










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.44 n.4 Buenos Aires oct./dic. 2007
Sigmodontinos (Rodentia: Cricetidae) del límite Pleistoceno-Holoceno en el Valle de Tafí (Tucumán, Argentina): taxonomía, tafonomía y significación paleoambiental
Pablo E. Ortiz1 y J. Pablo Jayat2
1Cátedra de Paleontología II, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán, Miguel Lillo 205, 4000 San Miguel de Tucumán, Tucumán, Argentina. peortiz@uolsinectis.com.ar
2Laboratorio de Investigaciones Ecológicas de las Yungas (LIEY), Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán, Casilla de Correo 34, 4107 Yerba Buena, Tucumán, Argentina.
Abstract. Sigmodontines (Rodentia: Cricetidae) from the Pleistocene-Holocene Boundary of the Tafí Valley (Tucumán, Argentina): Taxonomy, Taphonomy and Paleoenvironmental significance. A fossilsample of sigmodontine rodents from Tafí del Valle, Tucumán, Argentina (26º 51' 43'' S, 65º 43' 06'' W, 2007 m) was studied. A radiocarbon datation on bone fragments indicates that the sample can be referred to the Pleistocene-Holocene boundary (10.25 - 9.65 cal. ky BP). The sample is dominated by the phyllotines Calomys cf. C. musculinus (Thomas) and Phyllotis sp., followed by Akodon spegazzinii Thomas and Reithrodon auritus (Fischer). At the microscopy the surface of bones and teeth shows evidences of corrosion and digestion suggesting the assemblage was generated by the predatory activity of owls. Most of the species from the assemblage correspond to common elements of high altitude grasslands environments from northwestern Argentina, except for R. auritus. All species are currently sympatric in an area about 1000 meters above the fossiliferous locality, in a colder and drier environment than the study site. These facts allowed to infer drier and colder paleoenvironmental conditions at the deposition time than the current ones during the Pleistocene-Holocene boundary in Tafí del Valle.
Resumen. Se estudió una muestra de roedores sigmodontinos fósiles provenientes de las proximidades de Tafí del Valle, provincia de Tucumán, Argentina (26º 51' 43'' S, 65º 43' 06'' O, 2007 m de altitud). Una datación radiocarbónica efectuada sobre restos óseos permite referir la muestra al límite Pleistoceno- Holoceno (10,25 - 9,65 ka cal. AP). El conjunto está dominado por los filotinos Calomys cf. C. musculinus (Thomas) y Phyllotis sp., seguidos por Akodon spegazzinii Thomas y Reithrodon auritus (Fischer). El análisis de los restos al microscopio ha revelado la presencia de marcas de corrosión en huesos y el esmalte dentario, sugiriendo que la acumulación fue generada por la actividad trófica de lechuzas. La mayor parte de las especies registradas corresponden a elementos típicos de los ambientes de pastizales de neblina del noroeste argentino, con la excepción de R. auritus. Todas las especies tienen un área de simpatría a una altitud aproximada de 1000 metros por arriba de la localidad fosilífera, en un ambiente caracterizado por condiciones climáticas más frías y secas que las actuales en el área del yacimiento. Esto permite inferir condiciones paleoambientales de mayor aridez y menor temperatura para Tafí del Valle durante el límite Pleistoceno-Holoceno.
Key words. Pleistocene-Holocene boundary; Sigmodontinae; Owl pellets; Northwestern Argentina; Tucumán; Palaeoenvironments; Taxonomy.
Palabras clave. Límite Pleistoceno-Holoceno; Sigmodontinae; Egagrópilas; Noroeste de Argentina; Tucumán; Paleoambientes; Taxonomía.
Introducción
El noroeste de Argentina comprende un área con una riqueza particularmente alta de roedores sigmodontinos, con algo más de 45 especies vivientes reconocidas en la actualidad (Mares et al., 1997; Díaz, 1999; Díaz et al., 2000; Barquez et al., 2006; Jayat et al., 2006). Entre las posibles causas de esta riqueza puede señalarse la gran heterogeneidad ambiental de la región (Mares et al., 1996, 1997), consecuencia, en gran medida, de la exposición diferencial de los elementos del paisaje a los vientos húmedos provenientes del anticiclón del atlántico sur (Bianchi y Yáñez, 1992; Minetti et al., 2005). El noroeste argentino se caracteriza por la presencia de importantes cordones montañosos de diferente altitud y posición geográfica separados por valles, bolsones y numerosos cursos de agua (Bobba y Hernández, 2005). Las comunidades de sigmodontinos del noroeste han sido escasamente estudiadas en numerosos aspectos de su biología, aunque en los últimos años se han realizado algunos avances que incluyen relevamientos preliminares sobre diversidad y distribución (Barquez et al., 1991; Mares et al., 1989, 1997; Díaz, 1999; Díaz et al., 2000; Ortiz et al., 2000a; Jayat et al., 2006, 2007) así como aspectos ecológicos y biogeográficos (e.g., Mares, 1977; Ojeda y Mares, 1989; Tabeni et al., 2004; Jayat y Pacheco, 2006). Un tópico especialmente poco conocido es el registro fósil del grupo en la región, restringido a escasas contribuciones limitadas al lapso Pleistoceno tardío-Holoceno (Ortiz et al., 2000b; Ortiz, 2001; Ortiz y Pardiñas, 2001; Pardiñas y Ortiz, 2001; Teta y Ortiz, 2002; Ortiz y Jayat, 2007).
Un área de particular importancia paleontológica es el valle de Tafí, provincia de Tucumán, donde se ha estudiado un importante agregado de micromamíferos proveniente del Pleistoceno medio-superior en la localidad de La Angostura (Ortiz y Pardiñas, 2001). Recientes actividades de prospección paleontológica determinaron el hallazgo de una muestra de roedores asignable al límite Pleistoceno-Holoceno. El estudio de estos restos ha permitido el análisis y la comparación de los ensambles de sigmodontinos provenientes de una misma área en tres momentos del Cuaternario: Pleistoceno medio-superior, límite Pleistoceno-Holoceno y presente. A partir del estudio taxonómico, el análisis del origen tafonómico de los restos, la evaluación de los requerimientos de hábitat de las especies involucradas y su comparación con los agregados del Pleistoceno medio-superior y el actual, se discuten algunos aspectos relativos a la historia paleoambiental de la región.
Área de estudio y contexto ambiental
El yacimiento portador de los sigmodontinos estudiados se encuentra ubicado aproximadamente 2 km al sudoeste de la localidad de Tafí del Valle, departamento homónimo, provincia de Tucumán (26º 51' 43'' S y 65º 43' 06''O; 2007 m de altitud). El valle de Tafí es una depresión intermontana de unos 300 km2 al pie de la vertiente oriental de la Sierra del Aconquija. Las áreas más bajas del valle, donde está localizado el yacimiento, se sitúan entre 1900 y 2100 m de altitud y están rodeadas por formaciones montañosas que superan los 3200 m hacia el sur y el este y por cumbres que hacia el oeste y el norte superan los 4400 m (figura 1).

Figura 1. Ubicación geográfica del área del valle de Tafí (provincia de Tucumán), pisos altitudinales de vegetación y localidades fosilíferas: 1, Yacimiento de Tafí del Valle (provincia de Tucumán, límite Pleistoceno-Holoceno); 2, Yacimiento de La Angostura (provincia de Tucumán, Pleistoceno medio-superior) / geographic location of the Tafí valley area (Province of Tucumán), vegetation altitudinal belts and fossil localities: 1, Tafí del Valle fossil locality (Province of Tucumán, Pleistocene-Holocene boundary); 2, La Angostura fossil locality (Province of Tucumán, Middle-Upper Pleistocene).
El tipo de vegetación predominante en la actualidad corresponde a una comunidad herbácea biestratificada dominada por gramíneas conocida como Praderas Montanas (Cabrera, 1976), Pastizales de Altura (Vervoorst, 1982) o Pastizales de Neblina (Brown et al., 2001). Esta vegetación limita hacia el este y el sur con el estrato superior de la Selva de Yungas, dominado por Alnus acuminata Kunth (aliso del cerro) y conocido como Bosque Montano (Brown et al., 2001). El límite superior de este bosque oscila entre 1800 y 2900 m (Grau, 1985) y se desarrolla principalmente sobre laderas expuestas a los vientos húmedos del sudeste (Bell, 1991). La particular conformación topográfica del valle y las variaciones de humedad relacionadas con la misma determinan la presencia de parches de este bosque inmersos en una matriz de pastizales (figura 1). Hacia el norte y el oeste, a una altitud aproximada de 3000 m, los pastizales limitan en amplio ecotono con comunidades vegetales de tipo semiárido correspondientes a la estepa puneña y vegetación altoandina (Vervoorst, 1982), ambientes caracterizados por una menor cobertura vegetal del suelo, un sustrato rocoso y una mayor aridez.
La precipitaci ón anual promedio en el área del valle es de 430 mm, concentrada en el verano, y la temperatura media anual de 13,2°C (Torres Bruchmann, 1972; Sesma et al., 1998). Sin embargo, estos valores varían de acuerdo a la altitud y a la exposición de las laderas en relación a la posición del sol y de la dirección de los vientos húmedos predominantes del sudeste. Grandes áreas del valle muestran en la actualidad, particularmente sus zonas de menor altitud, un elevado grado de degradación antrópica por sobrepastoreo y actividades agrícolas.
Entre los sigmodontinos dominantes en ambientes de pastizales de neblina en Tucum án se incluye a Akodon spegazzinii Thomas, Necromys lactens (Thomas) y Phyllotis osilae J. A. Allen. En zonas de transición con el bosque montano se encuentran especies no registradas en pastizales puros como Abrothrix illuteus Thomas y Oxymycterus paramensis Thomas. En el ecotono con ambientes semiáridos, a mayor altitud, se registran Phyllotis xanthopygus (Waterhouse), Reithrodon auritus (Fischer) y Calomys musculinus (Thomas), normalmente ausentes en pastizales húmedos. Esta última especie es dominante en aquellas áreas afectadas por la agricultura y el sobrepastoreo (Bellocq, 1990; Pardiñas, 1999; Pardiñas et al., 2000).
Contexto geológico
Antecedentes
El agregado estudiado fue recuperado en sedimentos correspondientes a lo que Collantes et al. (1993) han caracterizado como Formación Tafí del Valle, un complejo de loess y paleosuelos que, de acuerdo a estos autores, se depositó durante el Pleistoceno superior y principios del Holoceno. En el paraestratotipo de esta unidad geológica el perfil ex- puesto (42,5 m de espesor) contiene 28 paleosuelos intercalados con capas de loess (Sayago et al., 1998; Zinck y Sayago, 1999, 2001). Estos autores refirieron una edad radiocarbónica de 27,6 ka antes del presente (en adelante AP) para la base del perfil y de 17,5 ka AP a 5 metros de la parte cuspidal, identificando para ese intervalo 20 niveles de paleosuelos. Asumiendo una evolución similar para los depósitos de la sección superior señalaron que dicha formación se desarrolló durante el Pleistoceno tardío y culminó a principios del Holoceno. Dataciones radiocarbónicas recientes realizadas por Sayago et al. (2005) determinaron un intervalo temporal para la formación entre 47 ka y 8,6 ka. Sin embargo, la parte basal del paraestratotipo fue reexaminada por Kemp et al. (2003), quienes a través de dataciones por luminiscencia estimulada ópticamente (OSL) brindaron una edad de 195 ka. Dataciones adicionales efectuadas por Kemp et al. (2004) y Carter-Stiglitz et al. (2006) a partir de perfiles realizados en otros sectores del valle indicaron edades incluidas en el intervalo entre 190,5 a 64 ka AP. Además, estudios magnetoestratigráficos realizados por Schellenberger et al. (2003) en una sección próxima al paraestratotipo determinaron una edad mínima de 1,15 millones de años para el comienzo de depositación de la formación. Claramente, estos nuevos datos aparecen como más confiables desde un punto de vista pedosedimentario ya que extienden en gran medida el lapso de sedimentación para toda la secuencia, brindando tasas más lentas y realistas que aquellas originalmente sugeridas por Zinck y Sayago (1999, 2001).
Entre los fósiles de mamíferos recuperados en la Formación Tafí del Valle se han registrado Chaetophractus vellerosus (Gray) (Herrera y Ortiz, 2003), Glyptodon reticulatus (Owen), G. clavipes (Owen) (R. Duarte, com. pers., 1999), Megatherium americanum Cuvier (Rufino y Mulé, 1994), Paraglossotherium elmollarum Esteban (Esteban, 1993), Cuvieronius cf. C. hyodon (Fischer), Hippidion sp. y Paleolama sp. (Powell, 1998).
Descripción del perfil estratigráfico
Los sedimentos portadores del agregado estudiado se encuentran hacia la parte alta de la Formación Tafí del Valle, en un afloramiento de orientación norte- sur. Los sedimentos, en particular el nivel con los fósiles de sigmodontinos, se relacionan con depósitos fluviales, en donde se observan estructuras en artesa y lenticulares. El perfil relevado fue descripto de base a techo y dividido en cuatro secciones con las siguientes características (figura 2).

Figura 2. Perfil estratigráfico de los niveles portadores de los restos de sigmodontinos estudiados (Tafí del Valle, provincia de Tucumán, límite Pleistoceno-Holoceno) / stratigraphical section of the fossil sigmodontines bearing levels (Tafí del Valle, Province of Tucumán, Pleistocene- Holocene boundary).
Sección 1. La base no es visible y presenta un espesor de 74 cm en promedio. Está compuesto por un conjunto de arenas finas con lentes de arenas medias a gruesas. Los rodados están compuestos por clastos metamórficos y graníticos de 4 a 10 cm. El color es pardo amarillento (10YR7/4). Presenta estratificación en artesa y contacto superior irregular. Los restos de roedores estudiados fueron exhumados en esta sección.
Sección 2. Está compuesta por guijarros y guijones. El espesor varía entre 16 y 23 cm. La fábrica es clasto soporte y el tamaño de éstos varía entre 4 y 13 cm, con predominancia de aquellos compuestos por materiales limo arcillosos castaños claros (10YR8/2) sobre los metamórficos y graníticos. La estructura es maciza y presenta un color 10YR7/4. El contacto superior es irregular.
Sección 3. Presenta un espesor de 15 cm. Compuesta por areniscas muy finas a finas. Muestra estratificación lenticular y rodados dispersos de guijarros (2 a 3 cm). Esta capa se encuentra cementada y muestra un contacto superior irregular.
Sección 4. Con un espesor de 90 a 120 cm, es una intercalación de arenas finas y medias con lentes de arenas gruesas de color 10YR5/4. Se observan marcas de raíces, material carbonoso y su aspecto es macizo y edafizado de color marrón amarillento (10YR7/4).
Edad de la muestra
La posición estratigráfica del agregado estudiado, en la parte superior de la Formación Tafí del Valle, permitió originalmente referir la muestra al Pleistoceno más tardío u Holoceno inferior. Esto fue corroborado por una datación radiocarbónica realizada por Espectometría de Masa por Acelerador (AMS) sobre un hueso de roedor, la que brindó una edad radiocarbono convencional de 8830 ± 70 años AP, calibrada en años calendario sobre la base de los últimos datos atmosféricos provistos por Reimer et al. (2004) entre 10,25 y 9,65 ka AP con 95,4% de probabilidad.
Materiales y métodos
Los restos estudiados conformaban un agregado constituido por poco más de 770 especímenes (sensu Grayson, 1984) craneanos y postcraneanos y fueron recolectados por los autores y auxiliares de campo de la Universidad Nacional de Tucumán mediante tamizado en seco de los sedimentos portadores.
Las determinaciones taxonómicas se efectuaron a partir del estudio de 128 restos cráneodentarios, sobre la base de la comparación con material actual de referencia proveniente del área del valle de Tafí y de otros sectores del noroeste argentino (Apéndice). Los ejemplares utilizados en la comparación provienen principalmente de los catálogos de campo de J. Pablo Jayat (JPJ) y Pablo E. Ortiz (PEO), de material recuperado en egagrópilas de lechuzas recolectadas en el mismo valle (PEO-E) y del estudio de especímenes depositados en las siguientes colecciones: Colección de Mamíferos del Instituto-Fundación Miguel Lillo (CML), Tucumán; Museo de Ciencias Naturales "Bernardino Rivadavia" (MACN), Buenos Aires y Museo Municipal de Ciencias Naturales "Lorenzo Scaglia" (MMPMa), Mar del Plata. Los restos fósiles de sigmodontinos estudiados se encuentran depositados en la Colección de Paleontología de Vertebrados del Instituto-Fundación Miguel Lillo (PVL), Tucumán.
La datación radiocarbónica por AMS fue llevada a cabo sobre hueso de una hemimandíbula de Phyllotis sp. (muestra LTL669A) y realizada en el Centro di Datazione e Diagnostica del Dipartimento di Ingegneria dell'Innovazione, Università di Lecce, Italia (CEDAD).
El criterio taxonómico adoptado corresponde al de Barquez et al. (2006). Para la descripción de la superficie oclusal de los molares se siguió la terminología propuesta por Reig (1977). Las medidas, expresadas en milímetros, fueron tomadas con calibre digital (con precisión de 0.02 mm) y ocular micrométrico adosado a lupa binocular. Las longitudes para las series molares son alveolares y los largos y anchos de los molares corresponden a sus valores máximos.
Las regiones fitogeográficas reconocidas aquí corresponden a las provincias fitogeográficas de Cabrera (1976), con algunas excepciones aclaradas en el texto.
Con el propósito de establecer el origen del agregado se procedió a la categorización de marcas y alteraciones superficiales de los huesos y dientes siguiendo a Andrews (1990) y Fernández Jalvo y Andrews (1992).
Como parámetro actualístico de comparación para efectuar inferencias paleoambientales se utilizaron datos provenientes de muestras de egagrópilas de Tyto alba (Scopoli) y Athene cunicularia (Molina) recuperadas en diversas localidades del valle de Tafí (véase Ortiz y Pardiñas, 2001) y de otros sectores en el noroeste argentino. Se tuvieron en cuenta las preferencias de hábitat conocidas para las poblaciones actuales de las especies registradas en el yacimiento (Jayat y Pacheco, 2006; Jayat et al., 2006), así como también las características ambientales de las áreas donde las mismas se encuentran en simpatría.
Los valores de riqueza específica para los agregados estudiados se expresan como el índice alfa derivado del modelo de serie logarítmica de Fisher et al. (1943) y tratado en detalle por Magurran (1988). Este índice enfatiza la medición de la riqueza de especies de una comunidad, presenta una gran capacidad de discriminación entre muestras, baja sensitividad a tamaños muestrales diferentes y ha sido ampliamentre utilizado en estudios de biodiversidad (Magurran, 1988).
Otras abreviaturas utilizadas en el texto y en las tablas incluyen: MD: hemimaxilar derecho; MI: hemimaxilar izquierdo; HD: hemimandíbula derecha; HI: hemimandíbula izquierda; M1, M2, M3, m1, m2 y m3: primer, segundo y tercer molar superior e inferior respectivamente; MNI = número mínimo de individuos; NISP = número de especímenes identificados. PF: Provincia Fitogeográfica.
Resultados
Los sigmodontinos registrados en el agregado fósil de Tafí del Valle corresponden a seis especies vivientes, las cuales forman parte de la fauna actual del área del valle o sus proximidades. Se destaca en la asociación la dominancia de los filotinos Phyllotis sp. y Calomys cf. C. musculinus. En la muestra se registraron también los roedores caviomorfos Galea musteloides Meyen y una especie indeterminada de Ctenomys Blainville.
Tafonomía
En el agregado, altamente concentrado en su yacencia original, se han recuperado todos los elementos del esqueleto en buen estado de preservación y elevado grado de integridad. Parte del material presentó claras señales de digestión (Andrews, 1990), con el 35% de los 43 incisivos examinados afectados por este proceso. Un porcentaje menor de corrosión también se observó en unas pocas mandíbulas y elementos postcraneanos. Estas evidencias sugieren que la asociación fue generada por la actividad trófica de lechuzas. La frecuencia de representación de los incisivos afectados ubica a la muestra de Tafí del Valle en la categoría 2 de Andrews (1990), quien incluyó aquí a una especie del género Bubo Duméril y a una del género Asio Brisson. Por otro lado, la intensidad de la digestión en los incisivos es comparable a lo documentado para Tyto alba y Asio flammeus (Pontoppidan), incluidas por Andrews (1990) en su categoría 1.
Comparación con agregados cuaternarios y actuales
La muestra fósil estudiada está dominada por Phyllotis spp. y Calomys cf. C. musculinus, seguidos por Akodon spegazzinii y Reithrodon auritus (tabla 1; figura 5). Las características más notorias de la muestra en comparación con la comunidad actual de sigmodontinos de la misma localidad, documentada a partir de egagrópilas de Tyto alba (MNI = 75), radican en la ausencia de Oligoryzomys flavescens y Abrothrix illuteus y en la presencia de R. auritus, actualmente ausente a la altitud del yacimiento (tabla 1; figura 5). Las especies dominantes en la muestra actual son Calomys cf. C. musculinus y A. spegazzinii seguidas por Phyllotis osilae. Por otra parte, en el agregado del Pleistoceno medio-superior de La Angostura, al sudeste del valle (figura 1), se han registrado 12 especies de sigmodontinos (Ortiz y Pardiñas, 2001). Este ensamble está dominado por una especie extinta, Tafimys powelli Ortiz et al., seguido por A. spegazziinii (como Akodon sp. en Ortiz y Pardiñas, 2001) y Andinomys edax. Además de la alta frecuencia de la especie extinta, el agregado de La Angostura se destaca por la presencia de R. auritus y de Neotomys ebriosus Thomas, esta última registrada en la provincia de Tucumán sólo para ese lapso (tabla 1; figura 5).
Tabla 1. MNI, % MNI, NISP y % NISP para los roedores registrados en los agregados fósiles de La Angostura (provincia de Tucumán, Pleistoceno medio-superior) y de Tafí del Valle (provincia de Tucumán, límite Pleistoceno-Holoceno) y de muestras actuales de egagrópilas de La Angostura y de Tafí del Valle. Los datos de las muestras fósiles y actuales de La Angostura fueron tomados de Ortiz y Pardiñas (2001) y Ortiz (2001) / MNI, MNI%, NISP and NISP% for registered rodents in La Angostura (Province of Tucumán, Middle-Upper Pleistocene) and Tafí del Valle (Pleistocene-Holocene boundary) fossil assemblages and recent samples of owl pellets of La Angostura and Tafí del Valle. The data of the fossil and recent samples of La Angostura are from Ortiz and Pardiñas (2001) and Ortiz (2001).

Paleontología sistemática
Orden RODENTIA Bowdich, 1821
Familia CRICETIDAE Fisher, 1817
Subfamilia SIGMODONTINAE Wagner, 1843
Tribu AKODONTINI Vorontzov, 1959 (sensu D'Elía, 2003)Género Akodon Meyen, 1833
Especie tipo. Akodon boliviensis Meyen, 1833.
Akodon spegazzinii Thomas, 1897
Figuras 3.8, 4.1 y 4.2; tablas 1 y 2

Figura 3. Vistas labiales de hemimandíbulas y hemimaxilares de algunos sigmodontinos del yacimiento de Tafí del Valle (provincia de Tucumán, límite Pleistoceno-Holoceno) / labial views of hemimandibles and hemimaxillae of some sigmodontines from Tafí del Valle (Province of Tucumán, Pleistocene-Holocene boundary). 1-2, Reithrodon auritus (Fischer). 1, rama mandibular izquierda (PVL 5752) / left mandible; 2, hemimaxilar izquierdo con placa cigomática (PVL 5753) / left maxilla with zygomatic plate. 3, Andinomys edax Thomas, rama mandibular derecha (PVL 5735) / right mandible. 4, Calomys cf. C. musculinus (Thomas), rama mandibular izquierda (PVL 5771) / left mandible. 5, Necromys cf. N. lactens (Thomas), hemimaxilar derecho con placa cigomática (PVL 5734) / right maxilla with zygomatic plate. 6-7, Phyllotis sp. 6, rama mandibular derecha (PVL 5793) / right mandible; 7, hemimaxilar derecho con placa cigomática (PVL 5823) / right maxilla with zygomatic plate. 8, Akodon spegazzinii Thomas, rama mandibular derecha (PVL 5843) / right mandible. Escala =10 mm / scale bar = 10 mm.

Figura 4. Vistas oclusales de los molares de los sigmodontinos representados en el yacimiento de Tafí del Valle (provincia de Tucumán, límite Pleistoceno-Holoceno) / molar occlusal views of the sigmodontines from Tafí del Valle (Province of Tucumán, Pleistocene-Holocene boundary). 1-2, Akodon spegazzinii Thomas. 1, M1-2 derechos (PVL 5847) / right M1-2; 2, m1-m2 derechos (PVL 5840) / right m1-m2. 3, Andinomys edax Thomas, m1 derecho (PVL 5735) / right m1. 4, Calomys cf. C. musculinus (Thomas) m1-2 izquierdos (PVL 5773) / left m1-2. 5, Phyllotis sp., m1 derecho (PVL 5793) / right m1. 6, Phyllotis osilae J. A. Allen, M1-M3 derechos (PVL 5827) / right M1-M3. 7- 8, Reithrodon auritus (Fischer). 7, m1-m3 derechos (PVL 5755) / right m1-m3; 8, M1-M3 izquierdos (PVL 5753) / left M1-M3. 9, Necromys cf. N. lactens (Thomas), M1-2 derechos (PVL 5734) / right M1-2. Escala = 2 mm / scale bar = 2 mm.
Tabla 2. Medidas de los molares y series dentarias de los sigmodontinos fósiles de Tafí del Valle (provincia de Tucumán, límite Pleistoceno-Holoceno). Se indica, cuando corresponde, media, desvío estándar entre paréntesis y número de ejemplares / measurements of the molars and toothrows of the sigmodontine rodents of Tafí del Valle (Province of Tucumán, Pleistocene-Holocene boundary). Media, standard deviation between parentheses, and sample size are given.

Material referido. PVL 5733: MD con M2; PVL 5836: HI con m1- m2; PVL 5837: HI con m1-m3; PVL 5838: HI con incisivo; PVL 5839: MD con M1, M3 y placa cigomática; PVL 5840: HD con incisivo y m1-m2; PVL 5841: HI con incisivo y m1-m2; PVL 5842: HD con m1; PVL 5843: HD con m1; PVL 5844: HD con incisivo y m1; PVL 5845: fragmento de HI con m1; PVL 5846: HD con incisivo; PVL 5847: MD con M1-M2 y placa cigomática.
Descripción. Placa cigomática con borde anterior recto y ángulo superior redondeado. Mandíbula grácil, con proceso coronoides que alcanza mayor altura que el cóndilo, el cual se extiende más atrás que el proceso angular. Proyección capsular situada por debajo de la región anterior de la escotadura sigmoidea o de la parte posterior del proceso coronoides. Cresta masetérica suavemente marcada que alcanza, hacia adelante, el nivel del protofléxido del m1. M1 con flexo anteromediano bien desarrollado, anteroflexo bien marcado que permite distinguir un corto anterolofo y posteroflexo superficial. Protoflexo de M2 vestigial y posteroflexo prácticamente obliterado. El m1 con procíngulo biconulado, cíngulo anterolabial bien desarrollado y posterofléxido penetrante. En m2 se observa un protofléxido y en m3 un hipofléxido transverso y penetrante.
Comentarios taxonómicos. De las especies de Akodon Meyen reconocidas para Argentina, al menos once habitan en la región del noroeste (Pardiñas et al., 2006) y con una taxonomía alfa aún confusa. Siguiendo a Myers et al. (1990) se considera aquí a A. tucumanensis J. A. Allen como subespecie de A. spegazzinii. Mediante estudios en curso que incluyeron series representativas y material topotípico de ambas formas nominales se pudo establecer la ausencia de diferencias morfométricas y morfológicas en caracteres craneanos y dentarios así como genéticas (gen cyt b) entre las mismas, resultados que apoyan la postura taxonómica de Myers et al. (1990). Otra especie asociada con frecuencia a A. spegazzinii ha sido A. alterus Thomas, con localidad tipo en Otro Cerro (Catamarca). Si bien Myers et al. (1990) consideraron que "it seems likely that alterus is properly allied with spegazzinii and tucumanensis" no tomaron una postura taxonómica al respecto sobre este taxón. Autores que trabajaron en la región han tratado a A. alterus como especie válida (Barquez et al., 1991; Mares et al., 1997; Díaz, 1999) o como subespecie de A. spegazzinii (Díaz et al., 2000) y ha sido citada, inclusive, para el valle de Tafí (Blaunstein et al., 1992). Los ejemplares que sustentan esta última afirmación provienen del área de El Infiernillo, a 3000 m de altitud. Sin embargo, especímenes colectados por los autores en la misma zona son indiferenciables en morfología craneana y dentaria, así como en caracteres genéticos, de topotipos de A. spegazzinii. El material fósil estudiado es referido a A. spegazzinii sobre la base de su morfología y su morfometría, que coinciden con las de ejemplares topotípicos.
Distribución y hábitat. Ampliamente distribuida en el noroeste argentino, A. spegazzinii, incluyendo a A. tucumanensis, habita en Tucumán en una variedad de ambientes, desde áreas de bosques a sectores con pastizales. En ambientes áridos y semiáridos como las PF del Monte y Prepuna habita en pastizales asociados a cursos de agua. En la PF de las Yungas se encuentra en todos los pisos altitudinales, desde la selva basal hasta los pastizales de neblina, donde es muy abundante. Su rango altitudinal se extiende desde los 400 m hasta algo más de 3300 m. En el valle de Tafí se conocen numerosos registros, desde los sectores más bajos hasta las cumbres que rodean el valle, a partir de capturas (Myers et al., 1990; Blaustein et al., 1992) y de material recuperado en egagrópilas de lechuzas (Ortiz, 2001; Ortiz y Pardiñas, 2001).
Género Necromys Ameghino, 1889
Especie tipo. Necromys conifer Ameghino, 1889.
Necromys cf. N. lactens (Thomas, 1918) Figuras 3.5 y 4.9; tablas 1 y 2
Material referido. PVL 5734: MD con M1-2; PVL 5852: fragmento de MI con M1-M3.
Descripción. Placa cigomática robusta y foramen incisivo que alcanza el nivel del protocono del M1. Molares superiores anchos, el M1 con procíngulo en forma de abanico. Anterolofo de M2 se extiende casi hasta el margen labial del molar.
Comentarios taxonómicos. Revisiones en curso permiten reconocer para el noroeste de Argentina al menos tres especies del género: N. amoenus (Thomas) (Jayat et al., 2006; Pardiñas et al., 2006), N. lactens (Barquez et al., 1991; Mares et al., 1997; Díaz, 1999; Díaz et al., 2000; Jayat y Pacheco, 2006) y N. lasiurus (Lund) (Jayat et al., 2006; Pardiñas et al., 2006). La longitud de la serie molar y el tamaño de los molares de los especímenes fósiles permitió descartar su pertenencia a N. amoenus. La morfología del material sugiere que podría corresponder a N. lactens, pero el carácter fragmentario de los restos y la ausencia de rasgos diagnósticos impidió su asignación inequívoca. Necromys lactens es, en la actualidad, la única especie registrada en el valle de Tafí (Ortiz y Pardiñas, 2001).
Distribución y hábitat. N. lactens se encuentra principalmente asociada a ambientes de pastizales de neblina y áreas de ecotono con bosques de la PF de las Yungas y ambientes semiáridos de altura, entre 1500 y 3100 m de altitud, en las provincias de Jujuy, Salta, Tucumán y Catamarca (Jayat y Pacheco, 2006). Aparentemente es más abundante en sectores húmedos a menor altitud que en las proximidades del límite altitudinal superior de su distribución. Esta especie ha sido registrada en el valle de Tafí a partir de ejemplares capturados (Jayat y Pacheco, 2006) y de restos recuperados en egagrópilas de lechuzas (Ortiz, 2001; Ortiz y Pardiñas, 2001), entre los 1900 y 2800 m.
Tribu PHYLLOTINI Vorontzov, 1959
Género Calomys Waterhouse, 1837
Especie tipo. Mus bimaculatus Waterhouse, 1837.
Calomys cf. C. musculinus (Thomas, 1913) Figuras 3.4 y 4.4; tablas 1 y 2
Material referido. PVL 5739: HI con m1-m2; PVL 5740: HD con m1-2; PVL 5741: MD con M1-M2; PVL 5742: MD con M1; PVL 5743: MD edéntulo; PVL 5768: fragmento de cráneo con M1-M2 izquierdo; PVL 5769: fragmento de MD con placa cigomática; PVL 5770: fragmento de MI edéntulo con placa cigomática; PVL 5771: HI con incisivo y m1; PVL 5772: HD con incisivo; PVL 5773: HI con incisivo y m1-m2; PVL 5774: HI con incisivo; PVL 5775: MD con M1-M3; PVL 5776: HI edéntula; PVL 5777: MD edéntulo con placa cigomática; PVL 5778: HI con incisivo y m1; PVL 5779: MD con M1-M2 y placa cigomática; PVL 5780: HI con incisivo y m1; PVL 5781: HD con m1; PVL 5782: MD edéntulo con placa cigomática; PVL 5783: HD con incisivo y m1; PVL 5784: fragmento de MD con M1 y placa cigomática; PVL 5785: MI con M1 y placa cigomática; PVL 5786: HI con incisivo; PVL 5787: fragmento de MD con placa cigomática; PVL 5788: fragmento de MD con placa cigomática; PVL 5855: MD edéntulo con placa cigomática; PVL 5856: fragmento de HD con m1-m3; PVL 5857: MD con M1.
Descripción. Bordes de la región supraorbitaria divergentes hacia atrás. Placa cigomática con borde anterior recto o levemente cóncavo y una proyección aguda en su ángulo superior. Forámenes incisivos alcanzan el nivel del protoflexo del M1. Mandíbula pequeña y relativamente alta, con proceso coronoides de mayor altura que el cóndilo. Proyección capsular situada por debajo del sector anterior de la escotadura sigmoidea o por debajo del proceso coronoides. Cresta masetérica proyectada por delante del borde dorsal de la mandíbula, terminando en una protuberancia robusta. Molares de corona crestada. M1 con un estilo anteromediano (sensu Hershkovitz, 1962) en su cara anterior, por debajo del flexo anteromediano. El m1 con fléxido anteromediano vestigial y cíngulo anterolabial bien desarrollado. Protofléxido de m2 penetrante y posterofléxido menos desarrollado que en m1.
Comentarios taxonómicos. La situación taxonómica de Calomys Waterhouse se ha mantenido confusa durante muchos años. Aportes recientes han brindado evidencias sobre la monofilia del género y han considerado la existencia de al menos doce especies (Salazar- Bravo et al., 2001, 2002). Hasta hace pocos años se reconocían para el noroeste argentino la presencia de C. callosus (Rengger), C. laucha (Fischer), C. lepidus (Thomas) y C. musculinus (e.g., Barquez et al., 1991; Ojeda y Mares, 1989), pero Díaz (1999) y Díaz et al. (2000) mencionan siete especies para las provincias de Jujuy y Salta en conjunto. Estudios recientes han considerado a las poblaciones del noroeste argentino referidas previamente a C. callosus como C. fecundus (Thomas) (Salazar-Bravo et al., 2002; Dragoo et al., 2003). Entre las especies de Calomys de pequeño tamaño, Barquez et al. (1991) mencionan para Tucumán a C. laucha para áreas bajas de llanura y a C. lepidus y C. musculinus para áreas de altura. Los restos fósiles son morfológica y morfométricamente indiferenciables de especímenes actuales del valle de Tafí asignados a C. musculinus. Sin embargo, debido al carácter fragmentario de los restos estudiados y a la carencia de rasgos diagnósticos (criterio también discutido por Pardiñas y Lezcano, 1995), referimos el material fósil a Calomys cf. C. musculinus.
Distribución y hábitat. Ejemplares de Calomys musculinus han sido capturados en la mayoría de los ambientes presentes en el noroeste argentino (e.g., Thomas, 1913; Díaz, 1999; Díaz et al., 2000) y es la especie más abundante del género en áreas de pastizales de neblina. Observaciones de campo en este ambiente indican que C. musculinus es especialmente abundante en áreas con escasa cobertura vegetal. En el valle de Tafí se registraron numerosos especímenes a través de capturas y de material proveniente de egagrópilas (Ortiz, 2001; Ortiz y Pardiñas, 2001).
Género Phyllotis Waterhouse, 1837
Especie tipo. Mus darwini Waterhouse, 1837.
Phyllotis osilae J. A. Allen, 1901 Figura 4.6; tablas 1 y 2
Material referido. PVL 5794: MI edéntulo con placa cigomática; PVL 5796: MI edéntulo y placa cigomática; PVL 5800: MI con M1- M2 y placa cigomática; PVL 5825: MD con M1 y placa cigomática; PVL 5827: fragmento de MD con M1-M3; PVL 5832: MI con M1- M2 y placa cigomática.
Descripción. Placa cigomática con borde anterior recto o levemente cóncavo. Forámenes incisivos alcanzan el nivel del procíngulo del M1. Fosetas posteropalatales pequeñas y situadas posteriormente o al mismo nivel del margen anterior de la fosa mesopterigoides, por detrás del borde posterior del alvéolo del M3. M1 sin evidencias de flexo anteromediano, con protoflexo transverso y paraflexo levemente dirigido hacia atrás. M2 sin anteroflexo y con meso e hipoflexo, penetrantes y oblicuos, que determinan una forma de "S" o "Z" para el molar.
Comentarios taxonómicos. Aunque en la región noroeste P. osilae coexiste con otras tres especies del género (P. caprinus Pearson, P. xanthopygus y P. anitae Jayat et al.), puede distinguirse por el tamaño y posición de las fosetas posteropalatales con respecto al borde anterior de la fosa mesopterigoides y por caracteres dentarios que incluyen la morfología y el desarrollo de la hipsodoncia.
Distribución y hábitat. En el noroeste argentino P. osilae es una especie restringida principalmente a pastizales de neblina y áreas ecotonales desde la provincia de Jujuy hasta el sur de Catamarca (Jayat y Pacheco, 2006). Ha sido capturada en bosques de aliso situados por encima de los 1500 m hasta áreas de transición con ambientes semiáridos de altura apenas por encima de los 3000 m. Es la especie de Phyllotis más común en pastizales de neblina, coexistiendo con P. xanthopygus en zonas ecotonales de gran altitud y con P. anitae en zonas más bajas linderas con el bosque (Jayat y Pacheco, 2006; Jayat et al., 2006, 2007). Existen numerosos registros de capturas y restos provenientes de egagrópilas de rapaces en el valle de Tafí, desde zonas bajas del valle hasta localidades ubicadas a 3300 m (Mares, 1977; Ortiz y Pardiñas, 2001; Jayat y Pacheco, 2006).
Phyllotis sp. Figuras 3.6, 3.7 y 4.5; tablas 1 y 2
Material referido. PVL 5744: HI con m1; PVL 5745: HI edéntula; PVL 5746: MD con M1; PVL 5747: HD edéntula; PVL 5748: MI con M1-M2: PVL 5749: MI con M1-M3; PVL 5750: fragmento de MD con fracción de M1; PVL 5751: m1 aislado; PVL 5789: HD con incisivo; PVL 5790: MI con M1-M3 y placa cigomática; PVL 5791: HI con incisivo y m1; PVL 5792: HI con incisivo y m1-m3; PVL 5793: HD con m1; PVL 5795: HD con incisivo y m1-m3; PVL 5797: HI con incisivo y m1; PVL 5798: HD con m1-m3; PVL 5799: HD con incisivo y m1; PVL 5801: HI con incisivo y m1-m2; PVL 5802: HD edéntula; PVL 5803: HD con incisivo y m1-m2; PVL 5804: MD con M1-M2 y placa cigomática; PVL 5805: HD con m1-m2; PVL 5806: HI edéntula; PVL 5807: MI con M1 y placa cigomática; PVL 5808: MI con M1-M3 y placa cigomática; PVL 5809: fragmento de HD con incisivo; PVL 5810: premaxilar con incisivo izquierdo; PVL 5811: fragmento de MI edéntulo y con placa cigomática; PVL 5812: fragmento de HD con m1; PVL 5813: fragmento de HI con incisivo y m1-m2; PVL 5814: HD con incisivo; PVL 5815: placa cigomática izquierda; PVL 5816: placa cigomática derecha; PVL 5817: placa cigomática derecha con M1; PVL 5818: fragmento de HD edéntu- la; PVL 5819: fragmento de MI con M1 y fragmento de placa cigomática; PVL 5820: HI con incisivo; PVL 5821: fragmento de MD con M1-M2; PVL 5822: HI con m1-m2; PVL 5823: MD con M1-M3 y placa cigomática; PVL 5824: MI con M1, M3 y placa cigomática; PVL 5826: MD con M1-M2 y placa cigomática; PVL 5828: HD con incisivo; PVL 5829: HD edéntula; PVL 5830: fragmento de MD con M1 y placa cigomática; PVL 5831: fragmento de MD con M3; PVL 5833: M1 derecho y fragmento de placa cigomática; PVL 5834: fragmento de MD edéntulo y placa cigomática; PVL 5835: fragmento de MI con M1; PVL 5858: HI con m1-m2; PVL 5859: HI edéntula.
Descripción. Placa cigomática con borde anterior recto o levemente cóncavo. Forámenes incisivos extendidos por detrás del nivel del protoflexo del M1. Mandíbula con proceso coronoides bien desarrollado que alcanza mayor altura que el cóndilo, el cual se extiende posteriormente más atrás que el proceso angular. Cresta masetérica supera anteriormente el nivel del margen anterior del m1. M1 de dos ejemplares juveniles con flexo anteromediano vestigial y una muesca indicadora del protoflexo. M2 de uno de estos ejemplares con para-anteroflexo bien desarrollado. M3 de un individuo adulto joven con foseta central de gran tamaño correspondiente al metaflexo. El m1 en ejemplares juveniles con remanentes de fléxido anteromediano y de posterofléxido. Cíngulo anterolabial bien desarrollado. Metafléxido e hipofléxido oblicuos y penetrantes determinando un m2 con forma de "S". Hipofléxido de m3 penetra hasta la línea media en forma perpendicular al eje del molar. Entofléxido menos penetrante y dirigido hacia adelante.
Comentarios taxonómicos. Rasgos de la dentición de los ejemplares fósiles, incluyendo una hipsodoncia moderada en los molares, permiten descartar su pertenencia a P. anitae. Si bien es probable que parte de este material corresponda a P. osilae, la ausencia de rasgos diagnósticos en los restos estudiados impide descartar su pertenencia a P. xanthopygus y obliga a mantener una hipótesis taxonómica abierta.
Distribución y hábitat. Otra especie de amplia distribución en el noroeste argentino es P. xanthopygus, particularmente en ambientes de las PF de la Puna, Prepuna y Monte (Mares, 1977; Steppan, 1998; Kramer et al., 1999). En áreas donde coexiste con P. osilae se encuentra limitada a zonas más xéricas y de mayor altitud. En el valle de Tafí ha sido capturada en zonas rocosas sólo en los sectores más altos a partir de los 3000 m. Muestreos intensivos durante casi una década en ambientes de pastizales de neblina sugieren que P. anitae se encuentra restringida a los bosques de A. acuminata (Jayat et al., 2007).
Tribu REITHRODONTINI Vorontzov, 1959
Género Reithrodon Waterhouse, 1837
Especie tipo. Reithrodon typicus Waterhouse, 1837.
Reithrodon auritus (Fischer, 1814) Figuras 3.1, 3.2, 4.7 y 4.8; tablas 1 y 2
Material referido. PVL 5737: HI edéntula; PVL 5738: incisivo superior izquierdo; PVL 5752: HI con m1-m2; PVL 5753: premaxilar y MI con incisivo, serie molar completa y placa cigomática; PVL 5754: premaxilar y MD con incisivo, serie molar completa y placa cigomática; PVL 5755: HD con m1-m3; PVL 5756: fragmento anterior de cráneo con premaxilar y maxilar y ambas series molares completas; PVL 5757: HD con m1-m2; PVL 5758: HD con m1-m3; PVL 5759: HI con m2 y m3; PVL 5760: fragmento de HI edéntula; PVL 5761: fragmento de premaxilar izquierdo con incisivo; PVL 5762: fragmento de placa cigomática derecha; PVL 5860: HI con m1-m3; PVL 5861: MD con M1, M3 y placa cigomática; PVL 5862: MI con M2-M3 y placa cigomática.
Descripción. Sutura premaxilo-maxilar dispuesta en ángulo en vista lateral. Borde anterior de placa cigomática profundamente excavado y con un proceso a manera de espina en su extremo anterodorsal. Forámenes incisivos alcanzan el nivel del protocono del M1. Incisivos superiores con un surco medial poco desarrollado y otro conspicuo hacia el borde labial (véase Steppan, 1995). Mandíbula con proceso coronoides muy corto situado por debajo del nivel del cóndilo. Cresta masetérica baja con extremo anterior que alcanza el foramen mentoniano. Molares hipsodontes, los superiores con flexos penetrantes que superan la línea media. M1 con patrón trilofodonte y sin evidencias de posteroflexo. M2 con procíngulo bien desarrollado y para-anteroflexo muy penetrante, rasgos que dan al molar un patrón trilofodonte. M3 con hipoflexo corto y fondo del metaflexo expandido y dirigido hacia atrás. Procíngulo de m1 de forma romboidal. Posterolófido bien desarrollado que alcanza el margen lingual. El m2 con ento- y posterofléxido penetrantes y posterolófido bien desarrollado. Hipofléxido de m3 recto y más penetrante que en m1 y m2.
Comentarios taxonómicos. Se considera actualmente que la especie presente en el noroeste argentino corresponde a Reithrodon auritus (Pardiñas y Galliari, 2001). No obstante, Thomas (1920) describió para Otro Cerro, Catamarca, otra taxón nominal, R. caurinus Thomas, actualmente referida como subespecie de R. auritus (Pardiñas y Galliari, 2001). La relación taxonómica de R. caurinus con las poblaciones pampeanas y patagónicas de R. auritus aún no ha sido estudiada.
Distribución y hábitat. Reithrodon auritus presenta una amplia distribución en Argentina, desde Tierra del Fuego hasta el noroeste, región en donde está restringido a unas pocas localidades aisladas situadas a más de 2000 m de altitud (Pardiñas y Galliari, 2001; Jayat et al., 2006). Los escasos registros en el noroeste provienen de sectores pertenecientes a la PF de la Puna y del límite altitudinal superior de los pastizales de neblina (Dalby y Mares, 1974; Pardiñas y Galliari, 2001; Jayat et al., 2006). En las localidades correspondientes a este último ambiente (El Infiernillo y Otro Cerro) sólo fue capturada en los sectores más altos, a 3000 m, donde las comunidades vegetales muestran una composición de especies empobrecida en relación con los pastizales situados a menor altitud. En El Infiernillo existen áreas con densa cobertura vegetal junto a sectores con suelo parcialmente desnudo y sustrato rocoso. En Patagonia estos ambientes de escasa cobertura han sido mencionados como especialmente propicios para la presencia de esta especie (Pearson, 1988; Monjeau, 1989). Cabe destacar que R. auritus no ha sido registrado en ninguna de las muestras de egagrópilas recuperadas en el área baja y media del valle de Tafí.
Sigmodontinae incertae sedis
Género Andinomys Thomas, 1902
Especie tipo. Andinomys edax Thomas, 1902.
Andinomys edax Thomas, 1902 Figuras 3.3 y 4.3; tablas 1 y 2
Material referido. PVL 5735: HD con m1; PVL 5763: HI con incisivo y m1; PVL 5764: HD con m1; PVL 5765: fragmento de HI edéntulo; PVL 5766: fragmento de HI edéntulo; PVL 5767: frontal aislado.
Descripción. Mandíbula robusta, con proceso coronoides alcanzando mayor altura que el cóndilo. Extremo anterior de cresta masetérica alcanza el nivel del protocónido del m1. Este molar presenta corona plana, con fléxido anteromediano y posterofléxido penetrantes.
Comentarios taxonómicos. Andinomys edax pertenece a un género monotípico para el cual se han reconocido dos subespecies A. edax edax y A. edax lineicaudatus (Yepes, 1935). Díaz (1999) trató a ambas formas como especies diferentes en virtud de algunas diferencias morfológicas y por su ocurrencia en simpatría en áreas de la PF de la Prepuna en Jujuy. Sin embargo, Díaz y Barquez (1999) mantuvieron el rango subespecífico de lineicaudatus dado originalmente por Yepes (1935). Hasta tanto se lleven a cabo estudios detallados de series representativas de ejemplares de distintas poblaciones, se sigue aquí el criterio de D´Elía et al. (2006) de reconocer una única especie.
Distribución y hábitat. Andinomys edax presenta una amplia distribución en la región de los Andes centrales. Aunque la mayoría de los registros provienen de altitudes por encima de los 3000 m (e.g., Anderson, 1997), en el noroeste argentino se ha citado desde aproximadamente 600 m de altitud en la PF de las Yungas en Tucumán (Capllonch et al., 1997) hasta más de 4500 m en ambientes de la PF de la Puna en Jujuy (Díaz, 1999). Esta notable plasticidad ecológica es sugerida también por el registro fósil de la especie (Ortiz y Jayat, 2007). En el valle de Tafí ha sido capturado en pastizales de neblina, en muchos casos asociados a rocas, en las zonas altas del valle (Barquez et al., 1991; Ortiz y Jayat, obs. pers.), y se han recuperado restos contenidos en egagrópilas en zonas próximas a los bosques, a menor altitud (Ortiz y Pardiñas, 2001).
Discusión y conclusiones
Aspectos tafonómicos
Las aves rapaces del Orden Strigiformes constituyen los contribuyentes más importantes en la formación de agregados fósiles de microvertebrados (e.g., Andrews, 1990; Fernández Jalvo, 1996; Stahl 1996; Pardiñas, 1999, 2000). El análisis integral del agregado fósil estudiado sugiere que el mismo se generó a partir de la acumulación de egagrópilas de lechuzas. El excelente estado de preservación de los restos ha permitido descartar a mamíferos carnívoros y aves falconiformes como los generadores del agregado, depredadores que fragmentan y corroen severamente los huesos (Andrews, 1990). La evidencia más confiable sobre el origen de la muestra corresponde a la presencia de marcas de corrosión en elementos óseos y dentarios debido a la digestión por los ácidos estomacales de las lechuzas. Por su alto grado de especificidad y persistencia a través de la sucesión de eventos tafonómicos se ha reconocido a estas marcas como las mejores señales para identificar el origen de un agregado de microvertebrados (e.g., Andrews, 1990; Fernández-Jalvo y Andrews, 1992; Denys et al., 1997). El escaso grado de digestión y la proporción de piezas afectadas señalan, una vez más, a las lechuzas como generadores de la muestra (Andrews, 1990).
Tres especies de estrigiformes que habitan en el valle de Tafí pueden señalarse como posibles responsables del agregado -Tyto alba, Asio flammeus y Bubo virginianus (Gmelin)- ya que incluyen en su dieta roedores sigmodontinos de tamaño mediano a relativamente grande, como los registrados en la muestra fósil (Ortiz et al., 2000a; Cirignoli et al., 2001; Ortiz y Pardiñas, 2001). No obstante, el porcentaje y el grado de digestión en los incisivos examinados no han permitido identificar inequívocamente a la especie responsable del yacimiento. Esta misma fuente de evidencia permite descartar a Athene cunicularia, con su dieta compuesta principalmente por insectos, anfibios y roedores sigmodontinos pequeños (Alvarez, 1992).
Reconstrucción paleoambiental
Contribuciones relativamente recientes han brindado valiosa información sobre las características ambientales durante el Pleistoceno tardío y el Holoceno en el centro y norte de Argentina. A partir de diversas fuentes de evidencias, que incluyen estudios geomorfológicos, sedimentológicos y paleopedológicos, numerosos autores coinciden en destacar condiciones más frías y secas que las actuales en torno al límite Pleistoceno-Holoceno (desde 18 ka hasta aproximadamente 9 ka AP). Iriondo (1993) e Iriondo y García (1993) indican estas condiciones para las tierras bajas del chaco occidental y todo el flanco oriental de las sierras subandinas. Estudios adicionales más puntuales realizados en los alrededores de San Salvador de Jujuy, Jujuy (Alcalde y Kulemeyer, 1999), laguna de Mar Chiquita, Córdoba (Kröhling e Iriondo, 1999) y localidades adicionales en esa provincia (Carignano, 1999) apoyan estas conclusiones. Iriondo (1993) sostiene que, como consecuencia de estas condiciones más frías y secas, los pisos de vegetación se ubicaron a niveles altitudinales inferiores a los del presente. En contraste, otras contribuciones han destacado condiciones de mayor humedad que las actuales para el fin del Pleistoceno y principios del Holoceno (Markgraf, 1985, 1989; Lupo, 1998; Zipprich et al., 1999). Sin embargo, es necesario destacar que estos estudios, basados sobre evidencias geomorfológicas y palinológicas abarcando el lapso 10,5-7,5 ka A.P, han estado circunscriptos a la región del altiplano de la provincia de Jujuy y sectores próximos en la provincia de Salta. Estas áreas de gran altura, junto con el altiplano y la puna de Atacama en Chile, han experimentado condiciones más húmedas que las actuales durante el período considerado (e.g., Messerli et al., 1993; Geyh y Grosjean, 1999).
El análisis exhaustivo de los rasgos que imponen los depredadores a los agregados fosilíferos es un requisito esencial para la realización de interpretaciones paleoambientales. La determinación del tamaño del área de caza del predador permite inferir los posibles parches de forrajeo así como las distancias aproximadas de procedencia de los individuos de la muestra en relación con el yacimiento. La consideración de estas características es importante para descartar posibles artefactos generados por la depredación en diferentes pisos altitudinales de vegetación (Ortiz y Pardiñas, 2001). Poblaciones europeas de T. alba presentan un territorio de caza que varía desde menos de 1 hasta 3 km y aquellas de A. flammeus un máximo de alrededor de 1,5 km (Andrews, 1990). Estudios realizados en poblaciones de Bubo bubo Linnaeus de Suecia demostraron que esta especie presenta un área de acción de mayor tamaño, con un máximo aproximado de 5 km de radio (Andrews, 1990). Un área de este tamaño, centrada en el yacimiento, no excede los límites del fondo del valle, en un relieve relativamente homogéneo que no supera los 250 m de rango altitudinal.
El hallazgo en la misma área del valle de Tafí de un yacimiento del Pleistoceno medio-superior y de otro cercano al límite Pleistoceno-Holoceno, sumado al conocimiento de la fauna actual del lugar, resulta de particular interés para el estudio de la evolución de las comunidades de micromamíferos de la región y los ambientes en que se desarrollaron. A partir del análisis del ensamble de La Angostura, más diverso que el de Tafí del Valle y sin análogo moderno (agregado no análogo sensu Barnosky, 1993), Ortiz (2001) y Ortiz y Pardiñas (2001) sugirieron condiciones climático- ambientales diferentes a las actuales para el área durante el Pleistoceno tardío. Sin embargo, las nuevas dataciones impiden ahora referir los sedimentos de la Formación Tafí del Valle al Pleistoceno más superior (Kemp et al., 2003, 2004; Schellenberger et al., 2003; Carter-Stiglitz et al., 2006), por lo que, ante la ausencia de dataciones directas sobre el material óseo de la muestra, el ensamble de La Angostura debe ser referido al intervalo Pleistoceno medio-superior. De acuerdo a Ortiz (2001), esta asociación habría sido el resultado de una mayor heterogeneidad ambiental como consecuencia de una menor amplitud estacional, de un modo similar al planteado para comunidades de micromamíferos de América del Norte y otros continentes (Graham, 1985; Graham y Lundelius, 1984; FAUNMAP Working Group, 1996), y a la contracción de los pisos altitudinales de vegetación. Asimismo, Ortiz (2001) y Ortiz y Pardiñas (2001) plantearon el carácter contemporáneo del agregado de La Angostura a partir de una baja promediación temporal durante el Pleistoceno más tardío. Sin embargo, la nueva cronología disponible para la Formación Tafí del Valle permite considerar que la muestra estudiada por Ortiz (2001) y Ortiz y Pardiñas (2001) se generó durante un intervalo temporal más amplio que el planteado originalmente, dando cuenta así de su mayor diversidad con relación a la muestra puntual del límite Pleistoceno-Holoceno. El ensamble de La Angostura cuenta con un MNI casi seis veces mayor que el agregado de Tafí del Valle y cuatro veces mayor que la muestra actual (tabla 1).
En términos generales, las evidencias indican que la composición taxonómica actual de la comunidad de roedores sigmodontinos de las áreas baja y media del valle de Tafí estaba ya establecida desde el límite Pleistoceno-Holoceno y persistió, con ligeras modificaciones, hasta el presente. Lo más destacable es un empobrecimiento en el elenco de especies a partir del fin del Pleistoceno con la extinción de Tafimys powelli, conocida para el Pleistoceno medio-superior de la Formación Tafí del Valle (Ortiz et al., 2000b) y el Pleistoceno superior de Las Juntas, Catamarca; la presencia de la asociación dominante Phyllotis + Calomys ya en el límite Pleistoceno-Holoceno, lo que indica un primer establecimiento del ensamble actual; la ausencia de Neotomys ebriosus, sin registro moderno para Tucumán (Pardiñas y Ortiz, 2001); y la actual retracción de Reithrodon auritus a las áreas más altas del valle, aunque presente hasta el Holoceno temprano (Ortiz y Pardiñas, 2001).
En comparación con la fauna actual de roedores sigmodontinos conocida para el área de baja y media altitud del valle de Tafí, se puede considerar a la muestra del límite Pleistoceno-Holoceno como de una baja riqueza específica. En el agregado fósil se encuentran representadas las especies más comunes para los ambientes de pastizales de neblina conocidas para esta latitud, que incluyen a Akodon spegazzinii, Phyllotis osilae y Calomys cf. C. musculinus como las dominantes. Sin embargo, faltan las especies de Oligoryzomys, Abrothrix illuteus y Oxymycterus cf. O. paramensis, registradas tanto para el Pleistoceno medio- superior como para el presente en el mismo valle (Ortiz y Pardiñas, 2001; Ortiz y Jayat, datos no publicados). En principio, la baja riqueza específica puede ser explicada por el tamaño de muestra relativamente reducido (MNI= 51). No obstante, al contrastar el conjunto fósil con una muestra actual de la misma localidad (MNI= 75) se observan importantes diferencias en la composición taxonómica cualitativa y cuantitativa entre ambas, si bien la riqueza específica es la misma (figura 5; tabla 1). Estas diferencias sugieren para el límite Pleistoceno-Holoceno condiciones paleoambientales diferentes a las actuales, de modo tal que las especies presa disponibles para las rapaces eran distintas en cada época en esa área del valle. Esta interpretación recibe sustento adicional al comparar la muestra actual de Tafí del Valle con una muestra de T. alba de La Angostura (MNI= 114) (Ortiz y Pardiñas, 2001; figura 5; tabla 1), localidad actualmente situada en áreas de pastizales pero muy próxima a zonas de bosque montano (figura 1). La muestra de La Angostura se diferencia de la de Tafí del Valle por la adición de especies que son más frecuentes en ambientes con cobertura arbórea (Abrothrix illuteus, Oxymycterus cf. O. paramensis y Oligoryzomys spp.).
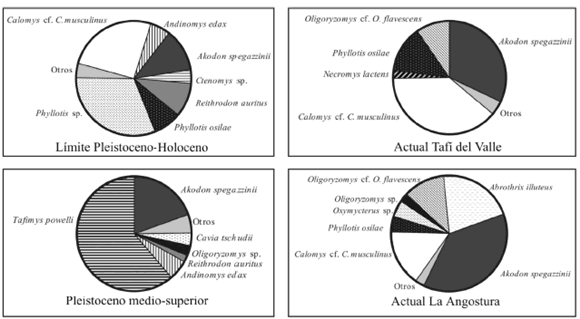
Figura 5. Comparación de la abundancia relativa (sólo para aquellos taxones con un MNI > 2%) en las muestras fósiles y actuales estudiadas / comparison of the relative abundance (only for taxa with MNI > 2%) in the studied fossil and modern samples.
Phyllotis osilae es una especie asociada a ambientes de pastizales relativamente húmedos aunque también ha sido registrada en zonas de mayor altitud en transición con ambientes más xéricos. Parte de los especímenes asignados a Phyllotis sp. pueden corresponder a P. xanthopygus, especie asociada a ambientes de mayor aridez y que coexiste con P. osilae en la zona de El Infiernillo (Mares, 1977) y otras áreas ambientalmente similares en el noroeste argentino (P. Jayat, obs. pers.). Necromys lactens es típica de los pastizales de neblina con alto grado de humedad por encima de los 1500 m (Ortiz y Pardiñas, 2001; Jayat y Pacheco, 2006) pero también ha sido registrada en áreas más secas, aunque en densidades mucho más bajas (P. Jayat, obs. pers.). Un claro indicador de áreas de pastizales con escasa cobertura vegetal del suelo es la presencia de Calomys cf. C. musculinus con una frecuencia relativamente alta (25%). En la actualidad esta especie alcanza valores de frecuencia elevados, tanto en Tafí del Valle como en La Angostura, particularmente en egagrópilas de A. cunicularia de la última localidad. Estos altos porcentajes pueden explicarse por factores relacionados con el incremento de superficies cultivadas y el sobrepastoreo, así como por el comportamiento trófico oportunista de estas lechuzas. Estos fenómenos fueron mencionados para explicar el aumento en la frecuencia de Calomys laucha y C. musculinus en agroecosistemas de las regiones pampeana y norpatagónica (Bellocq, 1990; Pardiñas, 1999; Pardiñas et al., 2000). Muestreos realizados en los últimos ocho años en áreas de pastizales de neblina en el noroeste argentino demuestran una neta preferencia de C. musculinus por microambientes con escasa cobertura vegetal (P. Jayat, obs. pers.). La presencia de Reithrodon auritus en el ensamble fósil, registrada en el noroeste en pastizales de áreas xéricas con escasa cobertura vegetal, sugiere condiciones paleoambientales algo más secas para el área baja del valle. Cabe destacar que entre las muestras de egagrópilas modernas de Tafí del Valle y La Angostura, con más de 230 individuos registrados, no se recuperaron especímenes de esta especie. La ausencia en la muestra del límite Pleistoceno-Holoceno de especies comunes en bosques montanos como Abrothrix illuteus, Oxymycterus paramensis y de ambientes húmedos como Oligoryzomys spp., refuerzan la interpretación de un paleoambiente algo más xérico.
La evidencia más concluyente sobre las condiciones paleoambientales reinantes durante la formación del agregado es la existencia en la actualidad de un área de simpatría de todas las especies registradas en la muestra fósil, en los sectores más elevados del valle a 3000 m de altitud. En esta área de simpatría el ambiente está caracterizado por comunidades empobrecidas de pastizales en transición con zonas de suelo desnudo y ambientes semiáridos altoandinos (Vervoorst, 1982), con temperaturas notablemente más frías que en los sectores bajos del valle y nevadas ocasionales durante el invierno casi todos los años.
En este trabajo se aportan evidencias concretas de que los rasgos generales de la composición taxonómica actual de la comunidad de roedores sigmodontinos de los pastizales de neblina de la provincia de Tucumán estaban ya establecidos, al menos, desde el final del Pleistoceno. No obstante, las frecuencias relativas de los taxones implicados parecen ser el producto de las condiciones ambientales actuales bajo la influencia del impacto antrópico durante los últimos 350 años, con el inicio de las actividades agropecuarias intensivas en el valle (Adris y Chocobar, 1999). Estos factores habrían provocado el aumento en la frecuencia de las especies de roedores oportunistas y una pérdida generalizada de la diversidad. En este sentido, la presencia en la actualidad en el valle de Calomys fecundus (Thomas), una especie frecuente en ambientes de ecotono Chaco-Yungas a baja altitud, constituye un posible ejemplo de esta tendencia. Prospecciones adicionales de restos de micromamíferos en sedimentos del Pleistoceno superior y Holoceno permitirán contrastar estas hipótesis y clarificar aspectos de la evolución de las comunidades de sigmodontinos en esta región.
Apéndice. Listado de especímenes actuales examinados y empleados como material comparativo / list of modern specimens used as comparative material.
Abrothrix illuteus Thomas, 1925. CATAMARCA: Las Chacritas, aproximadamente 28 km al NNO de Singuil, sobre Ruta Provincial Nº 1 (JPJ 520, 549). TUCUMÁN: aproximadamente 10 km al S de Hualinchay, sobre el camino a Lara, 2300 m (JPJ 481, 501, 763, 1220); El Mollar, valle de Tafí (PEO-E 8); La Angostura, Tafí del Valle (PEO-E 25); Las Agüitas, cumbre del Taficillo (CML 4020, 4021, 4036); quebrada de los Sosa (MACN 17458); Tafí Viejo, cerros occidentales (CML 297, 301); Zanjón de Tafí, 2 km al sudoeste de Tafí del Valle (PEO-E 178).
Akodon lutescens J. Allen, 1901. JUJUY: León, Arroyo La Tablada (MMPMa 1241, 1243, 1254). SALTA: Cañada Ojo de Agua, 10 km al sur de Rosario de la Frontera (PEO-E 80); TUCUMÁN: aproximadamente 10 km al S de Hualinchay, sobre el camino a Lara, 2300 m (JPJ 492); Hualinchay, sobre el camino a Cafayate, 1861 m (JPJ 172, 179-181).
Akodon simulator Thomas, 1916. CATAMARCA: aproximadamente 1 km al sudeste de El Espinillo, Campo del Pucará (Las Estancias), extremo sur de la Sierra del Aconquija, 1950 m (PEO-E 121). TUCUMÁN: Villa Nougués, aproximadamente 2 km al N, sobre Ruta Provincial Nº 338, a la altura de la antena repetidora del Canal 10 de televisión (JPJ 262, 265, 266).
Akodon spegazzinii spegazzinii Thomas, 1897. SALTA: Cachi Adentro, sobre río Cachi, 2490 m (JPJ 1206, 1209); aproximadamente 2 km al NNE de Cachi Adentro, sobre el camino a Las Pailas, 2567 m (JPJ 1194, 1199, 1203, 1204, 1205)
Akodon spegazzinii tucumanensis J.A. Allen, 1901. TUCUMÁN: 7 km al NO de El Infiernillo, por camino al electroducto a La Alumbrera (PEO 4); cerro Muñoz, arriba del Puesto de Ibáñez, subiendo por Santa Cruz, 3300 m (PEO 54, 55, 56, 57); Hualinchay, sobre el camino a Cafayate, 1861m (JPJ 157, 160, 162); La Angostura, 1 km al Norte (PEO 1); La Florida (Reserva), Pueblo Viejo (CML 6090 ); Reserva la Florida, 7 km al O. de Ibatín, sobre Río Pueblo Viejo (CML 5007, 5009, 5010); Reserva Provincial Santa Ana, sobre río El Saltón (CML 6070, 6072, 6074, 6081); Villa Nougues, aproximadamente 2 km al N, sobre Ruta Provincial Nº 338, a la altura de la antena repetidora del Canal 10 de televisión (JPJ 257, 261, 263); Zanjón de Tafí, 2 km al sudoeste de Tafí del Valle (PEO-E 179).
Necromys lactens (Thomas, 1918). JUJUY: Bárcena, aproximadamente 3 km al S, sobre Ruta Nacional Nº 9, 1808 m (JPJ 630, 633, 647, 670, 672); cerro Hermoso (MACN 19.517); El Duraznillo, cerro Calilegua, 2600 m (CML 1736, 1737); León (MMPMa 2158, 2161, 2169, 2185, 2190); Termas de Reyes, aproximadamente 15 km al N, sobre Ruta Provincial Nº 4 (JPJ 123, 124). TUCUMÁN: aproximadamente 10 km al S de Hualinchay, sobre el camino a Lara, 2300 m (JPJ 447); El Rincón, al oeste de la Loma Pelada, valle de Tafí (PEOE 162); Hualinchay, sobre el camino a Cafayate (JPJ 154, 195, 203); La Angostura, Tafí del Valle (PEO-E 27); Ruta 307, km 72 (MMPMa 2985); Tafí del Valle, 2800 m (CML 2062); Tafí del Valle, km 78, 2700 m (CML 1836); Zanjón de Tafí, 2 km al sudoeste de Tafí del Valle (PEO-E 180).
Oligoryzomys cf. O. flavescens (Waterhouse, 1837). CATAMARCA: aproximadamente 10 km al E de Los Varela, sobre Ruta Provincial Nº 4, Sierra de Humaya, 2006 m (JPJ 1306); aproximadamente 2 km al SE de Huaico Hondo, sobre Ruta Provincial Nº 42, al E del Portezuelo, 1992 m (JPJ 424, 802); Las Chacritas, aproximadamente 28 km al NNW de Singuil, sobre Ruta Provincial Nº 1, 1888 m (JPJ 525, 541); unión entre las rutas provinciales Nº 9 y 18, 3,4 km al S, sobre Ruta Provincial Nº 18, 1529 m (JPJ 9, 11, 13). TUCUMÁN: aproximadamente 10 km al S de Hualinchay, sobre el camino a Lara, 2300 m (JPJ 987, 997); Cumbre del Taficillo, al NE de Las Agüitas, Parque Biológico Sierra de San Javier (JPJ 120); El Mollar, valle de Tafí (PEO-E 11); Hualinchay, sobre el camino a Cafayate, 1861 (JPJ 247); Villa Nougues, aproximadamente 2 km al N, sobre Ruta Provincial Nº 338, a la altura de la antena repetidora del Canal 10 de televisión (JPJ 256); Zanjón de Tafí, 2 km al sudoeste de Tafí del Valle (PEO-E 181).
Andinomys edax Thomas, 1902. TUCUMÁN: Aconquija, 3000 m (MACN 29.250-29.253, 30.72, 33.178); Camino a Amaicha, km 83 (CML 670); Carapunco, km 81 de la Ruta Provincial N° 307, 2960 m (JPJ 1064); Castillo de las Brujas, 15 km al sudeste de Amaicha del Valle (PEO-E 50); cerro Muñoz, arriba del Puesto de Ibáñez, subiendo por Santa Cruz, 3300 m (PEO 37); cerro San Javier, a 2000 m (MACN 26.147, 26.148); El Infiernillo (MACN 17.566); Hualinchay, sobre el camino a Cafayate, 1861 m (JPJ 254); Infiernillo, 3000 m (CML 688, 689); sierra de Tafí Viejo, Tucumán, 2000 m (MACN 33.85); Zanjón de Tafí, 2 km al sudoeste de Tafí del Valle (PEO-E 182).
Calomys musculinus (Thomas, 1913). JUJUY: Bárcena, aproximadamente 3 km al S, sobre Ruta Nacional Nº 9, 1808 m (JPJ 131, 667). TUCUMÁN: Carapunco, km 81 de la Ruta Provincial N° 307, 2960 m (JPJ 1070, 1075).
Phyllotis osilae J.A. Allen, 1901. CATAMARCA: Unión entre las rutas provinciales Nº 9 y 18, 3,4 km al S, sobre Ruta Provincial Nº 18, 1529 m (JPJ 19, 24, 31, 1137, 1149). TUCUMÁN: 1 km al N de El Rincón, 26° 57´ S 65° 45' O, 2450 m (PEO 2, 3); cerro Muñoz, arriba del Puesto de Ibáñez, subiendo por Santa Cruz, 3300 m (PEO 49, 50, 51, 52, 53); Zanjón de Tafí, 2 km al sudoeste de Tafí del Valle (PEO-E 184).
Phyllotis xanthopygus (Waterhouse, 1837). CATAMARCA: Agua del Gauchi, aproximadamente 1 km al E del Mogote Los Cocos, 2024 m (JPJ 1286); Chumbicha, 1 km NO por el camino del balneario, 2600 pies (CML 3451). JUJUY: 4 km al O de Rinconada, 3860 m (PEO 11, 13); Abra Pampa (CML 1276, 1277, 1278, 1283); La Ciénaga, Abra Pampa (CML 1280); Laguna de Leandro (PEO 14). SAN JUAN: Estancia Leoncito, 2 km E observatorio Astronómico (CML 3624); San Guillermo (CML 1080).
Reithrodon auritus (Fisher, 1814). CATAMARCA: aproximadamente 1 km al sudeste de El Espinillo, Campo del Pucará (Las Estancias), extremo sur de la sierra del Aconquija, 1950 m (PEO-E 127).
Agradecimientos
Los autores desean agradecer a J. Babot, J. Aguilar, J. Powell, D. Miotti y G. Ortiz Tempel por la asistencia en las tareas de campo. H. Grau y D. García López colaboraron con la obtención de las fotografías. E. Pieroni y S. Moyano prestaron su ayuda en la confección del perfil estratigráfico. Este trabajo fue parcialmente solventado con fondos del Consejo de Investigaciones de la Universidad Nacional de Tucumán (subsidio a J. Powell), del Consejo Nacional de Investigaciones Científicas y Técnicas y de la Agencia Nacional de Promoción Científica y Tecnológica (subsidios a P. Ortiz). Agradecemos a los revisores U. F. J. Pardiñas y E. Tonni por sus comentarios y sugerencias que mejoraron sensiblemente la primera versión del manuscrito.
Bibliografía
1. Adris, S.I y Chocobar, M.F. 1999. El valle de Tafí y su inserción en la economía regional (fines del XVII - Siglo XVIII). IV Jornadas de Comunicaciones, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán (San Miguel de Tucumán, 1999), Resúmenes: 35. [ Links ]
2. Alcalde, J.A.y Kulemeyer, J.J. 1999. The Holocene in the South- Eastern region of the Province Jujuy, North-West Argentina. Quaternary International 57/58: 113-116. [ Links ]
3. Allen, J.A. 1901. New South American Muridae and a new Metachirus. Bulletin of the American Museum of Natural History, 14: 405-412. [ Links ]
4. Alvarez, M.E. 1992. [Sobre la alimentación de la lechucita de las vizcacheras (Athene cunicularia). Seminario de Licenciatura, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán, 53 pp. Inédito.]. [ Links ]
5. Ameghino, F. 1889. Contribución al conocimiento de los mamíferos fósiles de la República Argentina. Actas de la Academia Nacional de Ciencias en Córdoba, 6: 1-1027 y Atlas: 98 pliegos. [ Links ]
6. Anderson, S. 1997. Mammals of Bolivia, taxonomy and distribution. Bulletin of the American Museum of Natural History 231: 1- 652. [ Links ]
7. Andrews, P. 1990. Owls, caves and fossils. Predation, preservation, and accumulation of small mammal bones in caves, with an analysis of the a Pleistocene cave faunas from Westbury-Sub-Mendip, Somerset, UK. University of Chicago Press. Chicago, 231 pp. [ Links ]
8. Barnosky, A.D. 1993. Faunal response to glacial-interglacial transitions in the western United States. VI International Theriological Congress (Sydney), Abstracts 1: 14. [ Links ]
9. Barquez, R.M., Mares, M.A. y Ojeda, R.A. 1991. Mamíferos de Tucumán - Mammals of Tucumán. Oklahoma Museum of Natural History. Norman, 282 pp. [ Links ]
10. Barquez, R.M, Díaz, M.M. y Ojeda, R.A. 2006. Mamíferos de Argentina: sistemática y distribución. Sociedad Argentina para el Estudio de los Mamíferos. Tucumán, 356 pp. [ Links ]
11. Bell, D.A. 1991. Distribución del bosque de aliso del cerro, Alnus acuminata (Betulaceae) en la provincia de Tucumán, Argentina. Boletín de la Sociedad Argentina de Botánica 27: 21-30. [ Links ]
12. Bellocq, M.I. 1990. Composición y variación temporal de la dieta de Tyto alba en ecosistemas agrarios pampeanos, Argentina. Vida Silvestre Neotropical 2: 32-35. [ Links ]
13. Bianchi, A. y Yañez, C. 1992. Las precipitaciones en el Noroeste Argentino. Segunda Edición. INTA. Estación Experimental Agropecuaria Salta, 61 pp. [ Links ]
14. Blaustein, S.A., Liascovich, R.C., Apfelbaum, L.I., Daleffe, L., Barquez, R.M. y Reig, O.A. 1992. Correlates of systematic differentiation between two closely related allopatric populations of the Akodon boliviensis group from NW Argentina (Rodentia, Cricetidae). Mammalian Biology 57: 1-13. [ Links ]
15. Bobba, M.E. y Hernández, C.M. 2005. Aspectos geográficos del noroeste argentino. En: J.L. Minetti (ed.), El clima del noroeste argentino, Editorial Magna, Laboratorio Climatológico Sudamericano, San Miguel de Tucumán, pp. 9-25. [ Links ]
16. Brown, A.D., Grau, H.R., Malizia, L.R. y Grau, A. 2001. Argentina. En: M. Kappelle y A.D. Brown (eds.), Bosques Nublados del Neotrópico, Instituto Nacional de Biodiversidad (INBio), Costa Rica, pp. 623-659. [ Links ]
17. Cabrera, A.L. 1976. Territorios fitogeográficos de la República Argentina. Enciclopedia Argentina de Agricultura y Jardinería 2: 1-85. [ Links ]
18. Capllonch, P., Autino, A., Díaz, M.M., Barquez R. y Goytia, M. 1997. Los mamíferos del Parque Biológico Sierra de San Javier, Tucumán, Argentina: observaciones sobre su sistemática y distribución. Mastozoología Neotropical 4: 49-71. [ Links ]
19. Carignano, C. 1999. Late Pleistocene to recent climate change in Córdoba Province, Argentina: geomorphological evidence. Quaternary International 57/58: 117-134. [ Links ]
20. Carter-Stiglitz, B., Banerjee, S.K., Gourlan, A. y Oches, E. 2006. A multi-proxy study of Argentina loess: marine oxygen isotope stage 4 and 5 environmental record from pedogenic hematite. Palaeogeography, Palaeoclimatology, Palaeoecology 239: 45-62. [ Links ]
21. Cirignoli, S., Podestá, D.H. y Pardiñas, U.F.J. 2001. Diet of the short-eared owl in Northwestern Argentina. Journal of Raptor Research 35: 68-69. [ Links ]
22. Collantes, M., Powell, J.E. y Sayago, J.M. 1993. Formación Tafí del Valle (Cuaternario superior), provincia de Tucumán (Argentina): litología, paleontología y paleoambientes. 12° Congreso Geológico Argentino y 2° Congreso de Exploración de Hidrocarburos (Buenos Aires), Actas 2: 200-206. [ Links ]
23. Dalby, P.L. y Mares, M.A. 1974. Notes on the distribution of the coney rat, Reithrodon auritus, in Northwestern Argentina. American Midland Naturalist 92: 205-206. [ Links ]
24. D'Elía, G. 2003. Plylogenetics of sigmondontinae (Rodentia, Muroidea, Cricetidae) with special reference to the akodont group, and with additional comments on historical biogeography. Cladistics 19: 307-323. [ Links ]
25. D'Elía, G., Teta, P. y Pardiñas, U.F.J. 2006. Sigmodontinae incertae sedis. En: R. Barquez, M. Díaz y R. Ojeda (eds.), Mamíferos de Argentina: sistemática y distribución, Sociedad Argentina para el Estudio de los Mamíferos, Tucumán, pp. 197-202. [ Links ]
26. Denys, D., Dauphin, Y. y Fernández-Jalvo, Y. 1997. Apports biostratigraphiques et paléoécologiques de l'étude taphonomique des assemblages de micromammifères. Bilan et perspectives. Geobios 30, Supplement 1: 197-206. [ Links ]
27. Díaz, M.M. 1999. [Mamíferos de la Provincia de Jujuy: sistemática, distribución y ecología. Tesis doctoral, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán, Tucumán, Argentina, 563 pp. Inédita] [ Links ]
28. Díaz, M.M. y Barquez, R.M. 1999. Contributions to the knowledge of the mammals of Jujuy Province, Argentina. The Southwestern Naturalist 44: 324-333. [ Links ]
29. Díaz, M.M., Braun, J.K., Mares, M.A. y Barquez, R.M. 2000. An update of the taxonomy, systematics, and distribution of the mammals of Salta Province, Argentina. Occasional Papers, Sam Noble Oklahoma Museum of Natural History 10: 1-52. [ Links ]
30. Dragoo, J.W., Salazar-Bravo, J., Layne, L. y Yates, T. 2003. Relationships within the Calomys callosus species group based on amplified fragment length polymorphisms. Biochemical Systematics and Ecology 31: 703-713. [ Links ]
31. Esteban, G.I. 1993. A new genus of Mylodontinae from the Pleistocene of Northwestern Argentina (El Mollar, Tafí del Valle, Tucumán). Quaternary of South America and Antarctic Peninsula 8: 29-37. [ Links ]
32. FAUNMAP Working Group. 1996. Spatial response of mammals to Late Quaternary environmental fluctuations. Science 272: 1601-1606. [ Links ]
33. Fernández Jalvo, Y. 1996. Small mammal taphonomy and the middle Pleistocene environments of Dolina, Northern Spain. Quaternary International 33: 21-34. [ Links ]
34. Fernández Jalvo, Y. y Andrews, P. 1992. Small mammal taphonomy of Gran Dolina, Atapuerca (Burgos), Spain. Journal of Archaeological Sciences 19: 407-428. [ Links ]
35. Fischer [Von Waldheim], G. 1814. Zoognosia tabulis synopticis illustrata, in usum praelectionum Academiae Imperialis Medico- Chirurgicae Mosquensis Edita. Nicolai Sergeidis Vsevolozsky, Moscow, 3, Quadrupedum reliquorum, cetorum et monotrymatum descriptionem continents, xxiv+694 pp. [ Links ]
36. Fisher, R.A., Corbet, A.S. y Williams, C.B. 1943. The relation between the number of species and the number of individuals in a random sample of an animal population. Journal of Animal Ecology 12: 42-58. [ Links ]
37. Geyh, M.A. y Grosjean, M. 1999. Establishing a reliable chronology of lake level changes in the Chilean Altiplano: a result of close collaboration between geochronologists and geomorphologists. Zentralblatt für Geologie und Paläontologie 7/8: 985- 995. [ Links ]
38. Graham, R. 1985. Response of mammalian communities to environmental changes during the late Quaternary. En: J. Diamond y T.J. Case (eds.), Community Ecology, Harper and Row, New York, pp. 300-313. [ Links ]
39. Graham R. y Lundelius, E. 1984. Coevolutionary disequilibrium and Pleistocene extinctions. En: P.S. Martin y R.G. Klein (eds.), Quaternary extinctions: a prehistoric revolution, University of Arizona Press, Tucson, pp. 223-249. [ Links ]
40. Grau, A. 1985. La expansión del aliso del cerro (Alnus acuminata H.B.K. subsp. acuminata) en el Noroeste de Argentina. Lilloa 36: 237-247. [ Links ]
41. Grayson, D.K. 1984. Quantitative Zooarchaeology. Topics in the analysis of archaeological faunas. Studies in Archaeological Sciences, Academic Press Inc., New York, 202 pp. [ Links ]
42. Herrera, C.M. y Ortiz, P.E. 2003. Primer registro de Chaetophractus vellerosus para el Pleistoceno Tardío de la provincia de Tucumán, Argentina. 18º Jornadas Argentinas de Mastozoología (La Rioja), Actas 1: 30-31. [ Links ]
43. Hershkovitz, P. 1962. Evolution of neotropical cricetine rodents (Muridae) with special reference to the Phyllotine group. Fieldiana Zoology 46: 1-524. [ Links ]
44. Iriondo, M.H. 1993. Cambios climáticos en el noroeste durante los últimos 15,000 años. En: M. Iriondo (ed.), El Holoceno en la Argentina, II, Cadincua, pp. 35-44. [ Links ]
45. Iriondo, M.H. y García, N. 1993. Climatic variations in the Argentine plains during the last 18,000 years. Palaeogeography, Palaeoclimatology, Palaeoecology 101: 209-220. [ Links ]
46. Jayat, J.P. y Pacheco S.E. 2006. Distribución de Necromys lactens y Phyllotis osilae (Rodentia: Cricetidae: Sigmodontinae) en el noroeste argentino: modelos predictivos basados en el concepto de nicho ecológico. Mastozoología Neotropical 13: 69-88. [ Links ]
47. Jayat, J.P., D'Elia, G., Pardiñas, U.F.J. y Namen, J.G. 2007. A new species of Phyllotis (Rodentia, Cricetidae, Sigmodontine) from the upper montane forest of the Yungas of northwestern Argentina. En: D. Kelt, E. Lessa y J. Salazar-Bravo (eds.), The Quintessential Naturalist. Honoring the Life and Legacy of Oliver P. Pearson, California University Press, pp. 775-798. [ Links ]
48. Jayat, J.P., Ortiz, P.E., Teta, P., Pardiñas, U.F.J. y D'Elía, G. 2006. Nuevas localidades argentinas para algunos roedores sigmodontinos (Rodentia: Cricetidae). Mastozoología Neotropical 13: 51-67. [ Links ]
49. Kemp, R.A., Toms, P.S., Sayago, J.M., Derbyshire, E., King, M. y Wagoner, L. 2003. Micromorphology and OSL dating of the basal part of the loess-paleosol sequence at La Mesada in Tucumán province, Northwest Argentina. Quaternary International 106-107: 111-117. [ Links ]
50. Kemp, R.A., King, M., Toms, P.S., Derbyshire, E., Sayago, J.M. y Collantes, M. 2004. Pedosedimentary development of part of a Late Quaternary loess-palaeosol sequence in northwest Argentina. Journal of Quaternary Science 19: 567-576. [ Links ]
51. Kramer, K., Monjeau, A., Birney, E. y Sikes, R. 1999. Phyllotis xanthopygus. Mammalian Species 617: 1-7. [ Links ]
52. Kröhling, D.M. e Iriondo, M. 1999. Upper Quaternary palaeoclimates of the Mar Chiquita, North Pampa, Argentina. Quaternary International 57/58: 149-163. [ Links ]
53. Lupo, L.C. 1998. [Estudio sobre la lluvia polínica actual y la evolución del paisaje a través de la vegetación durante el Holoceno en la cuenca del río Yavi. Borde oriental de la Puna, Noroeste argentino. Tesis doctoral, Fakultät für Geschichts- und Geowissenschaften, Universität Bamberg, Alemania, 87 pp. Inédita.]. [ Links ]
54. Magurran, E. 1988. Ecological Diversity and its Measurement. Princeton University Press, Princeton, 179 pp. [ Links ]
55. Mares, M.A. 1977. Water balance and other ecological observations on three species of Phyllotis in northwestern Argentina. Journal of Mammalogy 58: 514-520. [ Links ]
56. Mares, M.A., Ojeda, R.A. y Barquez, R.M. 1989. Guide to the mammals of Salta Province, Argentina - Guía de los mamíferos de la Provincia de Salta, Argentina. University of Oklahoma Press, Norman, 303 pp. [ Links ]
57. Mares, M.A., Barquez, R.M., Braun, J.K. y Ojeda, R.A. 1996. Observations on the mammals of the Tucumán province, Argentina. I. Systematics, distribution, and ecology of the Didelphimorphia, Xenarthra, Chiroptera, Primates, Carnivora, Perissodactyla, Artiodactyla, and Lagomorpha. Annals of Carnegie Museum 65: 89-152. [ Links ]
58. Mares, M.A., Ojeda, R.A., Braun, J.K. y Barquez, R.M. 1997. Systematics, distribution and ecology of the mammals of Catamarca province, Argentina. En: T. Yates, W. Gannon y D. Wilson (eds.), Life among the Muses: Papers in Honor of James S. Findley, Museum of Southwestern Biology, University of New Mexico, Albuquerque, pp. 89-141. [ Links ]
59. Markgraf, V. 1985. Paleoenvironmental history of the last 10,000 years in Northwestern Argentina. Zentralblatt für Geologie und Palaontologie 11-12: 1739-1749. [ Links ]
60. Markgraf, V. 1989. Paleoclimates in Central and South America since 18,000 BP based on pollen and lake-level records. Quaternary Science Reviews 8: 1-24. [ Links ]
61. Messerli, B., Grosjean, M., Bonani, G., Bürgi, A., Geyh, M., Graf, K., Ramseyer, K., Romero, H., Schotterer, U., Schreier, H. y Vuille, M. 1993. Climate change and natural resource dynamics of the Atacama altiplano during the last 18,000 years: a preliminary synthesis. Mountain Research and Development 13: 117-127. [ Links ]
62. Meyen, F.J.F. 1833. Beitrage zur Zoologie, gesammelt auf einer reise um die Erde. Zweite Abhandlung. Saugethiere. Nova Acta Physico-Medica Academiae Caesariae Leopoldino-Carolinae Naturae Curiosorum (Verhandlungen der Kaiserlichen Leopoldinisch- Carolinischen Akademie der Naturforscher) 16: 599-600. [ Links ]
63. Minetti, J.L., Vargas, W.M., Hernández, C.M. y López, C.R. 2005. La circulación regional estacional en Sud América. Su incidencia en el clima del noroeste argentino. En: J.L. Minetti (ed.), El clima del noroeste argentino, Editorial Magna, Laboratorio Climatológico Sudamericano, Tucumán, pp. 39-75. [ Links ]
64. Monjeau, J.A. 1989. [Ecología y distribución geográfica de los pequeños mamíferos del Parque Nacional Nahuel Huapí y áreas adyacentes. Tesis doctoral, Universidad Nacional de La Plata, Buenos Aires, 253 pp. Inédita.]. [ Links ]
65. Myers, P., Patton, J.L. y Smith, M.F. 1990. A review of the boliviensis group of Akodon (Muridae: Sigmodontinae), with emphasis on Peru and Bolivia. Miscelaneous Publications, Museum of Zoology, University of Michigan 177: 1-104. [ Links ]
66. Ojeda, R.A. y Mares, M.A. 1989. A biogeographic analysis of the mammals of Salta Province, Argentina. Patterns of species assemblage in the neotropics. Special Publication, Museum Texas Tech University 27: 1-66. [ Links ]
67. Ortiz, P.E. 2001. [Roedores del Pleistoceno superior del valle de Tafí (Provincia de Tucumán), implicancias paleoambientales y paleobiogeográficas. Tesis doctoral, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán, Tucumán, Argentina, 230 pp. Inédita.]. [ Links ]
68. Ortiz, P.E. y Pardiñas, U.F.J. 2001. Sigmodontinos (Mammalia, Rodentia) del Pleistoceno tardío del Valle de Tafí (Tucumán, Argentina): taxonomía, tafonomía y reconstrucción paleoambiental. Ameghiniana 38: 3-26. [ Links ]
69. Ortiz, P.E. y Jayat, J.P. 2007. Fossil record of the Andean rat, Andinomys edax (Rodentia: Cricetidae), in Argentina. Mastozoología Neotropical 14: 77-83, [ Links ]
70. Ortiz, P.E., Cirignoli, S., Podestá, D. y Pardiñas, U.F.J. 2000a. New records of sigmodontine rodents (Mammalia: Muridae) from high-Andean localities of Northwestern Argentina. Biogeographica 76: 133-140. [ Links ]
71. Ortiz, P.E., Pardiñas U.F.J. y Steppan S.J. 2000b. A new fossil phyllotine (Rodentia: Muridae) from Northwestern Argentina and the relationships of Reithrodon group. Journal of Mammalogy 81: 37-51. [ Links ]
72. Pardiñas, U.F.J. 1999. [Los roedores muroideos del Pleistoceno Tardío- Holoceno en la región pampeana (sector este) y Patagonia (República Argentina): aspectos taxonómicos, importancia bioestratigráfica y significación paleoambiental. Tesis doctoral, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Argentina, 283 pp. Inédita.]. [ Links ]
73. Pardiñas, U.F.J. 2000. Tafonomía de microvertebrados en yacimientos arqueológicos de Patagonia (Argentina). Arqueología 9: 265-340. [ Links ]
74. Pardiñas, U.F.J. y Galliari, C.A. 2001. Reithrodon auritus. Mammalian Species 664: 1-8. [ Links ]
75. Pardiñas, U.F.J. y Lezcano, M. 1995. Cricétidos (Mammalia: Rodentia) del Pleistoceno tardío del nordeste de la provincia de Buenos Aires (Argentina). Aspectos sistemáticos y paleoambientales. Ameghiniana 32: 249-265. [ Links ]
76. Pardiñas, U.F.J. y Ortiz, P.E. 2001. Neotomys ebriosus, an enigmatic South American rodent (Muridae, Sigmodontinae): its fossil record and present distribution in Argentina. Mammalia 65: 244-250. [ Links ]
77. Pardiñas, U.F.J., Moreira, G.J., García-Esponda, C.M. y De Santis, L.J.M. 2000. Deterioro ambiental y micromamíferos durante el Holoceno en el nordeste de la estepa patagónica (Argentina). Revista Chilena de Historia Natural 73: 9-21. [ Links ]
78. Pardiñas, U.F.J., D´Elía, G., Teta, P., Ortiz, P.E., Jayat, J.P. y Cirignoli, S. 2006. Akodontini Vorontzov, 1959 (sensu D´Elía, 2003). En: R. Barquez, M. Díaz y R. Ojeda (eds.), Mamíferos de Argentina: sistemática y distribución, Sociedad Argentina para el Estudio de los Mamíferos, Mendoza: 146-166. [ Links ]
79. Pearson, O.P. 1988. Biology and feeding dynamics of a South American herbivorous rodent, Reithrodon. Studies on Neotropical Fauna and Environment 23: 25-39. [ Links ]
80. Powell, J.E. 1998. Los vertebrados fósiles. En: M. Gianfrancisco, M. E. Puchulu, J. Durango de Cabrera y G. F. Aceñolaza (eds.), Geología de Tucumán, Publicación Especial del Colegio de Graduados en Ciencias Geológicas de Tucumán, pp. 227-240. [ Links ]
81. Reig, O.A. 1977. A proposed unified nomenclature for the enamelled components of the molar teeth of the Cricetidae (Rodentia). Journal of Zoology 181: 227-241. [ Links ]
82. Reimer, P., Baillie, M., Bard, E., Bayliss, A., Beck, J., Bertrand, C., Blackwell, P., Buck, C., Burr, G., Cutler, K., Damon, P., Edwards, R., Fairbanks, R., Friedrich, M., Guilderson, T., Hogg, A., Hughen, K., Kromer, B., McCormac, G., Manning, S., Ramsey, C., Reimer, R., Remmele, S., Southon, J., Stuiver, M., Talamo, S., Taylor, F., van der Plicht, J. y Weyhenmeyer, C. 2004. IntCal04 terrestrial radiocarbon age calibration, 0-26 cal kyr BP. Radiocarbon 46: 1029-1058. [ Links ]
83. Rufino, S.D. y Mulé, P. 1994. Un Megatheriinae (Mammalia- Edentata) en un valle de altura en la provincia de Tucumán, Argentina. Acta Geológica Leopoldiana 39: 415-427. [ Links ]
84. Salazar-Bravo, J., Dragoo, J.W., Tinnin, D.S. y Yates, Y.L. 2001. Phylogeny and evolution of the Neotropical rodent genus Calomys: inferences from Mitochondrial DNA sequence data. Molecular Phylogenetics and Evolution 20: 173-184. [ Links ]
85. Salazar-Bravo, J., Dragoo, J.W., Bowen, M.D., Peters, C.J., Ksiazek, T.G. y Yates, Y.L. 2002. Natural nidality in Bolivian hemorrhagic fever and the systematics of the reservoir species. Infection, Genetics and Evolution 1: 191-199. [ Links ]
86. Sayago, J.M., Powell, J.E., Collantes, M.M. y Neder L.V. 1998. Cuaternario. En: M. Gianfrancisco, M.E. Puchulu, J. Durango de Cabrera y G.F. Aceñolaza (eds.), Geología de Tucumán, Publicación Especial del Colegio de Graduados en Ciencias Geológicas de Tucumán, pp. 111-128. [ Links ]
87. Sayago, J.M., Zinck, J.A., Collantes, M. y Toledo, M. 2005. Environmental changes in the pre-Andean valleys and Chaco Plain (Northwest Argentina) during the Late Pleistocene and Holocene. Neues Jahrbuch für Geologie und Paläontologie 236: 245-265. [ Links ]
88. Schellenberger, A., Heller, F. y Veit, H. 2003. Magnetostratigraphy and magnetic suceptibility of the Las Carreras loess-paleosol sequence in Valle de Tafí, Tucumán, NW-Argentina. Quaternary International 106-107: 159-167. [ Links ]
89. Sesma, P.J., Guido, E. y Puchulu, M.E. 1998. Clima de la provincia de Tucumán. En: M. Gianfrancisco, M.E. Puchulu, J. Durango de Cabrera y F.G. Aceñolaza (eds.), Geología de Tucumán, Publicación Especial del Colegio de Graduados en Ciencias Geológicas de Tucumán, pp. 41-46. [ Links ]
90. Stahl, P.W. 1996. The recovery and interpretation of microvertebrate bone assemblages from archaeological contexts. Journal of Archaeological Method and Theory 3: 31-75. [ Links ]
91. Steppan, S.J. 1995. Revision of the tribe Phyllotini (Rodentia: Sigmodontinae), with a phylogenetic hypothesis for the Sigmodontinae. Fieldiana, Zoology, n. s., 80: 1-112. [ Links ]
92. Steppan, S.J. 1998. Phylogenetic relationships and species limits within Phyllotis (Rodentia: Sigmodontinae): concordance between MTDNA sequence and morphology. Journal of Mammalogy 79: 573-593. [ Links ]
93. Tabeni, M.S., Bender, J.B. y Ojeda, R.A. 2004. Puntos calientes para la conservación de mamíferos en la provincia de Tucumán, Argentina. Mastozoología Neotropical 11: 55-67. [ Links ]
94. Teta, P. y Ortiz, P.E. 2002. Micromamíferos andinos holocénicos del sitio arqueológico Inca Cueva 5, puna de Jujuy, Argentina: tafonomía, zoogeografía y reconstrucción paleoambiental. Estudios Geológicos 58: 113-135. [ Links ]
95. Thomas, O. 1897. On some small mammals from Salta, N. Argentina. Annals and Magazine of Natural History, series 6, 20: 214-218. [ Links ]
96. Thomas, O. 1902. On mammals collected by Mr. Perry O. Simmons in the southern part of the Bolivian Plateau. Annals and Magazine of Natural History, series 7, 9: 222-230. [ Links ]
97. Thomas, O. 1913. On small Mammals collected in Jujuy by Señor E. Budin. Annals and Magazine of Natural History, series 8, 11: 136-143. [ Links ]
98. Thomas, O. 1916. Two new species of Akodon from Argentina. Annals and Magazine of Natural History 8: 334-336. [ Links ]
99. Thomas, O. 1918. On small Mammals from Salta and Jujuy collected by Mr. E. Budin. Annals and Magazine of Natural History, series 9, 1: 186-193. [ Links ]
100. Thomas, O. 1920. New species of Reithrodon, Abrocoma, and Scapteromys from Argentina. Annals and Magazine of Natural History (London), series 9, 5: 473-478. [ Links ]
101. Thomas, O. 1925. On some Argentine mammals. Two new rodents from Tucumán Province. Annales and Magazine of Natural History 9: 582-584. [ Links ]
102. Torres Bruchmann, E. 1972. Mesoclimas de la provincia de Tucumán. Revista Agronómica del Noroeste Argentino 9 (3-4): 527-544. [ Links ]
103. Vervoorst, F. 1982. Noroeste. En: 18 ° Jornadas Argentinas de Botánica, Simposio Conservación de la vegetación natural en la República Argentina, Serie Conservación de la Naturaleza (Fundación Miguel Lillo) 2: 9-23. [ Links ]
104. Waterhouse, G.R. 1837. [Characters of new species of the genus Mus, from the collection of Mr. Darwin.] Proceedings of the Zoological Society of London 1837: 15-21, 27-32. [ Links ]
105. Yepes, J. 1935. Consideraciones sobre el género "Andinomys" (Cricetinae) y descripción de una forma nueva. Anales del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" 38: 333-348. [ Links ]
106. Zinck, J.A. y Sayago, J.M. 1999. Loess-paleosol sequence of La Mesada in Tucumán Province, north west Argentina. Characterization and paleoenvironmental interpretation. Journal of South American Earth Sciences 5: 1-18. [ Links ]
107. Zinck, J.A. y Sayago, J.M. 2001. Climatic periodicity during the late Pleistocene from a loess-paleosol sequence in northwest Argentina. Quaternary International 78: 11-16. [ Links ]
108. Zipprich, M., Reizner, B., Zech, W., Stingl, H. y Veit, H. 1999. Upper Quaternary landscape and climate evolution in the Sierra de Santa Victoria (north-western Argentina) deduced from geomorphologic and pedogenic evidence. Zentralblatt für Geologie und Paläontologie 7/8: 997-1011. [ Links ]
Recibido: 19 de octubre de 2005.
Aceptado: 18 de setiembre de 2007.

