










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.44 n.4 Buenos Aires oct./dic. 2007
Nuevo carpoideo de la clase Cincta Jaekel, 1918 del norte de España: inferencias sobre la morfología funcional del opérculo
Samuel Zamora1, Eladio Liñán1, José A. Gámez Vintaned1, Patricio Domínguez Alonso2 y Rodolfo Gozalo3
1Área y Museo de Paleontología. Departamento de Ciencias de la Tierra. Universidad de Zaragoza. E-50009 Zaragoza. España. samuel@unizar.es , linan@unizar.es, gamez@unizar.es
2Departamento de Paleontología. Facultad de Ciencias Geológicas. Universidad Complutense. E-28040 Madrid. España. patricio@geo.ucm.es
3Departamento de Geología. Universitat de València. E-46100 Burjasot (Valencia). España. Rodolfo.Gozalo@uv.es
Abstract. A new carpoid (class Cincta Jaekel, 1918) from northern Spain and its bearing on the functional morphology of the operculum. Cinctan carpoids from Middle Cambrian (Languedocian) rocks of northern Spain are described. Sampled specimens probably belong to a new species of the genus Gyrocystis Jaekel; nevertheless, the lack of some key features prevents against erecting formally a new taxon at this moment. A functional interpretation of the operculum is made, and the evolutionary meaning of some structures related with its movement is analysed herein. When the opercular area of different cinctans (Asturicystis, Sotocinctus and Gyrocystis) are compared, it is concluded that several features in Gyrocystis sp. L were probably related to a better mobility of the operculum. These are the lesser number of postopercular plates anchored to the operculum, the reduction of the postopercular facet, a more rigid lintel, and the existence of quite big adopercular facets on the inner side of the adopercular processes. All these features permitted Gyrocystis sp. L a higher efficiency in expelling water out of the theca, and they can be interpreted as apomorphies in the evolutionary history of cinctans. The term postopercular facet is defined here as the articulation structure between the operculum and the postopercular plates.
Resumen. Se estudia nuevo material de carpoideos Cincta del Cámbrico Medio (Languedociense) del norte de España. Los ejemplares probablemente pertenecen a una nueva especie del género Gyrocystis Jaekel, que ante la no conservación de algunos elementos se prefiere mantener en nomenclatura abierta. Además, se realiza una interpretación funcional del opérculo y se analiza el significado evolutivo de algunas estructuras relacionadas con su articulación y movimiento. Comparando el complejo portaopercular en diferentes Cincta (Asturicystis, Sotocinctus y Gyrocystis), se observa que el descenso en el número de placas postoperculares ancladas al opérculo, la reducción de la faceta postopercular, el desarrollo de un dintel más rígido y la presencia de grandes facetas adoperculares en la cara interna de los tubérculos adoperculares podrían estar relacionados con una mayor movilidad del opérculo. Todas estas características permitirían una mayor eficacia en el proceso de expeler el agua fuera de la teca y pueden ser consideradas como apomorfías en la historia evolutiva de los Cincta. Se acuña el término faceta postopercular para la estructura que sirve de articulación del opérculo con las placas postoperculares.
Key words. Cincta; Middle Cambrian; Spain; Functional morphology.
Palabras clave. Cincta; Cámbrico Medio; España; Morfología funcional.
Introducción
Uno de los grupos fósiles del Cámbrico más extraño y enigmático son los carpoideos de la clase Cincta Jaekel, 1918. Aunque fueron descubiertos hace más de un siglo (Barrande, 1859), su anatomía no ha empezado a entenderse bien hasta tiempos más recientes con los trabajos de Ubaghs (1967) y, sobre todo, los de Friedrich (1993) y Sdzuy (1993).
Los Cincta fueron organismos con una organización corporal única entre todos los grupos de carpoideos. Presentan un cuerpo en forma de raqueta cuyo esqueleto de naturaleza calcítica es asimétrico y está formado por dos partes principales. Una está constituida por grandes placas que forman un anillo marginal denominado cinctus, y otra compuesta por un apéndice caudal en forma de espadilla llamado estela. Cerrando el cinctus por arriba y por debajo aparecen dos tegumentos de placas centrales más pequeñas denominados supracentralia e infracentralia. En la parte anterior aparece una cavidad (cámara distal) cubierta por una gran placa denominada opérculo que representa la principal autapomorfía de este grupo.
La interpretación de estos organismos ha diferido a lo largo de la historia y sigue siendo controvertida hoy en día. Las discrepancias radican principalmente en la interpretación de las diferentes aberturas de la teca, su relación con la anatomía interna y las diferentes homologías reconocidas. Existen dos escuelas que situarían a los Cincta, bien como equinodermos o bien como hemicordados. Dentro de la primera también existen importantes discrepancias. Según Ubaghs (1975), Friedrich (1993), Jefferies et al. (1996), Domínguez (1999) y Smith (2005), se trataría del grupo más basal del stem group Echinodermata; pero David et al. (2000) los incluyen como un grupo muy derivado de blastozoos. La otra escuela, sobre la base de estudios cladísticos de biología molecular y por la anatomía de los fósiles de deuteróstomos primitivos (Domínguez y Jefferies, 2005), sitúa los Cincta como un grupo basal de hemicordados, muy relacionado con los carpoideos ctenocistoideos (Friedrich, 1993; Jefferies et al., 1996; David et al., 2000).
Los objetivos de este trabajo son dos principalmente. Por una parte, presentar nuevo material de cincta del Cámbrico Medio (Languedociense) del norte de España. Por otra, describir la excepcional conservación de la zona opercular en uno de los ejemplares y compararla con las variaciones morfológicas de esta estructura a lo largo de la historia evolutiva del grupo, para de esa manera inferir algunos aspectos sobre la funcionalidad del opérculo.
Situación geográfica y geológica
Los Barrios de Luna es una pequeña localidad de los Montes de León (Cordillera Cantábrica) situada en la provincia de León (norte de España). Desde el punto de vista geológico, se localiza en la parte meridional de la Zona Cantábrica, una de las divisiones tectonoestratigráficas del Macizo Ibérico, y dentro de ella en el denominado Dominio tectónico de Somiedo-Correcilla (Zamarreño, 1972) (figura 1).

Figura 1. Situación geológica del yacimiento de Los Barrios de Luna (provincia de León, N de España)(*), según Sdzuy y Liñán (1993) / geological setting of the fossiliferous site at Los Barrios de Luna (León Province, Cantabrian Mountains, N Spain) (*). Based on Sdzuy and Liñán (1993).
La región de Los Barrios de Luna es una de las más estudiadas y emblemáticas del Cámbrico de la Cordillera Cantábrica (figura 1). La sección estudiada (Los Barrios de Luna 1, BL1) se localiza paralela al río Luna, a lo largo de la carretera de su margen derecha (aguas arriba), que va desde las inmediaciones del pueblo de Mora hasta la presa de Los Barrios de Luna. En esta sección aparecen representadas, en orden estratigráfico ascendente, la Formación Mora (constituida por lutitas y areniscas alternantes con rocas volcánicas, de edad neoproterozoica), la Formación Herrería (formada por cuarcitas, lutitas pizarrosas, areniscas groseras y conglomerados; Cámbrico Inferior), la Formación Láncara (que es una potente sucesión de calizas y dolomías, Cámbrico Inferior-Medio), la Formación Oville (representada por una sucesión de lutitas pizarrosas con esporádicos nódulos carbonatados en la base que culmina con areniscas finas; Cámbrico Medio) y la Cuarcita de Barrios (una sucesión de cuarcitas y areniscas groseras; Cámbrico Medio-Superior).
El C ámbrico de Los Barrios de Luna fue citado por Gómez de Llarena y Rodríguez Arango (1948). Pero fueron Lotze y Sdzuy (1961) los que describieron el primer perfil bioestratigráfico de la sucesión y destacaron su gran importancia en el contexto del Cámbrico europeo. Los datos bioestratigráficos y litoestratigráficos fueron ampliados por Sdzuy (1969).
Zamarre ño (1972) realizó un estudio detallado de la estratigrafía y sedimentología de la Formación Oville de esta localidad (figura 2), en la que se encuentran los Cincta aquí estudiados. Desde un punto de vista facial, el Cámbrico que nos ocupa corresponde a lo que Zamarreño (1972) define como facies Los Barrios dentro del Cámbrico de la Cordillera Cantábrica, y que posteriormente relacionó con el resto de las facies del Cámbrico español (Zamarreño, 1983). La "facies Los Barrios" fue estudiada posteriormente por Sdzuy y Liñán (1993) desde un punto de vista megafacial y en relación a su contexto paleogeográfico y paleoecológico dentro del Cámbrico del norte de España. Recientes síntesis del Cámbrico de la Zona Cantábrica se encuentran en Aramburu et al. (1992, 2004).
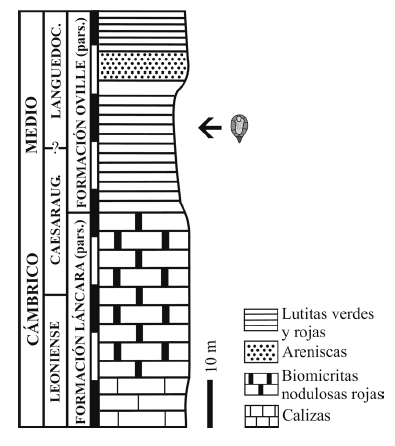
Figura 2. Posición estratigráfica del material estudiado (basado en Zamarreño, 1972) / stratigraphy of the fossiliferous site (based on Zamarreño, 1972).
La sucesi ón estratigráfica de la sección BL1 se encuentra muy tectonizada en los niveles inferiores donde aflora la Formación Herrería. La parte superior formada por los materiales de las Formaciones Láncara y Oville es muy continua. Allí la sucesión comienza con 60 metros de dolomías con laminaciones algales, a las que siguen 30 metros de calizas glauconíticas rosadas, sobre las que se sobreimponen 20 metros de calizas rojas nodulosas. Todas ellas constituyen parte de la Formación Láncara. La suprayacente Formación Oville, donde fueron encontrados los Cincta presentados en este trabajo (figura 2), es una unidad compuesta, en esta región, casi exclusivamente por lutitas pizarrosas entre las que se intercala, a casi treinta metros de su base, un nivel arenoso conocido regionalmente como la Arenisca de simula, nombre que hace referencia a una especie del género de trilobites Solenopleuropsis, del Cámbrico Medio. Los carpoideos estudiados en el presente trabajo se localizan a unos 20 metros de la base de la Formación Oville. Junto a ellos también está presente el trilobite Solenopleuropsis marginata Sdzuy, 1958, que indica la biozona de extensión homónima, o bien la biozona suprayacente (S. thorali) donde también aparece esta especie.
La existencia de Cincta en la Cordillera Cant ábrica se conoce desde muy antiguo (Prado et al., 1860), y multitud de trabajos han nombrado o descrito formas de este grupo (Schröeder, 1973, y referencias allí recogidas; Sdzuy, 1993; Gil Cid y Domínguez Alonso, 1995a). Sin embargo, en el área de Los Barrios de Luna sólo se ha descrito un taxón en nomenclatura genérica y específica abiertas (Friedrich, 1993). Éste parece estar muy relacionado con los Trochocystidae, a diferencia del que se describe en el presente trabajo, que pertenece al género Gyrocystis (Jaekel). De la misma localidad fue descrito el eocrinoideo Ubaghsicystis segurae Gil Cid y Domínguez Alonso, 2002.
Paleontología sistemática
El material está depositado en el Museo Paleontológico de la Universidad de Zaragoza con las siglas MPZ 2006/15 y MPZ 2006/16.
Metodología y terminología empleada
Ya que los fósiles aparecen disueltos y se conservan como moldes naturales, se han realizado dibujos utilizando cámara clara de réplicas en látex de los ejemplares, con el fin de estudiar su anatomía.
Conviene aclarar que, aunque a lo largo de la historia ha habido numerosas propuestas para nombrar las diferentes placas de la marginalia de los Cincta (Jaekel, 1918; Gislén, 1927; Meléndez, 1952; Ubaghs, 1967; Termier y Termier, 1973), en este trabajo se sigue la propuesta de Friedrich (1993), adoptada por Sdzuy (1993) y Fatka y Kordule (2001), en la que la placa marginal que forma parte de la base de la cámara distal (situada en el extremo anterior del cuerpo y sobre el plano sagital) se denomina placa M0 y mantendría siempre una posición central. A partir de ella se van numerando las sucesivas placas marginales (M1l, M1r, M2l, M2r…) según se localicen a la izquierda (l) o a la derecha (r) de la M0. Sdzuy (1993) destaca que de esta manera se evita el problema que existía con otras nomenclaturas antiguas, en las que se daban distintos nombres a placas situadas en la misma posición en diferentes especies
La bibliografía existente sobre Cincta no es muy abundante; además los términos morfológicos empleados en los diferentes trabajos han sido sustituidos repetidamente, en parte por cuestiones lingüísticas y en parte por cambios en la interpretación morfológica (Sdzuy, 1993). Los términos empleados aquí para la descripción se encuentran ampliamente desarrollados en los trabajos de Ubaghs (1967), Friedrich (1993) y Sdzuy (1993). Una traducción al castellano de la mayoría de ellos puede verse en Gil Cid y Domínguez Alonso (1995b). Aún así, es preciso definir algunos de ellos que son utilizados por primera vez en español e inglés en este trabajo:
Complejo portaopercular (porta-operculum complex, sensu Sdzuy, 1993): estructura compuesta por el porta, el opérculo y el dintel.
Porta (porta, sensu Sdzuy, 1993): es el gran orificio del cinctus, situado en la parte anterior de la cámara distal. Aparece cubierto por el opérculo.
Dintel (lintel, sensu Sdzuy, 1993): arco de placas supracentrales (normalmente 3 ó 4) situado entre las placas M1l y M1r. El dintel se articula al opérculo, a las facetas adoperculares y al resto de la supracentralia.
Facetas adoperculares (suropercular facets) (suroperculare Facetten, sensu Friedrich, 1993): facetas de articulación situadas en la cara interna de la base de los tubérculos adoperculares y que sirven para articular el dintel.
Tubérculos adoperculares (adopercular proceses) (sensu Gil Cid y Domínguez Alonso, 1995a): protuberancias en forma de maza o solapa que aparecen sobre las placas M1r y M1l de algunos Cincta. En ocasiones presentan ornamentación.
Además, se acuña aquí el término faceta postopercular (postopercular facet) para la estructura que sirve de articulación del opérculo con las placas postoperculares.
Clase CINCTA Jaekel, 1918
Familia GYROCYSTIDAE Jaekel, 1918
Discusión. Friedrich (1993) establece como principal criterio diagnóstico para diferenciar las tres familias de Cincta la longitud de los surcos epitecales. El material estudiado aquí no presenta surco epitecal derecho, mientras que el izquierdo no se encuentra conservado. La ausencia del surco epitecal derecho es típica de la familia Gyrocystidae, aunque ocasionalmente algún representante de la familia Sucocystidae (ej. Sucocystis theronensis Cabibel et al., 1959) también carece de él. Aún así, la morfología general de la teca, la unión de la estela al cinctus, el número de placas marginales y la falta de ornamentación en las supracentrales nos llevan a incluir nuestro material dentro de la familia Gyrocystidae.
Género Gyrocystis Jaekel, 1918
Especie tipo. Gyrocystis platessa Jaekel, 1918.

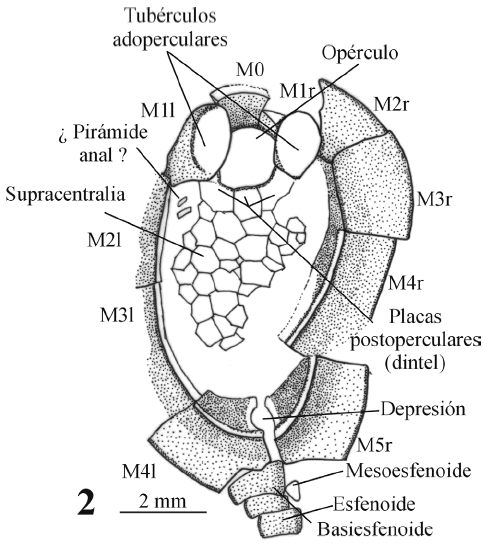
Figura 3. Dibujos con cámara clara de los dos ejemplares estudiados. Ambos aparecen en vista dorsal y se indican las principales características anatómicas. Dibujos realizados a partir de moldes de látex. 1: ejemplar MPZ 2006/15. 2: ejemplar MPZ 2006/16 / camera lucida drawings (made after latex casts) of the two specimens studied in this paper. Both are shown in dorsal view; main anatomical features are indicated. 1: specimen MPZ 2006/15. 2: specimen MPZ 2006/16.
Material y modo de conservación. Dos ejemplares incompletos en vista dorsal conservados como moldes internos y externos, en limolitas de grano fino y color gris-verdoso. Los ejemplares aparecen ligeramente desarticulados en algunas porciones pero no deformados. El tegumento de placas supracentrales se encuentra colapsado sobre las placas infracentrales.
Descripción. La teca de Gyrocystis sp. L es de contorno ovalado (figura 3). Su tamaño es de unos 9 mm de longitud y 6 mm de anchura en el ejemplar menor y de 18 mm x 13 mm en el ejemplar mayor. Hay que tener en cuenta que estas medidas son simplemente orientativas y no representativas del animal original ya que, por una parte, los fósiles están parcialmente desarticulados y, por otra, no se encuentran completos. El cinctus está formado por diez placas marginales (M5r, M4l), que son muy anchas en relación con la altura. La estela se encuentra incompleta en su parte distal y presenta la base muy ensanchada.
Un pequeño orificio (boca) está situado entre las placas M1r y M2r y carece de surco epitecal derecho. El surco epitecal izquierdo no se ha conservado, por lo que no se puede determinar su longitud. La placa M0 se sitúa, como en otros cincta, en la parte anterior y coincidiendo con el plano sagital. Presenta una morfología cóncava y lisa en su cara interna y con una anchura que se reduce ligeramente desde la parte anterior a la posterior. Las placas del cinctus M1r y M1l tienen unos tubérculos adoperculares muy desarrollados (figura 4.1) y con ornamentación de un enrejado en la porción terminal de éstos (figuras 4.1, 5.4). Se orientan hacia delante.
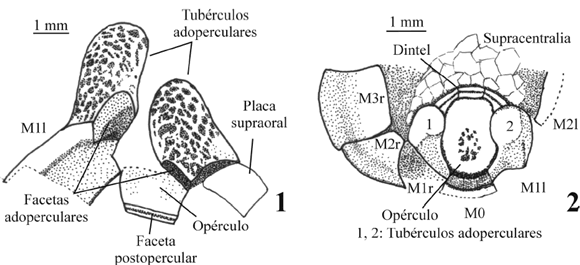
Figura 4. Dibujos con cámara clara de detalle de las estructuras relacionadas con la zona opercular, realizados a partir de los moldes de látex. 1, detalle de la parte posterior de los tubérculos adoperculares. En ellos se observan las facetas adoperculares muy desarrolladas que permiten el anclaje de las placas post-operculares izquierda y derecha del dintel. También se observa la faceta postopercular de articulación del opérculo al dintel. Ejemplar MPZ 2006/15. 2, se observa el opérculo en posición anatómica unido al dintel en su parte posterior. Las placas postoperculares del dintel se articulan con los tubérculos adoperculares. Ejemplar MPZ 2006/16 / close-up, camera lucida drawings (made after latex casts) of structures related to the opercular area. 1, close-up view of the adopercular processes. They show well developed, adopercular facets which allow postopercular plates to attach. Specimen MPZ 2006/15. 2, the operculum appears as in anatomical position, and is bounded with the lintel along the posterior margin. The postopercular plates of the lintel articulate with the adopercular processes. Specimen MPZ 2006/16.
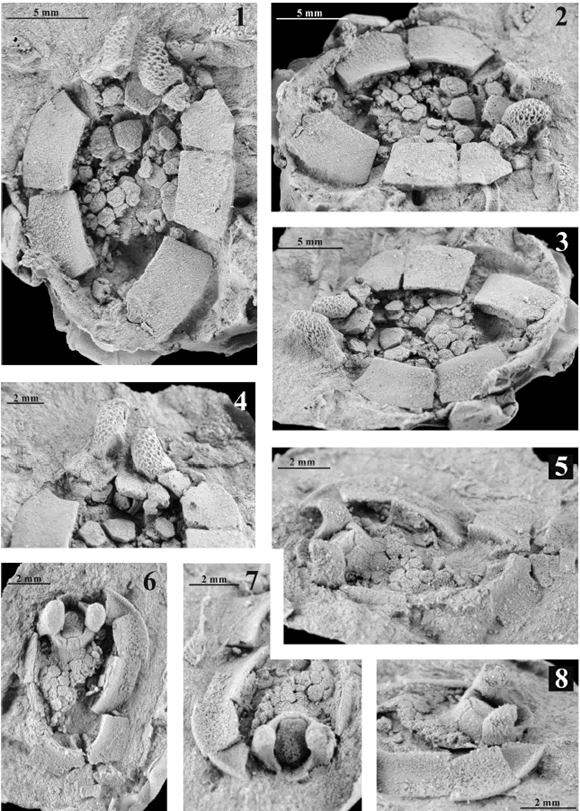
Figura 5. Gyrocystis sp. L. 1-4, ejemplar MPZ2006/15; 1, vista dorsal de una teca casi completa, en ella se observa el marco de placas marginales que está incompleto en la parte posterior; en la parte anterior se observan los tubérculos adoperculares y entre ellos la parte posterior del opérculo; 2, vista oblicua derecha de la teca, en la zona anterior derecha se observa la placa supraoral que se articula al tubérculo adopercular derecho y a la placa M2r del cinctus; 3, vista oblicua izquierda de la teca; 4, detalle de la parte posterior de los tubérculos adoperculares, se observa la faceta adopercular grande, profunda y ovalada en el tubérculo adopercular izquierdo, que permitiría el anclaje de la placa postopercular izquierda del dintel.; además, se aprecia la ornamentación en forma de enrejado de los tubérculos adoperculares y la faceta de articulación postopercular. 5-8, ejemplar MPZ2006/16); 5, vista lateral izquierda de una teca casi completa donde se observan los prominentes tubérculos adoperculares; 6, vista dorsal de la teca de un cincta donde se observa la mayor parte del cinctus y la parte anterior de la estela; la supracentralia aparece parcialmente completa en su parte anterior, el complejo portaopercular aparece conservado de manera excepcional; 7, detalle frontal del complejo portaopercular excelentemente conservado, se aprecia la articulación del opérculo al dintel y de éste a los tubérculos adoperculares; la zona ambulacral está rota por lo que no se observan los surcos epitecales; 8, detalle en vista oblicua derecha del complejo portaopercular y de parte de la teca. Todas las fotografías están realizadas a partir de réplicas en látex de los ejemplares que han sido blanqueadas mediante cloruro de amonio / Gyrocystis sp. L. 1-4, specimen MPZ 2006/15; 1, almost complete theca in dorsal view (the frame of marginal plates -or cinctus- is uncomplete on its rear part; the two adopercular processes are shown on the anterior part, and the rear end of the operculum is visible between them); 2, right, oblique view of the theca (the supraoral plate, shown on the right, anterior part, articulates to the right adopercular process and to the plate M2r of the cinctus); 3, left, oblique view of the theca; 4, detail of the rear part of the adopercular processes (the big, deep and oval adopercular facet of the left process -permitting to attach to the left postopercular plate of the lintel- is shown; moreover, the cancellate ornamentation of the adopercular processes and the articulating suropecular facet is shown). 5-8, specimen MPZ 2006/16; 5, left, side view of the almost complete theca where the prominent adopercular processes are visible; 6, dorsal view of the theca showing a major part of the cinctus, the anterior part of the stele, the supracentralia (well preserved only on its anterior part) and the exceptionally preserved porta-operculum complex; 7, detail of the porta-operculum complex in frontal view (articulation of the lintel with both the operculum and the adopercular processes is shown); since the anterior area is broken, marginal grooves are not visible; 8, detail in oblique view of the porta-operculum complex and part of the theca. All photographs are taken on ammonium chloride-whitened, latex casts.
La placa supraoral se conserva sólo en el ejemplar MPZ2005/15. Presenta un contorno subrectangular y se articula a la parte externa de la base del tubérculo adopercular derecho y a la cara interna de la placa M2r. El complejo portaopercular (figura 4.2) está compuesto como en otros Cincta por el porta, el dintel y el opérculo. El opérculo es piriforme, ligeramente convexo en sección y fuertemente ornamentado (figura 5.7). Se une en su parte posterior, mediante la faceta postopercular, a tres placas postoperculares que forman un arco bien definido o dintel. Las placas postoperculares izquierda y derecha se articulan respectivamente con la cara interna de la base de los tubérculos adoperculares mediante las facetas adoperculares, que están extremadamente desarrolladas.
La placa M2r es de contorno subtriangular. Las placas M3r, M4r, M2l y M3l son m ás largas que anchas y con una relación longitud/anchura de 1,3, 1,4, 1,8 y 1,5 respectivamente (relaciones tomadas a partir del ejemplar MPZ 2006/15). Todas las placas marginales presentan una amplia plataforma marginal y poca altura. Las placas supracentrales son muy grandes (diámetro hasta 1,5 mm) comparadas con otros Cincta, muy planas y de contorno hexagonal. Su tamaño se reduce desde la parte anterior hacia la posterior. Las placas infracentrales no se conservan en ninguno de los dos ejemplares estudiados.
La uni ón de la estela al cinctus se realiza mediante las placas M5r y M4l. En la cara interna de éstas aparece una fuerte depresión en forma de embudo. La estela está incompleta y sólo se conserva la parte proximal en uno de los ejemplares (figura 5.6). Se articula al cinctus mediante dos placas basiesfenoides y una mesoesfenoide. Las basiesfenoides son mucho más anchas que largas, con una relación longitud/anchura de 2,3 que es observable en la basiesfenoide izquierda del ejemplar MPZ2006/16. Además, se encuentran ligeramente arqueadas hacia la parte posterior.
Discusión. Dentro de la familia Gyrocystidae existen dos géneros, Gyrocystis con varias especies y Progyrocystis, cuya única especie es P. disjuncta Friedrich, 1993. Sus principales diferencias son: el cinctus de Progyrocystis aparece abierto por las placas marginales, mientras que en Gyrocystis está cerrado (esto es observable sólo en la cara ventral); el dintel de Gyrocystis está bien definido y presenta tres o más (raramente cuatro) placas postoperculares y el de Progyrocystis al menos cinco (según la reconstrucción de Friedrich, 1993, pág. 102, fig. 12). El material aquí estudiado se atribuye al género Gyrocystis por la morfología del dintel (compuesto por tres osículos postoperculares) y la presencia de placas supracentrales con un tamaño relativo mayor que las que muestra Progyrocystis.
Comparación. Se ha comparado el material con otros Cincta, preferentemente de la familia Gyrocystidae. Los tubérculos adoperculares tan desarrollados hacen que Gyrocystis sp. L se aleje mucho de taxones como G. platessa Jaekel, 1918, G. testudiformis Friedrich, 1993, G. erecta Friedrich, 1993 o G. cruzae Gil Cid y Domínguez, 1995a. Únicamente G. badulesiensis Friedrich, 1993 podría llegar a tener unos tubérculos adoperculares tan prominentes, aunque el taxón descrito aquí se diferencia bien de esta especie por la forma de anclaje del opérculo a las placas postoperculares y por la forma general de las marginales.
Tubérculos adoperculares tan desarrollados se observan en taxones como Elliptocinctus barrandei (Munier-Chalmas y Bergeron, 1889) o Elliptocinctus vizcainoi Friedrich, 1995, incluidos en la familia Sucocystidae por Friedrich (1993) y en Gyrocystidae por Sdzuy (1993). Gyrocystis sp. L se diferencia bien de las especies del género Elliptocinctus, entre otras cosas, por el número de placas marginales, once en E. vizcainoi Friedrich, doce en E. barrandei (Munier- Chalmas y Bergeron) y diez en Gyrocystis sp. L.
Existen características suficientes (como los grandes tubérculos adoperculares o el complejo portaopercular) para pensar que Gyrocystis sp. L podría tratarse de una especie nueva de Cincta, sin embargo, la escasez de material, así como la no conservación de algunas partes como la superficie ventral, aconsejan clasificarlo, de momento, en nomenclatura abierta.
Interpretación de las partes blandas
Entre los fósiles estudiados se pueden observar ciertas estructuras que pueden indicar la posición de algunas partes blandas en los organismos vivos, tales como músculos, ligamentos u órganos internos. El reconocimiento e interpretación de estas estructuras permite la reconstrucción de la anatomía interna, lo que ha supuesto en el estudio de los carpoideos un punto de discusión importante. En la cara interna de la unión entre las placas M5r y M4l existe una fuerte depresión, también presente en otros Cincta como por ejemplo Trochocystites bohemicus Barrande, 1887 o Sucocystis undata Friedrich, 1993, entre otros. Esta depresión en forma de embudo podría tratarse de una impresión debida a la inserción de los músculos de la faringe (cf. Domínguez, 1999). La cavidad corporal de los cincta estaría ocupada por una gran faringe que captaría las partículas nutritivas. La contracción de este paquete muscular comprimiría la faringe, deformaría la teca y expulsaría el agua a través del opérculo. La relajación de estos músculos permitiría la recuperación elástica de la teca y la succión de gran cantidad de agua para captar las partículas nutritivas (cf. Sdzuy, 1985; Friedrich, 1993; Domínguez, 1999).
Los márgenes laterales de las placas supracentrales no son rectos sino que presentan una acanaladura. Esta acanaladura podría estar ocupada en vida por ligamentos o algún tipo de tejido. De esta manera, el tegumento supracentral sería flexible y podría contraerse o distenderse en función de la acción de los músculos faríngeos y de la cantidad de agua absorbida. Esto estaría de acuerdo con la interpretación paleobiológica que sugieren autores previos (Sdzuy, 1985; Friedrich, 1993; Domínguez, 1999) para estos organismos.
En uno de los ejemplares estudiados (figuras 4.1 y 5.4) se observa el opérculo desarticulado. En la parte anterior, y coincidiendo con la parte que se articula con las placas postoperculares, se observa una acanaladura (faceta postopercular). Ésta podría estar ocupada en vida por un ligamento que permitiera el movimiento del opérculo. En este mismo ejemplar se observan facetas adoperculares en la base de los tubérculos adoperculares que podrían estar cubiertas en vida por ligamentos, permitiendo la articulación con el dintel (figura 4.1).
Interpretación funcional del opérculo
Los Cincta presentan tres aberturas principales en la teca (figura 6). La primera, que da paso a la cámara distal, está cubierta por una gran placa de morfología ovalada llamada opérculo y aparece localizada en el plano sagital, zona anterior, y sobre la placa M0. La segunda se sitúa en la parte superior anterior izquierda, al final de una pirámide compuesta por pequeñas plaquitas y la tercera en la parte frontal derecha entre las placas M1r y M2r. Existe una cuarta abertura situada en la unión de la supracentralia con el cinctus, en la parte superior derecha de la supracentralia tras la placa supraoral.
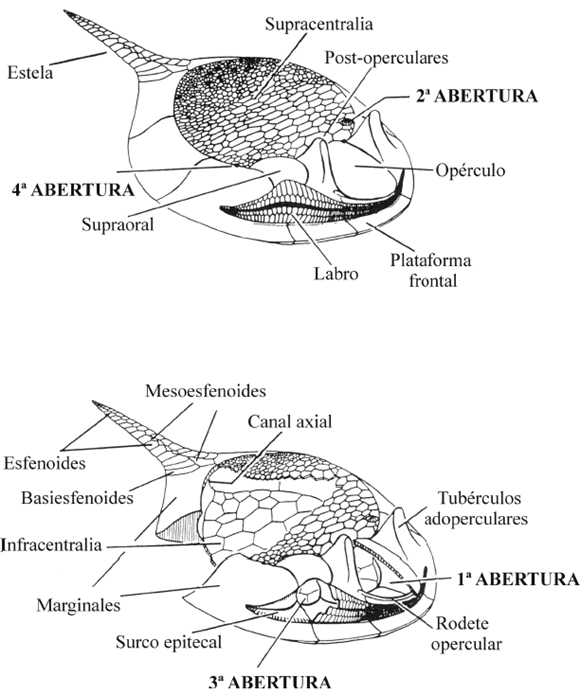
Figura 6. Dibujo de un Cincta donde aparece la posición de las principales aberturas en la teca. Modificado de Gil Cid y Domínguez Alonso (1995b) / location of main apertures in the theca of a schematic Cincta (modified from Gil Cid and Domínguez Alonso, 1995b).
La primera abertura se interpreta como el orificio anal (Ubaghs, 1967; Parsley, 1999), una hendidura branquial (Domínguez, 1999; Jefferies et al,. 1996) o como la abertura principal del atrio (Smith, 2005). Lo que parece estar claro es que la mayoría de los autores proponen que su función pudiera ser la de expeler el agua sobrante del interior de la teca (Friedrich, 1993; Sdzuy, 1993; Domínguez, 1999; Smith, 2005). La segunda, situada al final de una gran pirámide, es interpretada como una gonopirámide (Parsley, 1999) o una verdadera pirámide anal (Friedrich, 1993; Sdzuy, 1993; Domínguez, 1999; Smith, 2005). Esta estructura, en la mayoría de los casos, no se conserva porque durante el enterramiento las placas supracentrales colapsan sobre las infracentrales y se desarticula fácilmente en el proceso. No obstante, en ejemplares muy bien conservados (Ubaghs, 1967, p. 570, fig. 367-1; Sdzuy, 1993, p. 203, lám. 1, fig. 3) se observa que presenta una forma idéntica a las pirámides anales encontradas en diferentes grupos de equinodermos (Smith, 2005).
La mayoría de los autores están de acuerdo en que la boca coincidiría con la tercera abertura (Bather, 1913; Jaekel, 1900, 1918; Gislén, 1930; Nichols, 1972; Jefferies, 1990; Friedrich, 1993; Sdzuy, 1993; Domínguez, 1999; Parsley, 1999). Sobre la cuarta abertura, Smith (2005) plantea dudas incluso de su existencia, aunque para otros autores su existencia es real y se continúa interiormente con una serie de acanaladuras en la superficie interna del cinctus. Friedrich, (1993), Jefferies et. al. (1996) y Domínguez (1999) la consideran como un hidroporo-gonoporo.
Considerando que la función de la primera abertura es la de una válvula de descarga de agua (independientemente de que se trate de una hendidura branquial o de la abertura del atrio), y que la tercera abertura es la boca, la posición de vida de los Cincta sería con la cara inferior apoyada sobre el sustrato y la estela dirigida en dirección opuesta a las corrientes. La funcionalidad de esta disposición ha sido demostrada gracias a experimentos realizados en canales hidráulicos (Friedrich, 1993), en los que se observa cómo flujos de agua cargada de partículas se detienen al llegar a la supracentralia y las partículas en suspensión caen verticalmente, por pérdida de la capacidad de carga, hacia la zona anterior donde se encuentra la boca. Esta disposición frente a las corrientes y al sustrato sugiere que estos organismos eran filtradores, en los que parte del intestino funcionaba como una enorme faringe (Sdzuy, 1985; Domínguez, 1999; Smith, 2005), siendo su modo de vida análogo a los tunicados solitarios actuales (Sdzuy, 1985; Smith, 2005).
A grandes rasgos, el mecanismo de alimentación de los Cincta sería el de inhalar agua por la boca hasta que la faringe estuviera llena. Los Cincta más derivados, como el que se estudia en este trabajo, presentan un ambulacro izquierdo muy corto que segura- mente tuvo poca funcionalidad a la hora de captar la comida y llevarla hacia la boca. Sin embargo funcionaría de manera eficaz canalizando el flujo de agua hacia esta abertura. El labro, que se encarga de cubrir los ambulacros mediante una serie de filas de placas muy pequeñas, sería seguramente una estructura de protección que impediría el paso de grandes partículas hacia la boca que pudieran dañar al organismo. El aumento de presión en la faringe, causado por la compresión de la musculatura faríngea, produciría la abertura del opérculo y la expulsión del agua sobrante. En este mecanismo, la morfología del complejo portaopercular, compuesto por el opérculo, el porta, las facetas de los tubérculos adoperculares y el dintel, juega un papel fundamental.
El estudio, en este trabajo, de un Cincta con el complejo portaopercular extremadamente bien conservado, además de observaciones realizadas en material adicional en proceso de estudio y del material ya publicado por autores previos (principalmente Friedrich, 1993 y Sdzuy, 1993), permite hacer algunas aproximaciones al funcionamiento del opérculo.
En el ejemplar (MPZ 2006/16) el opérculo (figura 7.3) aparece en conexión anatómica. El dintel, compuesto por tres placas postoperculares, aparece unido a la parte posterior del opérculo de manera que el resto de éste queda libre. Las dos placas postoperculares laterales aparecen fuertemente unidas a los dos tubérculos adoperculares de las placas M1r y M1l. La foseta de la articulación (figura 5.4), situada en la cara interna de dichos tubérculos, es muy marcada, profunda y presenta una morfología ovalada. En este caso, el mecanismo de cierre del opérculo se produciría de manera similar al de una puerta, quedando la cámara distal totalmente cerrada. Esta descripción y función del complejo portaopercular sería válida, con diferentes modificaciones, para la mayoría de las especies conocidas de Cincta, pero no para los Cincta primitivos (Sdzuy, 1993).
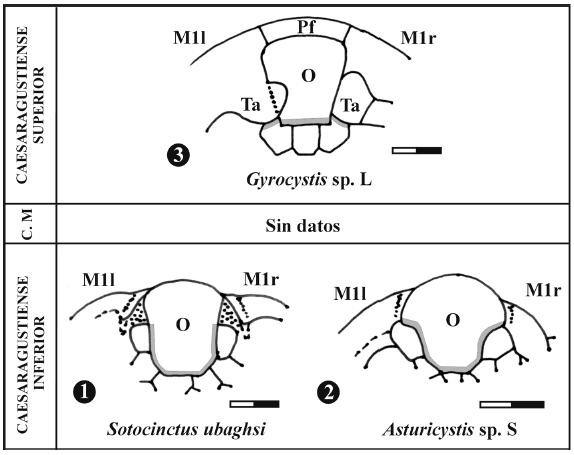
Figura 7. Interpretación de la articulación del dintel al opérculo. Ejemplares 1 y 2 correspondientes a Sotocinctus ubaghsi Sdzuy y Asturicystis sp. S (modificado de Sdzuy, 1993); escala = 2 mm. Ejemplar 3 correspondiente a Gyrocystis sp. L; escala = 1 mm. En color gris aparece la zona de articulación. Se observa cómo en los dos primeros la superficie articular ocupa la mitad del margen del opérculo por lo que esta estructura tendría una movilidad limitada. En el ejemplar 3, la articulación se restringe al margen posterior del opérculo con las tres placas postoperculares. Además las dos placas postoperculares laterales izquierda y derecha se articulan con los tubérculos adoperculares que presentan unas facetas adoperculares muy desarrolladas. Esto permitiría una gran movilidad del opérculo. O: Opérculo, Pf: Plataforma frontal, Ta: Tubérculo adopercular / interpretation of articulation of the dintel with the operculum. Cases shown in 1 and 2 belong respectively to Sotocinctus ubaghsi Sdzuy and Asturicystis sp. S (modified from Sdzuy, 1993); scale bar = 2 mm. Case shown in 3, to Gyrocystis sp. L; scale bar = 1 mm. The articulation area appears in grey colour. In 1 and 2, the articulation surface (or postopercular facet) runs along half of the margin of the operculum, which in consequence must have had only limited mobility. In 3, the articulation surface is restricted to the rear margin of the operculum with the three postopercular plates; moreover, the right and left postopercular plates articulate with the respective adopercular processes (which show pronounced adopercular factes). Such constructional design of the operculum must have provided a much higher mobility to it. O: operculum; Pf: outer edge of the cinctus ("marginale Lippe"); Ta: adopercular process ("suroperculare Fortsätze").
Sdzuy (1993) realiza un análisis muy detallado del opérculo en Asturicystis Sdzuy (figura 7.2) y Sotocinctus Sdzuy (figura 7.1), los cincta más antiguos conocidos hasta el momento. Ambos géneros presentan un opérculo que aparece unido a las placas postoperculares en la parte anterior y gran parte del lateral (Sdzuy, 1993, text-figs. 1, 2). En estos casos la unión se hace mediante, al menos, seis placas postoperculares y no tres como en el caso estudiado. Esto hace que los opérculos en los Cincta más antiguos tengan una menor libertad de movimiento, ya que aparecen articulados en una mayor porción. Sdzuy (1993) soluciona la falta de movilidad sugiriendo que la supracentralia debería ser flexible para permitir la apertura del opérculo.
En el Cincta MPZ 2006/16 parece ser que el opérculo, al estar articulado únicamente en su parte posterior, presentaría una mayor libertad de movimiento que en Asturicystis o Sotocinctus. Esto seguramente se traduciría en una mayor movilidad del opérculo y un incremento de las tensiones sobre el dintel. Para soportarlas, este cincta desarrolla un dintel que parece ser muy rígido, además de fuertes articulaciones de las placas postoperculares laterales con los tubérculos adoperculares.
Una vez comparados los complejos portaoperculares de los diferentes Cincta, se puede suponer que la reducción del número de placas postoperculares ancladas al opérculo, el desarrollo de un dintel más rígido y la presencia de grandes fosetas articulares en la cara interna de tubérculos adoperculares, que son extremadamente desarrollados, podrían estar relacionados con una mayor movilidad del opérculo y el soporte de la tensión asociada a dicho movimiento. Todas estas características permitirían una mayor eficacia en el proceso de expeler el agua y podrían ser consideradas como apomorfías en la historia evolutiva de la clase Cincta.
Conclusiones
El estudio de nuevo material de Cincta procedente de Los Barrios de Luna (Cordillera Cantábrica, norte de España) permite identificar la presencia del género Gyrocystis en el Cámbrico Medio (Languedociense inferior) de la Formación Oville. Tras el estudio de la morfología funcional del complejo portaopercular, se concluye que Gyrocystis sp. L presentaba un movimiento del opérculo muy eficaz. Éste se conseguía mediante una articulación restringida a la parte posterior del opérculo (únicamente en tres placas postoperculares), el desarrollo de un dintel rígido y la presencia de grandes fosetas articulares en la cara interna de los tubérculos adoperculares. Todas estas características permitirían una mayor eficacia en el proceso de expeler el agua y podrían ser consideradas como apomorfías en la historia evolutiva de la clase Cincta. No se descarta que tal configuración desembocara, con el tiempo, en una función de propulsión a chorro (ya planteada por Bonik et al., 1978), un mecanismo ya propuesto por Domínguez (1999) para los carpoideos ctenocistoideos.
Agradecimientos
Nuestro más sincero agradecimiento a R.P.S. Jefferies (Natural History Museum, Londres), siempre dispuesto a discutir numerosos aspectos sobre los cincta y a enseñarnos a mirar más allá de su extraña morfología externa. A D.F. Gracia Sevilla que donó amablemente los cincta estudiados en este trabajo al Museo Paleontológico de Zaragoza (Universidad-Gobierno de Aragón). Un especial agradecimiento a los revisores B. Lejebvre (Université Claude Bernard, Lyon, Francia) y R. Haude (Universität Göttingen, Alemania), cuyos comentarios y sugerencias han ayudado a mejorar notablemente este manuscrito. A T.M. Sánchez por sus correcciones editoriales pertinentes. Este trabajo es una contribución al Proyecto Consolíder CGL2006-12975/BTE ("MURERO") del Ministerio de Educación y Ciencia de España, y al Grupo Consolidado E-17 ("Patrimonio y Museo Paleontológico") de la Consejería de Ciencia, Tecnología y Universidad del Gobierno de Aragón. S. Zamora goza de una beca predoctoral financiada por el Departamento de Ciencia, Tecnología y Universidad del gobierno de Aragón. Se agradece también al servicio de fotografía paleontológica de la Universidad de Zaragoza.
Bibliografía
1. Aramburu, C., Méndez-Bedia, I., Arbizu, M. y García-López, S. 2004. La secuencia preorogénica. En: J.A. Vera (ed.), Geología de España, SGE-IGME, Madrid: 27-34. [ Links ]
2. Aramburu, C., Truyols, J., Arbizu, M., Méndez Bedia, I., Zamarreño, I., García Ramos, J.C., Suárez de Centi, C. y Valenzuela, M. 1992. El Paleozoico Inferior de la Zona Cantábrica. En: J.C. Gutiérrez Marco, J. Saavedra e I. Rábano (eds.), El Paleozoico Inferior de Ibero-América, Universidad de Extremadura: 397-421. [ Links ]
3. Barrande, J. 1859. État actuel des connaissances acquises sur la fauna primordiale. Bulletin Societé Géologique de la France, 2: 516-546. [ Links ]
4. Barrande, J. 1887. Système Silurien du centre de la Bohème. Vol. VII. Classe des échinodermes, ordre des Cystidées, Praga-Leipzig: 233 pp. [ Links ]
5. Bather, F.A. 1913. Caradocian Cystidea from Girvan. Royal Society of Edinburgh, Transactions XLIX, 2: 359-521. [ Links ]
6. Bonik, K., Gutmann, W.F. y Haude, R. 1978. Stachelhäuter mit Kiemen-Aparat: Der Beleg für die Ableitung der Echinodermen von Chordatieren.- Natur u. Museum 108: 211-214. [ Links ]
7. Cabibel, J., Termier, H. y Termier, G. 1959. Les echinodermes mesocambriens de la Montagne Noire (sud de la France). Annales de Paléontologie 44: 281-294. [ Links ]
8. David, B., Lefebvre, B., Mooi, R. y Parsley, R. 2000. Are homalozoans echinoderms? An answer from the extraxial-axial theory. Paleobiology 26: 529-555. [ Links ]
9. Domínguez, P. 1999. Early evolution of echinoderms: Class Ctenocystoidea revisited. En: M.D. Candia Carnevali y F. Bonasoro (eds.), Echinoderm Research. A. A. Balkema, Rotterdam, pp. 263-268. [ Links ]
10. Domínguez, P. y Jefferies, R.P.S. 2005. A Cladogram for the Deuterostomia based on molecular-biological and fossil evidence. En: Abstracts of the 53º Symposium on vertebrate palaeontology and comparative anatomy, p. 30. [ Links ]
11. Fatka, O. y Kordule, V. 2001. Asturicystis havliceki sp. nov. (Echinodermata, Homostelea) from the Middle Cambrian of Bohemia (Barrandian area, Czech Republic). Casopis Ceské Geologické Spolecnosti (=Journal of the Czech Geological Society), 46: 189-193. [ Links ]
12. Friedrich, W.P. 1993. Systematik und Funktionsmorphologie mittelkambrischer Cincta (Carpoidea, Echinodermata). Beringeria 7: 3-190. [ Links ]
13. Friedrich, W.P. 1995. Neue Nachweise mittelkambrischer Cincta (Carpoidea, Echinodermata) aus Marokko, Sardinien und Süd-Wales. En: Morocco'95-The Lower-Middle Cambrian standard of western Gondwana; introduction, field guide, abstracts, and proceedings of the First conference of the Lower Cambrian Stage Subdivision Working Group and I.G.C.P. Project 366 Ecological Aspects of the Cambrian Radiation. G. Geyer y E. Landing (eds.), Beringeria, Sonderheft 2: 255-269. [ Links ]
14. Gil Cid, M.D. y Domínguez Alonso, P. 1995a. Gyrocystis cruzae, una nueva especie de Cincta (Echinodermata Carpoidea) del Cambrico Medio del Ferredal de Quintana (Asturias, España). Boletín Geológico y Minero 106: 517-531. [ Links ]
15. Gil Cid, M.D. y Domínguez Alonso, P. 1995b. Presencia de Gyrocystis Jaekel,1918 en el Cámbrico Medio de Zafra (Badajoz). Revista de la Sociedad Geológica de España 8: 99-110. [ Links ]
16. Gil Cid M.D. y Domínguez Alonso, P. 2002. Ubaghsicystis segurae nov. gen. y sp., nuevo Eocrinoide (Echinodermata) del Cámbrico Medio del Norte de España. Coloquios de Paleontología 53: 21-32 [ Links ]
17. Gislén, T. 1927. A new Spanish carpoid. Arkiv zoologi 19 B(2): 1-3. [ Links ]
18. Gislén, T. 1930. Affinities between the Echinodermata, Enteropneusta, and Chordonia. Zoologiska Bidrag från Uppsala. 12: 199-304. [ Links ]
19. Gómez de Llarena, J. y Rodríguez Arango, C. 1948. Datos para el estudio geológico de la Babia Baja (León). Boletín del Instituto Geológico y Minero de España 61: 79-206. [ Links ]
20. Jaekel, O. 1900. Über Carpoideen: eine neue Klasse von Pelmatozoen. Zeitschrift der Deutschen Geologischen Gesellschaft 52: 661-677. [ Links ]
21. Jaekel, O. 1918. Phylogenie und System der Pelmatozoen. Paläontologische Zeitschrift 3: 1-128. [ Links ]
22. Jefferies, R.P.S. 1990. The solute Dendrocystoides scoticus from the upper Ordovician of Scotland and the ancestry of chordates and echinoderms. Palaeontology 33: 631-679. [ Links ]
23. Jefferies, R.P.S., Brown, N. y Daley, P. 1996. The early phylogeny of chordates and echinoderms and the origin of chordate leftright asymmetry and bilateral symmetry. Acta Zoologica (Stockholm) 77-2: 101-122. [ Links ]
24. Lotze, F. y Sdzuy, K. 1961. Das Kambrium Spaniens. Teil I: Stratigraphie. Akademie der Wissenschaften und der Literatur, Abhandlungen der mathematisch-naturwissenschaftlichen Klasse, 1961 (6): 283-498; Teil II: Trilobiten. Akademie der Wissenschaften und der Literatur, Abhandlungen der mathematisch- naturwissenschaftlichen Klasse, 1961: 499-690. [ Links ]
25. Meléndez, B. 1952. Los Carpoideos de España. Las Ciencias 17: 497-516. [ Links ]
26. Munier-Chalmas, M. y Bergeron, J. 1889. En: J. Bergeron, Étude géologique du Massif ancien situé au Sud du Plateau central. Annales de la Société Géologique du Nord 22: 338 pp. Paris. [ Links ]
27. Nichols, D. 1972. The water-vascular system in living and fossil echinoderms. Palaeontology 15: 519-538. [ Links ]
28. Parsley, R.L. 1999. The Cincta (Homostelea) as blastozoans. En: M.D. Candia Carnevali y F. Bonasoro (eds.), Echinoderm Research. A. A. Balkema, Rotterdam, pp. 369-375. [ Links ]
29. Prado, C., de Verneuil, E. y Barrande, J. 1860. Sur l'existence de la faune primordiale de la chaîne cantabrique. Bulletin de la Societé Géologique de la France 2, 17: 516-554. [ Links ]
30. Schröeder, R. 1973. Carpoideen aus dem Mittelkambrium Nordspaniens. Palaeontographica, Abteilung A, 141: 119-142. [ Links ]
31. Sdzuy, K. 1958. Neue Trilobiten aus dem Mittelkambrium von Spanien. Senckenbergiana lethaea 39: 235-253. [ Links ]
32. Sdzuy, K. 1969. Bioestratigrafía de la Griotte Cámbrica de Los Barrios de Luna (León) y de otras sucesiones comparables. Trabajos de Geología, Universidad de Oviedo 2 (año 1968): 45-58. [ Links ]
33. Sdzuy, K. 1985. La morfología de carpoideos del orden Cincta. 1º Jornadas de Paleontología (Zaragoza): 51. [ Links ]
34. Sdzuy, K. 1993. Early Cincta (Carpoidea) from the Middle Cambrian of Spain. Beringeria 8: 189-207. [ Links ]
35. Sdzuy, K. y Liñán, E. 1993. Rasgos paleogeográficos del Cámbrico Inferior y Medio del Norte de España. Cuadernos del Laboratorio Xeolóxico de Laxe 18: 189-215. [ Links ]
36. Smith, A. B. 2005. The pre-radial history of echinoderms. Geological Journal 40: 255-280. [ Links ]
37. Termier, H. y Termier, G. 1973. Les Échinodermes Cincta du Cambrien de la Montagne Noire (France). Geobios 6: 243-266. [ Links ]
38. Ubaghs, G. 1967. Homostelea. En: R.C. Moore (ed.), Treatise on Invertebrate Paleontology. Echinodemata 1: S565-S581. New York, Geological Society of America and University of Kansas Press. [ Links ]
39. Ubaghs, G. 1975. Early Palaeozoic echinoderms. Reviews in Earth and Planetary Science 3: 79-98. [ Links ]
40. Zamarreño, I. 1972. Las litofacies carbonatadas del Cámbrico de la zona cántábrica (NW España) y su distribución paleogeográfica. Trabajos de Geología, Universidad de Oviedo 5: 1-118. [ Links ]
41. Zamarreño, I. 1983. El Cámbrico en el Macizo Ibérico. En: Libro Jubilar J.M. Ríos. IGME, Madrid. Geología de España. 1: 117-191. [ Links ]
Recibido: 30 de agosto de 2006.
Aceptado: 18 de julio de 2007.

