










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.45 n.2 Buenos Aires abr./jun. 2008
Stegomastodon platensis (Proboscidea, Gomphotheriidae) en el Pleistoceno de Santiago del Estero, Argentina
María Teresa Alberdi1, Esperanza Cerdeño2 y José Luis Prado3
1Departamento de Paleobiología. Museo Nacional de Ciencias Naturales, CSIC, José Gutiérrez Abascal 2, 28006 Madrid, España. malberdi@mncn.csic.es
2Departamento de Geología y Paleontología, IANIGLA, CCT-CONICET- Mendoza. Avda. Ruiz Leal s/n. 5500 Mendoza, Argentina. espe@lab.cricyt.edu.ar
3INCUAPA-Departamento de Arqueología, Universidad Nacional del Centro. Del Valle 5737. 7400 Olavarría, Argentina. jprado@soc.unicen.edu.ar
Resumen. Los mamíferos fósiles de la Provincia de Santiago del Estero (Argentina) son escasamente conocidos. En este trabajo, se describen los restos mejor conservados de gonfoterios (Mammalia, Proboscidea) encontrados en los niveles pleistocenos aflorantes a lo largo del Río Dulce, cerca de la ciudad de Santiago del Estero. El estudio comparativo con otros gonfoterios provenientes de distintas localidades de América del Sur, en su mayoría argentinas y brasileñas, permite identificarlos como Stegomastodon platensis. Una muestra de esmalte dentario fue datada por 14C AMS y proporciona una edad de 19.900 ± 120 AP, confirmando su asignación al Pleistoceno superior. Se analizó la composición isotópica del carbono y del oxígeno del esmalte dentario con el fin de obtener nuevos datos sobre las preferencias alimenticias y de hábitat de los gonfoterios sudamericanos. Los valores isotópicos del carbono indican una dieta exclusiva de plantas C4. El análisis de los isótopos de oxígeno indica que el clima era más frío que en la actualidad.
Abstract. Stegomastodon platensis (Proboscidea, Gomphotheriidae) from Pleistocene levels of Santiago del Estero, Argentina. Mammal fossil remains from Santiago del Estero Province (Argentina) are very poorly known. In this paper, best preserved specimens of Gomphotheriidae (Mammalia, Proboscidea) are presented. They come from Quaternary beds outcropping along the Río Dulce, near of the city of Santiago del Estero. The comparative study with other South American gomphotheres remains, mainly from Argentina and Brazil, allow their identification as Stegomastodon platensis, being among the largest forms of this species. Previous studies referred this locality to the late Pleistocene. Enamel tooth sample was dated to 19.900 ± 120 BP by AMS 14C, supporting that age for this fauna. Carbon and oxygen isotopic composition of the tooth enamel has been analyzed to get new data on the paleodiet and habitat preference of South American gomphotheres. Carbon isotopic values indicate an adaptation to an exclusively C4 diet. The oxygen isotopic analysis indicates cooler climatic conditions than present.
Palabras clave. Gomphotheriidae; Stegomastodon platensis; Pleistoceno Superior; Santiago del Estero; Argentina; América del Sur.
Key words. Gomphotheriidae; Stegomastodon platensis; Late Pleistocene; Santiago del Estero; Argentina; South America.
Introducción
La familia Gomphotheriidae Hay, 1922 es un grupo ancestral del cual se originan otros grupos de proboscídeos. Los representantes de esta familia tuvieron una radiación adaptativa en Europa, Asia y América del Norte desde el Eoceno superior al Pleistoceno superior.
En América del Sur, los restos fósiles asignados a la familia Gomphotheriidae se registran desde el Pleistoceno medio hasta el Pleistoceno superior y se reconocen como dos géneros distintos, Cuvieronius y Stegomastodon (Alberdi et al., 2002; Prado et al., 2002). Se considera que los gonfoterios pasaron desde América del Norte a América del Sur a través del istmo de Panamá, como integrantes del Gran Intercambio Biótico Americano (Webb, 1991). El registro sudamericano más antiguo incuestionable corresponde a los mastodontes de la localidad boliviana de Tarija (1,0-0,7 Ma, MacFadden et al., 1983; MacFadden, 2000).
Casamiquela et al. (1996) han sugerido que los gonfoterios habrían ingresado en América del Sur durante el Plioceno o comienzos del Pleistoceno, pero no hay registros fiables que confirmen esta hipótesis, ya que la posición estratigráfica de los restos del Mioceno de Madre de Dios (Brasil; Campbell et al., 2000, 2001) no está suficientemente clara. Por su parte, Alberdi y Prado (1995) y Prado et al. (2003, 2005) plantean dos posibles vías migratorias, siguiendo las ideas de Webb (1991): una por la Cordillera de los Andes para el género Cuvieronius y que podría llegar hasta Chile; otra por las planicies brasileñas, que seguiría el género Stegomastodon y que llegaría a su nivel más austral en las Pampas de Argentina. Recientemente, Frassinetti y Alberdi (2005) han revisado los gonfoterios chilenos y concluyen que parte del material asignado por los mismos autores en 2000 a Cuvieronius hyodon corresponde realmente a Stegomastodon. Prado et al. (2003) mantienen la validez de los géneros Cuvieronius y Stegomastodon en América del Sur y resaltan la homogeneidad de su dentición.
Por lo que concierne a la provincia argentina de Santiago del Estero, los hermanos Wagner recuperaron, a finales del siglo XIX, numerosos restos de vertebrados en las riveras del río Dulce, en los departamentos de Río Hondo, La Banda y Santiago del Estero capital. Esta fauna pleistocena fue dada a conocer por los hermanos Wagner en colaboración con Carlos Rusconi (Rusconi, 1967), indicando la presencia de numerosos taxones y citando la existencia de huesos trabajados tanto de animales actuales como extinguidos. Los restos de gonfoterios fueron mencionados preliminarmente (Gramajo, 1992), pero no habían sido estudiados en detalle.
El objetivo principal de este trabajo es el estudio sistemático de los restos de gonfoterios procedentes del entorno de la ciudad de Santiago del Estero, realizando una comparación morfométrica con los datos existentes de otras localidades en América del Sur. Asimismo, se han realizado análisis isotópicos que contribuyen a conocer las adaptaciones tróficas y ambientales de estos mastodontes.
Contexto estratigráfico
La provincia de Santiago del Estero se sitúa en el centro-norte de Argentina (figura 1). Los restos de gonfoterios estudiados proceden de los niveles de terraza del Pleistoceno superior de Río Dulce en la zona de La Banda, Río Hondo y Santiago del Estero capital (Rusconi, 1967). Esta región forma parte de una unidad morfológica denominada Llanura Chaco Pampeana, cuyo rasgo más característico es la ausencia casi total de relieve. El espesor de la sucesión sedimentaria no se mantiene uniforme a través de toda el área, identificándose varias subcuencas. La zona de la que provienen los restos estudiados se la conoce clásicamente como subcuenca de Alhuampa (Russo et al., 1979). El Río Dulce corre a lo largo de la falla de Huyamampa en sentido noroeste-suroeste. Al oeste de la falla, los niveles asignados al Mio-Plioceno están cubiertos por aproximadamente 30 metros de loess pampeanos (figura 2). Estos sedimentos consisten, en parte, en sedimentos terciarios removidos y, en parte, rodados del zócalo cristalino y rocas precámbricas. Esta sedimentación de material grueso está interrumpida por intercalaciones de depósitos de arenas finas y loess, seguida nuevamente de acumulaciones gruesas o rodados poco gastados. Los fósiles provienen de los niveles de loess (Gramajo, 1992).
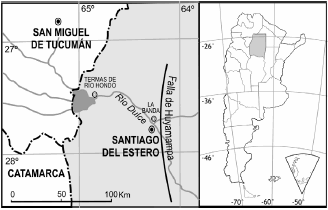
Figura 1. Situación geográfica de los afloramientos pleistocenos a lo largo del río Dulce, en el área de Río Hondo, La Banda y Santiago del Estero capital / geographic location of the Pleistocene outcrops along Dulce river in the area of Río Hondo, La Banda, and Santiago del Estero.

Figura 2. Corte esquemático de la geología del Río dulce y columna estratigráfica en la cantera de Los Morales (Río Dulce, Departamento La Banda, provincia de Santiago del Estero). Modificado de Gramajo (1992) / geological section and stratigraphic column at Los Morales quarry (Dulce river, La Banda Department, Santiago del Estero Province). Modified from Gramajo (1992).
Material y método
El material estudiado se encuentra depositado en el Museo de Ciencias Naturales y Antropológicas"Hermanos Emilio y Ducan Wagner" de Santiago del Estero (MCNA-SE), el Laboratorio de Antropología de la Universidad Nacional de Santiago del Estero (UNSE), el Museo de Ciencias Naturales y Antropológicas"Juan Cornelio Moyano" de Mendoza (MCNAM) y el Museo Argentino de Ciencias Naturales"Bernardino Rivadavia" de Buenos Aires (MACN), Argentina.
Los restos del MCNA-SE consisten en: cráneo bastante completo con colmillo izquierdo y fragmento derecho (n° 1), mandíbula sin ramas ascendentes (n° 8), fragmento de defensa sin extremos (n° 3), fragmento terminal de defensa robusta (n° 9), fragmento de defensa pequeña (n°s. 10 ó 12), fragmento de paladar con ambas series dentarias (puede corresponder al n° 13), cráneo incompleto con molariformes parcialmente destruidos y sin defensas, muy grande (n° 2 del listado y tiene además el n° 00165 del Museo), cráneo bastante deteriorado del que no se han podido tomar dimensiones porque era imposible tener acceso a él, fragmento de mandíbula derecha con restos de molar (n° 18), M3 superior derecho fragmentado (quizá n° 21), sínfisis y fragmento de mandíbula derecha con m2 muy gastado y m3 que puede corresponder al n° 6, fragmento de paladar con dos molares, dos M3 (quizá n° 14), mandíbula inferior izquierda con m3 (n° 23 del listado y n° 00151), fragmento de rama mandibular derecha con sínfisis y sin rama ascendente, con m3 (n° 19), mandíbula con la base del m3 roto (quizá n° 4 si se hubiera perdido el resto, ya que indicaba una mandíbula completa con ambas series dentarias), rama mandibular izquierda con restos de m2 y m3 (n° 28 y n° 00130), fragmento de sínfisis que conserva restos del m3 (n° 5), fragmento de mandíbula (n° 7 y n° 00143), maxilar con restos de M3 izquierdo y derecho (n° 30), fragmento de maxilar con ambas series dentarias completas (n° 15). Además, hay una mandíbula inferior izquierda, con restos de m2 y tres colinas de m3, y una mandíbula inferior izquierda con un m3 que no están en el listado facilitado por E.P. Tonni.
En el Laboratorio de Antropología de la UNSE, hay dos fragmentos de mandíbula, uno derecho con el alvéolo del m2 y uno izquierdo con el m3.
En el MCNAM, procedentes de La Banda: fragmento de cráneo juvenil (MCNAM-PV-87), fragmento de defensa (MCNAM-PV-35), fragmento de mandíbula juvenil (MCNAM-PV-82), m1 inferior izquierdo (MCNAM-PV-80), m2 inferior derecho (MCNAM- PV-78), m3 inferior izquierdo (MCNAM-PV- 79), m3 inferior izquierdo (MCNAM-PV-77), atlas (MCNAM-PV-81).
Los restos depositados en el MACN proceden del río Dulce en la zona de Río Hondo y La Banda: m2 inferior (MACN-6726), fragmento de mandíbula derecha, con la sínfisis, el alvéolo de m1 y el m2 (MACN- 19164), fragmento de mandíbula con los bordes del m3 izquierdo y m3 derecho bastante gastado (MACN- 18176), fragmento de maxilar con M3 izquierdo y fragmento de M2 (MACN-14016), fragmento de sínfisis relativamente alargado (MACN-5267), fragmento de mandíbula izquierda con un resto de m3 y el alvéolo de m2 (MACN-11689), rama mandibular izquierda muy alterada con restos de la sínfisis, el alvéolo de m2 y m3 saliendo (MACN-13009), rama mandibular derecha con un enorme alvéolo anterior y restos de un diente muy gastado, quizá un m3 (MACN-13011), dos M3 superiores derecho e izquierdo incompletos y dos fragmentos de defensa rectos que corresponden al mismo individuo (MACN- 14815).
Los materiales incluidos en la comparación corresponden a localidades de Brasil (Pains, Minas Gerais; Toca dos Ossos, Bahia; y Bonito, Mato Grosso) y Argentina (fundamentalmente de la región Pampeana), con buenos datos estratigráficos, y se encuentran depositados en las siguientes instituciones: Argentina: Museo de La Plata (MLP), Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" (MACN), y Museo Municipal "Dámaso Arce" Olavarria (MMO-MEDA), Buenos Aires; Brasil: Museo de Historia Naturais de Belo Horizonte (MHNBH) y Museo de la Pontifical Universidad Católica de Belo Horizonte (MUCBH); Francia: Museum National d'Histoire Naturelle de Paris (MNHNP); Dinamarca: Museum Zoologish de Copenhage (MZK); y Alemania: Naturkunde Museum de Berlín (NMB). Además se incluyen algunos restos de Uruguay, depositados en la Colección de Vertebrados del Departamento de Paleontología de la Universidad Nacional de la República, Montevideo (FCDPV).
En el estudio realizado, se sigue la metodología de Alberdi et al. (2002). Las dimensiones están tomadas con un calibre digital, van expresadas en milímetros y se detallan en las tablas 1 a 3. Las abreviaturas utilizadas son: L, longitud; A, anchura; 1 a 5, número de lofos/lófidos o colinas; T, talón; M/m, molar superior/ inferior; ca, cálculo aproximado; sn, sin número.
Se han realizado análisis bivariantes y análisis discriminante (AD) para los dientes. Concretamente, se analizaron 74 M3, 124 m3, 50 M2 y 58 m2, utilizando el programa SPSS versión 11.5 (para más datos sobre este método ver Reyment, 1991). Asimismo, se ha aplicado el índice de robustez de Osborn (1936) para los molares M2/m2 y M3/m3 (A máxima x 100 / L).
Los análisis de isótopos de oxígeno y carbono se realizaron a partir de una muestra de esmalte de diente, extraída de la parte anterior basal del primer lofo de M3. El estándar para los resultados de carbono es PDB y para el oxígeno V-SMOW con una precisión analítica de 0,2 ‰. También se realizó un análisis de 14C por AMS para obtener dataciones. Todos los análisis se llevaron a cabo en el Centrum voor Isotopen Onderzoek, Groninguen, Holanda.
Tabla 1 Dimensiones de los cráneos de Stegomastodon de Santiago del Estero. Parámetros seleccionados según Alberdi et al. (2002) / measurements (mm) of the Stegomastodon skulls from Santiago del Estero. Selected parameters following Alberdi et al. (2002): dorsal view: 1, longitud máxima del cráneo en el plano sagital / maximal length of the skull in the sagittal plane; 2, distancia entre el vértex y el plano superior de los orificios nasales / distance between the vertex and the upper plane of the nasal apertures; 3, altura máxima de los orificios nasales / maximal nasal height; 4, longitud del premaxilar en el plano sagital / premaxillary length in the sagittal plane; 5, anchura mínima entre las crestas temporales / minimum width between temporal crests; 6, anchura máxima suborbitaria / maximal suborbital width; 7, anchura de los premaxilares a nivel del orificio infraorbitario / premaxillary width at the level of the infra-orbital foramen; 8, anchura del proceso nasal / width of the nasal process; 9, anchura máxima del orificio nasal / maximal width of the nasal aperture; ventral view; 10, anchura entre los pseudomeatos auditivos externos / width between the external auditive pseudo-meatus; 11, anchura entre los bordes externos de las fosas de articulación mandibular / width between external margins of the mandibular articulation fossae; 12, anchura entre los bordes internos de las fosas de articulación mandibular / width between internal margins of the mandibular articulation fossae; 13, anchura entre los bordes antero-internos de los molares / width between antero-internal margins of the molars; 14, anchura entre los bordes postero-internos de los molares / width between postero-internal margins of the molars; 15, anchura de las coanas / width of the choana; 16, distancia sagital entre el plano anterior de los molares y la espina palatina / distance between the molar anterior plane and the palatine spine, sagittal plane; 17, distancia entre la espina palatina y el borde anterior del foramen mágnum / distance between the palatine spine and the anterior margin of the foramen magnum; 18, distancia entre las apófisis pterigoideas del esfenoides y el borde anterior del foramen mágnum / distance between the pterygoid process of the sphenoid and the anterior margin of the foramen magnum; 19, diámetro máximo antero-posterior de la fosa temporal a nivel de la arcada yugal / maximal antero-posterior diameter of the temporal fossa from yugal arcade; lateral view; 20, distancia entre el punto más elevado de la cresta temporal y el borde antero-inferior del pseudomeato auditivo externo / distance between the highest point of the temporal crest and the antero-inferior margin of the external auditive pseudo-meatus; 21, distancia entre el punto ventral de la órbita y el borde anteroinferior del pseudomeato auditivo / distance between the orbital ventral point and the antero-inferior margin of the external auditive pseudo meatus; 22, altura máxima de la órbita / maximal orbital height; 23, distancia entre el punto más elevado de la cresta temporal y el borde alveolar / distance between the highest point of the temporal crest and the alveolar margin; 24, distancia entre el borde antero-inferior del premaxilar y la apófisis pterigoidea / distance between the premaxillary antero-inferior margin and the pterygoid process; 25, distancia entre la punta del proceso zigomático del frontal y la apófisis pterigoidea / distance between the extreme of the frontal zygomatic process and the pterygoid process; 26, distancia entre el vértex y la apófisis pterigoidea / distance between the vertex and the pterygoid process; 27, distancia entre el borde posterior de los cóndilos occipitales y la apófisis pterigoidea / distance between the posterior margin of the occipital condyles and the pterygoid process; 28, diámetro máximo antero-posterior de los premaxilares / Antero-posterior maximal diameter of the premaxilla; posterior view or external cranial vault; 29, anchura máxima de la caja craneana / maximal width of the cranial vault; 30, anchura de la fosa del ligamento nucal / width of the nucal ligament fossa; 31, anchura entre los bordes latero-externos de los cóndilos occipitales ; 32, anchura entre los bordes latero-internos de los cóndilos occipitales / width between latero-external margins of the occipital condyles; 33, altura máxima del cráneo (del vértex a la parte externo-inferior de los cóndilos occipitales / maximal height of the skull (from the vertex to the externo-inferior border of the occipital condyles); 34, distancia entre el vértex y la cima de la fosa del ligamento nucal / distance between vertex and the top of the nucal ligament fosse; 35, longitud de la cresta de la fosa del ligamento nucal / length of the crest of the nucal ligament fossa.
Tabla 2 Dimensiones de las mandíbulas de Stegomastodon de Santiago del Estero. Parámetros seleccionados siguiendo a Alberdi et al. (2002) / measurements (mm) of the Stegomastodon mandibles following the nomenclature in Alberdi et al. (2002): 7, longitud de la serie dentaria / dental row length; 8, forámenes mentonianos / mentonial foramina; 9, sínfisis mandibular / mandibular symphysis; 10, longitud de la sínfisis en el plano sagital / symphysis length in sagittal plane; 10 bis, altura al mismo nivel / height at the same level; 11, fosa pterigoidea / pterigoid fossa; a, longitud máxima en el plano sagital / maximal length in sagittal plane; b, altura a nivel anterior de la serie molar / mandibular height anterior to molar series; c, altura a nivel posterior de la serie molar / mandibular height posterior to molar series; d, anchura máxima de la rama ascendente / maximal width of the ascending ramus; e, distancia mínima entre el borde antero interno de los molares / minimal distance between antero-internal molar edges; a, ángulo que forma el borde lateral de la sínfisis o cresta interalveolar con la superficie oclusal / angle formed by the lateral edge of the symphysis and the occlusal surface; f, distancia mínima entre los bordes postero-internos de los molares / minimal distance between postero-internal molar edges.
Tabla 3 Dimensiones de los molares de Stegomastodon de Santiago del Estero. Parámetros seleccionados siguiendo a Alberdi et al. (2002) / measurements (mm) of the upper and lower molars of Stegomastodon from Santiago del Estero (Argentina), following Alberdi et al. (2002): L, longitud máxima / maximal length; A1, A2, A3, A4 y A5, anchuras máximas de los respectivos lofos/lófidos / width of the respective lophids; T: talón; A>, máxima anchura / talon; maximum width.
Paleontología sistemática
Antecedentes
Los primeros estudios sistemáticos sobre gonfoterios sudamericanos se remontan a Cuvier (1806) quien los agrupó, según su procedencia geográfica, en "mastodonte des cordillères" y "mastodonte humboldien". Cabrera (1929) incluyó las formas bunodontas y brevirrostras sudamericanas en la Familia Gomphotheriidae, dentro de la Superfamilia Elephantoidea, y creó la Subfamilia Cuvieroniinae para las que presentan torsión y banda de esmalte en los colmillos superiores. Hoffstetter (1952) también consideró la presencia de dos subfamilias entre los gonfoterios sudamericanos: Cuvieroniinae Cabrera, 1929 y Anancinae Hay, 1922. Sin embargo, Simpson y Paula Couto (1957) consideraron una sola subfamilia, Anancinae, pero apuntando la posibilidad de incluirlos todos directamente en la Familia Gomphotheriidae, ya que la división a nivel subfamiliar no era satisfactoria con los datos disponibles. Más tarde, Tassy y Shoshani (1996: Apéndice B) incluyeron los gonfoterios de América del Sur en la Familia Gomphotheriidae y en la Subfamilia Cuvieroniinae. Al mismo tiempo, Tassy (1996) presentó un análisis filogenético en el que reconoce dos grupos de gonfoterios: los del Viejo Mundo y los del Nuevo Mundo, considerando que las formas americanas, en general, se relacionan más estrechamente con los gonfoterios tetralofodontos que con los trilofodontos de Eurasia. Asimismo, propone a Sinomastodon de China como posible antecesor de las formas brevirrostrinas de América del Sur siguiendo a Tobien et al. (1986, 1988), postura no compartida por Chen (1999). Por otra parte, los trabajos de Lambert (1996) y Dudley (1996) indican claramente que el género Anancus nunca llegó a América del Norte, lo que obviamente excluye la presencia de anancinos en América del Sur.
Nosotros mantenemos la postura expresada por Alberdi y Prado (1995) y Prado et al. (2002, 2003) de que no hay evidencias significativas que sustenten la separación de las formas sudamericanas en dos subfamilias diferentes.
Los gonfoterios de América del Sur están caracterizados por la presencia de un cráneo braquicéfalo con una tendencia a alcanzar una forma elefantoidea y una mandíbula de tipo brevirrostrina. La longitud y torsión de las defensas superiores son variables. Pueden presentar o no banda de esmalte, que muchas veces desaparece en los especimenes adultos. Los molares son bunodontos y braquidontos o subhipsodontos y suelen presentar una disposición angular entre los conos principales -pretrite y postrite- en las últimas colinas (lofos/lófidos), con figuras treboladas en la superficie oclusal, más o menos complicadas, producidas por el desgaste. Los M2 son trilofodontos con un talón a veces desarrollado. El M3 varía desde tetralofodonto a heptalofodonto.
Hasta ahora, los restos de gonfoterios de Argentina se han identificado como Stegomastodon platensis (Alberdi et al., 2002, 2004; Prado et al., 2003, 2005).
Familia GOMPHOTHERIIDAE Cabrera, 1929
Género Stegomastodon Pohlig, 1912
1842. Mastodon; Lesson, p. 157.
1906. Mastodon; Winge, p. 48.
1928. Bunolophodon; Spillmann, p. 67.
1929. Notiomastodon Cabrera, p. 90-96, fig. 2 y 4.
1950. Stegomastodon (Haplomastodon) Hoffstetter, p.22, fig. 2 y 3.
1952. Haplomastodon (Aleamastodon) Hoffstetter (en parte), p.208.
1957. Haplomastodon Hoffstetter; Simpson & Paula Couto, p. 166.
Especie tipo. Mastodon mirificus Leidy, 1858, p. 10.
Distribución estratigráfica y geográfica. El género se registra desde el Plioceno superior hasta el Pleistoceno inferior en las regiones centrales y occidentales de América del Norte. En América del Sur, está presente en el Pleistoceno medio (?) y superior de Brasil, Argentina, Paraguay y Uruguay y en el Pleistoceno superior de Ecuador, Perú, Chile, Colombia y Venezuela.
Diagnosis. Gomphotheriidae con cráneo corto y alto, tipo elefantoideo, menos deprimido que Cuvieronius. Mandíbula brevirrostra con dientes intermedios bunodontos y trilofodontos (P3/p3, P4/p4, M1/m1, M2/m2). M3/m3 con moderada alternancia de los conos externos e internos sobre las colinas o lofos/lófidos posteriores, moderadamente oblicuos. Patrón de figuras de desgaste en doble trébol que puede variar desde tréboles sencillos a complicados. Las defensas varían de ligeramente curvadas a casi rectas, sin esmalte, aunque puede aparecer en algunos ejemplares juveniles.
Stegomastodon platensis (Ameghino), 1888
1845. Elephas humboldtii Blainville, p. 249, nec Cuvier, 1824, p. 527.
1888. Mastodon platensis Ameghino, p. 7.
1888. Mastodon superbus Ameghino, p. 7; Casamiquela et al., 1996, p. 316.
1888. Mastodon bonaerensis Moreno, p.17, nomen nudum.
1889. Mastodon rectus Ameghino, p.643.
1891. Mastodon maderianus Ameghino, p. 243.
1929. Notiomastodon ornatus Cabrera, p. 91; Casamiquela et al., 1996, p. 316.
Véase Cabrera (1929) y Simpson y Paula Couto (1957) para ampliar las sinonimias.
Holotipo. MLP-8-63, una defensa adulta de San Nicolás de los Arroyos (provincia de Buenos Aires). Notificado por Ameghino (1888:7) e ilustrado por Ameghino (1889).
Distribución estratigráfica. Edades mamífero Ensenadense y Lujanense (Pleistoceno medio y superior).
Distribución geográfica. Es el gonfoterio más austral, típico de la Región Pampeana, Argentina, en especial en las provincias de Buenos Aires, Córdoba, Santa Fe, Entre Ríos y Santiago del Estero. También está citado en Uruguay (Mones y Francis, 1973) y posiblemente esté presente en Paraguay (Cabrera, 1929; Simpson y Paula Couto, 1957).
Diagnosis. Cráneo corto y alto, similar al de la especie tipo. Los molares, M3/m3, tetralofodontos o pentalofodontos; la morfología oclusal de los molares varía desde simple a complicada debido a la presencia de cónulos y coneletes centrales. Las defensas son largas y relativamente finas, sin banda de esmalte y algo más curvadas en los machos que en las hembras.
Descripción. Los restos de Gomphotheriidae provenientes de la zona de Santiago del Estero están representados por fragmentos de cráneos, mandíbulas, defensas y dientes aislados (figuras 3 y 4).
Figura 3. Stegomastodon platensis del entorno de la ciudad de Santiago del Estero / Stegomastodon platensis from Santiago del Estero city environments. 1, cráneo, vista dorsal o facial (MCNA-SE n° 1) / skull, dorsal or facial view (MCNA-SE n° 1); 2, idem, vista lateral izquierda / idem, left lateral view; 3, idem, M3 en vista oclusal / idem, M3, occlusal view; 4, cráneo, vista ventral (MCNA-SE n° 2 y 00165) / skull, ventral view (MCNA-SE n° 2 and 00165); 5, fragmento de maxilar con los M3, vista oclusal (MCNA-SE n° 14) / maxillary fragment with M3, occlusal view (MCNA-SE n° 14); 6, fragmento central de defensa sin banda de esmalte (MCNA-SE nº 3) / central tusk fragment without enamel band (MCNA-SE n° 3); 7, fragmento terminal de defensa sin banda de esmalte (MCNA-SE nº 9) / terminal tusk fragment without enamel band (MCNA-SE n° 9).
Figura 4. Stegomastodon platensis del entorno de la ciudad de Santiago del Estero / Stegomastodon platensis from Santiago del Estero city environments. . 1, mandíbula sin ramas ascendentes, vista oclusal (MCNA-SE n°. 8) / mandible without ascending rami, occlusal view (MCNA-SE n°. 8); 2, idem, vista lateral izquierda / idem, left lateral view; 3, fragmento de paladar juvenil con M1 y M2, vista oclusal (MCNA- SE n° 3) / juvenile palatal fragment with M1-M2, occlusal view (MCNA-SE n° 3); 4, fragmento mandibular derecho con restos de la sínfisis (MCNA-SE s/n) / right mandibular fragment with part of the symphysis (MCNA-SE s/n); 5, idem, vista lateral derecha / idem, right lateral view; 6, m3 derecho, vista oclusal (MCNA-SE s/n) / right m3, occlusal view (MCNA-SE s/n); 7, fragmento de m3 derecho, vista oclusal (MCNAM-PV-79) / right m3 fragment, occlusal view; (MCNAM-PV-79; 8, idem, vista lateral / idem, lateral view; 9, M3 izquierdo, vista oclusal (MACN-14016/96) / left M3, occlusal view (MACN-14016/96); 10, fragmentos de M3 izquierdo y derecho, vista oclusal (MACN- 14815/93) / left and right M3 fragments, occlusal view (MACN-14815/93); 11, extremos apicales de defensas sin banda de esmalte (MACN- 14815/93) / apical tusk fragments without enamel band (MACN-14815/93).
El cráneo MCNA-SE nº 1 pertenece a un individuo adulto, pero relativamente pequeño, con los dos M3, de cuatro colinas y un talón pequeño, con las figuras prácticamente perdidas por desgaste; las medidas de los dientes son aproximadas por tener la mayor parte de los bordes del esmalte perdidos (tablas 1 y 3, figuras 3.1, 3.2 y 3.3). Conserva un resto de la defensa derecha con una sección de 55 x 61 mm. El cráneo tiene el perfil tipo elefantoideo, no tan pronunciado como en algunos grandes cráneos de la Pampa argentina, con los premaxilares paralelos entre sí, ligeramente divergentes. El lado izquierdo está prácticamente completo, incluido el arco zigomático; el derecho conserva un resto de la defensa y la fosa y el ligamento nucal. El ejemplar, MCNA-SE nº 2 y n° 00165, es un cráneo de tamaño muy grande, con la base del molar derecho y el izquierdo muy desgastado, pero completo con cinco colinas y talón, con el cíngulo mesial e interno fuerte, cemento abundante y ángulo entre petrites y postrites en las tres últimas colinas (tabla 3). El fragmento de la defensa izquierda conservado en el alvéolo tiene un diámetro máximo de 126 mm; el alvéolo derecho tiene una sección redondeada de 133 mm de diámetro. El cráneo es muy ancho y difícil de medir en posición dorsal (tabla 1). La parte ventral del cráneo tiene una morfología similar a las de los restos procedentes de la Pampa Argentina (figura 3.4). El fragmento de paladar juvenil, MCNA-SE nº 13, tiene los M1, M2 y las dos primeras colinas de los M3 aflorando (tabla 3; figura 4.3). Dorsalmente, se observan las huellas de las defensas prácticamente paralelas. Los dientes conservan restos de cemento abundante, sobre todo los M2. Estos tienen muy desgastada la primera colina en la que se observan cónulos centrales a derecha e izquierda de la línea media y figuras de desgaste complicadas (más en el derecho que en el izquierdo). Los M1 están muy gastados, teniendo perdida casi una colina. Las dos colinas primeras de los M3 tienen división ternaria y gran cantidad de cónulos centrales, sin desgastar. Otro fragmento de paladar, MCNA-SE nº 14 (figura 3.5), muestra los M3 con cinco colinas y talón fuerte (tabla 3); hay algo de pticodontia sobre el molar izquierdo; figuras, línea media y cíngulo mesial perdidos por el desgaste.
Otros fragmentos maxilares están más incompletos. El MCNA-SE nº 30 muestra la base del alvéolo del M3 derecho con una longitud aproximada de 200 mm. En el MCNA-SE nº 15, fragmento de maxilar con ambas series dentarias completas y fuertes, los M3 tienen cuatro colinas y talón fuerte. Sobre la tercera y la cuarta colina se observa anancoidía más acusada sobre el diente izquierdo, línea media marcada y cónulos centrales a ambos lados de la línea media; tienen cíngulo mesial fuerte y no se observa cíngulo lateral (tablas 1 y 3).
Entre los molares aislados, el M3 MCNA-SE nº 21 (?) es robusto, ancho y corto y conserva las tres primeras colinas; la 3ª presenta una cierta anancoidía, figuras treboladas a ambos lados de la línea media y esmalte muy grueso. El M3 MACN 14016-96 es de gran tamaño (tabla 3), con desgaste iniciado en las dos primeras colinas; línea media, figuras treboladas y cemento abundante. Este molar figura en el inventario como Cuvieronius, pero no hay ningún dato que sustente esta afirmación (figura 4.9). Con el número MACN 14815-93 se incluyen dos fragmentos de M3, derecho e izquierdo, y dos fragmentos distales de incisivos. Los M3 están incompletos, con talón fuerte; las figuras treboladas a ambos lados de la línea media; no se ven restos de cíngulo y tienen cemento muy abundante (figura 4.10). Las defensas son rectas. El fragmento más largo tiene una longitud de 590 mm; sus diámetros mayores son 89 x 75 mm, en la mitad 83 x 65 mm y en el ápice 38 x 33 mm. El fragmento más corto de defensa, con el ápice alterado, tiene una longitud de 490 mm, diámetros máximos 80 x 68 mm y en el ápice 51 x 37,5 mm. En ambos casos, el ápice es subtriangular y afilado con su superficie pulida por desgaste (figura 4.11).
Entre otros fragmentos de defensas, el terminal MCNA-SE nº 9 tiene una longitud de 820 mm, es robusto y ligeramente curvado; sus diámetros en la base son 148 x 135 mm y en el ápice 78 x 90 mm, también pulido por desgaste (figura 3.7). El fragmento central MCNA-SE Nº 3 es ligeramente curvado, de 718 mm de longitud y 140 x 110 mm de diámetro en el extremo basal; en la zona de rotura los diámetros son 114 x 113 mm (figura 3.6). El fragmento MCNASE Nº 10 (ó 12) es el ápice de una defensa pequeña, de aspecto juvenil. El fragmento MCNAM-PV 35 es robusto, de 166,5 mm de longitud, sin torsión, con un diámetro > 98 mm y una banda de esmalte de 44 mm.
La mandíbula MCNA-SE nº 8 está casi completa, sin ramas ascendentes (figuras 4.1 y 4.2), con los dos m3 gastados, con cinco colinas y talón y los alvéolos de los m2 (tabla 3). Los m3 presentan figuras treboladas más complicadas externamente; las colinas 4ª y 5ª poco gastadas y con una cierta angulación entre los pretrite y los postrites cónulos centrales, línea media perdida en las primeras colinas. El cíngulo mesial se ha perdido por el desgaste, el labial es fuerte hasta el talón. Hay restos de cemento. La mandíbula conserva dos orificios mentonianos a nivel de la parte mesial del alvéolo del m2 (tabla 2). El fragmento MCNA- SE nº 18 (tabla 2) tiene restos del m3, posiblemente de cuatro colinas y talón (tabla 3), con figuras complicadas; tiene cíngulo externo muy robusto hasta el talón y abundante cemento. En la hemimandíbula MCNA-SE nº 6 (figura 4.4 y 4.5; tabla 2), el m2, muy gastado, tiene tres colinas, y el m3, cinco colinas y talón (tabla 3), parcialmente rotas en los postrites. En el fragmento MCNA-SE s/n (figura 4.6), el m3 tiene cinco colinas y talón pequeño; presenta una línea media más o menos perdida por el desgaste, figuras complicadas y cemento abundante (tablas 2 y 3). En otro fragmento s/n, se conservan el alvéolo del m2 y las tres primeras colinas del m3, con cíngulo mesial muy acusado y figuras sólo iniciadas en la primera colina. En el ejemplar MCNA-SE nº 23 y 00151, el m3 tiene cinco colinas y talón pequeño y gran número de cónulos centrales; línea media marcada, el cíngulo externo es fuerte, hasta el talón, presentando coneletes laterales en el borde de los valles internos (tabla 3). Presenta dos forámenes mentonianos (tabla 2), en contraste con el fragmento MCNA-SE nº 19 que presenta uno, obturado (tablas 2 y 3). El m3 de este ejemplar es enorme; tiene cinco colinas y talón, con coneletes en el borde interno de los valles y cíngulos mesial y lateral fuertes hasta el talón. Las dimensiones del alvéolo del m3 y algunos parámetros mandibulares del fragmento MCNA-SE s/n están incluidos en las tablas 3 y 2, respectivamente. En la rama ascendente, incompleta, se observa una fosa pterigoidea profunda y un orificio mentoniano justo en el nivel de rotura de la parte más mesial. Este fragmento podría corresponder a alguna de las mandíbulas relacionadas por E.P. Tonni que no hemos podido localizar en el Museo. En MCNA-SE nº 28 y 00130, se han podido tomar algunas dimensiones de la mandíbula (tabla 2) que presenta dos orificios mentonianos a nivel mesial del m2; el m3 (tabla 3) tiene cinco colinas y un talón pequeño, se observa la línea media y las figuras parecen algo complicadas. En MCNA-SE nº 5, se observan dos orificios mentonianos y se han podido tomar algunas dimensiones de la mandíbula (tabla 2). En MCNA-SE nº 7 y 00143 (tablas 2 y 3), también hay dos forámenes mentonianos a nivel mesial del m2 (alvéolo); el m3, enorme, tiene figuras de desgaste muy complicadas.
Los fragmentos mandibulares s/n de la UNSE (tabla 2) muestran un m3 grande, con línea media marcada, el cíngulo mesial perdido por el desgaste y el lateral externo hasta el talón, abundante cemento y ángulo entre el petrite y postrite a partir de la 4ª colina; un m2 muy gastado y un m3 saliendo, con tres colinas; en este ejemplar se observar una fosa pterigoidea prácticamente obturada; el cóndilo articular y el proceso coronoides están recubiertos de sedimentos y cantos rodados.
Los fragmentos mandibulares MACN 19164-27 (tabla 2), MACN 11689-24 (tabla 2), MACN 13009-25 (tablas 2 y 3), MACN 13011-27 y MACN 13661-97 (tablas 2 y 3) están más o menos completos, con restos de m2 y/o m3 con distintos grados de desgaste. En el m2 se observa la línea media, los cónulos centrales formando parte de las figuras treboladas; cíngulo fuerte, más reducido el externo que el interno; y cemento. Los m3 tienen cuatro colinas y un talón muy robusto; el más completo presenta línea media clara, cónulos centrales cerrando los valles, figuras treboladas enormes, más las externas que las internas, cemento abundante, cíngulo mesial y algo de reborde externo. En el último ejemplar hay un foramen mentoniano grande.
Los especimenes MCNAM-PV 82 y PV 87 son fragmentos de mandíbula y cráneo juveniles, respectivamente, posiblemente del mismo individuo, el primero con los alvéolos de p2 y p3 derechos y el inicio de la rama izquierda (tabla 2) y el segundo con restos de raíces quizá correspondientes al P3.
El MCNAM-PV 80 es un m1 izquierdo con tres colinas y talón enorme, con cíngulo mesial, línea media y figuras perdiéndose. El m2 MCNAM-PV 13 tiene el cíngulo mesial perdido por el desgaste y el lateral no se observa; talón con las figuras desapareciendo por desgaste. El m2 MCNAM-PV 78 sólo conserva las dos últimas colinas (2ª y 3ª), con escaso cemento. El MCNAM-PV 79 es un m3 muy curvado, con línea media marcada, cónulos centrales externos a la línea media y con cemento abundante; no se observa cíngulo (figuras 4.7 y 4.8). El MACN 6726 es un m2 de tres colinas y talón roto, con línea media, cónulos centrales formando figuras treboladas muy desgastadas y restos de cemento (tabla 3). El m3 MCNAM- PV 77, muy gastado e incompleto anteriormente, tiene la parte distal con figuras muy complicadas, con un cíngulo interno muy desarrollado que refuerza el talón, línea media y cónulos centrales perdidos, cemento abundante. El m3 MACN 18176- 94 presenta cuatro lófidos y talón enorme con dos coneletes; el cíngulo, no muy claro, es más acusado en el borde interno (tabla 3).
El único elemento postcraneal revisado es un atlas típico de gonfoterio (MCNAM-PV 81), con un foramen cerrado y otro abierto.
Resultados estadísticos
Los análisis bivariante y multivariante de los M3 y m3 estudiados, comparados con los datos de otras localidades sudamericanas de Argentina, Chile, Bolivia, Perú, Ecuador y Brasil (Alberdi et al., 2002, 2004; Prado et al., 2002), sitúan la mayoría de los restos dentarios de los gonfoterios de Santiago del Estero entre los más grandes de América del Sur y a la vez los más grandes de Argentina, Brasil e incluso Bolivia (figuras 5.1, 5.2 y 5.3). Los rasgos morfológicos descritos en el material estudiado, especialmente la zona mesial del premaxilar, donde afloran los alvéolos de las defensas apenas divergentes, y las defensas rectas y sin banda de esmalte en los adultos, permiten atribuirlos al género Stegomastodon. Los análisis bivariantes de los molares, muy variables en forma y tamaño, presentan una gran dispersión de los restos de esta localidad (figuras 5.1 y 5.2). Esto se debe a la gran variabilidad observada en todos los tipos de dientes analizados, similar a la observada en estudios previos de proboscideos y de otras poblaciones de América del Sur.

Figura 5. Análisis bivariante y discriminante de los últimos molares superiores e inferiores (M3/m3) de Stegomastodon platensis de Santiago del Estero comparados con los de otras localidades de Argentina, Brasil y Bolivia (Tarija) / Bivariant and discriminant analyses of the upper and lower last molars (M3/m3) of Stegomastodon platensis from Santiago del Estero compared with those from other localities in Argentina, Brazil, and Bolivia (Tarija). 1, diagrama bivariante de longitud y anchura mayor de M3 / bivariant analysis maximun length/width of M3; 2, diagrama bivariante de longitud y anchura mayor de m3 / bivariant analysis maximum length/width of m3; 3, análisis discriminante de los m3. AR, ejemplares de localidades argentinas; BR, ejemplares de Brasil; BOL y TAR, ejemplares de Tarija (Bolivia); SdE, restos del entrono de Santiago del Estero. ��, centroides / discriminant analysis of m3. AR, Argentinean remains; BR, Brazilian remains; BOL and TAR, remains from Tarija (Bolivia); SdE, remains from Santiago del Estero. ��, centroid
En el caso de los análisis discriminantes realizados para ver como se agrupan los restos dentarios de las localidades de Santiago del Estero con respecto a las poblaciones de proboscideos ya estudiadas de América del Sur, se observa que los terceros molares inferiores (m3), los más numerosos (9 ejemplares enteros), se asocian mayoritariamente con las poblaciones de Stegomastodon platensis (figura 5.3), quedando clasificados correctamente el 71,7% de los casos (tabla 4). Estos resultados confirman la presencia de Stegomastodon platensis en esta localidad. Sin embargo, hay que hacer constar que no es fácil establecer las diferencias entre las dos especies del género Stegomastodon: S. waringi y S. platensis, ya que la morfología del cráneo es bastante similar y la variabilidad morfológica y métrica de los molares es muy alta, por lo que sólo se pueden diferenciar teniendo una buena representación de los mismos. No obstante, en el conjunto de las poblaciones ricas en restos, se observan dos tendencias morfológicas claras. En el caso de las poblaciones de Stegomastodon platensis, la morfología de la superficie oclusal indica una mayor proporción de dientes con figuras treboladas muy complejas y pticodontia marcada en el esmalte de las mismas. En el caso de Stegomastodon waringi, las figuras treboladas son más simples en general y la pticodontia es mucho más reducida. A la vista de estas diferencias, identificamos los restos de Santiago del Estero como Stegomastodon platensis debido al alto número de dientes con figuras treboladas complicadas.
Tabla 4. Resultados de clasificación del análisis discriminante de los m3 (casos desagrupados) de Stegomastodon de Santiago del Estero frente a los de Stegomastodon platensis (1) de Argentina, Stegomastodon waringi (2) de Brasil y Cuvieronius hyodon (3) de Tarija (Bolivia) / classification results of the discriminant analysis of m3 of Stegomastodon from Santiago del Estero with respect to Stegomastodon platensis (1) Argentina, Stegomastodon waringi (2) from Brazil y Cuvieronius hyodon (3) from Tarija (Bolivia).

Los análisis comparativos de los índices de robustez de los molares, elaborados siguiendo a Osborn (1936) y comparados con los de otras poblaciones de América del Sur, indican en el caso de los m3 (los mejor representados) una distribución y una media similares a las de otras poblaciones de Argentina (figura 6). Debido a que todos los restos encontrados hasta ahora en Argentina corresponden a Stegomastodon platensis y que los caracteres morfológicos de los restos de Santiago del Estero son muy similares consideramos estos últimos como pertenecientes a esta especie.

Figura 6. Índice de robustez de los m3 de Santiago del Estero, según Osborn (1936), comparado con los restos de otras localidades de Argentina, Brasil y Bolivia (Tarija) / robustness index of the m3 from the Santiago del Estero, following Osborn (1936), compared with other samples from Argentina, Brazil and Bolivia (Tarija).
Resultados isotópicos
Los resultados de los análisis isotópicos de 14C e isótopos estables del oxígeno y del carbono han permitido datar los restos e inferir características ambientales y dietarias de estos animales.
Entre el material revisado, la datación de un fragmento de esmalte de molar superior por medio del análisis AMS 14C dio un resultado de 19.900 ± 120 AP, confirmando la edad Pleistoceno superior de estos niveles fosilíferos.
En cuanto a los análisis isotópicos, los estudios recientes de Sánchez et al. (2004) han puesto de manifiesto las preferencias dietarias de las especies de Stegomastodon de América del Sur. Los datos obtenidos para los ejemplares de Santiago del Estero indican, por primera vez, una adaptación de S. platensis a una dieta estricta de plantas C4, similar a la de S. waringi de Ecuador (en Brasil y Uruguay se infiere una dieta mixta - C3 para esta especie). Por su parte, S. platensis de la provincia de Buenos Aires presenta una especialización desde el Pleistoceno medio, con una dieta mixta, hasta el Pleistoceno tardío, con una dieta exclusiva C3 (tabla 5), presentando una estrategia dietaria oportunista (Sánchez et al., 2004), de forma similar al elefante africano actual (Bocherens et al., 1996). El resultado obtenido para Santiago del Estero se ajusta al modelo de variación latitudinal propuesto por Sánchez et al. (2004), quienes sugieren una transición entre plantas C4 a C3 en torno a 35° y 41º de latitud Sur. Los datos isotópicos obtenidos sobre pastizales actuales del Hemisferio Norte sugieren un cambio transicional entre pastos C4 y C3 en torno a 40-45º N (Ehleringer et al., 1997). Recientemente, MacFadden et al. (1999), en base al registro de Equus de ambas Américas, verifican que el gradiente latitudinal es simétrico hacia ambos lados del ecuador.
Tabla 5. Estadísticos descriptivos de los resultados de los análisis de δ18O y δ13C de Stegomastodon platensis y S. waringi, comparados con los datos de Sánchez et al., (2003, 2004). DS, desviación standard / univariate statistics for Stegomastodon platensis and S. waringi from the results of d18O and d13C analyses, compared with data from Sánchez et al. (2003, 2004). SD, standard deviation; O, oxígeno / oxygen; C, carbono / carbon.
Por su parte, los valores de los isótopos del oxígeno varían de acuerdo con la latitud y son fuertemente influenciados por la altitud. En la tabla 5, se observa que los valores del oxígeno para S. platensis de la provincia de Buenos Aires varían desde una media de 29,46 en el Pleistoceno medio hasta una media de 30,27 en el Pleistoceno tardío. Por la latitud de Santiago del Estero y considerando que se encuentra a 200 metros sobre el nivel del mar, cabría esperar un valor superior al del Pleistoceno tardío de Buenos Aires. Sin embargo, el bajo valor obtenido (27,73) se corresponde seguramente con un intervalo cercano al del máximo glacial, lo cual es coherente con la datación obtenida por AMS 14C.
Conclusiones
A la vista de los datos existentes y las discusiones previas, podemos identificar los restos de gonfoterios procedentes del área de río Dulce, en el entorno de La Banda, Río Hondo y Santiago del Estero capital, como Stegomastodon platensis debido a los caracteres de los restos craneales existentes, así como a la mayor proporción de restos dentarios que presentan figuras de desgaste treboladas complicadas y acompañadas de pticodontia. Además, el índice de robustez de los m3 inferiores es equivalente a los resultantes de las distintas poblaciones de Stegomastodon platensis conocidas en Argentina.
La datación radiométrica de 14C del esmalte de diente (sistema AMS) confirma una edad Pleistoceno tardío para estos depósitos. Los datos de isótopos estables del carbono obtenidos para Santiago del Estero indican, por primera vez, una adaptación de Stegomastodon platensis a una dieta estricta de plantas C4, corroborando la variación latitudinal en la dieta propuesta previamente para los gonfoterios sudamericanos. La medición de los isótopos del oxígeno indica, en función de la latitud y la altitud, un momento más frío que el actual.
Agradecimientos
Expresamos nuestro sincero agradecimiento a E.P. Tonni y a J. Laza (Universidad y Museo de La Plata) por proporcionarnos el listado de piezas revisadas por ellos en el Museo "Emilio y Duncan Wagner" de Santiago del Estero. A M.D. Drube, director del Museo "Emilio y Duncan Wagner", por su amabilidad y disposición en todo momento. A A. Kramarz del Museo Argentino de Ciencias Naturales, Buenos Aires, por facilitar el estudio del material a su cargo. A J. Muñoz, del Laboratorio de Fotografía del MNCN, por la realización de las fotografías y a J. Arroyo por los dibujos. A G. Farías por el arreglo de las figuras 1 y 5. Este trabajo ha sido financiado por el Proyecto Conjunto de Investigación AECI España-Argentina (2000-2002), los Proyectos PB94-0071, PB97- 1250, BTE2001-1684 y CGL2004-00400/BTE de la DGICYT, España, y las Subvenciones de la Universidad Nacional del Centro de la Provincia de Buenos Aires y el Proyecto PIP-02773, CONICET y ANPCYT PICTO 04-11503, Argentina.
Bibliografía
1. Alberdi, M.T., Cartelle, C. y Prado, J.L. 2002. El registro de Stegomastodon (Mammalia, Gomphotheriidae) en el Pleistoceno superior de Brasil. Revista Española de Paleontología 17: 217-235. [ Links ]
2. Alberdi, M.R. y Prado, J.L. 1995. Los mastodontes de América del Sur. En: M.T. Alberdi, G. Leone y E.P. Tonni (eds.), Evolución biológica y climática de la Región Pampeana durante los últimos 5 millones de años. Un ensayo de correlación con el Mediterráneo occidental. Monografías Museo Nacional de Ciencias Naturales, CSIC, España 12: 277-292. [ Links ]
3. Alberdi, M.T., Prado, J.L. y Salas, R. 2004. The Pleistocene Gomphotheriidae (Proboscidea) from Peru. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 231: 423-452. [ Links ]
4. Ameghino, F. 1888. Rápidas diagnosis de algunos mamíferos fósiles nuevos de la República Argentina. Obras Completas, Buenos Aires V: 469-480. [ Links ]
5. Ameghino, F. 1889. Contribución al conocimiento de los mamíferos fósiles de la República Argentina. Academia Nacional Ciencias (Córdoba), Actas 6: 1-1027. [ Links ]
6. Ameghino, F. 1891. Mamíferos y aves fósiles argentinos. Especies nuevas, adiciones y correcciones. Revista Argentina de Historia Natural 1: 240-259. [ Links ]
7. Blainville, H.M.D. de 1845. Ostéographie ou description iconographique comparée du squelette et du système dentaire des mammifères. III. Des éléphants. Paris, J. B. Baillière et Fils 1-367. [ Links ]
8. Bocherens, H.P., Koch, L., Mariotti, A., Geraads, D. y Jaeger, J.J. 1996. Isotopic biogeochemistry (13C, 18O) of mammalian enamel from African Pleistocene hominid sites. Palaios 11: 306- 318. [ Links ]
9. Cabrera, A. 1929. Una revisión de los Mastodontes Argentinos. Revista del Museo de La Plata 32: 61-144. [ Links ]
10. Campbell, K. E. Jr., Fraily, C.D. y Romero-Pittman, L. 2000. The late Miocene Gomphothere Amahuacatherium peruvium (Proboscidea: Gomphotheriidae) from Amazonian Peru: implications for the Great American faunal Interchange. Boletín de Estudios Regionales, IGEMMET D 23: 1-152. [ Links ]
11. Campbell, K.E. Jr., Heizler, M., Fraily, C.D., Romero-Pittman, L. y Prothero, D.R. 2001. Upper Cenozoic chronostratigraphy of the southwestern Amazon Basin. Geology 29: 595-598. [ Links ]
12. Casamiquela, R.M., Shoshani, J. y Dillehay, T.D. 1996. South American proboscidean: general introduction and reflections on Pleistocene extinctions. En: J. Shoshani y P. Tassy (eds.), The Proboscidea. Evolution and Palaeoecology of Elephants and their relatives, Oxford Univ. Press, Oxford, New York, Tokyo, pp. 316-320. [ Links ]
13. Cuvier, G. 1806. Sur différentes dents du genre des mastodontes, mais d'espèces moindres que celle de l'Ohio, trouvées en plusieurs lieux des deux continents. Annales du Muséum d'Histoire Naturelle 7: 401-420. [ Links ]
14. Chen, G. 1999. Sinomastodon Tobien et al., 1986 (Proboscidea, Mammalia) from the Pliocene and early-middle Pleistocene of China. Proceeding of the Seven Annual Meeting of the Chinese Society of Vertebrate Paleontology 1999: 179-187. [ Links ]
15. Dudley, J.P. 1996. Mammoths, gomphotheres, and the Great American Faunal Interchange. En: J. Shoshani y P. Tassy (eds.), The Proboscidea. Evolution and Palaeoecology of Elephants and their relatives, Oxford Univ. Press, Oxford, New York, Tokyo, pp. 289-295. [ Links ]
16. Ehleringer, J.R., Cerling, T.E. y Helliker, B.K. 1997. C4 photosynthesis, atmospheric CO2, and climate. Oecologia 112: 285-299. [ Links ]
17. Frassinetti, D. y Alberdi, M.T. 2000. Revisión y estudio de los restos fósiles de Mastodontes de Chile (Gomphotheriidae): Cuvieronius hyodon, Pleistoceno superior. Estudios Geológicos 56: 197-208. [ Links ]
18. Frassinetti, D. y Alberdi, M.T. 2005. Presencia del género Stegomastodon entre los restos fósiles de Mastodontes de Chile (Gomphotheriidae), Pleistoceno Superior. Estudios Geológicos 61: 101-107. [ Links ]
19. Gramajo, A.J. 1992. La fauna del Cuaternario y sus yacimientos en la Llanura Santiagueña. Museo de Ciencias Antropológicas y Naturales "Emilio y Duncan Wagner", Serie Estudio 4: 75-93. [ Links ]
20. Hay, O.P. 1922. Further Observations on some extinct elephants. Proceedings of the Biological Society of Washington 35: 97-102. [ Links ]
21. Hoffstetter, R. 1950. Observaciones sobre los mastodontes de Sud América y especialmente del Ecuador. Haplomastodon, subgn. nov. de Stegomastodon. Publicaciones de la Escuela Politécnica Nacional 1: 1-51. [ Links ]
22. Hoffstetter, R. 1952. Les mammifères Pléistocènes de la République de l'Equateur. Mémoires de la Societa de Géologie de France 66: 1-391 [ Links ]
23. Lambert, W.D. 1996. The biogeography of the gomphotheriid proboscideans of North America. En: J. Shoshani y P. Tassy (eds.), The Proboscidea. Evolution and Palaeoecology of Elephants and their relatives, Oxford Univ. Press, Oxford, New York, Tokyo, pp. 143-148. [ Links ]
24. Lesson, R.P., 1842. Nouveau tableau du règne animal: mammifères. Paris, Arthus-Bertrand, 204 pp. [ Links ]
25. Leidy, J. 1858. On new species of mastodon and elephant from Nebraska, Mastodon mirificus, Elephas imperator. Proceedings of the Academy of Natural Science of Philadelphia 10: 10. [ Links ]
26. MacFadden, B.J. 2000. Middle Pleistocene Climate Change recorded in Fossil Mammal Teeth from Tarija, Bolivia, and Upper limit of the Ensenadan Land-Mammal Age. Quaternary Research 54: 121-131. [ Links ]
27. MacFadden, B.J., Cerling, T.E., Harris, J.M. y Prado, J.L. 1999. Ancient latitudinal gradients of C3/C4 grasses interpreted from stable isotopes of New World Pleistocene horses. Global Ecology and Biogeography 8: 137-149. [ Links ]
28. MacFadden, B.J., Siles, O., Zeitler, P., Johnson, N.M. y Campbell, K.E,Jr. 1983. Magnetic Polarity Stratigraphy of the Middle Pleistocene (Ensenadan) Tarija Formation of Southern Bolivia. Quaternary Research 19: 172-187. [ Links ]
29. Mones, A. y Francis, J.C. 1973. Lista de los Vertebrados fósiles del Uruguay, II. Comunicaciones Paleontológicas del Museo de Historia Natural de Montevideo 1: 39-97. [ Links ]
30. Moreno, F.J.P. 1888. Informe preliminar de los progresos del Museo La Plata durante el primer semestre de 1888. Buenos Aires, 1-35. [ Links ]
31. Osborn H.F. 1936 Proboscidea. A monograph of the discovery, evolution, migration and extinction of the mastodons and elephants. 1: Moeritherioidea, Deinotheiroidea, Mastodontoidea. American Museum of Natural History 1: 1-802. [ Links ]
32. Pohlig, H. 1912. Sur une vieille mandibule de "Tetracaulodon ohioticum" Blum., avec défense in situ. Bulletin de la Société Belge de Géologie 26: 187-193. [ Links ]
33. Prado, J.L., Alberdi, M.T., Azanza, B., Sánchez, B. y Frassinetti, D. 2005. The Pleistocene Gomphotheriidae (Proboscidea) from South America. Quaternary International 126-128: 21-30. [ Links ]
34. Prado, J.L., Alberdi, M.T. y Gómez, G.N. 2002. Late Pleistocene gomphothere (Proboscidea) remains from the Arroyo Tapalqué locality (Buenos Aires, Argentina) and their taxonomic and biogeographic implications. Neues Jarhbuch Geologie und Paläontologie, Abh. 225: 275-296. [ Links ]
35. Prado, J.L., Alberdi, M.T., Sánchez, B. y Azanza, B. 2003. Diversity of the Pleistocene Gomphotheres (Gomphotheriidae, Proboscidea) from South America. Deinsea 9: 347-363. [ Links ]
36. Reyment, R.A. 1991. Multidimensional paleobiology. Pergamon Press, 416 pp. [ Links ]
37. Rusconi, C. 1967. Animales extinguidos de Mendoza y de la Argentina. Publicación Oficial del Gobierno de Mendoza 489 pp. [ Links ]
38. Russo, A., Ferello, R. y Chebli, G. 1979. Llanura Chaco Pampeana. Academia Nacional de Ciencias. 2º Simposio de Geología Regional Argentina 1: 139-183. [ Links ]
39. Sánchez-Chillón, B., Prado, J.L. y Alberdi, M.T. 2003. Paleodiet, ecology, and extinction of Pleistocene gomphotheres (Proboscidea) from Pampean Region (Argentina). Coloquios de Paleontología, Volumen Extraordinario 1: 617-625. [ Links ]
40. Sánchez-Chillón, B., Prado, J.L. y Alberdi, M.T. 2004. Feeding ecology, dispersal, and extinction of South American Pleistocene gomphotheres (Gomphotheriidae, Proboscidea). Paleobiology 30: 146-161. [ Links ]
41. Simpson, G.G. y Paula Couto, C. 1957. The Mastodonts of Brazil. Bulletin of the American Museum of Natural History 112: 125-190. [ Links ]
42. Spillmann, F. 1928. Tetrabelodon nov. spec. ayora Spillman. El Ecuador Comercial, Quito, Ecuador 6: 1-70. [ Links ]
43. Tassy, P. 1996. Who is who among the Proboscidea?.- In: J. Shoshani y P. Tassy (eds.), The Proboscidea. Evolution and Palaeoecology of Elephants and their relatives. Oxford University Press, Oxford, New York, Tokyo, pp. 39-48. [ Links ]
44. Tassy, P. y Shoshani, J. 1996. Historical overview of classification and phylogeny of the Proboscidea. En: J. Shoshani y P. Tassy (eds.), The Proboscidea. Evolution and Palaeoecology of Elephants and their relatives, Appendix B, Oxford Univ. Press, Oxford, New York, Tokyo, pp. 352-353. [ Links ]
45. Tobien, H., Chen, G.F. y Li, Y.Q. 1986. Mastodonts (Proboscidea. Mammalia) from the late Neogene and early Pleistocene of the People's Republic of China. Part.1. Historical account: the genera Gomphotherium, Choerolophodon, Synconolophus, Amebelodon, Platybelodon, Sinomastodon. Mainzer geowissenschaftlich Mitteilungen 15: 119-181. [ Links ]
46. Tobien, H., Chen, G.F. y Li, Y.Q. 1988. Mastodonts (Proboscidea. Mammalia) from the late Neogene and early Pleistocene of the People's Republic of China. Part.2. The genera Tetralophodon, Anancus, Stegotetrabelodon, Zygolophodon, Mammut, Stegolophodon. Some generalities on the Chinese mastodonts. Mainzer geowissenschaftlich Mitteilungen 17: 95-220. [ Links ]
47. Webb, S.D. 1991. Ecogeography and the Great American Interchange. Paleobiology 17: 266-280. [ Links ]
48. Winge, H. 1906. Jordfundne og nulevende Hovdyr (Ungulata) fra Lagoa Santa, Minas Geraes, Brasilien. Copenhagen, E. Museo Lundii 3: 1-9. [ Links ]
Recibido: 12 de mayo de 2006.
Aceptado: 10 de abril de 2008.

