










Serviços Personalizados
Journal

Artigo
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAmeghiniana
versão On-line ISSN 1851-8044
Ameghiniana v.45 n.2 Buenos Aires abr./jun. 2008
Asociaciones de foraminíferos bentónicos recientes de la Plataforma Continental Argentina (39°- 43° S y 58°- 65° O) analizadas con métodos estadísticos para datos composicionales
Marta Alperin1, Emiliana Bernasconi2 y Gabriela Catalina Cusminsky2
1Cátedra de Estadística. Facultad de Ciencias Naturales y Museo. UNLP. Edificio de Institutos y Laboratorios Calle 64 y 120, 1900 La Plata, Buenos Aires, Argentina. alperin@fcnym.unlp.edu.ar
2Centro Regional Universitario Bariloche. Universidad Nacional del Comahue. Quintral 1250. 8400 Bariloche / Consejo Nacional de Investigaciones Científicas y Técnicas. Argentina. bernasconie@crub.uncoma.edu.ar; gcusmins@crub.uncoma.edu.ar
Resumen. Se presenta el análisis de foraminíferos bentónicos en sedimentos superficiales de un sector de la Plataforma Continental Argentina y el golfo San Matías. El estudio cuantitativo se efectuó empleando métodos de análisis estadístico de datos composicionales. Mediante el Análisis de Componentes Principales (ACP), Análisis de Agrupamiento y Sistema de Información Geográfica (SIG) se pudo inferir: a) la presencia de seis grupos de especies cuyas características responderían a las condiciones oceanográficas del lugar, cuatro de ellos indicarían el predominio del sistema de agua costera y los dos restantes, la influencia de la corriente de Malvinas; b) la vinculación entre las variaciones conjuntas que experimenta la composición de 13 especies con la temperatura del agua, las características del fondo y las condiciones energéticas en el área y c) la correlación existente entre las relaciones Cibicides dispars/Bulimina patagonica y Cibicides mckannai/Bulimina patagonica y la temperatura del agua.
Abstract. Recent benthic foraminifera of the Argentinian Continental Shelf (39°- 43° S and 58°- 65° W) analysed with statistical methods for compositional data. The analysis of recent benthic foraminifera from the continental shelf and San Matías Gulf is presented. Quantitative analysis was performed using methods from compositional data analysis theory. Principal Components Analysis, Cluster analysis and Geographic Information System (GIS) were used to infer a) six groups of species that respond to oceanographic conditions, four of which were related to coastal waters and the others could be influenced by the Malvinas current system; b) the relationship between 13 species and the water temperature, bottom sediments, and energetic conditions of the area c) a correlation between Cibicides dispars/Bulimina patagonica and Cibicides mckannai/Bulimina patagonica and water temperature.
Palabras clave. Análisis composicional; Foraminíferos bentónicos; Plataforma continental argentina.
Key words. Compositional Data Analysis; Benthic foraminifers; Argentinean continental shelf.
Introducción
Los foraminíferos constituyen uno de los grupos de organismos más ampliamente utilizado para el análisis de ambientes marinos y la reconstrucción de paleoambientes costeros y de plataforma. Es bien sabido que la presencia de sus diferentes especies está relacionada, y a la vez controlada, por las variaciones de las propiedades físicas y químicas del agua de mar: temperatura, salinidad, concentración de nutrientes, contenido de oxígeno disuelto, a las que se agregan, entre otras, las variables del sustrato. Las características mencionadas han conducido a que, en muchos trabajos sobre este grupo se aborden, directa o indirectamente, problemas relacionados con la ecología de comunidades. Estudios de este tipo requieren la inferencia de las relaciones dentro de la composición de la comunidad en asociación con las variables del hábitat. Para investigar las grandes matrices de datos formadas con los registros de la abundancia de las especies y la información sobre el hábitat se suelen utilizar técnicas estadísticas como el Análisis Factorial, Análisis Canónico de Correspondencia y el análisis de agrupamiento entre otros. Por otra parte, muchos análisis ecológicos incluyen investigaciones sobre patrones de distribución de especies o de asociación de especies diferentes (Pielou, 1977). Ambos patrones suelen reflejar dependencia espacial (Rossi et al., 1992).
Es práctica general que los datos sean convertidos en proporciones o en porcentajes del total. Así, en cada muestra la suma de todas las especies es un valor fijo de 100%, una composición en el sentido de Aitchison (1986). Esta estandarización, que se utiliza para remover las diferencias en el número total de ejemplares entre las muestras, conlleva la dependencia entre los componentes o partes de la composición impuesta por la restricción de la suma constante del 100%. Tempranamente, Pearson (1897), Chayes (1960, 1971) y posteriormente Aitchison (op. cit.) advierten la imposibilidad de interpretar correctamente las matrices de correlación y de varianza-covarianza entre las partes de la composición debido a que, en datos de este tipo, se presentan falsas correlaciones negativas establecidas justamente por la mencionada restricción. De esta forma, las correlaciones espurias producto de la estandarización, inadvertidamente podrían interpretarse de manera errónea como inherentes a las relaciones ecológicas y/o a los patrones entre muestras si no se analizaran en un marco teórico y metodológico apropiado (Jackson, 1997).
Aitchison (1986) y posteriormente Aitchison (1997, 1999), Martín-Fernández et al. (1998), Aitchison et al. (2000), Aitchison y Greenacre (2002), Pawlowsky- Glahn y Egozcue (2001, 2002) entre otros, formularon una nueva teoría y presentaron metodologías de análisis que contemplan la restricción impuesta por la suma constante. Estos métodos, entre otros, tienen la ventaja de subsanar el problema de las falsas correlaciones. Su propuesta requiere una transformación adecuada de los datos composicionales, situados en el espacio muestral restringido o cerrado, llamado simplex "d" dimensional Sd (donde "d" indica el número de partes de la composición), hacia el espacio sin restricciones de los números reales Rd. Este procedimiento posibilita la aplicación de numerosos métodos estadísticos en los datos que no sufren esta restricción como los que se utilizan para describir variabilidad y estructuras de correlación y clasificación, entre otros.
Los antecedentes de análisis de datos micropaleontológicos en este marco teórico se limitan a pocos trabajos. Kucera y Malmgren (1998) realizaron un estudio comparativo entre datos sin transformar y transformados sobre foraminíferos planctónicos de 36 muestras superficiales de testigos de aguas subtropicales a subpolares del sector austral del océano Índico; Monechi et al. (2000), estudiaron nanoflora del Eoceno Superior de la sección del Massignano para delinear semejanzas y diferencias entre taxones y agrupar muestras con asociaciones similares. Más recientemente Buccianti y Esposito (2004), emplearon estos métodos para investigar los cambios de abundancia de nanoplancton calcáreo en un testigo del Campaniano del golfo de Salerno (Italia).
El registro de los foraminíferos, en el océano Atlántico Sudoccidental, se encuentra bien documentado. En base a la distribución de foraminíferos recientes se han establecido las provincias zoogeográficas y las masas de agua para la región de América del Sur (Boltovskoy, 1979; 1981 y referencias allí citadas; Boltovskoy et al., 1996 y Giussani de Khan y Watanabe, 1980). Además se han publicado trabajos enmarcados en aspectos bioestratigráficos y paleobiogeográficos tales como Bertels (1984); Boltovskoy (1957, 1978); Boltovskoy y Watanabe (1977); Boltovskoy y Giussani de Khan (1982) y Cusminsky (1991, 1994) y cuantitativos (Boltovskoy y Totah, 1985; Bertels, 1995; Laprida, 1998; Caramés y Malumián, 2000; Laprida y Bertels-Psotka, 2003; Bernasconi y Cusminsky, 2005a, 2007). En ambientes engolfados de la Plataforma Continental Argentina los trabajos no son tan numerosos. Boltovskoy (1954a) analiza los foraminíferos del golfo San Jorge mientras que Bernasconi y Cusminsky (2005a) y Bernasconi (2006) investigan los foraminíferos de los golfos San Matías y Nuevo.
En función de lo expuesto, el estudio de las relaciones de abundancia relativa de los foraminíferos bentónicos dentro del ambiente nerítico resulta relevante. En este trabajo se estudiaron los foraminíferos del sector de la Plataforma Continental Argentina ubicado entre los 39° y 43° Sur y 58° y 65° Oeste así como las del golfo San Matías. En particular se investigó la existencia de asociaciones entre especies, subrayando sus aspectos cuali y cuantitativos, todo ello en un contexto espacial y en relación con las características del hábitat. El énfasis estuvo puesto en la perspectiva cuantitativa adoptada, que corresponde a las bases teórico-metodológicas propuestas por Aitchison para el análisis de datos expresados como proporciones o porcentajes.
Área de estudio: características geomorfológicas, sedimentológicas y oceanográficas
El área estudiada, ubicada entre los paralelos 39° y 43° de latitud Sur y los meridianos 58° y 65° de longitud Oeste, abarca un sector de la Plataforma Continental Argentina propiamente dicha y el golfo San Matías (figura 1.1).
Figura 1. 1, Mapa de localización / location map. 2, Posición geográfica de las estaciones / geographic position of stations.
El sector de la plataforma continental corresponde a un tramo submarino de la cuenca del Colorado que tiene conexión directa con el océano Atlántico Sudoccidental. El relieve submarino presenta pendientes en dirección noroeste-sudeste, suaves hasta profundidades de algo más de 70 m, desde allí hasta aproximadamente la isobata de 140 m el relieve se torna ligeramente ondulado y a profundidades mayores las irregularidades son más marcadas (Cortelezzi y Mouzo, 1979; Gelós et al., 1988) (figura 1.2). Los sedimentos de la plataforma son preponderantemente arenosos y composicionalmente homogéneos, mineralógicamente son inmaduros y texturalmente bien seleccionados y maduros. Su mecanismo deposicional se ha atribuido a los procesos de regresióntransgresión del Pleistoceno-Holoceno, en tanto la actual dinámica distributiva estaría vinculada a las corrientes de plataforma que circulan con sentido general norte y noreste (Gelós et al., op. cit.). Conforme a la nomenclatura batimétrica propuesta por Boltovskoy (1965) en este ambiente se diferencia la plataforma interna, que sigue a continuación de la zona litoral y ocupa una franja hasta las profundidades de 50 a 70 m, y la plataforma externa a la cual corresponden profundidades mayores.
El golfo San Matías, situado en la provincia de Río Negro (40° 30' y 42° 30' S y 63° 30' y 65° O), es una cuenca semicerrada que posee profundidades mayores que la plataforma continental que lo rodea y se comunica con ella a través de un umbral; su porción central supera los 190 m de profundidad (Gelós et al., 1988; Parker et al., 1996). La depresión central del golfo está cubierta por sedimentos arcillosos de coloración verde, composicionalmente homogéneos y mineralógicamente inmaduros, cuyo origen es incierto (Gelós et al., op. cit.).
La hidrología del área presenta considerables variaciones espaciales y temporales. Las masas de agua de la Plataforma Continental son de origen subantártico y la circulación media es de sur a norte. Sin embargo, en el golfo San Matías y en el área costera semi- protegida El Rincón se generan localmente características particulares (Guerrero y Piola, 1997).
Para estas latitudes Martos y Piccolo (1988) diferencian dos dominios: uno costero o interno y otro externo. El dominio costero se extiende desde la zona litoral hasta la isobata de 40-50 m. Allí, Guerrero y Piola (1997) reconocen un Agua Costera con baja salinidad (S<33,4‰) en la desembocadura del río Negro al sur del Rincón y un Agua Costera con alta salinidad (entre 33,8 y 34,00‰) en la desembocadura del golfo San Matías y en la zona central y sur de la provincia de Buenos Aires. El dominio externo se extiende desde la isobata de 40-50 m hasta los 100 m de profundidad aproximadamente, posición donde se ubica el cambio de pendiente correspondiente al talud. Guerrero y Piola (1997) describen en este sector el Agua de la Plataforma Media (S entre 33,4 a 33,6/33,7‰) que es interpretada como un flujo hacia el norte denominado corriente Patagónica. Las aguas presentan una fuerte estacionalidad, siendo homogéneas en invierno y estratificadas durante la época estival. La estructura de temperatura y densidad observada en la época de primavera-verano muestra una capa superficial y una de fondo, ambas separadas por una pronunciada termoclina y picnoclina entre los 30 y 40 m. Las aguas superficiales se describen fluyendo en dirección suroeste con valores de salinidad y temperatura característicos del Río de la Plata (Tº > 15ºC y S<33‰) (Martos y Piccolo, 1988). Al progresar el otoño e invierno las aguas superficiales se enfrían, la columna de agua se mezcla verticalmente por convección y se percibe una fuerte influencia de la corriente de Malvinas, la temperatura media es de 12ºC, los valores de salinidad son típicos de plataformas continentales. Por último, fuera de la plataforma continental pero con influencia sobre la hidrología, Guerrero y Piola (1997) diferencian el agua de Plataforma Externa o Talud. Esta agua se caracteriza por temperaturas menores a 15ºC con rango de salinidad entre 33,7 y 34,0‰ se ubicarían entre el Agua de Plataforma Media y la corriente de Malvinas.
Materiales y métodos
El material utilizado para el presente trabajo proviene de 35 muestras de sedimentos superficiales (en adelante estaciones) obtenidos mediante un muestrador tipo Shipek con el buque Oceanográfico Puerto Deseado en 1984 por el Instituto Argentino de Oceanografía (IADO). Se emplearon también los datos sedimentológicos de las mismas estaciones obtenidos por Gelós et al. (1988) y Gelós y Spagnuolo (1992) y los parámetros fisicoquímicos del agua de fondo registrados durante la campaña oceanográfica por Pucci et al. (1985) (tabla 1).
Tabla 1. Ubicación geográfica; características físicas del agua, del sustrato de las estaciones estudiadas; número de especies y número de ejemplares. *: estaciones sin foraminíferos / geographic locations; physical characteristics of the water, the subtrate of each studied stations; species and specimens numbers. *: stations without foraminifers.

Procesamiento de las muestras, determinación y recuento de foraminíferos
Los sedimentos fueron lavados con agua y se retuvo el material de tamaño mayor a 63 mm utilizando un tamiz n° 230 del Tyler Screen System. Se revisaron 5g de sedimento seco y se extrajo la totalidad de los ejemplares presentes bajo lupa binocular. La determinación sistemática de los individuos a nivel genérico se basó en Loeblich y Tappan (1988) mientras que a nivel específico se usaron los trabajos de Boltovskoy (1954a, 1954b, 1959), Boltovskoy et al. (1980); Cusminsky (1992); Laprida (1998); Laprida y Bertels-Psotka (2003), entre otros.
Los foraminíferos bentónicos son abundantes en casi todas las estaciones estudiadas y el estado de preservación de los ejemplares es en general bueno. Se identificaron y contaron 4287 especímenes pertenecientes a 100 especies diferentes en los sedimentos de las 35 estaciones. Cabe destacar que la aparición de muchas de estas especies consiste apenas de unos pocos ejemplares. Más del 75% de las especies estuvieron representadas por 4 ejemplares o menos en el total de foraminíferos identificados, por lo que podrían considerarse especies raras. Siguiendo a Clifford y Stephenson (1975) y a Boesch (1977) se consideró que la probabilidad de ocurrencia de una especie rara en una muestra de sedimentos es muy pequeña y por ende, las co-ocurrencias de éstas carecen de patrones, por lo que no sería posible relacionarlas a requerimientos de hábitat debido a que se producirían al azar. En las estaciones 29, 30, 40 y 43 se encontraron sólo 2 a 4 ejemplares; en la estación 46 los 12 especimenes identificados pertenecen a las especies consideradas raras por lo que el material de estas cinco estaciones fue descartado del presente análisis.
Las razones expuestas justifican que, antes de emprender el análisis estadístico, se hayan eliminado las especies que aparecen en menos del 1% del total de especimenes contado en la totalidad de las muestras, centrando este estudio en las variaciones de las especies más abundantes. De este modo, se redujo la matriz original de datos y el análisis composicional se circunscribió a las asociaciones que forman las siguientes especies: Ammonia beccarii (Linné, 1758); Buccella peruviana f. campsi (Boltovskoy, 1954); Bulimina marginata d´ Orbigny, 1826; Bulimina patagonica d´ Orbigny, 1839; Cibicides dispars (d´ Orbigny, 1839); Cibicides fletcheri Galloway y Wissler, 1927; Cibicides mckannai Galloway y Wissler, 1927; Epistominella exigua (Brady, 1884); Globobulimina affinis (d´ Orbigny, 1839); Globocassidulina subglobosa (Brady, 1881); Nonionella auris (d´ Orbigny, 1839); Quinqueloculina seminula Linné, 1758 y Uvigerina peregrina Cushman, 1923.
Análisis estadísticos multivariantes
Siguiendo las metodologías estándares para análisis micropaleontológicos, se construyó una matriz de datos (W) recalculando los porcentajes de las 13 especies que reunieron el criterio fijado de abundancia y permanencia para cada estación de muestreo, es decir se expresan como una composición de 13 partes (d=13) (tabla 2). El análisis cuantitativo de las asociaciones de foraminíferos bentónicos se realizó mediante análisis estadístico de datos composicionales, lo que implica interesarse por las relaciones relativas entre las especies más que en las diferencias absolutas o de cantidad.
Tabla 2. Abundancias relativas (%) de las 13 especies de foraminíferos consideradas; número de especimenes; riqueza (RS): cantidad de especies presentes en cada estación; Grupo del análisis de agrupamiento al que pertenece la estación (Grupo) y estaciones en la que se encontró la especie (NºE) / relative abundance (%) of 13 foraminifers species considerated; specimens numbers; richness (RS); Analysis cluster group in wich the station is founded (Group) and stations where the specie was present (NºE).
Aun después de haber eliminado las especies raras, la inspección de la matriz de datos mostró un remanente importante de valores cero (60% de la matriz). Ninguna especie estuvo presente en todas las estaciones. La especie con mayor cantidad de ceros fue Uvigerina peregrina (presente sólo en 5 estaciones) en tanto que la especie más ampliamente distribuida fue Epistominella exigua (presente en 25). Dado que esta situación no admite ningún análisis composicional, se utilizó la estrategia de reemplazo multiplicativa de ceros (Martín-Fernández, 2001 y Martín- Fernández et al., 2003). Se eligió como valor de reemplazo del cero, la mitad de un ejemplar (0,5% = 0,005); así la composición se recalculó utilizando la siguiente expresión:
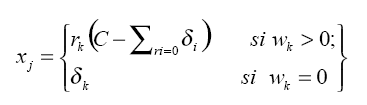
donde xj es el nuevo valor que toma cada especie luego de realizado el reemplazo de los ceros; δk es el valor que se atribuye en la parte original (wk) (δk = 0,005), C es la constante de la composición (C=1); Σri=0 δi es la suma de los valores de reemplazo de cero que se efectuaron en la composición (en la tabla 3 se ejemplifica el procedimiento para la estación 1).
Tabla 3. Ejemplo de la estrategia de reemplazo de "0" en la composición original de la estación 1 utilizando como valor de reemplazo 0=0,005; C-Σk/δk=0=1-(5.0,005)=0,975 / example of the replacement strategy for "0" in the original station 1 composition using 0=0.005 as replacement value y vector composicional de la estación 1 clr-transformada, g(x)=0,0270 / Clr-tranformed vector of station 1, g(x)=0,0270.
Luego, cada fila de la matriz X de proporciones recalculada de especies se transformó utilizando la transformación log-cociente centrada (clr). Para ello se utilizó la media geométrica de cada estación como divisor del log-cociente de la proporción de cada especie. Cada fila de la matriz de proporciones tiene un divisor diferente según:

donde ![]() es la media geométrica de cada estación (tabla 3). El resultado de la transformación de la matriz de datos (X), fue una matriz (Z) con el mismo número de columnas que la matriz original. Los datos clr-transformados mantienen las distancias entre partes, de modo que pueden emplearse cuando se requiere utilizar métodos de análisis multivariante basados en esta propiedad, tales como el análisis de agrupamiento. Por otra parte también permiten resolver problemas de descripción del grado de correlación entre componentes (especies) (Kucera y Malmgren, 1998; Martín-Fernández et al., 1998; Aitchison et al., 2000, entre otros).
es la media geométrica de cada estación (tabla 3). El resultado de la transformación de la matriz de datos (X), fue una matriz (Z) con el mismo número de columnas que la matriz original. Los datos clr-transformados mantienen las distancias entre partes, de modo que pueden emplearse cuando se requiere utilizar métodos de análisis multivariante basados en esta propiedad, tales como el análisis de agrupamiento. Por otra parte también permiten resolver problemas de descripción del grado de correlación entre componentes (especies) (Kucera y Malmgren, 1998; Martín-Fernández et al., 1998; Aitchison et al., 2000, entre otros).
Siguiendo a Aitchison y Greenacre (2002) se realizó un Análisis de Componentes Principales (ACP) de Z utilizando la matriz de covarianza y luego se construyó un biplot con las 2 primeras Componentes Principales (CP) calculadas (figura 2). El biplot es una representación de la matriz de varianza-covarianza entre componentes y en tal sentido puede usarse para describir las relaciones entre especies, entre estaciones y entre ambas. El centro del biplot es el promedio de la composición. La longitud de los rayos, segmentos que unen el centro del biplot con el vértice de cada especie, reproduce el desvío estándar de la especie: rayos más largos indican mayor variabilidad. La distancia entre pares de vértices de especies (enlace) representa la variabilidad de la relación entre ambas ![]() : distancias más largas indican mayor variabilidad. Por otra parte, la distancia entre las estaciones de muestreo es una aproximación de las diferencias que existen entre ellas. La proyección ortogonal de una estación sobre un enlace xi-xj es una representación del valor aproximado del log-cociente entre ese par de especies [ln(xi/xj)] en esa estación (Aitchison y Greenacre, op. cit.).
: distancias más largas indican mayor variabilidad. Por otra parte, la distancia entre las estaciones de muestreo es una aproximación de las diferencias que existen entre ellas. La proyección ortogonal de una estación sobre un enlace xi-xj es una representación del valor aproximado del log-cociente entre ese par de especies [ln(xi/xj)] en esa estación (Aitchison y Greenacre, op. cit.).

Figura 2. Biplot de las dos primeras Componentes Principales (CP) con ubicación de especies y estaciones. Entre paréntesis se indica el % de reconstrucción de cada especie / biplot of species and stations. Numbers indicate percentage of reconstruction of each species. Ab, Ammonia beccarii (57); Bpc, Buccella peruviana f. campsi (72); Bm, Bulimina marginata (45); Bp, Bulimina patagonica (66); Cd, Cibicides dispars (52); Cf, Cibicides fletcheri (4); Cm, Cibicides mckannai (66); Ee, Epistominella exigua (67); Ga, Globobulimina affinis (38); Gs, Globocassidulina subglobosa (44); Na, Nonionella auris (41); Qs, Quinqueloculina seminula (24); Up, Uvigerina peregrina (29). Los números identifican cada estación. Los símbolos de cada estación corresponden a los grupos de estaciones determinados con el análisis de agrupamiento. ![]() Grupo 1;
Grupo 1; ![]() Grupo 2, + Grupo 3, * Grupo 4, ♦ Grupo 5,
Grupo 2, + Grupo 3, * Grupo 4, ♦ Grupo 5, ![]() Grupo 6 / number: labels for each station. Symbol shows the group of stations defined with cluster analysis.
Grupo 6 / number: labels for each station. Symbol shows the group of stations defined with cluster analysis.
Para describir la matriz de datos composicionales se utilizaron las medidas propuestas por Aitchison (1986, 1997): el Centro (gm(X)) calculado como la media geométrica de cada una de las especies a la que se aplicó la operación de clausura (es decir los valores se recalcularon a 100%) y la varianza total (Var total( X)) igual a la traza de la matriz de covarianza del conjunto de datos clr-transformados. Una de las hipótesis a considerar es la existencia de diferentes asociaciones de foraminíferos. El método estadístico usado para validarla fue un análisis de Agrupamiento. Para la formación de los aglomerados jerárquicos se eligió el método de Ward utilizando los datos clrtransformados y el cuadrado de la distancia euclidiana como medida de distancia entre el centro de los grupos (Aitchison, 1992; Martín-Fernández et al., 1998 y Aitchison et al., 2000).
Una vez determinados los grupos de estaciones formados en el análisis de Agrupamiento, se describió la asociación de foraminíferos de cada grupo caracterizándola mediante el Centro y la varianza total. Además se consideraron la riqueza, expresada como el número de especies promedio (RS), y la constancia (Co). Para cada grupo se calculó la constancia de especie iésima con el cociente entre el número de estaciones del grupo donde la especie iésima fue efectivamente registrada y el número total de estaciones del grupo. Un valor Co=1 indica presencia en todas las estaciones del grupo; un valor Co=0 que la especie no está presente en ninguna estación del grupo (Boesch, 1973).
Paralelamente, se desarrolló un Sistema de Información Geográfico (SIG) en el que se volcaron los datos individuales de cada especie, las variables ambientales especializadas y paulatinamente, a medida que se generaron, los resultados parciales de los análisis estadísticos. En este contexto el uso del SIG permitió no sólo mapear las diferentes variables de interés, sino también convertirse en una herramienta para interpretar las relaciones entre especies, entre asociaciones de especies y sus relaciones con las variables ambientales, todos ellos en su contexto espacial.
Los cálculos y gráficos estadísticos se realizan con el software Excel-base CoDaPack (Thio, 2004). Se utilizó Quantum GIS (QGIS) 0.8 TITAN (Sutton et al., 2006).
Resultados
Generales para el área de estudio
El análisis de los datos, previo a la eliminación de las especies raras, indicó que, en promedio, se registraron 10 especies por estación (mínimo 2 en las estaciones 29, 30, 40 y 53 y máximo 36 en la estación 52) identificadas sobre la base de un recuento promedio de 122 ejemplares por estación (mínimo de 2 en las estaciones 29 y 40 y máximo 860 en la estación 50). El cálculo de la correlación entre la cantidad de especies identificadas y el número de ejemplares sugiere que ambas variables no están linealmente correlacionadas (r de Pearson = 0,403; p<0,05). Las estaciones con mayor número de especies se ubican preferentemente en una franja próxima a la costa. Las estaciones con mayor número de ejemplares muestran un patrón más errático, algunas son más litorales, otras más oceánicas (tabla 1).
Del análisis composicional efectuado con las 13 especies y las 30 estaciones que reúnen los criterios de constancia y abundancia fijados surge que: la composición promedio (Centro g(X)) es: Epistominella exigua (31,65), Buccella peruviana f. campsi (17,99); Cibicides dispars (9,22); Bulimina patagonica (7,02); Globobulimina affinis (5,20); Cibicides mckannai (5,03); Cibicides fletcheri (4,22); Nonionella auris (4,16); Ammonia beccarii (4,10); Bulimina marginata (3,04); Quinqueloculina seminula (2,98); Uvigerina peregrina (2,74), Globocassidulina subglobosa (2,65). Epistominella exigua es la especie con constancia mayor (Co = 0,8), seguida de Buccella peruviana f. campsi (Co = 0,6), Cibicides dispars y Bulimina patagonica (Co = 0,5) y las demás especies se encuentran con constancia 0,4 o menor (tabla 5).
El ACP realizado y el biplot de las dos primeras componentes (figura 2) recogen sólo el 52% de la variabilidad total para las 2 primeras CP (tabla 4), lo que indica que sólo algunas especies están bien representadas en este plano (figura 2, porcentaje de reconstrucción). Sin embargo, se aprecian tres grupos de especies, el A, integrado por C. dispars-U. peregrina- G. subglobosa-C. mckannai-E. exigua y C. fletcheri en el semieje negativo de la CP1; el B, integrado por A. beccarii-B. peruviana f. campsi-Q. seminula en el semieje positivo de las CP1 y CP2 y el C, B. patagonica-B. marginata-N. auris-G. affinis en el semieje positivo de la CP1 y semieje negativo de la CP2. La especie más variable es B. peruviana f. campsi, seguida de C. dispars, N. auris y E. exigua, y las menos variables son G. subglobosa, B. marginata y U. peregrina (tabla 5). Las relaciones más variables son las que mantienen entre sí y con el resto de las especies B. peruviana f. campsi, N. auris y C. dispars. Las relaciones menos variables son las que mantienen U. peregrina con G. subglobosa. Además existen gradientes en las relaciones C. dispars/ B. patagonica, C. mckannai/B. patagonica y E. exigua/ B. peruviana f. campsi. Al mismo tiempo, se diferencian dos agrupamientos de estaciones, uno en el semieje negativo de la CP1 cercano a las especies del grupo A y el otro en el semieje positivo de la CP1 en torno a las especies de los grupos B y C.
Tabla 4. Vectores propios, valores propios y proporción de la variación explicada por cada componente principal (CP) utilizando la matriz de covarianza de los datos clr-transformados / eigen-vectors, eigen-values, and the proportion of variance explained by each of the principal components (PC) using the clr-transformed data covarience matrix.
Tabla 5. Estadísticos composicionales de los grupos definidos en el dendrograma. Centro: media geométrica clausurada; Var-clr: varianza de los datos clr-transformados y Co: Constancia; Riqueza de especies y Profundidad, Temperatura y Salinidad promedios / compositional descriptive statistics of the cluster of samples defined in the dendrogram. Centro (geometric mean), Var-clr (clr-transformed data Variance) and Co (Constancy); species Richness and Deeph, Temperature and Salinity average.
Distribución espacial de abundancia de las especies
Los análisis cuantitativos revelaron distintas tendencias y fluctuaciones de abundancia de los taxa seleccionadas para el análisis (figura 3.1 y 3.2).
Figura 3.1. Mapas de las variaciones de abundancia relativa de las especies de foraminíferos seleccionadas (1 a 10) / relative abundance fluctuations of the selected foraminifera species maps.
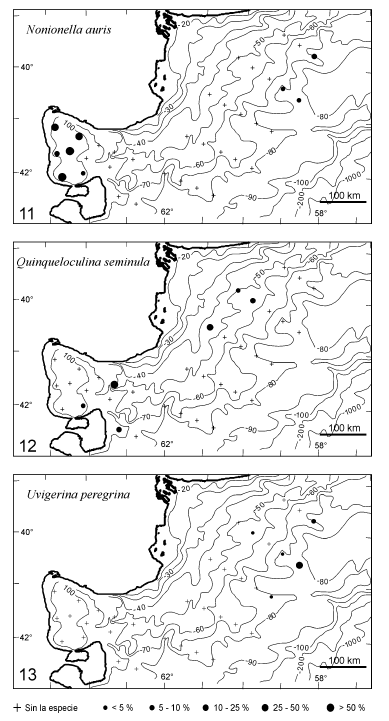
Figura 3.2. Mapas de las variaciones de abundancia relativa de las especies de foraminíferos seleccionadas (11 a 13) / relative abundance fluctuations of the selected foraminifera species maps.
Ammonia beccarii se encuentra circunscripta a 8 estaciones, la mayoría de ellas ubicadas en la boca del golfo San Matías y en el sector más austral del área de estudio a profundidades que oscilan entre los 30 y 90 m. En la boca del golfo constituye entre el 50 y 80% de la asociación presente, y en las restantes estaciones su abundancia no supera el 10% (figura 3.1).
Bucella peruviana f. campsi está presente en 18 estaciones con una vasta distribución latitudinal, se halla prácticamente restringida al sector de plataforma interna y al sector occidental y sur del golfo San Matías, entre las isobatas 40 y 160 m. Compone entre el 20 y el 78% de la asociación de la mayoría de las estaciones en las que aparece (figura 3.1).
Bulimina marginata se localiza en 8 estaciones ubicadas en dos sectores aislados, uno en la zona norte de la plataforma interna entre las isobatas 40 y 60 m y el otro en el sector del golfo entre las isobatas 75 y 160 m, en ambos casos con abundancias que no superan el 8% (figura 3.1).
Bulimina patagonica se registra en 15 estaciones, posee una amplia distribución latitudinal aunque limitada a la zona de plataforma interna, con profundidades entre 40 y 70 m y a la zona del golfo, entre las isobatas 75 y 160 m, su abundancia en ningún caso supera el 22% (figura 3.1).
Cibicides dispars posee una extendida distribución latitudinal presentándose en 15 estaciones. Se observa un gradiente de abundancia creciente en relación con la profundidad que oscila entre 0,1-1% entre los 40 y 50 m hacia valores entre 10-65% entre las isobatas 50 y 80 m. Está prácticamente ausente en el golfo San Matías (figura 3.1).
Cibicides fletcheri se encuentra en 9 estaciones cuya distribución es dispersa. Está presente en algunas estaciones de la plataforma entre los 40 y 80 m de profundidad con proporciones del orden del 5% o menor y con porcentajes entre 10 y 25 en algunas estaciones ubicadas entre las isobatas 50 y 90 m. Está bien representada en una muestra del golfo con casi 40% de abundancias a unos 106 m de profundidad (figura 3.1).
Cibicides mckannai se halla en 10 estaciones. Si bien su distribución latitudinal es amplia, está caracterizada por abundancias menores al 12% en las estaciones cuyas profundidades están entre los 40 y 82 m, y entre 60 y 80% en las ubicadas a profundidades entre 80 y 90 m (figura 3.1).
Epistominella exigua tiene la distribución espacial más amplia puesto que se presenta en 25 estaciones. Se caracteriza por abundancias que varían entre 1,2 y 87%, en las estaciones de plataforma interna y externa, entre las isobatas 40 y 80 m. En la zona del golfo se encuentra entre 90 y 125 m de profundidad con proporciones menores al 9% (figura 3.1).
Globobulimina affinis se reconoce en 12 estaciones. Su distribución es algo dispersa pues se la encuentra en la plataforma interna entre las isobatas de 40 y 70 m y en la zona del golfo a profundidades entre 75 y 120 m. Su abundancia registra valores entre el 1 y 30% (figura 3.1).
Globocassidulina subglobosa se halla en 7 estaciones. Su presencia se restringe fundamentalmente a las estaciones de plataforma externa, entre las isobatas 50 y 90 m, con proporciones que oscilan entre el 1 y 9% (figura 3.1).
Nonionella auris se localiza en 9 estaciones, principalmente en el golfo San Matías entre las isobatas 80 y 160 m, en proporciones que varían entre 35 y 94%. En el sector norte de la plataforma externa, entre 70 y 85 m, se registra en unas pocas estaciones en proporciones que no superan el 5% (figura 3.1).
Quinqueloculina seminula es identificada en sólo 6 estaciones dispersas de la plataforma interna y de la entrada al golfo, a profundidades entre 40 y 106 m, con proporciones entre 1 y 35% (figura 3.2).
Uvigerina peregrina su presencia se restringe a 5 estaciones ubicadas en la parte norte de la zona de plataforma, entre 50 y 90 m, con abundancias menores a 10%, a excepción de una estación a 82 m donde alcanza el 40% de la asociación (figura 3.2).
Asociaciones de especies
En la figura 4 se representó el dendrograma de estaciones obtenido al aplicar el método de Ward empleando el cuadrado de la distancia euclidiana con los datos clr-transformados. Se optó por un nivel de corte de árbol que da lugar a 6 grupos, cada uno con una asociación de foraminíferos que permitió diferenciarlos entre sí. Posteriormente, estos grupos de estaciones se estudiaron respecto a su posición geográfica (figura 5), variables ambientales, riqueza de especies y constancia, aspectos que no se utilizaron para realizar el análisis estadístico. De estos grupos se destacan las siguientes características:
Grupo 1. Las 6 estaciones que pertenecen a este grupo (3, 13, 14, 21, 22 y 39) tienen porcentajes altos de C. dispars y E. exigua, alcanzando valores promedio para el grupo de 42% y 34% respectivamente, con la mayor proporción de C. dispars de toda el área estudiada (figura 6, tabla 5). Estas características también se ponen de manifiesto en el biplot (figura 2), ya que las estaciones se agrupan próximas a los vértices de ambas composiciones de especies y alejadas del centro del diagrama. Por otra parte, la presencia de C. mckannai se destaca por su valor promedio y constancia altos (10,73% y 0,8 respectivamente). Las demás especies presentes poseen todas porcentajes inferiores al 4%, U. peregrina y G. subglobosa con constancias altas (0,7 y 0,8) y N. auris, B. peruviana f. campsi y C. fletcheri con constancias de 0,5 o menores. La varianza total del grupo es 10,3, indicando que es uno de los más homogéneos de todo el conjunto; las especies más variables son U. peregrina y C. mckannai. Las estaciones del grupo se ubican en la plataforma externa, a una profundidad promedio de 78 m, el sustrato es arenoso (más de 92% de arena y el resto de fango). Las aguas presentan temperatura media (T ½) de 7,95ºC, la más fría de todos los grupos y salinidad media (S‰) de 33,70 (figura 5, tabla 5).
Figura 4. Dendrograma de estaciones con los foraminíferos obtenido utilizando los datos clr-transformados, el cuadrado de la distancia euclidiana y el método de Ward / stations dendrogram with foraminifers obtained using clr-transformed data, squared Euclidean distances and the Ward´s method.
Figura 5. Grupos de estaciones definidas en/por el análisis de agrupamiento / groups of station defined with the cluster analysis.
Figura 6. Diagrama de barras de medias geométricas composicionales de los grupos de estaciones con foraminíferos determinados con el análisis de agrupamiento / bar graph of multivariate geometric compositional means of station groups of foraminifers defined with the cluster analysis. Ab, Ammonia beccarii; Bpc, Buccella peruviana f. campsi; Bm, Bulimina marginata; Bp, Bulimina patagonica; Cd, Cibicides dispars; Cf, Cibicides fletcheri; Cm, Cibicides mckannai; Ee, Epistominella exigua; Ga, Globobulimina affinis; Gs, Globocassidulina subglobosa; Na, Nonionella auris; Qs, Quinqueloculina seminula; Up, Uvigerina peregrine.
Grupo 2. Este grupo está formado por las estaciones 2, 15, 17, 20, 31 y 41. Se caracteriza por la presencia constante de E. exigua, especie dominante del grupo con porcentaje promedio de 77,5, el más alto del área estudiada (figura 6, tabla 5). Esta característica se aprecia también en el biplot (figura 2) pues las estaciones se agrupan cerca del vértice de esta componente. C. mckannai es también aquí una especie constante (0,8) aunque con valores promedio de 7,34% y alta varianza (tabla 5). G. affinis, C. dispars, C. fletcheri, B. patagonica; G. subglobosa, B. marginata y Q. seminula se encuentran en porcentajes inferiores a 4% (figura 6) y constancia de 0,5 o menores. Es el grupo con la variabilidad total mayor (15,2) producida por la gran varianza de las 9 especies presentes; en especial C. mckannai, C. dispars, C. fletcheri y G. affinis. Es importante mencionar la elevada variabilidad de C. mckannai y de la relación E. exigua/C. mckannai. En casi todas las estaciones predomina E. exigua por sobre C. mckannai, en tanto en la estación 31 la relación se invierte (tabla 2). Las estaciones de este grupo se ubican principalmente en la plataforma interna, a una profundidad promedio de 67 m, T ½ =12,63ºC, más cálidas que el grupo 1 y S‰ media =33,70 (figura 5, tabla 5). El sustrato es, al igual que el del grupo 1, esencialmente arenoso (más de 94%) con bajo contenido
en fango (tabla 1).
Grupo 3. Las 4 estaciones que integran el grupo (1, 16, 27 y 49) se distinguen por la presencia de B. peruviana f. campsi con valores promedio muy altos (56%), uno de los mayores del área estudiada para esta especie, E. exigua con valores también altos (20%), aunque menores que el promedio del área estudiada, G. affinis con porcentaje promedio de 8,54 (ligeramente superiores a la media de la región) y B. patagonica con porcentajes promedio cercanos al media de la región (7,13%) (figura 6). Otras especies presentes, aunque con menor constancia son C. fletcheri, U. peregrina, A. beccarii, Q. seminula, B. marginata, G. subglobosa y N. auris con proporciones menores a 1,2%. La variabilidad total del grupo es 11,6. Las estaciones de este grupo se ubican principalmente en la plataforma interna (figura 5), a profundidades promedio de 61 m, donde el sustrato es esencialmente arenoso con escaso fango. La estación 49 presenta 18% de fango y la estación 27 además posee 10% de grava. Las aguas tienen T ½= 16,97ºC, siendo las más cálidas del área estudiada y S‰ media =33,90 (tabla 5).
Grupo 4. Las 7 estaciones de este grupo (18, 19, 28, 42, 52, 54 y 55) se caracterizan por tener valores promedios altos de B. peruviana f. campsi de 65%, el más alto del área estudiada (figura 6), y valores de C. dispars y B. patagonica cercanos a los valores medios de la región. E. exigua se encuentra casi siempre presente, aunque con valores de composición muy inferiores al promedio de la zona. Otras especies de este grupo que se presentan con constancias de 0,7 o menores son C. fletcheri, A. beccarii, G. affinis, Q. seminula, B. marginata y N. auris, siempre con porcentajes menores a 5 (figura 6, tabla 5). En el biplot de Componentes Principales (figura 2) las estaciones que integran este grupo se ubican en torno a los vértices Q. seminula-A. beccarii. La variabilidad total del grupo es 12,5; la especie con la abundancia más variable es C. dispars. Las estaciones de este grupo se ubican en un ambiente de plataforma interna (figura 5), a profundidades promedio de 65 m. En la mayoría de las estaciones el sustrato es preponderantemente arenoso, con bajo porcentaje de fango, a excepción de las estaciones 54 y 55 que poseen entre 20 y 40% de grava. Las aguas tienen T ½=15,94ºC y S‰=33,78 (tabla 5).
Grupo 5. Este grupo está formado por 3 estaciones (44, 45 y 53) cuya característica principal es que en ellas A. beccarii alcanza un valor promedio alto, 81,13%, el mayor de toda la región bajo estudio. Acompañan a esta especie B. peruviana f. campsi, con una constancia de 0,7 y porcentajes promedio de 10 y Q. seminula (Co=0,3) con porcentajes cercanos a la media de la región (figura 6, tabla 5). La variabilidad total del grupo es de 11,4, relativamente baja, debido a la variabilidad de Q. seminula y B. peruviana f. campsi. Las estaciones de este grupo se ubican en la desembocadura del golfo San Matías (figura 5), a profundidades promedio 62 m, el sustrato es predominantemente areno-gravoso en las estaciones 44 y 45 y areno-limoso en la estación 53. Las aguas poseen T ½=16,27ºC, siendo apreciablemente más cálidas que las del área del golfo y salinidad media S‰=34,07, la mayor de toda el área de estudio (tabla 5).
Grupo 6. Este grupo está formado exclusivamente por estaciones del golfo San Matías (47, 48, 50 y 51) (figuras 4 y 5). Se distingue por la presencia de N. auris en todas las estaciones, que alcanza valores de 72,36%, el mayor de toda la región estudiada para esta especie. B. patagonica y B. marginata también se encuentran en todas las estaciones con porcentajes similares a los promedios de la región. Las otras 3 especies presentes, B. peruviana f. campsi, E. exigua y G. affinis, presentan índices de constancia elevados (0,8) pero porcentajes menores a 5%, muy inferiores a la media de la región (figura 6, tabla 5). En el biplot (figura 2) dos de las estaciones se ubican en torno a los vértices que forman el grupo de especies G. affinis-B. marginata-N. auris y B. patagonica. La variabilidad total del grupo es 8,8, indicando que es el más homogéneo de todo el conjunto. Las estaciones de este grupo se ubican a una profundidad promedio de 130 m, la mayor del área estudiada, T ½ =12,72ºC, más cálida que el grupo 1 y parecido a la del grupo 2 y S‰=34,06, similar a la del grupo 5. El sustrato es esencialmente fangoso (85% o más) con contenidos menores de arena, a excepción de la estación 48 que posee fango y arena en partes iguales.
Interpretación y discusión de los resultados
Generales para el área de estudio
Uno de los resultados más destacados es el agrupamiento de especies y estaciones en diferentes sectores del biplot (figura 2). En el semieje negativo de la CP1 el grupo de especies A, reúne algunos exponentes típicos de ambiente de plataforma externa y de la corriente de Malvinas. En el semieje positivo de la CP1 se ubican los grupos B y C que incluyen las especies características de ambientes de plataforma interna. La intensa correlación lineal (r de Pearson) que existe entre el valor de la Coordenada Principal 1 (CoP1) de cada estación con la temperatura (T ºC) del agua (corr(CoP1, T ºC) = 0,750, p<0,01), señala que sería uno de los factores responsables de esta separación. Valores negativos de la Coordenada Principal 1 corresponden a temperaturas cercanas a 12ºC, mientras que valores positivos concuerdan con temperaturas mayores.
Asimismo, en el ambiente de plataforma los cambios en las relaciones entre las especies Cibicides dispars/ Bulimina patagonica y Cibicides mckannai/ Bulimina patagonica también están vinculados con la temperatura (corr r de Pearson (ln(Cd/Bp), T ºC) = -0,785 y corr r de Pearson (ln(Cm/Bp), T ºC) = -0,852; p<0,01).
En el biplot (figura 2) se observa que el grupo A se halla representado por las especies más características de aguas frías como C. mckannai y C. dispars. Los grupos B y C se hallan definidos por especies de amplia distribución como B. patagonica y Q. seminula en la plataforma interna caracterizando un ambiente costero. Esto mostraría que en el sector de la plataforma donde C. mckannai y C. dispars están presentes las aguas tienen temperaturas más bajas que aquellas donde se encuentran presentes las especies de los grupos B y C.
Por otra parte, para toda el área estudiada, los cambios en la relación Epistominella exigua/Buccella peruviana f. campsi no están correlacionados con la temperatura, la profundidad ni con las características del sustrato medidas como ln(arena/limo) (r de Pearson: corr(ln(Ee/Bpc), T ºC) = -0,563; corr(ln(Ee/Bpc), Profundidad) = 0,028; corr(ln(Ee/Bpc); ln(arena/limo)) = 0,071; p>0,05).
La relación de abundancia de individuos entre Epistominella exigua y Buccella peruviana f. campsi fue reconocida por Boltovskoy y Totah (1985) quienes señalan una zona de transición entre la corriente Patagónica y la corriente de Malvinas, representada por una mezcla de especies de la zona costera y de la corriente de Malvinas. Por su parte, sobre la base de frecuencias relativas, Ferrero (2006) reconoce en los niveles superiores de dos testigos de la plataforma continental, altas proporciones de E. exigua y bajas proporciones de B. peruviana f. campsi, sugiriendo una profundización del área analizada.
Distribución geográfica de las especies
El género Ammonia Brünnich es representativo de ambientes de plataforma interna (Brock, 1999), donde está ampliamente distribuido y presenta gran variabilidad morfológica. Las distintas especies viven en sedimentos fangosos a arenosos, en aguas salobres, marinas e hipersalinas (Murray, 1991), con temperaturas de templado cálidas a tropicales (0 a 30ºC) y profundidades entre los 0 y 50 m. Ammonia beccarii se presenta en ambientes salobres a marinos; se halla en bahías, golfos, estuarios y en ambientes marino marginales (Boltovskoy, 1976; Laprida, 1998; Bernasconi y Cusminsky, 2005b, 2007; Bernasconi, 2006; Cusminsky et al., 2006). Se distribuye a lo largo de la provincia biogeográfica Argentina (Boltovskoy et al., 1980). Se la cita también en sedimentos gruesos como arena y grava (Bernasconi y Cusminsky, 2005b, 2007). Nuestros hallazgos corresponden a la boca del golfo San Matías y al sector más austral del área de estudio a profundidades que oscilan entre los 30 y 90 m y están en concordancia con los de otros autores.
El género Buccella Anderson es mencionado por Murray (1991) como característico de plataforma interna (0-100 m) a zona batial, de temperatura templado- fría. Bucella peruviana f. campsi tiene una amplia distribución. Esta especie es encontrada a lo largo de toda la Plataforma Continental Argentina (Boltovskoy, 1976), tanto en áreas engolfadas (Bernasconi y Cusminsky, 2005a, Bernasconi, 2006) como en la plataforma continental abierta (Bernasconi y Cusminsky, 2005b, 2006, 2007; Cusminsky et al., 2006). Se la encuentra en aguas de la corriente de Malvinas (Boltovskoy et al., 1980; Giussani de Khan y Watanabe, 1980). También ha sido registrada en ambientes marino- marginales así como en lagunas y estuarios (Boltovkoy y Totah, 1985; Laprida, 1998; Laprida y Bertels-Psotka 2003; Gómez et al., 2005, 2006). Nuestros hallazgos son concordantes con los de los autores mencionados. En el área estudiada la mayor abundancia se produce entre 46 y 50 m, aunque se la registró hasta la isobata de 80 m. Estos datos coinciden con lo hallado por Boltovkoy y Totah (1985) quienes la encuentran entre los 40 y 68 m de profundidad.
Bulimina d´ Orbigny vive en medios fangosos a arenas finas y aguas frías a templadas en un ambiente de plataforma interna a batial (Murray, 1991). La especie Bulimina marginata se distribuye en la Plataforma Continental Argentina (Boltovskoy et al., 1980). Se la cita en ambientes de plataforma abierta (Bernasconi y Cusminsky, 2005b, 2007) y en áreas engolfadas (Bernasconi y Cusminsky, 2005a; Bernasconi, 2006), también en aguas salobres del litoral argentino y en la laguna Dos Patos (Brasil) (Boltovskoy et al., op. cit.). En este trabajo B. marginata se encuentra principalmente en toda la zona del golfo y en la zona de plataforma interna en proporciones que no superan el 10%. B. patagonica habita en ambientes salobres a marinos y de temperatura moderada a fría. Se halla en plataforma y también en la desembocadura del río Quequén y río de La Plata (Boltovskoy et al.,op. cit). En el sector de la plataforma bajo estudio tiene una amplia distribución pues se encuentra en toda el área del golfo y en la zona de plataforma interna.
El género Cibicides Montfort es de aguas frías a cálidas, se presenta en ambientes de plataforma a zona batial entre 0 y más de 2000 m (Murray, 1991). Las especies Cibicides mckannai, C. dispars y C. fletcheri se distribuyen a lo largo de la provincia biogeográfica Argentina, en aguas de la corriente de Malvinas (Boltovskoy et al., 1980; Ferrero, 2006). C. dispars se cita en áreas engolfadas (Bernasconi y Cusminsky, 2005a; Bernasconi, 2006) y en plataforma abierta (Bernasconi y Cusminsky, 2006, 2007). El gradiente de abundancia de nuestros hallazgos, con un aumento relativo hacia la zona más profunda, es coincidente con lo hallado por Boltovkoy y Totah (1985), lo que estaría indicando una preferencia de estas especies por aguas frías.
Epistominella Musezima y Maruhasi es un género que vive en ambientes marinos de plataforma a zona batial (Murray, 1991), propio de aguas de templadas a frías. Lena (1976 en Ferrero, 2006) señala que la zona Costera Argentina, que se encuentra limitada al este por la corriente de Malvinas, está caracterizada por la dominancia de Epistominella exigua. Se cita en aguas salobres del río de la Plata y río Quequén. (Boltovskoy et al., 1980), fue reconocida entre los 68 y 120 m de profundidad por Boltovskoy y Totah (1985) y en áreas engolfadas por Bernasconi y Cusminsky (2005a). Se trata de una especie que prefiere aguas frías y bien oxigenadas (Ferrero, 2006). Las mayores abundancias se encontraron en el sector más septentrional del área estudiada.
Globobulimina Cushman vive en sedimentos fangosos, se encuentra en aguas templadas a frías, en un ambiente de plataforma a zona batial (Murray, 1991). Globobulimina affinis es citada en la Plataforma Continental Argentina, en áreas engolfadas de los golfos San Jorge, San Matías y Nuevo y en bahía San Blas (Boltovskoy, 1954a, 1954b; Bernasconi y Cusminsky, 2005a; Bernasconi, 2006). En el área bajo estudio se halló principalmente en el sector del golfo donde registró la mayor abundancia y en la zona de plataforma interna.
Globocassidulina Voloshinova es un género que vive en medios fangosos, a temperaturas templadas a frías y se la encuentra en ambiente de plataforma hasta zona batial (Murray, 1991). Globocassidulina subglobosa tiene una amplia distribución en la Plataforma Continental Argentina, se la cita en las zonas de influencia del río de la Plata (Boltovskoy, 1957; Boltovskoy et al., 1980) y en áreas engolfadas (Boltovskoy, 1954a; Bernasconi y Cusminsky, 2005a; Bernasconi, 2006) como así también en la zona de Mar Chiquita (Ferrero, 2006). En este trabajo se halló a G. subglobosa fundamentalmente distribuida en las estaciones ubicadas en la zona de la plataforma externa.
Nonionella Cushman se encuentra entre los 10-1000 m de profundidad; puede habitar ambientes marinos de salinidad normal hasta salobre (Murray, 1991). Es citado como un género que vive en un medio fangoso (Murray, op. cit.; Harloff y Mackensen, 1997). Hayward et al. (2002) sugieren que el fango y la profundidad son los factores que más influyen en su distribución. Nonionella auris se menciona en sedimentos fangosos (Bernasconi y Cusminsky, 2005a); se halla tanto en ambientes de plataforma (Malumián, 1972; Boltovskoy et al., 1980) como en ambientes engolfados (Boltovskoy, 1954a; Bernasconi y Cusminsky, 2005a; Bernasconi, 2006), de estuarios (Boltovskoy, 1957) y de bahías (Boltovskoy, 1954b). En este trabajo se la encuentra prácticamente restringida al golfo San Matías en medio fangoso y a una profundidad promedio de 130 m, la mayor de todo el sector estudiado.
Quinqueloculina d´ Orbigny es citado como de ambientes marinos a hipersalinos de plataforma y raramente batial (Murray, 1991). Es representativo de ambientes de plataforma interna (Boltovskoy, 1965; Brock, 1999). Quinqueloculina seminula tiene una amplia distribución a lo largo de toda la Plataforma Continental Argentina (Boltovskoy, 1976; Boltovskoy et al., 1980). Es una especie de ambientes de poca profundidad, capaz de soportar cambios muy bruscos de las condiciones ecológicas en la zona litoral, donde tienen lugar los movimientos de agua con diferente carácter y fuerza (Boltovskoy, 1965; Gómez et al., 2005; Cusminsky et al., 2006). En el sector estudiado, esta especie es relativamente más abundante en las estaciones de la entrada al golfo, caracterizadas por profundidades entre 30 y 90 m y un sustrato gravoso, lo cual indicaría un ambiente sometido a un régimen de mareas y corrientes costeras de alta energía. Esto se explica por las características que, en general, poseen los caparazones de miliólidos que se encuentran principalmente en plataforma continental (Brock, 1999). Según Usera y Blázquez (1997) la resistencia que ofrecen a la erosión caparazones de este tipo favorece el aumento de la abundancia relativa que experimentan en sustratos gravosos.
Uvigerina d´ Orbigny habita en sedimentos fangosos, entre los 100 m y más de 4500 m (Murray, 1991). Uvigerina peregrina se encuentra en la Plataforma Continental Argentina y en áreas engolfadas (Bernasconi y Cusminsky, 2005a). Se distribuye a lo largo de la provincia biogeográfica Argentina (Boltovskoy et al., 1980) y es de aguas frías (Kafescioglu, 1975). En el sector estudiado se encuentra principalmente en las mayores profundidades en la zona de plataforma externa.
Las asociaciones de especies
El análisis de los cambios en la composición de la asociación de las 13 especies de foraminíferos en los grupos de estaciones permite distinguir claramente dos ambientes marinos: un ambiente de plataforma externa caracterizado por la influencia de la corriente de Malvinas y un ambiente de plataforma interna con predominio de las aguas de Plataforma Media (Guerrero y Piola, 1997) y de las aguas costeras de alta salinidad (Guerreo y Piola op. cit.). Esta interpretación se fundamenta tanto en el análisis del biplot de estaciones y especies como en la conformación de grupos de estaciones como resultado del análisis de agrupamiento (figuras 2, 4 y 5). Por un lado, se disponen las estaciones de los grupos 1 y 2 en el semieje negativo de la CP1 y cercanas a los vértices de las especies que se hallan en aguas de la corriente de Malvinas (E. exigua, C. dispars, U. peregrina, G. subglobosa, C. mckannai), y por el otro, en el semieje positivo de la CP1 se disponen los grupos 3, 4, 5 y 6 que se caracterizan por la presencia de especies de plataforma interna (A. beccarii, B. peruviana f. campsi, N. auris, Q. seminula, G. affinis y B. patagonica). Esto además se evidencia en la separación en dos grandes ramas en el dendrograma, la de la izquierda con los grupos 1 y 2 y la de la derecha con los grupos 3, 4, 5 y 6.
Los grupos 1 y 2 están caracterizados por la presencia de especies de la corriente de Malvinas; sin embargo en el grupo 2 hay una predominancia neta de E. exigua por sobre las otras especies. Esto podría sugerir una disminución del área de influencia de la corriente de Malvinas, indicando que entre el límite occidental de esta masa de agua y la masa de agua costera o corriente Patagónica se ubica un sector de transición. Este sector de transición podría estar relacionado con el quiebre de pendiente descrito por Martos y Piccolo (1988). Esta inferencia está de acuerdo a lo propuesto por Boltovskoy y Totah (1985).
Por otra parte, tanto en el semieje positivo de la CP1 (figura 2) como en la rama derecha del dendrograma (figura 4), se pueden reconocer tres ambientes: el sector de la plataforma interna, el del golfo San Matías y un tercero, de transición entre ambos.
En la plataforma interna predomina B. peruviana f. campsi con más de 55% de la composición total en los grupos 3 y 4. En el grupo 3 esta especie está acompañada por E. exigua, G. affinis y B. patagonica y en el grupo 4 por C. dispars, B. patagonica, E. exigua y C. fletcheri. La presencia de estas especies sumada a la ubicación de las estaciones de ambos grupos indicaría la influencia del Agua de Plataforma Media fluyendo como la corriente Costera Patagónica.
El ambiente de transición se ubica en la zona de entrada al golfo San Matías donde prevalece A. beccarii acompañada por B. peruviana f. campsi y por pequeñas proporciones de Q. seminula (grupo 5). La presencia de sedimentos predominantemente gruesos estaría indicando un ambiente sometido a un régimen de mareas y corrientes costeras de alta energía.
En la zona del golfo San Matías la especie más importante es Nonionella auris que está acompañada por B. patagonica, B. marginata, B. peruviana f. campsi, E. exigua y G. affinis en menor proporción (grupo 6). La preponderancia de N. auris fue analizada por Bernasconi y Cusminsky (2005a) quienes señalan que su predilección por los sedimentos finos indican ambientes de baja energía. Las características de las aguas de los grupos 5 y 6 sugieren la influencia del Agua Costera con Alta Salinidad definida por Guerrero y Piola (1997).
Conclusiones
Los análisis estadísticos realizados se fundaron en la geometría composicional y en la premisa de que los estudios deben orientarse hacia las diferencias relativas entre especies. Los resultados obtenidos con el empleo del ACP Composicional, el análisis de Agrupamiento, las relaciones señaladas con las Coordenadas Principales y con diferentes log-cocientes entre especies han colaborado a interpretar los cambios de abundancia de foraminíferos bentónicos en este sector de la Plataforma Continental Argentina. A través de las variaciones conjuntas que experimenta la composición de las 13 especies se distinguieron diferentes asociaciones. Dichos cambios están vinculados con las variaciones de la temperatura y salinidad del agua, de las características del fondo y de las condiciones energéticas en el área. A partir del análisis microfaunístico, se identifica la influencia de las aguas de la corriente de Malvinas en el ambiente de plataforma externa; por otro lado, el Agua de Plataforma Media fluyendo como la corriente Costera Patagónica y el Agua Costera con Alta Salinidad se reconoce en el ambiente de plataforma interna y golfo San Matías.
Agradecimientos
Este trabajo fue financiado por la Agencia de Promoción Científica y Tecnológica (Proyectos BID PICT - 09659; 14653 y 26057; CONICET PIP 02040 y 6416 y UNC B 940. Las autoras desean agradecer al Instituto Argentino de Oceanografía (IADO) por facilitar las muestras. Especial reconocimiento para M. Mena y el árbitro anónimo por las pertinentes sugerencias que han permitido mejorar sustancialmente este trabajo.
Bibliografía
1. Aitchison, J. 1986. The statistical analysis of compositional data. Monographs on Statistics and applied Probability. Chapman and Hall Ltd., London (UK), 416 pp. [ Links ]
2. Aitchison, J. 1992. On criteria for measures of compositional difference. Mathematical Geology 24: 365-379. [ Links ]
3. Aitchison, J. 1997. The one-hour course in compositional data analysis or compositional data analysis is simple. In: V. Pawlowsky- Glahn (ed.) Proceedings of IAMG`97. The 3° Annual Conference of International Associations for Mathematical Geology International Center for Numerical Methods in Engineering (CIMNE) (Barcelona España), 1, 3-35. [ Links ]
4. Aitchison, J. 1999. Logratios and natural laws in compositional data analysis. Mathematical Geology 131: 563-580. [ Links ]
5. Aitchison, J. y Greenacre, M. 2002. Biplots of compositional data. Applied Statistics 51, 4: 375-392. [ Links ]
6. Aitchison, J., Barceló-Vidal, C., Martín-Fernández, J.A. y Pawlowsky-Glahn, V. 2000. Logratio analysis and compositional distance. Mathematical Geology 32: 563-580. [ Links ]
7. Bernasconi, E. 2006. [Los foraminíferos del Holoceno de testigos de la plataforma continental argentina (40º 30´- 42º 48´ LS y 59º 25´- 64º 40´LO). Tesis Doctoral, Universidad Nacional del Comahue, Bariloche, 210 pp, 7 pl. Inédito]. [ Links ]
8. Bernasconi, E. y Cusminsky, G. 2005a. Distribución de Nonionella auris (d'Orbigny) (Foraminiferida) en el golfo San Matías, Provincia de Río Negro, Argentina. Ameghiniana 42: 167-174. [ Links ]
9. Bernasconi, E. y Cusminsky, G. 2005b. Foraminíferos de un testigo de la plataforma continental argentina. 16° Congreso Geológico Argentino (La Plata), Actas 3: 279- 284. [ Links ]
10. Bernasconi, E. y Cusminsky G. 2006. Análisis preliminar cuantitativo del Orden Foraminiferida de un testigo de la Plataforma Continental argentina: Tafonomía y paleoecología. 3° Congreso Argentino de Cuaternario y Geomorfología (Córdoba), Actas 2: 681- 690. [ Links ]
11. Bernasconi, E. y Cusminsky, G.C. 2007. Foraminíferos bentónicos de un testigo holocénico de la plataforma continental argentina.(40º 30' 00'' S y 60º 59'05'' O). Ameghiniana 44: 271-278. [ Links ]
12. Bertels, A. 1984. Ciclos de disolución de foraminíferos planctónicos en el océano Atlántico Sudoccidental Austral: significancia paleoocenaográfica durante el Mioceno Superior-Plioceno Inferior. 3° Congreso Latinoamericano de Paleontología, Actas: 525-545. [ Links ]
13. Bertels, A. 1995. Cenozoic marine sedimentary basins of Argentina. Some palaeoenvironmental features based on foraminifera. En: S. Jardiné y de Klasz y J. P. Debeany (ed.) Geologie de l`Afrique et de l' Amerique Sud Complete Rendu des Colloques de Geologie d'Angers. Elf Aquitaine (Pau, Francia), pp. 351-359. [ Links ]
14. Boesch, D.F. 1973. Classification and community structure of macrobenthos in the Hampton Roads area, Virginia. Marine Biology 21: 226-224. [ Links ]
15. Boesch, D.F. 1977. Applications of numerical classification in ecological investigations of water pollution. Virginia Institute Marine Science, Special Science Reports, 144 pp. [ Links ]
16. Boltovskoy, E. 1954a. Foraminíferos del Golfo San Jorge. Revista Instituto Nacional de Investigación de las Ciencias Naturales y Museo Argentino de Ciencias Naturales Bernardino Rivadavia 3: 85-246. [ Links ]
17. Boltovskoy, E. 1954b. Foraminíferos de la Bahía San Blas. Revista del Instituto Nacional de Investigación de las Ciencias Naturales y Museo Argentino de Ciencias Naturales Bernardino Rivadavia 3: 247-300. [ Links ]
18. Boltovskoy, E. 1957. Los Foraminíferos del Estuario del Río de La Plata y su zona de influencia. Revista del Instituto Nacional de Investigación de las Ciencias Naturales y Museo Argentino de Ciencias Naturales Bernardino Rivadavia 6: 84 pp. [ Links ]
19. Boltovskoy, E. 1959. Foraminíferos Recientes del Sur de Brasil y sus relaciones con los de Argentina e India del Oeste. Revista del Servicio Hidrografía Naval 1005, 124 pp. [ Links ]
20. Boltovskoy, E. 1965. Los Foraminíferos recientes. Editorial Universitaria de Buenos Aires, 510 pp. [ Links ]
21. Boltovskoy, E. 1976. Distribution of Recent Foraminifera of the South American Region. En: Hedley, R.H. y Adams, C.G. (eds.) Foraminifera. Academic Press (London) 2: 171-236. [ Links ]
22. Boltovskoy, E. 1978. Estudio bioestratigráfico y paleontológico (foraminíferos bentónicos) del Cenozoico superior al este de las Islas Malvinas (DSDP, Crucero 36, sitios 327 y 329). Revista Instituto Nacional de Investigaciones Naturales y Museo de Ciencias Naturales Bernardino Rivadavia 8: 19-70. [ Links ]
23. Boltovskoy, E. 1979. Paleoceanografia del Atlántico Sudoccidental desde el Mioceno, según estudios foraminiferológicos. Ameghiniana 16: 357-389. [ Links ]
24. Boltovskoy, E. 1981. Masas de agua en el Atlántico Sudoccidental. En: D. Boltovskoy (ed.) Atlas de Zooplancton del Atlántico Sudoccidental. INIDEP pp. 227-236. [ Links ]
25. Boltovskoy, E. y Giussani de Khan, G. 1982. Foraminíferos bentónicos calcáreos uniloculares del Cenozoico Superior del Atlántico Sur. Revista de la Asociación Geológica Argentina 37: 408-448. [ Links ]
26. Boltovskoy, E. y Totah, V. 1985. Diversity, Similarity and Dominance in benthic foraminiferal fauna along one transect of the argentine shelf. Revue de Micropaléontologie 28: 23-31. [ Links ]
27. Boltovskoy, E. y Watanabe, S. 1977. Foraminíferos calcáreos uniloculares de profundidades grandes del Atlántico Sur y del Índico (Neógeno-Reciente). Revista Instituto Nacional de Investigaciones Naturales y Museo de Ciencias Naturales Bernandino Rivadavia 5: 41-64. [ Links ]
28. Boltovskoy, E., Boltovskoy, D., Correa, N. y Brandini, F. 1996. Planktic foraminifera from the southwestern Atlantic (30º-60º S): species-specific patterns in the upper 50 m. Marine Micropaleontology 28: 53-72. [ Links ]
29. Boltovskoy, E., Giussani, G., Watanabe, S. y Wright, R. 1980. Atlas of benthic shelf foraminifera of the southwest Atlantic. W. Junk. Publishers, The Hague, 147 pp. [ Links ]
30. Brock, G. 1999. Application of Foraminiferida in earth sciences. GEOS 39: 1-6. [ Links ]
31. Buccianti, A. y Esposito, P. 2004. Insights into Late Quaternary calcareous nannoplankton assemblages under the theory of statistical analysis for compositional data. Palaeogeography, Paleoclimatology, Palaeoecology 202: 209-227. [ Links ]
32. Caramés, A. y Malumián, N. 2000. Foraminíferos bentónicos del Cenozoico del pozo Estrella x-1, Cuenca del Colorado, plataforma continental argentina. Ameghiniana 37: 387-419. [ Links ]
33. Chayes, F. 1960. On correlation between variables of constant sum. Journal Geophysical Research 65: 4185-4193. [ Links ]
34. Chayes, F. 1971. Ratio Correlation: A Manual for Students of Petrology and Geochemistry. The University of Chicago Press, Chicago, 112 pp. [ Links ]
35. Clifford, H.T. y Stephenson, W. 1975. An Introduction to Numerical Classification. Academic Press (New York), 229 pp. [ Links ]
36. Cortelezzi, C. R. y Mouzo, F.H. 1979. Estudio sedimentológico de testigos del Margen Continental Argentino sobre un perfil a los 40º Latitud Sur. 6° Congreso Geología Argentina (Buenos Aires), Actas 2: 249-264. [ Links ]
37. Cusminsky, G. 1991. Foraminíferos planctónicos de testigos Cenozoicos del océano Atlántico sudoccidental austral. Ameghiniana 28: 225-240. [ Links ]
38. Cusminsky, G. 1992. Foraminíferos bentónicos provenientes de testigos del océano Atlántico Sudoccidental Austral. Revista Española de Micropaleontología 24: 5-32. [ Links ]
39. Cusminsky, G. 1994. Estudio micropaleontológico (Foraminiferida) de dos testigos extraídos del océano Atlántico sudoccidental austral. Revista Española de Micropaleontología 26: 109-123. [ Links ]
40. Cusminsky, G., Martínez, D. y Bernasconi, E. 2006. Foraminíferos y ostrácodos de sedimentos recientes del estuario de Bahía Blanca, Argentina. Revista Española de Micropaleontología, 38: 395-410. [ Links ]
41. Ferrero, L. 2006. [Micropaleontología y Paleoecología del Cuaternario del sudeste de la provincia de Buenos Aires. Tesis Doctoral, Universidad Nacional de Mardel Plata, Facultad de Ciencias Exactas y Naturales, 373 pp. Inédito]. [ Links ]
42. Gelós, E. y Spagnuolo, J. 1992. Relación área de aporte-ambiente tectónico de las psamitas de la plataforma continental argentina entre los 39º y 43º de latitud sur y el golfo San Matías. Revista de la Asociación Geológica Argentina 47: 141-146. [ Links ]
43. Gelós, E., Spagnuolo, J. y Lizasoain, G. 1988. Mineralogía y caracterización granulométrica de sedimentos actuales de la plataforma argentina entre los paralelos 39° y 43° de latitud Sur y del golfo San Matías. Revista de la Asociación Geológica Argentina 43: 63-79. [ Links ]
44. Giussani de Khan, G. y Watanabe, S. 1980. Foraminíferos bentónicos como indicadores de la corriente de Malvinas. Revista Española de Micropaleontología, 12: 169-177. [ Links ]
45. Gómez, E., Martínez, D., Borel, M., Guerstein, G.R. y Cusminsky, G.C. 2005. Submarine evidences of Holocene sea-level fluctuations in the Bahía Blanca estuary, Argentina. Journal of South America Earth Sciences 20: 139-155. [ Links ]
46. Gómez, E. A., Martínez, D., Borel, C.M., Guerstein, G.R. y Cusminsky, G.C. 2006. Negative sea-level oscillation at the Bahía Blanca estuary related to a ca. 2650 yr BP global climatic change. Brasil. Journal of Coastal Research, Special Issue, 39: 181-185. [ Links ]
47. Guerrero, R.A y Piola, A.R. 1997. Masas de agua en la Plataforma Continental. En: El Mar Argentino y sus recursos pesqueros, Contribución INIDEP Nº 998, 1: 107-118. [ Links ]
48. Harloff, J. y Mackensen, A. 1997. Recent benthic foraminiferal associations and ecology of the Scotia Sea and Argentine Basin. Marine Micropaleontology 31: 1-29. [ Links ]
49. Hayward, B., Neil, H., Carter, R., Grenfell, H. y Hayward, J. 2002. Factors influencing the distribution patterns of Recent deepsea benthic foraminifera, east of New Zealand, Southwest Pacific Ocean. Marine Micropaleontology 46: 139-176. [ Links ]
50. Jackson, D. A. 1997. Compositional data in community ecology: the paradigm or peril of proportions?. Ecology 78: 929-940 [ Links ]
51. Kafescioglu, I. 1975. Quantitative distribution of foraminifera on the continental shelf and uppermost slope off Massachusetts. Micropaleontology 21: 261-305. [ Links ]
52. Kucera, M. y Malmgren, B.A. 1998. Logratio transformation of compositional data-a resolution of the constant sum constraint. Marine Micropaleontology 34: 117-120. [ Links ]
53. Laprida, C. 1998. Micropaleontological assemblages (Foraminiferida and Ostracoda) from Late Quaternary marginal marine environments (Destacamento Río Salado Formation), Salado Basin, Argentina. Revue de Paléobiologie 17: 461-478. [ Links ]
54. Laprida, C. y Bertels-Psotka, A. 2003. Benthic foraminifers and paleoecology of a Holocene shelly concentration, Salado Basin, Argentina. Geobios, 36: 559-572. [ Links ]
55. Loeblich, A, Jr. y Tappan, H. 1988. Foraminiferal Genera and their Classification. Van Nostrand Reinhold Company. New York, 1- 2: 1182 pp. [ Links ]
56. Malumián, N. 1972. Foraminíferos del Oligoceno y Mioceno del subsuelo de la provincia de Buenos Aires. Ameghiniana 9: 97- 137. [ Links ]
57. Martín-Fernández, J.A. 2001.[Measures of difference and non-parametric classification of Compositional Data. Tesis Doctoral, Universitat Politècnica de Catalunya, 233 pp. Inédito.] [ Links ]
58. Martín-Fernández, J.A., Barceló-Vidal, C. y Pawlowsky-Glahn, V. 1998. Measures of Difference for Compositional Data and Hierarchical Clustering Methods. En: Buccianti, A., Nardi, G. y Potenza, R., Naples (eds.) Proceedings of the Fourth Annual Conference of the International Association for Mathematical Geology 1: 526-531. [ Links ]
59. Martín-Fernández, J.A., Barceló-Vidal, C. y Pawlowsky-Glahn, V. 2003. Dealing with zeros and missing values in compositional data sets using nonparametric imputation. Mathematical Geology 35: 253-277. [ Links ]
60. Martos, P. y Píccolo, M.C. 1988. Hydrography of the Argentine continental shelf between 38º and 42º S. Continental Shelf Research, vol. 8: 1043-1056. [ Links ]
61. Monechi, S., Buccianti, A. y Gardin, S. 2000. Biotic signals from nannoflora across the iridium anomaly in the upper Eocene of the Massignano section: evidence from statistical analysis. Marine Micropaleontology 39: 219-237. [ Links ]
62. Murray, J. W. 1991. Ecology and paleoecology of benthic foraminifera. Logman Scientific Technical, 397 pp. [ Links ]
63. Parker, G., Violante, R. y Paterlini, M. 1996. Fisiografía de la Plataforma Continental. En: Ramos, V. y Turic, A. (eds.), Geología y Recursos Naturales de la Plataforma Continental Argentina. 8° Congreso Geológico Argentino y 3º Congreso de Exploración de Hidrocarburos (Buenos Aires), Relatorio 1: 1-16. [ Links ]
64. Pawlowsky-Glahn, V. y Egozcue, J.J. 2001. Geometric approach to statistical analysis on the simplex. Stochastic Environmental Research and Risk Assessment 15: 384-398. [ Links ]
65. Pawlowsky-Glahn, V. y Egozcue, J.J. 2002. BLU estimators and compositional data. Mathematical Geology 34: 259-274. [ Links ]
66. Pearson, K. 1897. Mathematical contributions to the theory of evolution on a form of spurious correlation which may arise when indices are used in the measurements of organs. Proccedings Royal Society 60: 489-498. [ Links ]
67. Pielou, E.C. 1977. Mathematical ecology. John Wiley and Sons, New York, USA, 385 pp. [ Links ]
68. Pucci, E., Asteasuain, R., Villa, N., Lara, R., Rusansky, C., García, O., Marcovecchio, J., Asteasuain, A. y Cejas, J. 1985. Nutrientes, factores químicos y físicos relacionados y perfiles en aguas de la plataforma continental argentina. Informe técnico IADO 11, Bahía Blanca, 179 pp. [ Links ]
69. Rossi, R.E., Mull, D.J. y Franz, E.H. 1992. Geostatistical tools for modeling and interpreting ecological spatial dependence. Ecological Monographs 62:277-314. [ Links ]
70. Sutton, T., Dhar, T. y Conteras, G. 2006. Quantum GIS 0.8 'Titan' Free and Open Source Software for Geographic Information System for Windows. http://download.qgis.org/qgis/ [ Links ]
71. Thio, S. 2004. CoDaPack software package. Universidad de Girona, España. http://ima.udg.es/~thio/#Compositional Data [ Links ]
72. Usera, J. y Blázquez, A. 1997. Influencia del substrato en la distribución de los foraminíferos bentónicos de la plataforma continental interna entre Valencia y Alicante (España). Revista Española de Micropaleontología 29: 85-104. [ Links ]
Recibido: 10 de abril de 2008.
Aceptado: 11 de mayo de 2007.

