










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.45 n.3 Buenos Aires jul./sep. 2008
Gastrolitos en un plesiosaurio (Sauropterygia) de la Formación Allen (Campaniano-Maastrichtiano), provincia de Río Negro, Patagonia, Argentina
Ignacio A. Cerda1 y Leonardo Salgado1
1CONICET-INIBIOMA- Museo de Geología y Paleontología, Universidad Nacional del Comahue, Buenos Aires 1400, 8300 Neuquén, Argentina. nachocerda6@yahoo.com.ar, lsalgado@uncoma.edu.ar
Abstract. Gastroliths in a plesiosaur (Sauropterygia) from the Allen Formation (Campanian-Maastrichtian), Río Negro Province, Patagonia, Argentina. A group of 389 gastroliths associated to a partially preserved skeleton of a plesiosaur from the Allen Formation (Campanian-Maastrichtian), found in the Río Negro Province (Patagonia, Argentina), is described. The gastroliths were in situ in the abdominal region of the animal. With respect to their form, spheroidal and discoidal gastroliths, predominated over the laminar. The mean index of sphericity is of 0,686 (d.s ± 0,108). These two last data suggest that the plesiosaur from Río Negro, as many others found in other localities of the world (for instance, Kansas and Montana, in the United States), would have ingested the pebbles in the mouths of the rivers. Estimations on this and others plesiosaur specimens suggest that the gastroliths would have had a digestive function, more than hydrostatic.
Resumen. Se describe un conjunto de 389 gastrolitos asociados a un esqueleto parcialmente preservado de plesiosaurio proveniente de la Formación Allen (Campaniano-Maastrichtiano), hallado en la provincia de Río Negro (Patagonia, Argentina). Los gastrolitos se hallaban in situ, contenidos en la región abdominal del animal. Con respecto a su forma, predominan los guijarros esferoidales y discoidales por sobre los laminares. El índice de esfericidad promedio es de 0,686 (d.s ± 0,108). Estos dos últimos datos sugieren que el plesiosaurio de Río Negro, al igual que otros ejemplares encontrados en otras partes del mundo (por ejemplo, Kansas y Montana, en Estados Unidos), habrían ingerido las rocas en la desembocadura de los ríos. Cálculos realizados en éste y en otros especímenes de plesiosaurios, sugieren que los gastrolitos habrían desempeñado una función digestiva, más que hidrostática.
Key words. Gastroliths; Lithophagy; Plesiosaur; Late Cretaceous; Patagonia.
Palabras clave. Gastrolitos; Litofagia; Plesiosaurio; Cretácico Tardío; Patagonia.
Introducción
La litofagia o ingesta intencional de rocas, es una conducta ampliamente difundida entre los vertebrados, tanto actuales como extintos (Taylor, 1993; Whittle y Everhart, 2000). En los grupos extintos, la evidencia más concluyente a favor de ese comportamiento es la asociación íntima de piedras, generalmente redondeadas, a restos óseos.
Entre los grupos fósiles que cuentan con una mayor cantidad de especies litófagas, se encuentran los plesiosaurios (Williston, 1893, 1894, 1904; Brown, 1904; Darby y Ojakangas, 1980; Storrs, 1993; Taylor, 1993, y referencias allí citadas; Whittle y Everhart, 2000, y referencias allí citadas; Everhart, 2000, 2004, 2005; Gasparini y Salgado, 2000; Sato y Storrs, 2000; Cicimurri y Everhart, 2001, y referencias allí citadas; Sato, 2003; Thompson et al., 2004; Hiller et al., 2005; McHenry et al., 2005; Schmeisser et al., 2005). No obstante, a pesar del abundante registro de gastrolitos asociados a esos reptiles marinos, son relativamente escasos los trabajos en donde se desarrolle un análisis detallado de los mismos y en los cuales se discuta su significado paleobiológico. Entre los trabajos más relevantes sobre el tema se encuentran los de Darby y Ojakangas (1980), Everhart (2000), y Cicimurri y Everhart (2001).
El material de estudio presentado en este trabajo corresponde a un conjunto de clastos asociados a un espécimen subadulto de plesiosaurio. El mismo fue extraído en el año 1996 de una explotación minera de arcilla montmorillonítica (bentonita), ubicada en depósitos correspondientes al miembro medio de la Formación Allen (Campaniano-Maastrichtiano; Ballent, 1980). El sitio de extracción se encuentra ubicado en el ángulo noroccidental de la Cuenca Vidal (noroeste de la provincia de Río Negro), ocupada actualmente por el cuerpo de agua artificial del Lago Pellegrini. El ejemplar (MCS-Pv 4) se encuentra representado por una serie de vértebras cervicales, pectorales, dorsales y pélvicas, las cinturas pélvica y escapular, así como distintos elementos apendiculares y restos de costillas. De acuerdo con los estudios efectuados por Gasparini y Salgado (2000), este espécimen es asignado a la familia Elasmosauridae (Cope, 1869, sensu O'Keefe, 2001), un grupo de plesiosauroideos ampliamente registrado en prácticamente todo el mundo (O'Keefe, 2001).
El MCS-Pv 4 presentaba una gran cantidad de clastos alojados de forma apiñada en su región abdominal, los cuales fueron interpretados como gastrolitos in situ por Gasparini y Salgado (2000). Los clastos asociados al MCS-Pv 4 constituyen un rasgo litológico excepcional con relación al sedimento circundante (arcilita montmorillonítica), lo cual, sumado a sus rasgos tafonómicos (grado de articulación del esqueleto, posición de los clastos, etc.), muestra una clara concordancia con los criterios establecidos por Wings (2004: 84) para el reconocimiento de piedras gástricas en ejemplares fósiles.
En este trabajo se describen e interpretan los guijarros incluidos en el MCS-Pv 4, comparándolos con probables piedras gástricas asociadas a otros restos de plesiosaurios. Además, se discuten los nuevos aportes que brinda este hallazgo con respecto a la posible función de los gastrolitos en plesiosaurios. Esta contribución es la primera en donde se describen detalladamente las piedras gástricas de un plesiosaurio sudamericano.
Abreviaturas. MCS-Pv, Museo de Cinco Saltos, Paleovertebrados, Río Negro, Argentina. MUCPv, Museo de la Universidad Nacional del Comahue, Paleovertebrados, Neuquén, Argentina.
Métodos
Durante las tareas de preparación del MCS-Pv 4, los clastos fueron removidos de su posición original (la región abdominal del ejemplar) sin que se tomara un registro fotográfico. De todos modos, prácticamente todos los guijarros, incluso los de menor tamaño, fueron recuperados.
Para el análisis de los clastos se ha tenido en cuenta una serie de parámetros y propiedades, los cuales figuran en otros trabajos sobre piedras estomacales, tanto de reptiles marinos (Darby y Ojakangas, 1980; Everhart, 2000; Cicimurri y Everhart, 2001), como de dinosaurios noavianos (Kobayashi et al, 1999; Sanders et al, 2001). Esos parámetros y propiedades incluyen: el número total de clastos, la longitud de los tres ejes principales de cada uno de éstos [a: mayor; b: medio y c: menor (figura 1)] (medición realizada con un calibre milimetrado), el índice de esfericidad de máxima proyección (ψ) = (Blatt et al, 1972), el grado de redondeamiento de cada clasto (siguiendo las seis categorías propuestas por Powers [1953]), sus formas (siguiendo la clasificación de Zingg., la cual toma en cuenta la relación entre los ejes a, b y c) (Pettijohn, 1980) y composición litológica. Otras magnitudes tenidas en cuenta son el peso y el volumen del conjunto total de clastos.

Figura 1. Concepto y medida de los diámetros de un clasto. Izquierda, el eje b en posición; centro, el eje c en posición; derecha, el clasto en perspectiva. Modificado de Pettijohn, 1980 / concept and measurement of diameters of a clast. Left, the axis b in position; center, the axis c in position; right, clast in perspective. Modified from Pettijohn, 1980.
Descripción
El conjunto de clastos asociados al ejemplar MCS-Pv 4 está integrado por un total de 389 elementos (figura 2), en su mayoría de aspecto lustroso. El volumen de la muestra fue de 680 cm3 y su peso total de 1,46 kgr. El guijarro de mayor tamaño alcanzó los 113,7 grs.

Figura 2. Conjunto de guijarros asociados a MCS-Pv 4 / conjunct of pebbles associated to MCS-Pv 4. Escala / scale bar equals 10 cm.
El grado de redondeamiento del conjunto de clastos fluctuó entre el subredondeado y el muy redondeado, de acuerdo con las categorías establecidas por Powers (1953). En cuanto a la distribución de frecuencias de tamaños de los clastos (figura 3), el largo promedio de los ejes mayores (a) fue de 16, 45 mm (d.s ± 8,71). El rango de tamaño de los clastos varió desde los 5,1 mm hasta los 78,9 mm, abarcando un rango granulométrico comprendido entre los tamaño guija a guijón.

Figura 3. Distribución de frecuencias de tamaños de los gastrolitos asociados a MCS-Pv 4 / size distribution of the gastroliths associated to MCS-Pv 4.
El valor de esfericidad (ψ) obtenida (figura 4) arrojó una media de 0,686 (d.s ± 0,108). En cuanto a la forma de los clastos (figura 5), se observó un alto predominio de las formas discoidal y esferoide (40,05% y 34,55% respectivamente) por sobre la laminar (9,16%).

Figura 4. Distribución de frecuencias de esfericidad de los gastrolitos asociados a MCS-Pv 4 / sphericity distribution of the gastroliths associated to MCS-Pv 4.

Figura 5. Distribución de formas de gastrolitos asociados a MCS-Pv 4 según la clasificación de Zingg, donde a, b y c se corresponden con los ejes mayor, intermedio y menor respectivamente. Ambos ejes se encuentran cortados en 2/3 / distribution of the shape of the gastroliths associated to MCS-Pv 4 according to the classification of Zingg, where a, b and c are the major, the medium and the smaller axes. Both axes were sectioned in 2/3.
Composicionalmente, sólo se pudieron reconocer dos categorías litológicas a través de un examen macroscópico y mediante lupa binocular. En general, más del 88 % de los elementos corresponden a clastos de sílice criptocristalino (chert). Tres de los clastos obtenidos son fragmentos de leños fósiles, identificados como gimnospermas (Martínez comunicación personal, 2007). El resto de los elementos rocosos fueron identificados como clastos de sílice cuarzoso. La inspección mediante aumento de los rasgos superficiales de los clastos analizados, ha permitido observar en quince de estos elementos, una serie de marcas o muescas semicirculares de pequeño tamaño; con longitudes promedio de aproximadamente 1 mm (figura 6).

Figura 6. Detalle de superficie de algunos gastrolitos donde pueden observarse diversas marcas semicirculares. / detail of the surface of the gastroliths where semicircular marks can be observed. Escala / scale bar =5 mm.
Discusión
Wings (2004) ha propuesto una serie de criterios tafonómicos y sedimentológicos para la identificación de posibles gastrolitos. Según este autor, los clastos deben ser encontrados acumulados e in situ, ubicados en la región abdominal de esqueletos, relativamente completos y articulados. Si además su composición litológica difiere completamente del sedimento circundante, suman evidencias que permitirían identificarlos como gastrolitos. De esta manera, conforme a los criterios establecidos por Wings (2004), los rasgos observados en los clastos asociados al ejemplar MCS-Pv 4 permitirían adjudicarlos como gastrolitos.
Como se observa en la tabla 1, el número de gastrolitos registrados en distintos ejemplares de plesiosaurios es sumamente variable. No obstante, la gran cantidad de gastrolitos preservados en el MCS-Pv 4 es mucho mayor que en cualquier otro ejemplar de plesiosaurio conocido, con excepción de los 2.626 gastrolitos reportados por Thompson et al. (2004) para un plesiosaurio proveniente del la Península Antarctica (Formación López de Bertodano, Cretácico Superior). Probablemente, el escaso número de gastrolitos en la mayoría de los casos se deba a pérdidas ocurridas con posterioridad a la muerte del animal debido a procesos tafonómicos. De todas maneras, el número de gastrolitos no parece ser, en última instancia, un parámetro relevante en el estudio de formas litófagas como los plesiosaurios, ya que el mismo varía notablemente dentro de una misma especie. La situación se repite en la actualidad, donde individuos de un mismo taxón y grado de desarrollo exhiben una notable diferencia en cuanto a la cantidad de gastrolitos presentes en su tubo digestivo (Wings, 2004).
Tabla 1. Comparación de distintos parámetros obtenidos en gastrolitos de plesiosaurios citados en la literatura / comparison between different parameters obtained in gastroliths of plesiosaurs cited in the literature.
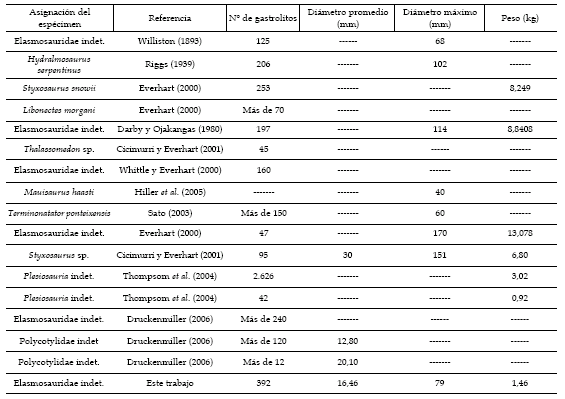
En cuanto al largo del eje mayor (a), el promedio estimado en MCS-Pv 4 es menor al determinado por Cicimurri y Everhart (2001) para un elasmosáurido asignado a Styxosaurus sp. Welles (16,46 contra 30 mm). Lo mismo ocurre con el tamaño del gastrolito más grande (figura 7), cuyo valor (79 mm), es menor al del gastrolito de mayor tamaño que se tiene registro para un plesiosaurio: 170 mm (Everhart, 2000).

Figura 7. Vértebra dorsal de MCS-Pv 4 (1) en comparación con el gastrolito de mayor tamaño (2) / Dorsal vertebra of MCS-Pv 4 (1), compared with the largest gastrolith (2). Escala / scale bar = 1 cm.
De acuerdo con Cicimurri y Everhart (2001), el largo de los ejes medio (b) y menor (c) del gastrolito de mayor tamaño puede indicar la sección transversal máxima de una presa. En el caso del MCS-Pv 4, los ejes medio y menor del gastrolito más largo miden 38,8 mm y 31 mm respectivamente, lo que daría un área transversal de aproximadamente 9,45 cm2. Esta medida indicaría que animal se alimentaba de peces e invertebrados de un tamaño relativamente pequeño.
La esfericidad de los gastrolitos tiene especial importancia en especies litófagas acuáticas, ya que permite inferir el ambiente del cual fueron obtenidos (e ingeridos) los clastos (Darby y Ojakangas, 1980). En principio, aquellos gastrolitos provenientes de un río mostrarán valores de esfericidad mayores que los de una playa marina (Dobkins y Folk, 1970, citado en Darby y Ojakangas, 1980). Asimismo, los clastos fluviales tendrán una esfericidad más variable que los marinos, lo que estará reflejado en una mayor desviación estándar (Dobkins y Folk, 1970). La esfericidad promedio y desviación estándar obtenidas en la muestra alojada en el MCS-Pv 4 (? = 0,686, ds ± 0,108) se corresponden con los índices tabulados para cantos de ríos (ψ = 0,684, ds ± 0,105) (Dobkins y Folk, 1970.). Estos valores sugieren que los clastos asociados al MCS-Pv 4 corresponderían a clastos de depósitos fluviales, de forma similar a los gastrolitos reportados por Darby y Ojakangas (1980) para un plesiosaurio del Campaniano de Bearpaw Shale (Montana, Estados Unidos). El probable paleoambiente de la Formación Allen en la zona de Lago Pellegrini es perfectamente compatible con esta posibilidad, ya que los niveles bentoníticos en los cuales fue hallado el ejemplar MCS-Pv 4 fueron depositados en una zona intermareal, próxima a la costa (Gasparini et al., 2007).
La condición subadulta de MCS-Pv 4, así como las características osteohistológicas de un segundo Elasmosauridae indet. con la misma morfología y probablemente correspondiente a un mismo taxón (MUCPv-92) (Gasparini et al., 2007), también son coincidentes con el ambiente propuesto para la Formación Allen, en esa localidad. Efectivamente, el ejemplar MUCPv-92, al igual que otros especimenes subadultos de plesiosaurios (Wiffen et al., 1995), muestra una organización osteohistológica de tipo pachyosteosclerótica, común en formas pobremente adaptadas a la natación rápida y sostenida, que viven en ambientes marinos poco profundos. Este segundo espécimen, sobre el cual se realizó el estudio paleohistológico (Salgado et al., 2007), no preservó gastrolitos.
Cicimurri y Everhart (2001) han referido que los gastrolitos de Styxosaurus sp. se clasifican en tres categorías principales en cuanto a su geometría: oblada (= discoidal), equidimensional (= esferoide) y prolada (= cilíndrica); aunque estos autores no mencionan los índices con los cuales los guijarros fueron agrupados en esas categorías.Por otro lado, en cuanto a su forma, los gastrolitos descriptos por Darby y Ojakangas (1980, figura 4) se asemejan notablemente a los de MCS-Pv 4 (figura 5), con una supremacía de los guijarros discoidales y esferoides. El predominio de las formas discoidales y esferoides, y un índice de esfericidad relativamente alto, como el observado en los gastrolitos del ejemplar patagónico y en los del ejemplar de Bearpaw Shale descritos por Darby y Ojakangas (1980), sugiere que el organismo pudo obtener sus gastrolitos en un ambiente de tipo fluvial distal.
Con respecto al conjunto de gastrolitos asociados al MCS-Pv 4, tanto el peso total de la muestra (1.460 grs.), como el del guijarro de mayor tamaño (113,7 grs.), son significativamente menores que aquellos reportados por Everhart (2000) (13,078 kgs. y 1,49 kgs., respectivamente). Esto último podría obedecer a los diferentes tamaños que presentan ambos especimenes (9 metros en el ejemplar reportado por Everhart y 7 metros para el MCS-Pv 4).
Probable función de los gastrolitos
En reptiles marinos, existen dos hipótesis principales sobre la posible función de los gastrolitos. Por un lado, algunos autores como Darby y Ojakangas (1980) y Taylor (1993), sostienen que los mismos pudieron haber funcionado como lastre, incrementando la gravedad específica del animal, o bien ayudando a su estabilidad durante el desplazamiento acuático. Otros sugieren que, al igual que en las aves actuales, los gastrolitos habrían sido utilizados como auxiliares durante el procesamiento mecánico del alimento (Williston, 1894; Brown, 1904; Cicimurri y Everhart, 2001; Mc Henry et al., 2005).
En cuanto a la hipótesis hidrostática, lo que en primer lugar debe tenerse en cuenta es la relación entre el peso de los gastrolitos y la masa corporal del animal, y luego juzgar si aquel puede influir significativamente en su flotabilidad. Para estimar la masa corporal del MCS-Pv 4, se debió primero determinar su longitud corporal. A partir del largo promedio de sus vértebras dorsales (4,5 cm) y asumiendo una semejanza morfológica básica con Elasmosaurus platyurus Cope, cuyas vértebras dorsales poseen un largo promedio de 8,9 cm (Everhart, 2005) y para cuyo ejemplar se calculó largo corporal de aproximadamente 14 m; se estima una longitud de 7 m para el ejemplar MCS-Pv 4. Este cálculo fue utilizado posteriormente para inferir la masa corporal del espécimen de Lago Pellegrini, a partir de las relaciones de longitud y masa para elasmosáuridos tabuladas por Everhart (2000). La masa corporal estimada es de 1.307 kgs.
La relación entre el peso total de los gastrolitos (cuya cantidad, rango de tamaños y tafonomía sugieren fuertemente que todas las rocas ingeridas por el animal al momento de morir están representadas en la muestra), y la masa corporal estimada del animal, es algo mayor al 0,1%. Por lo tanto, dicho porcentaje no pudo afectar significativamente la flotabilidad del animal, ya que, para que esto ocurra, el peso de la masa de gastrolitos debe ser superior 4 % como indican algunos modelos computarizados realizados para plesiosaurios (Henderson, 2005). Esta conclusión coincide con la de Everhart (2000) y Henderson (2005), quienes entienden que las posibilidades de que los gastrolitos de los plesiosaurios hayan sido utilizados como lastre son poco probables.
Con respecto a la hipótesis digestiva, para que los alimentos puedan ser macerados por el conjunto de gastrolitos, es necesario que exista un importante movimiento dentro del órgano que alojaba dichos elementos líticos. Tal como se describiera anteriormente, sobre la superficie de diversos gastrolitos pudo observarse una serie de marcas o muescas semicirculares (figura 6), muy similares a las descriptas por Everhart (2005) para un conjunto de supuestos gastrolitos. Este tipo de marcas pudo originarse a partir de movimientos rotatorios de los gastrolitos dentro del tracto digestivo, generando efectos de atrición y choque entre los mismos. Por otro lado, cuatro de los gastrolitos pertenecientes a la muestra, los cuales en primera instancia fueron contados como piezas distintas, resultaron ser las mitades de dos gastrolitos fracturados. Llamativamente, los bordes de fractura de esos fragmentos no son agudos, sino que, por el contrario, están notablemente desgastados y redondeados (figura 8). Nuestra interpretación es que el gastrolito fue quebrado dentro del animal y el constante rozamiento con otras superficies duras provocó el posterior desgaste de los bordes. Si se tiene en cuenta la gran dureza de estos clastos, compuestos mayormente por sílice, se puede inferir que los mismos estuvieron sometidos a un constante movimiento durante un lapso de tiempo más o menos prolongado. Estos hechos e interpretaciones sugieren que, de las dos hipótesis, aquella más aceptable es la que adjudica a los gastrolitos una función digestiva. Sin embargo, para poder corroborar esta hipótesis, deberían investigarse otros aspectos de la biología del animal, como su locomoción, su alimentación, entre otros. Además, es fundamental estudiar con mayor profundidad el comportamiento litófago de ciertos grupos actuales, en particular aquellos con hábitos acuáticos marinos (como los pinnípedos y las aves esfenisciformes), para poder realizar comparaciones y obtener conclusiones más fundamentadas.

Figura 8. Uno de los gastrolitos quebrados. 1, 2, ambas mitades del gastrolito en vista interna, donde puede verse el grado de desgaste y pulimento interno. 3, ambas mitades unidas. Se observa los bordes notablemente desgastados / one of the broken gastroliths. 1, 2, both halves of the gastrolith in internal view, where the worn and polished internal surface can be observed. 3, Both halves united. The corroded borders can be observed. Escala / scale bar = 1 cm.
Agradecimientos
Este trabajo fue financiado parcialmente por la National Geographic Society (grant 7757/04 a Z. Gasparini) y la Agencia de Promoción de Ciencia y Técnica (PICT- 25276 a Z. Gasparini). A. Garrido hizo útiles comentarios sobre la litología de los gastrolitos. L. Martínez identifico los leños fósiles presentes en la muestra. La asociación del Museo de Cinco Saltos ofreció su colaboración durante la extracción y preparación del material. Z. Gasparini y M. Fenández brindaron una constructiva revisión del manuscrito. Las mediciones de peso y volumen fueron realizadas en instalaciones del CEM nº 1 "Armando Novelli".
Bibliografía
1. Blatt, H., Middleton, G. y Murray, R. 1972. Origin of Sedimentary Rocks. Prentice-Hall, New Yersey, 634 pp. [ Links ]
2. Ballent, S.C. 1980. Ostrácodos de ambiente salobre de la Formación Allen (Cretácico Superior) en la Provincia de Río Negro (Republica Argentina). Ameghinana 17: 67-82. [ Links ]
3. Brown, B. 1904. Stomach stones and food of plesiosaurs. Science 20: 184-185. [ Links ]
4. Cicimurri, D.J. y Everhart, M.J. 2001. An elasmosaur with stomach contents and gastroliths from the Pierre Shale (Late Cretaceous) of Kansas. Transactions of the Kansas Academy of Science 104: 129-143. [ Links ]
5. Cope, E.D. 1869. Synopsis of the Extinct Batrachia and Reptilia of North America, Part I. Transactions of the American Philosophical Society, New Series, 14: 1-235. [ Links ]
6. Darby, D.G. y Ojakangas, R.W. 1980. Gastroliths from an Upper Cretaceous Plesiosaur. Journal of Paleontology 54: 548-556. [ Links ]
7. Druckenmiller, P.S. 2006. [Early Cretaceous plesiosaurs (Saurpterygia: Plesiosauria) from northern Alberta: palaeoenvironmental and systematic implications. Ph.D. Thesis, University of Calgary, 558 pp. Inédito.]. [ Links ]
8. Everhart, M. J. 2000. Gastroliths associated with plesiosaur remains in the Sharon Springs Member of the Pierre Shale (Late Cretaceous), Western Kansas. Transactions of the Kansas Academy of Science 103: 58-69. [ Links ]
9. Everhart, M.J. 2004. Plesiosaurs as the food of mosasaurs; new data on the stomach contents of a Tylosaurus proriger (Squamata; Mosa-sauridae) from the Niobrara Formation of western Kansas. The Mosasaur 7: 41-46. [ Links ]
10. Everhart, M.J. 2005. Elasmosaurid remain from the Pierre shale (Upper Cre-taceous) of estern Kansas. Possible missing elements of the type of Elasmosaurus platyurus Cope 1868?. http://www.Palarch.-nl/Paleontology/paleontology, 4,3,:19-32 (3 de agosto). [ Links ]
11. Gasparini, Z. y Salgado, L. 2000. Elasmosáuridos (Plesiosauria) del Cretácico tardío del norte de Patagonia. Revista Española de Paleontología 15: 13-21. [ Links ]
12. Gasparini, Z., Salgado, L. y Parras, A. 2007. Late Cretaceous plesiosaurs from northern Patagonia, Argentina. Geological Journal 42: 185-202. [ Links ]
13. Henderson, D.M. 2005. Floating point: a computational study of buoyancy, equilibrium, and gastroliths in plesiosaurs. Journal of Vertebrate Paleontology, supplement to volume 25: 68-69A. [ Links ]
14. Hiller, N., Mannering, A.A., Jones, C.M. y Cruickshank, A.R.I. 2005. The nature of Mauisaurus haasti Hector, 1874 (Reptilia: Plesiosauria). Journal of Vertebrate Paleontology 25: 588-601. [ Links ]
15. Kobayashi, Y., Lü, J.-C., Dong, Z.-M., Barsbold, R., Azuma, Y. y Tomida, Y. 1999. Herbivorous diet in an ornithomimid dinosaur. Nature 402: 480-481. [ Links ]
16. McHenry, C.R., Cook, A.G. y Wroe, S. 2005. Bottom-feeding plesiosaurs. Science 310: 75. [ Links ]
17. O'Keefe, F.R. 2001. A cladistic analysis and taxonomic revision of the Plesiosauria (Reptilia: Sauropterygia). Acta Zoologica Fennica 213: 1-63. [ Links ]
18. Pettijohn, F.J. 1980. Rocas Sedimentarias. EUDEBA, Buenos Aires, 731 pp. [ Links ]
19. Powers, M.C. 1953. A new roundness scale for sedimentary particles. Journal of Sedimentary Petrology 23: 117-119. [ Links ]
20. Riggs, E. 1939. A specimen of Elasmosaurus serpentinus. Geological Series of Field Museum of Natural History 6: 385-391. [ Links ]
21. Salgado, L., Fernández, M. y Talevi, M. 2007. Observaciones histológicas en reptiles marinos (Elasmosauridae y Mosasauridae) del Cretácico Tardío de Patagonia y Antártida. Ameghiniana 44: 513-523. [ Links ]
22. Sanders, F., Manley, K. y Carpenter, K. 2001. Gastroliths from the Lower Cretaceous sauropod Cedarosaurus weiskopfae. En: T. Tanke, K. Carpenter y M.W. Skrepnick, (eds.), Mesozoic Vertebrate Life, Indiana University Press, Indiana, pp. 166-180. [ Links ]
23. Sato, T. 2003. Terminonator ponteixensis, a new elasmosaur (Reptilia, Sauropterygia) from the Upper Cretaceous of Saskatchenwan. Journal of Vertebrate Paleontology 23: 83-103. [ Links ]
24. Sato, T. y Storrs, G.W. 2000. An early policotylid plesiosaur (Reptilia, Sauropterigya) from the Cretaceous of Hokkaido, Japan. Journal of Paleontology 61: 1242-1246. [ Links ]
25. Schmeisser, R., Gillette, D.D., Albright, L.B. y Titus, A.L. 2005. Plesiosaur gastroliths from the tropic shale (Cenomanian-Turonian) of Southern Utah. Geological Society of America, Abstracts with Programs 67: p. 132. [ Links ]
26. Storrs, G.W. 1993. Function and phylogeny in sauropterygian (Diapsida) evolution. American Journal of Science 293-A: 63-90. [ Links ]
27. Taylor, M.A. 1993. Stomach stones for feeding or buoyancy? The occurrence and function of gastroliths in marine tetrapods. Royal Society Philosophical Transactions, Biological Sciences 1296: 163-175 [ Links ]
28. Thompsom, W.A., Martin, J.E. y Reguero, M. 2004. Comparison of gastroliths within plesiosaurs from the Late Cretaceous marine deposits of Vega Island, Antartic Peninsula, and The Missouri River Area, South Dakota. Geological Society of America, Abstracts with Programs 36: p. 80. [ Links ]
29. Whittle, C.H. y Everhart, M.J. 2000. Apparent and implied evolutionary trends in lithophagic vertebrates from New Mexico and elsewhere. En: S.G. Lucas y A.B.Heckert (eds.), Dinosaurs of New Mexico, New Mexico Museum of Natural History and Science Bulletin 17: pp. 75-82. [ Links ]
30. Williston, S.W. 1893. An interesting food habit of the plesiosaurs. Transactions of the Kansas Academy of Science 13: 121-122. [ Links ]
31. Williston, S.W. 1894. A food habit of the plesiosaurs. The American Naturalist 28: 50. [ Links ]
32. Williston, S.W. 1904. The stomach stones of the plesiosaurs. Science 20: 565. [ Links ]
33. Wings, O. 2004. [Identification, Distribution, and Function of Gastroliths in Dinosaurs and Extant Birds with Emphasis on Ostriches (Struthio camelus). Ph.D. Thesis, University of Bonn, Bonn, 187 pp. Inédito.]. [ Links ]
34. Wiffen, J., de Buffrenil, V., de Ricqlès, A. y Mazin, J.-M. 1995. Ontogenetic evolution of bone structure in the late cretaceous plesiosauria from New Zealand. Geobios 28: 625-640. [ Links ]
Recibido: 23 de junio de 2006.
Aceptado: 2 de junio de 2008.

