










Serviços Personalizados
Journal

Artigo
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAmeghiniana
versão On-line ISSN 1851-8044
Ameghiniana v.45 n.2 Buenos Aires abr./jun. 2008
Cenozoic trace fossils of the Cruziana, Zoophycos and Nereites ichnofacies from the Fuegian Andes, Argentina
María I. López Cabrera1, Eduardo B. Olivero1, Noelia B. Carmona1 and Juan J. Ponce1
1Centro Austral de Investigaciones Científicas (CADIC-CONICET): Bernardo A. Houssay 200, 9410 Ushuaia Tierra del Fuego, Argentina. emolivero@gmail.com
Abstract. The systematics, paleoenvironmental implications, and diversity of Cenozoic trace fossils from the Fuegian Andes are studied. The relatively complete Paleocene-Miocene stratigraphic column includes ichnoassemblages of the Cruziana (Leticia Formation, late Middle Eocene), Zoophycos and Nereites (Early Eocene-Early Miocene turbidite systems) ichnofacies. The last two ichnoasemblages contain the only known deep marine Cenozoic trace fossils in Argentina. The late Middle Eocene Leticia Formation represents a transgressive-regressive cycle and bears the ichnogenera Curvolithus, Diplocraterion, Gyrochorte, Rosselia, Patagonichnus, Asterosoma, Palaeophycus, Paradictyodora, Planolites, Rhizocorallium, Schaubcylindrichnus, Taenidium, and Teichichnus. Ichnogenera of the Early Eocene-Early Miocene turbidite systems include Scolicia, Chondrites, Gyrophyllites, Nereites, Phycodes, Phycosiphon, Phymatoderma, Stelloglyphus, Zoophycos, Ophiomorpha and graphoglyptids. Graphoglyptids are dominated by Paleodictyon, Helicolithus, Helminthorhaphe, Desmograpton and Megagrapton. They are recorded in thin-bedded turbidites and mudstones (lobe deposits) and assigned to the Paleodictyon ichnosubfacies (Nereites ichnofacies). Ophiomorpha rudis and O. annulata are common at the contact between thick-bedded turbidites and mudstones, with abundant plant fragments. In sandrich, proximal channel-lobe deposits, they characterize the Ophiomorpha rudis ichnosubfacies (Nereites ichnofacies). Scolicia prisca and Nereites isp. are common in rippled fine-grained sandstones interbedded with thin mudstones. Zoophycos ispp. are dominant in slope mudstones with synsedimentary slumping. The maximum ichnodiversity is recorded in the late Middle-Late Eocene; which is concomitant with a marked cooling trend. The basal Oligocene displays an abrupt drop in diversity, whereas the Early Miocene shows a moderate diversity. These data do not support the alleged control of increased Eocene ichnodiversity by global warming during the Cenozoic thermal maximum. Specialized food competition, particularly for the graphoglyptid organisms, and generalized oligotrophy seem to offer a better explanation.
Resumen. Trazas fósiles de la Ichnofacies de Cruziana, Zoophycos y Nereites del Cenozoico de los Andes Fueguinos. Se dan a conocer resultados de estudios sistemáticos, implicancias paleoambientales y diversidad de trazas fósiles del Paleoceno- Mioceno de los Andes Fueguinos. Se reconocen las icnofacies de Cruziana (Formación Leticia, Eoceno Medio alto), y de Zoophycos y Nereites (sistemas de turbiditas del Eoceno-Mioceno temprano). Las dos últimas icnofacies constituyen las únicas icnoasociaciones marinas profundas del Cenozoico conocidas en la Argentina. La Formación Leticia representa un ciclo transgresivo- regresivo e incluye a Curvolithus, Diplocraterion, Gyrochorte, Rosselia, Patagonichnus, Asterosoma, Palaeophycus, Paradictyodora, Planolites, Rhizocorallium, Schaubcylindrichnus, Taenidium, y Teichichnus. Los sistemas de turbiditas del Eoceno temprano-Mioceno temprano incluyen a Scolicia, Chondrites, Gyrophyllites, Nereites, Phycodes, Phycosiphon, Phymatoderma, Stelloglyphus, Zoophycos, Ophiomorpha, y grafoglíptidos. Paleodictyon, Helicolithus, Helminthorhaphe, Desmograpton y Megagrapton son típicos grafoglíptidos de la icnosubfacies de Paleodictyon (icnofacies de Nereites) y se encuentran en turbiditas finas (depósito de lóbulos). Ophiomorpha rudis y O. annulata caracterizan la icnofacies de O. rudis (icnofacies de Nereites), en turbiditas medias a gruesas, con abundante materia vegetal (depósitos arenosos proximales de canales y lóbulos). Scolicia prisca y Nereites isp. son comunes en areniscas finas con ondulitas e intercaladas con pelitas. Zoophycos ispp. domina en fangolitas de talud con deslizamientos sinsedimentarios. Concordantemente con una marcada tendencia de enfriamiento, el Eoceno Medio superior- Eoceno Superior registra la máxima icnodiversidad, el Oligoceno basal una caída abrupta en la diversidad y el Mioceno temprano una moderada recuperación de la diversidad. Estos datos no concuerdan con la teoría de aumento de icnodiversidad debido al calentamiento global durante el máximo termal (Paleoceno/Eoceno) del Cenozoico. La especialización trófica desarrollada por los organismos productores de grafoglíptidos y una oligotrofía generalizada ofrecen una mejor explicación para estas tendencias.
Key words. Fuegian Andes; Cenozoic; Ichnology; Graphoglyptids; Turbidites.
Palabras clave. Andes Fueguinos; Cenozoico; Icnología; Grafoglíptidos; Turbiditas.
Introduction
The Fuegian Andes region has a relatively complete, about 4 km thick stratigraphic column of Paleocene to Miocene sedimentary rocks, which includes the only known record of deep-marine Cenozoic deposits in Argentina. These circumstances make the Fuegian Andes as one of the most important areas of southern South America for the study of Cenozoic rocks (Olivero and Malumián, 1999, 2002; Olivero et al., 2002, 2003; Malumián and Olivero, 2005, 2006).
Except for few taxonomic mentions and brief localized studies (e.g. Olivero and López C., 2001; Olivero et al., 2005; Carmona et al., 2006; López C. et al., 2006, and Ponce et al., 2007) the ichnology of this important area is almost unknown. In this paper we attempt to present a general ichnological study of the Cenozoic of the Fuegian Andes. The study covers basinal and/or slope turbidite systems and marginal marine and shelf settings. Turbidite systems include the Zoophycos and Nereites ichnofacies of the Lower Eocene Punta Torcida Formation; upper Middle to Upper Eocene Cerro Colorado Formation; Lower Oligocene Estancia María Cristina beds; uppermost Oligocene-Lower Miocene Desdémona Formation; and Lower Miocene Cabo Ladrillero and Cabo San Pablo beds. Marginal marine and shelf settings include the Cruziana ichnofacies of the late Middle Eocene Leticia Formation.
The objectives of this study are to document partly the ichnological record of the Cenozoic of the Fuegian Andes and to discuss its implications for general characterization of paleoenvironmental conditions and contribution to global ichnodiversity trends, in particular to the known increase in deepmarine trace fossil diversity during the Eocene. The systematic description covers mainly the ichnology of the turbidite systems, which includes several ichnospecies distributed in 16 ichnogenera that were mostly unknown in the Cenozoic of Argentina. The systematics of the bulk of trace fossils from the Leticia Formation (Cruziana ichnofacies) is the subject of an ongoing study by the authors. Thus, most of them will only be mentioned here except for some well preserved ichnotaxa, such as Curvolithus and Gyrochorte, which are barely known in the Cenozoic.
Late Cretaceous-Cenozoic stratigraphic framework
In southeastern Tierra del Fuego (figure 1), the Late Cretaceous-Cenozoic stratigraphy of the foreland Austral Basin consists of four major synorogenic clastic wedges, which are mainly characterized by thick, deep marine siliciclastic deposits (Olivero and Malumián, 2002). Within the fold and thrust belt, the deposits of the older clastic wedges include a Maastrichtian- Danian succession, the Late Paleocene-Early Eocene Río Claro Group, and the late Middle to Late Eocene La Despedida Group. These deposits are highly deformed and reach an aggregate thickness in excess of 3.500 m (Olivero et al., 2002; 2003). The youngest, Oligocene-Miocene clastic wedge is principally exposed outside the folded belt and consists mainly of subhorizontal strata, the c. 1 km thick Cabo Domingo Group (Malumián and Olivero, 2006).
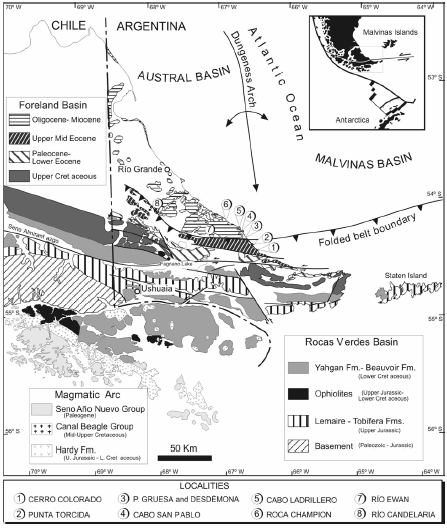
Figure 1. Geological sketch map of Tierra del Fuego, Argentina with location of studied sections / bosquejo geológico de Tierra del Fuego, Argentina con la situación de las secciones estudiadas.
At the Cabo Campo del Medio-Punta Torcida anticline (figure 1), a very thick (c. 1.600 m) Eocene sedimentary succession comprises the Punta Torcida Formation, Early Eocene; the Leticia Formation, late Middle Eocene; and the Cerro Colorado Formation, late Middle to Late Eocene (figure 2).

Figure 2. Schematic Early Eocene-Early Miocene stratigraphic column with location of main trace fossil bearing horizons and generalized paleoenvironmental conditions / columna estratigráfica esquemática del Eoceno Temprano-Mioceno Temprano con la situación de los principales horizontes con trazas fósiles y paleoambientes generalizados.
The Punta Torcida Formation of the Río Claro Group is dominated by dark gray mudstones deposited under dysaerobic conditions, with sparse intercalation of thin-bedded turbidite sandstones (c. 220 m thick). At the northern limb of the anticline, the upper part of the Punta Torcida Formation includes an additional, unnamed sedimentary package (c. 225 m thick) consisting of thick-bedded turbidite sandstones, tuffs, and distorted mudstones. Trace fossils, including graphoglyptids, are common in some horizons of the unnamed, upper package. These deposits conform a turbidite system that includes the following elements: basin (basal, thin bedded turbidites); base of slope (mid, thick-bedded turbidites); and slope (upper, distorted mudstones). Planktonic foraminifers indicate a middle Early Eocene age equivalent to the G. wilcoxensis Zone from New Zealand, or the tropical equivalent Zone P7 to P8, or Zone AP6 from Antarctica (Olivero and Malumián, 1999).
La Despedida Group rests over the Río Claro Group on a deeply incised unconformity. The basal Leticia Formation is composed of three main sandstone- dominated intervals, which have a complex architecture and display variable thickness along the northern and southern limbs of the anticline, c. 200 m and 500 m, respectively. The lower and upper intervals consist of thick, cross-bedded and parallel-laminated, channelized sandstone bodies, with minor, highly bioturbated heterolithic beds, which represent deltaic and estuarine settings (Olivero and López C., 2001). The scarcity of foraminifers and the absence of planktonic foraminifers suggest marginal, shallow, and restricted marine conditions for the lower and upper intervals of the Leticia Formation (Olivero and Malumián, 1999). The middle interval consists of glauconitic, fossiliferous, and highly bioturbated fine-grained sandstones, representing shelf settings. Minor horizons bear planktonic foraminifers and nannoplankton indicative of the G. index Zone to G. inconspicua Zone of New Zealand, or the equivalent to the interval upper Zone P12 to lower Zone P14 of tropical areas, i.e., late Middle Eocene (Olivero and Malumián, 1999).
The overlying Cerro Colorado Formation (c. 855 m thick) consists of a vertical stacking of four coarsening and thickening upwards successions, included in the CCa, CCb, CCc, and CCd members, respectively. Each member is composed of dark gray mudstones at the base, regular intercalation of mudstones and turbidites at the mid part, and thick sandstones and pebbly sandstones at the top. The uppermost CCd member bears abundant radiolarians and planktonic foraminifera typical of an oxygen minimum habitat, indicative of a Late Eocene age (upper Zone P15 to upper Zone P16). The microfauna and stratigraphic position of the lower three members are consistent with a latest Middle Eocene age (mid Zone P14 to lower Zone P15, Olivero and Malumián, 1999). Foraminifers of the three lower members of the Cerro Colorado Formation represent neritic settings, whereas those in the uppermost CCd member record a marked deepening (Olivero and Malumián, 1999; Malumián and Olivero, 2006). The CCa, CCc, and CCd members have several horizons with abundant graphoglyptids.
The Cabo Domingo Group (Malumián and Olivero, 2006) started in the latest Eocene with a major deepening phase represented in the basin depocenter by the uppermost part of the Cerro Colorado Formation, which records the last appearance datum (LAD) of Globigerinatheka index and the first appearance datum (FAD) of Praetenuitella insolita and Isthmolithus recurvus. The following Estancia María Cristina beds (claystones and sandstones, earliest Oligocene) and Puesto Herminita beds (claystones, Oligocene), with residual agglutinated foraminifera, were deposited below the calcite compensation depth during the maximum deepening of the basin that allowed for the entrance of corrosive Antarctic waters. The Desdémona Formation (tuffaceous, bioturbated mudstones deposited at or near the lisocline, with large slumped blocks), the Cabo Ladrillero beds (glauconitic sandstones and mudstones with outer neritic foraminifers, and carbonaceous mudstones, fine-grained sandstones and claystones with residual assemblages of Spirosigmoilinella- Martinottiella), and the Cabo San Pablo beds (sandstones and claystones) represents the latest Oligocene-early Miocene high sea level conditions. An extended unconformity and generalized regression is represented at c. 21 Ma. The overlying Cabo Viamonte beds (coarse breccias, conglomerates, sandstones and mudstones) represent the Neogene high sea level and climatic optimum (Malumián and Olivero, 2006), but are not the subject of this paper.
Ichnotaxonomy
Trace fossils are ordered alphabetically and descriptions are restricted to selected taxa, which were chosen by their paleoenvironmental significance or rarity of distribution in the Cenozoic of southern South America. Most of the material was studied in the field; the repository of collected material is the Laboratorio de Geología Andina, Centro Austral de Investigaciones Científicas, and it is identified with the labels CADIC PI 20, and CADIC PI 46 to 58.
Ichnogenus Cardioichnus Smith and Crimes, 1983
Cardioichnus isp.
Figure 3.1
Figure 3. 1, Cardioichnus isp. Cabo San Pablo beds. CADIC PI 46. Positive hyporelief / hiporelieve positivo. 2, Cosmorhaphe isp. Cerro Colorado Formation. Positive hyporelief / hiporelieve positivo. 3-4, Curvolithus simplex. Leticia Formation. 3, Full relief preservation / preservación en relieve completo. 4, Lower bedding plane with bilobate, positive hyporelief and upper trilobate surface in negative hyporelief / vista basal en hiporelieve positivo bilobado y superficie trilobada en hiporelieve negativo. 5, Desmograpton ichthyforme. Cerro Colorado Formation. Positive hyporelief / hiporelieve positivo. 6, Gyrophyllites rehsteineri. Cabo Ladrillero beds. Full relief / relieve completo. 7, Gyrochorte comosa. Leticia Formation. Basal view showing two bilobate furrows separated by a narrow crest (arrows) crosscutting Curvolithus simplex (Cu) / vista basal mostrando dos surcos separados por una cresta (flechas) cortando a Curvolithus simplex (Cu). 8, Helicolithus ramosus. Cerro Colorado Formation. Positive hyporelief / hiporelieve positivo. 9, Helicolithus isp. Punta Torcida Formation. Positive hyporelief / hiporelieve positivo. 10, Helicorhaphe isp. Punta Torcida Formation. Positive hyporelief / hiporelieve positivo. Scale bar: 1 cm. Field phothographs except figure 3.1 / Escala: 1 cm. Fotografías de campo, excepto figure 3.1.
Material. Many field specimens and one collected slab (CADIC PI 46) with one specimen.
Description. Resting trace, ovoid to subquadrate in outline, continuous with slightly sinuous burrow, preserved as positive hyporelief.
Remarks. Poor preservation of central V-shaped area and the lack of spine impressions avoid ichnospecific assignment. Cardioichnus ranges from Lower Cretaceous (Monaco et al., 2005) to early Pliocene (Gibert and Martinell, 1994).
Occurrence. Cerro Colorado Formation, CCc member, late Middle Eocene and Cabo San Pablo beds, Early Miocene.
Ichnogenus Cosmorhaphe Fuchs, 1895
Cosmorhaphe isp.
Figure 3.2
Material. Field picture of one incomplete specimen.
Description. Hypichnial trace fossil with two orders of meanders. The strings are 2.7-2.8 mm in diameter.
Remarks. Incomplete preservation in a weathered surface avoids ichnospecific assignment. This graphoglyptid burrow is commonly found in the deepmarine Nereites ichnofacies. It ranges from Cambrian (Narbonne et al., 1987) to Holocene (Ekdale, 1980).
Occurrence. Cerro Colorado Formation, CCa member, late Middle Eocene.
Ichnogenus Curvolithus Fritsch, 1908
Curvolithus simplex Buatois, Mángano, Mikulás and Maples, 1998
Figures 3.3-4
Material. Abundant specimens studied in the field and one polished slab (CADIC PI 47) with several specimens.
Description. Horizontal to oblique, straight to slightly curved endichnial burrows. Upper surface is trilobate, with a central lobe wider than the marginal ones. The central lobe is commonly divided by a furrow. Lower surface is bilobate. Burrows are about 10 mm in width.
Remarks. Buatois et al. (1998) distinguished two ichnospecies of Curvolithus: C. multiplex Fritsch 1908, with a convex quadralobate lower surface, and C. simplex, with a uni- to trilobate concave or convex lower surface. Curvolithus differs from similar trace fossils, such as Psammichnites, Aulichnites and Gyrochorte, by its trilobate upper surface. It occurs from Precambrian to Miocene (Buatois et al., 1998).
Occurrence. Upper part of the Leticia Formation, late Middle Eocene.
Ichnogenus Desmograpton Fuchs, 1895
Desmograpton ichthyforme (Macsotay, 1967)
Figure 3.5
Material. Three specimens studied in the field.
Description. Hypichnial narrow parallel ridges corresponding to angular semi-meanders joined at the center in an alternate position, connecting bars are not preserved. The string is 1.5 to 2 mm in diameter.
Remarks. Seilacher (1977) includes Desmograpton in the graphoglyptid group of biramous meanders. It occurs from Silurian (McCann, 1989) to Miocene (Uchman, 1995).
Occurrence. Cerro Colorado Formation, CCa member, late Middle Eocene.
Ichnogenus Gyrophyllites Glocker, 1841
Gyrophyllites rehsteineri (Fischer-Ooster, 1858)
Figure 3.6
Material. Three field specimens.
Description. Radial trace fossil with horizontal and overlapping lobes that radiate from a vertical shaft. Lobes unbranched, drop-like in outline and with a rounded margin at the distal parts. In vertical section, lobes radiate at several levels. Total diameter is 50 mm. Endichnial preservation.
Remarks. The swollen, radial feeding galleries organized around a vertical axis, which do not branch or cut across each other, were considered as diagnostic features of Gyrophyllites (Fu, 1991). These features differentiate the present material from other ichnogenera with radial structure. Gyrophyllites is recorded from the Lower Paleozoic of Argentina (Mángano et al., 2005) to the Miocene (this paper).
Occurrence. Cabo Ladrillero beds, Early Miocene.
Ichnogenus Gyrochorte Heer, 1865
Gyrochorte comosa Heer, 1865
Figure 3.7
Material. Abundant specimens studied in the field and one polished slab (CADIC PI 48) with several specimens.
Description. Hypichnial, curved burrows composed of two bilobate furrows separated by a narrow crest. Polished sections show that the burrow is vertically extended.
Remarks. All the specimens are only preserved in basal view. Gyrochorte has a discontinuous stratigraphic record (cf. Gibert, 2003), and the material from Tierra del Fuego is the only one recorded in the Paleogene.
Occurrence. Lower and upper parts of the Leticia Formation, late Middle Eocene.
Ichnogenus Helicolithus Azpeitia-Moros, 1933
Helicolithus ramosus (Vialov, 1971)
Figure 3.8
Material. Many specimens studied in the field and two slabs (CADIC PI 49-50) with two specimens.
Description. Hypichnial meandering rows of small dots corresponding to a meandering and helicoidal string.
Remarks. Uchman (2001) referred this type of preservation to washed-out parts of Helicolithus when only the apices of the turns are preserved. Helicolithus is a very common graphoglyptid in the Eocene flysch of Tierra del Fuego. This ichnogenus ranges from Late Cretaceous to Miocene (Uchman, 2003).
Occurrence. Cerro Colorado Formation, CCa and CCd members, late Middle Eocene and Late Eocene, respectively.
Helicolithus isp.
Figure 3.9
Material. One field specimen and a plaster cast (CADIC PI 51).
Description. Small segment of a horizontal winding trace fossil, with second order of helicoidal turns comprising two rows of four strings of small bars displaying an alternate and parallel pattern. Preserved at the sole of a sandstone bed.
Remarks. The alternate and parallel pattern of the two rows of bars suggests the "echelon" pattern (Seilacher, 1977) of Helicolithus. Incomplete preservation avoids ichnospecific assignment.
Occurrence. Punta Torcida Formation, Early Eocene.
Ichnogenus Helicorhaphe
, 1961
Helicorhaphe isp.
Figure 3.10
Material. One field specimen and a plaster cast (CADIC PI 52).
Description. Hypichnial, helicoidal string twisted along a horizontal axis. The trace is 60-70 mm long and the helicoidal ring is 1.5 mm in diameter.
Remarks. Helicorhaphe is a relatively rare graphoglyptid reported only from the Eocene of Poland and probably the Miocene of Iran (cf. Uchman, 1998).
Occurrence. Punta Torcida Formation, Early Eocene.
Ichnogenus Helminthorhaphe Seilacher, 1977
Helminthorhaphe isp.
Figure 4.1
Figure 4. 1, Helminthorhaphe isp. Cerro Colorado Formation. Positive hyporelief / hiporelieve positivo. 2, Megagrapton submontanum. Punta Torcida Formation. Positive hyporelief / hiporelieve positivo. 3, Nereites isp. Cabo San Pablo beds. Positive epirelief / epirelieve positivo. 4, Ophiomorpha rudis (Or) and Ophiomorpha annulata (Oa). Cerro Colorado Formation. Dense association at the sole of a sandstone bed / asociación densa en la base de un banco de arenisca. 5, Paleodictyon strozzi. Cerro Colorado Formation. Positive hyporelief / hiporelieve positivo. 6, Paleodictyon maximum. Punta Torcida Formation. Positive hyporelief / Hiporelieve positivo. 7, Paleodictyon majus. Cerro Colorado Formation. Positive hyporelief / hiporelieve positivo. 8, Paleodictyon minimum. Cabo San Pablo beds. Positive hyporelief / hiporelieve positivo. 9, Paradictyodora antarctica. Leticia Formation. Upper bedding plane view of a sandstone bed with two specimens / vista en planta de la parte superior de un banco de arenisca con dos especímenes. 10, Scolicia strozzi. Cabo San Pablo beds. Positive hyporelief / hiporelieve positivo. 11, Scolicia prisca. Cabo San Pablo beds. Negative epirelief / epirelieve negativo. Scale bar: 1cm. Field phothographs / escala: 1 cm. Fotografías de campo.
Material. One specimen (CADIC PI 53).
Description. Horizontal, non-branching, meandering strings 2.2 to 2.8 mm in diameter, with high amplitude of irregular meanders, preserved as convex hyporelief on the soles of sandstone turbidites. Strings are filled with sand grains.
Remarks. Uchman (1995) discussed Helminthoida and Helminthorhaphe (Seilacher, 1977) and included Helminthoida crassa Schafhäutl within Nereites irregularis MacLeay.
He also proposed Helminthorhaphe japonica (Tanaka, 1970) as the type ichnospecies of Helminthorhaphe, and Helminthorhaphe flexuosa exclusively for convex hypichnial forms such as Helminthoida crassa Schafhäutl sensu ![]() (1970, 1977) and Helminthorhaphe crassa (Schafhäutl) sensu Seilacher (1977). The material of Helminthorhaphe isp. from Tierra del Fuego has an irregular meandering pattern similar to that of Helminthorhaphe flexuosa Uchman (1995) and apparently lacks the typical bulging turns of Helminthorhaphe japonica (Tanaka). String diameter is considerably bigger than those of specimens described in the literature (e.g. Uchman, 1995), but size is not a useful ichnotaxobase in this case. Only one incomplete specimen was recorded and does not allow ichnospecific assignment. Helminthorhaphe occurs from Early Cretaceous to Miocene (Uchman, 2003).
(1970, 1977) and Helminthorhaphe crassa (Schafhäutl) sensu Seilacher (1977). The material of Helminthorhaphe isp. from Tierra del Fuego has an irregular meandering pattern similar to that of Helminthorhaphe flexuosa Uchman (1995) and apparently lacks the typical bulging turns of Helminthorhaphe japonica (Tanaka). String diameter is considerably bigger than those of specimens described in the literature (e.g. Uchman, 1995), but size is not a useful ichnotaxobase in this case. Only one incomplete specimen was recorded and does not allow ichnospecific assignment. Helminthorhaphe occurs from Early Cretaceous to Miocene (Uchman, 2003).
Occurrence. Cerro Colorado Formation, CCa member, late Middle Eocene.
Ichnogenus Megagrapton
Megagrapton submontanum (Azpeitia-Moros, 1933)
Figure 4.2
2001. Protopaleodyction isp. Olivero and López C.
Material. Many specimens studied in the field and one specimen cast (CADIC PI 54).
Description. Irregular nets of winding strings with short lateral appendages. Preserved as positive hyporelief. The string is 3-4 mm in diameter.
Remarks. Differences between Protopaleodictyon and Megagrapton were discussed by Uchman (1998). M. submontanum occurs in flysch deposits from Cenomanian to Eocene (Uchman, 1998).
Occurrence. Punta Torcida Formation, Early Eocene, and Cerro Colorado Formation, CCa member, late Middle Eocene.
Ichnogenus Nereites MacLeay, 1839
Nereites isp.
Figure 4.3
Material. Many specimens studied in the field.
Description. Horizontal, meandering burrows consisting of a meniscate backfilled median tunnel surrounded by a poorly developed, commonly not preserved, reworked zone. Meniscate area about 4.2-4.5 mm wide. Meanders irregular, but generally with partial overlapping turns.
Remarks. Nereites isp. is preserved at the top of finegrained, organic rich sandstone beds interbedded with thin mudstone beds (Carmona et al., 2006). Nereites isp. is similar to N. irregularis (Schafhäutl) but the holotype of the latter has more regular, guided meanders. N. missuriensis (Weller, 1899) contrasts by loose, winding meanders with a wide central backfilled area and enveloped zone of similar thickness (cf. Uchman, 1995). Nereites ranges from the late Precambrian to the Miocene (cf. Uchman, 1995). Modern forms, similar to Nereites are also known in the deep-sea bottoms (Wetzel, 2002).
Occurrence. Cerro Colorado Formation, CCc member, late Middle Eocene, and Cabo San Pablo beds, Early Miocene.
Ichnogenus Ophiomorpha Lundgren, 1891
Ophiomorpha annulata (
Figure 4.4
Material. Several field specimens.
Description. Straight to winding, horizontal, cylindrical, lined and branched trace fossil. Y-shaped branching at acute angles and dead ends branches are common. Tubes are 5.5-6.8 mm in diameter. Pellets mostly elongate, transversal to burrow axis and not always well developed.
Remarks. Preserved on top of sandy turbidites and distributed along bedding surfaces rich in plant debris. Uchman (1995, 1998) considered Ophiomorpha annulata and O. rudis as normal components of the Nereites ichnofacies.
Occurrence. Cerro Colorado Formation, CCa and CCb members, late Middle Eocene.
Ophiomorpha rudis (
Figure 4.4
Material. Several burrow segments (CADIC PI 55) and many field specimens.
Description. Horizontal to oblique, cylindrical branched burrows, commonly with chevron-like backfill. Exterior surface partly covered with small pellets and bioglyphs. Single burrow segments with changes in diameter ranging from 5.7 to 20 mm. Some horizontal burrows segments as long as 200 mm. When present, branching is Y-shaped, with marked swellings at the point of bifurcation. Overcrossing and anastomosing are common.
Remarks. Preserved as full relief or as positive hyporelief at the base of fine-grained sandstone beds (López Cabrera et al., 2006). Uchman (2004) typifies the Ophiomorpha rudis ichnosubfacies of the Nereites ichnofacies for medium- and thick-bedded flysch deposits, mainly of channels and proximal lobes.
Occurrence. Punta Torcida Formation, Early Eocene; Cerro Colorado Formation, CCa and CCb members, late Middle Eocene; and Desdémona Formation and Cabo Ladrillero beds, Early Miocene.
Ichnogenus Paleodictyon Meneghini in Savi and Meneghini, 1850
Remarks. Uchman (1995) reviewed Paleodictyon and his morphometric classification is followed here. Seilacher (1974) proposed the Paleodictyon ichnosubfacies within the deep-sea Nereites ichnofacies. Paleodictyon was recorded from Cambrian to recent. The latter record includes comparable traces recorded by several authors in the modern deep sea (Ekdale, 1980; Miller, 1991; Rona et al., 2003). Seilacher (1977) and Roder (1971) interpreted Paleodictyon as a burrow system made to cultivate or trap microorganisms. Garlick and Miller (1993) suggest a small elongate crustacean as the tracemaker.
Paleodictyon strozzi Meneghini in Savi and Meneghini, 1850
Figure 4.5
Material. One specimen studied in the field.
Description. Small hexagonal net 0.81-0.88 mm in string diameter, mesh-size 4.9-4.8 mm; preserved as positive hyporelief.
Occurrence. Cerro Colorado Formation, CCa member, late Middle Eocene.
Paleodictyon maximum (Eichwald, 1868)
Figure 4.6
Description. Hypichnial horizontal net of medium size. String diameter 2.8 mm and total mesh size 11.8 mm.
Occurrence. Cerro Colorado Formation, CCa member late Middle Eocene.
Paleodictyon majus Meneghini in Peruzzi, 1880
Figure 4.7
Material. Several specimens studied in the field.
Description. Horizontal medium size hexagonal net, string diameter ranging from 1.2 to 1.7 mm; meshsize ranging from 6.1 to 8.6 mm. Preserved as positive hyporelief.
Occurrence. Punta Torcida Formation, Early Eocene, Cerro Colorado Formation, CCa, CCc, and CCd members, the latter includes specimens partly covered by thin pyrite lamina, late Middle Eocene and Late Eocene.
Paleodictyon minimum Sacco, 1888
Figure 4.8
Material. Two specimens studied in the field.
Description. Very small, hypichnial horizontal net. String diameter 0.46-0.5 mm and total mesh size 2 mm.
Occurrence. Cabo San Pablo beds, Early Miocene.
Ichnogenus Paradictyodora Olivero, Buatois and Scasso, 2004
Paradictyodora antarctica Olivero, Buatois and Scasso, 2004
Figure 4.9
Material. Two collected specimens (CADIC PI 20) and several field specimens.
Description. Conical, upward enlarging vertical spreite structure with subvertical folded laminae produced by the migration of a J-shaped tube. Full relief preservation.
Remarks. The upward opening conical shape corresponds to one of the four morphotypes of Paradictyodora antarctica (Olivero et al., 2004). Paradictyodora has been recorded from the Upper Cretaceous of Antarctica and the Eocene to Miocene of Tierra del Fuego.
Occurrence. Punta Torcida Formation, Early Eocene; Leticia and Cerro Colorado Formations, late Middle Eocene; Estancia María Cristina beds, Oligocene; and Cabo Ladrillero and Cabo San Pablo beds, Early Miocene.
Ichnogenus Scolicia de Quatrefages, 1849
Scolicia strozzi (Savi and Meneghini, 1850)
Figure 4.10
Material. Several specimens studied in the field and one collected slab (CADIC PI 56) with two specimens.
Description. Hypichnial, meandering to straight bilobate ridge separated by a median furrow, about 13 mm wide. Ridge width ranging from 2 to 8 mm.
Remarks. S. strozzi represents washed out and casts of Scolicia burrows. Straight to gently winding specimens are assigned to S. strozzi var. vagans (![]() , 1977) and may correspond to locomotion activity of echinoids (Uchman, 1995).
, 1977) and may correspond to locomotion activity of echinoids (Uchman, 1995).
Occurrence. Cabo Ladrillero and Cabo San Pablo beds, Early Miocene.
Scolicia prisca de Quatrefages, 1849
Figures 4.11
Material. Abundant specimens studied in the field. One collected slab (CADIC PI 57) with three specimens.
Description. Epichnial trilobate furrows, 40.3-50.2 mm wide, with concave, semicircular bottom and oblique slopes. Two parallel grooves occur along the edges of the bottom. Preserved at the top of sandstone beds with linguoid ripples, interbedded with thin mudstone laminae.
Remarks. Uchman (1995) reviewed this ichnogenus and included Taphrhelminthopsis, Laminites, Subphyllochorda and Taphrhelminthoida as toponomic variants of Scolicia. The producers are irregular echinoids of the Spatangus group, with two drainage tufts that produce the drain channels (cf. Smith and Crimes, 1983; Uchman, 1995).
Occurrence. Cabo San Pablo beds, Early Miocene.
Scolicia isp.
Material. Several specimens studied in the field.
Description. Horizontal, cylindrical, straight to sinuous trace fossil with backfill menisci. Full relief preservation.
Remarks. Incomplete preservation of these specimens precludes ichnospecific assignment.
Occurrence. Cerro Colorado Formation, CCd member, Late Eocene and Cabo Ladrillero beds, Early Miocene.
Ichnogenus Stelloglyphus Vialov, 1964
Stelloglyphus isp.
Figure 5.1
Figure 5. 1, Stelloglyphus isp. Cabo San Pablo beds. Incomplete specimen. Note lobes with well-defined meniscus in the filling of the rosette structure / espécimen incompleto. Note el relleno meniscado de los lóbulos en la estructura en roseta. 2, Urohelminthoida appendiculata. Cerro Colorado Formation. Positive hyporelief / hiporelieve positivo. 3-4, Zoophycos ispp. 3, Zoophycos isp. Form B, Cerrro Colorado Formation. Note radial ribs across planar spreite / note las costillas radiales perpendiculares al spreite planar. 4, Zoophycos isp Form C, Desdémona Formation. Note minor lamellae separated by mayor lamellae. Endichnial preservation / note las láminas menores limitadas por láminas radiales mayores. Preservación endicnia. Scale in centimeters. Field phothographs / escala en centímetros. Fotografías de campo.
Material. Many specimens studied in the field.
Description. Rosette structure composed of several crowded lobes radiating at different levels from a central point. Lobes overlap and are generally curved at distal points. Internal fill with a well-defined meniscus-like structure. Rosette c. 92.5 mm in diameter. Endichnial preservation.
Remarks. Burrow morphology shows close similarity to Gyrophyllites Glocker but internal fill of Gyrophyllites lacks menisci (Uchman pers. comm.). Branched radial lobes, as diagnostic of Cladichnus fischeri (Heer, 1877), are absent in Stelloglyphus isp.
Occurrence. Cabo San Pablo beds, Early Miocene.
Ichnogenus Urohelminthoida Sacco, 1888
Urohelminthoida appendiculata Heer, 1877
Figure 5.2
Material. One collected specimen (CADIC PI 58) and several specimens studied in the field.
Description. Very small trace fossil consisting of hypichnial strings with tight meanders and lateral appendages protruding from the apical part of the meanders. Strings are 1.3 mm in diameter.
Remarks. Among graphoglyptids, Urohelminthoida was placed into the group of uniramous meanders, with a diagnostic lateral branch on the apex of each undulation (Seilacher, 1977). U. dertonensis Sacco, 1888 displays lateral appendages which pass straight into one arm of the meander but forms an angle with the second arm (Uchman, 1995). Urohelminthoida is recorded from the Jurassic to the Miocene (cf. Uchman, 1995). Modern traces similar to Urohelminthoida were observed in the deep-sea floor (Gaillard, 1991).
Occurrence. Cerro Colorado Formation, CCd member, Late Eocene.
Ichnogenus Zoophycos Massalongo, 1855
Zoophycos ispp.
Figures 5.3-4
Material. Many field specimens.
Description. The material includes three main forms. Form A, includes relatively simple, circular spreite structure. Form B (figure 5.3) consists of large planar spreite trace fossil with well developed radial ribs separating bundles of minor, sigmoidal or U-shaped spreite lamellae. Form C (figure 5.4), consists of large, planar, lobate spreite trace fossil with well developed bundles of minor sigmoidal lamellae limited by a major, radial, sub-parallel lamella. Spreite filled with large, ovoid to elongate pellets.
Remarks. Form B, characterized by radial ribs, is similar to the Echinospira group (cf. Uchman, 1995) and Form C, without radial ribs, to Zoophycos rhodensis Bromley and Hanken, 2003.
Occurrence. Form A was recorded from the Punta Torcida Formation, Early Eocene; Form B in the Cerro Colorado Formation, late Middle to Late Eocene and Cabo San Pablo beds, Early Miocene; and Form C in the Estancia María Cristina beds, Oligocene and Desdémona Formation, Early Miocene.
Discussion and conclusions
Two distinctive trace fossil assemblages are recognized in the studied Cenozoic deposits from Tierra del Fuego (figure 6). One is characterized by ichnogenera commonly found in the Cruziana ichnofacies and is restricted to the marginal marine and shelf deposits of the upper Middle Eocene Leticia Formation. The other one is characterized by ichnogenera common to the Zoophycos and Nereites ichnofacies, and is found in turbidite systems of the Eocene, Oligocene, and Miocene. Their paleoenvironmental significance and implication for the diversity trends of Cenozoic deep-sea trace fossils are briefly discussed below.
Figure 6. Eocene-Miocene stratigraphy, distribution of ichnogenera grouped by ethological categories, and total ichnogeneric diversity / estratigrafía del Eoceno-Mioceno, distribución de icnogéneros agrupados por categorías etológicas y diversidad icnogenérica total.
Trace fossils of the Leticia Formation
The following ichnogenera are restricted to the Leticia Formation: Asterosoma, Rhizocorallium, Curvolithus, Gyrochorte, Palaeophycus, Patagonichnus, Rosselia, Schaubcylindrichnus, Taenidium and Teichichnus (figure 6). They are not distributed uniformly and three distinctive packages located at the basal, middle, and upper parts of the formation are recognized. The lower and upper parts of the Leticia Formation are dominated by Curvolithus, Diplocraterion, Gyrochorte, Rosselia, Patagonichnus and Tasselia. The middle part is dominated by Asterosoma, Palaeophycus, Paradictyodora, Planolites, Rhizocorallium, Schaubcylindrichnus, Taenidium and Teichichnus. The ichnofabric features are also distinctive: whereas the lower and upper parts of the Leticia Formation includes a few centimeter thick bioturbated horizons, the middle part of the Formation includes meter thick, often several tens of meters thick, totally bioturbated packages. These two contrasting bioturbation patterns and ichnoassemblages of the Leticia Formation were interpreted as a symmetric, transgressive-regressive cycle, with the lower and upper parts representing shallow marine, and the middle part representing shelf settings (Olivero and López C., 2001).
Trace fossils from the turbidite systems
The trace fossil associations from the Early Eocene, Mid-Late Eocene, Oligocene and Miocene turbidite systems differ markedly from those of the Leticia Formation. The main characteristic components are Scolicia, Chondrites, Gyrophyllites, Nereites, Phycodes, Phycosiphon, Phymatoderma, Stelloglyphus, Zoophycos, Ophiomorpha rudis, O. annulata and several graphoglyptid ichnogenera (figure 6). These trace fossils, however, are not always found together, instead distinctive ichnoassemblages are recurrent in particular sedimentary facies.
Graphoglyptids are more common and diverse in fine grained, thin-bedded turbidites interbedded with thick mudstone beds, particularly in the basal unnamed strata of the Punta Torcida Formation (Early Eocene) and the CCa, CCc, and CCd members of the Cerro Colorado Formation (late Middle to Late Eocene). Paleodictyon, Helicolithus, Helminthorhaphe, Desmograpton and Megagrapton, particularly the two first ichnogenera, are relatively common, whereas Cosmorhaphe, Helicorhaphe and Urohelminthoida are rare. The thin-bedded turbidites and thick mudstone beds are interpreted as lobe deposits of turbidite systems and the trace fossil assemblage is assigned to the Paleodictyon ichnosubfacies (cf. Seilacher, 1974) within the Nereites ichnofacies. Graphoglyptids are considered diagnostic trace fossils of the Nereites ichnofacies, which is generally interpreted as indicative of deep-sea paleoenvironments, particularly in the Mesozoic and Cenozoic (cf. Seilacher, 1974, 1977; Uchman, 2004). In contrast to this interpretation, the foraminifers of the graphoglyptid-bearing deposits of the CCa and CCc members of the Cerro Colorado Formation suggest shallower, neritic settings (Olivero and Malumián, 1999; Malumián and Olivero, 2006). Some graphoglyptids, particularly Paleodictyon are recorded at the sole of event beds generated in shallower settings (see Fürsich et al., 2007 and references therein). However, the graphoglyptids from Tierra del Fuego are not associated with typical elements of the Cruziana ichnofacies, as those described by Fürsich et al. (2007) for the Jurassic Paleodictyon-bearing storm and prodelta beds from Iran. The sedimentary facies and graphoglytid association from the Cerro Colorado Formation of Tierra del Fuego seems to be partially comparable to the Eocene Kusuri Formation in Turkey, where the occurrence of graphoglyptid elements in neritic deposits was interpreted as originated in a rapidly-shoaling, restricted basin (Uchman et al., 2004). Nevertheless, the analysis of this seemingly abnormal paleobathymetric record for the graphoglyptids of the Cerro Colorado Formation is out the scope of this paper and it requires a specific sedimentologic and paleoecologic study of the graphoglyptid- bearing strata.
Ophiomorpha rudis and O. annulata are associated commonly at the contact between thick-bedded turbidite sandstones and mudstones with high content of plant debris, where they could form dense mazes distributed preferentially along the bedding plane. These Ophiomorpha ichnospecies are relatively common in the sandstone-dominated intervals of the CCa and CCb members, Cerro Colorado Formation. Other thick-bedded sandstones, especially in the Cabo San Pablo beds, are characterized only by Paleodictyon and Scolicia strozzi. The occurrence of these trace fossils, and the sedimentary facies in which they are recorded, are consistent with the establishment of the ichnosubfacies of Ophiomorpha rudis for sand-rich, proximal channel- lobe deposits within the Nereites ichnofacies (cf. Uchman, 2001).
Scolicia prisca and Nereites isp. are commonly associated with thin, fine-grained sandstones with ripple lamination, rhythmically interbedded with thin mudstone beds rich in plant fragments (Carmona et al., 2006). This ichnoassemblage is common in the Cerro Colorado Formation in distal, interbedded sandstone and mudstone, within the Nereites ichnofacies, and in the Cabo San Pablo beds in more proximal fine-grained beds.
Mudstone-dominated deformed beds involved in synsedimentary slumping are generally dominated by Zoophycos isp. This is the dominant trace fossil in the upper part of the unnamed strata of the Punta Torcida Formation and the lower part of the Desdémona Formation, both of which have been interpreted as representing slope deposits (cf. Olivero and López C., 2001; Malumián and Olivero, 2006), and thus they are included within the Zoophycos ichnofacies.
Diversity trends through Eocene to Miocene deep marine trace fossils
Uchman (2003, 2004) made a detailed analysis of Late Mesozoic-Cenozoic global trends in diversity of deep-sea trace fossils, mainly from sedimentary basins around the Tethys. After the Albian he found a rapid and continuous increase in diversity up to an optimum in the Eocene, then a marked drop at the Eocene-Oligocene boundary, and finally a reduced diversity in the Miocene. He also considered both the Eocene peak and the Oligocene drop in diversity as very distinctive.
In the Fuegian Andes, with the exception of the Maastrichtian Policarpo Formation, the Upper Cretaceous to Paleocene successions bear scarce trace fossils of low diversity (Olivero et al., 2002, 2003) and ichnodiversity trends can only be studied for the Eocene-Miocene rocks. Within the turbidite successions (figure 6), the Early Eocene contains 10 ichnogenera (of which four are graphoglyptids); the late Middle Eocene-Upper Eocene 19 (of which seven are graphoglyptids); the Early Oligocene six (of which one is a graphoglyptid); the Oligocene one (without graphoglyptids); the uppermost Oligocene-basal Miocene three (without graphoglyptids); the basal Early Miocene nine (without graphoglyptids); and the late Early Miocene 15 ichnogenera (of which one is a graphoglyptid). Regarding the Eocene-Miocene trend in ichnodiversity, our data are partly in agreement with the findings of Uchman (2003, 2004), and they show a late Middle-Late Eocene optimum, an abrupt drop in the Oligocene, and a reduced ichnodiversity in the Early Miocene.
Uchman (2004) interpreted that these changes were controlled by competition and food supply, major paleoceanographic changes particularly bottomwater temperatures, and general tectonic conditions that favored an increased area of flysch deposits. Increased food supply, particularly phytodetritus, may have been favorable for graphoglyptids, which are mainly interpreted as specialized farmers that used phytodetritus for microbial cultivation. In an early study, Seilacher (1974) already noted that the increased graphoglyptid diversity in the Late Cretaceous might have been related to increased phytodetritus flux to the oceans associated with the terrestrial radiation of the angiosperms. Uchman (2004) related his ichnodiversity peak near the Paleocene-Eocene boundary to the advent of oligotrophic conditions associated to the Cenozoic thermal maximum and global warming of bottom waters. He also assigned the distinct Early Oligocene drop in ichnodiversity to the global crisis related to marked drop in seawater temperatures.
Whereas the ichnodiversity data from the Fuegian Andes seem to support Uchman's explanation for the Oligocene crisis in ichnodiversity, they are in contrast to his explanation for the Eocene optimum in diversity. Regarding the latter point, the distribution of trace fossils throughout the Early Eocene Punta Torcida Formation is not uniform. The lower and mid parts of the Formation are dominated by mudstones deposited in dysoxic to anoxic and warm bottom conditions (Olivero and Malumián, 1999). Trace fossils are very scarce and consist of Planolites, Paradictyodora and Chondrites. Trace fossils, including graphoglyptids, are more abundant in the upper, unnamed strata of the Punta Torcida Formation (figure 6). However, the Eocene peak in ichnodiversity in the Fuegian Andes, including the graphoglyptid peak in diversity, is found in the Cerro Colorado Formation of late Middle to Late Eocene age. This Formation bears different foraminiferal benthic assemblages that reflect cooling waters by replacement of the species known in the Leticia Formation by species of typical genera of temperate-cool water, such as Ammoelphidiella, Antarcticella, and Buccella (Malumián and Olivero, 2006). Thus, our data do not support the idea that increased Eocene ichnodiversity was controlled by global warming of bottom waters during the Cenozoic thermal maximum. On the contrary, ichnodiversity is higher in the upper Middle Eocene-Late Eocene of the Fuegian Andes, accompanying a cooling trend in the water masses (Malumián et al., 2007).
A similar situation of increased diversity associated with a cooling trend of bottom waters is also found in the Maastrichtian of the Policarpo Formation, just above the inoceramid extinction level in the Fuegian Andes (Olivero et al., 2003). The inoceramids were apparently adapted to warm and poorly oxygenated waters and their global extinction pulses, during the mid-Maastrichtian, were related to cooling and enhanced bottom ventilation, promoted by circulation of deep Antarctic waters. The strong contrast in the intensity of bioturbation between the upper Campanian-lower Maastrichtian Bahía Thetis Formation, almost devoid of trace fossils, and the highly bioturbated Maastrichtian-Danian Policarpo Formation reflects a change from dysoxic-anoxic to well ventilated bottom conditions, probably associated with the postulated paleoceanographic changes in the austral deep oceans (Olivero et al., 2003).
Thus, in the Fuegian Andes the peak in ichnodiversity is not coincident with the Cenozoic thermal maximum and global warming of bottom waters. The relative lowered diversity of the Late Paleocene- Early Eocene in the Fuegian Andes is probably a local signal related to the generalized anoxic-dysoxic bottom conditions. Increased ichnodiversity is achieved during the late Middle Eocene to Late Eocene and the diversity maximum is accompanied by a cooling trend in the water masses as indicated by the occurrence of cold water foraminifers (Malumián et al., 2007). Consequently, this increased diversity challenges the concept that maximum diversity is related to warm bottom conditions. The producers of the deep-sea trace fossils from the Zoophycos and Nereites ichnofacies, particularly the graphoglyptids, were apparently highly specialized for food competition in oceanic areas characterized by relatively low food supply (Seilacher, 1974; Uchman, 2004). Contrary to the temperature trend that is highly variable during the Eocene, the δ13C trend, a proxy for productivity, is remarkable constant except for the extreme negative excursion at the Paleocene-Eocene boundary (cf. Luterbacher et al., 2004, and references cited therein). These trends suggest that the Eocene diversity optimum could be related to constancy of relative oligotrophy during the Eocene, a situation that favored the radiation of the graphoglyptid tracemakers.
Acknowledgments
We thank A. Uchman (Jagiellonian University, Poland) for helpful comments and constant support. D. Martinioni (CADIC) helps with field and laboratory work. Thin sections were made by A. Sobral (CADIC). Part of this research was funded by PIP 5100- CONICET. E.B.O. thanks DAAD (Germany) and CONICET (Argentina) for funding a Study Visit to Germany in 2005 that permitted the study together with the senior author of the large Cenozoic collection of flysch trace fossils deposited at the Geologische und Paläontologische Institut, Universität Tübingen. The help and advice of Dolf and Edith Seilacher during the stay at Tübingen is greatly appreciated. Constructive comments by R.A. Scasso and an anonymous reviewer help to improve this work.
References
1. Azpeitia-Moros, F. 1933. Datos para el estudio paleontológico de la costa Cantábrica y de algunos otros puntos de España. Boletín del Instituto Geológico y Minero de España 53: 1-65. [ Links ]
2. Bromley, R.G. and Hanken, N.M. 2003. Structure and function of large, lobed Zoophycos, Pliocene of Rhodes, Greece. Palaeogeography, Palaeoclimatology, Palaeoecology 192: 79-100. [ Links ]
3. Buatois, L.A., Mángano, M.G., Mikulá, R. and Maples, C.H.G. 1998. The Ichnogenus Curvolithus revisited. Journal of Paleontology 73: 758-769. [ Links ]
4. Carmona, N.B., Ponce, J.J., López Cabrera, M.I. and Olivero, E.B. 2006. Distribución y diversidad de trazas fósiles en hiperpicnitas: implicancias etológicas y comparación con patrones de trazas fósiles en turbiditas clásicas. 9° Congreso Argentino de Paleontología y Bioestratigrafía (Córdoba), Resúmenes: 279 pp. [ Links ]
5. Eichwald, E. 1868. Lethaea rossica ou paléontologie de la Russie. Décrite et figurée, Atlas. E. Schweizerbart, Stuttgart, 1657 pp. [ Links ]
6. Ekdale, A.A. 1980. Graphoglyptid burrows in modern deep-sea sediments. Science 207: 304-306. [ Links ]
7. Fischer-Ooster, C. 1858. Die fossilien Fucoiden der Schweizer Alpen, nebst Erörterungen über deren geologisches Alter. Huber, Bern: 72pp. [ Links ]
8. Fritsch, F.T. 1908. Problematica Silurica. In: J. Barrande (ed.), Systême Silurien du Centre de la Bohême, A.S. Oudin, Prague, pp. 1-28. [ Links ]
9. Fu, S. 1991. Funktion, Verhalten und Einteilung fucoider und lophocteniider Lebensspuren. Courier Forchungsinstitut Senckenberg 135: 1-79. [ Links ]
10. Fuchs, T. 1895. Studien über Fucoiden und Hieroglyphen. Denkschriften der Akademie der Wissenschaften zu Wien, mathematisch- naturwissenschaftliche 62: 369-448. [ Links ]
11. Fürsich, F.T., Taheri, J. and Wilmsen, M. 2007. New occurrences of the trace fossil Paleodictyon in shallow marine environments: examples from the Triassic-Jurassic of Iran. Palaios 22: 408-416. [ Links ]
12. Gaillard, C. 1991. Recent organism traces and ichnofacies on the deep-sea floor off New Caledonia, southwestern Pacific. Palaios 6: 302-315. [ Links ]
13. Garlick, G.D. and Miller, W.C. III. 1993. Simulations of burrowing strategies and construction of Paleodictyon. Journal of Geological Education 41: 159-163. [ Links ]
14. Gibert, J.M. de. 2003. Criterios icnológicos para reconocer comportamientos homólogos y homoplásicos en el registro fósil. In: L.A. Buatois and M.G. Mángano (eds.), Icnología: Hacia una convergencia entre Geología y Biología. Publicación Especial de la Asociación Paleontológica Argentina 9: 9-16. [ Links ]
15. Gibert, J.M. de and Martinell, J. 1994. Proximal-distal variations of trace fossil assemblages in a Pliocene Ria, Baix Llobregat, Northeastern Spain. Revista de la Societat Geológica de España 12: 209-214. [ Links ]
16. Glocker, F.E. 1841. Über die Kalkführende Sandsteinformation auf beiden Seiten der mittleren March, in der Gegend zwischen Kwassnitz und Kremsier. Nova Acta Acad. Caes. Leop. Carol. Germ. Nat. Cur. 19: 309-334. [ Links ]
17. Heer, O. 1865. Die Urwelt der Schweiz. Friedrich Schultze, Zurich, 662 pp. [ Links ]
18. Heer, O. 1877. Flora fossilis Helvetiae. Die vorweltliche Flora des Schweiz. J. Wurster & Co. Zurich, 182 pp. [ Links ]
19. Ksiazkiewicz, M. 1968. On some problematic organic traces from the Flysch of the Polish Carpathians. Part 3 (in Polish). Rocznik Polskiego Towarzystwa Geologicznego 38: 3-17. [ Links ]
20. Ksiazkiewicz, M. 1970. Observations on the ichnofauna of the Polish Carpatians. In: T.P. Crimes and J.C. Harper (eds.), Trace Fossils. Geological Journal Special Issue 3: 283-322. [ Links ]
21. Ksiazkiewicz, M. 1977. Trace fossils in the Flysch of the Polish Carpatians. Palaeontologica Polonica 36: 1-208. [ Links ]
22. López Cabrera, M.I., Olivero, E.B., Carmona, N.B. and Ponce, J.J. 2006. La icnosubfacies de Ophiomorpha rudis en el Eoceno de Tierra del Fuego. 9° Congreso Argentino de Paleontología y Bioestratigrafía (Córdoba), Resúmenes: 288. [ Links ]
23. Lundgren, B. 1891. Studier ofver fossilforande lösa block. Geologiska Föreningen i Stockholm Förhandlinger 13: 111-121. [ Links ]
24. Luterbacher, H.P., Ali, J.R., Brinkhuis, H., Gradstein, F.M. Hooker, J.J., Monechi, S, Ogg, J.G., Powell, J., Röhl, U., Sanfilippo, A. and Schmitz, B. 2004. The Paleogene Period. In: F.M. Gradstein, J.G. Ogg, and A.G. Smith (eds.), A Geological Time Scale 2004, Cambridge University Press: 384-408. [ Links ]
25. MacLeay, W.S. 1839. Note in the Annelida. In: R.I. Murchinson: The Silurian System. Part II. Organic Remains: 699-701. [ Links ]
26. Macsotay, O. 1967. Huellas problemáticas y su valor paleoecológico en Venezuela. Géos 16: 7. [ Links ]
27. Malumián, N. and Olivero, E.B. 2005. El Oligoceno-Plioceno marino del río Irigoyen, costa atlántica de Tierra del Fuego, Argentina: Una conexión atlántica-pacífica. Revista Geológica de Chile 32: 117-129. [ Links ]
28. Malumián, N. and Olivero, E.B. 2006. El Grupo Cabo Domingo, Tierra del Fuego: Bioestratigrafía, paleoambientes y acontecimientos del Eoceno-Mioceno marino. Revista de la Asociación Geológica Argentina 61: 139-160. [ Links ]
29. Malumián, N., López C., M.I., Nañez, C. and Olivero, E.B. 2007. Bioerosion patterns in Cretaceous-Cenozoic benthic foraminiferal tests from Patagonia and Tierra del Fuego Island, Argentina. In: R.G. Bromley, L.A. Buatois, M.G. Mángano, J.F. Genise and R.N. Melchor (eds.), Organism-sediment interactions: A multifaceted ichnology. SEPM Special Publication 88, 301-308. [ Links ]
30. Mángano, M.G., Buatois, L.A. and Muñiz Guinea, F. 2005. Ichnology of the Alfarcito Member (Santa Rosita Formation) of northwestern Argentina: animal-substrate interactions in a lower Paleozoic wave dominted shallow sea. Ameghiniana 42: 641-668. [ Links ]
31. McCann, T. 1989. The ichnogenus Desmograpton from the Silurian of Wales -first record from the Paleozoic. Journal of Paleontology 63: 950-953. [ Links ]
32. Massalongo, A. 1855. Monografía delle nereidi fossili del M. Bolca. Antonelli, Verona, 35 pp. [ Links ]
33. Miller, W.III. 1991. Paleoecology of graphoglyptids. Ichnos 1: 305- 312. [ Links ]
34. Monaco, P., Giannetti, A., Caracuel, J.E. and Yébenes, A. 2005. Lower Cretaceous (Albian) shell-armoured and associated echinoid trace fossils from the Sácaras Formation, Serra Gelada area, southeast Spain. Lethaia 38: 333-344. [ Links ]
35. Narbonne, G.M., Myrow, P., Landing, E. and Anderson, M.M. 1987. A candidate stratotype for the Precambrian-Cambrian boundary. Canadian Journal of Earth Sciences 24: 1277-1293. [ Links ]
36. Olivero, E.B. and Malumián N. 1999. Eocene Stratigraphy of Southeastern Tierra del Fuego, Argentina. Bulletin of the American Association of Petroleum Geologists 83: 295-313. [ Links ]
37. Olivero, E.B. and Malumián, N. 2002. Upper Cretaceous-Cenozoic clastic wedges from the Austral-Malvinas foreland basins, Tierra del Fuego, Argentina: Eustatic and tectonic controls. 3º European Meeting on the Palaeontology and Stratigraphy of Latin America (Toulouse, Francia), Addendum: 6-9. [ Links ]
38. Olivero, E.B. and López C., M.I. 2001. Ichnology of syntectonic turbidites, estuarine and shelf deposits, Eocene, Punta Torcida anticline, Isla Grande de Tierra del Fuego. 4° Reunión Argentina de Icnología y 2° Reunión de Icnología del Mercosur (S. M. Tucumán), Resúmenes: 62. [ Links ]
39. Olivero, E.B., Malumián, N. Palamarczuk, S. and Scasso, R. 2002. El Cretácico superior-Paleogeno del área del Río Bueno, costa atlántica de la Isla Grande de Tierra del Fuego. Revista de la Asociación Geológica Argentina 57: 199-218. [ Links ]
40. Olivero, E.B., Malumián, N. and Palamarczuk, S. 2003. Estratigrafía del Cretácico superior- Paleoceno del área de Bahía Thetis, Andes Fueguinos, Argentina: Acontecimientos tectónicos y paleobiológicos. Revista Geológica de Chile 30: 245- 263. [ Links ]
41. Olivero, E.B., Buatois, L.A. and Scasso, R.A. 2004. Paradictyodora antarctica: A new complex vertical spreite trace fossil from the Upper Cretaceous-Paleogene of Antarctica and Tierra del Fuego, Argentina. Journal of Paleontology 78: 783-789. [ Links ]
42. Olivero, E.B., López Cabrera, M.I., Carmona, N.B. and Ponce, J.J. 2005. Icnofacies de Nereites (Cretácico superior-Neógeno) en Tierra del Fuego: composición e implicancias paleoambientales. Ameghiniana Suplemento Resúmenes 42: 37R. [ Links ]
43. Peruzzi, D.G. 1881. Osservazioni sui generi Paleodictyon e Paleomeandron dei terreni cretacei ed eocenici dell´Appeninno sett. e centrale. Atti della Societá Toscana di Scienze Naturali Residente in Pisa. Memorie 5: 3-8. [ Links ]
44. Ponce, J.J., Olivero, E.B., Martinioni, D.R. and López C., M.I. 2007. Sustained and episodic gravity flow deposits and related bioturbation patterns in Paleogene turbidites (Tierra del Fuego, Argentina). In: R.G. Bromley, L.A. Buatois, M.G. Mángano, J.F. Genise and R.N. Melchor (eds.), Organism-sediment interactions: A multifaceted ichnology. SEPM Special Publication 88, 253- 266. [ Links ]
45. Quatrefages, M.A. de. 1849. Note sur la Scolicia prisca (A. de Q.) annelide fossile de la Craie. Annales des Sciences Naturelles, 3 sér., Zoologie 12: 265-266 [ Links ]
46. Roder, H. 1971. Gangsysteme von Paraonis fulgens Levinsen 1883 (Polychaeta) in ökologischer, ethologischer und aktuopaläontologischer Sicht. Senckenbergiana Maritima 3: 3-51. [ Links ]
47. Rona, P.A., Seilacher, A., Luginsland, H., Seilacher, E., de Vargas, C., Vetriani, C., Bernhard, J M., Sherrell, R M., Grassle, J., Low, S. and Lutz, R.A. 2003. Paleodictyon, a Living Fossil on the Deepsea Floor. EOS, Transactions, American Geophysical Union. 84, Fall Meet.Suppl: F828. [ Links ]
48. Sacco, F. 1888. Note di Paleoicnologia Italiana. Atti della Societá Toscana di Scienze Naturali 31: 151-192. [ Links ]
49. Savi, P. and Meneghini, G.G. 1850. Osservazioni strarigrafische e paleontologische concernati la geologia della Toscana e dei paesi limitrofi. In: R.I. Murchinson. Memoria sulla struttura geologica delle Alpi, degli Apenninni e dei Carpazi. Appendix: 246-528. [ Links ]
50. Schafhäutl, K.E. 1851. Geognostische Untersuchungen des südbayerischen Alpengebirges. Literarisch-artistische Anstalt, München: 208 pp. [ Links ]
51. Seilacher, A. 1974. Flysch trace fossils: Evolution of behavioural diversity in the deep-sea. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte: 233-245. [ Links ]
52. Seilacher, A. 1977. Patterns analysis of Paleodictyon and related trace fossils. In: T.P. Crimes and J.C. Harper (eds.), Trace Fossils 2. Geological Journal Special Issue 9: 289-334. [ Links ]
53. Smith, A.B. and Crimes, T.P. 1983. Trace fossils produced by heart urchins -a study of Scolicia and related traces. Lethaia 16: 79-92. [ Links ]
54. Tanaka, K. 1970. Trace fossils from the Cretaceous flysch of the Ikushumbetesu Area, Hokkaido, Japan. Geological Survey of Japan, Report 236: 1-102. [ Links ]
55. Uchman, A. 1995. Taxonomy and paleoecology of flysch trace fossils: The Marnoso-arenacea Formation and associated facies (Miocene, Northern Apennines, Italy). Beringeria 15: 115p. [ Links ]
56. Uchman, A. 1998. Taxonomy and ethology of flysch trace fossil: A revision of the Marian Ksiazkiewicz collection and studies of complementary material. Annales Societatis Geologorum Poloniae 68: 105-218. [ Links ]
57. Uchman, A. 2001. Eocene flysch trace fossils from the Hecho Group of the Pyrenees, northern Spain. Beringeria 28: 3-41. [ Links ]
58. Uchman, A. 2003. Trends in diversity, frequency and complexity of graphoglyptid trace fossils: Evolutionary and palaeoenvironmental aspects. Palaeogeography, Palaeoclimatology, Palaeoecology 192: 123-142. [ Links ]
59. Uchman, A. 2004. Phanerozoic history of deep-sea trace fossils. In: D. McIlroy (ed.) The Application of Ichnology to Palaeoenvironmental and Stratigraphic Analysis. Geological Society, London, Special Publications 228: 125-139. [ Links ]
60. Uchman, A., Janbu, N.E. and Nemec, W. 2004. Trace fossils in the Cretaceous-Eocene flysch of the Sinop-Boyabat Basin, central Pontides. Turkey. Annales Societatis Geologorum Poloniae 74: 197-235. [ Links ]
61. Vialov, O.S. 1964. Zvezdchatye ieroglify iz Triasa severovostoka Sibiri. Akademiya Nauk SSSR, Sibirskoe Otdielenie, Institut Geologii I Geofizki 5: 112-115. [ Links ]
62. Vialov, O.S. 1971. Redkie probliematiki iz Mezozoya Pamira i Kavkaza. Paleontologischeskiy Sbornik 7: 85-93. [ Links ]
63. Weller, S. 1899. Kinderbook faunal studies. I. The Fauna of the vermicular sandstone at Northeview, Webster country, Missouri. Transactions of the Academy of Science St. Louis 9: 9-51. [ Links ]
64. Wetzel, A. 2002. Modern Nereites in the South China Sea-Ecological Association with redox conditions in the sediment. Palaios 17: 507-515. [ Links ]
Recibido: 10 de mayo de 2007.
Aceptado: 7 de marzo de 2008.

