










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.44 n.3 Buenos Aires jul./sep. 2007
Trilobites de la Formación Volcancito (Miembro Filo Azul, Cámbrico Tardío) del Sistema de Famatina, La Rioja, Argentina: aspectos sistemáticos y paleoambientales
M. Franco Tortello1 y Susana B. Esteban2
1CONICET. División Paleontología Invertebrados, Museo de Ciencias Naturales, Paseo del Bosque s/n°, 1900 La Plata, Argentina. tortello@museo.fcnym.unlp.edu.ar
2Instituto Superior de Correlación Geológica, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán, Miguel Lillo 205, 4000 Tucumán, Argentina. susana_esteban2003@yahoo.com.ar
Resumen. El Miembro Filo Azul de la Formación Volcancito (sierra de Famatina, La Rioja, Argentina) consiste de un tramo inferior (30 m) de margas y lutitas calcáreas finamente laminadas con intercalaciones de areniscas finas, un tramo medio (85 m) de areniscas carbonáticas medianas a gruesas masivas, y un tramo superior (50 m) con pelitas que alternan con bancos de areniscas muy finas. Estudios previos sobre trilobites, graptolitos y conodontes asignaron la sección al Cámbrico Superior-Ordovícico Inferior. El tramo inferior del miembro (Biozona de Parabolina frequens argentina, sector inferior) contiene una profusa fauna de trilobites estudiada por Harrington y Leanza a mediados del siglo pasado. Desde entonces, se han obtenido nuevas colecciones, las cuales motivan este estudio. Se reilustran poliméridos originalmente descriptos con material escaso (Onychopyge riojana Harrington, Asaphellus riojanus Harrington y Leanza, Rhadinopleura eurycephala Harrington y Leanza, Plicatolina scalpta Harrington y Leanza) y se reconsidera la identidad de varios taxones (Angelina cf. hyeronimi (Kayser), Parabolinella sp. 1, Parabolinella sp. 2, Pseudokainella tosellii sp. nov., Conophrys sp.) (los trilobites agnóstidos, revisados recientemente, se excluyen del estudio). La fauna comprende varias especies endémicas. A su vez, el registro de Parabolina (Neoparabolina) frequens (Barrande) permite efectuar correlaciones con otras secciones del Cámbrico Superior. La abundancia de nutrientes en el fondo marino habría favorecido el desarrollo de una comunidad de trilobites diversa, con diferentes morfotipos y hábitos alimenticios. Varios taxones estaban especialmente adaptados a vivir en un ambiente con escaso oxígeno, algunos en probable simbiosis con bacterias del azufre.
Abstract. Trilobites from the Volcancito Formation (Filo Azul Member, Late Cambrian) of the Famatina System, La Rioja, Argentina: systematics and paleoenvironments. The Filo Azul Member of the Volcancito Formation (Famatina range, La Rioja, Argentina) consists of a lower part (30 m) of thinly laminated marls and calcareous shales with intercalations of fine-grained sandstones, a middle part (85 m) of massive middle to thick-grained calcareous sandstones, and an upper part (50 m) of shales with interbedded layers of fine sandstones. Previous studies on trilobites, graptolites and conodonts assigned the section to the Upper Cambrian-Lower Ordovician. The lower portion of the member (Parabolina frequens argentina Biozone, lower part) contains a diverse trilobite fauna that was studied by Harrington and Leanza in the 1950s. Since then, new collections have been obtained, which are the subject of the present study. New polymerid trilobites, originally described on the basis of scarce material, are re-illustrated (Onychopyge riojana Harrington, Asaphellus riojanus Harrington and Leanza, Rhadinopleura eurycephala Harrington and Leanza, Plicatolina scalpta Harrington and Leanza) and the assignations of several taxa are reconsidered (Angelina cf. hyeronimi (Kayser), Parabolinella sp. 1, Parabolinella sp. 2, Pseudokainella tosellii sp. nov., Conophrys sp.) (since the agnostoids have been recently revised, they are not included in this study). The fauna comprises several endemic species. On the other hand, the occurrence of Parabolina (Neoparabolina) frequens (Barrande) permits the outcrop to be correlated with other Upper Cambrian sections. The abundance of nutrients in the sea bottom must have favoured the development of a diverse trilobite community, which included different morphotypes and feeding habits. Several taxa were especially adapted to live under low-oxygen conditions, probably in symbiosis with sulfur bacteria.
Key words. Trilobites; Late Cambrian; Volcancito Formation; Argentina; Systematics; Paleoenvironment.
Palabras clave. Trilobites; Cámbrico Tardío; Formación Volcancito; Argentina; Sistemática; Paleoambiente.
Introducción
El Cámbrico Superior-Ordovícico basal del Sistema de Famatina (provincia de La Rioja) se encuentra bien representado en la sección Río Volcancito, en el Miembro Filo Azul de la Formación Volcancito (Harrington in Harrington y Leanza, 1957; Esteban, 1999, 2003a; Albanesi et al., 2005). Las primeras referencias de fósiles en esta localidad se remontan al inicio del siglo XX, ocasión en la que Bodenbender (1916) citó "calizas" con braquiópodos ("Obolus") y trilobites ("Agnostus"). Posteriormente Harrington (1938) estudió colecciones obtenidas por A. Flossdorf en 1913 que incluían una mayor diversidad de braquiópodos y trilobites (agnóstidos, olénidos, shumárdidos, asáfidos, ceratopígidos y kainélidos). Estas faunas, sumadas a nuevas colecciones obtenidas por H. Harrington en la década de 1940, fueron revisadas por Harrington y Leanza (1957) y asignadas a la Biozona de Parabolina (Neoparabolina) frequens argentina. Harrington y Leanza (1957) sentaron las bases del conocimiento del afloramiento, al reconocer varios géneros de trilobites (ej. Micragnostus Howell, Machairagnostus Harrington y Leanza, Gymnagnostus Robison y Pantoja-Alor, Rhadinopleura Harrington y Leanza, Onychopyge Harrington, Parabolina Salter, Parabolinella BrØgger, Plicatolina Shaw, Angelina Salter, Asaphellus Callaway, Shumardia Billings) y establecer afinidades faunísticas con la Cordillera Oriental.
Estudios recientes demostraron que la transición Cámbrico-Ordovícico se encuentra bien representada en Río Volcancito. Las margas de los tramos inferiores de la sección contienen agnóstidos característicos del Cámbrico Tardío [ej. Lotagnostus Whitehouse, Pseudorhaptagnostus (Machairagnostus) Harrington y Leanza, Gymnagnostus bolivianus (Hoek), Strictagnostus Shergold] asociados a numerosos trilobites poliméridos, mientras que las pelitas de los tramos superiores fueron asignadas al Ordovícico basal en base al registro del olénido Jujuyaspis keideli Kobayashi y el graptolito Rhabdinopora flabelliformis socialis (Salter) (Tortello y Esteban, 1997, 1998, 1999). El estudio de las conodontofaunas de la sección reveló que el límite Cámbrico-Ordovícico se sitúa a 85 m. de la base del miembro, signado por la primera aparición de Iapetognathus aengensis (Lindström) (Albanesi et al., 1999).
La mayoría del material estudiado por Harrington (1938) y Harrington y Leanza (1957) proviene de los tramos cámbricos de la sección, aflorantes directamente sobre el cauce del río Volcancito. En la década de 1970 Toselli (1977) obtuvo numerosos ejemplares adicionales de esos niveles que depositó en la Universidad Nacional de Tucumán, a los que se le sumó nuevo material obtenido por los autores. En el presente trabajo se dan a conocer estas colecciones, haciendo hincapié en los trilobites poliméridos (los trilobites agnóstidos, revisados por Tortello y Esteban, 1999, se excluyen de este estudio). De esta forma se amplía el conocimiento sistemático de algunos taxones originalmente descriptos con ejemplares escasos, y se proveen nuevos criterios para interpretar el ambiente de depositación y las implicancias paleoecológicas del material.
El Miembro Filo Azul de la Formación Volcancito
El Miembro Filo Azul de la Formación Volcancito se encuentra ubicado sobre la margen izquierda del río Volcancito, 2000 m aguas abajo del puesto homónimo, a 24 km al oeste de la localidad de Famatina, provincia de La Rioja (figura 1). En esta unidad se distinguen de base a techo tres intervalos con características sedimentológicas diferentes que permiten inferir importantes cambios en las condiciones ambientales durante su depositación (Esteban, 1999; Tortello y Esteban, 1999) (figura 2). Las facies 1 y 2 mencionadas en el presente trabajo surgen de una reevaluación de las facies de grano fino del tramo inferior de la secuencia descriptas en trabajos anteriores, siendo equivalentes a la "Facies A: margas finamente laminadas" de Tortello y Esteban (1999).
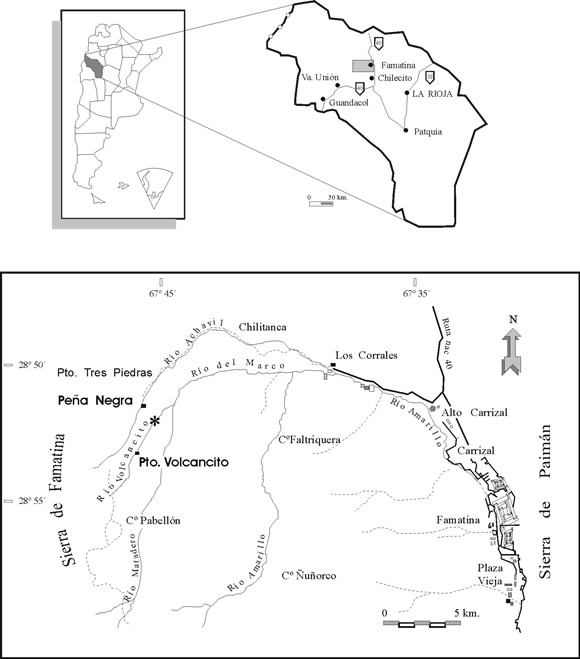
Figura 1. Mapa de ubicación / location map.

Figura 2. Columna estratigráfica del Miembro Filo Azul (Formación Volcancito) en río Volcancito, con las facies y los rangos estratigráficos de las faunas de trilobites / columnar section of the Filo Azul Member (Volcancito Formation) at Río Volcancito, showing facies and distribution of trilobites.
El intervalo inferior (Cámbrico Superior), de donde procede la fauna de trilobites estudiada, consiste de unos 30 m en los que predominan margas con laminación paralela (facies 1) y lutitas calcáreas con laminación ondulosa (facies 2). Ambos tipos de rocas se caracterizan por tener una coloración oscura, debido a su contenido relativamente alto de materia orgánica y sulfuros de hierro. Estudios al microscopio petrográfico han revelado que la laminación de estos sedimentos de grano fino obedece a la alternacia rítmica de capas claras ricas en silicatos y/o carbonatos y capas oscuras dominantemente orgánicas con material clástico subordinado. La laminación de la facies 2 está caracterizada por láminas que muestran superficies onduladas, con una longitud de onda de aproximadamente 1 mm y espesores que varían entre 0.1 y 1.3 mm (figura 3.1). Las láminas claras están constituidas por cuarzo, plagioclasa, delgadas láminas de muscovita flexurada y abundante material arcilloso identificado mediante Rayos X como caolinita. Las láminas oscuras presentan un material opaco dominante (materia orgánica no identificada) y material clástico subordinado correspondiente a pequeños granos de cuarzo, plagioclasa, muscovita y material arcilloso (caolinita) (figura 3.2). También se observa la presencia subordinada de material carbonático finamente dividido (micrita) y escasos cristales de opacos que corresponden a pirita, distribuidos irregularmente formando cristales euhedrales y subhedrales. La facies 1, con laminación paralela, se origina por decantación de material en suspensión, mientras que la facies 2, con laminación ondulosa, obedece a una depositación a partir de un régimen de flujo más alto representado por las láminas clásticas, que alterna con momentos durante los cuales la razón de sedimentación es extremadamente baja. El color oscuro de estas facies, la ausencia de bioturbación y el alto contenido de materia orgánica (especialmente en la facies 2) reflejan un bajo contenido de oxígeno en el fondo marino.
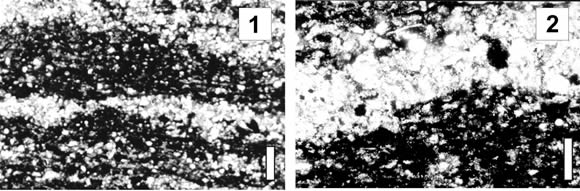
Figura 3. Fotografías de secciones delgadas de las lutitas calcáreas con laminación ondulosa de la facies 2 / thin section photomicrographs of calcareous shale with wavy lamination of facies 2. 1, Láminas claras ricas en cuarzo intercaladas con láminas oscuras con abundante materia orgánica / light quartz-rich laminae alternating with dark organic-rich laminae. Barra / bar: 0,14 mm; 2, Láminas oscuras donde se pueden observar los escasos granos de cuarzo incluidos en la materia orgánica, con sus ejes mayores paralelos a la estratificación / dark laminae showing single quartz grains within organic material with their longest axes parallel to the bedding planes. Barra / bar: 0.07 mm.
La depositación del material pelítico (facies 1 y 2) fue interrumpida en distintos momentos por depósitos más gruesos de génesis episódica, representados por bancos de areniscas limosas interestratificados (facies 3). Estas capas de evento presentan una estratificación delgada (2 a 10 cm de espesor), contactos netos y geometría tabular; generalmente son masivas en la base y finamente laminadas hacia el techo, en algunos casos con ondulitas asimétricas. La distribución de las estructuras internas en estos bancos indica una depositación a partir de un flujo unidireccional episódico y de corta duración asimilable a los originados por corrientes de turbidez inducidas por tormentas. La falta de estructuras indicadoras de retrabajo por oleaje permite inferir una depositación por debajo del nivel de olas de tormenta (Esteban, 1999; Tortello y Esteban, 1999).
En el intervalo medio del perfil, de 85 m de espesor, predomina una fracción más gruesa compuesta por areniscas carbonáticas grises muy finas a gruesas (facies 4 y 5), dispuestas en bancos de 5 a 20 cm de espesor, con escasas intercalaciones de lutitas y margas. Las capas arenosas de litología más gruesa son generalmente continuas, con bases netas y techos con suave ondulación, y no presentan estructura interna. Las mismas contienen abundantes fragmentos de trilobites y braquiópodos indeterminados. La falta de estructuras sedimentarias internas indica un modo de emplazamiento rápido posiblemente a partir de flujos gravitatorios, mientras que los topes ondulados sugieren una interacción con retrabajo oscilatorio por tormentas (Esteban, 1999). Por su parte, las capas arenosas de grano fino a muy fino, que a veces gradan a limolitas de grano grueso, presentan estratificación entrecruzada hummocky, lo que indica una de positación generada por tormenta. El espesor reducido de estos estratos (5 a 15 cm), la escasa amalgamación y la ausencia de ondulitas de olas en el techo sugieren una posición distal (tempestitas distales), ligeramente por encima del nivel de olas de tormenta (véase Brenchley et al., 1986). El límite Cámbrico-Ordovícico se sitúa en este intervalo, a 85 m de la base del miembro (Albanesi et al., 1999).
El intervalo superior del perfil (Ordovícico Inferior) comprende 50 m de pelitas verde grisáceas generalmente con laminación fina (facies 6) y delgados bancos interestratificados de areniscas de grano fino. En este tramo predomina la depositación por decantación a partir de material fino en suspensión, mayormente en ausencia de corrientes y olas. La presencia de bancos arenosos intercalados indica que el proceso de decantación alternaba con momentos de sedimentación episódica, a partir de corrientes inducidas por tormentas.
Paleoambiente y paleoecología
Ambiente de depositación
Las características sedimentológicas del Miembro Filo Azul sugieren un ambiente de plataforma externa, con transiciones hacia zonas más someras inmediatamente encima de la base de olas de tormenta (figura 4). A través de la secuencia se observa un cambio importante en la coloración, especialmente de los sedimentos finos, con depósitos mayormente oscuros hacia el tramo inferior y medio y verdosos en la parte superior. Esta diferencia de color estaría relacionada con variaciones en las condiciones de oxigenación de la cuenca, con los niveles más bajos de oxígeno hacia la base, donde se preservan mayores proporciones de materia orgánica. La presencia de abundante pirita y la ausencia de bioturbación en este tramo del perfil, son evidencias que corroboran la existencia de un bajo nivel de oxígeno, tanto en el sedimento como posiblemente en las aguas de fondo (Esteban, 2003b). El sulfuro de hierro presente, que constituye tanto cristales euhedrales a subhedrales como framboides de pirita, puede corresponder a una mayor actividad bacteriana y mayor aporte de sulfatos en el agua de mar. En el tramo superior de la secuencia, donde los depósitos se tornan más verdosos y el contenido orgánico es menor, es probable que las concentraciones de oxígeno disuelto hayan sido mayores.
La materia orgánica presente en las facies finas del Miembro Filo Azul ha sido considerada tanto de origen planctónico como bentónico (Esteban, 2003b). En el primer caso, las partículas orgánicas se acumulan por decantación formando láminas oscuras delgadas dispuestas paralelamente, las cuales son observadas en la mayoría de los niveles margosos. En tanto que el desarrollo de tapetes microbiales en el fondo marino es considerado como una causa probable de la laminación ondulosa documentada en el tramo inferior del perfil. El modelo de tapetes microbiales ha sido utilizado por diversos autores para explicar la laminación ondulosa en facies lutíticas tanto modernas (Bauld, 1981) como antiguas (Kauffman, 1981; Schieber, 1986, 1999; O´Brien y Slatt, 1990; O´Brien, 1996).

Figura 4. Modelo depositacional del Miembro Filo Azul / depositional model of the Filo Azul Member.
Desarrollo de matas microbiales: generalidades e implicancias
Las matas microbiales bentónicas desarrollaron un papel importante en la formación de sedimentos carbonosos durante el Proterozoico, debido a que en ese momento la biosfera estaba dominada por microorganismos (Schieber, 1986). Las matas o tapetes microbiales están formados por un entramado de filamentos bacteriales que recubren distintos tipos de sustrato. Normalmente presentan una superficie gelatinosa en la que pueden quedar adheridos sedimentos de grano fino. Una vez que el tapete queda cubierto por sedimentos, los filamentos crecen hacia arriba originando una nueva mata. La repetición de este proceso da lugar a un sedimento caracterizado por una fina laminación (Noffke et al., 2001).
Si bien la mayoría de los ejemplos estudiados corresponden a facies carbonáticas, se han descripto tapetes microbiales en ambientes clásticos terrígenos, los cuales corresponden principalmente a zonas marinas someras (e.g., Hagardon y Bottjer, 1997; Schieber, 1986, 1998; Noffke, 2000; Noffke et al., 2002). También se ha mencionado el desarrollo de matas microbiales en ambientes de mayor profundidad, por debajo de la zona fótica (Schieber, 1999) y en lugares donde el contenido de oxígeno es relativamente bajo (Oschmann, 2000).
El hallazgo de filamentos microbiales en posición de vida es un hecho infrecuente debido a que éstos son fácilmente degradados (Schieber, 1999). Por tal razón, la identificación y el estudio de tapetes en ambientes antiguos suele basarse en determinados rasgos que han quedado preservados en el sedimento como indicio de una actividad microbial en el momento de la depositación. Así por ejemplo, láminas wavy-crinkly, estructuras de sobreplegamiento y capas con sulfuros de hierro han sido consideradas evidencia de la colonización del sustrato por microorganismos (Schieber, 1998, 1999). Las estructuras wrinkle, estudiadas en detalle por Hagadorn y Bottjer (1997, 1999) y Pflüger (1999), también son interpretadas como típicas formas inducidas microbialmente. Noffke et al. (2001) investigaron en detalle distintas estructuras formadas por la actividad microbial, determinando que las mismas obedecen a un comportamiento diferente de la colonia según sea la dinámica sedimentaria que prevalece en el medio.
Otra evidencia que ha sido mencionada por Oschmann (2000) como indicadora de actividad bacterial es la presencia de precipitados, tales como sulfuros de hierro, interpretados como productos de la actividad metabólica de ciertas bacterias (e.g., reductoras del hierro y del sulfato).
La presencia de finas láminas onduladas oscuras en algunos niveles del tramo inferior de la sección Río Volcancito (facies 2) puede ser consecuencia del desarrollo de tapetes microbiales en el fondo marino. Aunque la laminación ondulada por sí sola no es un indicador seguro de la presencia de tapetes, la observación de secciones delgadas ha permitido determinar otras características que sugieren este origen. Por ejemplo, se ha identificado la abundancia de material opaco, que no reacciona ante la luz polarizada, dispuesto en capas del tipo de las láminas wavy-crinkly. Schieber (1986, 1998) describió este tipo de laminación en las lutitas limosas carbonosas de la Formación Newland en el Proterozoico medio de Montana, consistente en la alternancia de láminas carbonosas onduladas, "drapes" de una lutita arcillosa dolomítica y delgados lentes de limo (Schieber, 1986: figs. 3, 6, 7 y 8; Schieber, 1998: fig. 4), similares a las descriptas en matas microbiales modernas. Las capas de lutitas carbonosas pueden contener hasta un 30 o 40% de dolomita finamente cristalina. Por su parte, la microfábrica de la facies 2 (figura 3.1) muestra similitudes con la fábrica de tapetes cianobacteriales ilustrada por Noffke (2000: fig. 3.c) en el Ordovícico Inferior de la Montaña Negra (Francia), en donde se destaca el desarrollo de láminas milimétricas oscuras, onduladas, que presentan pequeños granos minerales (cuarzo) inmersos aisladamente en una matriz opaca, y láminas claras detríticas constituidas mayormente por granos de cuarzo en una matriz arcillosa. La disposición de los granos de cuarzo en las capas orgánicas, con sus ejes mayores generalmente orientados paralelos a la estratificación (figura 3.2), es otro rasgo que muestra que la materia carbonosa de esta facies no se habría acumulado por un proceso lento de decantación, ya que de haber sido así los granos de cuarzo presentarían una disposición al azar tal como se observa en las láminas claras detríticas (figura 3.2) (compárese con Noffke et al., 1997: fig. 3.2). De acuerdo a Noffke et al. (1997), la orientación preferencial de los granos de cuarzo por rotación de los mismos es factible dado que éstos se encuentran dentro de una matriz orgánica blanda, asignable a un tapete microbial.
A lo mencionado anteriormente podemos agregar que el estudio al microscopio electrónico ha permitido identificar, en muestras de la facies 2, la presencia de tubos inferiores a 20 µm de diámetro, de forma continua (figura 5.1) o bien adelgazados en distintos tramos (figura 5.2); sus superficies son lisas, aunque en algunos lugares se puede observar la incrustación de diminutos cristales minerales (figura 5.2). Estos cuerpos semejan filamentos de cianobacterias recientes tales como los ilustrados por Merz-Preiß (2000: figs. 1a y 3) y Gerdes et al. (1991, 2000). Los filamentos se disponen de una manera entrelazada (figura 5.1) o formando un ramillete de tubos en posición vertical (figura 5.2) que también recuerda la disposición de los filamentos de cianobacterias bentónicas actuales, perpendiculares a la superficie del tapete (véase Noffke et al., 2001: fig. 7.A). Por su parte, en la Facies 2 se han observado agregados microscópicos de sulfuro de hierro formando microesferas menores a 100 µm, constituidas por numerosos cristales diminutos de pirita (piritas framboidales). La formación de sulfuro de hierro en el medio sedimentario es considerado como un producto de desecho metabólico de ciertas bacterias (e.g., Desulfovibrio, Desulfobacter, entre otras), las cuales en muchos casos forman matas filamentosas (Oschmann, 2000).

Figura 5. Fotografías al microscopio electrónico (SEM) de las capas ricas en materia orgánica de la facies 2 / scanning electron photomicrographs of the organic-rich laminae of facies 2. 1, Entramado de filamentos microbiales / framework of microbial filaments. Barra / bar: 10 µm; 2, Filamentos microbiales orientados perpendicular a la superficie del tapete / microbial filaments oriented perpendicular to the mat surface. Barra / bar: 100 µm.
La presencia de algas calcáreas asignadas a los géneros Nuia y Girvanella ha sido citada por Astini (2001) para las facies más carbonáticas del Miembro Filo Azul. Según dicho autor, estas formas son abundantes en aquellos tramos de la secuencia donde el porcentaje de sedimentos siliciclásticos disminuye notablemente y comienzan a intercalarse rítmicamente las calcarenitas entre las margas laminadas. Nuia consiste en varas rectas, curvas, sinuosas o irregulares con un diámetro variable entre 0,07 y 0,3 mm, que se encuentran como individuos dispersos dentro del sedimento. En tanto que Girvanella se presenta como masas de tubos simples, muy pequeños (5 a 25 µm de diámetro), marcadamente sinuosos y entrelazados constituyendo fragmentos coloniales o restos de tapices (Astini, 2001).
Consideraciones paleoecológicas
La abundante materia orgánica disponible en el medio habría fomentado el desarrollo de una comunidad de trilobites compleja, con una alta diversidad taxonómica y diferentes morfotipos y hábitos alimenticios. Tal como indicaran Baldis y González (1986), los agnóstidos de río Volcancito son particularmente diversos (Tortello y Esteban, 1999, 2005). Los mismos incluyen Lotagnostus (Lotagnostus) sp., Micragnostus calviformis Harrington y Leanza, M. vilonii Harrington y Leanza, Strictagnostus micropeltis (Harrington y Leanza), Pseudorhaptagnostus (Machairagnostus) tmetus Harrington y Leanza, P. (M.) corrugatus (Suárez-Soruco), P. (Machairagnostus) sp., Gymnagnostus perinflatus (Harrington y Leanza) y G. bolivianus, las que caracterizan la Subzona de Pseudorhaptagnostus (Machairagnostus)-Gymnagnostus (Tortello, 2003) (figura 2). Aunque aún se debate intensamente sobre los posibles hábitos de vida de los agnóstidos (véase Fortey, 1985; Fortey y Owens, 1999; Tortello y Esteban, 2005), Robison (1972) postuló una vida pelágica basado en un estudio morfológico funcional, remarcando además la existencia de numerosas especies cámbricas de amplia distribución geográfica en ambientes de plataforma externa y talud. Estudios de detalle de Agnostus pisiformis (Linnaeus) y Trinodus elspethi (Raymond) del Cámbrico de Suecia y el Ordovícico de América del Norte respectivamente, indicaron que los agnóstidos habrían desarrollado la capacidad de nadar o flotar, parcialmente enrollados, en ámbitos próximos a un fondo marino rico en nutrientes ("flocculent zone") (Müller y Walossek, 1987; Bruton y Nakrem, 2005). En tal sentido, la alta diversidad de especies documentada en los niveles de margas oscuras del tramo inferior del perfil de río Volcancito avala tal hipótesis. Dado que presentan un tamaño reducido y un hipostoma desconectado del doblez ventral, la dieta de estos trilobites habría consistido en partículas orgánicas diminutas, incluyendo algas, detritos y organismos microscópicos (Fortey y Owens, 1999).
Las faunas de olénidos también encontraron un ámbito favorable para su desarrollo en esta facies, signada por escasa energía, baja presión de predación y abundante alimento disponible. Varios géneros de hábito bentónico presentan una morfología que puede interpretarse como una adaptación especial a condiciones de escaso oxígeno (Fortey, 1985). Parabolinella y Plicatolina poseen un exoesqueleto poco convexo y un tórax con un elevado número de segmentos y pleuras anchas (morfotipo "Olenus", Henningsmoen, 1957), sugiriendo la presencia de branquias numerosas, de buen desarrollo. Parabolinella sp. 1 muestra además un exoesqueleto particularmente delgado, aristas oculares de buen desarrollo, y numerosas aristas y surcos radiales sobre el campo preglabelar, caracteres que son compatibles con un hábito bentónico en ambientes exaeróbicos, ya documentados en otros olénidos [ej., Parabolinella? sp., del Cámbrico Superior de Gales (Rushton, 1982: lám. 3, figs. 23-26); Leptobenthos roartei Tortello y Esteban, del Tremadociano superior de Famatina (Tortello y Esteban, 2003a: text-figs. 4.G-H)]. Probablemente esta familia incluía formas detritívoras que se alimentaban de partículas de tamaño algo mayor que el de las partículas consumidas por los agnóstidos (Fortey y Owens, 1999), hecho que habría facilitado la coexistencia, en un ambiente muy especializado, de formas con estrategias alimentarias similares. Recientemente, Fortey (2000) ha postulado una posible relación simbiótica entre los trilobites olénidos y bacterias quimioautótrofas del azufre, las cuales podrían haber sido cultivadas sobre la membrana existente debajo de las pleuras y/o sobre los apéndices de aquéllos, en especial sobre las lamelas branquiales de los exopoditos. Esta relación simbiótica habría permitido el desarrollo de estos trilobites en ambientes con muy bajos niveles de oxígeno y altas concentraciones de sulfuro de hidrógeno, como es el caso aquí estudiado.
Entre los hapalopléuridos descriptos en la asociación, Rhadinopleura puede asimismo ser interpretado como una forma adaptada a vivir en ambientes disaeróbicos, por cuanto presenta un axis torácico extremadamente angosto (tr.) y numerosas pleuras expandidas lateralmente. Aunque algunos autores le asignan un posible hábito pelágico (Babcock y Smith, 2003), su morfología es compatible con un hábito bentónico, teniendo en cuenta además su escasa convexidad y el pobre desarrollo del aparato ocular. Se conocen otros representantes de la familia (Hapalopleura longirachis Tortello y Esteban, 2003a; H. aff. longicornis Harrington y Leanza, 1957) formando parte de una asociación atelóptica ("atheloptic assemblage") en otra unidad del Sistema de Famatina (Formación Bordo Atravesado, Tremadociano superior), asociados a trilobites bentónicos de ojos reducidos, característicos de un ambiente de plataforma externa profunda (Tortello y Esteban, 2003a).
Las asociaciones descriptas en el Miembro Filo Azul se completan con asáfidos, remopléuridos y ceratopígidos. Asaphellus riojanus constituye un elemento realmente abundante, representando alrededor de un tercio de los ejemplares coleccionados. Como es común en la familia, esta especie exhibe ocho segmentos torácicos; a su vez, cabe señalar que el axis del tórax es proporcionalmente angosto (tr.) y sus pleuras de buen desarrollo lateral, en especial si se compara con otras formas del género, como por ejemplo la especie tipo A. homfrayi (Salter). Asaphellus riojanus posee un hipostoma conspicuo, el cual habría facilitado la manipulación de partículas de alimento de tamaño mayor, relacionado con un hábito carroñero y/o predador. El género Onychopyge es otro polimérido bien representado en la asociación, que también se ha descripto en pelitas oscuras del Cámbrico Superior de la Cordillera Oriental (Zeballo y Tortello, 2005).
Otros géneros bentónicos de río Volcancito son taxones característicos de ambientes profundos de sustrato blando (Conophrys sp.; e.g., Fortey y Rushton, 1980; Fortey y Owens, 1987), mientras que algunos, de posible hábito nectobentónico (Fortey, 1985), se conocen de ambientes más diversos (Parabolina frequens) (véase abajo).
Observaciones bioestratigráficas y paleobiogeográficas
Parabolina (Neoparabolina) frequens es un nuevo elemento de amplia distribución que permite establecer correlaciones con perfiles clásicos de la Cordillera Oriental (Formaciones Santa Rosita, Lampazar y equivalentes; Biozona de Parabolina frequens argentina - tramo inferior). Entre las secciones mejor conocidas de este ámbito se encuentran las descriptas sobre el flanco occidental de la sierra de Cajas, en la provincia de Jujuy (Tortello y Esteban, 2003b). Tortello y Esteban (1999, 2003b) y Tortello (2003) correlacionaron las mismas con el perfil de río Volcancito sobre la base de las faunas de trilobites agnóstidos. Los tramos inferiores de la Formación Lampazar representan un ambiente de plataforma externa con facies de lutitas negras laminadas y condiciones de escasa energía y bajo oxígeno disuelto en el medio. En dichos niveles se documentan agnóstidos y olénidos acompañados por el shumárdido Akoldinioidia Zhou y Zhang, mientras que en Famatina se agregan asáfidos, ceratopígidos y remopleuridáceos, colonizadores de un medio que dio lugar a un rango amplio de adaptaciones en distintos niveles tróficos. Una asociación diversa, dominada por Agnostida, Olenidae, Ceratopygidae, Asaphidae, Pliomeridae, Catillicephalidae, Calymenidae, Lonchocephalidae y ?Conocoryphidae, también fue descripta por Benedetto (1977) en la región de Cajas, en tramos de la Formación Santa Rosita asignables al Cámbrico Superior.
La fauna estudiada en este trabajo es asimismo comparable a la documentada en la Formación Las Vicuñas en la Puna argentina (Moya et al., 1993). Esta unidad está compuesta por piroclastitas y lutitas negras con intercalaciones de calizas y coquinitas. Aunque no se conocen olénidos de esta formación, Asaphellus aff. communis, Onychopyge sp., "Kainella" sp. y Conophrys aff. alata (véase Moya et al., 1993) muestran afinidades con el material de Famatina.
El trilobite Parabolina frequens permite establecer relaciones con otros ámbitos. En tal sentido, se destacan las similitudes faunísticas y paleoambientales del Miembro Filo Azul con la Formación Tiñu, del Cámbrico Superior-Ordovícico Inferior de Oaxaca, Méjico. La parte inferior de la Formación Tiñu es una sucesión mixta clástico-carbonática, formada por calizas y margas negro azuladas, con lutitas y limolitas negras y pardo verdosas interestratificadas (Robison y Pantoja-Alor, 1968; Armella y Cabaleri, 1984). Dicha unidad se depositó en una cuenca profunda de aguas relativamente cálidas, con alto contenido de pirita idiomorfa y aglomeraciones globulares ferruginosas, las cuales pueden indicar una mayor actividad bacteriana (Armella y Cabaleri, 1984). Buena parte de los géneros documentados en la Formación Tiñu se registran en río Volcancito (Micragnostus, Gymnagnostus, Pseudorhaptagnostus, Parabolina (Neoparabolina), Angelina, Parabolinella, Plicatolina, Asaphellus, Onychopyge, Pseudokainella, Conophrys). Estos taxones, de amplia distribución geográfica, suelen aparecer asociados conformando comunidades de alta diversidad en ambientes ricos en nutrientes. Similares morfotipos (Lotagnostus, Micragnostus, Pseudorhaptagnostus, Parabolina frequens, Parabolinella, Plicatolina, Pseudokainella, Conophrys, Asaphidae, Hapalopleuridae) también se conocen en el Cámbrico Superior alto (Zonas de Peltura scarabaeoides y Acerocare) de Gales, en las calizas micríticas oscuras de la Formación Cwmhesgen (Miembro Dolgellau) (Rushton, 1982).
Paleontología sistemática
El material estudiado está depositado en la Colección Paleoinvertebrados de la Facultad de Ciencias Naturales e Instituto Miguel Lillo de la Universidad Nacional de Tucumán (PIL); y en el Departamento Paleontología Invertebrados del Museo de Ciencias Naturales de La Plata (MLP), Argentina.
Orden ASAPHIDA Salter, 1864 emend. Fortey y Chatterton, 1988
Superfamilia ASAPHOIDEA Burmeister, 1843
Familia ASAPHIDAE Burmeister, 1843
Género Asaphellus Callaway, 1877
Especie tipo. Asaphus homfrayi Salter, 1866, por designación original.
Asaphellus riojanus Harrington y Leanza, 1957
Figuras 6.1-6.14, 7.1-7.4, 7.15
1957. Asaphellus riojanus sp. nov. Harrington y Leanza (partim):
153, figs. 66.1-66.4 (non fig. 66.5).
Material. Un ejemplar completo, 2 cranidios, 2 librígenas, 5 hipostomas, 2 tórax-pigidios y 26 pigidios, PIL 11397, 11398, 11402, 11404b, 11404d-g, 11405a-k, 11407, 11408a-e, 11409, 11410b, 11418, 11421, 14274; MLP 28215, 28230.
Descripción. Céfalo moderadamente convexo. Glabela lisa, demarcada por surcos axiales angostos, suavemente constricta lateralmente al nivel de los ojos, con su porción anterior enangostada hacia delante y el extremo anterior redondeado a subacuminado. Borde cefálico anterior apenas demarcado, suavemente cóncavo. Sutura facial opistoparia. Áreas palpebrales de las fixígenas muy angostas (tr.), con los lóbulos palpebrales situados por detrás del punto medio del céfalo. Región posterior de las fixígenas ancha (tr.), con un surco del borde posterior apenas visible. Librígena con un borde angosto (tr.), delimitado por un surco del borde muy tenue. Campo genal ancho (tr.) y levemente convexo, con un zócalo ocular muy reducido bordeado por un surco subocular somero. Surco del borde posterior ausente. Espinas genales fuertes, de base ancha y longitud moderada, que se disponen sin quebrar la curvatura del margen de la librígena y alcanzan el nivel del 4°/5° segmento del tórax.
Hipostoma de contorno suboval, aproximadamente tan largo como ancho. Cuerpo medio grande, ovoide, convexo, enangostado hacia atrás, provisto de un par de surcos laterales profundos desconectados medialmente, con un lóbulo posterior pequeño que se prolonga posteriormente en un proceso triangular. Margen anterior fuertemente curvado hacia delante y alas anteriores reducidas. Bordes laterales de buen desarrollo (tr.), cóncavos, con su máxima anchura al nivel de la longitud media del hipostoma, enangostados hacia atrás para formar un borde posterior angosto (sag.), separados del cuerpo medio por un par de surcos anchos y profundos. Margen posterior entero, acuminado.
Tórax con ocho segmentos. Axis de lados subparalelos, proporcionalmente angosto (tr.), ocupando alrededor de 1/5 de la anchura máxima tóracica, bien delimitado por un par de surcos axiales profundos. Pleuras anchas (tr.), con surcos pleurales someros, fulcros distales y extremidades espinosas.
Pigidio de contorno semicircular, suavemente convexo. Axis angosto, largo, bien definido por surcos axiales profundos, con su porción anterior suavemente enangostada hacia atrás mostrando anillos axiales débilmente marcados, y la porción posterior de lados subparalelos, con el extremo distal bulboso. Campos pleurales con seis surcos pleurales muy débilmente marcados, suavemente arqueados hacia atrás. Borde pigidial angosto, levemente cóncavo.
Observaciones. Harrington y Leanza (1957) describieron Asaphellus riojanus sobre la base de un cranidio, una librígena y tres pigidios de río Volcancito. Se ilustra aquí abundante material adicional, que complementa la descripción original de la especie. De esta forma se agrega información sobre el hipostoma y el tórax, y se reconsidera la morfología de la librígena. Aunque los ejemplares muestran un grado de deformación elevado (compárese ejemplos figuras 6.7 y 6.8; 7.1 y 7.2), se aprecian claramente sus caracteres diagnósticos, entre los que se cuentan la glabela enangostada hacia adelante y suavemente constricta lateralmente al nivel de los ojos; lóbulos palpebrales muy angostos (tr.), situados por detrás del punto medio del céfalo; y el pigidio con un axis angosto y largo, seis surcos pleurales muy débilmente marcados y un borde angosto.
Harrington y Leanza (1957: fig. 66.5) ilustraron una librígena aislada de río Volcancito, provista de un borde muy convexo y un surco del borde profundo, que asignaron a A. riojanus. De acuerdo a la evidencia presentada aquí, dicho ejemplar no sería asignable a dicha especie, por cuanto ésta presenta un borde poco elevado y un surco del borde muy tenue.
Asaphellus riojanus es muy similar a Asaphellus communis Robison y Pantoja-Alor (1968: lám. 98, figs. 12-22), del Cámbrico Superior de Méjico, pues ambos poseen un tamaño moderadamente grande, el área frontal de buen desarrollo (sag.), la región anterior de la glabela enangostada hacia delante, ojos dispuestos por detrás del punto medio del cranidio, un pigaxis largo (sag.), surcos pleurales levemente impresos, el doblez pigidial ancho (tr.) y un borde pigidial reducido, levemente cóncavo. No obstante, la especie argentina se distingue porque presenta la región posterior de las fixígenas más amplia (exsag.) (cf. Robison y Pantoja-Alor, 1968), el área frontal con un borde y surco del borde casi imperceptibles, el proceso caudal del cuerpo medio del hipostoma más elongado (sag.), el pigaxis levemente más angosto, los surcos pleurales del pigidio algo más curvados hacia atrás, el doblez ventral pigidial más angosto y la región post-axial desprovista de una arista bien definida. Asaphellus riojanus se diferencia de A. aff. communis, de la Formación Las Vicuñas (Tremadociano inferior) en la Puna occidental de Argentina (Moya et al., 1993: lám. 1, figs. 11-12, 15-16; Waisfeld y Vaccari, 2003: lám. 34, fig. 6), porque el pigidio de este último presenta anillos axiales y surcos pleurales más marcados, un doblez ventral más ancho y una arista postaxial bien definida.
Asaphellus riojanus contrasta con A. emanuelensis (Legg, 1976: lám. 3, figs. 2, 4, 16, 17, 21-23; Laurie y Shergold, 1996: lám. 12, figs. 1-11), del Tremadociano de Australia, porque presenta lóbulos palpebrales más pequeños, un hipostoma con bordes laterales más anchos (tr.) y surcos pleurales pigidiales menos marcados. La especie de La Rioja se distingue más claramente de A. aspinus Robison y Pantoja-Alor, del Cámbrico Superior de Méjico y la Cordillera Oriental argentina (Robison y Pantoja-Alor, 1968: lám. 98, figs. 1-11; cf. Tortello y Esteban, 2003b: figs. 6.I-Q), por tener un tamaño proporcionalmente mayor, la región frontal del céfalo menos cóncava, lóbulos palpebrales más pequeños, librígenas con espina genal, el pigidio con una relación anchura/longitud menor, un pigaxis más angosto (tr.) y surcos pleurales visibles. A su vez, A. riojanus se diferencia de la especie tipo Asaphellus homfrayi Salter, del Tremadociano de Gran Bretaña (véase Fortey y Owens, 1991, figs. 3.l-u, 7.a-g, 8.a-b), porque exhibe ojos más grandes situados por detrás del punto medio del cranidio, un axis torácico más angosto (tr.) y los surcos pleurales del pigidio mejor definidos.

Figura 6. 1-14, Asaphellus riojanus Harrington y Leanza; 1, ejemplar completo, molde de látex / complete specimen, latex mould, PIL 11404d, X3,3; 2, tórax-pigidio / thorax and pygidium, PIL 14274, X4,2; 3, pigidio / pygidium, PIL 11404g, X2,3; 4, pigidio / pygidium, PIL 11418, X4,9; 5, pigidio / pygidium, PIL 11405i, X2; 6, pigidio / pygidium, PIL 11405e, X3; 7, pigidio / pygidium, MLP 28215, X3; 8, pigidio / pygidium, MLP 28230, X2,4; 9, pigidio / pygidium, PIL 11405c, X3,6; 10, pigidio / pygidium, PIL 11405b, X1,6; 11, pigidio / pygidium, PIL 11405d, X4,9; 12, pigidio / pygidium, PIL 11405j, X3,1; 13, pigidio, molde de látex / pygidium, latex mould, PIL 11405k, X2,9; 14, pigidio / pygidium, PIL 11405a, X2.
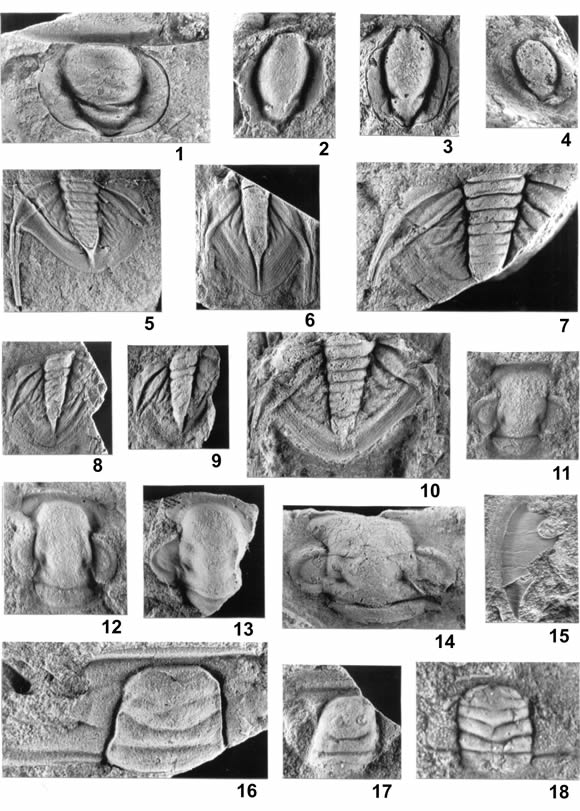
Figura 7. 1-4, Asaphellus riojanus Harrington y Leanza, hipostoma / hypostome; 1, PIL 11408e, X2,7; 2, PIL 11409, X3,3; 3, PIL 11408a, X3; 4, PIL 11408c, X2,2. 5-14, Onychopyge riojana Harrington; 5, pigidio, molde de látex / pygidium, latex mould, PIL 11411g, X3,5; 6, pigidio / pygidium, PIL 11411b, X2,2; 7, pigidio / pygidium, PIL 11411h, X3,7; 8, pigidio / pygidium, PIL 11411f, X2,3; 9, pigidio / pygidium, PIL 11411a, X2,1; 10, pigidio / pygidium, PIL 11411e, X4; 11, cranidio / cranidium, PIL 14275a, X5,6; 12, cranidio / cranidium, PIL 14275b, X3,6; 13, cranidio / cranidium, PIL 14275c, X3,4; 14, cranidio / cranidium, PIL 11411k, X3,6. 15, Asaphellus riojanus, librígena / free cheek, PIL 11404f, X3. 16-17, Parabolina (Neoparabolina) frequens (Barrande), cranidio / cranidium; 16, PIL 11399a, X5,4; 17, MLP 28242, X4,1. 18, Plicatolina scalpta Harrington y Leanza, cranidio / cranidium, PIL 11403b, X11,1.
Asaphellus sp.
Figura 8
Material. Un pigidio (MLP 28273).
Descripción. Pigidio semicircular, suavemente convexo, con una longitud (sag.) que representa un 60% de la máxima anchura. Axis largo, levemente enangostado hacia atrás, delimitado por un par de surcos axiales delgados, ocupando un 85% de la longitud pigidial y un 20% de la máxima anchura del pigidio, con evidencia de cinco anillos axiales en la porción anterior. Campos pleurales con cinco surcos pleurales tenues, suavemente arqueados hacia atrás. Borde angosto (tr., sag.), cóncavo. Doblez ventral de anchura moderada (tr. y sag.), provisto de 18 líneas en terraza subparalelas al margen pigidial, las cuales se curvan hacia delante en la línea sagital.
Observaciones. El pigidio se distingue de Asaphellus communis Robison y Pantoja-Alor (1968: lám. 98, figs. 12-22), del Cámbrico Superior de Méjico, por presentar un axis levemente más corto, un doblez ventral más angosto, y la región post-axial con líneas en terraza curvadas hacia delante, desprovista de quilla sagital.
Harrington (1938) y Harrington y Leanza (1957) reconocieron otro asáfido en la sección de río Volcancito, que asignaron a Asaphellus catamarcensis Kobayashi. No obstante, los dos pigidios ilustrados por estos autores (Harrington y Leanza, 1957: figs. 65.3, 65.5) difieren del material tipo de A. catamarcensis, del Tremadociano inferior de la región de Pascha, Cordillera Oriental (véase Waisfeld y Vaccari, 2003: lám. 22, figs. 3-6), por presentar surcos axiales más conspicuos. Uno de los ejemplares (Harrington y Leanza, 1957: 65.5) es muy similar al pigidio aquí descripto, pero hasta no contar con material adicional, las afinidades específicas de aquéllos permanecen inciertas.
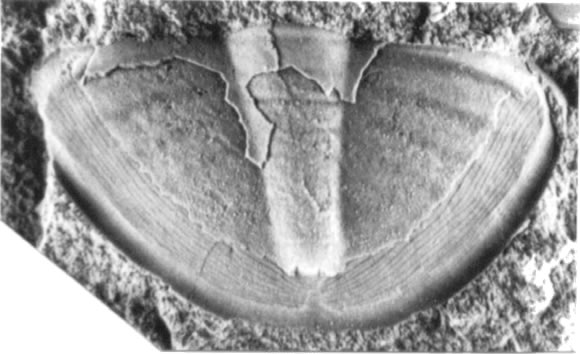
Figura 8. Asaphellus sp., pigidio / pygidium, MLP 28273, X4,6.
Familia CERATOPYGIDAE Linnarsson, 1869
Género Onychopyge Harrington, 1938
Especie tipo. Onychopyge riojana Harrington, 1938, por designación original.
Onychopyge riojana Harrington, 1938
Figuras 7.5-7.14
1938. Onychopyge riojana sp. nov. Harrington: 179, lám. 5, fig. 30.
1957. Onychopyge riojana Harrington. Harrington y Leanza: 187-
188, figs. 95, 96.2a-c. (Véase sinonimia).
?1957. Onychopyge plagiacantha sp. nov. Harrington y Leanza: 189, fig. 96.3.
Material. Cuatro cranidios y 17 pigidios (PIL 11399b, 11411a-k, 11416, 14275a-c, 14277).
Observaciones. Harrington (1938) describió originalmente el género Onychopyge [O. riojana (especie tipo) y O. minor] sobre la base de material escaso (dos pigidios) de río Volcancito. Posteriormente, Harrington y Leanza (1957) describieron algunos ejemplares adicionales de la misma localidad (un cranidio, una librígena y tres pigidios) y consideraron a O. minor como sinónimo junior de O. riojana.
La colección de nuevo material permite sumar algunos elementos a la descripción original de O. riojana. La glabela presenta líneas en terraza con un diseño Bertillon y un par de surcos laterales evidentes. El surco S1 es corto, oblicuo, bifurcado distalmente, situado próximo al surco axial, mientras que S2 es mucho más somero, representado apenas por una depresión corta y oblicua. Los cranidios mejor preservados muestran asimismo leves indicios de un surco S3 muy somero (figura 7.13). El anillo occipital, de imperfecta conservación en el cranidio ilustrado por Harrington y Leanza (1957: fig. 2.a), posee un buen desarrollo en los ejemplares ilustrados aquí; este segmento ocupa 1/7 de la longitud (sag.) del céfalo y es algo más ancho (tr.) que el resto de la glabela, y está surcado por líneas en terraza de trazo semicircular; el surco occipital, delgado y de profundidad moderada, está levemente curvado hacia atrás, al igual que el margen posterior del cranidio. Por su parte, el doblez ventral del pigidio es proporcionalmente ancho (tr., sag.) y exhibe 8-12 líneas en terraza profundas dispuestas en forma subparalela al margen pigidial; las espinas posterolaterales también presentan dichas líneas, en número de 7-8.
Varios ejemplares presentan un alto grado de deformación, razón por la cual se observan variaciones en las proporciones de algunos caracteres morfológicos. El pigidio holotipo de la especie (Harrington y Leanza, 1957: figs. 95, 96.2b) y algunos pigidios ilustrados aquí (ejemplo figura 7.6) están elongados (sag.), por lo que su axis exhibe una longitud proporcionalmente mayor y menor anchura que la presentada por ejemplares no deformados, y el margen posterior del pigidio se muestra exageradamente prolongado hacia atrás. El efecto de la deformación también se muestra en la secuencia de cranidios de las figuras 7.12-7.14, que incluye un ejemplar fuertemente "estirado" en sentido transversal (figura 7.14).
Onychopyge plagiacantha Harrington y Leanza (1957, p. 189, fig. 96.3) fue descripto sobre la base de un pigidio de río Volcancito que, de acuerdo a Harrington y Leanza (1957), se distinguiría de O. riojana por su pigaxis proporcionalmente ancho y la macropleura de disposición oblicua, orientada hacia atrás y afuera. Dado que el ejemplar se encuentra deformado transversalmente, estas diferencias pueden ser de naturaleza preservacional. Onychopyge plagiacantha es probablemente un sinónimo junior de O. riojana.
Aunque Onychopyge es un género bien documentado en el Cámbrico Superior alto (Biozona de Parabolina frequens argentina, parte inferior) de la Cordillera Oriental argentina, en general se lo conoce a través de material muy fragmentario. Harrington y Leanza (1957: figs. 96.1a-d; 96.4) describieron O. argentina y O. longispina en el extremo norte del país, cerca del límite con Bolivia; ambas se diferencian de O. riojana porque presentan un axis pigidial más corto, provisto de 5 anillos axiales (Harrington y Leanza, 1957). Por su parte O. harringtoni de la sierra de Cajas (provincia de Jujuy) (Benedetto, 1977: fig. 10; lám. 2, figs. 14-15) se distingue de O. riojana por su glabela más enangostada hacia adelante, un S2 glabelar más profundo y de disposición transversa, el anillo occipital más corto (sag.) y 6 anillos axiales en el pigidio. Onychopyge sp. de la región de Alfarcito (Jujuy) (Zeballo y Tortello, 2005: fig. 5.L) es muy similar a O. riojana, aunque el carácter fragmentario del primero no permite establecer una comparación adecuada. Onychopyge branisi Suárez Soruco (1975: lám. 1, fig. 7; lám. 2, figs. 7 y 8), del ?Cámbrico Superior de Bolivia, parece distinguirse de O. riojana por sus lóbulos palpebrales más conspicuos.
Onychopyge sp. aff. riojana, del Tremadociano de Nueva Zelandia (Wright et al., 1994: figs. 15.B-G) se diferencia más claramente de O. riojana porque posee la glabela truncada anteriormente y el área preglabelar de mayor desarrollo. De igual modo, muchas otras especies del género [O. sculptura Robison y Pantoja-Alor (1968: lám. 100, figs. 1-7, 10), del Cámbrico Superior de Méjico; O. depressa Quian (1986: lám. 80. figs. 1-6, 11, text.-fig. 116), de la transición Cámbrico-Ordovícico de China; O. austrina Peng (1984: lám. 6, figs. 1-3; lám. 7, figs. 1-2; Peng, 1990: lám. 18, figs. 8-10), del Tremadociano de China; O. parkerae Jell (1985: lám. 23, figs. 1-16; lám. 24, figs. 1-4), del Tremadociano de Australia; O. assula Shergold (1975: lám. 46, figs. 3-4), del Cámbrico Superior de Australia] se distinguen principalmente por su axis pigidial más corto, con un menor número de anillos axiales.
Superfamilia REMOPLEURIDIOIDEA Hawle y Corda, 1847
Familia RICHARDSONELLIDAE Raymond, 1924
Género Pseudokainella Harrington, 1938
Especie tipo. Pseudokainella keideli Harrington, 1938, por designación original.
Observaciones. Pseudokainella fue originalmente descripto por Harrington (1938) y su diagnosis emendada por Harrington y Leanza (1957). De acuerdo a Shergold (1972) y Taylor (1976), varias especies de Norteamérica y Méjico originalmente asignadas a Richardsonella Raymond, 1924 deben ser incluidas en Pseudokainella. El alcance del género fue posteriormente ampliado por Jell (1985), quien detectó una alta variabilidad en varios caracteres morfológicos (ej. dimensiones del campo preglabelar; diseño de las ramas preoculares de la sutura facial; grado de expresión de los surcos glabelares) de material procedente de Australia e incluyó a Elkanaspis Ludvigsen, 1982 y Fatocephalus Duan y An in Guo et al., 1982 en su sinonimia. Posteriormente Peng (1990) aceptó la diagnosis de Jell (1985), aunque consideró a Fatocephalus como un género válido en virtud de la ausencia de campo preglabelar y mejilla interocular.
En la actualidad existen otros criterios taxonómicos que contemplan la posibilidad de restringir Pseudokainella a la especie tipo P. keideli Harrington y revalidar Elkanaspis e incluso Parakainella Kobayashi 1953 (Waisfeld y Vaccari, 2003). Hasta no contar con una revisión integral de estos últimos taxones, que aporte elementos convincentes sobre sus respectivas diagnosis, en el presente trabajo se sigue el concepto amplio de Pseudokainella provisto por Jell (1985), parcialmente emendado por Peng (1990) (cf. Vaccari y Waisfeld, 2000).
Pseudokainella tosellii sp. nov.
Figuras 9.4-9.5, 9.7-9.9
1938. Kainella conica Kobayashi. Harrington: lám. 4, fig. 13.
Material. Un cranidio-tórax (holotipo, MLP 28258), 1 cranidio, 1 tórax y 5 pigidios (PIL 11399a, 11400, 11412, 11413; MLP 28257, 28265).
Procedencia geográfica y estratigráfica. Sección Río Volcancito, La Rioja, Argentina. Miembro Filo Azul (tramo inferior) de la Formación Volcancito; Biozona de Parabolina (Neoparabolina) frequens argentina; Cámbrico Tardío.
Derivación del nombre. En reconocimiento al Dr. Gustavo Toselli, quien coleccionó parte del material estudiado en este trabajo.
Diagnosis. Pseudokainella con un surco glabelar S1 de buen desarrollo y S2 casi imperceptible; pigidio elongado transversalmente, con cuatro anillos axiales (más una pieza terminal), cuatro pares de surcos pleurales profundos y seis pares de espinas marginales / Pseudokainella with a well developed S1 glabellar furrow and a very faint S2 furrow; pygidium transversely elongate, with four axial rings (and a terminal segment), four pairs of deep pleural furrows, and six pairs of marginal spines.
Descripción. Glabela convexa, bien elevada por sobre el nivel de las fixígenas, claramente delimitada por surcos axiales convergentes hacia delante, suavemente constricta al nivel del extremo anterior de los lóbulos palpebrales, con los ángulos anterolaterales redondeados, redondeada a truncada anteriormente, con una longitud que representa aproximadamente un 80% de la longitud cefálica; provista de dos pares de surcos laterales (S1 y S2). S1 muy profundo, recto a suavemente sigmoidal, de disposición oblicua hacia atrás, extendido desde el surco axial hasta las proximidades de la línea sagital; S2 casi imperceptible, más corto y de disposición menos oblicua que S1, en contacto con el surco axial apenas por detrás del extremo anterior del lóbulo palpebral. Surco occipital proporcionalmente ancho (sag., exsag.) y profundo, con su porción media suavemente arqueada y sus tramos laterales extendidos hacia delante. Anillo occipital ancho (sag.) y poco convexo, con un desarrollo transversal que excede la anchura del resto de la glabela, con sus tramos laterales extendidos hacia adelante. Margen anterior del cranidio curvado uniformemente hacia delante. Borde anterior convexo, con una anchura (sag.) similar a la del campo preglabelar, suavemente ensanchado (sag.) en su porción media. Surco del borde representado por una fila de hoyuelos profundos. Campo preglabelar deprimido y de desarrollo moderado, plano en el área sagital y suavemente inflado en las regiones laterales. Lóbulo palpebral localizado muy cerca de la glabela, levemente arqueado hacia fuera, dispuesto desde el surco occipital hasta sobrepasar el surco glabelar S2, ocupando alrededor de la mitad de la longitud (sag.) glabelar. Tramo anterior de la sutura facial divergente y suavemente curvilíneo, formando un ángulo de 45º con la línea sagital en su porción proximal. Área interocular de la fixígena muy reducida (tr.).
Tórax parcialmente preservado. Axis ocupando 1/3 de la anchura máxima del tórax en su porción anterior, suavemente enangostado hacia atrás, provisto de anillos lisos con la excepción del 8°, el cual presenta una espina axial de buen desarrollo orientada hacia atrás. Pleuras con fulcro proximal, surcos pleurales bien marcados y extremos espinosos.
Pigidio de contorno elíptico, elongado transversalmente, con una anchura que supera el doble de su longitud (sin considerar las espinas). Axis convexo, elevado sobre el nivel de los campos pleurales, enangostado hacia atrás, conformado por cuatro anillos separados por surcos transversos muy profundos más una pieza terminal prolongada en una pequeña arista axial que no alcanza el margen posterior del pigidio. Semianillo articulante de buen desarrollo, de contorno semicircular. Campos pleurales de convexidad moderada, con cuatro surcos pleurales profundos de disposición oblicua que contactan el surco del borde pigidial, separados por surcos interpleurales muy tenues. Surco del borde angosto y somero. Borde deprimido, liso, provisto de seis pares de espinas marginales fuertes de base ancha, triangulares, suavemente curvadas, que decrecen levemente de tamaño hacia atrás, espaciadas entre sí regularmente salvo el sexto par, que se sitúa muy próximo al quinto.
Observaciones. Harrington (1938) y Harrington y Leanza (1957) citaron kainélidos de la localidad de río Volcancito. El primero ilustró un cranidio, aunque deformado, que asignó a Kainella conica Kobayashi (Harrington, 1938: lám. 4, fig. 13), mientras que Harrington y Leanza (1957: 131, 250) mencionaron tres cranidios y un pigidio adicionales que asignaron al mismo taxón. Dicho material no se distingue de los ejemplares estudiados aquí, por lo que es incluido en la lista sinonímica de P. tosellii sp. nov.
La nueva especie de río Volcancito se diferencia de "Kainella" conica Kobayashi, del Tremadociano de la zona de Pascha, Salta (véase Harrington y Leanza, 1957: fig. 52.1-4; Tortello y Rao, 2000: figs. 4.C-F; Waisfeld y Vaccari, 2003: lám. 27, figs. 8-12) porque presenta el tramo anterior de la sutura facial menos divergente y un pigidio provisto de un borde mejor definido, con seis pares de espinas marginales. La especie tipo Pseudokainella keideli Kobayashi, del Tremadociano de la región de Alfarcito (Jujuy) (véase Harrington y Leanza, 1957: figs. 51-52.5-10; Waisfeld y Vaccari, 2003: lám. 27, figs. 13-16; Zeballo y Tortello, 2005: figs. 5.G-H), comparte con P. tosellii sp. nov. un pigidio expandido lateralmente que posee cuatro anillos axiales y espinas marginales de buen desarrollo. No obstante, esta última presenta un campo preglabelar de mayor desarrollo y una espina pigidial anterior más corta.
Pseudokainella tosellii sp. nov. se distingue de otras especies de Pseudokainella sensu lato descriptas para el Tremadociano de la Cordillera Oriental argentina por el hecho de presentar seis espinas marginales en el pigidio. De P. lata (Kobayashi) (Harrington y Leanza, 1957: figs. 53-54.1-9) se diferencia además porque exhibe un borde pigidial mejor definido; mientras que también difiere de P. pustulosa Harrington y Leanza (1957: fig. 55.1-8) por poseer un borde cefálico anterior más ancho (sag.), cuatro anillos axiales en el pigido, y por carecer de gránulos sobre la superficie del exoesqueleto. Pseudokainella sp., conocida a través de un cranidio del Cámbrico Superior-Tremadociano de la sierra de La Candelaria (Salta) (Tortello et al., 1996: fig. 2.A-B), apenas se distingue por su surco glabelar S1 menos conspicuo.
La nueva especie argentina contrasta con P. hunanensis Peng (1990: lám. 12, figs. 1-3, 8), del Tremadociano de China, por presentar un surco glabelar S2 imperceptible, el lóbulo glabelar anterior más angosto (tr.), el lóbulo palpebral menos arqueado hacia fuera y en consecuencia el área interocular de la fixígena más reducida. Por su parte, se diferencia de P. cristata (Billings, 1860), del Cámbrico Superior de Canadá (véase Rasetti, 1944: lám. 39, figs. 50-51; Ludvigsen et al., 1989: lám 28, figs. 9-15), por carecer de una constricción glabelar marcada al nivel del extremo anterior del lóbulo palpebral, y de una carena sagital en el sector posterior de la glabela. Pseudokainella tosellii sp. nov. es comparable a Pseudokainella quadrispinosa (Palmer, 1968: lám. 14, figs. 7-8), del Cámbrico Superior de Alaska, por cuanto ambas comparten un pigidio con cuatro anillos axiales, cuatro surcos pleurales, un surco del borde bien definido y espinas marginales fuertes. No obstante, el número de estas últimas es seis en la especie argentina y cuatro en la norteamericana.
Pseudokainella tosellii sp. nov. se diferencia claramente de un grupo de especies de Pseudokainella [P. variagranula (Robison y Pantoja-Alor, 1968: lám. 104, figs. 5-11), del Cámbrico Superior de Méjico; P. unisulcata (Rasetti), del Cámbrico Superior de Canadá (Rasetti, 1944: lám. 39, figs. 54-56; Ludvigsen et al., 1989: lám. 27, figs. 13-18); Pseudokainella sp., del Cámbrico Superior de Alaska (Palmer, 1968: lám. 14, figs. 5-6); P. diggerensis Jell (1985: lám. 24, figs. 5-14; lám 25, figs. 1-13), del Tremadociano de Australia] que exhibe un pigidio con el axis más ancho (tr.) y más corto (sag.), con menos anillos axiales, campos pleurales más reducidos transversalmente, y espinas marginales proporcionalmente más pequeñas, en número menor que seis.
Superfamilia TRINUCLEOIDEA Swinnerton, 1915
Familia HAPALOPLEURIDAE Harrington y Leanza, 1957
Género Rhadinopleura Harrington y Leanza, 1957
Especie tipo. Rhadinopleura eurycephala Harrington y Leanza, 1957, por designación original.
Rhadinopleura eurycephala Harrington y Leanza, 1957
Figura 9.15
1957. Rhadinopleura eurycephala sp. nov. Harrington y Leanza: 207, figs. 112.3a-b, 113.
Material. Dos cranidios (PIL 11410a).
Observaciones. Rhadinopleura posee un singular valor paleobiogeográfico por cuanto representa un hapalopléurido de particular distribución geográfica, restringida a Sudamérica y a la región de Hunan (China). El genotipo R. eurycephala Harrington y Leanza fue descripto sobre la base de un cranidio y un cranidio-tórax de río Volcancito (Harrington y Leanza, 1957: figs. 112.3a-b). El material estudiado aquí presenta una glabela piriforme muy convexa provista de tres pares de surcos glabelares cortos y profundos; área frontal y fixígenas muy anchas, sin borde anterior; lóbulos palpebrales pequeños situados muy lejos de la glabela; y aristas oculares largas y rectas dispuestas en forma perpendicular al eje axial, por lo que se lo asigna a dicha especie.
Babcock y Smith (2003: lám. 1, figs. 4, 5, 7, 8-10) describieron material del Temadociano (Biozona de Kainella meridionalis) de diferentes localidades del sur de Bolivia (Tambo Guanacuno, Cuesta de Erquis, Quebrada Palqui) que asignaron a R. eurycephala. No obstante, el mismo exhibe surcos glabelares largos, angostos y someros, una glabela de contorno variable y un campo preglabelar proporcionalmente reducido (sag.), por lo que podría pertenecer a otra especie.
Rhadinopleura sp.
Figura 9.14
Material. Un cranidio (MLP 28235).
Descripción. Cranidio subtrapezoidal. Glabela piriforme, muy convexa, bien elevada por sobre el nivel de las genas, más larga que ancha, bien definida por surcos axiales delgados y profundos, ocupando alrededor del 80% de la longitud cefálica, con su porción anterior enangostada hacia delante terminando en punta, provista de tres pares de surcos laterales; éstos son muy cortos aunque profundos, dispuestos en forma oblicua hacia atrás. Surco occipital angosto, levemente curvado hacia atrás. Área frontal amplia (sag.), con una quilla sagital extendida desde el extremo anterior de la glabela hasta el margen anterior del cranidio. Borde anterior no diferenciado. Fixígena ancha (tr.), poco convexa. Arista ocular muy larga (tr.), angosta (exsag.), recta y transversa, en contacto con el margen anterolateral de la glabela inmediatamente por delante de S3. Lóbulo palpebral pequeño, dispuesto lejos de la glabela. Ramas anteriores de la sutura facial suavemente convergentes hacia delante. La región posterior del cranidio se encuentra imperfectamente preservada.
Observaciones. El ejemplar podría ser considerado un representante de R. eurycephala extremadamente deformado en sentido anteroposterior. A su vez, es muy similar al cranidio asignado a Rhadinopleura sp. por Peng (1991: lám. 3, fig. 1) del Tremadociano inferior de Hunan oeste, China (el primero apenas se distingue porque posee surcos glabelares laterales menos evidentes). Hasta no contar con material adicional, el ejemplar es referido en nomenclatura abierta.
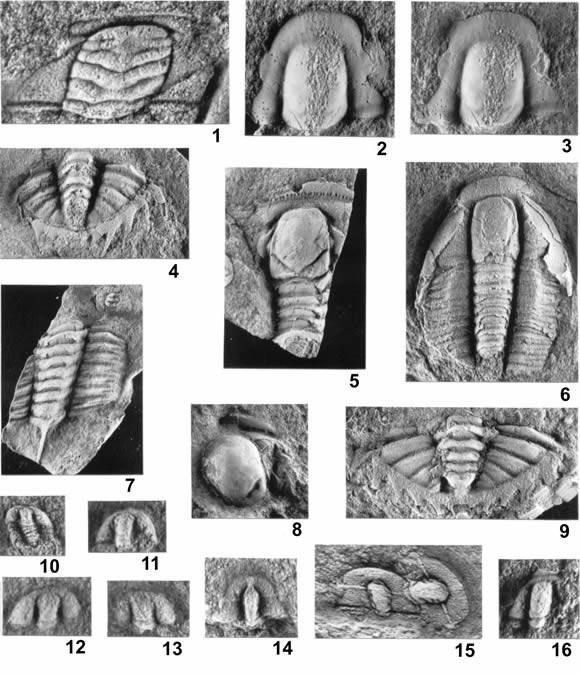
Figura 9. 1, Plicatolina scalpta Harrington y Leanza, cranidio / cranidium, PIL 14273a, X11. 2-3, Angelina cf. hyeronimi (Kayser); 2, cranidio, molde de látex / cranidium, latex mould, PIL 14276, X6; 3, cranidio (contramolde de ejemplar ilustrado en 2) / cranidium (contramould of specimen illustrated in 2), PIL 14272, X6. 4-5, Pseudokainella tosellii sp. nov.; 4, pigidio / pygidium, MLP 28257, X2,5; 5, cranidiotórax fragmentario (holotipo) / cranidium and fragmentary thorax (holotype), MLP 28258, X1,5. 6, Angelina cf. hyeronimi, céfalo-tórax / cephalon and thorax, MLP 28259, X3,2. 7-9, Pseudokainella tosellii sp. nov.; 7, tórax / thorax, PIL 11400, X2,3; 8, cranidio fragmentario / fragmentary cranidium, MLP 28265, X2,1; 9, pigidio / pygidium, PIL 11412, X3,3. 10-13, Conophrys sp.; 10, cranidio-tórax / cranidium and thorax, PIL 11400, X8; 11, cranidio / cranidium, MLP 28216, X10; 12, cranidio / cranidium, MLP 28187, X9,3; 13, cranidio / cranidium, MLP 28206, X9. 14, Rhadinopleura sp., cranidio / cranidium, MLP 28235, X7. 15, Rhadinopleura eurycephala Harrington y Leanza, cranidios / cranidia, PIL 11410a, X5,2. 16, (?) Bodenbenderia longifrons Harrington y Leanza, cranidio / cranidium, MLP 28237, X9.
Orden PTYCHOPARIIDA Swinnerton, 1915
Suborden OLENINA Burmeister, 1843
Familia OLENIDAE Burmeister, 1843
Subfamilia OLENINAE Burmeister, 1843
Género Parabolina Salter, 1849
Subgénero P. (Neoparabolina) Nikolaisen y Henningsmoen, 1985
Especie tipo. Parabolina frequens (Barrande, 1868), por designación original.
Parabolina (Neoparabolina) frequens (Barrande, 1868)
Figuras 7.16-7.17
Material. Tres cranidios (PIL 11399a, 11411f; MLP 28242).
Observaciones. Aunque el taxón fue citado por Harrington y Leanza (1957) para río Volcancito, se ilustran por primera vez ejemplares de Parabolina (Neoparabolina) frequens de esta localidad. El material consiste de tres cranidios, uno de ellos deformado transversalmente (figura 7.16). Los mismos se caracterizan por su glabela subtrapezoidal suavemente enangostada hacia delante, con el extremo anterior subtruncado, provista de un surco S1 transglabelar y un S2 con tendencia a estar conectado medialmente. Parabolina (Neoparabolina) frequens fue descripta en el Cámbrico Superior-Tremadociano inferior de Europa Central, Gran Bretaña, Noruega, Newfoundland, Méjico, Venezuela y la Cordillera Oriental de Argentina y Bolivia (véase ej. Harrington y Leanza, 1957; Sdzuy, 1955; Robison y Pantoja-Alor, 1968; Rushton, 1982; Nikolaisen y Henningsmoen, 1985; Zylinska, 2001; Zeballo y Tortello, 2005). La subespecie P. frequens argentina (Kayser) está ampliamente representada en numerosas localidades del Cámbrico Tardío de Sudamérica, caracterizando a la biozona que lleva su nombre (Harrington y Leanza, 1957). Este taxón se distingue de P. frequens frequens por su borde pigidial más reducido (Pribyl y Vanek, 1980). Dado que no se cuenta aún con pigidios de río Volcancito, el material estudiado aquí es determinado a nivel específico. De todos modos, se señala que éste se asemeja particularmente a los cranidios descriptos por Sdzuy (1955: lám. 3, figs. 58-61) en el Tremadociano inferior de Alemania.
Género Angelina Salter, 1859
Especie tipo. Angelina sedgwickii Salter, 1859 (Vodges, 1890).
Angelina cf. hyeronimi (Kayser, 1876)
Figuras 9.2-9.3, 9.6, ?10.9
1957. Angelina punctolineata Kobayashi (partim). Harrington y Leanza: figs. 36.2a-b.
Material. Un céfalo-tórax (incompleto) y 8 cranidios (PIL 11401h, 14272a-c, 14276, 14277; MLP 28233, 28234, 28259).
Descripción. Cranidio subtrapezoidal. Glabela proporcionalmente grande, de contorno subrectangular, 1,33 veces tan larga como ancha, bien definida por surcos axiales subparalelos a suavemente convergentes hacia delante, subtruncada anteriormente, ocupando 3/4 de la longitud del céfalo, lisa o con débiles indicios de un par de surcos laterales; anillo occipital liso, delimitado por un surco occipital somero, el cual posee su porción media transversa y los tramos laterales bifurcados o sólo extendidos hacia delante. Área frontal convexa, curvada hacia abajo, con un surco del borde anterior representado por una hilera de hoyuelos recta o levemente curvada hacia delante. Lóbulos palpebrales de tamaño mediano, dispuestos moderadamente cerca de la glabela y apenas por delante del punto medio del cranidio. Ramas anteriores de la sutura facial subparalelas a levemente convergentes. Área posterior de la fixígena triangular, con un borde posterior más angosto (exsag.) que el anillo occipital. Espina genal larga, que prolonga la curvatura del margen lateral de la librígena.
Observaciones. Angelina punctolineata Kobayashi fue descripta originalmente sobre la base de material coleccionado en el sur de Bolivia (véase Kobayashi, 1937: lám. 6, fig. 22; Harrington y Leanza, 1957: fig. 36.2c-d; Pribyl y Vanek, 1980: lám. 8, figs. 1-3). Asimismo Harrington y Leanza (1957: figs. 36.2a-b, e) asignaron a esa especie tres cranidios de río Volcancito, los que sin embargo no exhiben el surco del borde cefálico (hilera de hoyuelos) curvado hacia atrás característico del taxón. El material que se ilustra aquí tampoco muestra ese carácter, y aparenta tener un campo preglabelar algo más reducido que A. punctolineata.
Los ejemplares descriptos son muy similares a A. hyeronimi, del Cámbrico Superior de la Cordillera Oriental de Argentina y Bolivia, Méjico y Canadá (ej. Kayser, 1876: lám. 1, fig. 5; Harrington y Leanza, 1957: figs. 35.1-11; Robison y Pantoja-Alor, 1968: lám. 101, figs. 1-7; Tortello y Esteban, 2003b: figs. 6.A-D, E?, F?, G; Pratt, 1988: figs. 8.I-K) pero se diferencian levemente por tener una glabela de contorno más rectangular, definida por surcos axiales subrectos.
El ejemplar ilustrado en la figura 9.6 muestra las librígenas levemente desplazadas hacia atrás respecto del cranidio. Este tipo de configuración ("Harrington's configuration", véase Henningsmoen, 1975) es frecuente en trilobites con librígenas fusionadas medialmente y puede ser interpretada como el resultado del proceso de ecdisis. Durante dicho proceso, la sutura facial permitiría la apertura de la vieja cutícula, que el animal abandonaría en forma de exuvia desplazándose y emergiendo hacia delante. Ejemplos similares en la familia Olenidae se han documentado en el Paleozoico inferior de la Cordillera Oriental argentina, en Parabolina (Neoparabolina) frequens argentina (Kayser) (Tortello y Clarkson, 2006) y Parabolinella (Harrington y Leanza, 1957: figs. 38.8 y 39.3h; Henningsmoen, 1975: fig. 5).
Género Parabolinella BrØgger, 1882
Especie tipo. Parabolinella limitis BrØgger, 1882 (Bassler, 1915).
Parabolinella sp. 1
Figuras 10.1-10.2, 10.4, 10.7
Material. Seis cranidios (PIL 11399a, 11401b, 11401d, 11401f, 11401i; MLP 28279).
Descripción. Exoesqueleto delgado, muy poco convexo. Cranidio de contorno subtrapezoidal. Glabela subcuadrada, aproximadamente tan larga como ancha, poco elevada por sobre el nivel de las genas, apenas enangostada hacia delante, levemente constricta al nivel de los surcos laterales, bien definida por surcos axiales delgados y profundos, ocupando un 70-75% de la longitud (sag.) cefálica, con tres pares de surcos laterales desconectados medialmente. Surco preglabelar levemente curvado hacia delante, con una suave incisión en su porción sagital. Surco glabelar S1 profundo, sigmoidal, oblicuo hacia atrás y adentro, bifurcado distalmente; S2 profundo, recto a suavemente curvado hacia atrás, de disposición algo menos oblicua que S1, no bifurcado; S3 más somero y más corto que S1 y S2, dispuesto en forma perpendicular al eje axial. Anillo occipital provisto de un tubérculo axial, bien delimitado por un surco occipital transverso y profundo. Campo preglabelar de convexidad moderada y anchura (sag.) algo variable (compárese figuras 10.1, 10.2, 10.4), con aristas y surcos anastomosados. Borde anterior muy angosto (sag.), convexo, definido por un surco del borde profundo y delgado representado por una hilera de hoyuelos. Aristas oculares bien definidas, que se ensanchan progresivamente hacia fuera y se prolongan en lóbulos palpebrales de buen desarrollo, los cuales se disponen lejos de la glabela y apenas por delante del punto medio del cranidio. Ramas anteriores de la sutura facial levemente divergentes. Área posterior de la fixígena triangular, amplia (tr.), con un borde posterior angosto (exsag.) delimitado por un surco del borde delgado y profundo.
Observaciones. Harrington y Leanza (1957: 105) citaron la presencia de numerosos cranidios de Parabolinella en río Volcancito, que asignaron a P. argentinensis Kobayashi. Parabolinella sp. 1 es muy similar a P. argentinensis Kobayashi sensu Waisfeld y Vaccari (2003), del Tremadociano inferior de la Cordillera Oriental de Argentina (Kobayashi, 1936: lám. 15, figs. 1-5; Waisfeld y Vaccari, 2003: lám. 32, figs. 9-13), pero se distingue por su borde cefálico anterior más recto y levemente más angosto (sag.). Estos caracteres pueden carecer de valor específico, pero hasta no contar con ejemplares adicionales, que incluyan exoesqueletos completos, el material es referido en nomenclatura abierta.
Parabolinella sp. 1 también es comparable con P. hunanensis Peng, de la transición CámbricoOrdovícico de China (Peng, 1991: lám. 2, figs. 4-5), de la que apenas contrasta por sus lóbulos palpebrales más grandes y los tramos anteriores de la sutura facial levemente divergentes. A su vez, Parabolinella sp. 1 se diferencia de P. coelatifrons Harrington y Leanza, del Cámbrico Superior de la Cordillera Oriental (Harrington y Leanza, 1957: 39.3.a-h; Tortello y Esteban, 2003b: figs. 5.A-B), por presentar estrías más evidentes sobre el campo preglabelar, los surcos S2 y S3 más profundos, la glabela constricta al nivel de los surcos laterales, un surco preglabelar con su porción central curvada hacia atrás, y aristas oculares más conspicuas.
Parabolinella sp. 2
Figuras 10.3, 10.5-10.6, 10.8
Material. Seis cranidios (PIL 11399b, 11401g, 11405f; MLP 28182, 28197, 28199).
Descripción. Cranidio de contorno subtrapezoidal. Glabela rectangular, más larga que ancha, moderadamente convexa, de lados subparalelos, truncada anteriormente, bien definida por surcos axiales delgados y profundos, ocupando alrededor de un 65-70% de la longitud (sag.) cefálica, con dos pares de surcos laterales (S1 y S2) profundos que no están conectados medialmente. Éstos son rectos a levemente arqueados hacia delante y se disponen en forma oblicua hacia atrás y adentro; S1 algo más oblicuo que S2. En los ejemplares mejor conservados se observa un tercer par de surcos (S3) apenas esbozado, subparalelo a S2. Surco occipital angosto, recto en su porción media y levemente curvado hacia delante cerca de sus extremos. Anillo occipital con un tubérculo occipital conspicuo. Campo preglabelar ancho (sag.) y moderadamente convexo, ocupando alrededor de 1/4 de la longitud cefálica. Borde anterior convexo, angosto (sag.), bien definido por una hilera de hoyuelos que en conjunto conforman el surco del borde anterior. Ramas anteriores de la sutura facial levemente divergentes. Lóbulos palpebrales situados moderadamente cerca de la glabela y apenas por delante del punto medio del cranidio. Área posterior de la fixígena triangular, amplia (tr.), con un borde posterior más angosto (exsag.) que el anillo occipital, delimitado por un surco del borde profundo.
Observaciones. Parabolinella sp. 2 es comparable a P. triarthroides Harrington, del Tremadociano superior de Jujuy (Cordillera Oriental) (Harrington, 1938: lám. 7, figs. 10-11; Harrington y Leanza, 1957: fig. 39.1.a-b; Waisfeld y Vaccari, 2003: lám. 32, figs. 14-18), por cuanto exhibe una glabela subcuadrada y surcos glabelares con un diseño similar. No obstante, el primero presenta la glabela algo más angosta (tr.) y un campo preglabelar más amplio (sag.). Parabolinella sp. 2 se diferencia más claramente de Parabolinella sp. 1, con la cual aparece asociada en el Miembro Filo Azul, porque posee la glabela de contorno rectangular, el surco S3 vestigial o ausente, un campo preglabelar liso y aristas oculares de menor desarrollo.
Subfamilia PLICATOLININAE Robison y Pantoja-Alor, 1968
Género Plicatolina Shaw, 1951
Especie tipo. Plicatolina kindlei Shaw, 1951, por designación original.

Figura 10. 1-2, 4, 7, Parabolinella sp. 1, cranidio / cranidium; 1, PIL 11401i, X7; 2, PIL 11399a, X3,7; 4, PIL 11401b, X4; 7, PIL 11401f, X3. 3, 5-6, 8, Parabolinella sp. 2, cranidio / cranidium; 3, PIL 11405f, X5,5; 5, PIL 11401g, X5; 6, PIL 11399b, X 5,4; 8, MLP 28182, X5. 9, (?) Angelina cf. hyeronimi (Kayser), cranidio / cranidium, MLP 28233, X6,9.
Plicatolina scalpta Harrington y Leanza, 1957
Figuras 7.18, 9.1
1957. Plicatolina scalpta sp. nov. Harrington y Leanza: 111, 113, figs. 40, 41.1-9 (véase sinonimia).
2003b. Plicatolina scalpta Harrington y Leanza. Tortello y Esteban: 341, figs. 5.D-E.
Material. Cuatro cranidios y 1 tórax (PIL 11403a-b, 14273a-c).
Observaciones. Harrington y Leanza (1957) describieron Plicatolina scalpta sobre la base de varios ejemplares de la Cordillera Oriental, sumado a un cranidio aislado de río Volcancito (Harrington y Leanza, 1957: fig. 41.5). La especie exhibe cierta variabilidad en el grado de desarrollo del campo preglabelar. El material de río Volcancito incluye ejemplares con el campo preglabelar relativamente reducido, y en tal sentido se asemeja especialmente a los descriptos por Harrington y Leanza (1943: lám. 2, fig. 10) y Tortello y Esteban (2003b: figs. 5.A-D) para el Cámbrico Superior de Salitre (sur de Bolivia) y la sierra de Cajas (Jujuy, Argentina) respectivamente.
Suborden PTYCHOPARIINA Swinnerton, 1915
Familia SHUMARDIIDAE Lake, 1907
Género Conophrys Callaway, 1877
Especie tipo. Conophrys salopiensis Callaway, 1877, por designación original.
Conophrys sp.
Figuras 9.10-9.13
Material. Seis cranidios y 1 pigidio imperfectamente preservado (PIL 11400, 11410a; MLP 28187, 28206, 28216, 28227).
Descripción. Cranidio de contorno semicircular, moderadamente convexo. Glabela convexa, elevada sobre el nivel de las genas, de lados subparalelos en su porción posterior, bien definida por surcos axiales profundos; lóbulos anterolaterales muy pequeños; extremo glabelar anterior acuminado, delimitado por un surco preglabelar angosto y somero. S1 y S2 extremadamente cortos, apenas visibles en contacto con los surcos axiales. Surco occipital angosto y muy somero, curvado hacia atrás. Anillo occipital liso, de ancho uniforme (sag., exsag.). Fixígena subtriangular, convexa, extendida hacia adelante en un campo preglabelar muy corto (sag.). Surco del borde posterior profundo. Borde posterior imperfectamente preservado.
Observaciones. Harrington (1938: lám. 9, fig. 14) ilustró un shumárdido de río Volcancito que asignó a Shumardia erquensis Kobayashi, 1937. Posteriormente, Harrington y Leanza (1957) citaron la presencia de diez céfalos y dos pigidios adicionales. De acuerdo a la ilustración provista por Harrington (1938), dichos ejemplares serían coespecíficos de los que se ilustran aquí. Aunque el material no está perfectamente conservado, es posible asignarlo a un grupo de especies de Conophrys que posee lóbulos glabelares anterolaterales pequeños [ej. C. alata (Robison y Pantoja-Alor, 1968); C. fabiani Waisfeld et al. 2001; C. erquensis Kobyashi, 1937; C. rushtoni Waisfeld et al., 2001; C. changshanensis (Lu in Lu et al., 1976); C. sulcatus (Malanca, 1996)].
Conophrys sp. se diferencia de C. fabiani, del Tremadociano inferior de la Puna argentina (Moya et al., 1993: lám. 1, fig. 2; Waisfeld et al., 2001: figs. 13.1-12), por presentar el surco occipital levemente curvado hacia atrás y S1 y S2 menos desarrollados. Por su parte, contrasta levemente con C. alata (Robison y Pantoja-Alor, 1968: lám. 99, figs. 13-18), del Cámbrico Superior alto de Méjico, por su surco occipital arqueado hacia atrás.
Conophrys sp. se distingue de C. erquensis (Kobayashi, 1937: lám. 6, figs. 1-3), del Tremadociano inferior de Bolivia, porque el primero exhibe una glabela más larga, menos redondeada frontalmente. A su vez, se diferencia de C. rushtoni Waisfeld et al. (2001), del Tremadociano inferior de Gales (véase Rushton, 1982: lám. 3, figs. 13-17, 19, 21), por presentar la glabela más larga (sag.), surcos glabelares laterales menos conspicuos y el anillo occipital desprovisto de tubérculo axial. Conophrys changshanensis (Lu) (véase Peng, 1990: lám. 5, figs. 7-11), del Tremadociano inferior del sur de China, se diferencia de la forma argentina porque ésta posee una glabela más elongada (sag.).
Incertae Sedis
El material estudiado se completa con dos cranidios muy pequeños, de preservación regular, asignables a Bodenbenderia longifrons Harrington y Leanza (1957: 226, fig. 124.2) (véase figura 9.16).
Conclusiones
El examen de nuevas colecciones de trilobites del Miembro Filo Azul de la Formación Volcancito permite redescribir e ilustrar varios taxones originalmente propuestos con material escaso (Onychopyge riojana Harrington, Asaphellus riojanus Harrington y Leanza, Rhadinopleura eurycephala Harrington y Leanza, Plicatolina scalpta Harrington y Leanza) y reconsiderar la identidad de algunas formas [Angelina cf. hyeronimi (Kayser), Parabolinella sp. 1, Parabolinella sp. 2, Pseudokainella tosellii sp. nov., Conophrys sp.]. La presencia de Parabolina frequens (Barrande) constituye un nuevo criterio que, sumado a la información bioestratigráfica provista por los trilobites agnóstidos, permite efectuar correlaciones con otras secciones de edad cámbrica tardía alta.
El tramo inferior del miembro Filo Azul contiene niveles con alta concentración de materia orgánica y ciertos rasgos que sugieren el crecimiento de tapetes microbiales. La abundancia de nutrientes habría favorecido el desarrollo de una comunidad de trilobites compleja, que incluye diferentes morfotipos y hábitos alimentarios. Varios taxones estaban especialmente adaptados a vivir en ambientes con escaso oxígeno, algunos en probable simbiosis con bacterias del azufre. La diversidad de trilobites muestra mayores similitudes con las descriptas en la secuencia mixta (carbonático-siliciclástica) de la Formación Tiñu (Oaxaca, Méjico) y el Cámbrico Superior alto de Gales, Cordillera Oriental y Puna.
Agradecimientos
Nuestro agradecimiento a D. Ruiz Holgado y E. Gómez Hasselrot por la confección de los dibujos; a J. Rodríguez por el revelado y copiado de las fotografías; a F.G. Aceñolaza por el asesoramiento brindado; a los revisores E.N.K. Clarkson, B.G. Waisfeld y un árbitro anónimo por sus valiosos comentarios y sugerencias; y al Comité Editor de Ameghiniana por su eficiente labor editorial. Este trabajo fue financiado por el Instituto Superior de Correlación Geológica, el Consejo Nacional de Investigaciones Científicas y Técnicas y la Universidad Nacional de Tucumán.
Bibliografía
1. Albanesi, G.L., Esteban, S.B. y Barnes, C.R. 1999. Conodontes del intervalo del límite Cámbrico-Ordovícico en la Formación Volcancito, Sistema de Famatina, Argentina. Temas Geológico-Mineros, Instituto Tecnológico Geominero de España 26: 521-526. [ Links ]
2. Albanesi, G.L., Esteban, S.B., Ortega, G., Hünicken, M.A. y Barnes, C.R. 2005. Bioestratigrafía y ambientes sedimentarios de las Formaciones Volcancito y Bordo Atravesado (Cámbrico superior-Ordovícico inferior), Sistema de Famatina, provincia de La Rioja. Asociación Geológica Argentina, Serie D: Publicación Especial 8: 41-64. [ Links ]
3. Armella, C. y Cabaleri, N.G. 1984. Microfacies y paleoecología en la transgresión tremadociana de la Formación Tiñu, Oaxaca, Mexico. 3° Congreso Latinoamericano de Paleontología (Méjico), Memoria: 42-53. [ Links ]
4. Astini, R.A. 2001. Las algas calcáreas Nuia y Girvanella a través de la transición cambro-ordovícica (Formación Volcancito) en el Famatina: significado paleoambiental y paleogeográfico. Ameghiniana 38: 243-255. [ Links ]
5. Babcock, L.E. y Smith, L. 2003. Ordovician hapalopleurine trilobites (Trinucleidea: Orometopidae) from Tarija Department, Bolivia. Revista Técnica de Yacimientos Petrolíferos Fiscales Bolivianos 21: 235-240. [ Links ]
6. Baldis, B.A.J. y González, S.B. 1986. Australoharpes precordilleranus nov. sp. (Trilobita, Ptychopariida, Harpidae) del Ordovícico Inferior de la sierra de Villicum (Provincia de San Juan, Argentina). 4° Congreso Argentino de Paleontología y Bioestratigrafía (Mendoza), Actas 1: 73-79. [ Links ]
7. Barrande, J. 1868. Silurische Fauna aus der Umgebung von Hof in Bayern. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie 1868: 641-696, lám. 6-7. [ Links ]
8. Bassler, R.S. 1915. Bibliographic index of American Ordovician and Silurian fossils. Bulletin of the United States Natural Museum 92: 1-152. [ Links ]
9. Bauld, J. 1981. Occurrence of benthic microbial mats in saline lakes. Hydrobiology 81: 87-111. [ Links ]
10. Benedetto, J.L. 1977. Una nueva fauna de trilobites tremadocianos de la Provincia de Jujuy (sierra de Cajas), Argentina. Ameghiniana14: 186-214. [ Links ]
11. Billings, E. 1860. On some new species of fossils from the limestone near Point Levis opposite Quebec. Canadian Naturalist 5: 301-324. [ Links ]
12. Bodenbender, G. 1916. El Nevado de Famatina. Boletín de la Academia Nacional de Ciencias (Córdoba) 21: 100-182. [ Links ]
13. Brenchley, P.J., Romano, M. y Gutiérrez-Marco, J.C. 1986. Proximal and distal hummocky cross-stratified facies on a wide Ordovician shelf in Iberia. En: R.J. Knight y J.R. Mc Lean (eds.), Shelf sands and sandstones, Canadian Society of Petroleum Geologist Memoir 11: 241-255. [ Links ]
14. BrØgger, W.C. 1882. Die Silurischen Etagen 2 und 3 im Kristianiagebiet und auf Eker. Universitats Programm 32, semestre 1882, Kristiania, 376 pp. [ Links ]
15. Bruton, D.L. y Nakrem, H.A. 2005. Enrolment in a Middle Ordovician agnostoid trilobite. Acta Palaeontologica Polonica 50: 441-448. [ Links ]
16. Burmeister, H. 1843. Die Organisation der Trilobiten, aus ihren lebenden Verwandten entwickelt; nebst einer systematischen übersicht aller zeither beschriebenen Arten. Berlin, 147 pp. [ Links ]
17. Callaway, C. 1877. On a new area of Upper Cambrian rocks in south Shropshire, with a description of a new fauna. Quarterly Journal of the Geological Society of London 33: 652-672. [ Links ]
18. Esteban, S.B. 1999. [Estratigrafía, geología sedimentaria y paleontología del Ordovícico basal del Sistema de Famatina. Tesis Doctoral, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán, Argentina, 189 pp. Inédito. [ Links ]].
19. Esteban, S.B. 2003a. The Early Ordovician in the Sistema de Famatina: stratigraphic and geotectonic framework. Serie Correlación Geológica 16: 27-40. [ Links ]
20. Esteban, S.B. 2003b. Biogenic and physical sedimentary structures in latest Cambrian-earliest Ordovician mudrock facies (Famatina Range, Northwestern Argentina). Geologica Acta 1: 85-94. [ Links ]
21. Fortey, R.A. 1985. Pelagic trilobites as an example of deducing the life habits of extinct arthropods. Transactions of the Royal Society of Edinburgh 76: 219-230. [ Links ]
22. Fortey, R.A. 2000. Olenid trilobites: The oldest known chemoautotrophic symbionts? PNAS (Proceedings of the National Academy of Sciences of the United States of America) 97: 6574-6578. [ Links ]
23. Fortey, R.A. y Owens, R.M. 1987. The Arenig Series in South Wales: stratigraphy and palaeontology. Bulletin of the British Museum (Natural History), Geology 41: 69-307. [ Links ]
24. Fortey, R.A. y Owens, R.M. 1991. A trilobite fauna from the highest Shineton Shales in Shropshire, and the correlation of the latest Tremadoc. Geological Magazine 128: 437-464. [ Links ]
25. Fortey, R.A. y Owens, R.M. 1999. Feeding habits in trilobites. Palaeontology 42: 429-465. [ Links ]
26. Fortey, R.A. y Rushton, A.W.A. 1980. Acanthopleurella Groom 1902: origin and life habits of a miniature trilobite. Bulletin of the British Museum (Natural History), Geology 33: 79-89. [ Links ]
27. Guo, H., Duan, J. y An, S. 1982. Cambrian-Ordovician boundary in the north China platform with descriptions of trilobites. Papers for the Fourth International Symposium on the Ordovician System, Changchun College of Geology, Changchun: 1-31. [ Links ]
28. Gerdes, G., Krumbein, W. y Reineck, H. 1991. Biolaminations - Ecological versus depositional dynamics. En: E. Einsele, W. Ricken y A. Seilacher (eds.), Cycles and events in stratigraphy, Springer-Verlag, Berlin, pp. 592-607. [ Links ]
29. Gerdes, G., Krumbein, W. y Noffke, N. 2000. Evaporite microbial sediments. En: R.E. Riding y S.M. Awramik (eds.), Microbial sediments, Springer-Verlag, Berlin, pp. 196-208. [ Links ]
30. Hagardon, J. y Bottjer, D. 1997. Wrinkle structures: Microbially mediated sedimentary structures common in subtidal siliciclastic settings at the Proterozoic-Phanerozoic transition. Geology 25: 1047-1050. [ Links ]
31. Hagadorn, J.W. y Bottjer, D.J. 1999. Restriction of a Late Neoproterozoic biotope: suspect-microbial structures and trace fossils at the Vendian-Cambrian transition. Palaios 14: 73-85. [ Links ]
32. Harrington, H.J. 1938. Sobre las faunas del Ordoviciano Inferior del Norte argentino. Revista del Museo de La Plata (Nueva Serie), Sección Paleontología 1: 109-289. [ Links ]
33. Harrington, H.J. y Leanza, A.F. 1943. La fáunula del Tremadociano inferior de Salitre (Bolivia). Revista del Museo de La Plata (Nueva Serie), Sección Paleontología 2: 343-356. [ Links ]
34. Harrington, H.J. y Leanza, A.F. 1957. Ordovician trilobites of Argentina. Department of Geology, University of Kansas Special Publication 1: 1-276. [ Links ]
35. Henningsmoen, G. 1957. The trilobite family Olenidae, with description of Norwegian material and remarks on the Olenid and Tremadocian Series. Skrifter Utgitt av det Norske Videnskaps-Akademi i Oslo I Matematisk-Naturvidenskapelig Klasse 1957(1): 303 pp. [ Links ]
36. Henningsmoen, G. 1975. Moulting in trilobites. Fossils and Strata 4: 179-200. [ Links ]
37. Jell, P.A. 1985. Tremadoc trilobites of the Digger Island Formation, Waratah Bay, Victoria. Memoirs of the Museum Victoria 46: 53-88. [ Links ]
38. Kauffman, E.G. 1981. Ecological reappraisal of the German Posidonienschiefer (Toarcian) and the stagnant basin model. En: J. Gray, A.J. Boucot y W.B. Berry (eds.), Communities of the past, Hutchinson Ross, Stroudsburg, pp. 224-259. [ Links ]
39. Kayser, E. 1876. Uber primordiale und untersilurische Fossilien aus der Argentinischen Republik. Palaeontographica, Supplementun 3, Lieferung 2, Theil III. En: Actas de la Academia Nacional de Ciencias 8: 297-332, Tablas 1-5 (traducción al castellano). [ Links ]
40. Kobayashi, T. 1936. On the Parabolinella fauna from Province Jujuy, Argentina. Japanese Journal of Geology and Geography 13: 85-102. [ Links ]
41. Kobayashi, T. 1937. The Cambro-Ordovician shelly faunas of South America. Journal of the Faculty of Science, Imperial University of Tokyo, Section 2, 4: 369-522. [ Links ]
42. Kobayashi, T. 1953. On the Kainellidae. Japanese Journal of Geology and Geography 23: 38-61. [ Links ]
43. Lake, P. 1907. A monograph of the British Cambrian trilobites. Part 2. Monograph of the Palaeontographical Society, pp. 29-48, láms. 3-4. [ Links ]
44. Laurie, J.R. y Shergold, J.H. 1996. Early Ordovician trilobite taxonomy and biostratigraphy of the Emanuel Formation, Canning Basin, Western Australia. Palaeontographica A 240: 105-144. [ Links ]
45. Legg, D.P. 1976. Ordovician trilobites and graptolites from the Canning Basin, Western Australia. Geologica et Palaeontologica 10: 1-58. [ Links ]
46. Linnarsson, J.G.O. 1869. Om Vestergötlands Cambriska och Siluriska aflagringar. Kongliga Svenska Vetenskaps-Akademiens Handlingar 8: 1-89, láms. 1, 2. [ Links ]
47. Lu, Y., Chu, C., Chen, Y., Zhou, Z., Chen, J., Lu, G., Yu, W., Chen, X. y Xu, H. 1976. Ordovician biostratigraphy and palaeozoogeography of China. Memoirs of the Nanjing Institute of Geology and Palaeontology, Academia Sinica 7: 1-83. [ Links ]
48. Ludvigsen, R. 1982. Upper Cambrian and Lower Ordovician trilobite biostratigraphy of the Rabbitkettle Formation, western District of Mackenzie. Life Science Contributions, Royal Ontario Museum 134: 188 pp. [ Links ]
49. Ludvigsen, R., Westrop, S.R. y Kindle, C.H. 1989. Sunwaptan (Upper Cambrian) trilobites of the Cow Head Group, western Newfoundland, Canada. Palaeontographica Canadiana 6: 1-175. [ Links ]
50. Malanca, S. 1996. Morfología y ontogenia de un nuevo Shumardiidae (Trilobita) del Tremadociano de la Sierra de Mojotoro. 12° Congreso Geológico de Bolivia (Tarija), Memorias 1: 391-399. [ Links ]
51. Merz-Preiß, M. 2000. Calcification in cyanobacteria. En: R.E. Riding y S.M. Awramik (eds.), Microbial sediments, Springer-Verlag, Berlin, pp. 50-56. [ Links ]
52. Moya, M.C., Malanca, S., Hongn, F.D. y Bahlburg, H. 1993. El Tremadociano temprano en la Puna occidental, Argentina. 12° Congreso Geológico Argentino y 2° Congreso de Exploración de Hidrocarburos (Mendoza), Actas 2: 20-30. [ Links ]
53. Müller, K. y Walossek, D. 1987. Morphology, ontogeny, and life habit of Agnostus pisiformis from the Upper Cambrian of Sweden. Fossils and Strata 19: 1-124. [ Links ]
54. Nikolaisen, F. y Henningsmoen, G. 1985. Upper Cambrian and lower Tremadoc olenid trilobites from the Digermul peninsula, Finmark, northern Norway. NGU Norges Geologiske Undersokelse Bulletin 400: 1-49. [ Links ]
55. Noffke, N. 2000. Extensive microbial mats and their influences on the erosional and depositional dynamics of a siliciclastic cold water environment (Lower Arenigian, Montagne Noire, France). Sedimentary Geology 136: 207-215. [ Links ]
56. Noffke, N., Gerdes, G., Klenke, T. y Krumbein, W. 1997. A microscopic sedimentary succession of graded sand and microbial mats in modern siliciclastic tidal flats. Sedimentary Geology 110: 1-6. [ Links ]
57. Noffke, N., Gerdes, G., Klenke, T. y Krumbein, W. 2001. Microbially induced sedimentary structuresa new category within the classification of primary sedimentary structures. Journal of Sedimentary Research 71: 649-656. [ Links ]
58. Noffke, N., Knoll, A. y Grotzinger, J. 2002. Sedimentary controls on the formation and preservation of microbial mats in siliciclastic deposits: A case study from the Upper Neoproterozoic Nama Group, Namibia. Palaios 17: 533-544. [ Links ]
59. O´Brien, N. 1996. Shale lamination and sedimentary processes. En: A.E.S. Kemp (ed.), Palaeoclimatology and Palaeoceanography from laminated sediments, Geological Society Special Publication 116: 23-36. [ Links ]
60. O´Brien, N. y Slatt, R. M. 1990. Argillaceous Rock Atlas, Springer, New York, 141 pp. [ Links ]
61. Oschmann, W. 2000. Microbes and black shales. En: R.E. Riding y S.M. Awramik (eds.), Microbial sediments, Springer-Verlag, Berlin, pp. 137-148. [ Links ]
62. Palmer, A.R. 1968. Cambrian trilobites of East-Central Alaska. Geological Survey Professional Paper 559-B: 1-115. [ Links ]
63. Peng, S. 1984. Cambrian-Ordovician boundary in the CiliTaoyuan border area, northwestern Hunan with descriptions of relative trilobites. En: Stratigraphy and Palaeontology of Systemic Boundaries in China, Cambrian-Ordovician Boundary (1), Ashui Sciences Technology Press, Hebei, pp. 285-405. [ Links ]
64. Peng, S. 1990. Tremadoc stratigraphy and trilobite faunas of northwestern Hunan. 2. Trilobites from the Panjiazui Formation and the Madaoyu Formation in Jiangnan Slope Belt. Beringeria 2: 55-171. [ Links ]
65. Peng, S. 1991. Tremadocian trilobites from Guotang Formation, Luxi, Western Hunan. Acta Palaeontologica Sinica 30: 141-166. [ Links ]
66. Pflüger, F. 1999. Mat ground structures and Redox Facies. Palaios 14: 25-39. [ Links ]
67. Pratt, B.R. 1988. An Ibexian (Early Ordovician) trilobite faunule from the type section of the Rabbitkettle Formation (southern Mackenzie Mountains, Northwest Territories). Canadian Journal of Earth Sciences 25: 1595-1607. [ Links ]
68. Pribyl, A. y Vanek, J. 1980. Ordovician trilobites of Bolivia. Rozpravy Ceskoslovenske Akademie Ved 90: 1-90. [ Links ]
69. Quian, Y. 1986. Trilobites. En: J.Y. Chen (ed.), Aspects in CambrianOrdovician boundary in Dayangcha, China, China Prospect Publishing House, Beijing, pp. 255-313. [ Links ]
70. Rasetti, F. 1944. Upper Cambrian trilobites from the Lévis Conglomerate. Journal of Paleontology 18: 229-258. [ Links ]
71. Raymond, P.E. 1924. New Upper Cambrian and Lower Ordovician trilobites from Vermont. Proceedings of the Boston Society of Natural History 37: 389-446. [ Links ]
72. Robison, R.A. 1972. Mode of life of agnostid trilobites. International Geological Congress, 24th Session, Section 7: 33-40. [ Links ]
73. Robison, R.A. y Pantoja-Alor, J. 1968. Tremadocian trilobites from the Nochixtlán region, Oaxaca, Mexico. Journal of Paleontology 42: 767-800. [ Links ]
74. Rushton, A.W.A. 1982. The biostratigraphy and correlation of the Merioneth-Tremadoc Series boundary in North Wales. En: M. G. Basset y W.T. Dean (eds.), The Cambrian-Ordovician boundary: sections, fossil distributions, and correlations, National Museum of Wales, Geological Series 3: 41-59. [ Links ]
75. Salter, J.W. 1849. Figures and descriptions illustrative of British organic remains. Memoirs of the Geological Survey 2: 1-39, 10 pls. [ Links ]
76. Salter, J.W. 1859. En: R.I. Murchinson, Siluria (3º ed.), John Murray, London. [ Links ]
77. Salter, J.W. 1866. A monograph of British trilobites. Part 3. Monograph of the Palaeontographical Society, pp. 129-176, pls. 15-25. [ Links ]
78. Schieber, J. 1986. The possible role of benthic microbial mats during the formation of carbonaceous shales in shallow MidProterozoic basins. Sedimentology 33: 521-536. [ Links ]
79. Schieber, J. 1998. Possible indicators of microbial mat deposits in shales and sandstones: examples from the Mid-Proterozoic Belt Supergroup, Montana, U.S.A. Sedimentary Geology 120:105-124. [ Links ]
80. Schieber, J. 1999. Microbial mats in terrigenous clastics: the challenge of identification in the rock record. Palaios 14: 3-12. [ Links ]
81. Sdzuy, K. 1955. Die Fauna der Leimitz-Schefer (Tremadoc). Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft 492: 1-74. [ Links ]
82. Shaw, A.B. 1951. The paleontology of northwestern Vermont. I. New Late Cambrian trilobites. Journal of Paleontology 25: 97-114. [ Links ]
83. Shergold, J.H. 1972. Late Upper Cambrian trilobites from the Gola Beds, western Queensland. Bulletin of the Bureau of Mineral Resources of Australia, Geology and Geophysics 112: 126 pp., 19 lám. [ Links ]
84. Shergold, J.H. 1975. Late Cambrian and Early Ordovician trilobites from the Burke River Structural Belt, western Queensland. Bulletin of the Bureau of Mineral Resources of Australia, Geology and Geophysics 153: 251 pp, 58 lám. [ Links ]
85. Suárez Soruco, R. 1975. Nuevos trilobites del Tremadociano inferior del sur de Bolivia. Revista Técnica de Yacimientos Petrolíferos Fiscales Bolivianos 4: 129-146. [ Links ]
86. Taylor, M.E. 1976. Indigenous and redeposited trilobites from late Cambrian basinal environments of Central Nevada. Journal of Paleontology 50: 668-700. [ Links ]
87. Tortello, M.F. 2003. Biostratigraphic significance of the latest Cambrian-earliest Ordovician agnostoid trilobites from northwestern Argentina. Geologica Acta 1: 61-72. [ Links ]
88. Tortello, M.F. y Clarkson, E.N.K. 2006. Ontogenia, morfología y ecdisis de Parabolina frequens argentina (Kayser) (Trilobita, Olenidae) del Cámbrico Tardío del noroeste argentino. 9° Congreso Argentino de Paleontología y Bioestratigrafía (Córdoba), Resúmenes: p. 141. [ Links ]
89. Tortello, M.F. y Esteban, S.B. 1997. Significado bioestratigráfico de una asociación de trilobites del tramo basal de la Formación Volcancito (Sistema de Famatina, La Rioja, Argentina). Ameghiniana 34: 265-270. [ Links ]
90. Tortello, M.F. y Esteban, S.B. 1998. La transición Cámbrico-Ordovícica en la Formación Volcancito (Sistema de Famatina, La Rioja, Argentina). 7° Congreso Argentino de Paleontología y Bioestratigrafía (Bahía Blanca), Resúmenes: 93. [ Links ]
91. Tortello, M.F. y Esteban, S.B. 1999. La transición Cámbrico-Ordovícico en la Formación Volcancito (sierra de Famatina, La Rioja, Argentina). Ameghiniana 36: 371-387. [ Links ]
92. Tortello, M.F. y Esteban, S.B. 2003a. Lower Ordovician stratigraphy and trilobite faunas from the southern Famatina Range, La Rioja, Argentina. Special Papers in Palaeontology 70: 213-239. [ Links ]
93. Tortello, M.F. y Esteban, S.B. 2003b. Trilobites del Cámbrico Tardío de la Formación Lampazar (sierra de Cajas, Jujuy, Argentina). Implicancias bioestratigráficas y paleoambientales. Ameghiniana 40: 323-344. [ Links ]
94. Tortello, M.F. y Esteban, S.B. 2005. Trilobites agnóstidos del Cámbrico Tardío de la sierra de Famatina (La Rioja, Argentina). Significado bioestratigráfico y paleoambiental. Serie Correlación Geológica 19: 157-168. [ Links ]
95. Tortello, M.F. y Rao, R.I. 2000. Trilobites y conodontes del Ordovícico Temprano del Angosto de Lampazar (provincia de Salta, Argentina). Boletín Geológico y Minero 111: 61-84. [ Links ]
96. Tortello, M.F., Aceñolaza, G.F. y Muruaga, C. 1996. Presencia del género Pseudokainella (Trilobita, Remopleuridacea) en el Cambro-Ordovícico de la Sierra de La Candelaria, Salta, Argentina. Ameghiniana 33: 99-102. [ Links ]
97. Toselli, G.A. 1977. El Paleozoico Inferior y Medio de la región de Volcancito, Sierra de Famatina, Provincia de La Rioja, Rep. Argentina. Acta Geológica Lilloana 14: 83-104. [ Links ]
98. Vaccari, N.E. y Waisfeld, B.G. 2000. Trilobites tremadocianos de la Formación Las Vicuñas, Puna occidental, Provincia de Salta, Argentina. 14° Congreso Geológico Boliviano (La Paz), Memorias: 164-169. [ Links ]
99. Vogdes, A.W. 1890. A bibliography of Paleozoic Crustacea from 1698 to 1889 including a list of North American species and a systematic arrangement of genera. United States Geological Survey Bulletin 63: 1-177. [ Links ]
100. Waisfeld, B.G. y Vaccari, N.E. 2003. Trilobites. En: J.L. Benedetto (ed.), Ordovician Fossils of Argentina, Secretaría de Ciencia y Tecnología, Universidad Nacional de Córdoba, pp. 295-409. [ Links ]
101. Waisfeld, B.G., Vaccari, N.E., Chatterton, B.D.E. y Edgecombe, G. D. 2001. Systematics of Shumardiidae (Trilobita), with new species from the Ordovician of Argentina. Journal of Paleontology 75: 827-859. [ Links ]
102. Wright, A.J., Cooper, R.A. y Simes, J.E. 1994. Cambrian and Ordovician faunas and stratigraphy, Mt. Patriarch, New Zealand. New Zealand Journal of Geology and Geophysics 37: 437-476. [ Links ]
103. Zeballo, F. y Tortello, M.F. 2005. Trilobites del Cámbrico Tardío- Ordovícico Temprano del área de Alfarcito, Tilcara, Cordillera Oriental de Jujuy, Argentina. Ameghiniana 42: 127-142. [ Links ]
104. Zylinska, A. 2001. Late Cambrian trilobites from the Holy Cross Mountains, central Poland. Acta Geologica Polonica 51: 333-383. [ Links ]
Recibido: 16 de febrero de 2006.
Aceptado: 3 de marzo de 2007.

