










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.45 n.1 Buenos Aires ene./mar. 2008
Ontogeny, structure and moulting of Parabolina frequens argentina (Kayser) (Trilobita, Olenidae) from the Furongian of northwestern Argentina
M. Franco Tortello1 and Euan N.K. Clarkson2
1Departamento Paleozoología Invertebrados, Museo de Ciencias Naturales, Paseo del Bosque s/n°, 1900 La Plata, Argentina. tortello@museo.fcnym.unlp.edu.ar
2Grant Institute, School of Geosciences, University of Edinburgh, West Mains Road, Edinburgh EH9 3JW, United Kingdom. Euan.Clarkson@glg.ed.ac.uk
Abstract. Parabolina frequens argentina (Kayser) (Trilobita, Olenidae) is a guide fossil occurring in different assemblages and lithologies of the upper Cambrian (Furongian) of northwestern Argentina. The black and dark grey shales of the Lampazar Formation at sierra de Cajas and the Santa Rosita Formation (lower part) at Iruya and Alfarcito (Jujuy and Salta provinces) contain numerous well-preserved specimens at different stages of development. The juveniles represent anaprotaspis, metaprotaspis and degrees 0-11 meraspis. The latter exhibit a high variability related with the pattern of glabellar furrows and lobes, the sagittal length of the thoracic segments, the splay and length of the genal and thoracic spines, the relative width of the axis, and the relative development of the axial tubercles. As in other species of Parabolina, the meraspid stages of P. frequens argentina do not form well-defined instar groupings. Besides, in the adult specimens the length of the anterior cephalic border and the preglabellar field are also variable characters. The hypostome is characterized by having almost completely reduced anterior wings; it lies in close association with the ventral doublure (conterminant condition). Although some authors suggested that P. frequens argentina and P. frequens frequens (Barrande) could be synonyms, these taxa differ in having distinctive pygidia and hypostomes. Many exoskeletal configurations can be interpreted as exuviae. From a functional point of view, it is likely that the genal and macropleural spines on the 8th thoracic segment supported the body of the trilobite when resting on the sea floor. The exoskeleton can be reconstructed in two alternative postures, the "alert" and "relaxed" attitudes.
Resumen. Ontogenia, Estructura Y Muda De Parabolina Frequens Argentina (Kayser) (Trilobita, Olenidae) Del Furongiano Del Noroeste Argentino. Parabolina frequens argentina (Kayser) (Trilobita, Olenidae) se encuentra ampliamente representado en el Cámbrico superior (Furongiano) del noroeste argentino, en distintas asociaciones y litologías, constituyendo un elemento valioso para datar afloramientos y establecer correlaciones. Las lutitas oscuras de la Formación Lampazar en la sierra de Cajas y de la Formación Santa Rosita (parte inferior) en Iruya y Alfarcito (provincias de Jujuy y Salta, Argentina) contienen numerosos ejemplares bien preservados, en diferentes fases de desarrollo. Los especímenes juveniles representan estadios anaprotaspis, metaprotaspis y meráspidos 0-11. Estos últimos exhiben una alta variabilidad relacionada con el grado de expresión de los surcos laterales de la glabela y lóbulos glabelares, la longitud exsagital de los segmentos torácicos, la curvatura y longitud de las espinas genales y torácicas, la anchura del axis y el desarrollo relativo de los nodos axiales. Como se ha documentado en otras especies de Parabolina, los estadios meráspidos de P. frequens argentina no conforman grupos de tamaño (clusters) bien definidos. Por su parte, los ejemplares adultos presentan un borde cefálico anterior y un campo preglabelar de longitudes algo variables. El hipostoma se dispone en contacto con el doblez cefálico anterior (condición coincidente) y posee alas anteriores muy reducidas. Aunque algunos autores consideran a P. frequens argentina y P. frequens frequens (Barrande) como sinónimos, estos taxones se distinguen por presentar pigidios e hipostomas distintivos. Numerosos especímenes con configuraciones particulares representan exuvias generadas durante el proceso de muda. Con relación al hábito de vida, las espinas genales y las macroespinas del octavo segmento torácico habrían constituido los principales puntos de apoyo del organismo sobre el sustrato. El exoesqueleto puede ser reconstruido en dos posturas alternativas, activa y de reposo.
Key words. Parabolina; Trilobita; Olenidae; Furongian; Argentina; Ontogeny; Morphology; Ecdysis.
Palabras clave. Parabolina; Trilobita; Olenidae; Furongiano; Argentina; Ontogenia; Morfología; Ecdisis.
Introduction
The first conspicuous lower Paleozoic fossils in South America were collected in Tincuya (southern Bolivia) and described by Kayser in 1876 ( "Agnostus" tilcuyensis Kayser (= Trilobagnostus tilcuyensis), "Arionellus" lorentzi Kayser and "A". hyeronimi Kayser (=Angelina hyeronimi), and "Olenus" argentinus Kayser (=Parabolina frequens argentina)). On the basis of their affinities with species from Scandinavia and England, Kayser (1876) originally assigned these trilobites to the Late Cambrian, indicating that P. frequens argentina was the commonest taxon in the assemblage.
During the 1910's, 1930's and 1940's, P. frequens argentina was successively documented in other localities of southern Bolivia and northwestern Argentina (Salitre, Tilcara, río Volcancito; Steinmann and Hoek, 1912; Kobayashi, 1937; Harrington, 1938; Harrington and Leanza, 1943). Finally, Harrington and Leanza (1957) provided a complete review on the morphology and synonymy of this taxon, showing that it is widely distributed in the Cordillera Oriental (Jujuy and Salta Provinces), as well as in the Famatina System (La Rioja Province). Since P. frequens argentina is a common fossil occurring in different assemblages and lithologies, it has proved to have great biostratigraphic value for establishing correlations within the latest Cambrian of the "Baltic Province" (Shergold, 1988) (e.g., Harrington and Leanza, 1957; Frederickson, 1958; Aceñolaza, 1992).
Sierra de Cajas (Aceñolaza, 1968), Iruya (Vilela, 1960) and Alfarcito (Harrington and Leanza, 1957) are representative Furongian localities of Salta and Jujuy, which contain numerous well-preserved specimens of P. frequens argentina at different stages of development. The aim of this paper is to describe the ontogeny of this subspecies, including some aspects of its holaspid morphology which, as originally indicated by Harrington and Leanza (1957), is highly variable. In addition, the record of different exoskeletal configurations provides interesting paleobiological data, related to possible techniques of ecdysis and mode of life of the taxon.
Stratigraphy and localities
Harrington and Leanza (1957) erected the Parabolina frequens argentina Biozone on the basis of numerous trilobites dominated by olenids and agnostoids. The unit is represented in many successions in the Cordillera Oriental, comprising the Lampazar, Casa Colorada and Santa Rosita (lower part) formations and equivalents. Though Harrington and Leanza (1957) originally referred the whole biozone to the early Tremadoc, subsequent studies demonstrated that its lower part is assignable to the Furongian (e.g., Rushton, 1982, p. 46; Ludvigsen, 1982, p. 150; Aceñolaza, 1983; Salfity et al., 1984; Tortello, 2003). Parabolina frequens argentina is particularly abundant in the lower part of the unit, below the first appearance of the lowest Ordovician Jujuyaspis keideli Kobayashi (Trilobita, Olenidae) and Rhabdinopora Eichwald (Graptolithina) (see Benedetto, 1977; Aceñolaza, 1983; Aceñolaza and Aceñolaza, 1992; Ortega and Rao, 1995; Tortello and Esteban, 1998, 1999; Tortello et al., 1999; Tortello, 2003).
The P. frequens argentina Biozone is mainly composed of shales, sandstones and rare interspersed marls and limestone layers, representing a wide range of sedimentary environments, including wavedominated and open marine conditions. Those sections representing deep outer shelf settings, with conditions of low energy, deposition of fine sediments and abundant organic matter, generally contain well-preserved fossils. In these settings P. frequens argentina commonly occurs associated with agnostoids (Lotagnostus Whitehouse, Gymnagnostus Robison and Pantoja-Alor, Pseudorhaptagnostus (Machairagnostus) Harrington and Leanza, Micragnostus Howell) and olenids (e.g., Parabolinella Kobayashi, Plicatolina Shaw, Beltella Lake, Angelina Salter), which comprise benthic species especially adapted to live in environments low in oxygen (Parabolinella, Plicatolina). Tortello (2003) described the Pseudorhaptagnostus (Machairagnostus)-Gymnagnostus Subzone (latest Furongian) in such a facies (="Olenid facies" of Fortey, 1975), and designated a composite stratotype that encloses the Quebrada Azul (Lampazar Formation, sierra de Cajas, Jujuy) (Aceñolaza, 1968; Tortello and Esteban, 2003).
Most specimens studied herein come from the Lampazar Formation outcropping at the Quebrada Azul, 5 km East of El Molino-Mina Aguilar (Humahuaca Department, Jujuy Province) (figure 1). This 180 m thick section represents an outer shelf setting with some transitions to shallower environments. Oxygen and energy levels were not uniform during deposition: fine grained sediments vary from laminated black and dark grey shales in the lower part of the formation to laminated greenish shales with intercalations of siltstones and sandstones in the middle and upper part (Tortello and Esteban, 2003). It lies on the quartzites of the Padrioc Formation (Late Cambrian) and is overlain by the sandstones, quartzites and interbedded shales and limestones of the Cardonal Formation (Aceñolaza, 1968). The columnar section showing the stratigraphic range of the trilobite faunas ( Gymnagnostus perinflatus (Harrington and Leanza), G. bolivianus (Hoek), Pseudorhaptagnostus (Machairagnostus) tmetus Harrington and Leanza, P. (M.) corrugatus (Suárez-Soruco), Micragnostus vilonii Harrington and Leanza, Parabolina frequens argentina (Kayser), Parabolinella coelatifrons Harrington and Leanza, Angelina hyeronimi (Kayser), Beltella ulrichi (Kayser), Plicatolina scalpta Harrington and Leanza, Akoldinioidia sp. and Asaphellus cf. aspinus Robison and Pantoja-Alor] is shown in figure 2. Parabolina frequens argentina largely dominates the assemblages, representing more than 80 per cent of the total fauna.

Figure 1. Location map / mapa de ubicación.
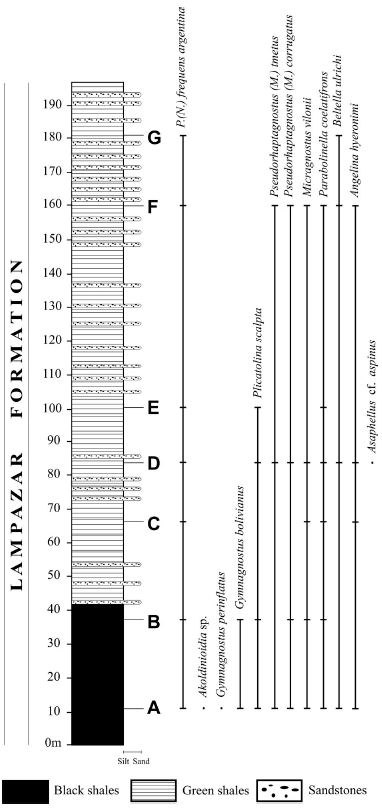
Figure 2. Columnar section of the Lampazar Formation (Furongian) at Quebrada Azul (Cajas Range), showing fossiliferous levels (A-G) and distributions of Parabolina frequens argentina and associated trilobites (after Tortello and Esteban, 2003) / columna estratigráfica de la Formación Lampazar (Furongiano) en la quebrada Azul (Sierra de Cajas), mostrando los niveles fosilíferos (A-G) y las distribuciones de Parabolina frequens argentina y trilobites asociados (basado en Tortello y Esteban, 2003).
Additional material was collected from the lower San Isidro River, 3 km north-east of Iruya Town, Salta Province (figure 1). This outcrop is mainly composed of dark shales assigned to the Santa Rosita Formation (lower part), interposed between the quartzites of the Mesón Group (Cambrian) and the sandstones of the Salta Group (Cretaceous-Paleogene) (Vilela, 1960; Turner, 1964; Turner and Mon, 1979). In this locality Parabolina frequens argentina is documented in association with the trilobites Angelina hyeronimi (Kayser), Plicatolina sp., Lotagnostus (Semagnostus) sp. and Micragnostus cf. vilonii Harrington and Leanza, and the hyolithid Tajinella? iruyensis Pagani, Sabattini and Tortello (see Pagani et al., 2005). Finally, some specimens come from the Alfarcito region, about 8 km east of Tilcara Town, Jujuy Province. In this locality the latest Cambrian is represented by the Casa Colorada Formation, which mainly consists of shales containing P. frequens argentina associated with Parabolinella, Plicatolina, and Onychopyge Harrington. The unit is interposed between the Mesón Group and the sandstones and siltstones of the Alfarcito Fm. (lower Tremadoc). Detailed information on this locality was provided by Harrington and Leanza (1957, p. 6-7) and Zeballo and Tortello (2005).
Description of the ontogeny of Parabolina frequens argentina
The descriptive methods used in this work are similar to those applied by Berard et al. (2000) and Tortello and Clarkson (2003) for specimens preserved in shales. Many latex replicas of external and internal moulds were prepared. For light macrophotography, the late meraspid and holaspid specimens were coated with magnesium oxide, whereas latex replicas of the early meraspides with gold-palladium coating were examined under SEM. Based on techniques by Clarkson and Taylor (1995a), Berard et al. (2000) and Clarkson and Ahlberg (2002), the reconstructions of the smaller stages were made from the SEM photographs by drawing a squared grid on each print and transposing this on to millimetre-squared graph paper to give a constant scale.
The material is housed in the Museo de Ciencias Naturales de La Plata (MLP), and the Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán (PIL), Argentina.
Order PTYCHOPARIIDA Swinnerton, 1915
Suborder OLENINA Burmeister, 1843
Family OLENIDAE Burmeister, 1843
Subfamily Oleninae Burmeister, 1843Genus Parabolina Salter, 1849
Subgenus P. (Neoparabolina) Nikolaisen and Henningsmoen, 1985
Type species. Parabolina frequens (Barrande, 1868), original designation.
Parabolina (Neoparabolina) frequens (Barrande, 1868)
Remarks. Following Pribyl and Vanek (1980), Parabolina argentina (Kayser, 1876) is regarded as a subspecies of the earlier described species Parabolina frequens (Barrande, 1868) (see below, Systematic position).
Parabolina (Neoparabolina) frequens argentina (Kayser, 1876) Figures 3-11
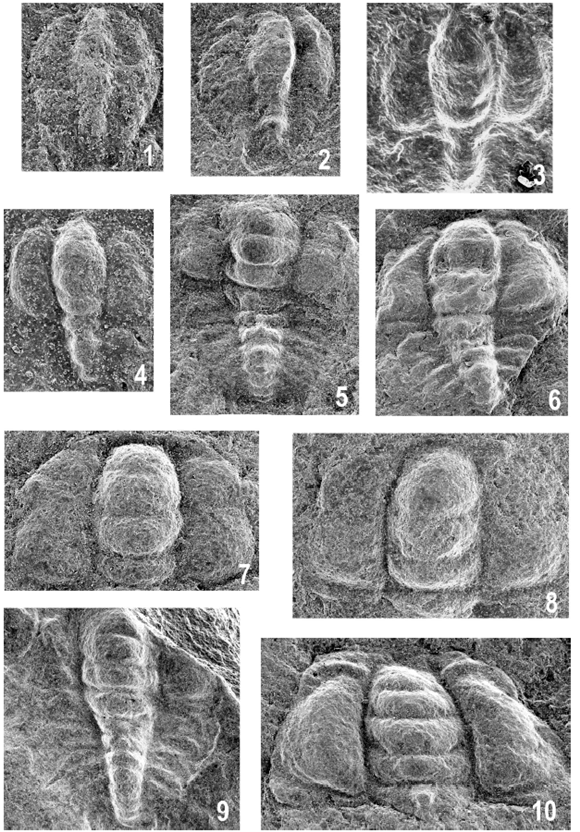
Figure 3. Parabolina frequens argentina (Kayser), from Quebrada Azul (Cajas Range), bed A / de la quebrada Azul (sierra de Cajas), nivel A. 1, ?Anaprotaspid stage / ?anaprotaspis, MLP 31577, x176. 2, Metaprotaspid stage / metaprotaspis, MLP 31580, x121. 3-4, Degree 0 meraspid specimens lacking librigenae / meráspidos 0 (sin librígenas); 3, MLP 31579a, x153; 4, MLP 31578a, x134. 5-8, Degree 1 meraspides / meráspidos 1; 5, Axial shield / escudo axial, MLP 31581, x80; 6, MLP 31582, axial shield slightly flattened / escudo axial suavemente deformado, x85; 7, Cranidium / cranidio, MLP 31583, x112; 8, Cranidium / cranidio, MLP 31584, x108. 9, Degree 2 meraspis, almost complete specimen damaged anteriorly / meráspido 2, ejemplar casi completo, mal preservado anteriormente, MLP 31579b, x70. 10, Probable degree 3-5 meraspis, cranidium / probable meráspido 3-5, cranidio, MLP 31585, x82.
1876. Olenus argentinus sp. nov. Kayser: 6, pl. 1, figs. 1-3.
1957. Parabolina argentina (Kayser). Harrington and Leanza: 81-85, figs. 25-26 (see for synonymy).
1957. Parabolina argentina (Kayser). Henningsmoen: 116-117 (see for synonymy).
1958. Parabolina argentina (Kayser). Frederickson: 541-543, pl. 80.
1965. Parabolina argentina (Kayser). Branisa: pl. 1, fig. 5.
1965. Parabolina sp. Branisa: pl. 1, fig. 9.
?1968. Parabolina cf. P. argentina (Kayser). Robison and Pantoja- Alor: 788, pl. 101, figs. 25-26.
1980. Parabolina frequens argentina (Kayser). Pribyl and Vanek: 14- 15, pls. 2-5 (see for synonymy).
2000. Parabolina (Neoparabolina) frequens argentina (Kayser). Tortello and Rao: 69, figs. 3.D-I.
2003. Parabolina (Neoparabolina) frequens argentina (Kayser). Tortello and Esteban: 338-340, figs. 5.G-I.
2005. Parabolina (Neoparabolina) frequens argentina (Kayser). Zeballo and Tortello: 134-135, figs. 4.D-E, K; 6.A.
Protaspid stages
We follow here Størmer's (1942, p. 56) division of the protaspid period of growth into anaprotaspid and metaprotaspid stages as used in Clarkson and Taylor (1995a) and Clarkson et al. (1997).
Two protaspids are present in our material. None of these specimens, however, is especially well preserved and some inferences have had to be made in the reconstruction. There is a rather large size gap between the first and second protaspid stages described below, so an intermediate instar is probably missing.
An ?anaprotaspis is a cambered disc, ovoid in form, 0.25 mm long (sag.) and 0.2 mm wide, slightly tapering posteriorly (figures 3.1, 4.1). A short intergenal spine is present on the right side. The axis is raised above the genal regions; it is long and narrow, less than 0.1 mm across, tapering anteriorly and posteriorly, with no clear indication of segmentation.
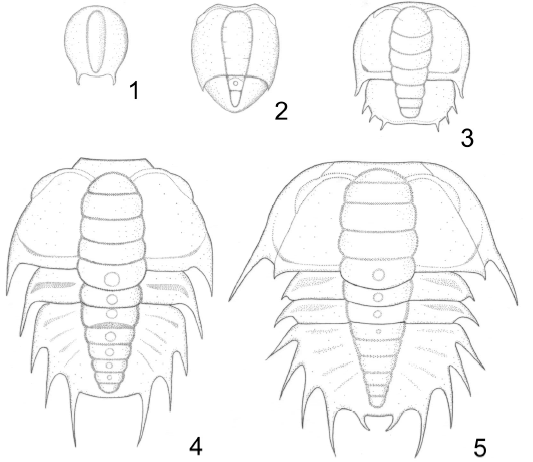
Figure 4. Parabolina frequens argentina (Kayser), protaspid and early meraspid stages, reconstructed from specimens in figure 3 / protáspidos y meráspidos tempranos; reconstrucciones basadas en los especímenes ilustrados en la figura 3. 1, Anaprotaspis. 2, Metaprotaspis. 3, Degree 0 meraspis / meráspido 0. 4, Degree 1 meraspis / meráspido 1. 5, Degree 2 meraspis / meráspido 2. All specimens x76 / todos los especímenes x76.
In addition, a slightly distorted metaprotaspis, 0.38 mm long (sag.) and 0.30 mm wide, is illustrated in figures 3.2, 4.2. It is somewhat pyriform in shape, distinctly tapering posteriorly. At 0.26 mm from the anterior border there is a distinctive transverse furrow, indicating the boundary between the cephalon from the protopygidium; the latter is not yet separated nor released. The protoglabella is elongated-barrel shaped, with indistinct traces of furrowing, and there is some trace of eye ridges and palpebral lobes at the antero-lateral margins. The transitory pygidium has a node on the first axial ring, and a smooth ring behind. The posterior margin is rounded and, in this specimen, no traces of spines are visible.
Meraspid stages
The specimens studied include axial shields with 0, 1, 2, 4… thoracic segments, as well as isolated tagmata. The traditional descriptive methodology by Raw (1925), consisting of the division of the meraspid period into degrees (1, 2, 3, etc.), marked by the addition of successive segments to the thorax, can be applied here.
Degree 0 meraspis. The best preserved complete individual representing this stage is an external mould illustrated in figure 3.3 (see also figure 4.3). As reconstructed, it is 0.44 mm in length and 0.40 mm at its widest point. The cephalon is 0.26 mm long (sag.), subcircular in outline, with a distinct flattened border and straight intergenal spines. Glabella occupying central third of the cranidium, slightly wider at its mid point, reaching anterior border, well defined by deep and narrow axial furrows. In this specimen S0 is deeply impressed, transglabellar, curved backward. S1 impressed laterally and oblique backward, fainter medially. S2 appears to be transglabellar but indistinct. S3 poorly defined. Occipital ring slightly narrower (tr.) than L0, smooth. Fixigenae quite inflated, with faintly defined palpebral lobes.
Transitory pygidium 0.15 mm long (sag.) and 0.30 mm wide, curled down posteriorly, semicircular in outline. Axis 0.08 mm across at its widest point, convex, tapering posteriorly, surrounded by narrow axial furrows; it consists of three segments lacking central nodes. Pygidial margin with three pairs of short spines.
A second, distorted specimen (figure 3.4) is of similar size and form. Likewise specimen illustrated in figure 3.3, it can be used to confirm the slightly barrel-shaped glabella and distinct intergenal spines, as well as the overall proportions described above.
Degree 1 meraspis. Two complete specimens and several other disarticulated or distorted individuals are assignable to this growth stage. The reconstruction (figure 4.4) is based on the specimen illustrated in figure 3.5, but some details have been added from other specimens (figures 3.6-3.8). As reconstructed this degree is 0.82 mm long (excluding spines) and 0.72 mm across at its widest point. Cephalic contour originally semicircular with a transverse anterior border. Glabella 0. 20 mm wide, long, rounded anteriorly, with subparallel lateral margins, well defined by deep and narrow axial furrows. Lateral furrows (S0, S1, S2, S3) subequally spaced, transverse to slightly curved backwards, each swollen medially, defining lobes L1-L4. S0 deeper and wider (sag.) than S1-S3. Occipital ring of similar length (sag.) and appearance to glabellar lobes, extending backwards over the anterior part of the single thoracic segment and provided with a medial node. Posterior border furrow deep and well defined. Fixigenae quite inflated, with a pair of ocular ridges curving anteriorly, and faintly defined palpebral lobes. Intergenal spines illustrated in the reconstruction are hypothetical. Librigenae and hypostome unknown.
Thoracic segment with a short (sag.) axial ring, which is partially covered by the projecting occipital ring, with a conspicuous median node. Pleura long (exsag.), about two thirds the total width of the thorax, with obliquely truncated outer terminations, each with a narrow spine. Pleural furrow slightly oblique, reaching the outer margin.
Transitory pygidium semicircular, 0.52 mm long (sag.) and 0.55 mm wide (excluding spines). It is slightly curled down posteriorly. Axis very convex, raised above the pleural fields, less than two thirds the total width of the pygidium, tapering backward; it is composed of six axial rings, each with a median node. In this specimen the rear edge of the anterior axial ring is slightly separated from the articulating half-ring of the succeeding segment, interpreted as a stage preceding the release of the second thoracic segment. Pleural furrows shallow, increasingly oblique posteriorly. Pygidial margin with at least three pairs of marginal spines, the rearmost pair prolonged posteriorly.
Degree 2 meraspis. Only a single specimen (figure 3.9), almost complete but damaged anteriorly was available. The reconstruction (figure 4.5) is accordingly tentative. The individual is 0.92 mm long and 0.80 mm across at its widest point (excluding spines). Cranidium 0.40 mm long (sag.) and 0.75 mm wide (tr.). Glabella occupying central third of cranidium, slightly barrel-shaped, rounded anteriorly. S0, S1 and S2 conspicuous, dividing the glabella into distinct lobes L1 and L2 of similar size. The anterior lobe is traversed by a faint S3. Occipital ring of similar dimensions and form to L1 and L2, projecting posteriorly. The eyes are set further back than in degree 1. The course of the posterior facial suture can be faintly discerned on the left side of this specimen; it is almost straight, making an angle of some 20° to the sagittal plane, thus defining a trapezoidal cranidium. Intergenal spines still present. The right side of the specimen shows a narrow librigenae, with a genal spine extended to about the rear of the first thoracic segment.
Thorax consisting of two similar segments, 0.20 mm long (sag.), the first being somewhat longer (sag.) than the second, terminating in recurved spines. Pygidium elongate, semicircular, axis tapering posteriorly with five to six axial rings, reaching almost to the posterior margin. Median nodes poorly developed. Margin with five pairs of spines, laterally splayed, becoming inwardly curved posteriorly.
Degree ?3 meraspis. Small juvenile cranidia were available to us, but to which degree they pertain is not easy to define. The specimen illustrated in figure 3.10 may be a degree 3 meraspid judged from its aspect, but it could possibly pertain to a degree 4 or even 5 meraspid because of its size. It is a slightly distorted cranidium 0.65 mm long and 1.1 mm broad, trapezoidal in form. The glabella, occupying the central third of the cephalic width, tapers forward almost reaching the anterior border, and is rounded quadrate anteriorly. Transglabellar furrows S0, S1, S2 and S3 are subequally spaced, transverse to slightly curved backward, defining lobes of almost equal length (sag.). S0 is deeper than S1-S3. The occipital ring of which the rear margin curves posteriorly bears a prominent central node. There is a narrow (sag.), straight anterior border. The eye ridges are strong, and the palpebral lobes prominent and curved. Intergenal spines apparently are absent.
Degrees 4-5 meraspis. The slightly distorted specimen figured in 5.1 is probably a degree 4 or 5 meraspis (see also figure 6). Since the posterior region is not particularly well-preserved it is not possible to state with certainty which it actually is. The rear part is thus reconstructed on the basis of complementary material (see below). The total length is about 1.5 mm. Cranidium trapezoidal, 0.60 mm long (sag.) and 1 mm broad. Facial suture sinuous, posterior branch making an angle of some 40° to the sagittal plane. Axial and posterior border furrows deeply impressed, the former defining a glabella which though broken seems to have possessed distinct S0, S1, S2 and probably S3 furrows, delimiting subequal rectangular lobes. Occipital ring large, curving posteriorly. Eye ridges strong, curving, laterally joining the large palpebral lobes subopposite S3. Librigenae distinct, still relatively narrow and bearing stout genal spines. Intergenal spines absent. Thorax of four or five segments, axis tapering backward. Pleurae bearing terminal spines which become progressively longer and more inclined posteriorly toward the rear. Pygidium unclear, tentatively reconstructed with reference to other specimens.
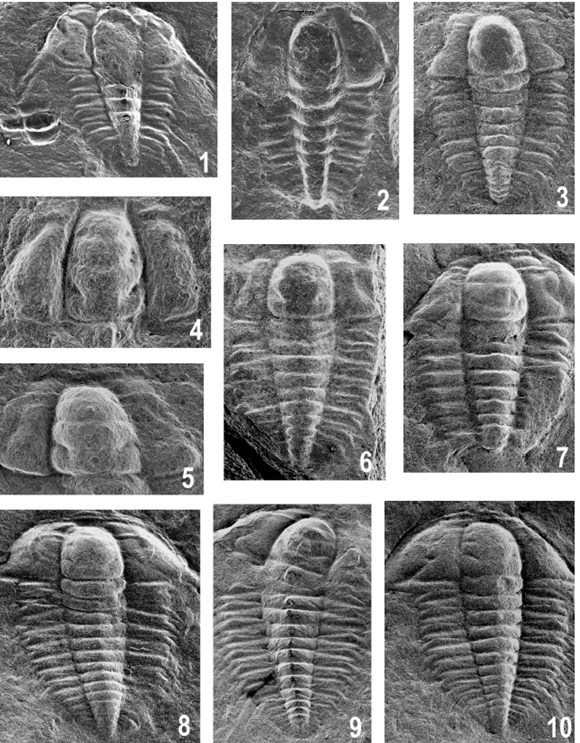
Figure 5. Parabolina frequens argentina (Kayser), from Quebrada Azul (Cajas Range) / de la quebrada Azul (sierra de Cajas). 1, Degree 4- 5? meraspis, complete specimen with glabella imperfectly preserved, bed A / meráspido 4-5?, ejemplar completo con la glabela preservada en forma imperfecta, nivel A, MLP 31578b, x35; note a degree 0 meraspid specimen on the left side / nótese un ejemplar meráspido 0 sobre el sector izquierdo. 2, Degree 4 meraspis, axial shield, bed F / meráspido 4, escudo axial, nivel F, MLP 31586, x40. 3, Degree 6 meraspis, axial shield, bed A / meráspido 6, escudo axial, nivel A, MLP 31588, x28. 4, Probable degree 4 meraspis, cranidium, bed A / probable meráspido 4, cranidio, nivel A, MLP 31587, x62. 5-7, Degree 6 meraspis / meráspidos 6; 5, cranidium, bed A / cranidio, nivel A, MLP 31589, x35; 6, axial shield, bed F / escudo axial, nivel F, MLP 31590, x25; 7, cephalon-thorax, bed D / céfalo-tórax, nivel D, MLP 31591, x27. 8, Degree 9 meraspis, complete exoskeleton, bed A / meráspido 9, exoesqueleto completo, nivel A, MLP 31593, x18. 9, Degree 8 meraspis, thorax-pygidium and displaced cephalon, bed B / meráspido 8, tórax-pigidio y céfalo desplazado, nivel B, MLP 31592, x22. 10, Degree 9 meraspis, complete exoskeleton, bed A / meráspido 9, exoesqueleto completo, nivel A, MLP 31594, x19.
The complete specimen illustrated in figure 5.2 has four thoracic segments, and is of length 1.5 mm. In the reconstruction (figure 6), pygidial spines and librigena-facial sutures are conjectural. The cranidium 0.70 mm long of figure 5.4 seems to be that of a degree 4 meraspis.
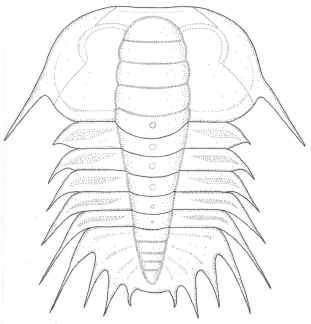
Figure 6. Parabolina frequens argentina (Kayser). Reconstruction of meraspid degree 5 / reconstrucción de un meráspido 5, x50.
Degree 6 meraspis. The primary description is mainly based on specimens illustrated in figures 5.3, 5.5- 5.7, all of which are of similar size and form. Total exoskeleton length (estimated) is 2.1 mm (without spines).
Cranidium trapezoidal, 0.87 mm long (sag.) and 1.6 mm broad. Glabella quite inflated, almost reaching anterior border, occupying slightly more than the central third. S0 deep, S1 faintly defined medially and deeply impressed laterally, oblique backward; S2 represented by a lateral pair of shallow excavations opposite centre of the large palpebral lobes. Occipital ring quite similar in form to the anterior thoracic axial ring; the axis is broadest here. Fixigenae are not especially broad in this stage. Course of facial suture sinuous, broadly inclined at about 45° to the sagittal plane.
Thorax 0.85 mm long, consisting of 6 segments. Axis markedly tapering posteriorly, the spinose tips of the thoracic spines increase in length and posterior inclination rearward. This backward swing continues into the pygidium, which has five pairs of spines so that the macropleural spines of the 8th segment are subparallel with the sagittal plane and the shorter posterior spines curve inwards. The total length of the pygidium (without spines) at this stage is 0.45 mm and the macropleural spines are already defined.
Later stages in ontogeny. Specimens assigned to these degrees are rather variable. Those representing degrees 7 and 8 in most cases resemble miniature adults although having fewer segments. That reconstructed here is 2.8 mm long (figure 5.9), whereas degree 9 meraspis (figures 5.8, 5.10) is 3.5 mm long. The smallest holaspides (figures 7.15, 10.12) in our material have attained a length of 4 mm. The time of origin and growth of the axial spine on segment 12 still remains unclear. There are indications of a short axial spine in some specimens but otherwise it seems to have grown at a later stage of development.

Figure 7. Holaspid specimens of Parabolina frequens argentina (Kayser). 1, Cranidium / cranidio, Alfarcito, MLP 31620b, x7.7. 2, Cranidium / cranidio, Quebrada Azul, bed B / nivel B, PIL 14270, x4. 3, Cranidium / cranidio, Quebrada Azul, bed D /nivel D, MLP 31608, x4.7. 4, Cranidium, latex mould / cranidio, molde de látex, Quebrada Azul, bed F / nivel F, MLP 31610, x8.7. 5, Cranidium, latex mould / cranidio, molde de látex, Alfarcito, MLP 31613, x6.5. 6, Hypostome / hipostoma, Quebrada Azul, bed A / nivel A, MLP 31627, x7. 7, Hypostome, latex mould / hipostoma, molde de látex, Quebrada Azul, bed A / nivel A, MLP 31612, x7.7. 8, Cranidium / cranidio, Quebrada Azul, bed A /nivel A, MLP 31596b, x8. 9, Cranidium / cranidio, Alfarcito, MLP 31604, x7.2. 10, Hypostome / hipostoma, Quebrada Azul, bed A/ nivel A, MLP 31618, x6.5. 11, Cranidium / cranidio, Iruya, MLP 31605, x5.1. 12, Cranidium, latex mould / cranidio, molde de látex, Quebrada Azul, bed D / nivel D, MLP 31616, x7.2. 13, Hypostome / hipostoma, Quebrada Azul, bed D / nivel D, PIL 13843, x10. 14, Cephalon and thorax, latex mould / céfalo y tórax, molde de látex, Alfarcito, MLP 31615, x4. 15, Three complete early holaspides, latex mould / tres especímenes completos, holáspidos tempranos, molde de látex, Alfarcito, PIL 12536, x6. 16, Thorax-pygidium, latex mould / tóraxpigidio, molde de látex, Quebrada Azul, bed C / nivel C, MLP 31611, x2.8.
The late holaspis morphology of P. frequens argentina is well known since Harrington and Leanza (1957, p. 81-85) provided an accurate description of the taxon, including a discussion on its synonymy and intraspecific variability. Thus, only a few complementary aspects will be pointed out here. In addition, some variable features of the adult are discussed below (see Ontogenetic Variability).
According to Nikolaisen and Henningsmoen (1985, p. 4), the genus Parabolina is characterized by a hypostome with almost completely reduced anterior wings. We describe some small holaspides and several adults of P. frequens argentina retaining their hypostomes in close association with the ventral doublure (conterminant condition) (figures 7.10, 7.13, 8.7, 8.10). On moulting it is shed along with the librigenae and may remain in its original position (figures 10.7, 10.9) or may become displaced (figures 7.6, 7.7) (see below). Originally briefly described by Kobayashi (1937, p. 477, pl. 4, fig. 13) and illustrated by Zeballo and Tortello (2005, figs. 4.K, 6.A), the hypostome is subtrapezoidal in outline, longer than wide. The contour of the anterior margin is a variable feature: in some specimens it is forwardly curved (figure 7.13), whereas in others it is virtually planar (figure 7.10), providing a more efficient surface of contact to the rear edge of the cephalic doublure. The middle body is prominent, ovoid, convex, with the anterior edge poorly defined, highest posteriorly. Anteriorly, the middle body virtually merges with the frontal margin, whereas it is separated from the posterior lobe by a deep, narrow and uniformly impressed middle furrow which is strongly bowed backward. The surface is smooth. Anterior wings small, very narrow (tr.). A thin external rim is present on each side of the hypostome. Posterior margin semicircular, surrounded by a very narrow (sag.), convex posterior border.

Figure 8. Holaspid specimens of Parabolina frequens argentina (Kayser). Most specimens from Quebrada Azul (Cajas Range) excepting 9 (Alfarcito) / la mayoría de los especímenes de la Quebrada Azul (sierra de Cajas) excepto 9 (Alfarcito). 1, Cranidium, thorax and displaced, inverted pygidium, bed A / cranidio, tórax, y pigidio desplazado e invertido, nivel A, PIL 13835, x2.4. 2, Cranidium, bed F / cranidio, nivel F, PIL 14263, x5.5. 3, Incomplete thorax, bed F / tórax incompleto, nivel F, MLP 31597, x4.3. 4, Cranidium and displaced fragmentary thorax, bed D / cranidio y tórax fragmentario, desplazado, nivel D, MLP 31623, x4.4. 5, Fragmentary thorax and pygidium, bed D / tórax fragmentario y pigidio, nivel D, MLP 31619, x4.2. 6, Fragmentary thorax and displaced, inverted pygdium, bed A / tórax fragmentario y pigidio desplazado e invertido, nivel A, MLP 31600, x4.2. 7, Thorax-pygidium + librigenae and hypostome, bed A / tórax-pigidio + librígenas e hipostoma, nivel A, MLP 31622, x3.5. 8, Fragmentary thorax, latex mould, bed D / tórax fragmentario, molde de látex, nivel D, MLP 31632, x6.2. 9, Cephalon and thorax / céfalo y tórax, MLP 31603, x8. 10, Thorax-pygidium + librigenae and hypostome, bed D / tórax-pigidio + librígenas e hipostoma, nivel D, PIL 13843, x3.6. 11, Cephalon and displaced thorax-pygidium, latex mould, bed F / céfalo y tórax-pigidio desplazado, molde de látex, nivel F, MLP 31614, x8.3. 12, Pygidium, latex mould, bed D / pigidio, molde de látex, nivel D, MLP 31617, x8. 13, Pygidium, bed A / pigidio, nivel A, MLP 31606, x6.2.
Some well-preserved fully adult specimens of P. frequens argentina show a delicate rounded protuberance in front of the occipital node, in the centre of the occipital ring (e.g., figures 7.1, 7.4, 7.5, 7.11, 9.6). In whitened specimens it is possible to note indications of its internal structure, which seems to consist of fine pits. This protuberance resembles the median occipital organ described from the upper Cambrian olenid Olenus wahlenbergi Westergård by Clarkson and Taylor (1995a). It is similar to dorsal organs in larval decapod crustaceans, which are combined glandular and sensory organs (Barrientos and Laverack, 1986). An occipital ring with "two tubercles" was also described in P. (Neoparabolina) frequens finmarchica Nikolaisen and Henningsmoen (1985, figs. 4, 11.C-J, 5.A-Ea), from the Lower Tremadocian of Norway.

Figure 9. Holaspides of Parabolina frequens argentina (Kayser). Most specimens from Quebrada Azul (Cajas Range) excepting 2 and 6 (Alfarcito) / la mayoría de los especímenes de la quebrada Azul (sierra de Cajas) excepto 2 y 6 (Alfarcito). 1, Axial shield, bed C / escudo axial, nivel C, MLP 31607, x6.8. 2, Complete specimen / ejemplar completo, MLP 31601, x6.6. 3, Librigenae and thorax, bed D / mejilla libre y tórax, nivel D, MLP 31628, x5.4. 4, Thorax-pygidium, bed D / tórax-pigidio, nivel D, MLP 31598, x3.5. 5, Thorax-pygidium, bed F / tórax-pigidio nivel F, MLP 31599, x4.3. 6, Complete specimen, latex mould / ejemplar completo, molde de látex, MLP 31609, x4.5. 7, Librigenae, thoraxpygidium and hypostome, bed D / mejilla libre, tórax-pigidio e hipostoma, nivel D, MLP 31595, x4.3. 8, Complete specimen, bed D / ejemplar completo, nivel D, MLP 31596a, x5.4. 9, 12th thoracic segment showing axial spine, bed F / último segmento torácico, con espina axial, nivel F, MLP 31602, x5. 10, Librigenae and thorax-pygidium, bed A / librígenas y tórax-pigidio, nivel A, PIL 14271, x5.3.

Figure 10. Holaspides of Parabolina frequens argentina (Kayser). 1, Thorax, latex mould / tórax, molde de látex, Quebrada Azul, bed F / nivel F, MLP 31634, x5.2. 2, Thorax-pygidium / tórax-pigidio, Quebrada Azul, bed B / nivel B, MLP 31626, x7.8. 3, Two early holaspides / dos holáspidos tempranos, Alfarcito, MLP 31621, x7.5. 4, Disarticulated cranidium, librigenae and thorax, latex mould / cranidio, librígenas y tórax desarticulados, molde de látex, Quebrada Azul, bed A / nivel A, MLP 31630, x6.3. 5, Thorax-pygidium / tórax-pigidio, Quebrada Azul, bed A / nivel A, PIL 13088, x2. 6, Thorax-pygidium, latex mould / tórax-pigidio, molde de látex, Quebrada Azul, bed F / nivel F, MLP 31633, x2.6. 7, Thorax-pygidia and displaced librigena with associated hypostome, latex mould / tórax-pigidios y librígena con hipostoma asociado, molde de látex, Alfarcito, PIL 12536, x2.5. 8, Displaced cranidium, librigenae and thorax / cranidio, librígenas y tórax, desplazados, Quebrada Azul, bed D / nivel D, MLP 31629, x3.3. 9, Thorax and displaced librigenae + hypostome / tórax y librígenas + hipostoma desplazados, Quebrada Azul, bed C / nivel C, MLP 31625, x5. 10, Cephalon and fragmentary thorax, showing Harrington's configuration / céfalo y parte del tórax, con las librígenas desplazadas, dispuestas debajo del cranidio, Quebrada Azul, bed D / nivel D, MLP 31624, x3.6. 11, Fragmentary thorax, latex mould / tórax fragmentario, molde de látex, Quebrada Azul, bed D / nivel D, MLP 31631, x5.7. 12, Axial shield with displaced cranidium, and fragmentary thorax / escudo axial con cranidio desplazado, y tórax fragmentario, Alfarcito, MLP 31620a, x6.
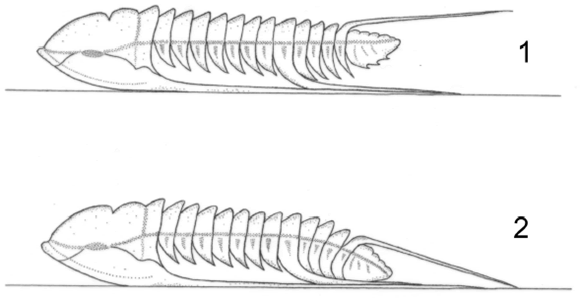
Figure 11. Parabolina frequens argentina (Kayser), reconstruction in lateral view in (1) active and (2) relaxed posture / reconstrucción en vista lateral de las posturas activa (1) y de reposo (2).
The present study has revealed no further details of the structure of the eye, and the nature of the lentiferous surface remains unknown.
Ontogenetic variability
In the majority of trilobites and, certainly in most olenids, morphology and size are closely coupled throughout development. In some cases it has proved possible, by measuring length/width ratios, to establish instar groupings which plot out on a graph as a sequence of clusters (Clarkson and Ahlberg, 2002; Clarkson et al., 2003, 2004). These clusters correspond to protaspid and meraspid degrees, and sometimes can be traced into the early holaspid stages. Within each degree, morphology and size are tightly constrained, and all the individuals forming a cluster are very similar in appearance. An isolated cranidium or pygidium can thus be confidently referred to its correct degree by comparison with intact specimens. Even if an ontogeny has been worked out on the basis of disarticulated sclerites alone, it may still be possible to determine, with a fair degree of certainty, to which degree an individual cranidium or pygidium belongs.
In some olenids, however, clusters are more open, or less evident (Tortello and Clarkson, 2003). Some instances have been described where, although most individuals follow a "normal" growth trajectory some others do not fall within the expected size range. Thus in Ctenopyge (Ctenopyge) gracilis Henningsmoen, from the Furongian of Sweden, degree 7 meraspides have been described which are the size of degree 6 or even degree 5 individuals (Clarkson et al., 2004). They have achieved a more mature form when still quite small.
The highest degree of variability in olenids described up until now is in Parabolina spinulosa (Wahlenberg), from the Furongian of Sweden, both in the juvenile stages and the adult. The ontogeny of P. spinulosa resembles that of P. frequens argentina, as would be expected, though the early stages of development are more clearly represented in the latter, and the macropleural spines are lacking in the former. But direct comparisons are difficult because of this variability, which affects both size and proportions as well as basic morphology (Clarkson et al., 1997, p. 83). The same kind of variability is true on Parabolina frequens argentina, but to an even greater degree. The problem of assigning disarticulated meraspid/young holaspid sclerites to their correct degree, noted by these authors in P. spinulosa, is more acute and has led to some problems in working out successive stages in development. As in P. spinulosa both size and shape are highly variable. Thus, specimen illustrated in figure 5.2 is an undoubted degree 4 meraspis 1.4 mm in length, while that of figure 5.1 is a degree 5 meraspis of very similar size. The degree 6 meraspid is considerably larger (specimen in figure 5.3 is nearly twice the size of that of figure 5.1, being 2.2 mm in length). Degree 7 meraspid is not much larger.
Shape and proportions are also greatly variable. This is particularly evident in the glabellar furrows. In "normal" early olenid development there are four subequal lobes in the glabella, defined by furrows S0, S1, S2 and S3. These become modified later in development, reducing to two or three. In P. frequens argentina the range of variation in juvenile specimens is formidable. Thus the small degree 5 meraspid has three transglabellar furrows, retaining a rather juvenile form while in a larger degree 6 meraspis (figure 5.3) only S1 is present, impressed laterally but hardly transglabellar. Because of this great range of variation it is not usually possible to assign disarticulated sclerites to their correct degree. Thus, a well-preserved cranidium 0.65 mm in length and 1.1 mm in width has four subequal glabellar lobes as in an early meraspid (figure 3.10). From its size it could represent any stage of development between a degree 3 and a degree 5 meraspid and it is just not possible to be more definite.
Other proportions of the developing exoskeleton are also singularly variable; the sagittal length of the thoracic segments, the splay and length of the genal and thoracic spines, the relative width of the axis, and the relative development of the axial tubercules all vary greatly. Thus, the evident differences between the reconstructions of degree 1 and degree 2 meraspis are actual, and not just the result of imperfect preservation.
In addition, Harrington and Leanza (1957) cited several variable features in the holaspid morphology of P. frequens argentina. For instance, no less than five gradational patterns in glabellar furrows have been defined for the adult: one, two or three furrows may be present and of variable form. Similarly, the cephalic anterior border can be, according to Harrington and Leanza (1957, p. 83), 1.7 to 3 times wider (sag.) than preglabellar field, a fact clearly exemplified in figures 7.1-7.5, 7.8-7.9, 7.11-7.12. The anterior cephalic border of many specimens from Alfarcito seems to be narrower (sag.) than that of individuals from sierra de Cajas, though the border of a few individuals from the former locality is proportionately wide (see figure 7.1). The width (sag.) of the preglabellar field is also a rather variable character in the material studied, which intergrades between 2% and 9% the length (sag.) of the cranidium.
As documented in other olenids, the nodes on the axis of the thorax and the pygidium are better presented in juveniles and small adults of P. frequens argentina, whereas the full-grown individuals can bear distinct, partially developed, or smooth axial nodes (e.g., compare figures 8.1, 9.1-9.8, 9.10). Clarkson et al. (1997) suggested that the axial nodes originated as juvenile structures, which disappeared in the later stages of development of many Ordovician olenids (e.g., Jujuyaspis keideli (Kobayashi); see Tortello and Clarkson, 2003), but were paedomorphically retained in the adults of Parabolina.
Pribyl and Vanek (1980) postulated a variability affecting number of marginal spines in the holaspid pygidium of P. frequens argentina, stating that some specimens can bear two pairs of spines instead of three as it is usual. Nevertheless, we could not find such variation in the material examined. All wellpreserved pygidia from Cajas, Iruya and Alfarcito show three pairs of marginal spines, suggesting that it is a stable condition (cf. Harrington and Leanza, 1957).
Systematic position
After Kayser (1876), Kobayashi (1937), Harrington (1938), and Harrington and Leanza (1957), Pribyl and Vanek (1980) illustrated additional material of P. argentina from Bolivia and pointed out its great morphological similarity with P. frequens (Barrande, 1868), a species previously described from the Lower Ordovician of Germany and Bohemia (Barrande, 1868; Sdzuy, 1955, pl. 3, figs. 58-70; text-fig. 10). Pribyl and Vanek (1980) noted that argentina hardly distinguishes from frequens in having "2-3 pygidial spines" and a pygidial axis reaching slightly nearer to the posterior margin of pygidium. Therefore, they regarded the two as subspecies of P. frequens.
Rushton (1982, pl. 2, fig. 15) illustrated additional material of P. frequens frequens from the Furongian of Wales and, in view of the variability reported in P. frequens argentina by Harrington and Leanza (1957), suggested that both taxa could be synonyms (cf. Nikolaisen and Henningsmoen, 1985). More recently, Zylinska (2001, text.-fig. 11; pl. 5, figs. 1-14) described new specimens from the upper Cambrian of Poland and made a comparative analysis of the subspecies of P. frequens ( P. frequens frequens, P. frequens argentina, as well as P. frequens finmarchica Nikolaisen and Henningsmoen (1985, figs. 4, 11.C-J, 12.A-Ea) from the lower Tremadocian of northern Norway). Zylinska (2001) evaluated the large morphologic variation within these subspecies, concluding that they fall into the variability of P. frequens and hence cannot be maintained.
Parabolina frequens is evidently a variable taxon. However, we consider that its described subspecies have some stable features which justify their validity. P. frequens argentina differs from P. frequens frequens in having a more transversely expanded pygidium, three pygidial marginal spines (2-4 may occur in P. frequens frequens), and a proportionately narrower (sag.) pygidial posterior border (cf. Pribyl and Vanek, 1980). In addition, the hypostome of the former has a more rounded middle body.
Parabolina frequens argentina and P. frequens finmarchica share a relatively narrow pygidial border, but the latter clearly differs in having forwardly located genal spines, stronger ocular ridges, convergent anterior branches of facial suture, and a hypostome with a narrower middle body.
Moulting
The studied material consists of numerous specimens representing both ecdysial remains and exoskeletons of dead individuals. Since ecdysial units are hardly attacked by scavengers, and each trilobite normally produced several exuviae, the chances of finding moulting configurations in low-energy environments should be greater than finding remains of dead animals (Henningsmoen, 1975). The latter are generally represented by complete specimens showing no fissures between exoskeletal plates, occasionally found in life attitude or tightly enrolled (individuals illustrated in figures 9.2 and 9.6 may represent such condition). On the other hand, the material at our disposal includes many exoskeletal remains that can be interpreted as exuviae. Olenids moulted by opening the facial suture and emerging forwards, leaving the exuvial parts on various distinctive arrangements (Henningsmoen, 1957, 1975; Chatterton and Ludvigsen, 1998). It is common to find the exoskeleton of P. frequens split in two main parts: axial shield (cranidium + thorax + pygidium) (e.g., figure 9.1) and librigenae (e.g., figure 10.7). They may lie close together or removed from each other. Similar configurations were described by Chatterton and Ludvigsen (1998) in the upper Steptoean trilobites Wujiajiania sutherlandi Chatterton and Ludvigsen (Olenidae), Labiostria Palmer (Aphelaspidae) and Pterocephalia Roemer (Pterocephaliidae).
The librigenae are fused into a single piece in Parabolina (Sdzuy, 1955; Henningsmoen, 1957; Clarkson et al., 1997). Several displaced librigenae of Parabolina frequens argentina retaining their hypostomes are illustrated in figures 10.7 and 10.9. In some cases the librigenae were displaced backwards below the axial shield during the ecdysis ("Harrington's configuration" Henningsmoen, 1975 - see figure 10.10-), a fact also documented by Harrington and Leanza (1957) in other olenids (e.g., Parabolinella Kobayashi) as well as in some kainellids (e.g., Pseudokainella Harrington, Apatokephalus Brogger) (Harrington and Leanza, 1957, figs. 38.8, 52.6, 58.3), and by Chatterton and Ludvigsen (1998, fig. 19.5) in Labiostria.
Other specimens of P. frequens argentina can be interpreted as typical exuviae. In some individuals the cranidium is missing, whereas the librigenae are preserved more or less in normal position (figures 8.7, 8.10, 9.3, 9.10). A similar ecdysial configuration has been previously documented in Parabolina (Harrington and Leanza, 1957, fig. 25.2) and many other olenids ( e.g., Cloacaspis Fortey (1974, pl. 12, fig. 1), Bienvillia Clark (Harrington and Leanza, 1957, fig. 43.1f), Saltaspis Harrington and Leanza (1957, fig. 32.1c), Acerocare Angelin (Henningsmoen, 1957, pl. 30.5; Henningsmoen, 1975, fig. 10.A)) and non-olenid trilobites (see McNamara and Rudkin, 1984; Chatterton and Ludvigsen, 1998). Similarly, the collections studied include specimens consisting of a thorax or a thoracopygon and a displaced cranidium (figures 8.4, 10.4, 10.12).
It is also common to find disarticulated axial shields and isolated fragmentary thoraxes (e.g., figures 8.4, 8.8, 9.4, 9.9, 10.1-10.4, 10.7-10.9, 10.11- 10.12), suggesting that in many cases moulting was a difficult process. In many cases thoracic segments are preserved displaced, separated, or telescoped under each other. Although exuviation in P. frequens argentina generally occurred by egression of the postecdysial trilobite from its exuvia without inversion of any exoskeletal elements, there are some examples consisting of an inverted pygidium close to its corresponding thorax (see figures 8.1, 8.6).
Figure 8.9 shows a specimen with "open" facial sutures. According to Henningsmoen (1975), it may represent either an exuvium or a dead trilobite with sutures opened at its site of death or after postmorten transport.
Most of these examples were collected from the Lampazar Formation in Sierra de Cajas, from levels representing a quiet-water, outer shelf setting (Tortello and Esteban, 2003). Moulting of P. frequens argentina took place on the sea floor. The preservation of arrangements of exoskeletal elements which have not suffered postecdysial disturbance, by bioturbation or by physical proceses, is compatible with a low-energy environment.
Life attitude
All the available material of P. frequens argentina is flattened, at least to some extent, and the original convexity of the exoskeleton remains unknown. Trilobites of the subfamily Oleninae, however, are never greatly convex, as exemplified by Olenus wahlenbergi Westergård reconstructed from material retaining its original form (Clarkson and Taylor, 1995a, fig. 16b). Here we assume a similar convexity for P. frequens argentina. From the functional point of view, this species differs from Olenus in having long genal spines, macropleural spines on the 8th thoracic segment, and the strong axial spine on the 12th and final thoracic segment. It is very likely that the genal and macropleural spines supported the body of the trilobite when resting on the sea floor. The exoskeleton can be reconstructed in two alternative postures, the "alert" and "relaxed" attitudes (figure 11). In the "alert" posture the body would rest on the horizontal genal and macropleural spines but the whole body would be stretched out horizontally. The extensor muscles would be contracted and all the apodemes would lie in a single horizontal plane.
The difference between the "alert" and the "relaxed" posture is that in the latter case the extensor muscles are no longer contracted. Thus the rigid frame provided by the genal and macropleural spines would hold the anterior part of the body as in the "alert" posture but the rear part posterior to the macropleural spines of the 8th thoracic segment would incline downward as the muscles relaxed.
The pleural extremities of the segments posterior to the macropleural spines are quite long and graduated in size toward the rear. In the "relaxed" attitude they may well have touched the seafloor but most importantly, strong support would be given to the posterior part of the body by the axial spine of the final thoracic segment. The angle which this spine made, in life, to the horizontal plane cannot be known with certainty since it is normally preserved broken or crushed against the thorax. If, however, the spine was parallel with the extended body or rose at an angle of only a few degrees, it would support the relaxed trilobite most effectively, as it is reconstructed.
In the "alert" attitude the trilobite would be ready for action, whereas when fully relaxed it would have been supported at all five points with minimal expenditure of energy. Fortey (1985) suggested that the morphology of Parabolina of P. frequens type (= Neoparabolina) is comparable to that of the Furongian genus Irvingella Ulrich and Resser, a taxon with possible pelagic habits. The occurrence of P. frequens argentina in different lithologies (Tortello and Esteban, 2003) is compatible with such a mode of life; however, the size of the eyes of Parabolina seems to be similar to that of most benthic olenids. As stated above, P. frequens argentina was able to rest on the sea floor supported by the genal and macropleural spines. Most leptoplastines (Clarkson and Taylor, 1995b; Clarkson et al., 2003, 2004) used their long genal spines to support the outstretched body above the water-sediment interfacies. In P. frequens argentina a similar objective has been achieved in a somewhat different way, testifying to the remarkable evolutionary flexibility among trilobites of the Family Olenidae.
Acknowledgements
We thank S. Esteban, G. Aceñolaza and P. Payrola for assistance in the field. B. Waisfeld and E. Vaccari provided additional specimens for comparison. Figures 1-2 were produced by D. Ruiz Holgado and E. Gómez-Hasselrot. We also thank J. Rodríguez for assistance with photography. The paper was greatly improved by constructive comments from B. Chatterton and S. Peng. Financial support was provided by the Consejo Nacional de Investigaciones Científicas y Técnicas (Argentina) and the Carnegie Trust for the Universities of Scotland, which enabled E. N. K. Clarkson to visit La Plata in August 2005.
Bibliography
1. Aceñolaza, F.G. 1968. Geología estratigráfica de la región de la sierra de Cajas, Dpto. Humahuaca (provincia de Jujuy). Revista de la Asociación Geológica Argentina 23: 207-222. [ Links ]
2. Aceñolaza, F.G. 1983. The Tremadocian beds and the Cambrian- Ordovician boundary problems in Latin America. In: Papers for the Symposium on the Cambrian-Ordovician and Ordovician- Silurian boundaries, Nanjing Institute of Geology and Palaeontology, Academia Sínica, Nanjing, pp. 88-93. [ Links ]
3. Aceñolaza, F.G. 1992. El Sistema Ordovícico de Latinoamérica. In: J.C. Gutiérrez Marco, J. Saavedra and I. Rábano (eds.), Paleozoico Inferior de Ibero-América, Universidad de Extremadura, pp. 85-118. [ Links ]
4. Aceñolaza, F.G. and Aceñolaza, G.F. 1992. The genus Jujuyaspis as a world reference fossil for the Cambrian-Ordovician boundary. In: B.D. Webby and J.R. Laurie (eds.), Global Perspectives on Ordovician Geology, Balkema, Rotterdam, pp. 115-120. [ Links ]
5. Barrande, J. 1868. Faune Silurienne des environs de Hof, en Baviere. Prague, pp. 31-110. [ Links ]
6. Barrientos, Y. and Laverack, M. S. 1986. The larval crustacean dorsal organ and its relationship to the trilobite median tubercle. Lethaia 19: 309-313. [ Links ]
7. Branisa, L. 1965. Los fósiles guía de Bolivia. Boletín del Servicio Geológico de Bolivia 6: 1-82. [ Links ]
8. Benedetto, J.L. 1977. Algunas consideraciones acerca de la posición del límite Cámbro-Ordovícico en América del Sur. Geos 23: 3-11. [ Links ]
9. Berard, P., Clarkson, E.N. K. and Taylor, C.M. 2000. The ontogeny of Taihungshania miqueli (Bergeron, 1893) from the Arenig of the Montagne Noire, southern France. Transactions of the Royal Society of Edinburgh: Earth Sciences 90: 147-163. [ Links ]
10. Burmeister, H. 1843. Die Organisation der Trilobiten, aus ihren lebenden Verwandten entwickelt; nebst einer systematischen übersicht aller zeither beschriebenen Arten. Berlin, 147 pp., 6 pls. [ Links ]
11. Chatterton, B.D.E. and Ludvigsen, R. 1998. Upper Steptoean (Upper Cambrian) trilobites from the McKay Group of southeastern British Columbia, Canada. The Paleontological Society Memoir [ supplement to Journal of Paleontology 72] 49: 1-43. [ Links ]
12. Clarkson, E.N.K. and Ahlberg, P. 2002. Ontogeny and structure of a new, miniaturised and spiny olenid trilobite from southern Sweden. Palaeontology 45: 1-22. [ Links ]
13. Clarkson, E.N.K. and Taylor, C.M. 1995a. Ontogeny of the trilobite Olenus wahlenbergi Westergård, 1922 from the upper Cambrian Alum Shales of Andrarum, Skåne, Sweden. Transactions of the Royal Society of Edinburgh: Earth Sciences 86: 13-34. [ Links ]
14. Clarkson, E.N.K. and Taylor, C.M. 1995b. The lost world of the olenid trilobites. Geology Today, July-August 1995: 147-154. [ Links ]
15. Clarkson, E.N.K., Taylor, C.M. and Ahlberg, P. 1997. Ontogeny of the trilobite Parabolina spinulosa (Wahlenberg) from the upper Cambrian Alum Shales of Sweden. Transactions of the Royal Society of Edinburgh: Earth Sciences 88: 69-89. [ Links ]
16. Clarkson, E.N.K., Ahlgren, J. and Taylor, C.M. 2003. Structure, ontogeny, and moulting of the olenid trilobite Ctenopyge (Eoctenopyge) angusta Westergård, 1922 from the Upper Cambrian of Västergötland, Sweden. Palaeontology 46: 1-27. [ Links ]
17. Clarkson, E.N.K., Ahlgren, J. and Taylor, C.M. 2004. Ontogeny, structure and functional morphology of some spiny Ctenopyge species (Trilobita) from the upper Cambrian of Vastergotland, Sweden. Transactions of the Royal Society of Edinburgh: Earth Sciences 94: 115-143. [ Links ]
18. Fortey, R.A. 1974. The Ordovician trilobites of Spitsbergen. I. Olenidae. Norsk Polarinstitutt Skrifter 160: 1-81. [ Links ]
19. Fortey, R.A. 1975. Early Ordovician trilobite communities. Fossils and Strata 4: 331-352. [ Links ]
20. Fortey, R.A. 1985. Pelagic trilobites as an example of deducing the life habits of extinct arthropods. Transactions of the Royal Society of Edinburgh 76: 219-230. [ Links ]
21. Frederickson, E.A. 1958. Lower Tremadocian trilobites from Venezuela. Journal of Paleontology 32: 541-543. [ Links ]
22. Harrington, H.J. 1938. Sobre las faunas del Ordoviciano Inferior del Norte argentino. Revista del Museo de La Plata, Nueva Serie (Sección Paleontología) 1: 109-289. [ Links ]
23. Harrington, H.J. and Leanza, A.F. 1943. La fáunula del Tremadociano inferior de Salitre (Bolivia). Revista del Museo de La Plata (Nueva Serie), Sección Paleontología 2: 343-356. [ Links ]
24. Harrington, H.J. and Leanza, A.F. 1957. Ordovician trilobites of Argentina. Department of Geology, University of Kansas Special Publication 1: 1-276. [ Links ]
25. Henningsmoen, G. 1957. The trilobite family Olenidae, with description of Norwegian material and remarks on the Olenid and Tremadocian Series. Skrifter Utgitt av det Norske Videnskaps-Akademi i Oslo I Matematisk-Naturvidenskapelig Klasse 1957(1): 303 pp. [ Links ]
26. Henningsmoen, G. 1975. Moulting in trilobites. Fossils and Strata 4: 179-200. [ Links ]
27. Kayser, E. 1876. Uber primordiale und untersilurische Fossilien aus der Argentinischen Republik. Palaeontographica, Supplementun 3, Lieferung 2, Theil III. En: Actas de la Academia Nacional de Ciencias 8: 297-332, Tablas 1-5 (Spanish version). [ Links ]
28. Kobayashi, T. 1937. The Cambro-Ordovician shelly faunas of South America. Journal of the Faculty of Science, Imperial University of Tokyo, Section 2, 4: 369-522. [ Links ]
29. Ludvigsen, R. 1982. The Cambrian-Ordovician boundary in the western District of Mackenzie, Canada. In: M.G. Basset and W. T. Dean (eds.), The Cambrian-Ordovician boundary: sections, fossil distributions, and correlations, National Museum of Wales, Geological Series 3: 141-153. [ Links ]
30. McNamara, K.J. and Rudkin, D. M. 1984. Techniques of trilobite exuviation. Lethaia 17: 153-173. [ Links ]
31. Nikolaisen, F. and Henningsmoen, G. 1985. Upper Cambrian and lower Tremadoc olenid trilobites from the Digermul peninsula, Finmark, northern Norway. NGU Norges Geologiske Undersokelse Bulletin 400: 1-49. [ Links ]
32. Ortega, G. and Rao, R.I. 1995. Nuevos hallazgos de graptolitos (Ordovícico Inferior) en la sierra de Cajas, Departamento Humahuaca, Provincia de Jujuy, Argentina. Boletín de la Academia Nacional de Ciencias (Córdoba) 60: 293-316. [ Links ]
33. Pagani, M.A., Sabattini, N.M. and Tortello, M.F. 2005. A new species of Hyolitha from the Late Cambrian of northwestern Argentina. Serie Correlación Geológica 19: 169-176. [ Links ]
34. Pribyl, A. and Vanek, J. 1980. Ordovician trilobites of Bolivia. Rozpravy Ceskoslovenske Akademie Ved 90: 1-90, pls. 1-24. [ Links ]
35. Raw, F. 1925. The development of Leptoplastides salteri and other trilobites. Quarterly Journal of the Geological Society of London 81: 233-284. [ Links ]
36. Robison, R.A. and Pantoja-Alor, J. 1968. Tremadocian trilobites from the Nochixtlán region, Oaxaca, Mexico. Journal of Paleontology 42: 767-800. [ Links ]
37. Rushton, A.W.A. 1982. The biostratigraphy and correlation of the Merioneth-Tremadoc Series boundary in North Wales. In: M. G. Basset and W. T. Dean (eds.), The Cambrian-Ordovician boundary: sections, fossil distributions, and correlations, National Museum of Wales, Geological Series 3: 41-59. [ Links ]
38. Salfity, J. A., Malanca, S., Moya, M. C., Monaldi, C. R. and Brandán, E. M. 1984. El límite Cámbrico-Ordovícico en el norte de la Argentina. 9° Congreso Geológico Argentino (San Carlos de Bariloche), Actas 1: 568-575. [ Links ]
39. Salter, J.W. 1849. Figures and descriptions illustrative of British organic remains. Memoirs Geological Survey U. K., decade 2, 39 pp. [ Links ]
40. Sdzuy, K. 1955. Die Fauna der Leimitz-Schefer (Tremadoc). Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft 492: 1-74. [ Links ]
41. Shergold, J. H. 1988. Review of the trilobite biofacies distributions at the Cambrian-Ordovician boundary. Geological Magazine 125: 363-380. [ Links ]
42. Steinmann, G. and Hoek, H. 1912. Das Silur und Cambriun des Hochlandes von Bolivia und ihre Fauna. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie 34: 176-252. [ Links ]
43. Størmer, L. 1942. Studies on trilobite morphology, Part II. The larval development, the segmentation and the sutures, and their bearing on trilobite classification. Norsk Geologisk Tidsskrift 21: 49-164. [ Links ]
44. Tortello, M.F. 2003. Biostratigraphic significance of the latest Cambrian-earliest Ordovician agnostoid trilobites from northwestern Argentina. Geologica Acta 1: 61- 72. [ Links ]
45. Tortello, M.F. and Clarkson, E.N.K. 2003. Ontogeny of the Early Ordovician olenid trilobite Jujuyaspis keideli Kobayashi from northwestern Argentina. Ameghiniana 40: 257-275. [ Links ]
46. Tortello, M.F. and Esteban, S.B. 1998. La transición Cámbrico- Ordovícica en la Formación Volcancito (Sistema de Famatina, La Rioja, Argentina). 7° Congreso Argentino de Paleontología y Bioestratigrafía (Bahía Blanca), Resúmenes: 93. [ Links ]
47. Tortello, M.F. and Esteban, S.B. 1999. La transición Cámbrico- Ordovícico en la Formación Volcancito (sierra de Famatina, La Rioja, Argentina). Ameghiniana 36: 371-387. [ Links ]
48. Tortello, M.F. and Esteban, S.B. 2003. Trilobites del Cámbrico Tardío de la Formación Lampazar (sierra de Cajas, Jujuy, Argentina). Implicancias bioestratigráficas y paleoambientales. Ameghiniana 40: 323-344. [ Links ]
49. Tortello, M.F. and Rao, R.I. 2000. Trilobites y conodontes del Ordovícico Temprano del Angosto de Lampazar (provincia de Salta, Argentina). Boletín Geológico y Minero 111: 61-84. [ Links ]
50. Tortello, M.F., Rábano, I., Rao, R.I. and Aceñolaza, F.G. 1999. Los trilobites de la transición Cámbrico-Ordovícico en la quebrada Amarilla (sierra de Cajas, Jujuy, Argentina). Boletín Geológico y Minero 110: 555-572. [ Links ]
51. Turner, J.C. 1964. Descripción geológica de la Hoja 2c, Santa Victoria (Provincias de Salta y Jujuy). Boletín del Instituto Nacional de Geología y Minería 104: 93 pp. [ Links ]
52. Turner, J.C. and Mon, R. 1979. Cordillera Oriental. 2° Simposio Geología Regional Argentina, Academia Nacional de Ciencias 1: 57-94. [ Links ]
53. Vilela, C.R. 1960. Algunos rasgos particulares de la geología de Iruya (Salta-Jujuy). Revista de la Asociación Geológica Argentina 20: 119-140. [ Links ]
54. Zeballo, F. and Tortello, M.F. 2005. Trilobites del Cámbrico Tardío - Ordovícico Temprano del área de Alfarcito, Tilcara, Cordillera Oriental de Jujuy, Argentina. Ameghiniana 42: 127- 142. [ Links ]
55. Zylinska, A. 2001. Late Cambrian trilobites from the Holy Cross Mountains, central Poland. Acta Geologica Polonica 51: 333-383. [ Links ]
Recibido: 6 de diciembre de 2006.
Aceptado: 18 de setiembre de 2007.

