webb (1987), P. laevis Cantrill y Webb (1987), P. macclymontae McLoughlin et al. (1995) y P. westralensis McLoughlin (1996); y una para el Cretácico (Albiano tardío) de Antártida: P. antarctica Cantrill y Nagalingum (2005). Los ejemplares de Río Fósiles y Bajo Comisión presentan semejanzas tanto con P. antarctica como con P. leavis, dos especies que a su vez son muy afines entre sí. Según Cantrill y Nagalingum (2005), ambos taxones se distinguen por la forma de las pínnulas y el ángulo de emergencia de las venas secundarias. Sin embargo, en los ejemplares patagónicos ambos caracteres son intermedios entre una y otra forma, en tanto el tipo de margen, presencia de una única vena por diente y densidad de venas son caracteres compartidos con ambas especies. No obstante la proximidad geográfica y edad de la especie antártica, por la presencia de venas laterales dicotomizadas hasta dos veces en P. laevis (Cantrill y Nagalingum, 2005 señalan hasta tres dicotomías en P. antarctica), el ejemplar de Bajo Comisión y los descriptos por Halle (como Asplenites lanceolatus Halle) son aquí referidos tentativamente como Phyllopteroides cf. P. laevis Cantrill y Webb sin descartar por ello que puedan ser afínes a la especie antártica. Una mayor cantidad de material es requerida con el objeto de determinar si efectivamente los ejemplares patagónicos pueden ser incluidos en uno u otro taxón o bien constituyen una entidad específica distinta.
P. laevis ha sido también reconocida para el Cretácico Inferior de India (Banerji, 1992; 1996). McLoughlin et al. (2002: 19) sugirieron la posibilidad de incluir bajo el género Phyllopteroides a los restos foliares descriptos como cf. Sagenopteris paucifolia (Phillips) Ward por Césari et al. (1999) para el Aptiano de Antártida. Sin embargo, la presencia de anastomosis en los ejemplares antárticos no se corresponde con la diagnosis del primero. La abundancia y riqueza de este género para el Cretácico Inferior de Australia hace suponer que no sería extraño que su límite de distribución durante el Cretácico alcance, al menos, el sur de India por un lado, y la Patagonia austral, por otro.
En cuanto a la afinidad botánica de Phyllopteroides ha sido primero clasificado como pteridosperma por Douglas (1969), pero luego Cantrill y Webb (1987) sugirieron su relación con las Osmundaceae basados en una asociación de frondes estériles de P. dentata Medwell con restos de esporangios (Caecumen expansa Cantrill y Webb) que pertenecerían a dicha familia.
Procedencia. Bajo Comisión 2 (BC2).
Material estudiado. BAPb 12521.
Incertae Sedis
Género Cladophlebis Brongniart 1849
Especie tipo. Cladophlebis albertsii (Dunker) Brongniart 1849.
Cladophlebis patagonica Frenguelli emend. Herbst 1966ª
Figura 4.4
Descripción. Fragmentos de pinnas, el mayor de 2,5 cm de largo; ancho máximo medido de la pinna de 13 mm. Raquis 1 mm de ancho. Pínnulas opuestas a subopuestas; insertas por toda su base con ángulos de 45-57º; simétricas, con márgenes enteros, rectos, culminando en ápice agudo. Pínnulas de 7-9 mm de largo y 2,5-3 mm de ancho máximo; relación largo:ancho 2,5-3,5: 1. La vena media es decurrente y vasculariza la pínnula con recorrido recto. Venas laterales mal preservadas, emergiendo en ángulo agudo; en número, aparentemente, no mayor a 10; dicotomizadas una vez, sin anastomosarse.
Comentarios. Los registros de C. patagonica (Frenguelli) Herbst se extienden a tafofloras de distintas localidades de Patagonia, cuyas edadades incluyen el Jurásico Medio tardío (Feruglio, 1951: 66; Herbst y Zalazar, 1998); Aptiano (Feruglio, 1951: 52; Herbst, 1966a) y Cretácico Inferior (Halle, 1913a; Longobucco et al., 1985; este trabajo).
Procedencia. Bajo Comisión (BC1), nivel fosilífero NF F.
Material estudiado. BAPb 12420 a (y su contraparte 12420a).
Género Aculea Douglas 1973
Especie tipo. Aculea bifida Douglas 1973.
Aculea bifida Douglas 1973
Figura 4.11
Descripción. Segmento fértil, el mayor conservado de 3,5 cm de longitud, pinnas aciculares, algo flexuosas; varias veces bifurcadas; con grosor no superior a 1-1,2 mm y disminuyendo a 0,5 mm en las últimas ramificaciones; ápice de las pínnulas expandido culminando en 1-3 soros terminales reniformes. Comparaciones. Segmentos con pínnulas profundamente disectas, lineares son característicos del Cretácico Inferior del sur de Australia y han sido asignadas a los géneros Aculea Douglas, Alamatus Douglas y Amanda Douglas (Douglas, 1973) básicamente por la posición diferencial de sus soros. Entre ellas, sólo Aculea presenta soros terminales semejantes al material aquí descripto. El registro fósil de Aculea bifida Douglas incluye el Neocomiano-Aptiano (Douglas, 1973; Drinnan y Chambers, 1986) y Neocomiano-Barremiano (McLoughlin, 1996) en cuencas del este y oeste de Australia, respectivamente, y Aptiano de Antártida (Cantrill, 2000). Su afinidad botánica es incierta. No obstante, por la posición de los soros y división de su fronde se ha señalado su similitud con miembros actuales de la familia Hymenophyllaceae (Drinnan y Chambers, 1986; McLoughlin et al., 2002) y Davalliaceae (Drinnan y Chambers, 1986).
Procedencia. Bajo Comisión 2 (BC2).
Material estudiado. BAPb 12518a y b; 12519.
Género Sphenopteris Sternberg 1825
Especie tipo. Sphenopteris elegans (Brongniart) Sternberg 1825.
cf. Sphenopteris sp. "f" Douglas 1973
Figuras 4.5-4.7, 9.16
Descripción. Fragmentos de frondes estériles, al menos, bipinnadas. Pinna primaria más larga (BAPb13460) de 7-8 cm de longitud. Lámina con 2-2,5 mm de ancho máximo, medido por sobre el punto de división de la misma. La pinna primaria se divide pseudomonopodialmente originando pinnas secundarias dispuestas alternadamente e insertas con ángulos de 10-20º respecto al raquis primario. Pinna secundaria más larga medida de 2-3 cm (BAPb 13492, figuras 4.5-4.7). Las pinnas secundarias se dividen pseudodicotómicamente, con ángulos que no superan los 10º, originando pínulas o "proyecciones foliares" de margen entero y ápice agudo a levemente redondeado y que, en general, no superan los 2 mm de largo y 0,75 mm de ancho (hasta 4 mm de largo y 1 mm de ancho en piezas BAPb 13466 y BAPb 13467). Las venas son muy delgadas y sin diferencia apreciable de grosor entre las que se hallan en la porción proximal y las de la porción distal de una pinna secundaria. Se dicotomizan, culminando en el ápice de las pínnulas. Número máximo de venas 4-6 en la pina primaria a 1 en cada pínnula.
Comentarios. Esta especie es muy abundante, especialmente en el nivel NF E, en el cual parece formar intercalaciones de horizontes monotípicos. Este tipo de cubiertas, dominadas numéricamente por hojas de helechos esfenopterídeos, es observado por Cantrill (1996) en paleosuelos del Cretácico (Albiano tardío) de Antártida y, en virtud de su abundancia, sugiere que podrían haber formado verdaderos matorrales.
Comparaciones. Siguiendo el criterio de otros autores, se opta aquí por incluir, dentro de Sphenopteris Brongniart, los ejemplares de Bajo Comisión, reservando el uso de otras denominaciones genéricas (Alamatus Douglas, Aculea Douglas, Amanda Douglas, Onychiopsis Yokoyama, entre otros) exclusivamente para frondes fértiles.
La mayor similitud, en cuanto al desarrollo y grado de disección de la fronde, se encuentra con Sphenopteris sp. "f" Douglas (1973) del Cretácico Inferior (Neocomiano) de Victoria (Australia); ejemplares que, en una reciente revisión, fueron referidos como Aculea sp. A (McLoughlin et al., 2002).
Frondes esfenopterídeas fueron descriptas por Halle (1913a) para la tafoflora de Río Fósiles. Las mismas fueron remitidas a entidades ya conocidas (Sphenopteris (Onychiopsis?) psilotoides (Stokes y Webb) Ward?; Sphenopteris (Ruffordia?) goepperti Dunker y Sphenopteris cf. naktongensis Yabe) o a una nueva especie (Sphenopteris patagonica Halle). Entre ellas, es comparable el ejemplar ilustrado por Halle (1913a, lám. 2, fig. 8) y que es asignado con dudas a Sphenopteris (Ruffordia?) goepperti Dunker. Esta especie fue también reconocida (como Sphenopteris cf. goepperti Dunker) por Archangelsky (1964) en depósitos aptianos de la Formación Anfiteatro de Ticó. Si bien presentan cierto parecido, los ejemplares de ambas localidades pueden ser distinguidos del material aquí analizado, por sus pínnulas con lámina algo más expandida. Sphenopteris (Onychiopsis?) psilotoides (Stokes y Webb) Ward es otra especie en común entre Río Fósiles (Halle, 1913a) y Anfiteatro de Ticó (Archangelsky, 1967a). Si bien podría parecer semejante a los ejemplares de Bajo Comisión existe cierta diferencia respecto al plan de división de la fronde, más claramente pinnado en Sphenopteris (Onychiopsis?) psilotoides.
Para el Cretácico de Antártida han sido reconocidas frondes comparables a las de Bajo Comisión. Tal es el caso de Aculea bifida Douglas y Aculea acicularis Cantrill. La primera de ellas fue descripta por Cantrill (2000) a partir de frondes fértiles pertenecientes a la Formación Cerro Negro (Aptiano). Esta especie presenta, además, un amplio registro para el Cretácico Inferior (Neocomiano) de Australia (Douglas, 1973; Drinnan y Chambers, 1986; McLoughlin, 1996; McLoughlin et al., 2002). Aculea acicularis Cantrill (1996), en tanto, fue definida a partir de segmentos estériles pertenecientes a la Formación Triton Point (Albiano tardío). Ambas especies, sin embargo, presentan una lámina más angosta que la de los ejemplares de Bajo Comisión. Procedentes de la Formación Triton Point, Cantrill y Nagalingum (2005) describieron fragmentos aislados de frondes estériles con dos formas diferentes, una con lámina reducida y otra con lámina expandida que, no obstante, atribuyeron a una única especie: Sphenopteris warragulensis McCoy. La misma ha sido ampliamente reconocida para el Neocomiano-Cenomaniano de Australia (e.g. Drinnan y Chambers, 1986; McLoughlin et al., 1995; McLoughlin, 1996). Si bien los ejemplares de Bajo Comisión no presentan mayores semejanzas con los espécimenes australianos, es destacable su similitud con las formas antárticas de lámina reducida descriptas por Cantrill y Nagalingum (2005: lám. 4, figs. B y C).
Procedencia. Bajo Comisión (BC1), niveles fosilíferos NF B, C y E.
Material estudiado. BAPb 12441, 12447, 12452, 12461, 12462b (NF B), BAPb 12473a, 12476, 12490 (NF C), BAPb 12435, 13421, 13450, 13454, 13460, 13461, 13462, 13463 (y su contra 13464), 13466, 13467, 13470 (y su contraparte 13471), 13479, 13492 (NF E).

Figura 4. 1-3, Nathorstia alata Halle. 1, aspecto general de pinnas estériles (flecha blanca) y pinnas fértiles (flecha negra) / general view of sterile pinnae (white arrow) and fertile pinnae (black arrow). BAPb 245 2, detalle de pinna fértil / detail of fertil pinnae. BAPb 245 3, detalle de pínnula / detail of pinnules. BAPb 245. 4, Cladophlebis patagonica Frenguelli emend. Herbst. BAPb 12420. 5-7, cf. Sphenopteris sp. "f" Douglas. 5-6, BAPb 13492. 7, BAPb 12435. 8, cf. Mohriopsis plastica Appert. BAPb 13467. 9-10, Phyllopteroides cf. P. laevis Cantrill y Webb. BAPb 12521. 9, aspecto general / general view. 10, detalle margen, las flechas indican sitios de dicotomia de venas laterales / margin detail, arrows indicate lateral vein dichotomous branched sites. 11, Aculea bifida Douglas. BAPb 12518. Escala / scale 2 = 0,1 cm; 3, 4, 8, 11 = 0,5 cm; 1, 5-7, 9 = 1 cm. 1913a. Cladophlebis australis (Mor.) Sew, Halle, pág. 25, lám. 1, figs. 10-13. 1913a. Cladophlebis cf. browniana (Dunk.) Sew, Halle, pág. 27, lám. 4, figs. 6-7. Para una sinonimia completa véase Herbst (1971).
Sphenopteris sp.
Figura 5.7
Descripción. Fragmentos de pínnulas pequeñas, de no más de 6-7 mm de largo; flabeladas; de base cuneada y ápice consistente de lóbulos redondeados. Venas paralelas con dicotomías de bajo ángulo culminando en los lóbulos apicales.
Comparaciones. Si bien el material es muy escaso vale señalar algunas semejanzas. Presenta parecido, en su aspecto general, con las pínnulas de Sphenopteris cf. naktongensis Halle (1913a: lám. 4, fig. 9) de Río Fósiles. También es similar en la forma y venación de las pínnulas con Ruffordia goepperti (Dunker) Watson ilustrada por Watson (1969: lám. 4, figs. 3, 4 y 7) y Ruffordia göpperti (Dunk.) var. latifolia Seward (1894: lám. 6, figs. 1-1) del Cretácico Inferior de Inglaterra.
Procedencia. Bajo Comisión (BC1), nivel fosilífero NF D.
Material estudiado. BAPb 12463a y b, 12464, 12465.
División PTERIDOSPERMOPHYTA
Orden CORYSTOSPERMALES Sternberg 1820
Género Pachypteris Brongniart emend. Harris 1964
Especie tipo. Pachypteris lanceolata Brongniart 1828.
Pachypteris crassa (Halle) Townrow 1965
Figuras 5.1-5.2, 9.14
Descripción. Fragmentos de fronde estéril, al menos, bipinnada. Segmento mayor medido de 3 cm de largo (figura 5.1). Raquis primario de 1 mm de ancho máximo y secundario de no más de 0,5 mm, este último finamente alado. Pinnas de disposición alterna u opuesta insertas al raquis con un ángulo de 45-60º; imparipinnadas. Pinnas contiguas no superpuestas; rectas o algo curvas en dirección al ápice. Pinnas más largas encontradas de 1-1,2 cm a 1,5 cm. Lámina pinatisecta (algo menos disecta apicalmente) originando 4-6 pares de pínnulas opuestas a subopuestas, además de una pínnula apical; insertas con un ángulo de 45º; de forma romboidal (sobre todo las basales) a elípticas, porción basal acroscópica fuertemente contraída y basiscópica algo contraída y decurrente; ápice agudo; márgenes enteros o lobados (especialmente en las pínnulas basales). Pínnulas contiguas separadas a algo confluentes apicalmente. Pínnulas disminuyen su tamaño hacia el ápice, dando a la pina un aspecto triangular a romboidal. Pínnulas basales de 3-4 mm de largo y 1,5-2 mm de ancho máximo y último par apical de 2 mm de largo y 1 mm de ancho. Relación largo:ancho cercana a 2: 1. Pínnula terminal de tamaño mayor o similar a las basales. Pínnulas vascularizadas por una vena media decurrente, dividida pseudodicotómicamente, originando 3-4? pares de laterales que emergen con ángulos de 45º y alcanzan el margen sin dicotomizarse o haciéndolo una vez. La vena media, de grosor similar al de las laterales, se bifurca próxima al ápice.
Comparaciones. Los caracteres fisonómicos foliares de la diagnosis de Pachypteris Brongniart enmendada por Harris (1964) se reconocen en los ejemplares de Bajo Comisión y permiten incluirlos en este morfogénero. La única salvedad reside en que en Pachypteris (Brongniart) Harris, la porción basiscópica de las pínnulas basales se desarrolla junto al raquis principal o bien directamente sobre el raquis entre las pinnas. Este carácter, contemplado en la diagnosis enmendada por Harris (1964) y observado en Pachypteris papilosa (Thomas y Bose) Harris, Pachypteris lanceolata Brongniart del Jurásico de Yorkshire (especie tipo del género) y Pachypteris elegans Archangelsky del Cretácico de Baqueró, no se encuentra presente en los ejemplares de Bajo Comisión o bien es apenas incipiente. Sin embargo, otras especies asignadas al mismo morfogénero (Halle, 1913b; Hernández y Azcárate, 1971; Gee, 1989; Césari et al., 1999) compartirían esta misma salvedad.
Pachypteris crassa (Halle) Townrow fue reconocida por otros autores en floras del Aptiano de Antártida (Hernández y Azcárate, 1971; Césari et al., 1999). En particular, los ejemplares de Bajo Comisión presentan gran similitud con los descriptos por Césari et al. (1999), aunque éstos no preservan detalles de venación, más allá de la presencia de una vena media hasta el tercio distal de la pínnula. Esta misma venación presentan los ejemplares descriptos por Gee (1989) para el Jurásico Superior/Cretácico Inferior de Antártida, las cuales fueron originalmente citadas por Halle (1913b) como Scleropteris crassa Halle. Para el Cretácico Inferior (Aptiano) de Patagonia fueron citadas Pachypteris (?) patagonica Feruglio (1951) y P. elegans Archangelsky (1966), esta última con un raquis de mayor grosor y pínnulas algo menos separadas, no obstante lo cual, es bastante similar a los ejemplares de Bajo Comisión.
Ticoa magallanica Archangelsky (1976), del Cretácico Inferior de la cuenca Magallánica de Chile, también presenta gran similitud en cuanto a la forma de la fronde. La ausencia de cutícula en los ejemplares de Bajo Comisión y de venación en los de Magallanes no permiten una comparación más exhaustiva. No obstante, como fue señalado por Archangelsky (1976), no ha de descartarse que sean especies afines. Finalmente, frondes de aspecto similar han sido citadas para el Jurásico Inferior (Liásico), como Scleropteris vincei Herbst (Herbst, 1966b; Arrondo y Petriella, 1980), y Jurásico Medio tardío (Caloviano), como Scleropteris lotenaense Baldoni (1980), todas ellas en la cuenca Neuquina. La mayor diferencia entre éstas y los ejemplares de Bajo Comisión reside en la forma de las pínnulas (algo más redondeadas en los ejemplares de Scleropteris), a la vez que en su disposición (algo superpuesta en S. vincei Herbst).
Procedencia. Bajo Comisión 1 (BC1), nivel fosilífero NF C.
Material estudiado. BAPb 12481a, 12482 al 12486.
Pachypteris auriculata sp. nov.
Figuras 5.3-5.6, 9.15
Holotipo. BAPb 12487.
Diagnosis. Sterile fronds, at least, bipinnate. Rachis with longitudinal striation. Pinnae inserted at wide angles (80º-90º), imparipinnate. Adjacent pinnae not superimposed. Pinnules numerous, opposite to sub-opposite; inserted at wide angles (near 90º). Adjacent pinnules scarcely spaced, inserted to rachis by contracted bases. Pinnules with one basal pair of rounded isodiametric auricles (absent or little differentiated in the apical pinnules) separated from the rest of the pinnule by a marked incision. Pinnular margins parallel, entires but with slight lobes and rounded apex. Venation open; each pinnule supplied by a middle vein of straight course, dichotomous once near apex; and lateral veins diverging with acute angles (45-50º) and forked dichotomously (once or twice) or trichotomously.
Etimología. El epíteto específico auriculata deriva del latín y alude a la presencia de aurículas en la base de las pínnulas.
Descripción. Fragmentos de fronde estéril, al menos bipinnada imparipinnada. Raquis de aspecto flexuoso, con estrías longitudinales (figura 5.4). Raquis primario de 2 mm de ancho y raquis secundario de no más de 1 mm de ancho máximo. Pinnas insertas al raquis con un ángulo de 80º-90º, imparipinadas de, al menos, 6 cm de largo y 2,5 cm de ancho máximo (figuras 5.3, 5.6). Pinnas contiguas no superpuestas. Pínnulas opuestas a subopuestas (aunque su filotaxis parece ser helicoidal); insertas con ángulos próximos a 90º, en número de, al menos, 14-15 pínnulas en las pinas mayores. Pínnulas contiguas separadas, pueden estar en contacto pero no imbricadas. Pínnulas más grandes de 1,4-1,5 cm de largo y 4 mm de ancho máximo; su tamaño se mantiene constante y disminuye rápidamente próximo al ápice de la pinna. Pínnulas de la porción basal y media de la pina insertas al raquis por medio de una base constricta. Presentan, en el primer quinto de su longitud total, una constricción de la lámina que llega casi hasta la vena media, generando un par de aurículas basales isodiamétricas redondeadas. El resto de la lámina se continúa con márgenes paralelos, enteros, con lóbulos apenas marcados (producto de muy suaves constricciones de la lámina) culminando en un ápice redondeado. En las pínnulas apicales las aurículas basales están ausentes o muy poco diferenciadas. Pínnula apical terminal de forma triangular. Pínnulas vascularizadas por una vena media que emerge con muy leve decurrencia; de grosor casi constante y recorrido recto, bifurcándose muy próximo al margen. Venas laterales apenas preservadas, aparentemente, no más de seis pares, que emergen con un ángulo de 45-50º, dicotomizándose rápidamente una o dos veces (esta última muy próxima al margen); también se ha observado una vena lateral dividida tricotómicamente.
Comparaciones. En la literatura disponible del Cretácico del sur de Gondwana no se han hallado formas en todo semejantes. Frondes similares, aunque claramente diferenciables, corresponden a pteridospermas mesozoicas del hemisferio norte. Por ejemplo, Pachypteris bendukidzei Doludenko y Svanidze (en Boureau y Doubinger, 1975: 679), del Jurásico Medio a Superior de Georgia, y Pachypteris papillosa (Thomas y Bose) emend. Harris, corystospermáceas del Jurásico de Yorkshire. Entre los ejemplares de Pachypteris papillosa, Harris señaló la fuerte lobación de uno de ellos (en Harris, 1964, fig. 51 H-I), el cual es comparable a los de Bajo Comisión. Sin embargo, el par de aurículas bien diferenciadas en la base de las pínnulas es un carácter distintivo de los especímenes patagónicos, que sumado a otras características, justifica su asignación a una nueva especie. En cuanto a su posición sistemática, no obstante la ausencia de estructuras fértiles y cutícula, por su textura cartácea a coriácea, la presencia de un raquis estriado y la posible emergencia helicoidal de las pínnulas (véase figuras 5.4-5.5) posiblemente corresponda a una pteridosperma.
Procedencia. Bajo Comisión 1 (BC1), nivel fosilífero NF C.
Material estudiado. BAPb 12477 al 12480, 12481a (y su contraparte 12481d), 12487a y b.
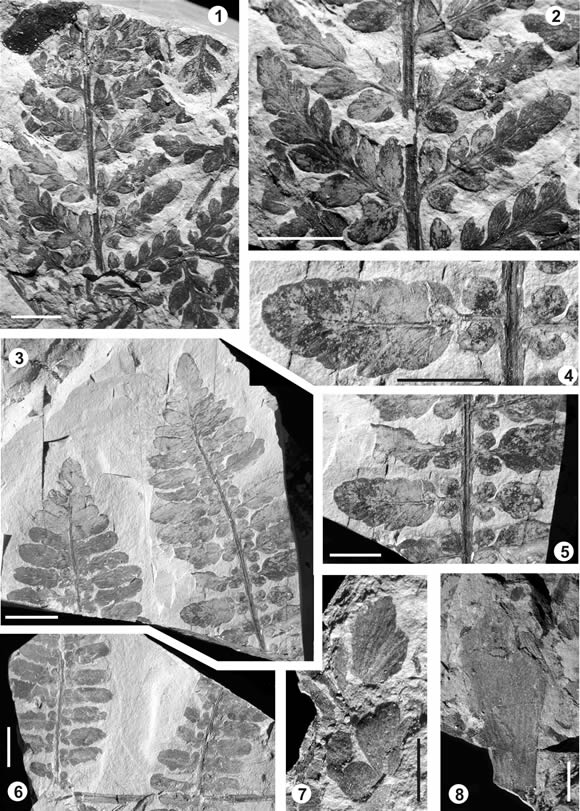
Figura 5. 1-2, Pachypteris crassa (Halle) Townrow. BAPb 12481; 1, aspecto general / general view; 2, detalle de pinnas / detail of pinnae; 3-6, Pachypteris auriculata sp. nov. 3, aspecto general / general view; BAPb 12487. 4, detalle de pinnas / detail of pinnae; BAPb 12487. 5, detalle de pínnula / detail of pinnules; BAPb 12487. 6, aspecto general / general view; BAPb 12480. 7, Sphenopteris sp., detalle de pínnula / detail of pinnules; BAPb 12873. 8, Cycadolepis sp., espécimen incompleto / incomplete specimen; BAPb 12507. Escala / scale 9 = 0,25 cm; 1, 2, 4, 5, 7, 8 = 0,5 cm; 3, 6 = 1 cm.
División CYCADOPHYTA
Orden BENNETTITALES sensu Taylor y Talor 1993
Género Ptilophyllum Morris 1840
Especie tipo. Ptilophyllum acutifolium (Morris) Bose y Kasat 1972.
Ptilophyllum acutifolium (Morris) Bose y Kasat 1972
Figuras 6.1-6.7
Descripción. Fragmentos de hojas monopinnadas, el mayor de 6 cm de largo (incompleto con extremos apical y basal no conservados) y 5 cm de ancho máximo. Raquis de 1-2 mm de ancho. Folíolos insertos a la cara superior del raquis, cubriéndolo, con ángulos de entre 60-80º, alternos a subalternos. Folíolos contiguos en contacto o con una separación mínima, no superior a 1 mm. Folíolos de la porción media de la hoja de 1,5-2,5 cm de largo y 0,25-0,33 cm de ancho máximo (relación largo: ancho típicamente 7-10:1, pudiendo ser incluso 5:1); lineales, de recorrido recto; base recta, contraída en su parte anadrómica y decurrente en la catadrómica, con márgenes paralelos, angostándose recién en la porción distal de la pínnula y culminando en un ápice agudo y falcado (especialmente en BAPb 12506, figura 6.1). La longitud de los folíolos disminuye en dirección basal y apical, siendo de 7 mm los menores conservados en ambos extremos (basal BAPb 12500, figura 6.4 y apical BAPb 12498, figura 6.5). Venación escasamente preservada; venas paralelas, en número de 9-11 (esto es, una concentración de alrededor de 30-40 por cm) (figura 6.6). Se obtuvieron fragmentos de cutícula. Ambas caras poseen células epidérmicas rectangulares, con alrededor de 30-40 x 80-100 µm, con paredes anticlinales muy sinuosas y periclinales lisas (figuras 6.2-6.3). No se pudieron observar aparatos estomáticos.
Comentarios. La cutícula de los ejemplares de Bajo Comisión muestra las células epidérmicas con las paredes anticlinales muy sinuosas características de las Bennettitales. Sin embargo, no se han podido observar aparatos estomáticos. Por esta razón, coincidiendo con Césari et al. (1999: 14), se sigue el criterio de Bose y Kasat (1972) de utilizar la especie P. acutifolium Morris para ejemplares preservados sólo como impresiones.
Comparaciones. P. acutifolium Morris ha sido descripta, por Halle (1913a), como parte de la flora de Río Fósiles. Este autor ilustró la especie con ejemplares que presentan una notable variabilidad morfológica, razón por la cual han sido objeto de posteriores revisiones (Frenguelli, 1935; Archangelsky y Baldoni, 1972: 244). Efectivamente, como señalaron estos últimos autores, los especímenes de Halle (1913a), ilustrados en lám. 3, figs. 1-4, parecerían corresponder a una especie distinta de aquellos ilustrados, por este autor, en lám. 3 figs. 5-9. De hecho, los nuevos ejemplares aquí descriptos presentan una mayor similitud con estos últimos.
Otros registros de P. acutifolium incluyen el Jurásico Superior-Cretácico Inferior de India (Bose y Kasat, 1972; Bose y Banerji, 1984; Banerji, 1996), Cretácico Inferior (Neocomiano-Barremiano) del oeste de Australia (McLoughlin, 1996), y Aptiano de Islas Shetland del Sur (Hernández y Azcárate 1971; Césari et al. 1999). Entre todos ellos, se destaca el notable parecido de los ejemplares de Bajo Comisión, con aquellos descriptos por Bose y Kasat (1972) (especialmente el lectotipo, lám. 14, fig. 115) y Césari et al. (1999). Coincidiendo con Césari et al. (1999), no se consideran pertenecientes a P. acutifolium los ejemplares del Jurásico Inferior (Liásico) descriptos por Arrondo y Petriella (1980, lám. 2: e y f) y Herbst (1964) procedentes de Neuquén y Mendoza respectivamente (este último no ilustrado por el autor), ni tampoco los de Longobucco et al. (1985) del Cretácico de Santa Cruz. Frenguelli (1935) asignó a Ptilophyllum hislopi (Oldham) Seward, impresiones foliares que atribuyó al Tithoniano (Jurásico Superior) y que provendrían de la localidad "a" de Halle (1913a). No obstante cierto parecido con los especímenes de Bajo Comisión, la leve separación de los folíolos y su base no decurrente los diferencia de P. acutifolium.
Otras especies de distribución austral, que presentan cierta similitud con P. acutifolium son: Zamites antarticus Halle y Ptilophyllum longipinnatum Menéndez, conocidas para el Jurásico de Antártida (Halle, 1913b; Gee, 1989) y Cretácico (Aptiano) de Patagonia (Menéndez, 1966) respectivamente. Zamites antarticus Halle se caracteriza por sus folíolos largos, estrechos, lineales, con una mayor relación largo/ancho y generalmente más separados que P. acutifolium; la venación es fina y densa y no presenta el carácter constante de los ápices curvados hacia el sector distal (Césari et al., 1999: 16). En tanto, P. acutifolium se distingue de P. longipinnatum, por tener folíolos con una menor concentración de venas y porción catadrómica notablemente decurrente, además, nuevamente de su extremo apical típicamente curvado en dirección acrópeta.
Procedencia. Bajo Comisión 1 (BC1), nivel fosilífero NF A.
Material estudiado. BAPb 12493-12510; BAPbPm 485-488.
Género Cycadolepis Saporta 1873
Especie tipo. Cycadolepis villosa Saporta 1873.
Cycadolepis sp.
Figura 5.8
Descripción. Fragmento de porciones basal y media de una bráctea de 2,5 cm de largo y 1 cm de ancho máximo. Base ancha, redondeada, margen entero. La venación no es clara pero se pueden apreciar, al menos, 10 venas paralelas. Cutícula muy alterada. Comentarios. Cycadolepis Saporta es el nombre dado, en un amplio sentido, a escamas circular-lanceoladas que podrían representar brácteas estériles de conos de bennettitales (Harris, 1969). En este caso aparece asociado (no en conexión orgánica) a numerosas hojas de Ptilophyllum acutifolium Morris.
Procedencia. Bajo Comisión 1 (BC1), nivel fosilífero NF A.
Material estudiado. BAPb 12507a (y su contraparte 12507b).
División GINKGOPHYTA
Orden GINKGOALES Engler 1897
Género Ginkgoites Seward 1919
Especie tipo. Ginkgoites obovata (Nathorst) Seward 1919.
Ginkgoites skottsbergii Lundblad 1971
Figuras 6.8-6.13
1913a. Baiera cf. australis McCoy, Halle, pág. 37, lám. 4, figs. 23-30, lám. 5, figs. 1-6.
Descripción. Hoja palmatilobada con pecíolo de, al menos, 2-4 mm de largo y 1-2 mm de ancho. Lámina con 2-3,2 cm de diámetro; ángulo basal de 90º-120º; dividida dicotómicamente en, al menos, 4-7 segmentos. Éstos son de forma linear lanceolada, de margen entero, con 2-5 mm de ancho máximo (figuras 6.8-6.10). Venas en número de 4-10 en cada segmento, equivalente a 20 (17-25) venas por cm. Hojas anfiestomáticas. Cutículas abaxial y adaxial muy similares, de 1-2 µm de espesor (medido en pliegues). Células epidérmicas, sobre venas, alargadas, rectangulares, dispuestas en hileras, con paredes anticlinales rectas y, entre venas, isodiamétricas a algo alargadas, poligonales, con paredes anticlinales básicamente rectas a finamente sinuosas (figura 6.11). Cara periclinal con características estriaciones finas radiales (figura 6.12). Estomas sin orientación definida (aunque preferentemente longitudinal a oblicua), aleatoriamente distribuidos. Aparato estomático haplocélico, oval a circular; con uno o dos ciclos de subsidiarias (el segundo incompleto), éstas en número de 6-8 (figura 6.13). El primer ciclo de subsidiarias puede presentar paredes engrosadas, generando un anillo oscuro alrededor del ostíolo.
Comparaciones. La suma de características mega y microscópicas permiten asignar estos ejemplares a Ginkgoites skottsbergii Lundblad (1971), originalmente citada por Halle (1913a) para Río Fósiles aunque bajo el nombre de Baiera cf. australis McCoy.
Procedencia.Bajo Comisión 2 (BC2).
Material estudiado. BAPb 12512a-b; 12520a-c; 12513; 12535. BAPbPm 489-494.
Orden CONIFERALES Engler 1897
Género Brachyphyllum Brongniart emend. Harris 1979
Especie tipo. Brachyphyllum mamillare Brongniart 1828.
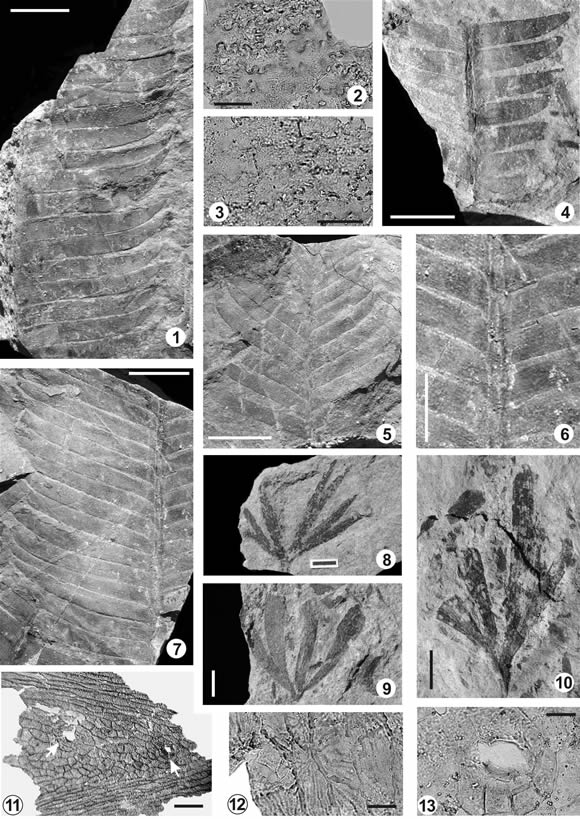
Figura 6. 1-7, Ptilophyllum acutifolium (Morris) Bose y Kasat. 1, aspecto general / general view; BAPb 12506. 2-3, vista interna de cutículas abaxial y adaxial mostrando células epidérmicas con paredes anticlinales onduladas / inner view of abaxial and adaxial cuticle showing epidermal cells with sinuous anticlinal walls; BAPbPm 485 (BAPb 12507). 4, porción basal de hoja / basal part of leaf; BAPb 12500. 5, porción apical de hoja / apical part of leaf; BAPb 12498. 6, detalle de la base de los folíolos, pueden verse venas paralelas / detail of basal part of leaflet, parallel veins are easly recognized; BAPb 12507. 7, aspecto general / general view; BAPb 12507. 8-13, Ginkgoites skottsbergii Lundblad; 8, BAPb 12513; 9, BAPb 12512. 10, BAPb 12520. 11-13, detalle de cutícula / detail of cuticle. 11, cutícula mostrando células epidérmicas alargadas (sobre la venas) e isodiamétricas (entre las venas) y dos estomas (flecha) / cuticle showing epidermal cell elongated (on veins) and isodiametric (between veins) and two stomata (arrow). BAPbPm 494 (BAPb 12520); 12, células epidérmicas con estriaciones finas / epidermal cell with thin striate. BAPbPm 490 (BAPb 12520). 13, detalle de estoma / detail of stomata. BAPbPm 490 (BAPb 12520). Escala / scale 12, 13 = 20 µm, 2, 3 = 50 µm; 11 = 100 µm; 6, 10 = 0,5 cm; 1, 4, 5, 7-9 = 1 cm.
Brachyphyllum kachaikensis sp. nov.
Figuras 7.1-7.9
Holotipo. BAPb 13436.
Otros materiales. BAPb 13424, 13439.
Diagnosis. Shoots branched at acute angle (near to 45º). Leaves scale-like, helically arranged; adpressed, with rhomboidal to oval leaf base cushion and acute apex. Free part of leaf 1/4 to 1/3 of total length. Leaf size 2-2,5 mm long and 1-1,4 mm broad (breadth/length ratio 1,5: 1 to 2: 1). Cuticle thick. Leaf margin macroscopically entire, but microscopically serrate. Epidermal cells isodiametric to rectangular (mostly elongated in longitudinal sense); periclinal cell walls usually smooth but occasionally short and slightly striae; anticlinal cell walls straight, cutinized. Hypodermal cell remains longitudinally elongated, strongly cutinized. Stomata present on both sides. On adaxial side only a few without an apparent order. On abaxial side stomata are uniform scattered but sometimes forming short ill-defined longitudinal rows, covering 2/3 of total leaf surface. Stomata typically monocyclic (sometimes imperfectly dicyclic); with a deep hypostomatical chamber. Subsidiary cells usually 5 (occasionally 4 or 6), isodiametric to polygonal. Subsidiary cells of neighboring stomata in contact. Guard cells slightly sunken. Mouth of pit basically oval to rounded, variably orientated.
Etimología. El epíteto específico kachaikensis alude a la unidad formacional (Kachaike) en que fue coleccionado.
Descripción. Rama de hasta 3 mm de ancho, ramificada en ángulo agudo (próximo a 45º). Hojas escamiformes, dispuestas helicoidalmente de manera adpresa a las ramas (su porción libre corresponde a 1/4 pudiendo llegar a 1/3 de la longitud total), con 2 a 2,5 mm de largo y 1 a 1,4 de ancho máximo (relación largo:ancho entre 1,5: 1 y 2: 1); alargadas, de forma romboidal y ápice agudo (figura 6.9). Margen macroscópicamente entero, aunque microscópicamente serrulado (figura 7.4). Células epidérmicas desde isodiamétricas a rectangulares, en este caso, mayormente elongadas en sentido longitudinal, con paredes periclinales lisas que pueden presentar, no obstante, engrosamientos cuticulares a modo de estrías cortas y poco marcadas. No se observan papilas. Paredes anticlinales rectas, de 6-7,5 µm de grosor (incluyendo ambas caras de células epidérmicas en contacto y laminilla media). Sus medidas oscilan entre 40 y 50 µm de largo y 20 a 25 µm de ancho. Restos de células hipodérmicas fuertemente cutinizadas. Estos engrosamientos de cutina pueden disponerse alrededor de los aparatos estomáticos formando un anillo de estrías (figura 7.6). Hojas anfiestomáticas. Estomas provistos de una profunda cámara hipoestomática. En cara adaxial sólo unos pocos estomas, distribuidos sin orden aparente. En cara abaxial, distribuidos uniformemente o en hileras cortas poco definidas, cubriendo más de 2/3 partes de la longitud total de la hoja y casi la totalidad del ancho de la misma. Son más escasos, e incluso ausentes, en una franja que se extiende de base a ápice y ocupa la porción media de la hoja, lo que sugiere la presencia de una vena media. Aparato estomático típicamente monocíclico (pudiendo presentarse como imperfectamente dicíclico), formado generalmente por 5 células subsidiarias (ocasionalmente 4 ó 6) isodiamétricas a poligonales, en general no diferenciadas en polares y laterales (figuras 7.5-7.8). No se observan células subsidiarias compartidas, pero sí subsidiarias vecinas en contacto. Ostíolo básicamente oval a circular, de 35-45 µm de largo y 25-30 µm de ancho, sin orientación definida. Células oclusivas hundidas (figura 7.9), de 30 µm de largo y 7 µm de ancho, con marcado engrosamiento de su cara interna (figura 7.7).
Comparaciones. Por su filotaxis helicoidal y forma de las hojas, puede incluirse en el género Brachyphyllum Brongniart emend. Harris (1979). Seis especies de Brachyphyllum (B. baqueroense Traverso, B. bretti Archangelsky, B. irregulare Archangelsky, B. mirandai Archangelsky, B. mucronatum Archangelsky y B. tigrense Traverso) han sido citadas para el Cretácico Inferior (Aptiano) de Patagonia (Archangelsky, 1966; Traverso, 1966, 1968). Entre éstas, B. baqueroense es asignable al género Pagiophyllum Heer. A su vez, ramas portadoras de hojas de este tipo han sido asignadas a las familias Araucariaceae (Nothopehuen brevis Del Fueyo, 1991) y Podocarpaceae (Squamastrobus tigrensis Archangelsky y Del Fueyo, 1989), demostrando que se trata de un grupo artificial. Si bien las hojas de Bajo Comisión presentan cierta semejanza con todas estas especies, es posible diferenciarlas de ellas. Las hojas de B. tigrense y S. tigrensis se distinguen por sus estomas dispuestos en hileras longitudinales. En B. bretti los estomas también tienden a formar hileras (aunque menos pronunciadas que en B. tigrense), y sus ostíolos son de menor dimensión y forma más elongada que los de Bajo Comisión; además presenta estrías características en la cara periclinal de las células epidérmicas, las que se hallan apenas desarrolladas a ausentes en nuestros ejemplares. Estas estrías aparecen aún más marcadas, pudiendo estar asociadas a papilas en B. mirandai diferenciándose así de los ejemplares de Bajo Comisión. B. mucronatum se distingue por sus hojas más elongadas, con hipodermis menos cutinizada y ostílolo de menores dimensiones. Con B. irregulare presenta similitudes; sin embargo, esta especie posee, como una característica constante y distintiva, células subsidiarias con numerosos surcos que atraviesan la cara interna de la pared anticlinal en dirección a la boca estomática, carácter ausente en nuestros especímenes. Además, todas estas especies poseen aparatos estomáticos formados generalmente por cuatro células subsidiarias, en tanto en los ejemplares de Bajo Comisión lo más usual es cinco y ocasionalmente cuatro o seis subsidiarias. Esta característica (típicamente cinco células subsidiarias) es compartida con Nothopehuen brevis, la cual se distingue por la presencia de papilas en la porción distal de la hoja. Brachyphyllum feistmantelii (Halle) Sahni emend. Archangelsky (1976) es una especie de distribución austral del Jurásico Superior a Cretácico Inferior (véase Archangelsky, 1976) y cuyas hojas se disponen perpendiculares al eje del tallo, carácter que lo distingue de los espécimenes de Bajo Comisión.
Baldoni (1980) describió dos nuevas especies, Brachyphyllum ramosum Baldoni y Brachyphylum lotenaense Baldoni, del Jurásico Medio de la cuenca Neuquina. B. ramosum se distingue principalmente por sus hojas hipostomáticas, con muy escasos estomas, generalmente en la base de la cara abaxial; en tanto B. lotenaense consiste sólo de impresiones de hojas de contorno rómbico con ángulos redondeados que parecen diferir de los ejemplares de Bajo Comisión.
Impresiones de ramas de coníferas, provenientes de depósitos del Jurásico Inferior a Medio de Santa Cruz, han sido asignadas al género Brachyphyllum por Herbst (1965) y Herbst y Salazar (1998). No obstante, las mismas difieren de B. kachaikensis por presentar hojas de contorno romboidal generalmente más anchas que largas.
Halle (1913a) describió, para Río Fósiles, abundantes ramas con hojas atribuibles, en principio, al género Brachyphyllum. Sin embargo, se encuentran en conexión con conos megasporangiados, por lo que fueron incluidos por el autor en la nueva especie Athrotaxites ungeri Halle. Esta especie ha sido reconocida por Archangelsky (1963) para otra localidad también cretácica (aptiana) de Santa Cruz, agregando características cuticulares a la diagnosis específica (ahora como Athrotaxis ungeri). Sus hojas presentan estomas principalmente en cara adaxial y muy raramente en la base de la cara abaxial por lo que se distingue claramente de nuestros especímenes.
En síntesis, los ejemplares de Bajo Comisión presentan gran similitud con las especies de Brachyphyllum descriptas para el Grupo Baqueró, especialmente con B. irregulare y B. mirandai. Sin embargo, presentan pequeñas diferencias en sus caracteres cuticulares que justifican la proposición de esta nueva especie. Procedencia. Bajo Comisión 1 (BC1), nivel fosilífero NF E.
Material estudiado. BAPb 12429, 12430-12432, 12436, 12438, 13410 y contraparte 13411, 13424 y contraparte 13425, 13429, 13432, 13434, 13436 y contraparte 13437, 13438, 13439, 13441, 13444, 13449, 13450, 13454 y contraparte 13489. BAPbPm 495 (BAPb 12432 y BAPb 12436), 496 (BAPb 12436), 497 (BAPb 12436), 498 (BAPb 13439), 499 (BAPb 13449), 500 (BAPb 13449), 501 (BAPb 13449), 502 (BAPb 13449), 503 (BAPb 13449), 504 (BAPb 13425).
Material estudiado. BAPb 12494 (NF A), BAPb 12469, 12490, 12491 (NF B), BAPb 12443, 12446, 12460 (NF C), BAPb 12536-12541 (NF D), BAPb 12426 (NF F).
Familia TAXODIACEAE sensu Page 1990
Género Athrotaxis (Unger) Florin 1940
Especie tipo. Athrotaxis lycopodioides (Unger) Florin 1940.
Athrotaxis ungeri (Halle) Florin 1940
Figuras 7.10-7.11
1913a. Athrotaxites ungeri Halle, pág. 40, lám. 3, figs. 13-18, 19?, 20?, 21?, lám. 5, figs. 10-13.
Descripción. Conos megasporangiados de forma oblonga en compresión lateral (longitudinal) (figura 7.10) y contorno cilíndrico, globoso en compresión apical (figura 7.11), de hasta 1,5 cm de largo y 0,8-1 cm de ancho máximo. Se observa bráctea tectriz de, al menos, 3 mm de largo, de base cuneada y distalmente expandida en forma de triángulo culminando en un corto ápice agudo. Las brácteas, algo separadas unas de otras parecen estar completamente fusionadas a la escama ovulífera. Uno de los especímenes se encuentra en conexión orgánica con una porción de rama portadora de hojas escamiformes, adpresas.
Comentarios. Generalmente, los conos se encuentran maduros y aislados, sugiriendo algún tipo de mecanismo de abscisión que funcionaría luego de la fecundación. En los niveles correspondientes al sitio BC2, los conos son muy numerosos debido, tal vez, a que la abscisión ocurrió bajo un control estacional (véase Miller y LaPasha, 1983).
La especie ha sido descripta por Halle (1913a), para Río Fósiles, como Athrotaxites ungeri Halle. Conos megasporangiados de Athrotaxis ungeri (Halle) Florin han sido también descriptos por Llorens y Del Fueyo (2003) provenientes de otra localidad cretácica (Ea. Kachaike) próxima a la localidad "b" de Halle (1913a) y algo más al sur de Bajo Comisión (cuyos depósitos corresponden también a la Formación Kachaike). Cantrill y Falcon-Lang (2001) describieron conos megasporangiados que asignaron a Athrotaxites ungeri Halle, procedentes de la Formación Triton Point (Albiano tardío) de Antártida. El registro más antiguo para la especie, aunque en este caso identificada sobre la base de material vegetativo, corresponde al Aptiano (Formación Anfiteatro de Ticó) de Santa Cruz (Archangelsky, 1963).
Procedencia. Bajo Comisión 1 (BC1) (nivel fosilífero NF E) y BC2.
Material estudiado. BAPb 13436 y su contraparte 13437 (BC1, NF E), BAPb 12514; 12516-12517; 12522a-b-12527; 12529; 12532-12534 (BC2).

Figura 7. 1-9, Brachyphyllum kachaikensis sp. nov. 1, último y penúltimo orden de ramificación, las flechas indican sitios de ramificación / ultimate and penultimate order branching, arrows indicate branched sites; BAPb 13436. 2, detalle de rama con hojas adpresas y dispuestas helicoidalmente / detail of branch with adpressed and helically arranged leaves; BAPb 13424. 3, aspecto general de hoja mostrando cara abaxial (en vista interna) y adaxial muy reducida (en vista externa) / general view of leaf showing abaxial surface (inner view) and very reduced adaxial surface (outer view); BAPbPm 504 (BAPb 13425). 4, detalle de margen dentado / detail of thooted margin. BAPbPm 495 (BAPb 12432). 5, disposición de estomas en cara abaxial / stomatal arrangement on abaxial surface; BAPbPm 504 (BAPb 13425). 6, remanentes de células hipodérmicas formando un anillo de estrías alrededor del estoma / remains of hypodermal cells forming a ring of striate around the stomata; BAPbPm 500 (BAPb 13449). 7-8, detalle de estoma en vista interna de cara abaxial / detail of stomata from inner view of abaxial surface; 7, estoma con 6 células subsidiarias, células oclusivas hundidas con engrosamiento longitudinal en cara proxima al poro / stomata with 6 subsidiary cells, sunken guard cells showing a longitudinal thickening on the proximal surface; BAPbPm 498 (BAPb 13439). 8, estoma mostrando cámara subestomática profunda y restos de células oclusivas / stomata showing a deep hypostomatical chamber and guard cells; BAPb 13453. 9, detalle de estoma, en vista externa de cara abaxial, mostrando células oclusivas hundidas / detail of stomata from external view of abaxial surface showing sunken guard cells. 10-11, Athrotaxis ungeri (Halle) Florin; 10, cono megasporangiado en compresión lateral / longitudinal compression of megasporangiate cone; BAPb 12523. 11, cono megasporangiado en compresión apical / apical compression of megasporangiate cone. 12-15, morfotipo A; 12, BAPb 13490; 13, BAPb13489; 14-15, detalle dientes / detail of teeth; BAPb13489. Escala / scale 9 = 10 µm; 4, 7, 8 = 20 µm; 6 = 50 µm; 5 = 100 µm; 2= 0,1 cm; 1, 3, 10-13 = 0,5 cm.
Magnoliopsida Incertae sedis
Thorhallenia dentata gen. et sp. nov.
Figuras 8.1-8.10, 9.1-9.6
Sintipos. 13418, 13424, 13436.
Thorhallenia gen. nov.
Diagnosis. Simple leaf (or leaflet), microphyll, elliptic to near ovate laminae, sligthly incised with one or two lateral pairs lobes. Base obtuse, convex to concave-convex shape; apex convex, acute to obtuse. Petiole marginal. Margin toothed. Teeth simple to compound with typically convex shape; angular sinus and apparently glandular apex. Each tooth irrigated by one middle main vein and one or two lateral accessories. Venation pinnate craspedodromous to actino/palinactinodromous. Primary vein straight. Secondary order veins (3-4 pairs) opposite to subopposite. Tertiary order venation with a combinated arrangement between alternate percurrent and reticulated.
Thorhallenia dentata sp. nov.
Diagnosis. Igual que para el género.
Etimología. El nombre genérico Thorhallenia constituye un reconocimiento al paleobotánico sueco Thor G. Halle. El epíteto específico dentata alude a la presencia de margen foliar provisto de dientes.
Descripción. Hoja simple, micrófila, de 3-5 cm de largo y 2-4 cm de ancho máximo (relación largo:ancho de 1-1,7:1). Lámina elíptica a algo ovada, básicamente simétrica a algo asimétrica (figuras 8.5-8.6; 9.3-9.4). Base obtusa, de forma convexa (figuras 8.5, 9.4) a cóncavo-convexa; ápice agudo (85º en figuras 8.6, 9.3) a obtuso (100º en figuras 8.2, 8.6, 9.1, 9.4) de forma convexa. Pecíolo marginal de hasta 1,2 cm de largo y 1,3 mm de ancho máximo (figuras 8.3, 9.2). Lámina algo incisa entre venas de 2º orden, determinando uno o dos pares de lóbulos laterales. Margen dentado. Dientes simples a compuestos (de 2º orden en figuras 8.1-8.2 y 9.1 y hasta 3º orden, en figuras 8.3-8.4 y 9.2), irregularmente espaciados, de forma típicamente convexa-convexa (tipo A1 sensu Hickey, 1973), con senos angulares y un capuchón o mamelón apical. Cada diente es vascularizado por una vena principal o media (de 2º o más alto orden) y una o dos laterales (de 3º o tal vez más alto orden pero de grosor relativo considerable) que se fusionan próximo al ápice del diente. Dependiendo del desarrollo relativo y recorrido del par basal de venas de segundo orden el plan de venación varía desde pinnado craspedódromo (figuras 8.2-8.3, 8.6; 9.1-9.2, 9.4) a actinódromo/palinactinódromo (figuras 8.7, 8.8, 8.10; 9.5-9.6). En este último caso, se considera el par de secundarias basales como venas primarias laterales. La vena de 1º orden (o vena primaria principal en los ejemplares actino/palinódromos) es de grosor moderado a típicamente fuerte (0,4-0,9 mm de grosor máximo), de recorrido recto, disminuyendo de grosor progresivamente en dirección apical. Venas de 2º orden, 3-4 pares, opuestas a subopuestas, con ángulos de emergencia agudos, de entre 30-45º el par basal y aumentando progresivamente a 50-60º los superadyacentes. Recorrido básicamente recto; grosor relativo moderado a grueso (0,4-0,8 mm para las venas basales a 0,15 mm para el último par apical). Las secundarias basales emergen del mismo punto que la vena media; presentan ramificaciones exmediales, casi ortogonales a la vena media y aproximadamente paralelas entre sí, culminando en el ápice de un diente (venas agróficas sensu Leaf Architecture Working Group, 1999: 32). La venación de 3º orden presenta un ordenamiento que combina modelos percurrente alterno y reticulado. Ángulo de emergencia perpendicular a obtuso respecto a la vena media.
Comentarios. A partir de un patrón arquitectural que incluye: hojas con cierta lobación, pecioladas, margen con dientes doblemente convexos, venación desde pinada craspedódroma a actino/palinactinódroma y venas de 2º orden opuestas a subopuestas, se han aceptado, dentro de Thorhallenia dentata gen. et sp. nov., ejemplares que podrían parecer, en primera instancia, morfológicamente distanciados. Ahora bien, emergiendo a mitad del recorrido del primer par de venas secundarias (o primarias laterales), se observa una ramificación apical, que coalesce con una ramificación basal del par superadyacente conformando una vena que se dirige al seno del lóbulo pero sin alcanzarlo. Este patrón de venación, con cierta variación, puede reconocerse también entre pares superadyacentes de venas secundarias. Si bien es frecuente en formas lobadas pertenecientes a distintas especies actuales, este patrón de venación es común a todos los ejemplares fósiles y sería un elemento que justificaría la inclusión bajo un mismo morfotipo de las formas fósiles aparentemente distintas según lo mencionado anteriormente. Otro carácter común a todos los ejemplares es la presencia de dientes con un capuchón o mamelón apical, vascularizados por una vena media y hasta dos laterales. Dependiendo del distinto grado de lobación de la lámina es posible observar cierta plasticidad en el desarrollo de la venación. Es de esperar que hojas con lóbulos basales más profundos refuercen el par basal de venas, aproximándose, por su grosor relativo, a venas primarias laterales y derivando en un plan de venación actino/palinactinódromo.
En definitiva, si bien es evidente cierto espectro de variación morfológica en Thorhallenia dentata sp. nov., que podría estar incluyendo más de una especie, no hay elementos que justifiquen claramente una separación entre especies o tipos morfológicos distintos. Por otro lado, durante la radiación inicial de poblaciones de organismos u órganos aislados, es de esperar que éstos ofrezcan una intergradación morfológica continua. Producto de hibridización, poliploidía, plasticidad fenotípica o una combinación de estos fenómenos, las asociaciones de angiospermas cretácicas reflejan una considerable variación de formas (Spicer, 1986).
Por otro lado, cierta asimetría en BAPb13429 (figuras 8.5, 9.3) y BAPb13409 (figuras 8.6, 9.4) podría sugerir que se trata de hojas compuestas, sin embargo no se han encontrado folíolos en conexión orgánica que refuercen esta idea.
Comparación con angiospermas fósiles. Thorhallenia dentata gen. et sp. nov. presenta gran similitud con la especie Cissus guidoensis (Kurtz) Hünicken 1995 de la flora maastrichtiana de cerro Guido (cuenca Austral). Coinciden en la forma general de la hoja, dientes doblemente convexos y venas secundarias opuestas. Esto es especialmente notorio entre BAPb13424 (figuras 8.1-8.2; 9.1) y los ejemplares tipo y paratipo de cerro Guido (Hünicken, 1995: lám. B, figs. 12 y 15, lám. F, figs. 10 y 11). No obstante, Thorhallenia dentata se diferencia de los ejemplares de cerro Guido principalmente por la ausencia de venas secundarias "congestionadas" en la base.
En depósitos pertenecientes a la Formación La Cantera (Aptiano tardío) en la provincia de San Luis, Argentina, se encontraron hojas de angiospermas pertenecientes, al menos, a tres morfotipos (Puebla, 2003). Uno de ellos, consistente en hojas compuestas trifoliadas con folíolos nanófilos, pinnatilobados (con ápices de los lóbulos aparentemente glandulares) y venación craspedódroma, presenta cierta semejanza con Thorhallenia dentata gen. et sp. nov. Sin embargo, se distingue principalmente por su tamaño, profundidad de los senos de los dientes y curso notablemente recto de sus venas secundarias.
Cantrill y Nichols (1996) propusieron un nuevo morfogénero (Gnafalea) con dos especies (G. binatus Cantrill y Nichols y G. jeffersonii Cantrill y Nichols), para hojas lanceoladas a ovadas, de margen dentado y venación pinnada craspedódroma, integrantes de una asociación de angiospermas del Cretácico (Albiano tardío) de Antártida. La primera de ellas (G. binatus) presenta dientes doblemente convexos, recorridos por una vena media y una accesoria, y venas secundarias opuestas. Estas características son observadas en Thorhallenia dentata gen. et sp. nov. y sugieren cierto parecido entre ambas especies, en particular con los ejemplares patagónicos menos lobados. No obstante, la especie antártica se diferencia, por un lado, por la presencia de venas secundarias marcadamente curvadas y, por otro, por el recorrido de las mismas en dirección al seno de cada diente (y no al ápice de los mismos como en Thorhallenia dentata gen. et sp. nov.), bifurcándose próximo al margen, de modo que cada rama vasculariza al diente sub y suprayacente. Además, en G. binatus los dientes se disponen algo más proyectados apicalmente (incluso los basales) respecto a lo observado en Thorhallenia dentata gen. et sp. nov. Por su parte, G. jeffersonii, que también presenta dientes convexo-convexos, se distingue por la ausencia de lobación, venas secundarias más próximas entre sí (y por consiguiente mayor número de las mismas) y por el margen variable, el cual puede ser parcialmente entero.
Hojas lobadas, dentadas, con venación actinódroma/craspedódroma han sido reconocidas en depósitos campaniano-maastrichtianos de Nueva Zelanda e incluidos por Pole (1992) dentro del parataxón TARA-22 ("Aralia tasmani" Ett.). El mismo contempla, sin embargo, algunos ejemplares de mayor tamaño y otros marcadamente más lobados que los ejemplares de Bajo Comisión.
McLoughlin et al. (1995) describieron pequeñas hojas dentadas, algo lobadas y venación craspedódroma (angiosperm sp. "H") en depósitos del Cenomaniano de Australia. Sin embargo, la ausencia de venación de mayor orden no permite una adecuada comparación.
Hojas con dientes vascularizados por una vena media y hasta dos laterales se encuentran presentes en ejemplares del Aptiano de Patagonia descriptos por Romero y Archangelsky (1986). Con estos ejemplares, Torhallenia dentata gen. et sp. nov. comparte, además, el tipo de venación craspedódroma. Estas características están presentes también en hojas del Cretácico (Aptiano?) de Colombia descriptas como Moutonia quetamensis Pons (1988) y en una dicotiledónea no identificada con venación craspedódroma reportada para el Albiano tardío-Cenomaniano temprano del Oeste-Noroeste de América del Norte (Crabtree, 1987: 730-31, fig. 36).
Procedencia. Bajo Comisión 1 (BC1), nivel fosilífero NF E.
Material estudiado. BAPb 13408, 13409, 13410 y contraparte 13411, 13412, 13418, 13419, 13421, 13422, 13424 y contraparte 13425-13429, 13433, 13434, 13436.
Morfotipo A
Figuras 7.12-7.15, 9.8-9.11
Descripción. Fragmento de porción apical (de 1,5 cm de largo y 1,7 cm de ancho) de una hoja dentada con venación craspedódroma a semicraspedódroma (figura 7.13). Venas de tercer orden dispuestas según un modelo entre percurrente alterno y reticulado. Dientes convexo-convexos (tipo A1, sensu Hickey, 1973), de contorno redondeado, con un capuchón o mamelón apical, vascularizados por una vena media y dos laterales accesorias (figuras 7.14-7.15, 9.10-9.11). Un depósito de materia orgánica en el ápice de los dientes sugiere un carácter glandular para los mismos. Puede observarse, también, que la vena media y las accesorias se fusionan justo sobre la presunta glándula. Exmedialmente a las venas laterales, y casi sobre el margen del diente, puede desarrollarse una segunda vena, ambas conectadas por venas transversales (figuras 7.15, 9.11).
Comentarios y comparaciones. Si bien se trata sólo de un fragmento, por su buena preservación y, principalmente por las características de sus dientes, merece especial atención. Este tipo de diente (con ápice glandular, vascularizado por una vena media y laterales accesorias) es referido por Hickey y Wolfe (1975) como "tipo chlorantoide". Todzia y Keating (1991) mediante una revisión de la arquitectura foliar de las Chloranthaceae identifican, al menos, seis tipos de dientes, de los cuales el "2B" es el que más se aproxima al del ejemplar fósil. Cabe destacar la particular similitud del material patagónico con el tipo de dentición de Chloranthus henryi Hemsl. (véase Hickey y Wolf, 1975, fig. 2.A). Sin embargo, la presencia de dientes del tipo chloranthoide no es exclusiva de Chloranthaceae, sino que es compartida por otros representantes de las Magnoliidae (Illiciales; Amborella), además de Hamamelidae (Cercidiphyllaceae, Trochodendraceae) y Ranunculidae (Ranunculaceae, Podophyllaceae, Hydrastidaceae, Glaucidiaceae) (Hickey y Wolf, 1975; Wolfe, 1989; Doyle y Endress, 2000). No obstante su acotada presencia entre los linajes de angiospermas actuales, este tipo de diente aparece de modo recurrente en hojas cretácicas de distintas regiones del mundo. Dientes convexo-convexos con ápice glandular están presentes en dos especies pertenecientes a la Zona I del Grupo Potomac (Aptiano temprano y medio de América del Norte): Proteaephyllum dentatum Fontaine y Quercophyllum tenuinerve Fontaine (Doyle y Hickey, 1976; Hickey y Doyle, 1977), siendo esta última especialmente semejante a nuestro ejemplar. Hydrocotylophyllum alexandri Cantrill y Nichols (1996), del Albiano tardío de Antártida, presenta crenaciones vascularizadas por una vena media y laterales subsidiarias fuertes. Dientes chlorantoideos están presentes en ejemplares del Aptiano de Patagonia (Romero y Archangelsky, 1986), Cretácico (Aptiano?) de Colombia (Pons, 1988) y Albiano tardío-Cenomaniano temprano de América del Norte (Crabtree, 1987: 730-731, fig. 36), entre otros.
En virtud de sus dientes apicales grandes y marcadamente redondeados este espécimen es tratado aquí como un tipo morfológico aparte. Por la forma y tamaño de los dientes se incluye bajo este morfotipo al ejemplar BAPb 13490 (figuras 7.12, 9.8) y su contraimpronta BAPb 13491. No obstante, es muy probable que corresponda a un fragmento apical de Thorhallenia dentata gen. et sp. nov. A este respecto, es interesante hacer notar la semejanza con la porción distal de los ejemplares BAPb 13429 (figuras 8.5, 9.3) y BAPb 13409 (figuras 8.6, 9.4). Pese a ello, por desconocerse su patrón de venación de primer orden y por no haberse encontrado dientes con idénticas características a las de este espécimen (aunque sí semejantes) en los ejemplares referidos a Thorhallenia dentata gen. et sp. nov., no se lo ha incluido en esta especie.
Procedencia. Bajo Comisión 1 (BC1), nivel fosilífero NF E.
Material estudiado. BAPb 13489, 13490 y contraparte 13491.
Morfotipo B
Figuras 8.11-12, 9.7
Descripción. Fragmento único de una hoja simple, pinnatilobada con al menos dos, tal vez tres, pares de lóbulos; nanófila, de 2 cm de largo y 1,5 cm de ancho máximo (relación largo: ancho de, al menos, 1,3: 1). Textura herbácea. Lámina básicamente simétrica. Extremos basal y apical no conservados. Lóbulos con margen entero y senos redondeados que alcanzan, aproximadamente, la mitad de la distancia a la vena media. Venación pinnada y aparentemente craspedódroma. Vena de 1º orden de grosor débil a moderado (0,2 mm de grosor máximo), de recorrido algo zigzagueante. Venas de 2º orden (al menos 3 pares) alternas, con ángulos de emergencia moderados, entre 50-65º. Grosor relativo grueso (en el orden de 0,15-0,2 mm); recorrido algo sinuoso que culmina, aparentemente, en el ápice de cada lóbulo. Venación de 3º orden sin un ordenamiento claro, pudiendo ser alterna u opuesta; con un ángulo de emergencia variable y recorrido recto a sinuoso; grosor similar a algo menor respecto a las venas de 2º orden. En los lóbulos, las venas de 3º orden están conectadas por arcos broquidódromos planos, constituyéndose una vena marginal de grosor similar al de las venas de 2º orden. Exmedialmente a ésta se desarrolla una vena fimbrial. Venación de orden más alto sin una clara disposición (percurrente a poligonal). Cabe destacar que la excesiva sinuosidad de algunas venas es, en parte, producto de la deformación post deposicional de la hoja.
Comparaciones. Entre las angiospermas cretácicas de Patagonia, sólo se conocen hojas pinnatilobadas en depósitos coniacianos de la Formación Mata Amarilla (Iglesias et al., 2004). Hojas pinnatilobadas con senos redondeados han sido descriptas para el Cenomaniano de Australia (como Angiosperm sp. "C", en McLoughlin et al., 1995) y Campaniano-Maastrichtiano de Nueva Zelanda (como Nothofagus praequercifolia (Ett.) Pole y N. melanoides Pole y parataxón TARA-20 "Fagus producta" Ett. en Pole, 1992) formas éstas, bastante diferentes a nuestros ejemplares. También están presentes en asociaciones eocretácicas de otras regiones paleoflorísticas. Tal es el caso de "hoja tipo 5" descripta por Mohr y Friis (2000) para la Formación Crato (Aptiano-Albiano?) del nordeste de Brasil. La misma presenta, al igual que el Morfotipo B, profundos lóbulos, venación pinnada craspedódroma y venas de tercer orden formando arcos broquidódromos en los lóbulos.
Procedencia. Bajo Comisión 1 (BC1), nivel fosilífero NF E.
Material estudiado. BAPb 13420.

Figura 8. 1-10, Thorhallenia dentata gen et. sp. nov. 1-2, sintipo BAPb 13424; 1, detalle dientes / detail of teeth; 3-4, sintipo BAPb 13418; 4, detalle diente / detail of tooth; 5, BAPb 13429; 6, BAPb 13409; 7, BAPb 13421; 8-9, sintipo BAPb 13436; 9, detalle venación / detail of venation; 10, BAPb 13411; 11-12, morfotipo B. BAPb 13420; 12, detalle venación / detail of venation. Escala / scale 1 = 0,1 cm; 9 = 0,2 cm; 5, 6, 8, 10, 11 = 0,5 cm; 2, 3, 7 = 1 cm.

Figura 9. 1-6, Thorhallenia dentata gen et. sp. nov. 1, sintipo BAPb 13424; 2, sintipo BAPb 13418; 3, BAPb 13429; 4, BAPb 13409; 5, BAPb 13421; 6, BAPb 13411; 7, morfotipo B. BAPb 13420; 8-11, morfotipo A; 10, detalle diente / detail of tooth; 12-16, corresponden a dibujos esquemáticos / correspond to schematic drawings; 12, Gleichenites juliensis Herbst, detalle pínnula / detail of pinnule; 13, Gleichenites sanmartini Halle emend. Herbst, detalle pínnula / detail of pinnule; 14, Pachypteris crassa (Halle) Townrow, detalle pinna / detail of pinnae; 15, Pachypteris auriculata sp. nov., detalle pínnula / detail of pinnule; 16, cf. Sphenopteris sp. 'f' Douglas, detalle sección de pinna primaria, rp = raquis primario, ps = pinna secundaria, pn = pínnula / detail of primary pinae section, rp = primary rachis, ps = secondary pinnae, pn = pinnule. Escala / scale 12, 13 = 0,5 mm; 14 = 0,2 cm; 1-9, 15, 16 = 0,5 cm.
Las angiospermas de Bajo Comisión
Los registros más tempranos de hojas de angiospermas en Patagonia corresponden a depósitos del Aptiano Superior temprano (Romero y Archangelsky, 1986; Passalia et al., 2003). La primera mención de posibles angiospermas en depósitos cretácicos en la zona de lago San Martín corresponde a Halle (1913a) quien describió dos pequeños fragmentos referidos con dudas a dicotiledóneas: uno con margen entero y otro dentado pero sólo conservando una vena media. Los mismos corresponden a la parte superior del valle del Río Fósiles y aparecen, al igual que en Bajo Comisión, asociados a frondes esfenopterídeas. Proveniente de Ea. Kachaike, localidad próxima a Bajo Comisión, Cúneo y Gandolfo (2005) describieron una asociación de hojas de angiospermas obtenida de sedimentitas cretácicas asignadas a la Formación Kachaike. Consta de hojas simples y, al menos, una compuesta, en su mayoría elípticas, con venación broquidódroma a eucamptódroma pero todas ellas con margen entero; características éstas completamente diferentes a las de las hojas de Bajo Comisión. Tampoco se encuentran claras semejanzas con impresiones foliares de angiospermas conocidas para otros depósitos cretácicos de Patagonia (eg. Berry, 1928, 1937; Frenguelli, 1953; Menéndez, 1959; Passalia et al., 2001). Así, con excepción de Cissus guidoensis (Kurtz) Hünicken, 1995 perteneciente a la flora maastrichtiana de cerro Guido (Chile), no se ha encontrado, en asociaciones cretácicas de Patagonia -y, hasta donde el autor conoce, del extremo sur de Gondwana-, registros foliares de angiospermas en todo comparables a los de Bajo Comisión. Sin embargo, alguno de los caracteres individuales de estos últimos puede reconocerse en angiospermas de varias asociaciones del Cretácico de ambos hemisferios. Este es el caso del tipo de dientes chloranthoidos presentes en "morfotipo B" y, aparentemente, Thorhallenia dentata gen. et sp. nov. Es interesante notar la recurrencia de este tipo de dientes (aunque con variaciones) asociados a distintos patrones arquitecturales de hojas eocretácicas (e.g. Romero y Archangelsky, 1986; Crabtree, 1987; Pons, 1988; Cantrill y Nichols, 1996). Sobre la base de su registro fósil, Hickey y Taylor (1989) postularon la presencia de dientes chloranthoidos como prototípico de las angiospermas.
Por otro lado, como una forma de representar el progresivo incremento en regularidad del patrón de venación foliar, Hickey (1977) definió cuatro rangos aplicables a hojas de angiospermas del Cretácico de América del Norte. Este incremento en rangos puede observarse, también, en las asociaciones eocretácicas de Patagonia. Así, tenemos hojas pertenecientes a un rango I (Passalia et al., 2003) y rango I alto (Romero y Archangelsky, 1986) para Formación Anfiteatro Ticó (Aptiano), rango II para hojas de Bajo de los Corrales (Albiano?) y rango II y III (Passalia et al., 2001) para las hojas de la Formación Castillo (Albiano tardío a Cenomaniano temprano). En el caso de Thorhallenia dentata gen. et sp. nov., la presencia de venas de segundo orden de curso usualmente regular, no decurrentes, la ubicaría en un rango distinto de I, en tanto la disposición más o menos irregular de las venas de tercer orden y su no tan clara resolución de las de cuarto orden sugieren un rango II, tal vez casi un rango III. En este sentido, y pese a corresponder a la misma unidad formacional, es notable la diferencia entre las angiospermas de Bajo Comisión y las descriptas para Ea. Kachaike (Cúneo y Gandolfo, 2005), cuyo grado de organización ciertamente es muy inferior (rango I). Ahora bien, considerando el incremento en rangos como el resultado de un proceso evolutivo que tiene lugar a través del tiempo (y dejando de lado la posibilidad de reversiones), la notable diferencia entre las hojas de Bajo Comisión y Ea. Kachaike estaría indicando o bien una marcada separación temporal entre ambas tafofloras, o un rápido proceso de organización de la venación, o bien la posibilidad de coexistencia de hojas de angiospermas con rangos disimiles de organización de la venación e incluso la simultaneidad de algunos de estos eventos.
Comparaciones con otras macrofloras cretácicas de Patagonia
El contenido fosilífero de ambas localidades (Bajo Comisión 1 y 2), y su distribución estratigráfica, se resumen en el cuadro 1. Del total de especies reconocidas hasta el momento, ocho han sido previamente citadas por Halle (1913a), para las localidades "b" y "c" de Río Fósiles (cuadro 2), ya sea bajo la misma o distinta ubicación sistemática (producto de ulteriores revisiones), pero pertenecientes a la misma entidad específica. La localidad "c" de Halle (1913a) es la más próxima geográficamente a Bajo Comisión y la que posee el mayor número de elementos en común. Los resultados aquí expuestos indican que muy probablemente la flora del presente trabajo corresponde a la misma secuencia deposicional estudiada por el paleobotánico sueco y posiblemente a los mismos o muy cercanos afloramientos. La lista paleoflorística de Halle (1913a) ha sido objeto de revisiones basadas en las ilustraciones de su publicación (Krassilov, 1964; Herbst, 1971; Baldoni, 1981) o bien sobre el material original (Lundblad, 1955, 1971; Van Konijnenburg-van Cittert, 1993) o nuevo (este trabajo). Por tal razón la misma se encuentra ahora levemente modificada (cuadro 2).
Cuadro 1. Distribución de las especies en cada nivel fosilífero (NF A - NF F) en el perfíl de Bajo Comisión 1 y especies presentes en Bajo Comisión 2. / distribution of species by fossiliferous level (NF A - NF F) in the Bajo Comisión 1 section and species recognized in Bajo Comisión 2.

Cuadro 2. Lista de especies reconocidas por Halle (1913a) en el área de lago San Martín y posteriores revisiones de la misma por (K) Krassilov 1964, (H) Herbst 1971, (L) Lundblad 1955 y 1971, (B) Baldoni 1981, (V) Van Konijnenburg-van Cittert (1993) y este trabajo. / Species recognized by Halle (1913a) in the surrounding area of San Martín Lake and later revision of them by (K) Krassilov 1964, (H) Herbst 1971, (L) Lundblad 1955 and 1971, (B) Baldoni 1981, (V) Van Konijnenburg-van Cittert (1993) and this study.
Con posterioridad al trabajo de Halle (1913a), se han realizado otros estudios paleobotánicos en afloramientos del Cretácico Inferior, cercanos a lago San Martín, asignados a la Formación Belgrano (Baldoni y Ramos, 1981) y Formación Kachaike (Baldoni y Ramos, 1981; Longobucco et al., 1985; Llorens y Del Fueyo, 2003; Cúneo y Gandolfo, 2005). El reducido número de especies identificadas en cada una de estas localidades no permite un análisis de sus relaciones estatigráficas; no obstante ello, entre las procedentes de la Formación Kachaike se destacan algunos elementos comunes con los de Bajo Comisión y Río Fósiles de Halle (1913a). Entre ellos: Gleichenites sanmartinii, identificada por Baldoni y Ramos (1981) en la localidad "d" (La Potranquita) y por Longobucco et al. (1985) en la localidad Cerro Mirador, Cladophlebis patagonica, citada por Longobucco et al., (1985) para la localidad La Florida y Athrotaxis ungeri y Nathorstia alata reconocidos para Ea. Kachaike por Llorens y Del Fueyo (2003) y Cúneo y Gandolfo (2005) respectivamente.
Estas dos últimas especies fueron también reconocidas por Piátnitzky (1938) para el área de lago Cardiel procedentes de los niveles inferiores de la Formación Piedra Clavada. Esta unidad sería un equivalente lateral de Formación Kachaike (Riccardi y Rolleri, 1980: 1210) o bien la suprayacería (Arbe, 2002).
En el centro-norte de Santa Cruz, en depósitos más antiguos correspondientes al Grupo Baqueró (Aptiano), se ha identificado más de un centenar de especies (Archangelsky, 1967b, 2003), en su mayoría de pteridófitas, pteridospermas, gimnospermas y uno de los más tempranos registros de angiospermas para Patagonia. Hasta el momento, la lista de especies de esta paleoflora compartidas con aquellas reconocidas para el área del lago San Martín, incluye:
Gleichenites sanmartini, Cladophlebis patagonica y Athrotaxis ungeri. A éstas se pueden agregar, si se consideran las especies originalmente identificadas por Halle (1913a): Sphenopteris patagonica Halle, Sphenopteris (Onychiopsis?) psilotoides (Stokes y Webb) Ward, Sphenopteris (Ruffordia?) goepperti Dunk (la identidad de estas dos últimas, para Río Fósiles, ha sido puesta en duda por Baldoni, 1981) y Elatocladus sp., la cual, según Archangelsky (1967b: 134), sería afín a Podocarpus? palissyafolia Berry del Grupo Baqueró. Se destaca, sin embargo, la ausencia de Nathorstia alata, Phyllopteroides cf. P. laevis y Ptilophyllum acutifolium en los depósitos baqueroenses. Más antiguas aún, las sedimentitas de la Formación Springhill (Neocomiano-Barremiano), contienen a Ticoa magallanica Archangelsky, la cual, si fuese considerada coespecífica de Pachypteris crassa (véase texto), sería un elemento en común con Bajo Comisión.
Por último, dos especies reconocidas en Bajo Comisión como Gleichenites juliensis y Cladophlebis patagonica se hallan presentes, en Patagonia, en depósitos correspondientes al Jurásico Medio (Herbst, 1962b, 1971).
Conclusiones
Se ha prospectado la zona donde Thor G. Halle (1913a) exhumó restos de plantas fósiles cretácicas, al norte del lago San Martín (provincia de Santa Cruz, Argentina), habiéndose localizado dos sitios fosilíferos: Bajo Comisión 1 y 2. Aunque ambos corresponden a la Formación Kachaike, la relación litoestratigráfica entre los depósitos de estas localidades es aún poco clara. En Bajo Comisión 1, un detallado perfil que abarca sedimentos de plataforma deltaica y canales fluviales con planicies de inundación, permitió reconocer seis niveles plantíferos, cuyo contenido megaflorístico incluye briófitas, pteridófitas, pteridospermas, bennettitales y coníferas, además de frondes de afinidad sistemática incierta y en el que se destaca la presencia de restos foliares de angiospermas.
En tanto, en Bajo Comisión 2 los niveles plantíferos son portadores de frondes de pteridófitas, ginkgoales y coníferas. Si bien no se ha reconocido el total de especies identificadas por Halle (1913a), existe una evidente similitud entre ambas listas paleoflorísticas, lo cual sugiere que estos depósitos son, en efecto, los que originalmente estudió el destacado paleobotánico sueco. Se reconocen y describen, además, nuevos taxones para la Formación Kachaike, algunos de los cuales corresponden a nuevas especies (Pachypteris auriculata sp. nov., Brachyphyllum kachaikensis sp. nov. y Thorhallenia dentata gen. et sp. nov.). También, se proponen reasignaciones sistemáticas para otros taxones (Gleichenites juliensis por Gleichenites cf. G. micromerus y Phyllopteroides cf. P. laevis por Asplenites lanceolatus) que se suman a revisiones previas del listado florístico original de Halle.
El interés de estos depósitos reside, por un lado, en la escasez de floras de esta edad para Patagonia, que se restringe a unas pocas localidades y taxones y, por otro, en la posibilidad de indagar en las características de las angiospermas durante su proceso de radiación en el Cretácico de Patagonia, como así también, en el contexto florístico en el que ello se llevó a cabo.
Agradecimientos
A S. Césari, A. Gandolfo y S. Archangelsky por la revisión crítica del manuscrito original, así como a R. Herbst y A. Troncoso, por sus comentarios en calidad de árbitros. A G. Cladera y O. Limarino por la elaboración del perfil estratigráfico de Bajo Comisión 1 (BC1). A A. González por la elaboración del mapa de ubicación. A G. del Fueyo, A. Archangelsky, G. Cladera, I. Escapa, O. Limarino y O. Cárdenas quienes participaron de los trabajos de campo. Al CONICET. Trabajo financiado por los Proyectos PICT 6044 y 10747 (CONICET).
Bibliografía
1. Aguirre-Urreta, M.B. 2002. Invertebrados del Cretácico Inferior. En: M.J. Haller (ed.), Geología y Recursos Naturales de Santa Cruz. 15º Congreso Geológico Argentino (Calafate), Relatorio: 439-459. [ Links ]
2. Appert, O. 1973. Die Pteridophyten aus dem Oberen Jura des Manamana in Südwest-Madagaskar. Schweizerische Paläontologische Abhandlungen Mémoires suisses de Paléontologie 94: 1-62. [ Links ]
3. Arbe, H.G. 2002. Análisis estratigráfico del Cretácico de la Cuenca Austral. En: M.J. Haller (ed.), Geología y Recursos Naturales de Santa Cruz. 15º Congreso Geológico Argentino (Calafate), Relatorio: 103-128. [ Links ]
4. Archangelsky, S. 1963. A new Mesozoic flora from Ticó, Santa Cruz province, Argentina. Bulletin of The British Museum (Natural History) Geology 8: 45-92. [ Links ]
5. Archangelsky, S. 1964. Notas sobre la flora fósil de la zona de Ticó, Provincia de Santa Cruz. 5. Sphenopteris cf. goepperti Dunker. 6. Cladophlebis sp. Ameghiniana 3: 280-284. [ Links ]
6. Archangelsky, S. 1966. New Gymnosperms from the Ticó Flora, Santa Cruz province, Argentina. Bulletin of The British Museum (Natural History) Geology 13: 259-295. [ Links ]
7. Archangelsky, S. 1967a. Notas sobre la Flora fósil de la zona de Ticó, provincia de Santa Cruz, 8. Seis especies del género Sphenopteris. Ameghiniana 5: 149-157. [ Links ]
8. Archangelsky, S. 1967b. Estudio de la Formación Baqueró, Cretácico Inferior de Santa Cruz, Argentina. Revista Museo de Ciencias Naturales de La Plata, Nueva Serie. Paleontología 5: 63-171. [ Links ]
9. Archangelsky, S. 1976. Vegetales fósiles de la Formación Springhill, Cretácico, en el subsuelo de la cuenca Magallánica. Ameghiniana 13: 141-158. [ Links ]
10. Archangelsky, S. 2003. La flora cretácica del Grupo Baqueró, Santa Cruz, Argentina. Monografías del Museo Argentino de Ciencias Naturales, nº 4. Buenos Aires, 14 pp. + CD. [ Links ]
11. Archangelsky, S. y Baldoni, A. 1972. Revisión de las Bennettitales de la Formación Baqueró (Cretácico inferior), Provincia de Santa Cruz. I. Hojas. Revista Museo de Ciencias Naturales de La Plata, Nueva Serie. Paleontología 7: 185-265. [ Links ]
12. Archangelsky, S. y Del Fueyo, G. 1989. Squamastrobus gen. nov., a fertile podocarp from the Early Cretaceous of Patagonia, Argentina. Review of Palaeobotany and Palynology 59: 109-126. [ Links ]
13. Arrondo, O.G. y Petriella, B. 1980. Alicurá, nueva localidad plantífera liásica de la Provincia de Neuquén, Argentina. Ameghiniana 17: 200-215. [ Links ]
14. Baldoni, A.M. 1980. Plantas fósiles jurásicas en una nueva localidad en la provincia del Neuquén. Ameghiniana 17: 255-272. [ Links ]
15. Baldoni, A.M. 1981. Análisis de algunas tafofloras jurásicas y eocretácicas de Argentina y Chile. 2º Congreso Argentino de Paleontología y Bioestratigrafía y 1º Congreso Latinoamericano de Paleontología (Buenos Aires, 1978), Actas 5: 41-65. [ Links ]
16. Baldoni, A.M. y Ramos, V. 1981. Nuevas localidades con plantas fósiles cretácicas en la Cordillera Patagónica (provincia de Santa Cruz, República Argentina). 8º Congreso Geológico Argentino (Buenos Aires), Actas 4: 472-759. [ Links ]
17. Baldoni, A.M., Askin, R.A. y Ragona, D. 2001. Palynology of the Lower Cretaceous Kachaike Fm, Santa Cruz Province, Argentina. En: D.K. Goodmanand y R.T. Clarke (eds.), Proceedings of the 9º International Palynological Congress (Houston, 1996), American Association of Stratigraphic Palynologists Foundation, pp. 191-200. [ Links ]
18. Banerji, J. 1992. Osmundaceous fronds in Lower Cretaceous beds at Chunakhal, Rajmahal Hills, Bihar, India. Alcheringa 16: 1-13. [ Links ]
19. Banerji, J. 1996. Early Cretaceous megaflora from Murlipahar, Rajmahal Basin, India. Geophytology 25: 41-46. [ Links ]
20. Barreda, V. y Archangelsky, S. 2006. The southernmost record of tropical pollen grains in the mid-Cretaceous of Patagonia, Argentina. Cretaceous Research 27: 778-787. [ Links ]
21. Bell, W.A. 1956. Lower Cretaceous floras of western Canadá. Geological Survey of Canada Memoir 285: 1-331. [ Links ]
22. Berry, E.W. 1928. Tertiary fossil plants from Argentine Republic. Proceedings of the United States National Museum 73: 1-27. [ Links ]
23. Berry, E.W. 1937. An Upper Cretaceous flora from Patagonia. En: E. Bennett Mathews (ed.), Studies in geology Nº 12. Contributions to Paleobotany of South America. Johns Hopkins Press, Baltimore, pp. 11-31. [ Links ]
24. Boureau, E. y Doubinger, J. 1975. Traité de Paléobotanique 4 (2) Filicophyta. Masson et Cie Éditeurs, Paris, 406 pp. [ Links ]
25. Bose, M.N. y Banerji, J. 1984. The fossil floras of Kachchh 1-Mesozoic megafossils. The Palaeobotanist 33: 1-189. [ Links ]
26. Bose, M.N. y Kasat, M.L. 1972. The genus Ptilophyllum in India. The Palaeobotanist 19: 115-145. [ Links ]
27. Brongniart, A. 1828. Histoire des Végéteaux Fossiles, ou Reserches Botaniques et Géologiques sur les Végétaux Renfermes dans les Divers Couches du Globe. G. Dufour y E. D'Ocagne (eds.), Paris, 488 pp. [ Links ]
28. Brongniart, A. 1849. Tableau des genres de végétaux fossiles considérés sous le point de vue de leur classification botanique et de leur distribution géologique: Dictionnaire universel histoire naturel 13: 1-127. [ Links ]
29. Cantrill, D.J. 1996. Fern thickets from the Cretaceous of Alexander Island, Antarctica containing Alamatus bifarius Douglas and Aculea acicularis sp. nov. Cretaceous Research 17: 169-182. [ Links ]
30. Cantrill, D.J. 2000. A Cretaceous (Aptian) flora from President Head, Snow Island, Antarctica. Palaeontographica B 253: 153-191. [ Links ]
31. Cantrill, D.J. y Falcon-Lang, H.J. 2001. Cretaceous (Late Albian) coniferales of Alexander Island, Antarctica. 2. Leaves, reproductive structures and roots. Review of Palaeobotany and Palynology 115: 119-145. [ Links ]
32. Cantrill, D.J. y Nagalingum, N.S. 2005. Ferns from the Cretaceous of Alexander Island, Antarctica: Implications for Cretaceous phytogeography of the Southern Hemisphere. Review of Palaeobotany and Palynology 137: 83-103. [ Links ]
33. Cantrill, D.J. y Nichols, G.J. 1996. Taxonomy and palaeoecology of Early Cretaceous (Late Albian) angiosperms leaves from Alexander Island, Antarctica. Review of Palaeobotany and Palynology 92: 1-28. [ Links ]
34. Cantrill, D.J. y Webb, J.A. 1987. A reappraisal of Phyllopteroides Medwell (Osmundaceae) and its stratigraphic significance in the Lower Cretaceous of eastern Australia. Alcheringa 11: 59-85. [ Links ]
35. Césari, S. 2006. Aptian ferns with in situ spores from the South Shetland Islands, Antarctica. Review of Palaeobotany and Palynology 138: 227-238. [ Links ]
36. Césari, S., Parica, C., Remesal, M. y Salani, F. 1999. Paleoflora del Cretácico Inferior de península Byers, islas Shetland del Sur, Antártida. Ameghiniana 36: 3-22. [ Links ]
37. Crabtree, D.R. 1987. Angiosperms of the Northern Rocky Mountains: Albian to Campanian (Cretaceous) magafossil floras. Annals of the Missouri Botanical Garden 74: 707-747. [ Links ]
38. Cúneo, R. y Gandolfo, M.A. 2005. Angiosperm leaves from the Kachaike Formation, Lower Cretaceous of Patagonia, Argentina. Review of Palaeobotany and Palynology 136: 29-47. [ Links ]
39. Del Fueyo, G.M. 1991. Una nueva Araucariaceae cretácica de la Patagonia, Argentina. Ameghiniana 28: 149-161. [ Links ]
40. Douglas, J.G. 1969. The Mesozoic floras of Victoria Parts 1 and 2. Geological Survey of Victoria, Memoirs 28: 1-310. [ Links ]
41. Douglas, J.G. 1973. The Mesozoic floras of Victoria Part 3. Geological Survey of Victoria, Memoirs 29: 1-185. [ Links ]
42. Doyle, J.A. y Endress, P.K. 2000. Morphological phylogenetic analysis of basal angiosperms: comparison and combination with molecular data. International Journal of Plants Science 161: S121-S153. [ Links ]
43. Doyle, J.A. y Hickey, L.J. 1976. Pollen and leaves from the MidCretaceous Potomac Group and their bearing on early angiosperm evolution. En: C.B. Beck (ed.), Origin and early evolution of angiosperms. Columbia University Press, New York, pp. 139-206. [ Links ]
44. Drinnan, A.N. y Chambers, T.C. 1986. Flora of the Lower Cretaceous Koonwarra Fossil Beds (Korumburra Group), South Gippsland, Victoria. En: P.A. Jell y J. Roberts (eds.), Plants and invertebrates from the Lower Cretaceous Koonwarra Fossil Bed, South Gippsland, Victoria. Association of Australasian Palaeontologists, Memoirs 3: 1-77. [ Links ]
45. Feruglio, E. 1951. Piante del Mesozoico della Patagonia. Pubblicazioni dell'Istituto Geologico della Università di Torino 1: 35-80. [ Links ]
46. Florin, R. 1940. The Tertiary fossil conifers of South Chile and their phytogeographical significance. Kungliska Svenska Vetenskapsakademiens Handlingar 19: 1-107. [ Links ]
47. Frenguelli, J. 1935. Ptilophyllum hislopi (Oldham) en los 'Mayer River Beds' del Lago San Martín. Notas del Museo de La Plata, Paleontología 1: 71-83. [ Links ]
48. Frenguelli, J. 1953. La flora fósil de la región del Alto Río Chalía en Santa Cruz (Patagonia). Notas del Museo de La Plata, Paleontología 16: 239-257. [ Links ]
49. Gee, C.T. 1989. Revision of the Late Jurassic/Early Cretaceous flora from Hope Bay Antarctica. Palaeontographica B 213: 149-214. [ Links ]
50. Goeppert, H.R. 1836. Die fossilen Farrenkräuter (Systema filicum fossilium). Nova Acta Leopoldina 17: 1-486. [ Links ]
51. Guler, M.V. y Archangelsky, S. 2002. Quistes de dinoflagelados de la Formación Kachaike (Cretácico Inferior), provincia de Santa Cruz. 8º Congreso Argentino de Paleontología y Biostratigrafía (Corrientes), Resúmenes: p. 71. [ Links ]
52. Guler, M.V. y Archangelsky, S. 2006. Quistes de dinoflagelados de la Formación Kachaike, Cretácico Inferior de la Cuenca Austral. 13º Simposio Argentino de Paleobotanica y Palinología (Bahía Blanca), Resúmenes: 41. [ Links ]
53. Halle, T.G. 1913a. Some mesozoic plant-bearing deposits in Patagonia and Tierra del Fuego and their floras. Kungliska Svenska Vetenskapsakademiens Handlingar 51: 1-58. [ Links ]
54. Halle, T.G. 1913b. The Mesozoic Flora of Graham Land. Wissenchaftliche Ergebnisse der Schwedischen Südpolar-Expedition, 1901-1903 3: 1-123. [ Links ]
55. Harris, T.M. 1964. The Yorkshire Jurassic Flora 2, Caytoniales, Cycadales and Pteridosperms. British Museum (Natural History), London, 191 pp. [ Links ]
56. Harris, T.M. 1969. The Yorkshire Jurassic Flora 3, Bennettitales. British Museum (Natural History), London, 186 pp. [ Links ]
57. Harris, T.M. 1979. The Yorkshire Jurassic Flora 5, Coniferales. British Museum (Natural History), London, 163 pp. [ Links ]
58. Heer, O. 1874. Die Kreide-Flora der Arctischen Zone. Kungliska Svenska Vetenskapsakademiens Handlingar 12: 1-138. [ Links ]
59. Heer, O. 1880. Nachträge zur Jura-Flora von Sachalin. Kungliska Svenska Vetenskapsakademiens Handlingar 18: 1-17. [ Links ]
60. Herbst, R. 1962a. Sobre las especies de Gleichenites de los sedimentos baqueroenses de Santa Cruz, Patagonia. Ameghiniana 2: 141-151. [ Links ]
61. Herbst, R. 1962b. Gleichenites juliensis n. sp. del Jurásico Medio de Santa Cruz, Argentina. Ameghiniana 2: 187-191. [ Links ]
62. Herbst, R. 1964. La flora liásica de la zona del Río Atuel, Mendoza, Argentina. Revista de la Asociación Geológica Argentina 19: 108-131. [ Links ]
63. Herbst, R. 1965. La flora fósil de la Formación Roca Blanca, prov. Santa Cruz, Patagonia. Con consideraciones geológicas y estratigráficas. Opera Lilloana 12: 7-101. [ Links ]
64. Herbst, R. 1966a. Nuevos elementos de la flora fósil de la Formación Baqueró, Santa Cruz, Patagonia. Acta Geológica Lilloana 8: 75-86. [ Links ]
65. Herbst, R. 1966b. Revisión de la flora Liásica de Piedra Pintada, prov. Neuquén, Argentina. Revista del Museo de La Plata (nueva serie), Paleontología 5:27-33. [ Links ]
66. Herbst, R. 1971. Palaeophytología kurtziana 3º, 7. Revisión de las especies argentinas del género Cladophlebis. Ameghiniana 8: 265-281 [ Links ]
67. Herbst, R. y Zalazar, E.B. 1998. Revisión de la Flora Matildense del Gran Bajo de San Julián, prov. de Santa Cruz, Argentina. Facena 14: 7-20. [ Links ]
68. Hernández, P.J. y Azcárate, V. 1971. Estudio Paleobotánico preliminar sobre restos de una tafoflora de la península Byers (Cerro Negro), Isla Livingston, Islas Shetlands del Sur, Antártica. Serie Científica del Instituto Antártico Chileno 2: 15-50. [ Links ]
69. Hickey, L.J. 1973. Classification of the arquitecture of dicotyledonous leaves. American Journal of Botany 60: 17-33. [ Links ]
70. Hickey, L.J. 1977. Stratigraphy and Paleobotany of the Golden Valley Formation (Early Tertiary) of Western North Dakota. The Geological Society of America, Memoirs 150: 157-164. [ Links ]
71. Hickey, L.J. y Doyle, J.A. 1977. Early Cretaceous fossil evidence for angiosperm evolution. The Botanical Review 43: 1-104. [ Links ]
72. Hickey, L.J. y Taylor, D.W. 1989. Reexamination of leaf architectural characters of basal angiosperms and their sister groups: implications for the origin and relationships of angiosperms. American Journal of Botany 76 (Suppl.) Abstracts: 245-246. [ Links ]
73. Hickey, L.J. y Wolf, J.A. 1975. The bases of angiosperm phylogeny: vegetative morphology. Annals of the Missouri Botanical Garden 62: 538-589. [ Links ]
74. Hünicken, M.A. 1995. Floras Cretácicas y Terciarias. En: P.N. Stipanicic y M.A. Hünicken (eds.), Revisión y actualización de la obra paleobotánica de Kurtz en la República Argentina. Atlas de la Academia Nacional de Ciencias (Córdoba) 10: 199-219. [ Links ]
75. Iglesias, A., Zamuner, A.B., Larriestra, F., Poiré, D.G. y Romero, E.J. 2004. Great diversity of angiosperms in the Late Cretaceous, Mata Amarilla Formation, Patagonia, Argentina. 7º International Organization of Paleobotany Conference (San Carlos de Bariloche), Abstracts: 58-59. [ Links ]
76. Klavins, S.D, Taylor, T.N. y Taylor, E.L. 2004. Matoniaceous ferns (Gleicheniales) from the Middle Triassic of Antarctica. Journal of Paleontology 78: 211-217. [ Links ]
77. Krassilov, V.A. 1964. On the fern genus Nathorstia. Paleontological Journal 2: 115-120 (en ruso). [ Links ]
78. Leaf Architecture Working Group. 1999. Manual of leaf architecture, morphological description and categorization of dicotyledonous and net-veined monocotyledonous angiosperms. Smithsonian Institution, Washington, 65 pp. [ Links ]
79. Longobuco, M.I., Azcuy, C.L. y Aguirre-Urreta, B. 1985. Plantas de la Formación Kachaike, Cretácico de la prov. de Santa Cruz. Ameghiniana 21: 305-315 [ Links ]
80. Lundblad, B. 1955. Contributions to the Geological History of the Hepaticae. II On a fossil member of the Marchantiineae from the Mesozoic plant-bearing deposits near Lago San Martín, Patagonia (Lower Cretaceous). Botaniska Notiser 108: 22-39. [ Links ]
81. Lundblad, B. 1971. A restudy of the Ginkgolean leaves of the Mesozoic Flora of Lago San Martín, Patagonia (Ginkgoites Skottsbergii n. sp.) Journal Indian Botanical Society Golden Jubilee 50A: 236-241. [ Links ]
82. Llorens, M. y Del Fueyo, G.M. 2003. Coníferas fértiles de la Formación Kachaike, Cretácico medio de la provincia de Santa Cruz, Argentina. Revista del Museo Argentino de Ciencias Naturales (nueva serie) 5: 241-244. [ Links ]
83. McLoughlin, S. 1996. Early Cretaceous macrofloras of Western Australia. Records of the Western Australian Museum 18: 19-65. [ Links ]
84. McLoughlin, S., Drinnan, A.N. y Rozefelds, A.C. 1995. A Cenomanian flora from the Winton Formation, Eromanga Basin, Queensland, Australia. Memoirs of the Queensland Museum 38: 273-313. [ Links ]
85. McLoughlin, S., Tosolini, A.M.P., Nagalingum, N.S. y Drinnan, A.N. 2002. Early Cretaceous (Neocomian) flora and fauna of the Lower Strzelecki Group, Gippsland Basin, Victoria. Association of Australasian Palaeontologists, Memoirs 26: 1-144. [ Links ]
86. Medwell, L.M. 1954. Fossil plants from Killara near Casterton, Victoria. Proceedings of the Royal Society of Victoria 66: 17-23. [ Links ]
87. Menéndez, C.A. 1959. Flora Cretácica de la Serie del Castillo, al Sur del Cerro Cachetamán. Revista de la Asociación Geológica Argentina 14: 219-238. [ Links ]
88. Menéndez, C.A. 1966. Fossil Bennettitales from the Tico Flora, Santa Cruz province, Argentina. Bulletin of The British Museum (Natural History) Geology 12: 1-42. [ Links ]
89. Miller, C.N. y LaPasha, C.A. 1983. Structure and affinities of Athrotaxites berryi Bell, an Early Cretaceous conifer. American Journal of Botany 70: 772-779. [ Links ]
90. Mohr, B.A.R. y Friis, E.M. 2000. Early angiosperms from the Lower Cretaceous Cretaceous Crato Formation (Brazil), a preliminary report. International Journal of Plant Science 161: S155-S167. [ Links ]
91. Morris, J. 1840. En: C.W.Grants, Memoir to illustrate the geological map of Cutch. Transactions of the Geological Society of London, Series 2, 5: 289-329. [ Links ]
92. Nagalingum, N.S. y Cantrill, D.J. 2006. Early Cretaceous Gleicheniaceae and Matoniaceae (Gleicheniales) from Alexander Island, Antarctica. Review of Palaeobotany and Palynology 138: 73-93. [ Links ]
93. Passalia, M.G., Romero, E.J. y Panza, J.L. 2001. Improntas foliares del Cretácico de Santa Cruz, Argentina. Ameghiniana 38: 73-84. [ Links ]
94. Passalia, M.G., Archangelsky, S., Romero, E.J. y Cladera, G. 2003. A new early angiosperm leaf from the Anfiteatro de Ticó Formation (Aptian), Santa Cruz Province, Argentina. Revista del Museo Argentino de Ciencias Naturales (nueva serie) 5: 245-252. [ Links ]
95. Piátnitzky, A. 1938. Observaciones geológicas en el oeste de Santa Cruz (Patagonia). Boletín de Informaciones Petroleras Yacimientos Petrolíferos Fiscales 165: 45-95. [ Links ]
96. Pole, M. 1992. Cretaceous macrofloras of Eastern Otago, New Zealand: Angiosperms. Australian Journal of Botany 40: 169-206. [ Links ]
97. Pons, D. 1988. Le Mésozoïque de Colombie macroflores et microflores. Éditions du Centre National de la Recherche Scientifique, París, 168 pp. [ Links ]
98. Puebla, G.G. 2003. Nueva angiosperma del Cretácico Inferior de la provincia de San Luis, Argentina. 29º Jornadas Argentinas de Botánica y 15º Reunión Anual de la Sociedad Botánica de Chile (San Luis), Resúmenes: 299. [ Links ]
99. Riccardi, A.C. 1971. Estratigrafía en el oriente de la Bahía de La Lancha, lago San Martín, Santa Cruz, Argentina. Revista Museo de Ciencias Naturales de La Plata, Geología 61: 245-318. [ Links ]
100. Riccardi, A.C. y Rolleri, E.O. 1980. Cordillera Patagónica Austral. En: 2º Simposio de Geología Regional Argentina (Córdoba, 1976), Academia Nacional de Ciencias (Córdoba), 2: 1173-1306. [ Links ]
101. Romero, E.J. y Archangelsky, S. 1986. Early Cretaceous angiosperms leaves from southern South América. Science 234: 1580-1582. [ Links ]
102. Ruiz, L.C. 1984. Plantas fósiles cretácicas procedentes de la zona del Lago Cardiel, provincia de Santa Cruz. 9º Congreso Geológico Argentino (San Carlos de Bariloche), Actas 4: 444-454. [ Links ]
103. Rushforth, S.R. 1970. Notes on the fern family Matoniaceae from the western United States. Brigham Young University Geology Studies 16: 1-34. [ Links ]
104. Saporta, G. 1873. Paléontologie française ou description des fósiles de France, plantes jurassiques 2. Cycadeés. París, 222 pp. [ Links ]
105. Seward, A.C. 1894. The Wealden Flora, Part 1 Tallophyta- Pteridophyta. Catalogue of the Mesozoic plants in the Department of Geology, British Museum (Natural History). London, British Museum, 179 pp. [ Links ]
106. Seward, A.C. 1919. Fossil Plants Volume 4. Cambridge Biological Series. London, 542 pp. [ Links ]
107. Seward, A.C. 1926. The Cretaceous plant-bearing rocks of western Greenland. Philosophical Transactions of the Royal Society of London 215B: 57-174. [ Links ]
108. Spicer, R.A. 1986. Comparative leaf architectural analysis of Cretaceous radiating angiosperms. En: R.A. Spicer y B.A. Thomas (eds.), Systematic and taxonomic approaches in palaeobotany, The Systematic Association, Special Volume 31, Oxford, pp. 221-232. [ Links ]
109. Sternberg, K.G. 1825. Versuch einer geognostisch-botanischen Darstellung der Flora der Vorwelt Teil 4, Leipzig, pp. 1-48. [ Links ]
110. Surange, K.R. 1966. Indian Fossil Pteridophytes. Botanical Monograph nº 4. Council of Scientific and Industrial Research. New Delhi, 209 pp. [ Links ]
111. Todzia, C.A. y Keating, R.C. 1991. Leaf architecture of the Chloranthaceae. Annals of the Missouri Botanical Garden 78: 476-496. [ Links ]
112. Townrow, J.A. 1965. A new member of the Corystospermaceae Thomas. Annals of Botany n.s. 29: 495-510. [ Links ]
113. Traverso, N.E. 1966. Brachyphyllum tigrense nueva conífera de la Formación Baqueró, Cretácico de Santa Cruz. Ameghiniana 4: 189-194. [ Links ]
114. Traverso, N.E. 1968. Brachyphyllum baqueroense otra nueva conífera de la Formación Baqueró, Cretácico de Santa Cruz. Ameghiniana 5: 374-378. [ Links ]
115. Van Konijnenburg-van Cittert, J.H.A. 1993. A review of the Matoniaceae based on in situ spores. Review of Palaeobotany and Palynology 78: 235-267. [ Links ]
116. Watson, J. 1969. A revision of the English Wealden Flora, I Charales-Ginkgoales. Bulletin of the British Museum (Natural History) Geology 17: 210-254. [ Links ]
117. Wolfe, J.A. 1989. Leaf architectural analysis of the Hamamelididae. En: P.R.Crane y S. Blackmore (eds.), Evolution, Systematics, and Fossil History of the Hamamelidae 1: Introduction and "Lower" Hamamelidae, Systematics Association Special 40A, Clarendon Press, Oxford, pp. 75-104. [ Links ]
Recibido: 15 de noviembre de 2005.
Aceptado: 17 de julio de 2007.

