










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.45 n.3 Buenos Aires jul./sep. 2008
Revision of Parastrapotherium (Mammalia, Astrapotheria) and other Deseadan astrapotheres of Patagonia
Alejandro G. Kramarz1 and Mariano Bond2
1Consejo Nacional de Investigaciones Científicas y Técnicas. Sección Paleontología de Vertebrados. Museo Argentino de Ciencias Naturales Bernardino Rivadavia. Av. Angel Gallardo 470, 1405 Buenos Aires, Argentina. agkramarz@macn.gov.ar
2Consejo Nacional de Investigaciones Científicas y Técnicas. Departamento Paleontología de Vertebrados. Museo de La Plata. Paseo del Bosque s/nro, 1900 La Plata, Argentina.
Abstract. Parastrapotherium Ameghino, 1895 is one of the most characteristic faunal elements of the Deseadan South American Land Mammal Age (Late Oligocene) of Patagonia, and seven species have been described for that Age. The genus was traditionally differentiated from Astrapotherium by its giant size and by having a higher number of upper and lower premolars. The revision of the dentition of Parastrapotherium revealed that this genus has only one more lower premolar than Astrapotherium. Other differential features are: lower crowned cheek teeth, cingulum absent at base of labial fold in all upper cheek teeth, upper premolars with labial fold less prominent and with broader base, crown of lower incisors proportionally more robust and buccolingually broader, lower molars with bunoid "pillar", lingual cingulum absent at the base of metaconid, and lower cheek teeth with very superficial labial flexid. Of the described Deseadan species, P. holmbergi and P. martiale Ameghino are herein interpreted as valid. The validity of P. ephebicum Ameghino is uncertain. Astrapotherium voghti Mercerat is valid and herein provisionally transferred to Parastrapotherium. Liarthrus copei Ameghino, previously considered as synonym of P. holmbergi, should be kept in a separate genus. The phylogenetic relationships of Parastrapotherium are still uncertain, because it has some features suggesting uruguaytheriine affinities (lower molar cingulum absent at the base of metaconid and very superficial labial flexids), whereas the dental formula and the degree of hypsodonty suggest a very basal position among the Oligocene - Miocene astrapotheres. Cladistic analysis is pending on further additional studies on the still poorly known Astraponotus and other astrapotheriids.
Resumen. Revisión de Parastrapotherium (Mammalia, Astrapotheria) y otros astrapoterios Deseadenses de Patagonia. Parastrapotherium Ameghino, 1895 es uno de los elementos faunístico más característicos de la Edad Mamífero Deseadense (Oligoceno tardío) de Patagonia, y siete especies fueron descriptas para esa Edad. El género fué tradicionalmente diferenciado de Astrapotherium por su tamaño gigantesco y por tener mayor cantidad de premolares superiores e inferiores. La revision de la dentadura de Parastrapotherium reveló que este género sólo presenta un premolar inferior más que Astrapotherium. Otros rasgos distintivos son: molarifirmes de corona más baja, cíngulo ausente en la base del pliegue labial de los molariformes superiores, premolares superiores con pliegue labial menos prominente y más ancho en su base, incisivos de corona proporcionalmente más robusta y bucolingualmente más ancha, molares inferiores con "pillar" bunoide, cíngulo lingual ausente en la base del metacónido y molariformes inferiores con fléxido labial muy superficial. De las especies deseadenses decriptas, se interpretan aquí como válidas a P. holmbergi y P. martiale Ameghino. La validez de P. ephebicum Ameghino es incierta. Astrapotherium voghti Mercerat es también válida y transferida aquí tentativamente a Parastrapotherium. Liarthrus copei Ameghino, previamente considerada sinónima de P. holmbergi, debería ser mantenida en un género aparte. Las relaciones filogenéticas de Parastrapotherium aún son inciertas, dado que presenta algunos caracteres que sugieren afinidades con los uruguayterinos (cíngulo de los molares inferiores ausente en la base del metacónido y fléxidos labiales muy superficiales), mientras que la formula dentaria y el grado de hipsodoncia sugieren una posición muy basal entre los astrapoterios oligomiocénicos. Un análisis cladístico está pendiente de nuevos estudios adicionales sobre el aún pobremente conocido Astraponotus y otros astrapotéridos.
Key words. Parastrapotherium; Astrapotherium; Liarthrus; Systematics; Oligocene; Patagonia.
Palabras clave. Parastrapotherium; Astrapotherium; Liarthrus; Sistemática; Oligoceno; Patagonia.
Introduction
Parastrapotherium Ameghino, 1895 is a conspicuous representative of the Patagonian Deseadan (Late Oligocene) paleomastozoological fauna, though it has also been reported for Colhuehuapian (Early Miocene) levels (Ameghino, 1902; Scott, 1937; Kramarz et al., 2005; Kramarz and Bond, in press). This genus includes some of the largest known astrapotheres, even larger than those described for the middle Miocene of La Venta, Colombia (Johnson and Madden, 1997), although several small to minute species have been also referred to Parastrapotherium (Ameghino, 1895, 1902).
Ameghino (1895) first described this genus based upon materials collected by Carlos Ameghino, at present housed at the MACN Collections. According to Ameghino, Parastrapotherium differs from the better-known Astrapotherium Burmeister from the Santa Cruz Formation (Santacrucian Age, late Early Miocene) basically by having higher number of upper and lower premolars. Gaudry (1904) interpreted the Deseadan astrapotheres as cogeneric with the Santacrucian species. Loomis (1914) described a few materials from the Deseadan Cabeza Blanca locality, considered Parastrapotherium as different from Astrapotherium, and accepted four of the eight species described by Ameghino (1889, 1895, 1901) as valid, but without direct comparisons with the type materials. Scott (1937) redescribed Parastrapotherium based on more complete materials housed at FMNH. Scott considered that most of the interpretations made by Ameghino on the basis of very fragmentary materials, were wrong in light of latter discoveries, but he agreed with Ameghino in distinguishing between the two taxa, with Parastrapotherium having a higher number of premolars than Astrapotherium. Kramarz and Bond (in press) revised the species of Parastrapotherium recognised by Ameghino (1899, 1902) for his "couches à Colpodon" (Colhuehuapian Age) and concluded that some should be transferred to Astrapotherium. However, at present there is no adequate characterisation of this genus to allow clear differentiation from other astrapotheres, except for its giant size. In this contribution the genus Parastrapotherium is re-examined and the dental characters that differentiate this genus from Astrapotherium and other astrapotheriids are evaluated, in order to propose an adequate diagnosis. The type specimens of all the Deseadan species referred to Parastrapotherium and other related genera are revised, and their validity and corresponding stratigraphic distributions are herein discussed.
Abbreviations. AMNH, American Museum of Natural History (USA); FMNH, Field Museum of Natural History (USA); MACN, Museo Argentino de Ciencias Naturales; MLP, Museo de La Plata; MPEF, Museo Paleontológico Egidio Feruglio (Trelew, Chubut Province); SALMA, South American Land Mammal Age.
Systematic paleontology
Order Astrapotheria Lydekker, 1894
Family Astrapotheriidae Ameghino, 1887
Parastrapotherium Ameghino, 1895
Traspoatherium Ameghino, 1895: 641.
Loxocoelus Ameghino, 1895: 653 (partim).
Helicolophodon Roth, 1903: 143.
Type species. Parastrapotherium holmbergi Ameghino, 1895
Geographic and stratigraphic distribution. Sarmiento Formation, Deseadan Age (Late Oligocene), Chubut and Santa Cruz provinces, and Colhuehuapian Age (Early Miocene) (Flynn y Swisher, 1995), Chubut province; Cerro Bandera Formation, Colhuehuapian Age (Kramarz et al., 2005), Neuquén province, Argentina.
Diagnosis. Large astrapotheriid, size ranging from comparable to Astrapotherium magnum, to larger than Astrapotherium giganteum. Dental formula in adult stages I?/3, C1/1, P2/2, M3/3. Very developed hypsodont canines; the lower ones strongly extroverted and rooted in adults, as in Astrapotherium. Crowns of cheek teeth lower than in Astrapotherium but higher than in Astraponotus. Upper premolars with labial fold less prominent and broader at the base than in Astrapotherium and Astrapothericulus. Labial cingula absent or reduced at the base of the labial fold in all upper cheek teeth. Crista of M2 isolated from the crochet during early and moderate stages of wear. Crown of lower incisors with base buccolingually much broader than in Astrapotherium. Lower cheek teeth with labial flexid shallower than in Astrapotherium. Lower molars with bunoid "pillar", as in Astrapothericulus.
Comparative description
Dentition. Ameghino (1895) described some isolated teeth and interpreted them as probable upper incisors of Parastrapotherium. The only available premaxillaries of this genus are incomplete anteriorly, and therefore it is not possible to unquestionably determine the presence of such teeth. All other known Astrapotheriidae with preserved premaxillaries lack incisors, suggesting that the same condition occurred in Parastrapotherium.
The upper canines are recurved, columnar, evergrowing as in Astrapotherium (Scott, 1928). Ameghino (1897) and Loomis (1914) pointed out that both the upper and lower canines of Parastrapotherium are comparatively smaller than in Astrapotherium. However, the size of the canines is very variable due to sexual dimorphism (Scott, 1937), though there is also an important ontogenetic component to this variability. The size of the canines is not a differential feature between Astrapotherium and Parastrapotherium. Juvenile upper canines are sharp-pointed, isosceles triangular in cross section, with the smaller side forming the anterior wall and the lower angle in the posterior side of the tooth. The enamel is continuous around the entire crown. The anteroposterior and transverse diameters increase with tooth growth. The enamel first disappears from the anterior wall, which generally exhibits a longitudinal groove that becomes deeper toward the base. A second groove is present on the lingual wall, generally on the anterior half of this surface. In adult stages the enamel is also reduced on the posterior angle and later on the lateral walls, though in some specimens the enamel persists on the labial wall even longer. At this stage the lateral walls become slightly convex, so that the tooth acquires a characteristic heart-like cross section (figure 1). This ontogenetic series does not differ from that known for Astrapotherium. The tip of the canine is strongly bevelled due to a conspicuous wear facet developed on the apex of the anterior surface, oblique with respect to the tooth axis. Some specimens show an accessory wear facet on the apex of the posterior angle of the tooth. This feature is also observed in Astrapotherium and Astrapothericulus Ameghino ("Astrapothericulan" stage, late Early Miocene), but its functional meaning is still unknown. The diastema separating the canine from the premolars is slightly larger than the entire P3-P4 length.

Figure 1. Parastrapotherium martiale, MACN A 52-604 (holotype/ holotipo). Left upper canine in schematic cross section / canino superior izquierdo en corte transversal esquemático.
All cheek teeth are brachydont, with high degree of unilateral hypsodonty, particularly the molars. The crowns are comparatively lower than in Astrapotherium, but higher than in Astraponotus Ameghino (Mustersan).
The anteriormost permanent premolar is the P3 (Gaudry, 1904; Loomis, 1914). Ameghino (1985) interpreted Parastrapotherium as having a complete premolar series, or at most with only P1 absent. His interpretation was based upon the specimen MACN A 52-509a, a maxillary fragment with two premolars (figure 2), which Ameghino considered as P1-P2 or P2-P3, because they are implanted very near to what he interpreted as the preserved posterior margin of the alveolus for the canine. However, this margin corresponds to the preserved dorso-mesial wall of the choanae, and in fact only the posterior portion of the diastema is preserved. Scott (1937) pointed out that Parastrapotherium has three upper premolars, and that the P2 is very small and frequently lost in full-grown skulls. Nevertheless, all the examined specimens lack P2 or an alveolus for this tooth, including the FMNH specimens listed and measured by Scott.

Figure 2. Parastrapotherium holmbergi, MACN A 52-509a (syntype/ sintipo). Right P3-P4. 1, in occlusal view/ en vista oclusal; 2, in labial view/ en vista labial.
P4 differs from P3 (figure 2) in being nearly 50% larger and with a more rectangular outline. Both have a single lingual conical cusp (protocone) and a labial wall (ectoloph); the anterior portion of this wall has a lingual projection that reaches the base of the protocone, forming a rudimentary protoloph. The posterior portion of the ectoloph shows a lingual projection, as a rudimentary metaloph, lower than the anterior one. In P3 the metaloph does not reach the base of the protocone, thus the central valley is open posterolingually. In P4 the metaloph is slightly more developed and reaches the base of the protocone; therefore the central valley is posterolingually closed at advanced stages of wear. The labial face of the ectoloph has a single vertical fold (figure 2.2), less prominent than in Astrapotherium and Astrapothericulus; this fold gradually broadens toward the base, much more so than in the latter genera. The base of the labial fold is comparatively broader in P4 than in P3, and occupies the labial base of the crown almost entirely (figure 2.2), in contrast to Astrapotherium and Astrapothericulus. The basal cingula are more marked than in Astrapotherium, but less than in Astrapothericulus. In contrast to both genera, the cingulum is absent or very reduced at the base of the labial fold (figure 2.2). The wear is more pronounced on the labial portion of the P3 and produces a large wear facet on the buccal surface of the ectoloph (figure 2.1), probably due to occlusion with the p3. This pattern is different from that of Astrapotherium and Astrapothericulus, which lack a p3. Both P3 and P4 have a small anterolabial cylindrical root and a large posterolingual root, which has a very deep vertical cleft on the anterior wall, and a much shallower cleft on the posterior one. Ameghino (1895, 1901, 1902) interpreted this feature as the result of the fusion of two individual roots. Astrapotherium and Astrapothericulus show the same condition, but the cleft on the posterior face of the posterolingual root is absent. The roots are generally divergent, but in some specimens they are parallel or convergent, and fused near the base of the crown or along their entire length. Such variations, interpreted by Ameghino as specific differences, are also observed among co-specific individuals in extant ungulates (e.g. Tayassu), and they would result from ontogenetic variations (Loomis, 1914), and due to the individual variations of available intralveolar space during formation of roots. Therefore, we interpret these variations of roots disposition as having no taxonomic value. M2 is nearly 30% larger than M1 (see table 1) and with less rectangular outline. Both show basically the same occlusal structure as in Astrapotherium and Astrapothericulus: the ectoloph forms the entire labial wall of the tooth, both parastyle and metastyle are well developed, the labial fold of the paracone is very conspicuous, the protoloph is wide and connects protocone with paracone, crista and crochet are present and delimit a small, ephemeral median fossette, the metaloph is narrow and connects metacone with hypocone, and the posterior cingulum is high and encloses the small posterofossette (figure 3.1). However, the protocone is more conical (walls are less vertical) and the central valley remains wider until more advanced stages of wear than in Astrapotherium, (compare figure 3.2 with 3.5). The triangular areas of exposed dentine at the base of the anterior and posterior faces of the tooth are comparatively lower than in Astrapotherium. The posterior end of the crista remains isolated from the crochet at more advanced stages of wear (figure 3.1). These characters would be related to a lesser degree of hypsodonty in Parastrapotherium. Moreover, the inflection of the anterior wall of the protoloph is not as penetrating and the base of the hypocone is more lingually pointed than in Astrapotherium. As in the premolars, the labial cingulum is interrupted at the base of the labial fold, in opposition to Astrapotherium.
Table 1. Compared dental measurements for the type specimens of the Deseadan species of Parastrapotherium, Traspoatherium convexidens, and Loxocoelus obturtus, in centimetres. Abbreviations: BL, bucco-lingual width; MD, mesio-distal length; TRW, transverse width / medidas dentarias comparadas de las especies deseadenses de Parastrapotherium, Traspoatherium convexidens y Loxocoelus obturtus, en centímetros. Abreviaturas: BL, ancho buco-lingual; MD, longitud mesio-distal; TRW, ancho transverso.


Figure 3. 1-2, Compared upper molars of Parastrapotherium holmbergi spp. and Astrapotherium magnum / molares superiores compardos de Parastrapotherium holmbergi spp. y Astrapotherium magnum. 1, MACN A 52-509b, P. holmbergi , M2 in occlusal view / en vista oclusal; 2, same in lingual view / el mismo en vista lingual; 3, MACN A 52-425, P. martiale, M3 in occlusal view / en vista oclusal; 4, MACN A 3223, A. magnum, M2 in occlusal view/ en vista oclusal; 5, same in lingual view / el mismo en vista lingual; 6, MLP 12-103, A. magnum, erupting M3, in occlusal view / M3 en erupción, en vista oclusal.
The M3 (figure 3.3) differs from that of Astrapotherium by the same character related to hypsodonty mentioned above for M1 and M2. Scott (1937) noted that in Parastrapotherium the crista of M3 projects inward nearly reaching the protocone, whereas this element is longer, recurved and parallel to the protoloph in Astrapotherium. Such difference is due to the fact that the crista in Parastrapotherium rises from the central portion of the ectoloph, instead of from the metacone as in Astrapotherium (figure 3.6), and the anterior portion of the central valley is wider.
The three lower incisors, not described by Scott (1937), have shovel-like crowns and are implanted radially and almost horizontally. They are propor-tionally larger and the base is anteroposteriorly much broader than in the remaining Astrapotheriidae (figure 4). The enamel covers the crowns except in the triangular areas at the base of the lateral walls (figure 4.2). Unworn incisors have a deep notch at their end that is worn away. However, some worn specimens show an apical notch secondary produced by the wear (figure 4.3); this feature is also observed in some specimens referred to Astrapotherium and Astrapothericulus. The lingual face is slightly concave and has a medial longitudinal furrow, but a prominent crest replaces the furrow in at least one Miocene species (Kramarz and Bond, in press). There is a conspicuous V-shaped cingulum on the base, crenulated in some specimens. The labial face is convex and has a medial longitudinal furrow shallower than the lingual one. The labial cingulum is less prominent, and in some specimens continuous with the lingual cingulum. The i1 and i2 have similar shape; i2 is larger and with relatively higher crown. The crown of i3 is lower and mesiodistally wider than that of i2.

Figure 4. Incisors of Parastrapotherium / insicivos de Parastrapotherium. 1, MACN A 52-515, i2? in lingual view / en vista lingual; 2, same in lateral view / el mismo en vista lateral. 3, MACN A 52-515, i3 in lingual view / en vista lingual.
Lower canines are also hypsodont but rooted in adult stages. Scott (1937) indicated that the lower canines of Astrapotherium are evergrowing. However, adult specimens referable to A. magnum (MACN Pv 14512) from Santa Cruz Formation have rooted lower canines, as in Parastrapotherium. As in the upper ones, size, shape, and implantation of the lower canines vary depending on the sex (Scott, 1937) and ontogenetic development. When erupted, the canines are curved backward, pointed, and compressed; the labial wall is more convex than the lingual one, and both form an anterior cutting edge. The enamel covering is continuous around the entire crown. During growth, the crown becomes more compressed, and a longitudinal depression appears at the anterior portion of the lingual face, gradually becoming deeper and wider toward the base. The enamel is excluded from the anterior and posterior faces. In adults the enamel is also excluded from the lateral walls. A wear facet is present at the tip of the posterior margin; some specimens have also an accessory, generally irregular (rippled) wear facet on the anterior edge. The trefoil cross-section indicated by Scott (1937) is not present in any of the specimens examined, and none have significant differences with that of Astrapotherium.
The anteriormost permanent lower premolar is the p3 (Gaudry, 1904; Scott, 1937). Loomis (1914) interpreted that this tooth fall out in old individuals, but at least in the adult (not senile) specimen AMNH 29573 the p3 is still in position. Other observed adult individuals lack p3 but preserved it alveolous. The diastema between canine and p3 is nearly as long as the entire m1-m2 length. In P. holmbergi the p3 is single rooted, but some specimens still retain a vertical sulcus on the lingual side. At least in the holotype of P. martiale the p3 is clearly two rooted. In both species the crown has a main central conical cusp with accessory anterior and posterior cusps (figure 5.1-5.4). The posterior one is connected with the posterolabial wall of the central cusp through a labial, crescentic crest, which delimits a small lingual valley (metaflexid). The anterior cusp varies according to the species. In specimens referred to P. holmbergi (figure 5.1-3) it is a small basal cuspid attached to the anterior slope of the central cusp. In P. martiale (figure 5.4) the anterior cusp is as developed as the posterior one, with a short labial crescent that contacts the anterolabial face of the central cusp, delimiting an anterolingual valley, so that the tooth has a more molariform aspect than in P. holmbergi. In both species the base of the lingual wall of the tooth shows a continuous cingulum connecting the bases of the anterior and posterior cusps. The labial cingulum is present but less marked than the lingual one. The labial wall of the tooth is strongly convex and lacks labial flexid. Scott (1937) stated that the p3 of Parastrapotherium is similar to the p2 of Albertogaudrya Ameghino (Casamayoran); however, the presence of an anterior cusp and a well-developed posterior crescent make it more similar to the p3 of this genus, whereas it only shares with the p2 the absence of labial flexid.

Figure 5. Compared lower premolars of Parastrapotherium spp. and Astrapotherium magnum / premolares inferiores comparados de Parastrapotherium spp. y Astrapotherium magnum. 1, MACN A 52-506, P. holmbergi, right p3 in occlusal view / p3 derecho en vista oclusal; 2, same in lingual view / el mismo en vista lingual; 3, idem, in labial view/ el mismo en vista labial; 4, MACN A 52-604, P. martiale (holotype/ holotipo), right p3 - p4/ p3 - p4 derechos; 5, MACN A 52-506, P. holmbergi, left p4 (reversed) / p4 izquierdo (invertido); 6, MACN A 3230, A. magnum, left p4 (reversed)/ p4 izquierdo (invertido). Arrows indicate anterior and lingual sides / las flechas indican los lados anterior y lingual.
The p4 (figure 5.4-5) is nearly 80% larger than p3 (see table 1). The central cusp is more columnar; the anterior and posterior cusps and their corresponding crescents are more developed and the lingual valleys are deeper and more penetrating, resembling the molars; the anterior crescent is higher and shorter than the posterior one. The labial flexid is absent or hardly insinuated, as in Astrapothericulus, but it is much shallower than in the p4 of Astrapotherium (figure 5.6). Some specimens show a small excrescence at the base of the posterior slope of the central cusp, resembling the "pillar" of the molars (figure 5.4). The wear is more intense on the talonid, thus the trigonid remains higher even at advanced stages of wear. The p4 have two slightly divergent roots, although in some specimens the roots are parallel and fused along their lingual margins, retaining a vertical labial sulcus. As in the upper premolars, such variations have no taxonomic value.
The lower molars are typically "bicrescentic", with well-developed metaflexid and paraflexid, of which the former is deeper and more penetrating. The hy-poflexid is represented by a slight inflection of the labial wall (figure 6.1), little more marked than in Uruguaytherium Kraglievich (Deseadan? from Uruguay) and Xenastrapotherium Kraglievich (Deseadan? -Laventan from Venezuela, Colombia, Ecuador, and Brazil). On the contrary, the hypoflexid is a deep cleft in Astrapotherium (figure 6.3) and Astrapothericulus. The crowns have columnar appearance less and the lingual valleys are wider than in Astrapotherium. The metaflexid is U-shaped in lingual view (figure 6.2), whereas it has a typical V-shape in Astrapotherium (figure 6.4). All the lower molars have a small column or "pillar" attached to the posterior face of the metaconid, near the anterior end of the hypolophid, as in Astrapothericulus. The base of the "pillar" merges with the posterior slope of the metaconid, thus this element is not distinguishable in very worn molars. The "pillar" is also present in Astrapotherium, but it is more "lophoid" and transversally elongated. According to the species, the labial cingulum is either delicate (P. holmbergi and P? vogthi) or well developed (P. martiale) as in Astrapotherium, but not as much as in Astrapothericulus. The labial cingulum is present at the opening of the lingual valleys, but it is absent or very reduced at the base of the metaconid (figure 6.2), as in Urugaytherium and Xenastrapotherium and in contrast to Astrapotherium (figure 6.4) and Astrapothericulus.

Figure 6. Compared lower molars of Parastrapotherium holmbergi and Astrapotherium magnum / molares inferiores comparados de Parastrapotherium holmbergi y Astrapotherium magnum. 1, MACN A 52-504, P. holmbergi, m3 in occlusal view/ en vista oclusal; 2, same in lingual view / el mismo en vista lingual; 3, MACN A 3239, A. magnum, m3 in occlusal view / en vista oclusal; 4, same in lingual view / el mismo en vista lingual.
Skull and mandible. Based upon an almost complete, crushed palate (but lacking the premaxillaries) and basicranium (FMNH 13329, here referred to P. holmbergi), Scott (1937) pointed out that the skull of Parastrapotherium does not differ from that of Astrapotherium magnum in any notably way, except that the premaxillaries of Parastrapotherium could have been longer and posteriorly narrower. However, in other cranial remains referable to P. holmbergi, not described by Scott (AMNH 29596, a crushed juvenile skull also lacking the premaxillaries, and FMNH 13413, an almost complete right maxilla with dentition) the maxillaries have an anterior triangular slit to receive the premaxillaries even shorter than in Astrapotherium. The preserved parts of these specimens, beside a restored partial skull (AMNH 25575), show neither significant differences with Astrapotherium, except the presence of a conspicuous posttemporal foramen (not described in the Santacrucian genus) and less prominent postorbital process of the jugal. In the holotype of Parastrapotherium martiale (MACN A 52-604, a distorted anterior portion of skull) the premaxillaries are as broad and have convex ventral surface as in Astrapotherium, but they are shorter posteriorly, as in P. holmbergi. Moreover, the maxillary sheaths for the tusks are proportionally more robust, the preorbital portion of the maxillary is less prominent, and the postorbital process of the jugal is more reduced than in A. magnum and P. holmbergi. The infraorbital foramen is slightly larger and located in a more anterior and ventral position than in A. magnum, thus it is well-exposed in lateral view; this feature is not preserved in the skulls referred to P. holmbergi. Other minor differences are probably attributable to post mortem distortion.
The mandible of P. holmbergi is essentially as in A. magnum, but with some minor differences that seem to be constant among the examined specimens: the diastema is shorter due the presence of an additional premolar, the horizontal ramus is slightly lower, and the anterior margin of the ascending ramus is slightly backwardly oriented, not so vertical and without a sigmoid-like profile. As in A. magnum, the symphysis is long, strongly procumbent, deeply concave on the dorsal side and becoming shallower anteriorly. Scott (1937) described a specimen (FMNH 13365) with a distinct, erected lower canine interpreted, as a female. Concordantly, in this specimen the symphysis is narrower and less procumbent than in specimens with extroverted canines. Similar variations, but not so marked, are also observed in A. magnum (e.g. FMNH 13173). The distorted mandible associated to the skull fragment of the holotype of Parastrapotherium martiale (MACN A 52-604, figure 7), has both articulated dentaries lacking the ascending rami. Even taking the distortion into account, the jaw is more slender than in Astrapotherium giganteum, which has similar p4-m3 length. The most conspicuous feature is the presence of a single and very large mental foramen.

Figure 7. Parastrapotherium martiale, MACN A 52-604 (holotype), mandible (partially distorted), in occlusal view / (holotipo), mandíbula (parcialmente deformada), en vista oclusal.
Species
Parastrapotherium holmbergi Ameghino, 1895 Figures 2.1-2, 3.1, 3.2, 5.1-3, 5.5, 6.1, 6.2, and 8.1-3
Parastrapotherium trouessarti Ameghino, 1895: 638.
Parastrapotherium lemoinei Ameghino, 1895: 640.
Traspoatherium convexidens Ameghino, 1895: 641.
Loxocoelus obturtus Ameghino, 1901: 404.
Helicolophodon giganteus Roth, 1903: 143.
Syntype. MACN A 52-509 a-f, right maxillary fragment with P3-P4, three P3, one P4 and one M2; MACN A 52-504, two p3, two p4, five complete low-er molars and fragment of another one (not associated); MACN A 52-515, eight isolated incisors; MACN A 52-502, one isolated i2?; MACN A 52-518, four fragments of lower canines and one fragment of upper canine (not associated).
Type of synonyms. MACN A 52-503, right mandibular fragment with incomplete p4-m2 and roots of m3, MACN A 52-506, five incisors, two p3, three p4, two m1?, one m2 and one m3, non-associated, and MACN A 52-436, isolated M2 (syntypes of P. trouessarti); MACN A 52-429, isolated upper deciduous premolar (holotype of P. lemoinei); MACN A 52-401, four isolated P3 (syntypes of T. convexidens); MACN A 52-527, isolated upper deciduous premolar (holotype of Loxocoelus obturtus); MLP 12-2285, upper premolar and incisor (holotype of Helicolophodon giganteus).
Diagnosis. Size similar to Astrapotherium magnum. Upper canines with flat anterior face or with a longitudinal furrow, according to ontogenetic stage. p3 with no developed trigonid. Lower molars with a distinct "pillar". Differs from the Colhuhuapian species P. symmetrum (Ameghino, 1902) by having incisors with median longitudinal furrow on the lingual side.
Distribution. The type specimens of P. holmbergi and referred materials come from the Deseadan La Flecha locality, Santa Cruz Province. P. holmbergi is also recorded in Deseadan levels at Pico Truncado (Santa Cruz Province) and Cabeza Blanca (Chubut Province) (for these localities and others mentioned in the text see Marshall et al., 1983). The provenance of Helicolophodon giganteus, here considered as junior synonym of P. holmbergi, is still uncertain. Roth (1903) indicated that the type came from beds of his "Cretáceo Superior del Lago Musters", Chubut Province. Simpson (1936) suggested that this locality was almost surely his Cerro del Humo Locality, and assigned the fossil bearing deposits to the Mustersan Age (Late Eocene). However, Simpson (1936) suggested that the type of Helicolophodon giganteus could be derived from Deseadan beds poorly exposed at this locality.
Comments. According to Ameghino (1895), Parastrapotherium trouessarti is similar to P. holmbergi, but differing because of the presence of a "pillar" in the lower molars (Ameghino, 1897: fig. 34). Evidently, Ameghino did not recognise this structure in P. holmbergi because all the lower molars that he assigned to the type species are much more worn. Ameghino also differentiated P. trouessarti by having p4 with diverging roots, whereas the only p4 with preserved roots referred to P. holmbergi (MACN 52-504) has parallel and partially fused roots. As we have mentioned above, this feature represents individual variation. Finally, Ameghino stated that upper molars of P. trouessarti have more rippled labial walls, but the only upper molar referred to this species in the Ameghino Collection, MACN A 52-436 (figured as P. martiale by Ameghino, 1904: fig. 224 and 230, and by Loomis, 1914: fig. 108) does not differ significantly from the M2 of the syntype of P. holmbergi, except for the fact that it is less worn.
Parastrapotherium lemoinei is based upon an isolated upper tooth (MACN A 52-426, figure8.1) interpreted by Ameghino (1895) as a M2 of a species much smaller than P. holmbergi. The figure of this tooth in Ameghino's work (1897: fig. 35) does not show the lingual projection of the crochet that contacts the pos-terolabial wall of the protocone, isolating the hypoflexus from the central valley. This pattern is characteristic of deciduous premolars, as seen in the FMNH 13566, and in other astrapotheriids (e.g. Astrapothericulus, see MACN 52-417, Ameghino, 1904: fig. 5). According to its size (table 1), the holotype of P. lemoinei corresponds to a DP4 of P. holmbergi.

Figure 8. 1, Parastrapotherium lemoinei, MACN A 52-426 (holotype), right DP4/ (holotipo) DP4 derecho; 2, Parastrapotherium convexidens, MACN A 52-401 (syntype), right P3 / (sintipo), P3 derecho; 3, Helicolophodon giganteus, MLP 12-2285 (holotype), right P4/ (holotipo), P4 derecho .
The holotype of Loxocoelus obturtus was originally described as a M1 assigned to a Homalodotheriidae (Ameghino, 1901). This tooth belongs to an Astrapotheriidae since it has vertically banded enamel; it shows the same structure than the type of P. lemoinei, and basically it does not differ from the DP3 of P. holmbergi.
Traspoatherium convexidens is based upon four isolated P3 (MACN A 52-401) with more convex labial wall and less prominet labial fold than in P. holmbergi (Ameghino, 1895) (figure 8.2). However, these features are also observed in more complete specimens positively referable to P. holmbergi. Ameghino also noted that these teeth have a single root, but in fact only one of them has its roots preserved, and they are parallel and fused on the external side, which is a mere individual variation.
Helicolophodon giganteus is based upon an upper premolar (figure 8.3) and an incisor. Roth (1903) compared the premolar with the P3 of P. crassum Ameghino. However this tooth corresponds to an almost unworn P4, and the morphological differences with those of the syntype of P. holmbergi are only due the lesser degree of wear. The premolar is slightly larger than the P4 of the syntype of P. holmbergi (table 1), but very similar to the P4 of other specimens referable to this species. The incisor has no relevant differences with the syntype of P. holmbergi. This synonymy was previously suggested by Simpson (1936).
Parastrapotherium martiale Ameghino, 1901 Figures 1, 3.3, 5.4, 7, 9.1 and 9.2
Parastrapotherium superabile Ameghino, 1901: 402.
Parastrapotherium insuperabile Ameghino, 1901: 403.
Holotype. MACN A 52-604, anterior portion of skull with the bases of both canines and right P3-P4, left maxillary fragment with P3-M3, and both incomplete mandibular rami with alveoli for both i1 and alveolus for left i2, complete right i2-i3 and left i3, both canines, canine, right p3-m2, and left p3-m3, all belonging to a single specimen.
Type of synonyms. MACN A 52-404a (holotype of P. superabile), isolated P4; MACN A 52-500 (holotype of P. insuperabile), isolated P4.
Diagnosis. Differs from P. holmbergi by its larger size (25% larger), proportionally smaller premolars, more developed trigonid in p3, and lower molars with more developed labial cingulum. Differs from P. crassum by having upper canines with flat anterior face or with a longitudinal furrow, according to the ontogenetic stage. Differs from P? vogthi by its larger size and by the distinct "pillar" in the lower molars. Differs from P. symmetrum by its larger size and by its incisors with median longitudinal furrow on the lingual side.
Distribution. The provenance of the holotype of P. martiale is very dubious. Ameghino (1901) indicated that it came from his "Astraponotus beds" (Mustersan). However, this must be wrong, since later (1900-1902) he included this species in the faunal list for his "Pyrotherium beds" (Deseadan), and he did not comment the presence of Parastrapotherium in older levels. According to the catalogue of the MACN Ame-ghino Collection, the type specimen comes, with doubts, from the "Yacimiento de Pinturas", the type locality of Ameghino's "Astrapothericulan beds" (Santa Cruz province, late Early Miocene). This must also be a mistake, since Ameghino never included either P. martiale or other species of Parastrapotherium within the faunal lists for these beds (Ameghino, 1900-1902, 1906). Very abundant recent collections from the "Astrapothericulan beds" at the Pinturas Formation, housed at MACN, show no occurrence of Parastrapotherium either. Specimens positively referable to P. martiale come from Deseadan levels at the southern cliff of Lake Colhue Huapí, although some fragmentary materials from La Flecha could also correspond to this species. P. martiale also occurs at the Colhuehuapian Cerro Bandera Formation in Neuquén Province (Kramarz et al., 2005) and at Colhuehuapian levels at the southern cliff of Lake Colhue Huapí (Kramarz and Bond, in press).
Comments. Although Ameghino (1901) described the mandible of the holotype and mentioned characters of the lower and upper dentition (figure 9.1), he did not mention the associated skull fragment, even though it is the one of the most complete specimens of Parastrapotherium known to date. Ameghino (1901) and Loomis (1914) pointed out that in this species the mandibular symphysis is narrow and excavated, the canines are little diverging, and the incisors are implanted almost juxtaposed. However, we interpret that this jaw is strongly modified by post-mortem deformation (figure 7), which has also affected the associated skull fragment, and that these features cannot be considered as diagnostic.

Figure 9. Parastrapotherium martiale. 1, MACN A 52-604 (holotype), left maxillary with P3-M3 / (holotipo), maxilar izquierdo con P3-M3; 2, MACN A 52-404a, (holotype of P. superabile), right P4/ (holotipo de P. superabile) P4 derecho.
With respect to the dentition, Ameghino (1901) and Loomis (1914) noted that P. martiale differs from P. holmbergi because it has stronger labial cingula in the lower molars and lingual cingula in the uppers. However, the development of these cingula is variable in both species. Moreover, the P3 and P4 of the holotype of P. martiale show lingual cingula as developed as in those of the syntype of P. holmbergi.
The type of Parastrapotherium superabile Ameghino, 1901 is an isolated P4 (MACN A 52-404a; figure 9.2), very similar in size to the type of P. martiale (table 1); the differences in the crown features noted by Ameghino (1901) are essentially due the lesser degree of wear. In agreement with Loomis (1914), it is herein considered as synonym of P. martiale.
Parastrapotherium insuperabile Ameghino, 1901 is also based on an isolated P4 (Ameghino, 1904: fig 393-394). According to Ameghino (1901), followed by Loomis (1914), it differs from P. martiale by being significantly larger (table 1), higher crowned, and with more pronounced basal cingulum. However, the crown height does not differ from that of the holotype of P. superabile, and the cingulum is as conspicuous as in other specimens similar in size, or even smaller (e.g. MACN Pv 14542) than the type of P. martiale. Ameghino also assigned an isolated p4 (MACN A 52-404b) to P. insuperabile, pointing out that the posterior crescent (talonid) is much lower than the anterior one (trigonid), which is a mere con-sequence of the characteristic wear pattern of the p4 in the Astrapotheriidae.
Parastrapotherium crassum Ameghino, 1902 Figure 10

Figure 10. Parastrapotherium crassum. 1, MACN A 52-526, (lectotype), right upper canine in lingual view / (lectotipo), canino superior derecho en vista lingual; 2, same in schematic cross section/ el mismo en corte transversal esquemático.
Parastrapotherium crassum Ameghino, 1902: 97.
Lectotype. MACN A 52-526, a fragment of upper canine and associated P3.
Diagnosis. Similar in size to P. martiale. Differs from P. holmbergi and P. martiale by having upper canines with very convex anterior face in the adult stage.
Distribution. P. crassum was described by Ameghino (1902) for his "Colpodon beds" (Colhuehuapian) at the southern cliff of Lake Colhue Huapí. However, the specimen herein designed as lectotype have black, glossy enamel and dark dentine. Fossils obtained in recent collections with stratigraphic control reveal that this kind of preservation is typical of the fossil teeth from Deseadan levels at the southern cliff of Lake Colhue Huapí, whereas all teeth from Colhuehuapian levels at this locality (Colhue Huapí Member of the Sarmiento Formation, Spalletti and Mazzoni, 1979) have very characteristic brown enamel and light coloured dentine. The remaining specimens described by Ameghino (1902) as P. crassum belong to other species provisionally referred to the genus Astrapotherium (Kramarz and Bond, in press) and, concordantly, show "colhuehuapian" preservation. Specimens referable to P. crassum also occur at La Flecha locality. Therefore, P. crassum should be listed among the Deseadan species.
Comments. This species was based by Ameghino (1902) on many specimens that we interpret as belonging to different species. We herein designate the upper canine fragment and associated P3 (MACN A 52-526) as lectotype, since it is the only specimen with differential features. According to Ameghino (1902), this canine differs by having very convex labial face, thus the tooth is ovoid in cross section (figure 10.2); the associated P3 indicates that this is an adult specimen. On the contrary, the upper canine of the remaining species is subtriangular in cross section in juveniles, or heart-shaped in adults (figure 1). The preserved proximal portion of the canine is rather smaller than the holotype of P. martiale (table 1), but the associated P3 is slightly larger and does not differ significantly in morphology. Isolated upper canines with identical morphology (MLP 95-III-10-103) coming from La Flecha locality are herein referred to P. crassum. We provisionally accept P. crassum as a valid species until further evidence is known, being the only diagnostic feature the characteristic shape of the upper canine. Ameghino (1902) also assigned to P. crassum a lower canine (MACN A 52-521), noting that it differs from other species by having the lingual face flat instead of concave as in the remaining species. This canine is associated with a mandibular fragment with p4-m1, which clearly lacks p3, and thus it is not cogeneric with the lectotype. This specimen is more probably derived from Colhuehuapian levels at the Southern cliffs of Lake Colhue Huapí and is provisionally referred to genus Astrapotherium (Kramarz and Bond, in press). The remaining specimens referred by Ameghino (1902) to this species P. crassum (an isolated upper molar, MACN A 52-517, and isolated incisors, MACN A 52-514), belong to Parastrapotherium but cannot be identified to specific level. Ameghino (1902) stated that P. crassum had three deciduous lower premolars, but the anteriormost was not replaced. Although this is the normal condition in Parastrapotherium species, the Ameghino collection contains no specimen with complete milk dentition to allow testing such condition.
Parastrapotherium? voghti (Mercerat, 1891) Figure 11.1
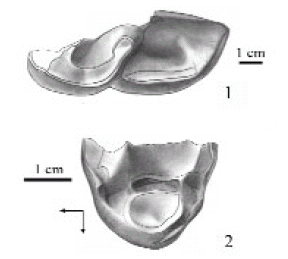
Figure 11. 1, Parastrapotherium? voghti, MLP 12-122 (syntype), left m3/ (sintipo), m3 izquierdo. 2, Liarthrus copei, MACN A 52-406 (lectotype), incomplete left P3? / (lectotipo), P3? izquierdo incompleto. Arrows show anterior and lingual sides / las flechas indican los lados anterior y lingual.
Syntype. MLP 12-122, left mandibular fragment with m2-m3 and probably associated right m1, left m2, m3 and right m3, two isolated upper molars and two fragments of upper molars
Diagnosis. Similar in size to P. holmbergi. Differs from P. holmbergi and P. martiale in having lower molars without distinct "pillar".
Distribution. The syntype of Parastrapotherium? voghti comes from an undetermined unit at "Carhué", Chubut Province (in schedula), probably located near Paso de Indios locality. Dental remains of Pyrotherium are also probably from "Carhué", suggesting a Deseadan age for this locality.
Comments. This species is herein tentatively transferred to Parastrapotherium because all the available cheek teeth are lower crowned than in Astrapotherium, the upper molars have no labial cingulum and the lowers have no lingual cingulum at the base of the metaconid, as in P. holmbergi. However, it differs from the species of Parastrapotherium by lacking the characteristic "pìllar" of the lower molars. The m3 are little worn (figure 11.1) and both show a distinct cuspule attached to the posterior portion of the lingual wall of the hypolophid that probably corresponds to a vestigial entoconid, which is not observed in the same stage of wear either in P. holmbergi or P. martiale. Ameghino (1891a and b, 1895) considered this species as synonym of his P. ephebicum, but we do not accept this proposal since the dental measurements for P.? voghti (table 1) are significantly larger than those of P. ephebicum (see section below).
Astrapotherium ephebicum Ameghino, 1889 species inquirenda
Comments. In the first description of A. ephebicum, Ameghino (1889) indicated only that this is a small species from the Pyrotherium beds at Neuquén Province, with m1 slightly more than 30 mm long. In 1891 Ameghino also described an upper canine, incisors and a M1, but indicated that the anteroposterior diameter of the m1 is 34 to 38 mm, according to the stage of wear, and that the species is known from Deseado and Chubut, in addition to Neuquén Province. However, in 1895 Ameghino transferred it to Parastrapotherium, stating that the species is known only through a m1 and a m2 of an old individual, and that the m1 is 31 mm long. Loomis (1914), who did not examine Ameghino's specimens, interpreted the original description as being based on a mandibular fragment with m1 and m2, but at present there are no mandibular remains referred to P. ephebicum in the Ameghino Collection, and no specimen agrees with the 1889 description of P. ephebicum. It is probable that the tooth described in 1889 is a deciduous lower premolar of a large species, but there is no current evidence to verify either the validity of this species or its distribution.
Liarthrus Ameghino, 1895
Type species. Liarthrus copei Ameghino, 1895.
Geographic and stratigraphic distribution. Sarmiento Formation, Deseadan Age (Late Oligocene), Santa Cruz Province.
Emended diagnosis. P3 with no developed anterior transverse crest (protoloph).
Liarthrus copei Ameghino, 1895 Figure 11.2
Holotype. MACN A 52-406, right astragalus, p4, and incomplete upper premolar.
Provenance. The type specimen Liarthrus copei comes from La Flecha locality, Santa Cruz. Province. Other isolated upper premolars referable to this species (housed at the FMNH) come from the same locality and from Pico Truncado Locality.
Comments. The astragalus described by Ameghino (1895) barely differs from that of Parastrapotherium by having a slightly flatter tibial facet, and it does not justify taxonomic distinction. Regarding the p4, Ameghino noted that the labial flexid is less marked than in A. magnum, a condition that we interpret as a diagnostic feature for the genus Parastrapotherium, and an accessory column in the posterior valley ("pillar"), which occurs variably both in Parastrapotherium and Astrapotherium. Ameghino (1895) indicated that the lingual cusp of the upper premolar is isolated from the outer wall, and in 1897 (:fig. 137 a) he interpreted this tooth as a P2. However, this tooth is more probably a P3 (Gaudry, 1904) (at least the preserved part is similar in size to the P3 of the P. holmbergi syntype) and both the description and the figure are inaccurate. The lingual cusp is not isolated, but connected with the labial wall through two very low and delicate transverse crests; the posterior one would correspond to the posterior lingual projection of the ectoloph (figure 11.2). Therefore, this tooth differs from all the remaining Astrapotheriidae in the absence of a true protoloph, suggesting a radically distinct structural condition. Moreover, the protocone is more conical and the enamel layer on the labial wall of the central valley is much broader than in the species of Parastrapotherium, Astrapotherium, and Astrapothericulus. In 1897 Ameghino assigned to L. copei an upper premolar (MACN A 52-407, see Ameghino, 1897: fig. 137 b), characterized by being antero-posteriorly compressed, which is probably an anomalous condition. Loomis (1914) considered L. copei as junior synonym of P. holmbergi based on the characters and size of the described P4 and P3. However, the P4 is not included in the original type series of L. copei; this tooth would actually belong to Parastrapotherium, clo-ser in size to P. martiale than to P. holmbergi. In summary, we provisionally accept L. copei as a valid species of still uncertain affinities.
Taxonomic conclusions
The genus Parastrapotherium Ameghino shows several dental characters distinct from Astrapotherium Burmeister: lower crowned cheek teeth, p3 present, basal cingulum absent at the base of the labial fold in all upper cheek teeth, upper premolars with a labial fold less prominent and with broader base, crown of the lower incisors proportionally more robust and buccolingually broader, lower cheek teeth with very shallow labial flexid, lower molars with bunoid "pillar", and lingual cingulum absent at the base of the metaconid. In view of the conservative nature of the astrapotheriids, these differences largely justify the separation of Parastrapotherium as a distinct genus.
Of the Deseadan species described for the genus Parastrapotherium, Parastrapotherium holmbergi Ameghino (= P. trouessarti, P. lemoinei, and Traspoatherium convexidens), Parastrapotherium martiale Ameghino (= P. superabile and P. insuperabile), and Parastrapotherium crassum Ameghino are herein accepted as valid. This proposal partially agrees with the synonymies indicated by Loomis (1914). P. martiale differs from the type species by its larger size, P3 and p3 proportionally smaller, and p3 with more complex trigonid. P. crassum differs by having upper canines with ovoid cross section, instead of subtriangular or heartlike as the former species.
Astrapotherium voghti Mercerat is also interpreted as a valid species, different from P. ephebicum, and it is herein transferred tentatively to Parastrapotherium. Liarthrus copei has a distinct structure of the P3 that suggests it is a valid species of still uncertain affinities. The validity of P. ephebicum is uncertain, as it is probably based on deciduous premolars not found in the Ameghino collection.
These conclusions complement a recent revision of the Colhuehuapian astrapotheres (Kramarz and Bond, in press): Parastrapotherium symmetrum (Ameghino, 1902) and Parastrapotherium herculeum (Ameghino, 1899) are the only valid species of the genus for this Age; Parastrapotherium ruderarium Ameghino, 1902 (= Parastrapotherium paucum Ameghino, 1902) should be transferred to Astrapotherium. These results suggest that Parastrapotherium includes a limited number of species, which range in size from large to giant; all the small species referred to this genus are based on deciduous dentition of large taxa or are referable to other genera.
Considerations on Parastrapotherium relationships
Before the recognition of the non-patagonian astrapotheriids as a distinct lineage within the astrapotheres (i.e. Urugaytheriinae, Kraglievich, 1928), Parastrapotherium has been considered as close related to Astrapotherium (Ameghino, 1895; Scott, 1928). Kraglievich (1928) did not include explicitly Parastrapotherium within his Uruguaytheriinae, but proposed that Parastrapotherium could be related to the origin of this subfamily because its dentition is not as specialized as in Astrapotherium. Subsequent phylogenetic proposals (Soria, 1984; Frailey, 1987; Cifelli, 1993) did not consider the uruguaytherine genera, and therefore the relationships between Parastrapotherium and the remaining Oligocene -Miocene astrapotheres were not tested. The more recent analysis on the astrapotheres (Johnson and Madden, 1997) concluded that Parastrapotherium is close to Astrapotherium, and that Astrapothericulus is the sister group of this clade plus the monophyletic Uruguaytheriinae, though noting that the position of the three patagonian genera is weakly supported. We interpret that this proposal implies that Parastrapotherium (which has two lower premolars) would have derived from an ancestor with only one lower premolar, as in Astrapothericulus. Moreover, Johnson and Madden (1997) did not consider that the cheek teeth of Parastrapotherium are positively lower crowned than those of Astrapotherium and Astrapothericulus. These characters suggest that Parastrapotherium would have a very basal position among the Oligocene - Miocene astrapotheres. On the other hand, Parastrapotherium has a poorly developed labial flexid in the lower molars, and this is a very distinct feature for the Uruguaytheriinae (Kraglievich, 1928; Johnson and Madden, 1997). Likewise, the absence of cingulum at the base of the metaconid is also shared with the Uruguaytheriinae. Certainly, the relationships of Parastrapotherium are still uncertain and a complete cladistic analysis is required to evaluate the polarity and phylogenetic relevance of these features. At present, most of the dental characters are not verifiable in the still poorly known Eocene Astraponotus, traditionally accepted as the sister group of all the younger astrapotheres (Simpson, 1967; Soria, 1984; Frailey, 1987; Cifelli, 1993). New and very complete cranial and mandibular materials of Astraponotus, as also a partial skull of Astrapothericulus from the Pinturas Formation, are now under study. These still undescribed materials will provide a more complete data set for a further cladistic analysis to evaluate the phylogenetic position of Parastrapotherium as well as the relationships among the astrapotheriids.
Acknowledgments
We thank M.G. Vucetich (MLP) and R. Madden (Duke University) for allowing us to study the materials collected during the MLP - Duke University expeditions to Patagonia supported by U. S. National Science Foundation and Consejo Nacional de Investigaciones Científicas y Técnicas. Special thanks to E. Ruigomez (MPEF), M. Reguero (MLP), J. Flynn (AMNH), and P. Makovicky (FMNH) for access to materials under their care. E. Bellosi (MACN) and A Carlini (MLP) provided significant information about the stratigraphy of Gran Barranca. To the reviewers E. Cerdeño (IANIGLA-CRICYT, Mendoza) and M. Reguero, for their useful comments. C. Morgan (MLP) for her help in the English version. Drawings were made by the artist J. González. This work was partially financed by BID 1728/0C-AR PICT 32344 awarded to V. Barreda (MACN) and by Consejo Nacional de Investigaciones Científicas y Técnicas (Res. Nro.1227/05) to A. Kramarz.
References
1. Ameghino, F. 1887. Ennumeración sistemática de las especies de mamíferos fósiles coleccionados por Carlos Ameghino en los terrenos eocenos de Patagonia austral y depositados en el museo de La Plata. Boletín del Museo de La Plata 1: 1-26. [ Links ]
2. Ameghino, F. 1889. Contribución al conocimiento de los mamíferos fósiles de la República Argentina. Actas de la Academia Nacional de Ciencias en Córdoba 6: xxxii + 1-1027. [ Links ]
3. Ameghino, F. 1891a. Mamíferos y Aves fósiles argentinas. Especies nuevas, adiciones y correcciones. Revista Argentina de Historia Natural 1: 240-259. [ Links ]
4. Ameghino, F. 1891b. Revista crítica y bibliográfica: "Sinópsis de la familia de los Astrapotheriidae.por Alcides Mercerat. Revista Argentina de Historia Natural 1: 275-280. [ Links ]
5. Ameghino, F. 1895. Prémière contribution a la connaissance de la faune mammalogique des couches a Pyrotherium. Boletín Instituto Geográfico Argentino 15: 306-660. [ Links ]
6. Ameghino, F. 1897. Mammifères crétacés de l´Argentine. Deuxième contribution a la connaissance de la faune mammalogique des couches a Pyrotherium. Boletín Instituto Geográfico Argentino 18: 406-429, 431-521. [ Links ]
7. Ameghino, F. 1899. Sinópsis geológico - paleontológica. Suplemento (Adiciones y Correcciones): 1-13. [ Links ]
8. Ameghino, F. 1900-1902. L´âge des formations sédimentaires de Patagonie. Anales de la Sociedad Científica Argentina 50: 109-131, 145-165, 209-229; 51: 20-39, 65-91, 52: 198-197, 244-250; 54: 161-180, 220-240, 283-342. [ Links ]
9. Ameghino, F. 1901. Notices prèliminaires sur des ongulés nouveaux des terrains Crétacés de Patagonie. Boletín de la Academia de Ciencias (Córdoba) 16: 349-426. [ Links ]
10. Ameghino, F. 1902. Première contribution à la connaissance de la faune mammalogique des couches à Colpodon. Boletín de la Academia Nacional de Ciencias (Córdoba) 17: 71-138. [ Links ]
11. Ameghino, F. 1904. Recherches de morphologie phylogénétique sur les molaires supérieures des ongulés. Anales del Museo Nacional de Buenos Aires 3: 1-541. [ Links ]
12. Ameghino, F. 1906. Les Formations sédimentaires du Crétacé supérieur et du Tertiaire de Patagonie. Anales del Museo Nacional de Buenos Aires 8: 1-358. [ Links ]
13. Cifelli, R.L. 1993. The phylogeny of the native South American ungulates. In: F.S. Szalay, M.J. Novacek y M.C. McKenna (eds.), Mammal Phylogeny, Springer-Verlag pp. 195-216. [ Links ]
14. Flynn, J.J. and Swisher, C.C. III 1995. Cenozoic South American Land Mammal Ages: Correlation to global geochronologies. Geochronology Times Scales and Global Stratigraphic Correlation. SEPM Special publication 54: 317-333. [ Links ]
15. Frailey, C.D. 1987. The Miocene Vertebrates of Quebrada Honda, Bolivia. Part I. Astrapotheria. Occasional Papers of the Museum of Natural History, the University of Kansas 122: 1-15. [ Links ]
16. Gaudry, A. 1904. Fossiles de Patagonie. Dentition de quelques mammifères. Mémoires de la Société Géologique de France. Paléontologie 12: 5-43. [ Links ]
17. Johnson, S.C. and Madden, R.H. 1997. Uruguaytheriine Astrapotheres of Tropical South America. In: R.F. Key, R.H. Madden, R.L. Cifelli, and J.J. Flynn (eds.), Vertebrate Paleontology in the Neotropics. The Miocene fauna of La Venta, Colombia, Smithsonian Institution Press, Washington and London, pp. 355-381. [ Links ]
18. Kraglievich, L. 1928. Sobre el supuesto Astrapotherium christi Stehlin, descubierto en Venezuela (Xenastrapotherium n. gen.) y sus relaciones con Astrapotherium magnum y Uruguaytherium beaulieui., pp. 1-16. La Editorial Franco-Argentina. Buenos Aires. [ Links ]
19. Kramarz, A. and Bond, M. (in press). Colhuehuapian Astrapotheriids (Mammalia) from Gran Barranca Sur del Lago Colhue Huapi. In: R. Madden, A. Carlini. M.G. Vucetich and R. Kay (eds.), ´The Paleontology of Gran Barranca: Evolution and Environmental Change through the Middle Cenozoic". Cambridge University Press. [ Links ]
20. Kramarz, A., Garrido, A., Forasiepi, A., Bond, M. and Tambussi, C. 2005. Estratigrafía y vertebrados (Mammalia - Aves) de la Formación Cerro Bandera, Mioceno Temprano de la provincia del Neuquén, Argentina. Revista Geológica de Chile 32: 273-291 [ Links ]
21. Loomis, F.B. 1914. The Deseado Formation of Patagonia. Rumford Press, Concord, N. H, Amherst, 232 pp. [ Links ]
22. Lydekker, R. 1894. Contribution to the knowledge of the fossil vertebrates of Argentina. 3. A study of extinct Argentine un-gulates. Anales del Museo de La Plata, Paleontología Argentina 2: 1-91. [ Links ]
23. Marshall, L.G., Hoffstetter, R., and Pascual, R. 1983. Mammals and stratigraphy: geochronology of continental mammals-bearing Tertiary of South America. Paleovertebrata, Mémoire Extraordinaire: 1-93. [ Links ]
24. Mercerat, A.1891. Sinópsis de la familia de los Astrapotheriidae (Eoceno de Patagonia). Revista del Museo de La Plata 1: 241-255. [ Links ]
25. Roth, S. 1903. Noticias preliminares sobre nuevos mamíferos fósiles del Cretáceo superior y Terciario inferior de la Patagonia. Revista del Museo de La Plata 11: 135-158. [ Links ]
26. Scott, W.B. 1928. Mammalia of the Santa Cruz beds. IV Astrapotheria. In: W.B. Scott (ed.), Reports of the Princeton University Expeditions to Patagonia, 1896-1899, Princeton University, E. Schweizerbart'sche Verlagshandlung (E. Nägele), Stuttgart, pp. 301-351. [ Links ]
27. Scott, W.B. 1937. The Astrapotheria. Proceedings of the American Philosophical Society 77: 300-393. [ Links ]
28. Simpson, G.G. 1936. Notas sobre los mamíferos más antiguos de la colección Roth. Instituto del Museo de la Universidad Nacional de La Plata. Obra del Cincuentenario 2: 63-94. [ Links ]
29. Simpson, G. G. 1967. The beginning of the Age of mammals in South America. Part 2. Bulletin of the American Museum of Natural History 137: 1-259, plates 1-46 [ Links ]
30. Soria, M.F. 1984. Eoastrapostylopidae: diagnosis e implicaciones en la sistemática y evolución de los Astrapotheria preoligocénicos. 2º Congreso Argentino de Paleontología y Bioestratigrafía (Corrientes),, Actas 1982: 175-182. [ Links ]
31. Spalletti, L.A. and Mazzoni, M.M. 1979. Estratigrafía de la Formación Sarmiento en la barranca sur del lago Colhue-Huapi, provincia del Chubut. Revista de la Asociación Geológica Argentina 34: 271-281. [ Links ]
Recibido: 1 de diciembre de 2006.
Aceptado: 10 de julio de 2008.

