










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.45 n.2 Buenos Aires abr./jun. 2008
Dasypodidae (Xenarthra, Cingulata) de la Formación Cerro Azul (Mioceno tardío) de la provincia de La Pampa, Argentina
Juan J. Urrutia1,2, Claudia I. Montalvo2 y Gustavo J. Scillato-Yané1,3
1CONICET
2Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa, Uruguay 151, 6300 Santa Rosa, Argentina. cmontalvo@exactas.unlpam.edu.ar
3División Paleontología de Vertebrados, Museo de La Plata, Paseo del Bosque s/nº, 1900 La Plata, Argentina. scillato@fcnym.unlp.edu.ar
Resumen. Se revisaron 5.185 restos fósiles de dasipódidos recuperados de 20 afloramientos huayquerienses (Micoceno tardío) de la Formación Cerro Azul en La Pampa (Argentina). Se registró la presencia de 16 taxones, todos pertenecientes a la Subfamilia Euphractinae: Chasicotatus ameghinoi Scillato-Yané, Chasicotatus sp., Doellotatus inornatus (Rovereto), Doellotatus chapadmalensis Bordas, Ringueletia simpsoni (Bordas), Chorobates villosissimus (Rovereto), Chorobates cf. C. villosissimus, Macroeuphractus morenoi (Lydekker), Macroeuphractus retusus Ameghino, Macroeuphractus sp., Vetelia perforata Scillato-Yané, Macrochorobates scalabrinii (Moreno y Mercerat), Macrochorobates chapalmalensis (Ameghino), Proeuphractus limpidus Ameghino, Proeuphractus sp. y Euphractini incertae sedis. Se mencionan por primera vez para la Formación Cerro Azul a M. chapalmalensis, P. limpidus y Euphractini incertae sedis. Se extiende el biocrón de M. chapalmalensis hasta la edad Huayqueriense. Las variaciones en la composición de las faunas de dasipódidos entre los afloramientos estudiados fueron atribuidas principalmente a diferencias temporales y se ajustó la posición relativa de algunas localidades dentro de un esquema bioestratigráfico de la Formación Cerro Azul previamente propuesto. En su conjunto, todos los dasipódidos recuperados de la Formación Cerro Azul presentan similitudes con aquéllos de las faunas mio-pliocénicas tanto del centro como del noroeste de Argentina; sin embargo, las asociaciones que sugieren menor antigüedad, muestran una creciente similitud con las faunas recuperadas de formaciones neógenas aflorantes en la costa atlántica bonaerense.
Abstract. Dasypodidae (Xenarthra, Cingulata) from the Cerro Azul Formation (late Miocene) of La Pampa province, Argentina. A total of 5,185 fossil remains of dasypodids were recovered from 20 outcrops of Huayquerian age (late Miocene) of the Cerro Azul Formation in La Pampa (Argentina). Sixteen taxa were recorded, all belonging in the Subfamily Euphractinae: Chasicotatus ameghinoi Scillato-Yané, Chasicotatus sp., Doellotatus inornatus (Rovereto), Doellotatus chapadmalensis Bordas, Ringueletia simpsoni (Bordas), Chorobates villosissimus (Rovereto), Chorobates cf. C. villosissimus, Macroeuphractus morenoi (Lydekker), M. retusus Ameghino, Macroeuphractus sp., Vetelia perforata Scillato-Yané, Macrochorobates scalabrinii (Moreno y Mercerat), Macrochorobates chapalmalensis (Ameghino), Proeuphractus limpidus Ameghino, Proeuphractus sp. and Euphractini incertae sedis. M. chapalmalensis, P. limpidus and Euphractini incertae sedis are first records from the Cerro Azul Formation. The range of M. chapalmalensis reaches down to the Huayquerian. Compositional variation of the dasypodid fauna among the surveyed outcrops are believed to be due mainly to temporal differences. The relative position of some of the localities was adjusted within a biostratigraphic frame previously proposed for the Cerro Azul Formation. The dasypodid association of the Cerro Azul Formation shows similarity with other Miocene and Pliocene faunas from central and north-western Argentina. However, those localities where associations hinting younger ages show faunal compositions increasingly similar to those contained in Neogene formations exposed along the coast of Buenos Aires province.
Palabras clave. Dasipódidos; Sistemática; Bioestratigrafía; Mioceno tardío; La Pampa; Argentina.
Key words. Dasypodids; Systematics; Biostratigraphy; Late Miocene; La Pampa; Argentina
Introducción
Los dasipódidos constituyen uno de los grupos de mamíferos de más amplia distribución cronológica en Argentina (Paleoceno medio-Reciente). En lo que respecta a su distribución geográfica, son elementos característicos de la región Neotropical (Scillato-Yané, 1982). Ortiz Jaureguizar (1998) propuso que los dasipódidos, junto con los roedores caviomorfos, habrían sido los grupos dominantes de la fauna sudamericana en el lapso Mioceno tardío- Plioceno ("edad de las planicies australes", Pascual y Bondesio, 1982).
En la provincia de La Pampa, los niveles aflorantes de la Formación Cerro Azul (Linares et al., 1980) han aportado importantes asociaciones faunísticas asignadas al Huayqueriense (Mioceno tardío; Abello et al., 2002; Cerdeño y Montalvo, 2001, 2002; Esteban et al., 2001, 2003; Goin et al., 2000; Goin y Montalvo, 1988; Montalvo y Casadío, 1988; Montalvo et al., 1995, 1996, 1998, 2008; Verzi et al., 1991, 1994, 1995, 1999, 2003, 2008). En estas asociaciones, los xenartros constituyen el grupo más abundante en cuanto a frecuencia de hallazgos y los dasipódidos, en general, la familia más abundante (Montalvo y Szelagowski, 1999). La primera cita para La Pampa la hizo Bordas (1933) quien fundó la especie Doellotatus chapadmalensis Bordas. Posteriormente, Zetti (1972) mencionó Macroeuphractus sp. para la zona del Valle Argentino, Naicó y Salinas Grandes de Hidalgo y además, para esta última localidad, Proeuphractus sp. y un Eutatini indeterminado. Scillato-Yané (1980), en su revisión de los dasipódidos fósiles conocidos de la República Argentina, mencionó para esta provincia a Vetelia perforata Scillato-Yané, Doellotatus chapadmalensis, Chorobates villosissimus (Rovereto) y Macroeuphractus morenoi (Lydekker). En trabajos más recientes, Esteban et al. (2001, 2003) revisaron parte de los dasipódidos recuperados de 11 localidades de la Formación Cerro Azul, incorporando a la lista los siguientes taxones: Chasicotatus ameghinoi Scillato-Yané, Doellotatus inornatus (Rovereto), Ringueletia simpsoni (Bordas), Macrochorobates scalabrinii (Moreno y Mercerat), Proeuphractus sp., Zaedyus pichiy (Desmarest) y Gen. nov. "A" Scillato-Yané, 1982. Finalmente, Urrutia y Scillato-Yané (2003) mencionaron la presencia de Macroeuphractus retusus Ameghino en Salinas Grandes de Hidalgo. Estos últimos estudios mostraron que la fauna de dasipódidos presenta diferencias composicionales entre localidades (Esteban et al., 2001, 2003; Urrutia y Scillato-Yané, 2003). Si bien se planteó que las mismas podrían obedecer a diferencias temporales entre los afloramientos, quedaba pendiente una revisión exhaustiva de todos los materiales recuperados hasta el momento en los afloramientos de la Formación Cerro Azul, a fin de descartar que esas diferencias se debieran a sesgos de muestreo. En este trabajo se realiza una revisión de los dasipódidos de la Formación Cerro Azul. Se actualizan las asociaciones de dasipódidos obtenidas de las distintas localidades de dicha formación, se comparan entre sí y con las de otras formaciones del Cenozoico tardío de Argentina y se infieren relaciones estratigráficas y paleozoogeográficas entre las localidades portadoras y otros yacimientos fosilíferos del Neógeno de Argentina.
Marco geológico y bioestratigráfico
Los sedimentos loéssicos aflorantes en el este y centro de la provincia de La Pampa presentan una gran similitud litológica y paleontológica con los de la "Formación" Epecuén (nunca definida formalmente), del oeste de la provincia de Buenos Aires. Los niveles portadores de vertebrados fósiles que afloran en el este de La Pampa, fundamentalmente los de Salinas Grandes de Hidalgo (Departamento Atreucó) se asignaron primeramente a la "Formación" Epecuén (Pascual y Bocchino, 1963; Pascual et al., 1965; Zetti, 1972; Scillato-Yané, 1975; Campbell y Tonni, 1980; Tambussi, 1987). Más tarde Goin et al. (2000), sobre la base de un análisis estratigráfico detallado en esa localidad, propusieron considerarlos como parte de la Formación Cerro Azul. Esta formación fue descripta por Linares et al. (1980) y a ella se asignaron, en trabajos paleontológicos posteriores, los niveles portadores de vertebrados en varias localidades del centro y este de La Pampa (Abello et al., 2002; Cerdeño y Montalvo, 2001, 2002; Esteban et al., 2001, 2003; Goin et al., 2000; Goin y Montalvo, 1988; Montalvo y Casadío, 1988; Montalvo et al., 1995, 1996, 1998, 2008; Verzi et al., 1991, 1994, 1995, 1999, 2003, 2008).
El análisis estratigráfico detallado de las sedimentitas de la Formación Cerro Azul permitió reconocer en la sucesión clástica tres asociaciones de facies sedimentarias: A, B y C (véase Goin et al., 2000), indicando que el ciclo depositacional aflorante comenzó con depósitos lacustres que conforman la asociación A; sobre ésta se ubican los estratos más potentes y de mayor distribución en la región, correspondientes a la asociación B, consistentes en depósitos eólicos con numerosas evidencias pedogenéticas. Finalmente, se depositaron facies propias de cursos fluviales que constituyen la asociación de facies C. Si bien los niveles de esta última asociación de facies fueron ubicados en la parte superior del perfil integrado, la asociación fósil recuperada fue asignada al Chasiquense (véase Verzi et al., 2008 y bibliografía allí citada) y presenta evidencias de reelaboración tafonómica (Montalvo et al., 2005). En las localidades analizadas en este trabajo, Salinas Grandes de Hidalgo, Laguna Chillhué y Laguna Guatraché presentan las asociaciones de facies A y B; en Cerro La Bota está representada la asociación de facies C y en el resto de las localidades aflora únicamente la asociación de facies B.
La Formación Cerro Azul posee una litología homogénea y una amplia distribución areal lo que ha dificultado una correlación estratigráfica precisa entre sus afloramientos. Verzi (1999, 2002) y Verzi et al. (2003, 2004, 2008) plantearon una hipótesis bioestratigráfica para algunos afloramientos de la Formación Cerro Azul en La Pampa y otras unidades en la provincia de Buenos Aires sobre la base del incremento en la hipsodoncia de los molares y otros caracteres dentarios observados en el linaje Chasichimys-Xenodontomys (Rodentia, Ctenomyidae). Los especímenes de este grupo de roedores recuperados de las localidades Cerro La Bota y Cerro Patagua sugirieron que las faunas de estas localidades son más antiguas que la de otras localidades analizadas y asignables al Chasiquense (Mioceno tardío). Para las asociaciones del resto de las localidades, consideradas de edad Huayqueriense, se ha propuesto la siguiente secuencia en orden decreciente de antigüedad: Laguna Chillhué y Naicó, Bajo Giuliani, Barrancas Coloradas, El Guanaco, Sección Seminario (Grünbein), "Formación Irene" (estas dos últimas en la provincia de Buenos Aires) y Caleufú. En particular, las especies nominales de Xenodontomys halladas en Barrancas Coloradas, El Guanaco y Caleufú se interpretaron como cronomorfos sucesivos (Verzi, 1999, 2002; Verzi et al., 2003, 2004, 2008).
Materiales y métodos
Se estudiaron los restos fósiles asignados a la Familia Dasypodidae recuperados de afloramientos de la Formación Cerro Azul procedentes de 20 localidades de la provincia de La Pampa (figura 1). Las mismas están ubicadas en los departamentos que se mencionan entre paréntesis: Barrancas Coloradas, Bajo Giuliani y El Guanaco (Capital); Puesto Colorado, Telén y La Salada (Loventué); Quehué y Laguna Chillhué (Guatraché); Salinas Grandes de Hidalgo, Cerro Patagua, Estancia Quiñi Malal, Cerro El Chancho y Cerro La Bota (Utracán); Laguna Guatraché (Hucal); Ruta 14, Estancia Don Mariano y Naicó (Toay); Estancia Ré (Conhelo); Cerro de los Guanacos (Chical Có) y Caleufú (Rancul).
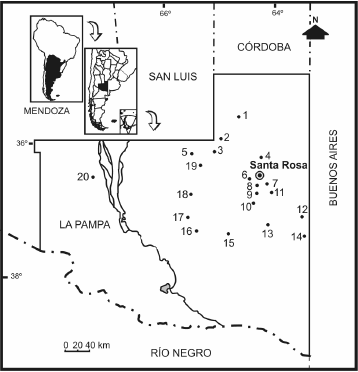
Figura 1. Mapa de ubicación con las localidades fosilíferas de la Formación Cerro Azul / location map showing fossiliferous localities of the Cerro Azul Formation. 1,. Caleufú, 2, Estancia Ré, 3, Puesto Colorado, 4, El Guanaco, 5, Telén, 6, Estancia Don Mariano, 7, Barrancas Coloradas, 8, Bajo Giuliani, 9, Naicó, 10, Quehué, 11, Ruta 14, 12, Salinas Grandes de Hidalgo, 13, Laguna Chillhué, 14, Laguna Guatraché, 15, Estancia Quiñi-Malal, 16, Cerro La Bota, 17, Cerro Patagua, 18, Cerro El Chancho, 19, La Salada, 20, Cerro de los Guanacos.
Se revisaron 5.185 restos de dasipódidos de la Formación Cerro Azul depositados en la colección de la Facultad de Ciencias Exactas y Naturales de la Universidad Nacional de La Pampa, cuyo acrónimo es GHUNLPam. En los casos en que un mismo número de colección incluye varios restos encontrados aislados en el afloramiento, se indican en los listados con números separados por barras. No se tuvieron en cuenta los pocos restos de dasipódidos recuperados de afloramientos pampeanos que se encuentran depositados en las colecciones de la Facultad de Ciencias Naturales y Museo de La Plata (MLP) y en la del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" (MACN) por no contar con datos precisos de su procedencia. Del total de restos revisados, sólo 1.450 pudieron determinarse hasta el nivel de tribu, género o especie; todos ellos pertenecen a la Subfamilia Euphractinae y corresponden a placas aisladas de la coraza y, en menor medida, a placas asociadas en pequeños grupos. En el anexo 1 se listan los ejemplares asignados a cada taxón, organizados de acuerdo a sus localidades de procedencia. No se incluyeron en el anexo 1 los ejemplares mencionados por Esteban et al. (2001, 2003) cuya asignación no es discutida en este trabajo.
Para realizar las determinaciones, los fósiles se compararon con el material tipo y otros restos depositados en las colecciones del MACN y del MLP.
Sistemática paleontológica
Superorden XENARTHRA Cope, 1889
Orden CINGULATA Illiger, 1811
Familia DASYPODIDAE Bonaparte, 1838
Subfamilia EUPHARACTINAE Pocock, 1924
Tribu EUTATINI Bordas, 1933Género Chasicotatus Scillato-Yané, 1979
Especie tipo. Chasicotatus ameghinoi Scillato-Yané, 1979.
Chasicotatus ameghinoi Scillato-Yané, 1979
Figura 2.1
Figura 2. 1, Chasicotatus ameghinoi GHUNLPam 8974/114; 2-3, Doellotatus inornatus GHUNLPam 2316/6, 2316/22; 4-5, Doellotatus chapadmalensis GHUNLPam 8317/5, 8317/21; 6-7, Ringueletia simpsoni GHUNLPam 21561/41, 21562/1; 8-9, Vetelia perforata GHUNLPam 9184/4, 8011/1; 10-11, Chorobates villosissimus GHUNLPam 22634/25, 8679/44; 12-13, Chorobates cf. C. villossisimus GHUNLPam 19239/19, 19239/35; 14-15, Macrochorobates scalabrinii GHUNLPam 2291/55, 8198/96; 16-17, Macrochorobates chapalmalensis GHUNLPam 5066/5, 5976/1; 18-19 Macroeuphractus morenoi GHUNLPam 9257/226, 9523/94; 20, Macroeuphractus retusus GHUNLPam 14000; 21-22, Proeuphractus limpidus GHUNLPam 21648/14, 14447/16 23-24, Proeuphractus sp. GHUNLPam 8807/43, 9058/48; 25-26, Euphractini incertae sedis GHUNLPam 12922/4, 14400/4. Escala: 1 cm. / scale bar: 1 cm.
Observaciones. El material asignado es un grupo de placas móviles y fijas que presentan una figura central más estrecha y alargada que en Doellotatus y Eutatus y cuatro a seis forámenes pilíferos grandes dispuestos en la zona posterosuperior.
Distribución. Chasicotatus ameghinoi se registra en la Formación Arroyo Chasicó (Chasiquense) en Buenos Aires (Scillato-Yané, 1982), en la Formación Chiquimil (Chasiquense) en Catamarca (Esteban y Nasif, 1999) y en la Formación Cerro Azul (Huayqueriense; tabla 1) en La Pampa (Esteban et al., 2001, 2003).
Tabla 1. Distribución geográfica de los taxones de dasipódidos presentes en la Formación Cerro Azul / geographic distribution of dasypodid taxa from the Cerro Azul Formation.
Chasicotatus sp.
Observaciones. Esteban et al. (2003) ubicaron en este taxón nueve fragmentos de placas recuperadas de Caleufú mencionando que las placas de borde presentan la porción distal transformada en una prominencia cónica. Agregaron que Chasicotatus sp. nov. Scillato- Yané, 1982: 79 y Ch. peiranoi Esteban y Nasif, 1996 presentan esta característica. El estado fragmentario en que se encuentran las placas impide determinarlas hasta nivel de especie. Sin embargo, en el holotipo de Ch. ameghinoi (MLP 76-VI-12-11) que conserva muy pocas placas marginales, la de sexta banda pelviana termina en un tubérculo agudo dirigido hacia atrás y afuera, similar al que presentan las demás especies y una de las placas de borde antes mencionada.
Género Doellotatus Bordas, 1932
Especie tipo. Eutatus inornatus Rovereto, 1914.
Sinonimia. Eutatus Rovereto, 1914 (partim: 37, 156 y 223, nec Eutatus Gervais 1867: 280); Eutatopsis Kraglievich 1934: 57.
Doellotatus inornatus (Rovereto) Bordas, 1932
Figuras 2.2-3
Observaciones. Los restos asignados corresponden a un grupo de placas de la coraza dorsal, fijas y móviles, con la figura central elíptica y bastante elevada. Forámenes pilíferos en el borde posterior, proporcionalmente más grandes que en Chasicotatus.
Distribución. Esta especie se registra en las formaciones Monte Hermoso (Montehermosense) e "Irene" (con respecto a la edad de esta "formación" véase Verzi et al., 2003) en Buenos Aires (Scillato-Yané, 1982) y en la Formación Cerro Azul (Huayqueriense; tabla 1) en La Pampa (Esteban et al., 2001, 2003).
Doellotatus chapadmalensis Bordas, 1933
Figuras 2.4-5
Observaciones. El material asignado es un conjunto de placas fijas y móviles que presenta una figura central más corta, ancha y aplanada que D. inornatus.
Distribución. Doellotatus chapadmalensis se registra en las formaciones Monte Hermoso (Montehermosense), Chapadmalal (Chapadmalalense) e "Irene" (con respecto a la edad de esta "formación" véase Verzi et al., 2003) en Buenos Aires (Scillato-Yané, 1982) y en la Formación Cerro Azul (Huayqueriense; tabla 1) en La Pampa (Esteban et al., 2001, 2003).
Género Ringueletia Reig, 1958
Especie tipo. Doellotatus simpsoni Bordas, 1933.
Sinonimia. Doellotatus Bordas, 1933 (partim: 611, nec Bordas, 1932: 167).
Ringueletia simpsoni (Bordas) Reig, 1958
Figuras 2.6-7
Observaciones. El material estudiado es un grupo de 12 placas de superficie muy rugosa, con una figura central lageniforme, aplanada y forámenes pilíferos del margen posterior de las placas dispuestos en dos filas bien diferenciables, siendo los anteriores más pequeños que los posteriores.
Distribución. Ringueletia simpsoni se registra en las formaciones Monte Hermoso (Montehermosense), Chapadmalal (Chapadmalalense), Barranca de los Lobos (Marplatense) e "Irene" (con respecto a la edad de esta "formación" véase Verzi et al., 2003) en la costa de la provincia de Buenos Aires (Scillato-Yané, 1982) y en la Formación Cerro Azul (Huayqueriense; tabla 1) en La Pampa (Esteban et al., 2003).
Tribu EUPHRACTINI Pocock, 1924
Género Vetelia Ameghino, 1891
Especie tipo. Vetelia puncta Ameghino, 1891.
Vetelia perforata Scillato-Yané, 1979
Figuras 2.8-9
Observaciones. Los restos asignados son placas fijas que presentan una figura central subelíptica, bien redondeada en su extremo anterior, rodeada adelante y a los lados por figuritas laterales menores, con numerosas perforaciones pilíferas en el margen posterior.
Distribución. Esta especie se registra en la Formación Collón Curá (Friasense) en la Patagonia; la Formación Arroyo Chasicó (Chasiquense) en Buenos Aires (Scillato-Yané, 1982) y en la Formación Cerro Azul (Huayqueriense; tabla 1) en La Pampa (Esteban et al., 2001).
Género Chorobates Reig, 1958
Especie tipo. Proeuphractus recens Ameghino, 1887.
Sinonimia. Proeuphractus Ameghino, 1887: 19 (nec Ameghino, 1886: 208); Dasypus Lydekker, 1894 (partim: 59, nec Linné, 1758: 50); Proeuphractus Rovereto, 1914 (partim: 107, 155, 224, nec Ameghino, 1886: 208).
Chorobates villosissimus (Rovereto) Reig, 1958
Figuras 2.10-11
Observaciones. Las placas móviles asignadas a esta especie presentan la figura central elevada, abiselada, desviada hacia el lado externo; surcos anchos y de fondo cóncavo, figuras laterales indivisas, no tan elevadas como en Proeuphractus. Las placas fijas presentan una figura central elevada, abiselada, desviada hacia el lado externo; figuritas periféricas muy convexas, globosas. Tanto las placas fijas como las móviles poseen perforaciones pilíferas en una sola fila y sobre los márgenes posterior y lateral externo.
Distribución. Scillato-Yané (1982) mencionó Chorobates villosissimus para la Formación Tunuyán en Mendoza; el "Araucanense" de las provincias de Catamarca y Tucumán y la "Formación" Epecuén en Buenos Aires. Tonni et al. (1992) lo incluyeron en la fauna del "Hermosense típico" y Esteban et al. (2001) agregaron a su distribución la Formación Cerro Azul (Huayqueriense; tabla 1) en La Pampa.
Chorobates cf. C. villosissimus (Rovereto) Reig, 1958
Figuras 2.12-13
Observaciones. Castellanos (1947) mencionó un resto (Nº 293 de la colección del Instituto de Fisiografía y Geología de Rosario) proveniente del "Araucanense" de la provincia de Catamarca asignado a Chorobates villosissimus, que es aproximadamente una vez y media mayor que el holotipo (MACN 2940) establecido por Rovereto (1914). La serie de restos provenientes de los afloramientos de Caleufú y El Guanaco se asemejan, en tamaño, al ejemplar de Chorobates villosissimus descripto por Castellanos (1947). El registro de C. villosissimus es relativamente abundante en el Neógeno de Argentina y el tamaño de la mayor parte de los restos es similar al del holotipo. Este grupo de placas se asigna a Chorobates cf. C. villosissimus hasta tanto se realice una revisión exhaustiva de los materiales.
Género Macrochorobates Scillato-Yané, 1980
Especie tipo. Proeuphractus scalabrinii Moreno y Mercerat, 1891.
Sinonimia. Proeuphractus Moreno y Mercerat, 1891: 226 (nec Ameghino, 1886: 208); Dasypus Lydekker, 1894 (partim: 60, nec Linné 1758: 50); Proeuphractus Ameghino 1908: 427 (nec Ameghino, 1886: 208); Acantharoidea Rovereto, 1914: 108; Rivadaviafractus Bordas, in schedis (partim vide Reig 1958: 250).
Macrochorobates scalabrinii (Moreno y Mercerat) Scillato-Yané, 1980
Figuras 2.14-15
Observaciones. El material asignado es un grupo de placas fijas y móviles con la figura central elevada, desviada hacia el lado externo. En las placas fijas, las figuras laterales son bastante convexas.
Distribución. Scillato-Yané (1982) mencionó Macrochorobates scalabrinii para la "Formación" Epecuén (Huayqueriense) en la provincia de Buenos Aires y el "Araucanense" de las provincias de Catamarca y Tucumán; Esteban et al. (2001) extendieron su distribución a la Formación Cerro Azul (Huayqueriense; tabla 1) en La Pampa.
Macrochorobates chapalmalensis (Ameghino) Scillato-Yané, 1980
Figura 2.16-17
Observaciones. Los restos estudiados corresponden a un conjunto de placas de tamaño algo menor y con las figuras menos convexas que las de M. scalabrinii.
Distribución. Macrochorobates chapalmalensis fue mencionada por Scillato-Yané (1982) para las formaciones Monte Hermoso (Montehermosense), "Irene" (con respecto a la edad de esta "formación" véase Verzi et al., 2003) y Chapadmalal (Chapadmalalense) en la provincia de Buenos Aires y la Formación Uquía ("Uquiense", Plioceno tardío-Pleistoceno) en Jujuy. En este trabajo se cita por primera vez para la Formación Cerro Azul (Huayqueriense; tabla 1) en La Pampa.
Género Macroeuphractus Ameghino, 1887
Especie tipo. Macroeuphractus retusus Ameghino, 1887.
Sinonimia. Dasypotherium Moreno, 1889: 38; Dasypus Lydekker, 1894 (partim: nec Linné 1758: 50); Proeuphractus Ameghino 1920: 677 (nec Ameghino, 1886: 208).
Macroeuphractus morenoi (Lydekker) Rovereto, 1914
Figuras 2.18-19
Observaciones. Las placas móviles asignadas a esta especie presentan la figura central estrecha, muy elevada y abiselada; las figuras laterales divididas en figuritas menores por surcos transversales. Las placas fijas presentan una figura central abiselada y figuritas periféricas convexas.
Distribución. Macroeuphractus morenoi fue citada en las provincias de Catamarca y Tucumán para el"Araucanense" y para la "Formación" Epecuén (Huayqueriense) en Buenos Aires (Scillato-Yané, 1982); Esteban et al. (2001, 2003) agregaron a la distribución conocida registros en la Formación Cerro Azul (Huayqueriense; tabla 1) en La Pampa.
Macroeuphractus retusus Ameghino, 1887
Figura 2.20
Observaciones. El material estudiado es un grupo de placas asociadas. Las placas fijas presentan las figuras más aplanadas y los surcos más superficiales que en M. morenoi. Se conservan dos fragmentos posteriores de placas móviles con la figura central plana ensanchada hacia atrás y las figuras laterales indivisas.
Distribución. Macroeuphractus retusus se registra en las formaciones Ituzaingó (Huayqueriense) y Monte Hermoso (Montehermosense) en las provincias de Entre Ríos y Buenos Aires respectivamente (Scillato- Yané, 1982) y en la Formación Cerro Azul (Huayqueriense; tabla 1) en La Pampa (Urrutia y Scillato-Yané, 2003).
Macroeuphractus sp.
Observaciones. Esteban et al. (2003) asignaron a este taxón cuatro placas del escudo cefálico procedentes de Caleufú.
Género Proeuphractus Ameghino, 1886
Especie tipo. Proeuphractus limpidus Ameghino, 1886.
Proeuphractus limpidus Ameghino, 1886
Figura 2.21-22
Observaciones. El material asignado consiste en placas móviles y fijas con orificios pilíferos ubicados en una sola fila sobre el margen posterior y ambos márgenes laterales. Las placas móviles presentan una figura central alargada y completamente recta; dos figuras laterales bastante elevadas separadas de la central por surcos anchos y de fondo cóncavo. En las placas fijas la figura central es muy elevada y fuertemente desviada hacia el lado externo y las figuritas periféricas muy convexas, casi globosas.
Distribución. Proeuphractus limpidus se registra en la Formación Ituzaingó en la provincia de Entre Ríos (Scillato-Yané, 1982), en la Formación Salicas en La Rioja (Tauber, 2005) y en niveles asignados al Huayqueriense de Uruguay (Perea y Scillato-Yané, 1995). En este trabajo, se menciona por primera vez para la Formación Cerro Azul (Huayqueriense; tabla 1) en La Pampa.
Proeuphractus sp.
Figuras 2.23-24
Observaciones. El género Proeuphractus tiene una sola especie válida, P. limpidus. Los fósiles estudiados son de mayor tamaño que los del ejemplar tipo, más del doble de ancho y de largo. Si bien en los restos están presentes las características diagnósticas del género, la ornamentación presenta algunas diferencias: en las figuras laterales de las placas móviles, se esboza una división en figuritas menores (figuras laterales indivisas en P. limpidus). Esteban et al. (2001, 2003) y Tauber (2005) notaron estas diferencias y propusieron esperar que se conozcan ejemplares más completos para fundar una nueva especie.
Euphractini incertae sedis
Figuras 2.25-26
Sinonimia. Zaedyus Esteban et al., 2001 (partim: 82; nec Ameghino, 1889: 867).
Observaciones. Los restos corresponden a placas móviles, semimóviles y fijas recuperadas aisladas en La Salada, Estancia Ré, Quehué y Telén. Esteban et al. (2001) incluyeron los materiales que consideramos aquí como Euphractini incertae sedis dentro de Zaedyus, sin embargo son de menor tamaño, las figuras de las placas fijas son menos convexas y todos los forámemes pilíferos del margen posterior son de tamaño similar. Las características preservadas en los restos permiten determinarlos como pertenecientes a la tribu Euphractini; no obstante, el material con que se cuenta es insuficiente como para realizar una asignación más precisa.
Discusión
Esteban et al. (2001, 2003) y Urrutia y Scillato-Yané (2003) plantearon que la fauna de dasipódidos de los diferentes afloramientos fosilíferos de la Formación Cerro Azul presenta diferencias composicionales. La tabla 1 muestra la distribución de los taxones en las localidades analizadas, todos representantes de la Subfamilia Euphractinae. Las primeras 8 localidades de esa tabla (Caleufú, El Guanaco, Barrancas Coloradas, Bajo Giuliani, Naicó, Laguna Chillhué, Cerro La Bota y Cerro Patagua) corresponden a aquellas en las que la ubicación temporal relativa de sus asociaciones faunísticas fue propuesta por Verzi (1999, 2002) y Verzi et al. (2003, 2004, 2008), desde las más antiguas (Cerros Patagua y La Bota) hasta la más moderna (Caleufú). Las asociaciones faunísticas de las demás localidades no han sido ubicadas aún en la secuencia temporal.
En los diferentes afloramientos, los restos hallados consisten principalmente en placas aisladas. La mayor frecuencia de hallazgos corresponde a Chasicotatus ameghinoi (552 restos) que es además, la especie más ampliamente distribuida, ya que está presente en 19 de las 20 localidades analizadas (tabla 1). También son abundantes Doellotatus inornatus y Chorobates villosissimus (277 y 278 placas asignadas, respectivamente). Scillato-Yané (1979) manifestó que Chasicotatus ameghinoi y Vetelia perforata habrían sido los dasipódidos más abundantes en el sudoeste de la provincia de Buenos Aires durante la edad Chasiquense. Ch. ameghinoi continúa siendo abundante en niveles de la Formación Cerro Azul, incluso hasta en aquellos considerados como los más modernos, asignados al Huayqueriense más tardío, mientras que el registro de V. perforata es escaso y con distribución reducida, ya que sólo está presente en Cerro La Bota, Cerro de los Guanacos, Telén y Cerro Patagua.
Esteban et al. (2001) asignaron varios ejemplares a Zaedyus pichiy; GHUNLPam 5550/1 corresponde a una placa fija rota que presenta sobre el margen posterior cuatro o posiblemente cinco forámenes pilíferos relativamente grandes. Su estado de conservación no permite determinarla hasta el nivel de género pero es probable que corresponda a una especie de la tribu Eutatini. Los ejemplares GHUNLPam 9688/49, 9688/52, 8320/9 y 8419/3 presentan características que admiten su inclusión en la tribu Euphractini, aunque éstas no son suficientes como para asignarlos a ningún género conocido. En cuanto a la placa GHUNLPam 12528, ilustrada por Esteban et al. (2001:83 figura 4 a), si bien su asignación es correcta, no proviene de la Formación Cerro Azul. Los ejemplares GHUNLPam 5433/13 y 8606/45 provenientes de Laguna Chillhué corresponden efectivamente a Zaedyus pichiy, pero presentan una fosilización algo diferente al resto de los materiales y dado que en esa localidad la Formación Cerro Azul está cubierta por sedimentos holocénicos, es probable que estos especímenes provengan en realidad de la unidad más moderna que se encuentra por encima y hayan sido mal catalogados en la colección; finalmente la placa GHUNLPam 5433/9 presenta características que indican que no es fósil. Una situación similar a esta ocurre con los ejemplares GHUNLPam 9892, 9893, 9894 y 2316/17 asignados por Esteban et al. (2001) a Gen nov. "A" Scillato-Yané, 1982 que no están fosilizados y posiblemente pertenezcan a Chaetophractus. De este análisis surge que no es posible confirmar la presencia de Zaedyus pichiy y Gen. nov. "A" en la Formación Cerro Azul.
Proeuphractus es un género monotípico y la única especie válida, P. limpidus, proviene de la Formación Ituzaingó ("Mesopotamiense") de edad Huayqueriense (Cione et al., 2000) y de sedimentos asignados a la misma edad de Uruguay (Perea y Scillato-Yané, 1995) y La Rioja (Tauber, 2005). Proeuphractus sp. fue mencionado previamente para las formaciones Arroyo Chasicó (Scillato-Yané, 1979; Cerdeño, 2000), Cerro Azul (Esteban et al., 2001, 2003), Salicas (Tauber, 2005) y también en el "Hermosense" típico (Tonni et al., 1992). Al menos para las formaciones Cerro Azul y Salicas, parte de los restos asignados a este género podrían corresponder a una nueva especie (Esteban et al., 2001, 2003 y Tauber, 2005). Su distribución en las distintas localidades de la Formación Cerro Azul es amplia por lo que aporta poca información bioestratigráfica.
Los afloramientos de Laguna Chillhué y Salinas Grandes de Hidalgo son los más potentes entre los analizados en este trabajo (Montalvo et al., 1995; Goin et al., 2000). En ambas localidades, muchos de los fósiles no se han recuperado in situ, sino aislados en la base de los afloramientos, por lo que es difícil realizar en esos casos una asignación estratigráfica correcta. De las especies de dasipódidos reconocidas en Laguna Chillhué, la mayoría tiene un amplio rango bioestratigráfico y está presente en gran parte de los afloramientos de la Formación Cerro Azul. De acuerdo al análisis realizado por Verzi (1999) y Verzi et al. (2004, 2008) la asociación faunística de Laguna Chillhué estaría ubicada cronológicamente entre aquéllas asignadas al Chasiquense (Cerro La Bota y Cerro Patagua) y las más modernas. Excepto por la presencia de Vetelia perforata, tiene la misma composición de dasipódidos que Cerro La Bota, lo que mostraría que este registro para Laguna Chillhué no contradice aquella hipótesis. Salinas Grandes de Hidalgo comparte con Laguna Chillhué varias especies de dasipódidos: Chasicotatus ameghinoi, Doellotatus inornatus, Chorobates villosissimus, Macrochorobates scalabrinii, Macroeuphractus morenoi y Proeuphractus sp.; además, Salinas Grandes de Hidalgo es la única localidad de la Formación Cerro Azul con Macroeuphractus retusus, especie abundante en el Montehermosense, si bien también está presente en el "Mesopotamiense" (Cione et al., 2000). Por otro lado, se registra Doellotatus chapadmalensis, presente también en Caleufú y, en la provincia de Buenos Aires, en las formaciones "Irene" y Chapadmalal. Otros taxones también avalan que la asociación de Salinas Grandes de Hidalgo sería más moderna que la de Laguna Chillhué. El equímido Reigechimys (Rodentia), que presenta un patrón de cambio anagenético (Verzi, 1999, 2002; Montalvo y Verzi, 2004), está representado en Laguna Chillhué por Reigechimys plesiodon y en Salinas Grandes de Hidalgo por la especie con caracteres más derivados Reigechimys octodontiformis (Verzi et al., 1994).
El Guanaco y Barrancas Coloradas son las únicas localidades donde aparece Macrochorobates chapalmalensis. Esta especie, que no se conocía para la Formación Cerro Azul, había sido registrada en las formaciones "Irene", Monte Hermoso y Chapadmalal en la provincia de Buenos Aires y en la Formación Uquía en Jujuy (Scillato-Yané, 1982). La presencia de dos cronomorfos diferentes del roedor Xenodontomys simpsoni Kraglievich en El Guanaco y Barrancas Coloradas indicaría la menor antigüedad de la asociación de El Guanaco. A su vez, las asociaciones de ambas localidades serían más antiguas que la de Caleufú, donde está presente X. elongatus (Verzi et al., 2003, 2004, 2008). Por otro lado, Verzi et al. (2003) mencionaron que la "Formación Irene" presenta un cronomorfo de X. ellipticus, derivado con respecto al de El Guanaco, pero primitivo con respecto a X. elongatus, de Caleufú. Sobre esta base, indicaron que al menos los niveles de "Formación Irene" portadores de Xenodontomys, serían asignables al Mioceno tardío (Verzi et al., 2003). La presencia de Macrochorobates chapalmalensis en las localidades de El Guanaco y Barrancas Coloradas es consistente con la propuesta de Verzi (1999) y Verzi et al. (2003, 2004, 2008).
Chorobates cf. C. villosissimus está presente solamente en El Guanaco y Caleufú. Con respecto a la asociación faunística de Caleufú, Esteban et al. (2003) mencionaron que, considerando los dasipódidos presentes, podría asignarse tanto al Plioceno temprano como al Mioceno tardío. Verzi et al. (2003) sugirieron que esta asociación correspondería a la parte final del Mioceno tardío, cercana, pero algo más antigua que la "edad mamífero" Montehermosense. Entre los dasipódidos se destaca la presencia de Ringueletia simpsoni, que no se ha registrado en otras localidades de la Formación Cerro Azul y de Chasicotatus ameghinoi y Macroeuphractus morenoi que no se conocen en edades más modernas que la Huayqueriense; aunque Mones y Mehl (1990) mencionan una especie afín a Macroeuphractus morenoi para sedimentos post-huayquerienses de Bolivia.
Como se mencionó, Verzi (1999) y Verzi et al. (2003, 2004, 2008) asignaron al Chasiquense las asociaciones registradas en Cerro La Bota y Cerro Patagua. En ambas está presente Vetelia perforata, que fue descripta originalmente para la Formación Arroyo Chasicó y ha sido considerada como un elemento relictual pan-santacrucense (Scillato-Yané, 1979). Sin embargo, esta especie también se registró en Telén y Cerro de los Guanacos en asociaciones asignadas a la edad Huayqueriense (Montalvo et al., 1998, 2008).
En las localidades restantes, sólo se recuperaron dasipódidos que tienen rangos bioestratigráficos y de distribución amplios.
Tonni et al. (1998) y Cione y Tonni (2005) propusieron un esquema bioestratigráfico para el Cenozoico superior en la provincia de Buenos Aires. Establecieron una biozona de Chasicotatus ameghinoi en el Miembro Las Barrancas de la Formación Arroyo Chasicó (Chasiquense tardío) y una de Macrochorobates scalabrinii en los sedimentos suprayacentes a la Formación Arroyo Chasicó, asignándolos al Huayqueriense temprano. En la Formación Cerro Azul tanto M. scalabrinii como Ch. ameghinoi se encuentran ampliamente distribuidos y asociados a taxones típicamente huayquerienses y otros más modernos, como en Caleufú. Esto pone en dudas su utilidad como indicadores bioestratigráficos, tal como fue propuesto por Verzi et al. (2004, 2008) y Tauber (2005).
El análisis del conjunto de los dasipódidos de la Formación Cerro Azul avala la propuesta de una conexión zoogeográfica importante de las faunas del Mio-Plioceno del centro de Argentina con las coetáneas de los valles intermontanos de las provincias de Catamarca, Tucumán y San Juan (Scillato-Yané et al., 1995). En los yacimientos del Neógeno de los valles del noroeste de la Argentina son abundantes los registros de la tribu Euphractini y escasos los de Eutatini (Scillato-Yané, 1979).
La presencia de grandes Euphractini (Vetelia, Macroeuphractus y Macrochorobates) en la Formación Cerro Azul, sugiere condiciones climáticas relativamente cálidas, deducida de las exigencias ecológicas conocidas de los representantes vivientes de mayor talla de la tribu citada (Scillato-Yané, 1979). Las evidencias sedimentológicas son concordantes con estos datos, en particular los paleosuelos de la Formación Cerro Azul preservados en la localidad de Telén presentan caracteres propios de ambientes de planicie, con clima semiárido y marcada estacionalidad en las precipitaciones (Melchor et al., 2000; Montalvo et al., 2008). Chorobates villosissimus es una especie muy común en las provincias de Catamarca y Mendoza. Vetelia está representado en sedimentitas asignadas al Mioceno superior en las provincias de Catamarca y San Juan (Esteban y Nasif, 1996; Georgieff et al., 2004). V. perforata se registra en la Formación Collón Curá (Friasense, Mioceno medio; Scillato Yané, 1982) y en la región pampeana, tanto en asociaciones asignadas al Chasiquense (i.e. Arroyo Chasicó, Cerro La Bota y Cerro Patagua, véase Verzi et al., 2008) como en otras asociaciones recuperadas de sedimentos huayquerienses de la Formación Cerro Azul (i.e. Telén y Cerro de los Guanacos).
La tribu Eutatini está representada en la Formación Cerro Azul por tres géneros: Doellotatus, Ringueletia y Chasicotatus. En las localidades portadoras de faunas más modernas es notable un aumento en la diversidad de especies de esta tribu, particularmente en Salinas Grandes de Hidalgo y Caleufú. En ambas asociaciones está presente Doellotatus chapadmalensis y en la última, Ringueletia simpsoni. En los niveles que contienen fauna de edades Chasiquense hasta Chapadmalalense en la provincia de Buenos Aires esta tribu está representada por esos tres géneros y sólo el último se registró en el noroeste argentino (Scillato- Yane, 1982; Esteban y Nasif, 1996). Esto indica una creciente similitud de las faunas más modernas de la Formación Cerro Azul con las asociaciones recuperadas de niveles neógenos de la provincia de Buenos Aires.
Conclusiones
La Formación Cerro Azul, presenta mayor número de especies de dasipódidos (16) que cualquier otra unidad estratigráfica asignada al Mioceno tardío.
Se agrega a la fauna conocida para la Formación Cerro Azul Macrochorobates chapalmalensis, Proeuphractus limpidus y Euphractini incertae sedis.
No se confirma la presencia de Zaedyus pichiy ni de Gen. nov. "A" Scillato-Yané, 1982, propuestas por Esteban et al. (2001).
Las amplias distribuciones biocronológicas de Chasicotatus ameghinoi y Macrochorobates scalabrinii en la Formación Cerro Azul no avalan las biozonas propuestas por Tonni et al. (1998) y Cione y Tonni (2005) para caracterizar al Chasiquense superior y el Huayqueriense basal.
El análisis de las diferencias en la fauna de dasipódidos de las localidades prospectadas de la Formación Cerro Azul, avala el esquema bioestratigráfico propuesto por Verzi (1999) y Verzi et al. (2003, 2004, 2008). En particular se comprueba la modernidad creciente de los yacimientos de Barrancas Coloradas, El Guanaco y Caleufú. Las dos primeras localidades son las únicas de la Formación Cerro Azul que presentan Macrochorobates chapalmalensis. El Guanaco comparte con Caleufú la presencia de Chorobates cf. C. villosissimus y, en la última, la exclusividad del registro de Ringueletia simpsoni dentro de la Formación Cerro Azul indica una mayor modernidad de la fauna.
El conjunto de dasipódidos de la Formación Cerro Azul sugiere la existencia de una conexión zoogeográfica importante con la región de los valles del noroeste de Argentina que durante el Mioceno tardío- Plioceno temprano presentaron condiciones de tipo chaqueño (Scillato-Yané, 1975). Pero, las asociaciones de dasipódidos de los afloramientos que presentan indicios de mayor modernidad sugieren una relación más estrecha con las faunas recuperadas de las formaciones aflorantes en la costa atlántica bonaerense.
Chasicotatus ameghinoi. Véase Esteban et al. (2001; 2003) y GHUNLPam 8041/2, 8041/3, 8041/11, 8047/1, 8047/2, 8047/3 y 8047/4 (Cerro Patagua); 2242/7, 2243/15, 2252/13, 2265/7, 2335/4, 2371/1, 2371/2, 2371/22, 2371/25, 2371/26, 2371/27, 2371/28, 2371/37, 2371/51, 2371/54, 2371/56, 2371/63, 2371/72, 2373/2, 2918/1, 5391, 5472/8, 5505/9, 5516/16, 5679/5, 5679/6, 5679/7, 5679/14, 5679/15, 5679/16, 5679/26, 5679/27, 5711/6, 5711/7, 8066/22, 8066/30, 9570/8, 9570/22, 12771/2, 14068/1, 14068/2, 21634/6, 21634/19, 21634/22, 21766/1 y 21766/3 (Bajo Giuliani); 2038/22 (Barrancas Coloradas); 19239/103, 21561/21 (Caleufú); 9716/1, 9716/4, 9716/5, 9716/9, 9716/12, 9716/13, 9716/23, 9716/27, 14246/2, 14246/7, 14246/13, 14246/16, 14246/19, 14246/22, 14246/40, 14280/5, 14280/9, 14280/11, 22634/1, 22634/7 y 22634/26 (Estancia Ré); 5372, 6919, 6971, 6988/4, 8011/18, 8011/21, 8011/25, 21746/4, 22001/11 y 22001/13 (Cerro La Bota); 2099/34, 2099/39, 2128, 2163/28, 2291/54, 2291/61, 2291/63, 2316/3, 2316/5, 2316/15, 2316/16, 2316/17, 2316/18, 2316/19, 2316/21, 2386/13, 2386/17, 5568/2, 5568/7, 5575/1, 5575/2, 5575/6, 5575/8, 5575/9, 5577/4, 5577/5, 5609/2, 5609/4, 5627/2, 5627/12, 5627/15, 8404/3, 8606/18, 8606/22, 8606/23, 8606/31, 8986/15, 14447/12, 14447/19, 14447/21, 14447/22, 14447/23, 14457/4, 14651/1, 14651/2, 19038/7, 19038/8, 19038/10, 21728/1, 21728/2 y 21740/1 (Laguna Chillhué); 2283/3 y 5465/2 (El Guanaco); 5512/8, 5651/3, 5651/4 y 5651/8 (Estancia Don Mariano); 2082, 2083, 2096/2, 2096/6 y 2096/9 (Laguna Guatraché); 9257/14, 9257/141, 9257/142, 9257/146, 9257/164, 9257/259, 9257/260, 9257/261, 9257/262, 9257/263, 9257/264, 9257/265, 9257/266, 9257/267, 9257/268, 9257/269, 9257/270, 9257/271, 9257/272, 9257/273, 9257/274, 9257/275, 9257/276, 9257/277, 14816/3, 14817/1, 14817/2, 14817/3, 14817/5, 14818/2, 14818/3, 14820/6, 21651/1, 21651/4, 21651/8 y 21652/14 (La Salada); 9184/13, 9184/17, 9184/22, 9184/25, 9184/26, 9184/30, 19063/1, 21615, 21617/1, 21617/2, 21617/3, 21617/4 y 21617/11 (Cerro de los Guanacos); 2111/1, 2111/5, 2111/6, 2111/8, 2111/9, 2111/11, 2111/15 y 2111/16 (Naicó); 14177/1, 14177/2, 14177/3, 14177/4, 14177/5, 14177/7, 14177/8, 14177/10, 14177/14, 14177/17, 14177/18, 14177/19, 14177/20, 14177/25, 14177/26, 14177/29, 14177/30, 14177/44, 14177/45, 14177/48, 14177/53, 14177/54, 14177/73, 14177/89, 14177/94, 14177/99, 14177/106, 14177/113, 14177/141, 14177/146, 14286/10 y 14286/13 (Puesto Colorado); 80, 5816/1, 5816/3, 5816/4, 5816/5, 5816/6, 5816/9, 5896/4, 5896/11, 5896/20, 5896/27, 5896/30, 5896/35, 5896/42, 6734/7, 6824/7, 6824/11, 8198/77, 8679/33, 8679/36, 8679/42, 8679/43, 8679/45, 8679/46, 8679/47, 8679/48, 9146/3, 9146/4, 9146/9, 12987/1, 12987/2, 12987/3, 12987/4, 12987/5, 12987/6, 14400/8 y 14400/10 (Quehué); 8078/10 y 8078/15 (Estancia Quiñi Malal); 5914/4, 8275/9, 8275/10, 8275/11, 8275/12, 8807/8, 8807/9, 8807/11, 8807/15, 8807/23, 8807/31, 8807/53, 8807/56, 8807/68, 8974/97, 8974/98, 8974/99, 8974/104, 8974/105, 8974/106, 8974/107, 8974/108, 8974/109, 8974/114, 8974/120, 8974/121, 8974/122, 8974/123, 8974/125, 8974/126, 8974/127, 8974/129, 8974/134, 8974/135, 8974/136, 8974/138, 89747/139, 8974/140, 8974/142, 8974/143, 8974/145, 8974/147, 8974/149, 8974/150, 8974/152, 8974/158, 8974/159, 8974/160, 8974/161, 8974/162, 8974/165, 8974/166, 9164/3, 14313, 14318/3, 14318/4, 14318/7, 14318/14, 14345/4, 14490/1, 14490/2, 14490/3, 14490/4, 14490/5, 14490/6, 14496/6 y 14592 (Salinas Grandes de Hidalgo); 8253/9, 8253/17, 8253/23, 8253/54, 8253/83, 8253/85, 8253/100, 8253/106, 8253/107, 8253/126, 8253/128, 8392/1, 8392/3, 8392/4, 8392/5, 8392/6, 8392/7, 8392/9 ,8392/10, 8392/11, 8392/12, 8392/13, 8392/15, 8392/17, 8392/18, 8392/19, 8392/20, 8392/24, 8453/7, 8453/11, 8453/28, 8453/29, 8453/37, 8453/42, 8453/43, 8453/46, 8453/50, 8453/59, 8453/64, 8453/65, 8453/69, 8453/79, 8453/81, 8453/83, 8453/92, 8453/94, 8453/99, 8453/100, 8453/106, 8453/116, 8453/117, 8453/118, 8453/120, 8453/126, 8453/127, 8453/140, 8453/143, 8453/145, 8453/148, 9061/1, 9062/22, 9064/2, 9064/9, 9064/12, 9064/13, 9064/18, 9066/11, 9066/17, 9067/40, 9067/60, 9067/62, 9067/71, 9067/74, 9067/76, 9067/137, 9067/140, 9093/1, 9093/3, 9093/4, 9093/10, 9093/12, 9093/14, 9093/15, 9093/16, 9446, 9475/4, 9478/11, 9478/12, 9478/14, 9478/15, 9478/17, 9478/18, 9478/20, 9478/23, 12922/8, 12922/9, 12922/13, 12922/18, 14107, 14623/10, 14623/11, 14623/14, 14623/30, 14623/35, 14862/3, 14862/4, 14862/5, 14862/7 y 14867/3 (Telén).
Doellotatus inornatus. Véase Esteban et al. (2001; 2003) y GHUNLPam 435/1, 633, 636, 2243/11, 2243/21, 2252/11, 2252/12, 2335/2, 2371/3, 2371/5, 2371/6, 2371/7, 2371/9, 2371/10, 2371/14, 2371/17, 2371/18, 2371/20, 2371/23, 2371/24, 2371/31, 2371/32, 2371/33, 2371/34, 2371/36, 2371/40, 2371/42, 2371/49, 2371/50, 2371/53, 2371/59, 2371/64, 2371/65, 2371/70, 2371/75, 2373/3, 2918/2, 5472/7, 5472/10, 5479/1, 5479/8, 5484/1, 5505/6, 5516/7, 5516/11, 5544/4, 5544/5, 5679/2, 5679/3, 5679/4, 5679/8, 5679/9, 5679/10, 5679/12, 5679/17, 5679/19, 5711/4, 5711/5, 5711/8, 5722/9, 5722/12, 5722/13, 5953/4, 6724/5, 8066/13, 8066/18, 8066/25, 8066/26, 8066/27, 9570/19, 12954/1, 12954/2, 12954/9, 14066/26, 14066/28, 14066/29, 14066/30, 14066/31, 14066/32, 14066/34, 14066/35, 14066/36, 14066/37, 14066/38, 14066/39, 21177/1, 21634/34, 21766/2, 22659/2, 22659/6 y 22659/8 (Bajo Giuliani); 2038/26, 2038/28, 2159, 2161, 2408/6 y 2408/7 (Barrancas Coloradas); 14246/1, 14246/4, 14246/9, 14280/3, 21779/1, 22634/9, 22634/36 y 22634/39 (Estancia Ré); 5378, 6988/9 y 22001/15 (Cerro La Bota); 2099/40, 2127, 2291/62, 2316/6, 2316/10, 2316/11, 2316/12, 2316/13, 2316/14, 2316/22, 2386/11, 2386/18, 2386/19, 2386/22, 2389/3, 2435/11, 5575/10, 5575/11, 5575/12, 5576/9, 5609/1, 5609/10, 5609/11, 5627/9, 5627/11, 5627/13, 5627/16, 8028/1, 8034/19, 8404/2, 8606/8, 8606/10, 8606/14, 8606/27, 8986/4, 8986/7, 8986/11, 8986/12, 14447/1, 14447/6, 14447/17, 14447/18, 14447/20, 14447/24, 14656/1, 19037/2, 19037/3, 19038/1, 19038/9, 21728/8 y 21740/5 (Laguna Chillhué); 2283/2, 5066/3, 12806/2, 12999/1, 12999/2 y 12999/3 (El Guanaco); 5512/3 y 5541 (Estancia Don Mariano); 9099/3 (Laguna Guatraché); 9257/165, 9257/299, 9257/300, 9257/301, 9257/302, 9257/303, 14815/3, 14815/5, 14816/2, 14817/6, 14818/5, 14818/6, 14819/6, 14819/8, 14820/3, 14820/4, 21652/1 y 21652/4 (La Salada); 14177/21, 14177/32, 14177/36, 14177/43, 14177/51, 14177/117, 14177/169, 14177/289, 14286/1 y 14286/3 (Puesto Colorado); 5816/2, 8679/25, 9688/57, 14400/7, 14400/9 y 14738/4 (Quehué); 12754/9; 12754/10 y 21167/2 (Ruta 14); 14496/4 (Salinas Grandes de Hidalgo); 8253/1, 8253/3, 8253/14, 8253/25, 8253/69, 8253/93, 8253/109, 8453/17, 8453/32, 8453/48, 8453/66, 8453/82, 8453/84, 8453/87, 8453/111, 8453/114, 9060/2, 9064/11, 9067/13, 9067/70, 9067/99, 9478/7, 9478/10, 9478/13, 9478/21, 9478/38, 9478/54, 9478/70, 14192/3, 14623/3, 14623/4, 14623/5, 14623/6, 14623/7 y 14623/8 (Telén).
Doellotatus chapadmalensis. Véase Esteban et al. (2001; 2003) y GHUNLPam 5656/1, 8275/8, 8275/15, 8275/17, 8807/5, 8807/6, 8807/14, 8807/16, 8807/17, 8807/24, 8807/26, 8807/27, 8807/36, 8807/51, 8807/61, 8807/85, 14008/14, 14008/15, 14318/1, 14318/2, 14478/2 y 14496/1 (Salinas Grandes de Hidalgo).
Vetelia perforata. Véase Esteban et al. (2001) y GHUNLPam 8041/1 (Cerro Patagua); 6987/2 y 8011/1 (Cerro La Bota); 9107 (Telén).
Chorobates villosissimus. Véase Esteban et al. (2001) y GHUNLPam 435/2, 2371/8, 2371/46, 5395, 5451/8, 5504/1, 5504/2, 5504/3, 5504/4, 5504/6, 5504/9, 5504/10, 5505/7, 5505/10, 5516/15, 5679/20, 5679/31, 5711/9, 8066/29, 9081/4, 9570/17, 9570/21, 9570/24, 12728/1, 12771/1, 14068/3, 14083/1 y 14083/3 (Bajo Giuliani); 2038/9, 2038/20 y 5535/2 (Barrancas Coloradas); 14246/21, 14246/38, 14280/4, 14280/8, 22634/16 y 22634/25 (Estancia Ré); 22001/1 (Cerro La Bota); 2099/16, 5568/8, 5572/8, 5575/3, 5575/4, 5575/5, 5575/7, 5627/14, 8404/1, 8606/17, 8606/34, 14447/13, 14651/4, 19037/7, 19038/2 y 21740/2 (Laguna Chillhué); 14815/4, 14816/4, 14816/5 y 19057/2 (La Salada); 14177/6, 14177/27, 14177/28, 14177/31, 14177/33, 14177/34, 14177/38, 14177/39, 14177/40, 14177/41, 14177/64, 14177/121, 14177/301 y 14286/9 (Puesto Colorado); 5816/17, 5816/18, 5816/19, 5896/14, 5896/15, 5896/21, 5896/23, 5896/43, 5896/45, 8198/54, 8679/12, 8679/20, 8679/21, 8679/22, 8679/23, 8679/24, 8679/26, 8679/27, 8679/28, 8679/29, 8679/44, 9146/11, 9701/1, 9701/2, 9701/3, 9701/4, 9701/5, 9701/7, 14400/1, 14400/2, 14738/1, 14738/2 y 21180/4 (Quehué); 8078/6 y 8087/6 (Estancia Quiñi Malal); 125, 5656/5, 5656/7, 8275/5, 8275/14, 8275/16, 8275/18, 8275/19, 8974/112, 8974/124, 8974/132, 14008/13 y 14496/7 (Salinas Grandes de Hidalgo); 8231/1, 8231/4, 8231/6, 8231/7, 8231/8, 8231/9, 8253/16, 8253/70, 8253/72, 8253/75, 8253/88, 8253/91, 8253/102, 8253/103, 8253/108, 8253/110, 8253/112, 8253/115, 8253/116, 8253/118, 8253/119, 8253/120, 8253/123, 8253/131, 8253/132, 8253/138, 8253/139, 8253/142, 8253/144, 8253/151, 8253/159, 8419/2, 8453/3, 8453/8, 8453/49, 8453/68, 8453/122, 8453/128, 8453/152, 8453/153, 8453/171, 8453/182, 8698/3, 8698/27, 8698/28, 8698/34, 8698/51, 8698/53, 8698/61, 9059/22, 9059/29, 9059/42, 9064/17, 9064/20, 9065/2, 9065/6, 9065/19, 9065/33, 9066/19, 9067/33, 9067/46, 9067/51, 9067/65, 9067/90, 9067/98, 9067/111, 9067/141, 9098, 9367/25, 9367/26, 9367/27, 9367/28, 9367/33, 9367/39, 9367/42, 9367/46, 9478/24, 9478/28, 9478/30, 9478/31, 9478/34, 9478/71, 9642/37, 9642/64, 9642/69, 9642/71, 9642/72, 9642/73, 9642/74, 9642/75, 12812/1, 12812/4, 12896/5, 12896/34, 12896/45, 12896/55, 12896/56, 12896/57, 12896/61, 12896/83, 12922/12, 12922/14, 12922/15, 12922/16, 14623/9, 14623/15, 14623/23, 14623/26, 14623/27, 21170/1, 21648/32, 21648/40, 21648/48, 21648/50, 21648/65, 21648/75, 22709/2, 22709/3, 22709/4, 22709/32, 22726/7, 22726/10 y 22726/12 (Telén).
Chorobates cf. C. villosissimus GHUNLPam 19596/50 (Caleufú); 2283/1, 5066/5, 5348, 5465/1, 5465/4, 12964/2, 14853/1, 14853/2, 14853/3 y 14853/4 (El Guanaco). Se incluyen aquí los ejemplares GHUNLPam 19239/1, 19239/19, 19239/35, 19239/73, 19239/97, 19239/100, 19596/19, 19596/20, 19596/23, 19596/26, 19596/52, 21561/1, 21561/2, 21561/3, 21561/4, 21561/5, 21561/6, 21561/7, 21561/8, 21561/9, 21561/10, 21561/11, 21561/12 y 21561/13 que fueron mencionados por Esteban et al. (2003) como Chorobates villosissimus.
Macrochorobates scalabrinii. Véase Esteban et al. (2001) y GHUNLPam 6731/3 y 8011/6 (Cerro La Bota); 2291/45, 2435/1 y 5574/1 (Laguna Chillhué); 5512/9, 5512/10, 5512/12, y 5512/13 (Estancia Don Mariano); 2109/5, 2109/21, 2109/23, 14318/19 y 14318/20 (Salinas Grandes de Hidalgo); 8453/130, 8453/132, 8453/147, 8453/168, 8453/173, 8453/175, 8453/181, 8453/184, 8453/191, 8453/194, 8453/251, 9065/26, 12896/4 y 12925/5 (Telén).
Macrochorobates chapalmalensis GHUNLPam 5976/1 y 12965 (El Guanaco); 2038/9 (Barrancas Coloradas).
Macroeuphractus morenoi. Véase Esteban et al. (2001; 2003) y GHUNLPam 2038/7 (Barrancas Coloradas); 9717/28 (Estancia Ré); 2315/21, 5434/2, 5434/3, 5577/1, 5627/3, 5627/5 y 21740/6 (Laguna Chillhué); 14177/56, 14177/59 y 14177/65 (Puesto Colorado); 5816/11, 5816/12, 5816/13, 5816/14, 5816/15, 8198/96, 8680/1 y 8680/7 (Quehué); 191, 2109/2, 2109/13, 2109/16, 2109/19, 5656/8, 5656/10, 8275/20, 8275/21, 9165/7, 9165/8, 9165/9, 9165/10, 9165/11, 9165/12, 14008/2, 14008/17, 14305/5, 14305/6, 14314, 14318/15, 14318/18, 14318/21, 14318/23, 14318/24 y 14318/25 (Salinas Grandes de Hidalgo); 8253/4, 8253/156, 8253/157, 8253/158, 8453/136, 8453/137, 8453/149, 8453/156, 8453/157, 8453/159, 8453/162, 8453/164, 8453/169, 8453/176, 8453/189, 9059/4, 9059/50, 9062/25, 9064/1, 9367/1, 9367/2, 9367/11, 9478/1, 14623/1, 14623/2, 14624/2, 14624/3, 22726/3 y 22732/1 (Telén).
Macroeuphractus retusus GHUNLPam 14000 (Salinas Grandes de Hidalgo).
Proeuphractus limpidus GHUNLPam 8445/6 (Cerro La Bota); 2291/66, 8606/35, 8606/36, 14447/16 y 19037/5 (Laguna Chillhué); 21648/14 (Telén).
Proeuphractus sp. Véase Esteban et al. (2001; 2003) y GHUNLPam 8445/3 y 8445/4 (Cerro La Bota); 8083/1 (Cerro Patagua); 2163/6, 2163/11, 2291/4, 2311/2, 2386/5 y 8606/3 (Laguna Chillhué); 2109/18 (Salinas Grandes de Hidalgo).
Euphractini incertae sedis GHUNLPam 9257/193 y 14818/4 (La Salada); 8679/50, 9688/49, 9688/52, 12987/9, 14400/3, 14400/4 y 14400/5 (Quehué); 8253/50, 8253/122, 8320/9 8419/3 9367/46, 12922/2, 12922/4, 12922/5, 12922/6, 12922/7, 14866/5, 22709/15 y 22709/16 (Telén); 14246/18 (Conhelo).
Agradecimientos
A M. Reguero, S. Bargo y A. Kramarz, quienes facilitaron la revisión de materiales a su cargo. A E. Brussa por la lectura crítica del manuscrito. A G. Esteban y D.H. Verzi, cuyos comentarios en la revisión contribuyeron a mejorar este trabajo. La Facultad de Ciencias Exactas y Naturales de la Universidad Nacional de La Pampa financió este trabajo.
Bibliografía
1. Abello, A., Montalvo, C.I. y Goin, F. 2002. Marsupiales del Mioceno superior de Caleufú (La Pampa, Argentina). Ameghiniana 39: 433-442. [ Links ]
2. Ameghino, F. 1886. Contribución al conocimiento de los mamíferos fósiles de los terrenos terciarios antiguos del Paraná. Boletín de la Academia Nacional de Ciencias en Córdoba 9: 5-228. [ Links ]
3. Ameghino, F. 1887. Apuntes preliminares sobre algunos mamíferos extinguidos del yacimiento de Monte Hermoso existentes en el Museo de La Plata. Boletín del Museo de La Plata 1: 1-20. [ Links ]
4. Ameghino, F. 1889. Contribución al conocimiento de los mamíferos fósiles de la República Argentina. Actas de la Academia Nacional de Ciencias (Córdoba) 6: 1-1028. [ Links ]
5. Ameghino, F. 1891. Caracteres diagnósticos de cincuenta especies nuevas de mamíferos fósiles argentinos. Revista Argentina de Historia Natural 1: 129-167. [ Links ]
6. Ameghino, F. 1908. Las formaciones sedimentarias de la región litoral de Mar del Plata y Chapadmalal. Anales del Museo Nacional de Buenos Aires 10: 343-428. [ Links ]
7. Ameghino, F. 1920. Sur les édentés fósiles de l'Argentine. (Examen critique, revisión et correction de l'ouvrage de Mr. R. Lydekker: "The extinct edentates of Argentina", etc.). Obras Completas y Correspondencia Científica de Florentino Ameghino 11: 447-909. La Plata. [ Links ]
8. Bonaparte, C.L.J.L. 1838. Synopsis vertebratorum systematic. Nuovi Annales Scientiae Naturae vol. 2, anno 1: 105-133. [ Links ]
9. Bordas, A.F. 1932. Proposición de nuevo género para Eutatus inornatus. Physis 11: 167-168 [ Links ]
10. Bordas, A.F. 1933. Notas sobre los Eutatinae. Nueva Subfamilia extinguida de Dasypodidae. Anales del Museo Nacional de Historia Natural 37: 583-614. [ Links ]
11. Campbell, K. y Tonni, E. 1980. A new genus of teraton from the Huayquerian of Argentina (Aves: Teratornithidae). Contributions in Science. Natural History Museum Los Angeles County 330: 59-68. [ Links ]
12. Castellanos, A. 1947. Nuevos restos de la coraza de los géneros Proeuphractus y Macroeuphractus Ameghino descubiertos en el "Araucanense" del Valle de Yocavil (provincias de Catamarca y Tucumán). Publicaciones del Instituto de Fisiografía y Geología de Rosario 32: 1-44. [ Links ]
13. Cerdeño, E. 2000. Los mamíferos de la Formación Arroyo Chasicó (Mioceno superior), provincia de Buenos Aires, del Museo "J. C. Moyano" de Mendoza. Ameghiniana 37: 503-508. [ Links ]
14. Cerdeño, E. y Montalvo, C.I. 2001. Los Mesotheriinae (Mesotheriidae, Notoungulata) del Mioceno superior de La Pampa, Argentina. Revista Española de Paleontología 16: 63-75. [ Links ]
15. Cerdeño, E. y Montalvo, C.I. 2002. Los Hegetotheriinae (Hegetotheriidae, Notoungulata) del Mioceno superior de la provincia de La Pampa, Argentina. Revista del Museo Argentino de Ciencias Naturales n. s. 4: 35-43. [ Links ]
16. Cione, A.L. y Tonni, E.P. 2005. Bioestratigrafía basada en mamíferos del Cenozoico superior de la provincia de Buenos Aires, Argentina. En: R. de Barrio, R. Etcheverry, M Cavallé y E. Llambías (eds.), Geología y recursos minerales de la provincia de Buenos Aires. 16º Congreso Geológico Argentino (La Plata) Relatorio, pp. 183-200. [ Links ]
17. Cione, A., Azpelicueta, M., Bond, M., Carlini, A., Casciota, J., Cozzuol, M., de la Fuente, M., Gasparini, Z., Goin, F., Noriega, J., Scillato-Yané, G., Soibelzon, L., Tonni, E., Verzi, D. y Vucetich, M. 2000. Miocene Vertebrates from Entre Ríos province, eastern Argentina. INSUGEO, Serie Correlación Geológica 14: 191-237. [ Links ]
18. Cope, E.D. 1889. The Edentata of North America. American Naturalist 32: 657-664. [ Links ]
19. Esteban, G. y Nasif, N. 1996. Nuevos Dasypodidae (Mammalia, Xenarthra) del Mioceno tardío del Valle del Cajón, Catamarca, Argentina. Ameghiniana 33: 327-334. [ Links ]
20. Esteban, G. y Nasif, N. 1999. Mamíferos fósiles de la Formación Chiquimil (Mioceno tardío), provincia de Catamarca, Argentina. Bioestratigrafía. Ameghiniana Suplemento Resúmenes 36: 11R. [ Links ]
21. Esteban, G., Nasif, N. y Montalvo, C.I. 2001. Nuevos registros de Dasypodidae (Xenarthra) del Mioceno tardío de la provincia de La Pampa (Argentina). Revista Española de Paleontología 16: 77-87. [ Links ]
22. Esteban, G., Nasif, N., Montalvo C.I. y Visconti, G. 2003. Nuevos registros de Dasypodidae (Xenarthra) en la Formación Cerro Azul (Mioceno tardío) de Caleufú, La Pampa, Argentina. Ameghiniana 40: 495-499. [ Links ]
23. Georgieff, S., Herbst, R., Esteban, G. y Nasif, N. 2004. Análisis paleoambiental y registro paleontológico de la Formación Desencuentro (Mioceno superior), Alto de San Nicolás, La Rioja, Argentina. Ameghiniana 41: 45-56. [ Links ]
24. Gervais, P. 1867. Sur une nouvelle colection d'ossements fossiles de Mammifères recueillé par M. Fr. Seguin dans la Confédération Argentine. Comptes rendus des séances de l'Académie des Sciences 65: 279-289. [ Links ]
25. Goin, F. y Montalvo, C.I. 1988. Revisión sistemática y reconocimiento de una nueva especie del género Thylatheridium Reig (Marsupialia, Didelphidae). Ameghiniana 25: 161-167. [ Links ]
26. Goin, F., Montalvo, C.I. y Visconti, G. 2000. Los marsupiales (Mammalia) del Mioceno superior de la Formación Cerro Azul (provincia de La Pampa, Argentina). Estudios Geológicos 56: 101-126. [ Links ]
27. Illiger, C. 1811. Prodromus Systematics Mammalium et Avium Additis Terminis Zoographicis Utriusque Cassis. Salfeld, Berlín, 301 pp. [ Links ]
28. Kraglievich, L. 1934. La antigüedad pliocena de las faunas de Monte Hermoso y Chapadmalal, deducidas de su comparación con las que le precedieron y sucedieron. Imprenta "El Siglo Ilustrado", 136 pp. [ Links ]
29. Linares, E., Llambías, E. y Latorre, C. 1980. Geología de la provincia de La Pampa, República Argentina y geocronología de sus rocas metamórficas y eruptivas. Revista de la Asociación Geológica Argentina 35: 87-146. [ Links ]
30. Linné, C. 1758. Systema naturae. L. Salvii, Stockholm, 10, 1, 824 p. [ Links ]
31. Lydekker, R. 1894. Contribution to a knowledge of the fossil vertebrates of Argentina. 2. The extinct Edentates of Argentina. Anales del Museo de La Plata, sección Paleontología 3: 1-118. [ Links ]
32. Melchor, R., Visconti, G. y Montalvo, C.I. 2000. Late Miocene calcic vertisols from central La Pampa, Argentina. 2° Congreso Latinoamericano de Sedimentología y 7° Reunión Argentina de Sedimentología, (Mar del Plata), Actas: 119-120. [ Links ]
33. Mones, A. y Mehl, J. 1990. La presencia de Macroeuphractus aff. moreni (Lydekker, 1895), en la Formación La Paz (Plioceno), Bolivia. Aspectos taxonómicos y bioestratigráficos (Mammalia: Cingulata: Dasypodidae). Comunicaciones Paleontológicas del Museo de Historia Natural de Montevideo 22: 17-31. [ Links ]
34. Montalvo, C.I. y Casadío, S. 1988. Presencia del género Palaeoctodon (Rodentia, Octodontidae) en el Huayqueriense (Mioceno tardío) de la provincia de La Pampa. Ameghiniana 25: 111-114. [ Links ]
35. Montalvo, C.I. y Szelagowski, M. 1999. Vertebrados del Mioceno superior en la Colección Paleontológica de la Universidad Nacional de La Pampa. 7° Jornadas Pampeanas de Ciencias Naturales (Santa Rosa), Actas: 233-242. [ Links ]
36. Montalvo, C.I. y Verzi, D.H. 2004. El registro de roedores Caviomorpha de la Formación Cerro Azul (Mioceno tardío, La Pampa, Argentina): sistemática, biocronología, evolución y paleoclimas. Ameghiniana Suplemento Resúmenes 41: 56R. [ Links ]
37. Montalvo, C.I., Cardonatto, M., Visconti, G., Verzi, D.H. y Vucetich, M.G. 1996. Vertebrados de la Formación Cerro Azul (Mioceno tardío) del Valle de Quehué, provincia de La Pampa, Argentina. 6° Jornadas Pampeanas de Ciencias Naturales (Santa Rosa), Actas: 159-165. [ Links ]
38. Montalvo C.I., Melchor, R.N., Visconti, G. y Cerdeño, E. 2008. Vertebrate taphonomy in loess-paleosol deposits: a case study from the Late Miocene of central Argentina. Geobios 41: 133- 143. [ Links ]
39. Montalvo, C.I., Urrutia, J.J., Visconti, G. y Verzi, D.H. 2005. Tafonomía de la asociación faunística de la Formación Cerro Azul en Cerro de la Bota (Mioceno tardío, La Pampa, Argentina), Ameghiniana Suplemento Resúmenes 42: 35R. [ Links ]
40. Montalvo, C.I., Verzi, D.H., Vucetich, M.G. y Visconti, G. 1998. Nuevos Eumysopinae (Rodentia, Echimyidae) de la Formación Cerro Azul (Mioceno tardío) de La Pampa, Argentina. 5° Jornadas Geológicas y Geofísicas Bonaerenses (Mar del Plata), Actas 1: 57-64. [ Links ]
41. Montalvo, C.I., Visconti, G., Púgener, L. y Cardonatto, M. 1995. Mamíferos de Edad Huayqueriense (Mioceno tardío), Laguna Chillhué, provincia de La Pampa. 4° Jornadas Geológicas y Geofísicas Bonaerenses (Junín), Actas 1: 73-79. [ Links ]
42. Moreno, F.P. 1889. Breve reseña de los progresos del Museo La Plata, durante el segundo semestre de 1888. Boletin del Museo LaPlata 3: 1-44. [ Links ]
43. Moreno, F.P. y Mercerat, A. 1891. Exploracion arqueológica de la provincia de Catamarca. Paleontología. Revista del Museo de La Plata 1: 222-236. [ Links ]
44. Ortiz Jaureguizar, E. 1998. Paleoecología y evolución de la fauna de mamíferos de América del Sur durante la "Edad de las Planicies Australes" (Mioceno superior-Plioceno superior). Estudios Geológicos 54: 161-169. [ Links ]
45. Pascual, R. y Bocchino, A. 1963. Un nuevo Borhyaeninae (Marsupialia) del Plioceno medio de Hidalgo (La Pampa). Ameghiniana 3: 97-107. [ Links ]
46. Pascual, R. y Bondesio, P. 1982. Un roedor Cardiatheriinae (Hydrochoeridae) de la Edad Huayqueriense (Mioceno tardío) de La Pampa. Sumario de los ambientes terrestres en la Argentina durante el Mioceno. Ameghiniana 19: 19-35. [ Links ]
47. Pascual, R., Pisano, J. y Ortega, E. 1965. Un nuevo Octodontidae (Rodentia, Caviomorpha) de la Formación Epecuén (Plioceno medio) de Hidalgo (provincia de La Pampa). Consideraciones sobre los Ctenomyinae Reig, 1958, y la morfología de sus molariformes. Ameghiniana 4: 19-30. [ Links ]
48. Perea, D. y Scillato-Yané, G.J. 1995. Proeuphractus limpidus Ameghino, 1886 (Xenarthra, Dasypodidae, Euphractini): Osteología comparada del cráneo y elementos de la coraza asociados (Neógeno del Uruguay). Boletín de la Real Sociedad Española de Historia Natural (Sección Geología) 90: 125-130. [ Links ]
49. Pocock, R. I. 1924. The external characters of the South American edentates. Procedures of the Zoological Society of London 65: 983- 1031. [ Links ]
50. Reig, O. 1958. Notas para una actualización del conocimiento de la fauna de la Formación Chapadmalal. I Lista faunística preliminar. Acta Geológica Lilloana 11: 241-253 [ Links ]
51. Rovereto, C. 1914. Los estratos araucanos y sus fósiles. Anales del Museo de Historia Natural de Buenos Aires 25: 1-250. [ Links ]
52. Scillato-Yané, G.J. 1975. Presencia de Macroeuphractus retusus (Xenarthra, Dasypodidae) en el Plioceno del área Mesopotamia (Argentina). Su importancia bioestratigráfica y paleobiogeográfica. Ameghiniana 12: 322-328. [ Links ]
53. Scillato-Yané, G.J. 1979. Notas sobre los Dasypodidae (Mammalia, Edentata) del Plioceno del territorio argentino. I Los restos de Edad Chasiquense (Plioceno inferior) del sur de la provincia de Buenos Aires. Ameghiniana 14: 133-144. [ Links ]
54. Scillato-Yané, G.J. 1980. Catálogo de los Dasypodidae fósiles (Mammalia, Edentata) de la República Argentina. 2° Congreso Argentino de Paleontología y Bioestratigrafía y 1° Congreso Latinoamericano de Paleontología (Buenos Aires, 1978), Actas 3: 7-36. [ Links ]
55. Scillato-Yané, G.J. 1982. [Los Dasypodidae (Mammalia-Edentata) del Plioceno y Pleistoceno de Argentina. Tesis Doctoral. Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, 159 pp. Inédita.]. [ Links ]
56. Scillato-Yané, G.J., Carlini, A.A., Vizcaíno, S.F. y Ortiz Jaureguizar, E. 1995. Los Xenartros. En: M.T. Alberti, G. Leone y E.P. Tonni (eds.), Evolución biológica y climática de la región pampeana durante los últimos cinco millones de años. Un ensayo de correlación con el Mediterráneo occidental. Monografías. Museo Nacional de Ciencias Naturales y Consejo Superior de Investigaciones Científicas, Madrid, pp. 181-209. [ Links ]
57. Tambussi, C. 1987. Catálogo crítico de los Tinamidae (Aves: Tinamiformes) fósiles de la República Argentina. Ameghiniana 24: 241-244. [ Links ]
58. Tauber, A.A. (h). 2005. Mamíferos fósiles y edad de la Formación Salicas (Mioceno tardío) de la sierra de Velasco, La Rioja, Argentina. Ameghiniana 42: 443-460. [ Links ]
59. Tonni, E., Prado, J., Fidalgo, F. y Laza, J. 1992. El Piso/Edad Montehermosense (Plioceno) y sus mamíferos. 3° Jornadas Geológicas Bonaerenses, (La Plata), Actas: 113-118. [ Links ]
60. Tonni, E., Scillato-Yané, G.J., Cione, A. y Carlini, A. 1998. Bioestratigrafía del Mioceno continental en el curso inferior del Arroyo Chasicó, provincia de Buenos Aires. 7° Congreso Argentino de Paleontología y Bioestratigrafía (Bahía Blanca), Actas: 135. [ Links ]
61. Urrutia, J. y Scillato-Yané, G.J. 2003. Registro de Macroeuphractus retusus (Dasypodidae, Euphractini) en la Formación Cerro Azul (Mioceno tardío) en Salinas Grandes de Hidalgo, La Pampa, Argentina. Ameghiniana Suplemento Resúmenes 40: 95R. [ Links ]
62. Verzi, D.H. 1999. The dental evidence on the differentiation of the ctenomyine rodents (Caviomorpha, Octodontidae, Ctenomyinae). Acta Theriologica 44: 263-282. [ Links ]
63. Verzi, D.H. 2002. Patrones de evolución morfológica en Ctenomyinae (Rodentia, Octodontidae). Mastozoología Neotropical 9: 309-328. [ Links ]
64. Verzi, D.H., Montalvo, C.I. y Deschamps, C. 2008. Biostratigraphy and biochronology of the Late Miocene of central Argentina: evidence from rodents and taphonomy. Geobios 41: 145-155. [ Links ]
65. Verzi, D.H., Montalvo, C.I. y Tiranti, S. 2003. Un nuevo Xenodontomys (Rodentia, Octodontidae) del Mioceno tardío de La Pampa. Patrón evolutivo y biocronología. Ameghiniana 40: 229-238. [ Links ]
66. Verzi, D.H., Montalvo, C.I. y Vucetich, M.G. 1991. Nuevos restos de Xenodontomys simpsoni Kraglievich y la sistemática de los más antiguos Ctenomyinae (Rodentia, Octodontidae). Ameghiniana 28: 325-331. [ Links ]
67. Verzi, D.H., Montalvo, C.I. y Vucetich, M.G. 1999. Afinidades y significado evolutivo de Neophanomys biplicatus (Rodentia, Octodontidae) del Mioceno tardío-Plioceno temprano de Argentina. Ameghiniana 36: 83-90. [ Links ]
68. Verzi, D.H., Vieytes, E.C. y Montalvo, C.I. 2004. Dental evolution in Xenodontomys and first notice on secondary acquisition of radial enamel in rodents (Rodentia, Caviomorpha, Octodontidae). Geobios 37: 795-806 [ Links ]
69. Verzi, D.H., Vucetich, M.G. y Montalvo, C.I. 1994. Octodontid-like Echimyidae (Rodentia): an upper Miocene episode in the radiation of the family. Palaeovertebrata 23: 199-210. [ Links ]
70. Verzi, D.H., Vucetich, M.G. y Montalvo, C.I. 1995. Un nuevo Eumysopinae (Rodentia Echimyidae) del Mioceno tardío de la provincia de La Pampa y consideraciones sobre la historia de la Subfamilia. Ameghiniana 32: 191-195. [ Links ]
71. Zetti, J. 1972. [Los mamíferos fósiles de edad Huayqueriense (Plioceno medio) de la región pampeana. Tesis Doctoral. Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, 86 pp. Inédita.]. [ Links ]
Recibido: 12 de julio de 2006.
Aceptado: 2 de junio de 2008.

