










Services on Demand
Journal

Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAmeghiniana
On-line version ISSN 1851-8044
Ameghiniana vol.45 no.3 Buenos Aires July/Sept. 2008
Pitekunsaurus macayai gen. et sp. nov., nuevo titanosaurio (Saurischia, Sauropoda) del Cretácico Superior de la Cuenca Neuquina, Argentina
Leonardo S. Filippi1 y Alberto C. Garrido2,3
1Museo Municipal "Argentino Urquiza", Jujuy y Chaco s/n, (8319) Rincón de los Sauces, Neuquén, Argentina. lsfilippi@yahoo.com.ar
2Museo Carmen Funes, Av. Córdoba 55, 8318 Plaza Huincul, Neuquén, Argentina.
3Dirección General de Minería, Elena de Vega 472, 8340 Zapala, Neuquén, Argentina. algene@copelnet.com.ar
Abstract. Pitekunsaurus macayai gen. et sp. nov., new titanosaur (Saurischia, Sauropoda) from Upper Cretaceous Neuquén Basin, Argentina. A new titanosaur is described, Pitekunsaurus macayai gen. et sp. nov., from mudstone levels asigned to Anacleto Formation (Lower - Middle Campanian), corresponding to the uppermost beds of the Neuquén Group (Upper Cretaceous of Neuquén Basin). The specimen is represented by braincase, left frontal, one tooth, four cervical vertebrae, three dorsal vertebrae, four caudal vertebrae, right ulna and scapula, proximal extreme of left femur, rib fragments and uncertain remains. Pitekunsaurus is characterized by the following autapomorphies: (1) basipterygoid processes broadly separated and parallelly projected, (2) anterior cervical vertebrae with small depressions or longitudinal grooves in the spinal sector of spinopostzygapophyseal lamina, (3) centropostzygapophyseal lamina forked proximally in anterior dorsal vertebrae, and (4) posterior centrodiapophyseal lamina with accessory lamina in anterior dorsal vertebrae. The existence of two types of articulations in the posterior caudal vertebrae, one amphicoelous and another biconvex, indicates a close relationship with Rinconsaurus caudamirus Calvo y González Riga, suggesting that the caudal morphology of titanosaurs is much more complex and more varied than previously supposed.
Resumen. Se da a conocer un nuevo titanosaurio, Pitekunsaurus macayai gen. et sp. nov., procedente de niveles fangolíticos asignados a la Formación Anacleto (Campaniano Inferior - Medio), correspondiente a los depósitos cuspidales del Grupo Neuquén (Cretácico Superior de Cuenca Neuquina). El ejemplar esta representado por basicraneo, frontal izquierdo, un diente, cuatro vértebras cervicales, tres dorsales y cuatro caudales, ulna y escápula derecha, extremo proximal de fémur izquierdo, fragmentos de costillas y restos indeterminados. Pitekunsaurus está caracterizado por las siguientes autapomorfías: (1) procesos basipterigoides ampliamente separados y proyectados paralelamente, (2) presencia en las cervicales anteriores de pequeñas depresiones o fosas longitudinales en el sector espinal de lámina espinopostzigapofiseal, (3) lámina centropostzigapofiseal bifurcada proximalmente en las dorsales anteriores y (4) lámina centrodiapofiseal posterior con una lámina accesoria en las dorsales anteriores. La existencia en las vértebras caudales posteriores de dos tipos de articulación, una anficélica y otra biconvexa, sugiere una relación mas estrecha con Rinconsaurus caudamirus Calvo y González Riga, sugiriendo que la morfología caudal de los titanosaurios es mucho más compleja y variada de lo sospechado.
Key words. Sauropoda; Titanosauria; Upper Cretaceous; Neuquén Basin; Argentina.
Palabras clave. Sauropoda; Titanosauria; Cretácico Superior; Cuenca Neuquina; Argentina.
Introducción
En los depósitos continentales del Cretácico Superior aflorantes en ambas márgenes del Río Colorado, en inmediaciones de la localidad neuquina de Rincón de los Sauces, se han realizado en las dos últimas décadas importantes hallazgos de dinosaurios saurópodos (Calvo et al., 1997; Coria y Salgado, 1999; González Riga y Calvo, 2001; Calvo, 2002; Calvo y González Riga, 2003; Calvo y González Riga, 2004; González Riga, 2003, 2005). Estos ejemplares pertenecen en todos los casos a restos de titanosaurios, el grupo de dinosaurios mejor representado en el Cretácico Superior de la Cuenca Neuquina (Powell, 2003; Salgado, 2003). Aun cuando el número de hallazgos realizados en esta región es importante, hasta el momento solo se han descripto dos especies: Rinconsaurus caudamirus (Calvo y González Riga, 2003) y Mendozasaurus neguyelap (González Riga, 2003, 2005).
En este trabajo se describe una nueva especie de titanosaurio, integrada por materiales craneanos y postcraneanos, los que fueron preliminarmente dados a conocer por Filippi y Garrido (2006). Los materiales de esta nueva especie proceden de la localidad de Aguada Grande, sitio ubicado a 6 km al sur de la ciudad de Rincón de los Sauces, en la provincia de Neuquén (figura 1). Con esta contribución, se pretende aportar nueva información que ayude a comprender mejor la diversidad de los titanosaurios del Cretácico Superior de Patagonia.

Figura 1. Mapa de ubicación del sitio donde fue hallado el holotipo de Pitekunsaurus macayai / location map of the place where the holotype of Pitekunsaurus macayai was found.
Abreviaturas institucionales. MAU-Pv, Paleontología de Vertebrados, Museo Municipal "Argentino Urquiza", Rincón de los Sauces, Neuquén, Argentina; ISI, Indian Statistical Institute, Calcuta, India; MCNA, Museo de Ciencias Naturales de Alava, España; MUCPv, Paleontología de Vertebrados, Museo Universidad Nacional del Comahue, Neuquén, Argentina; TMM, Texas Memorial Museum, Austin, Estados Unidos.
Nota. Las actuales siglas MAU-Pv, son utilizadas en reemplazo de las antiguas siglas MRS-Pv, con las que se denominaban las piezas del ex Museo Paleontológico Rincón de los Sauces, hoy Museo Municipal "Argentino Urquiza".
Abreviaturas de localidades. MAU-Pv-AC, Aguada Chivato; MAU-Pv-AG, Aguada Grande; MAU-Pv-CRS, Cañadón Río Seco; MAU-Pv-LL, Loma del Lindero.
Abreviaturas anatómicas. Para identificar las láminas neurales de las vértebras se ha utilizado la nomenclatura propuesta por Wilson (1999). ac, acromión; acpl, lámina centroparapofiseal anterior; apcdl, lámina accesoria de la lámina centrodiapofiseal posterior; cc, costilla cervical; ccr, cavidad craneana; cf, cabeza femoral; cgl, cavidad glenoidea; cn, canal neural; co, cóndilo occipital; cpol, lámina centropostzigapofiseal; cprl, lámina centroprezigapofiseal; dp, diapófisis; cpes, cresta paraesfenoidal; cpo, crista prootica; en, espina neural; eo, exoccipital; fepz, fosa espinopostzigapofiseal; fipz, fosa infrapostzigapofiseal; fn, faceta para el nasal; fo, fenestra ovalis; fp, facetas para el parietal; fprf, facetas para el prefrontal; fpt, fosa pituitaria; fr, fosa radial; fsc, fosa supracoracoidea; he, hoja escapular; lpze, lámina postzigapostespinal; ol, olecranón; pcdl, lámina centrodiapofiseal posterior; pcpl, lámina centroparapofiseal posterior; pbpt, procesos basipterigoides; pes, paraesfenoides; pl, pleurocelo; plat, protuberancia lateral; p.d., lámina paradiapofiseal; podl, lámina postzigodiapofiseal; posl, lámina postespinal; pp, parapófisis; ppo, proceso paraoccipital; prdl, lámina prezigo-diapofiseal; prpl, lámina prezigoparapofiseal; prsl, lámina prespinal; prz, prezigapófisis; pt, proceso transverso; pz, postzigapófisis; sar, superficie articular para el radio; so, supraoccipital; sls/obs, suturas lateroesfenoides/orbitoesfenoides; spdl, lámina espino-diapofiseal; spol, lámina espinopostzigapofiseal; sprl, lámina espinoprezigapofiseal; tb, tuberosidad basal; 4t, cuarto trocánter; vy, vena yugular; V-XII, forámenes de salida de los nervios craneales.
Contexto geológico
Los restos óseos de este ejemplar MAU-Pv-AG-446 fueron hallados desarticulados en depósitos de fangolitas arcillosas macizas, asociados a delgados horizontes de caliche y pequeñas geodas de calcita. Debido a la escasa dispersión de los materiales asociados y al hecho que no se han recuperado piezas repetidas, presentando morfologías y dimensiones comparables entre sí; se atribuyen los restos recuperados a un único ejemplar.
El sector sobre el que se ubica el sitio fosilífero se encuentra mayormente cubierto por una fina capa de depósitos aluviales y eólicos holocenos, los cuales enmascaran a gran parte de los afloramientos cretácicos. En este sentido, ante la imposibilidad de poder realizar un perfil estratigráfico más completo y representativo del área, se procedió a levantar una sección estratigráfica a lo largo de la pared oriental del cañadón en el sitio fosilífero (figura 2), reconociéndose un conjunto de cuerpos sedimentarios canaliformes y tabulares a subtabulares de espesor variable, con potencias siempre menores a los 1,50 m. En el cuadro 1, se expone sintéticamente las características e interpretación de las litofacies identificadas.
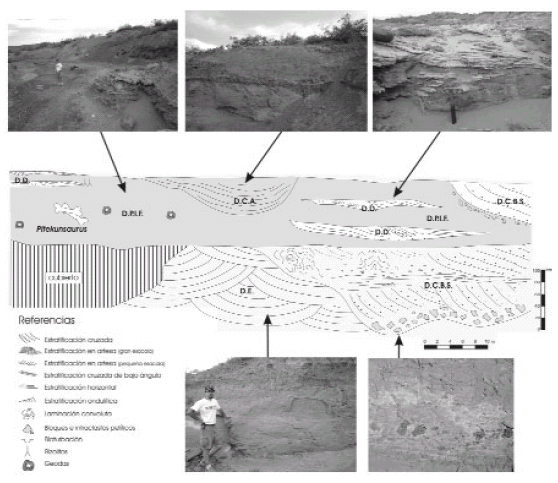
Figura 2. Sección estratigráfica de los depósitos fosilíferos de Aguada Grande donde fue hallado el holotipo de Pitekunsaurus macayai. Referencias: D.C.A., depósitos de canales abandonados, D.C.B.S., depósitos de canales de baja sinuosidad, D.D., depósitos de desbordes, D.E., depósitos eólicos, D.P.I.F., depósitos de planicie de inundación fangosa / stratigraphic section of the Aguada Grande fosiliferous deposits where the holotype of Pitekunsaurus macayai was found. References: D.C.A., abandoned deposits canals, D.C.B.S., low sinuosity deposits canals, D.D., overflowed deposits, D.E., eolic deposits, D.P.I.F., deposits of muddy flooded plain.
Cuadro 1. Litofacies reconocidas para la Formación Anacleto en el sitio fosilífero de Aguada Grande / recognice litofacies for the Ancleto Formation in the fosiliferous Aguada Grande site.
Los cuerpos canalizados de mayor magnitud (D.C.B.S. = depósitos de canales de baja sinuosidad, figura 2) conforman cuerpos tabulares, acuñados lateralmente, compuestos por areniscas cuarzolíticas friables de granulometría gruesa y pobre selección, presentando además una abundante concentración de intraclastos pelíticos con diámetros que van desde los 25 cm en la sección basal hasta los 5 mm en la sección media y cuspidal. En general estos cuerpos exhiben una sucesión granodecreciente, integrada por una asociación de depósitos tractivos (facies SP dominante, St, Sh, Sl y Sr) interpretados, sobre la base de su asociación, dimensiones, geometría y facies dominantes, como depósitos de barras arenosas longitudinales (Haszeldine, 1983; Walker y Cant, 1984). En los en los términos granulométricos más finos, resultan frecuentes la presencia de estructuras de deformación por licuefacción y escape de fluidos (facies Scl).
Los depósitos pelíticos (D.P.I.F. = depósitos de planicie de inundación fangosa, figura 2) son los que presentan una mayor continuidad y extensión lateral, desarrollándose en cuerpos tabulares compuestos principalmente por fangolitas macizas de coloración rojiza (facies Fm). En algunos sectores, estos depósitos presentan frecuentes rasgos de edafización, marcados principalmente por la presencia de trazas radiculares (facies Fr) y delgados niveles irregulares de caliche pedogénico (facies P). Variados restos fósiles, entre los que se encuentra el ejemplar MAU-Pv-AG-446, provienen de estos niveles.
Íntimamente asociados a los depósitos pelíticos, se encuentran pequeños cuerpos arenosos de geometría subtabular a lenticular (D.D. = depósitos de desborde, figura 2), con espesores menores a los 40 cm y una extensión lateral máxima de pocas decenas de metros. Los mismos se componen de areniscas cuarzolíticas de granulometría media y moderada selección, exhibiendo una asociación de estructuras sedimentarias tractivas compuestas por artesas de pequeña escala (facies St), estratificación horizontal (facies Sh) y estratificación cruzada de bajo ángulo (facies Sl). Un rasgo destacable en estos cuerpos es la presencia de una intensa bioturbación que comprende una alta densidad de trazas de tipo Taenidium y Scoyenia, las cuales obliteran en algunos sectores todo rasgo de estratificación primaria.
Formas canalizadas rellenas por depósitos de fangolitas arenosas laminadas (D.C.A.= depósitos de canales abandonados, figura 2), constituyen otra configuración de cuerpo sedimentario asociada a los depósitos de planicie de inundación fangosa. La tenue laminación de estos depósitos concordante a la base cóncava de la estratoforma (facies Fl), sugiere una sedimentación por acreción vertical en cuerpos de aguas estancas; rasgo particular de depósitos subácueos desarrollados en canales abandonados (Spalletti, 1987).
Completan la secuencia sedimentaria un conjunto de depósitos arenosos (D.E.= depósitos eólicos, figura 2) conformados por areniscas cuarcíticas medianas a finas, de granos subredondeados a redondeados y buena selección. El rasgo sedimentario dominante de estos depósitos lo constituye un conjunto de cosets de estratificación cruzada en artesas de mediana a gran escala, limitados por marcadas superficies de reactivación (facies St (2)). Sobre la base de sus atributos sedimentarios (granos redondeados, buena selección y desarrollo dominante de artesas de mediana a gran escala) se atribuyen estas sedimentitas a depósitos eólicos, posiblemente originados por aventamientos o blow outs (Spalletti, 1987).
Sobre la base de la presencia de cuerpos arenosos canalizados de geometría tabular, la ausencia de cuerpos de acreción lateral y una asociación de facies compleja con estratificación cruzada planar dominante; se establece un modelo sedimentario conformado por sistemas de canales arenosos de baja sinuosidad desarrollados en planicies de inundación fangosas (Collinson, 1986; Bentham et al., 1993; Miall, 1996). La intercalación de cuerpos producidos por flujos de desbordes, indicarían períodos de caudales excepcionales, los cuales inundarían algunas regiones de la planicie y colmatarían de agua sectores deprimidos y canales o brazos abandonados.
La presencia de depósitos eólicos y el desarrollo de horizontes de caliche pedogénico, sugerirían condiciones de sequedad ambiental o semiaridéz. Estas interpretaciones resultan concordantes con los datos aportados por Garrido (2000) para la mayor parte de la secuencia sedimentaria continental cretácica del área central de la Cuenca Neuquina, señalándose para esta un régimen climático templado cálido con marcadas alternancias de periodos estacionales secos y húmedos.
Respecto a la edad de estos depósitos, se asignan a la Formación Anacleto (Subgrupo Río Colorado, Grupo Neuquén) sobre la base de su posición y relaciones estratigráficas, con una edad estimada en el Campaniano Inferior - Medio (Legarreta y Gulisano, 1989).
Paleontología sistemática
Superorden Dinosauria Owen, 1842
Orden Saurischia Seeley, 1888
Infraorden Sauropoda Marsh, 1878
Superfamilia Titanosauriformes Salgado, Coria y Calvo, 1997
Familia Titanosauria Bonaparte y Coria, 1993
Pitekunsaurus gen. nov.
Etimología. Vocablo proveniente del idioma mapuche. Pitëkun: denunciar o descubrir algo (Espósito, 2003), en referencia a que el ejemplar fue hallado gracias a la denuncia realizada por el Sr. Luis Macaya; y saurus (Griego), reptil.
Especie tipo. Pitekunsaurus macayai sp. nov.
Diagnosis. La misma que la de la especie tipo.
Pitekunsaurus macayai sp. nov. Figuras 3-8

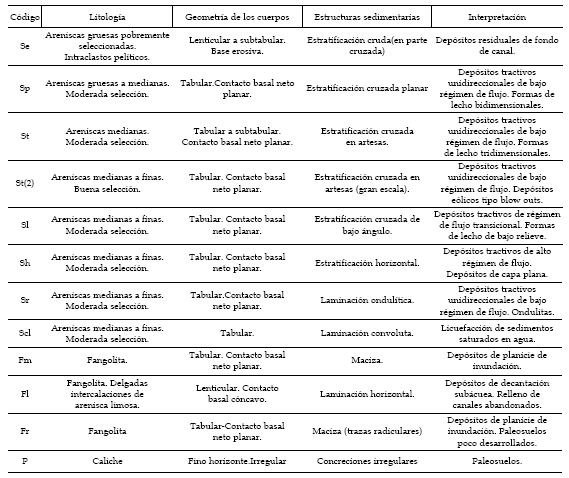
Figura 3. Pitekunsaurus macayai. 1-4, basicraneo en vista posterior (1), anteroventral (2), lateral (3) y anterodorsal (4) (MAU-Pv-AG-446/5) / Pitekunsaurus macayai. 1-4, braincase in posterior (1), anteroventral (2), lateral (3) and anterodorsal view (4). Escala gráfica: 10 cm / scale bar: 10 cm.
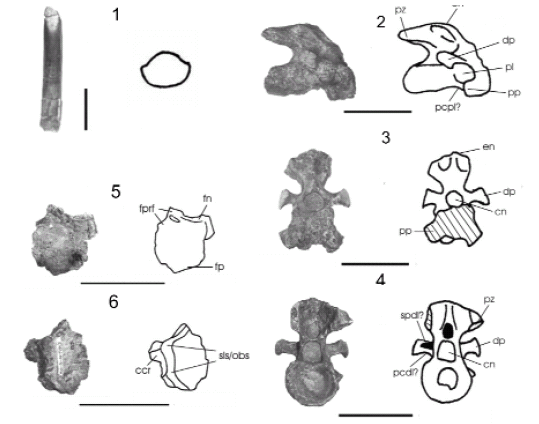
Figura 4. Pitekunsaurus macayai. 1, diente en vista lingual y sección (MAU-Pv-AG-446/2). 2-3, frontal izquierdo en vista dorsal (2) y ventral (3) (MAU-Pv-AG-446/5). 4-6, axis en vista lateral (4), anterior (5) y posterior (6) (MAU-Pv-AG-446/3)/ Pitekunsaurus macayai. 1, tooth in lingual view and section; 2-3, left frontal in dorsal (2) and ventral view (3); 4-6, axis in lateral (4), anterior (5) and posterior view (6). Escala gráfica: 10 cm, 1: 1 cm / scale bar: 10 cm, 1: 1 cm.
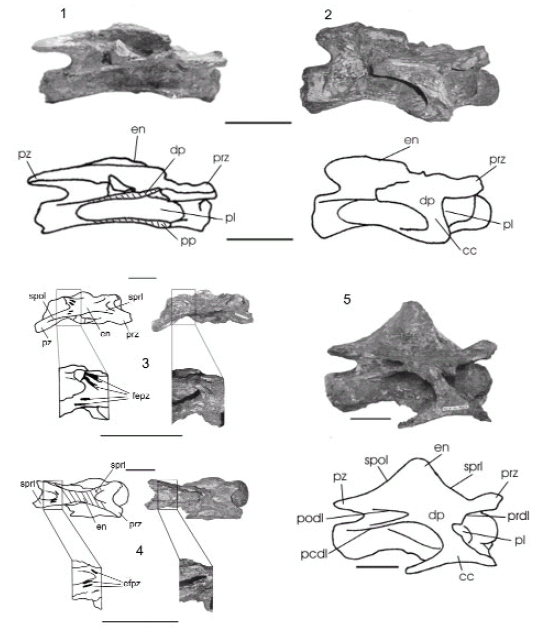
Figura 5. 1-2, Vértebras cervicales anteriores de Pitekunsaurus macayai en vista lateral (MAU-Pv-AG-446/4) y (MAU-Pv-AG-446/7) y 3-4, en vista dorsal. 5, vértebra cervical posterior en vista lateral (MAU-Pv-AG-446/6) / 1-2, Anterior cervical vertebra of Pitekunsaurus macayai in lateral view and 3-4, in dorsal view.5, Posterior cervical vertebra in lateral view. Escala gráfica / scale bar: 10 cm.
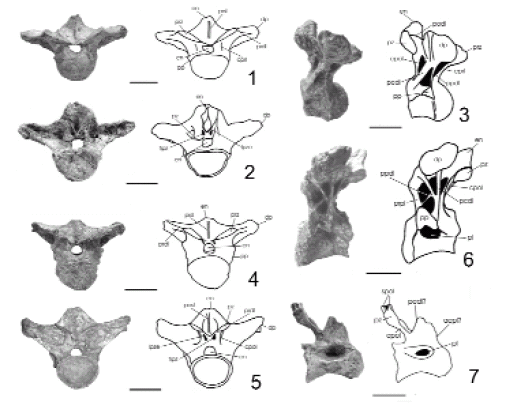
Figura 6. 1-6, Vértebras dorsales anteriores de Pitekunsaurus macayai (MAU-Pv-AG-446/8 y 9) en vista anterior (1 y 4), posterior (2 y 5) y lateral (3 y 6). 7, Vértebra dorsal posterior de Pitekunsaurus macayai (MAU-Pv-AG-446/30) en vista lateral. 1-6 Anterior dorsal vertebra of Pitekunsaurus macayai in anterior (1 y 4), posterior (2 y 5) and lateral view (3 y 6). 7 Posterior dorsal vertebra of Pitekunsaurus macayai y lateral view. Escala gráfica: 10 cm / scale bar: 10 cm.
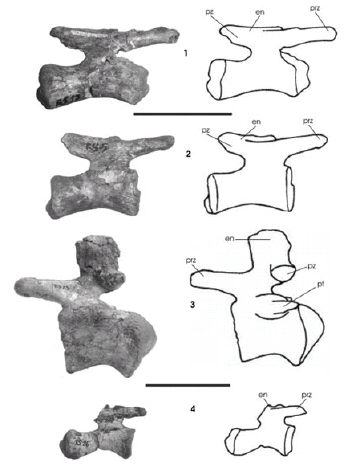
Figura 7. Pitekunsaurus macayai. 1, 2 y 4, vértebras caudales posteriores en vista lateral (MAU-Pv-AG-446/10, 11 y 24); 3, vértebra caudal anterior en vista lateral (MAU-Pv-AG-446/25). Pitekunsaurus macayai. 1, 2 and 4, posterior caudal vertebrae in lateral view; 3, anterior caudal vertebra in lateral view. Escala gráfica / scale bar: 10 cm.
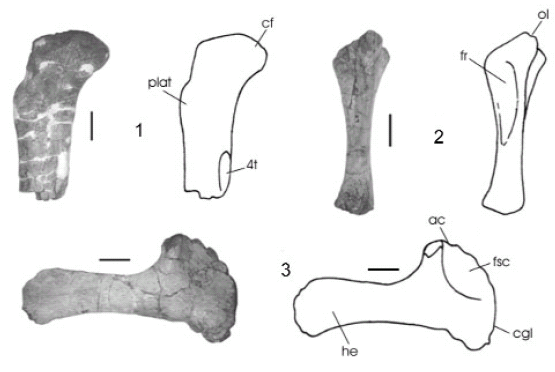
Figura 8. Pitekunsaurus macayai. 1, porción proximal de fémur izquierdo en vista posterior (MAU-Pv-AG-446/1); 2, ulna derecha en vista anterolateral (MAU-Pv-AG-446/27); 3, escápula derecha en vista lateral (MAU-Pv-AG-446/28). Pitekunsaurus macayai. 1, proximal portion of left fémur in posterior view; 2, right ulna in anteroventral view; 3, right scapula in lateral view. Escala gráfica / scale bar: 10 cm.
Etimología. Macayai, en reconocimiento al Sr. Luis Macaya, quien desinteresadamente denunció los restos al Museo Municipal "Argentino Urquiza" en abril del 2004.
Diagnosis. Esta especie se basa en las siguientes autapomorfías: (1) procesos basipterigoides ampliamente separados y proyectados paralelamente, (2) presencia en la unión de la base de la espina neural y lámina espinopostzigapofiseal de pequeñas depresiones o fosas longitudinales en vértebras cervicales anteriores, (3) lámina centropostzigapofiseal bifurcada proximalmente en vértebras dorsales anteriores, y (4) lámina centrodiapofiseal posterior con lámina accesoria en vértebras dorsales anteriores.
Diagnosis. This genus is characterized by the following autapomorphies: (1) basipterygoid processes broadly separated and parallely projected, (2) presence of small depressions or longitudinal grooves in the spinal sector of spinopostzygapophyseal lamina in anterior cervical vertebrae, (3) centropostzygapophyseal lamina forked proximally in anterior dorsal vertebrae, and (4) posterior centrodiapophyseal lamina with accessory lamina in anterior dorsal vertebrae.
Holotipo. Basicraneo y frontal izquierdo, MAU-Pv-AG-446/5; un diente, MAU-Pv-AG-446/2; axis MAU-Pv-AG-446/3; dos vértebras cervicales anteriores, MAU-Pv-AG-446/4 y MAU-Pv-AG-446/7; una vértebra cervical posterior, MAU-Pv-AG-446/6; dos vértebras dorsales anteriores, MAU-Pv-AG-446/8 y MAU-Pv-AG-446/9; una vértebra dorsal posterior, MAU-Pv-AG-446/30; una vértebra caudal anterior, MAU-Pv-AG-446/25; tres vértebras caudales posteriores, MAU-Pv-AG-446/10, MAU-Pv-AG-446/11 y MAU-Pv-AG-446/24; un fémur incompleto, MAU-Pv-AG-446/1; una escápula derecha, MAU-Pv-AG-446/28; y una ulna derecha MAU-Pv-AG-446/27.
Localidad. Cañadón de Aguada Grande (37°27'08" Sur; 68°56'36" Oeste), área Rincón de los Sauces, norte de la provincia del Neuquén, Argentina. Horizonte y edad. Grupo Neuquén (Cretácico Superior), Subgrupo Río Colorado, Formación Anacleto (Campaniano Inferior - Medio).
Descripción
Frontal. Se ha colectado un frontal izquierdo MAU-Pv-AG-446/5, algo incompleto pero bien preservado (figuras 4.2 y 4.3). Pese a no haber conservado su borde lateral izquierdo, se estima que es anteroposteriormente corto, similar a los de Rapetosaurus krausei (Curry Rogers y Forster, 2001, 2004) y Bonatitan reigi (Martinelli y Forasiepi, 2004). En el borde lateral interno, coincidente con la línea de sutura entre los frontales, existe una prominencia longitudinal similar a la presente en Antarctosaurus wichmannianus (Huene, 1929). En vista ventral, se observa la sutura para el complejo lateroesfenoides-orbitoesfenoides, que separa la cavidad craneana de la orbital, como en Rapetosaurus krausei.
Basicraneo. Se ha recuperado un basicráneo MAU-Pv-AG-446/5 incompleto pero bien preservado. Este material presenta los siguientes elementos constitutivos: supraoccipital, exoccipital-opistótico, proceso paraoccipital izquierdo, basioccipital, cóndilo craneal, tuberosidades basales, proótico, paraesfenoides, basiesfenoides y proceso basipterigoides (figura 3.1-4).
El supraoccipital exhibe una tuberosidad medial robusta, la cual se encuentra enmarcada por dos depresiones laterales. A diferencia de lo que se observa en Saltasaurus loricatus (Bonaparte y Powell, 1980; Powell, 1986, 1992), Quaesitosaurus orientalis (Kurzanov y Bannikov, 1983), Rapetosaurus krausei (Curry Rogers y Forster, 2001; 2004) Bonatitan reigi (Martinelli y Forasiepi, 2004) y Titanosauria indet. MAU-Pv-LL-207 (Calvo y González Riga, 2004), Pitekunsaurus macayai no presenta una depresión o cresta longitudinal sobre el plano sagital. El basicráneo MUCPv-334 descripto por Calvo y Kellner (2006) posee un supraoccipital similar, sin embargo, la tuberosidad medial de este último es mucho más prominente. Debido a la fusión que presenta los elementos del basicráneo MAU-Pv-AG-446/5, rasgo que comparten la mayoría de los titanosaurios (Calvo y Kellner, 2006), no pueden distinguirse fácilmente líneas de suturas entre los elementos mencionados. Sin embargo, se infiere que el supraoccipital forma el borde superior del foramen mágnum, además de la región posterodorsal de la caja craneana.
El exoccipital y opistótico se encuentran fusionados, resultando igualmente difícil interpretar los límites de cada uno de ellos. El foramen del nervio craneal V es grande, de forma elíptica y elongado dorsoventralmente, hallándose delimitado por el laterosfenoides y proótico. El nervio VI posee un pequeño foramen, ubicado ventralmente al foramen del nervio craneal V. La fisura metótica es de forma elíptica, con el eje mayor ubicado dorsoventralmente, pasando por ella los nervios craneales IX, X, XI y la vena yugular. Esto último es una diferencia con lo que se observa en el espécimen TMM 40435 correspondiente a un titanosauriforme indeterminado descripto por Tidwell y Carpenter (2003), en el cual el nervio accesorio XI, posee un pequeño foramen aparte, situado posteriormente a la fisura metótica. Como en otros titanosaurios, MAU-Pv-AG-446/5 presenta una fenestra ovalis ubicada anteriormente a la fisura metótica, como así también una crista prootica poco expandida lateralmente. El foramen del nervio craneal XII se ubica posteriormente cerca del borde inferior del exoccipital, entre la base de éste y el cuello del cóndilo occipital. El proceso paraoccipital es corto, presentándose proyectado lateroventralmente. Aunque su extremo distal esta incompleto, se estima la presencia de un ensanchamiento dorsoventral, similar al observado en Nemegtosaurus mongoliensis (Nowinski, 1971) y Brachiosaurus brancai (Janesch, 1935-36).
El basioccipital presenta un cóndilo de forma subesférica, con una pequeña concavidad en la cara dorsal similar al observado en Antarctosaurus wich-mannianus (Huene, 1929) y el titanosaurio indet. ISI R 162 descripto por Chatterjee y Rudra (1996), cuyo cóndilo occipital es de tamaño proporcional al foramen mágnum. Las tuberosidades basales se encuentran marcadamente expandidas o proyectadas lateralmente, presentando una forma subcircular como en Nemegtosaurus mongoliensis (Nowinski, 1971); con el diámetro mayor orientado transversalmente. Entre el foramen mágnum y las tuberosidades basales existe una amplia depresión, observándose posterodistalmente una suave escotadura triangular que separa ambas tuberosidades. Anteroventralmente a las tuberosidades basales se observa una depresión semilunar que se extiende lateralmente hacia los procesos basipterigoides, encontrándose separada medialmente por la cresta paraesfenoidal. En MAU-Pv-AG-446/5 solo se ha preservado la base del proceso cultriforme, siendo mucho menos elongado que el observado en el basicraneo MUCPv-334 descripto por Calvo y Kellner (2006).
El basiesfenoides exhibe procesos basipterigoides gráciles, de sección subcilíndrica y proyectados paralelamente entre sí, presentando en su región proximal una amplia separación subcuadrangular. En vista anteroventral se observa la fosa pituitaria (figuras 3.2 y 3.4), la cual posee un contorno dorsal semicircular y ventral triangular, con una delgada cresta que se continúa hacia la base del proceso cultriforme. En aquellos titanosauriformes que han preservado materiales craneanos como Brachiosaurus brancai (Janesch, 1935-36), Antarctosaurus wichmannianus (Huene, 1929), Nemegtosaurus mongoliensis (Nowinski, 1971) Titanosauria indet. (Coria y Salgado, 1999), Rapetosaurus krausei (Curry Rogers y Forster, 2001, 2004) Malawisaurus dixeyi (Gomani, 2005) y Bonatitan reigi (Martinelli y Forasiepi, 2004), la divergencia entre los procesos basipterigoides está siempre presente, variando en mayor o menor grado. La ausencia en Pitekunsaurus macayai de esta divergencia, sumada a la amplia separación entre los procesos, es considerada como autapomorfía para la especie.
Diente. El material MAU-Pv-AG-446/2 corresponde a una pieza delgada, de sección subcilíndrica y ligeramente curvada hacia la cara lingual (figura 4.1). Las carenas longitudinales se hallan escasamente desarrolladas. El ápice de la corona presenta una compresión labiolingual, manifestando una superficie de desgaste en bisel en su cara lingual. De acuerdo con Calvo (1994), este diente presentaría morfologías del tipo "lápiz-cincel" común entre los titanosaurios como Nemegtosaurus (Nowinski, 1971), Quaesitosaurus (Kurzanov y Banikov, 1983), y probablemente Salta-saurus (Bonaparte y Powell, 1980; Powell, 1986, 1992).
Axis. El material MAU-Pv-AG-446/3 (figura 4.4-6) corresponde a una pieza relativamente pequeña, corta y alta, presentando un centro opistocélico y espina neural robusta; muy similar al material correspondiente al Titanosauridae indeterminado Serie A de Peirópolis (Powell, 1986). La espina neural se encuentra más inclinada hacia atrás que en Saltasaurus loricatus (Bonaparte y Powell, 1980; Powell, 1986, 1992), exhibiendo una lámina prespinal bien definida y procesos laterales prominentes. El arco neural se ubica en la porción anterior de la vértebra, ocupando prácticamente toda la longitud de la misma. En la región posterior de la espina neural, entre las postzigapófisis y por encima del canal neural, se observa una fosa postespinal de diámetro proporcional al del canal neural. Las postzigapófisis son cortas, mostrando que la faceta articular preservada posee un contorno elíptico en sentido anteroposterior, encontrándose inclinada levemente hacia el plano axial. Las parapófisis se presentan como procesos bien desarrollados proyectados lateroposteriormente, exhibiendo pequeños pleurocelos sobre éstas en el sector anterior del centro vertebral. Las carillas articulares de estas últimas son subcirculares, hallándose notoriamente inclinadas medialmente. En la cara ventral, entre las parapófisis, se observa una amplia depresión, enmarcada por una lámina denominada aquí como centroparapofiseal posterior, la cual se continúa posteriormente sobre el borde lateroventral del centro vertebral. Sin embargo, la lámina centroparapofiseal posterior es considerada por Wilson (1999), como perteneciente solo a vértebras dorsales, por lo que dicha denominación puede no ser la adecuada. Las diapófisis se presentan como procesos subcirculares prácticamente planos, ubicados casi en posición vertical respecto del plano axial. En vista posterior, se observa del lado izquierdo una pequeña fosa subcircular, formada por las láminas espinodiapofiseal? y centrodiapofiseal posterior?
Vértebra cervical anterior. Se han recuperado dos vértebras cervicales anteriores MAU-Pv-AG-446/4 y MAU-Pv-AG-446/7 (figuras 5.1 y 5.2; 5.3 y 5.4), las cuales, de acuerdo con la morfología observada en el Titanosauridae indeterminado de la Serie A de Peirópolis (Powell, 1986), podrían corresponder a la 4ta? y 7ma? cervical respectivamente. Los cuerpos vertebrales son opistocélicos, con los extremos articulares bien definidos, más anchos que altos. En la 4ta? cervical se observa en la región anterior de la cara ventral una ligera concavidad, mientras que hacia el sector posterior se desarrolla una superficie ligeramente convexa; características que se expresan más suavemente en la 7ma? cervical. Sólo se han preservado la diapófisis y parapófisis derechas correspondientes a la 7ma? cervical junto a la porción proximal de la costilla cervical. La espina neural en la 4ta? cervical es baja y ha experimentado una deformación por aplastamiento del lado derecho, mientras que en la 7ma? cervical se encuentra anteriormente incompleta y meteorizada. Pese a su estado de preservación, se estima que la espina era relativamente baja. El arco neural es bajo y abarca prácticamente toda la longitud del centro vertebral. Las fosas pre y postespinales son profundas, siendo esta última proporcionalmente mayor. Sólo se han preservado los sectores proximales de las prezigapófisis y postzigapófisis en la 4ta? y 7ma? cervical, estando presente en la primera únicamente las del lado derecho. Tanto la prezigapófisis como la postzigapófisis son relativamente cortas, proyectándose lateralmente respecto del plano axial. La prezigapófisis se halla unida a la espina por una lámina espinoprezigapofiseal, que junto a la lámina interprezigapofiseal, une ambas sobre el techo del canal neural delimitando la fosa prespinal. La postzigapófisis preservada en 4ta? cervical supera levemente la cara articular posterior, exhibiendo una lámina interpostzigapofiseal que hace de piso a la fosa postespinal. La faceta articular de la postzigapófisis es subcircular y se inclina levemente hacia el plano axial. En el sector espinal de lámina espinopostzigapofiseal (figura 5.3 y 5.4) se observan pequeñas depresiones o fosas longitudinales, rasgo considerado aquí como autapomórfico de Pitekunsaurus macayai.
Vértebra cervical posterior. Se ha preservado la mitad derecha de una cervical posterior MAU-PV-AG-446/6 (figura 5.5), que se asigna a la 11ra cervical según comparaciones con el ejemplar de Titanosauridae indeterminado Serie A de Peirópolis (Powell, 1986). Presenta un centro vertebral opistocélico, relativamente más largo que los observados en otros titanosaurios como Isisaurus colberti (Jain y Bandyopadhyay, 1997) y Mendozasaurus neguyelap (González Riga, 2005), quienes poseen centros cervicales cortos asociados con altas espinas neurales. Lateralmente se observa una marcada depresión longitudinal, en la cual se ubica el pleurocelo. La espina neural es alta y robusta, de contorno triangular en vista lateral, con tuberosidades laterales bien marcadas delimitadas ventralmente por dos crestas longitudinales someramente definidas. Si bien la preservación del material es pobre, se observa que las fosas prespinal y postespinal son profundas, siendo esta última la de mayor tamaño. El sector anterior de la cara ventral del centro es ancho y anteroposteriormente cóncavo. La parapófisis y diapófisis se presentan como proyecciones laminares laterales subtriangulares, ubicadas en la mitad anterior del centro vertebral y orientadas levemente hacia abajo. La diapófisis esta reforzada por la lámina prezigodiapofiseal, postzigodiapofiseal y centrodiapofiseal posterior, encontrándose ubicada en un nivel levemente inferior al de las prezigapófisis. La lámina centrodiapofiseal anterior no se ha preservado. En vista lateral (figura 7), como ocurre en Trigonosaurus pricei (Campos et al., 2005), las láminas postzigodiapofiseal y centrodiapofiseal posterior divergen delimitando una profunda fosa. En Rinconsaurus caudamirus (Calvo y González Riga, 2003), la lámina centrodiapofiseal posterior diverge respecto de la lámina postzigodiapofiseal, con un ángulo mayor, que le permite contactarse con la lámina centroparapofiseal posterior en el borde posteroventral del centro vertebral. Las láminas centrodiapofiseal posterior y centroparapofiseal posterior se proyectan anteroposteriormente de forma paralela. En este nuevo ejemplar, y en otros titanosaurios como Rinconsaurus, el ejemplar MAU-PV-AC-01, Malawisaurus dixeyi (Gomani, 2005) y Trigonosaurus, presenta prezigapófisis largas y postzigapófisis cortas. Las facetas articulares de la prezigapófisis y postzigapófisis son subcirculares, encontrándose la primera posteromedialmente orientada, y la segunda anterolateralmente dirigida.
Vértebra dorsal anterior. Se colectaron dos vértebras dorsales anteriores en buen estado de preservación, las cuales, de acuerdo con la morfología observada en el ejemplar de Titanosauridae indeterminado de la Serie A de Peirópolis (Powell, 1986), son interpretadas como la 1ra dorsal MAU-Pv-AG-446/9 (figura 6.1-3) y 2da? dorsal MAU-Pv-AG-446/8 (figura 6.4-6). Ambas poseen un centro vertebral fuertemente opistocélico, con profundos pleurocelos acuminados. El arco neural es corto y ancho, situado en la mitad del centro vertebral. La parapófisis se ubica en la cara lateral del mismo y sobre la base del arco neural, disponiéndose dorsoventralmente extendida con el desarrollo de una corta lámina centroparapofiseal anterior. Las prezigapófisis y postzigapófisis son de forma subcircular, con el diámetro lateromedial mayor que el anteroposterior e inclinadas medialmente. Las diapófisis son largas y robustas, encontrándose proyectadas lateralmente en la 2da? dorsal en un ángulo levemente mayor que en la 1ra dorsal. Éstas se encuentran reforzadas por las láminas espinodiapofiseales, prezigodiapofiseales, paradiapofiseales y centrodiapofiseales posteriores. La lámina centropostzigapofiseal se encuentra bifurcada proximalmente en la 2da? dorsal (figura 6.5), carácter que es considerado como una autapomorfía de Pitekunsaurus macayai. La lámina postzigodiapofiseal no es claramente distinguible. En la 1ra dorsal (figura 6.3), la lámina centrodiapofiseal posterior y paradiapofiseal delimitan una fosa infradiapofiseal de contorno subtriangular, de forma similar a Mendozasaurus neguyelap (González Riga, 2003; figura 4.B). La lámina centrodiapofiseal posterior de la 2da? dorsal (figura 6.6) presenta una lámina accesoria que se proyecta desde la mitad de la misma hasta conectar distalmente en el sector anterior a la diapófisis, observándose lateralmente entre ambas láminas, una profunda fosa subtriangular; carácter que es considerado como una autapomorfía para Pitekunsaurus macayai. La lámina paradiapofiseal se halla unida a esta última lámina accesoria (figura 6.6). En Rinconsaurus caudamirus (Calvo y González Riga, 2003) se observa también una lámina accesoria, pero ubicada paralelamente a la lámina centrodiapofiseal. La lámina centroprezigapofiseal es simple, más reducida en la 2da? dorsal. En ambas dorsales existe una reducción del espacio entre el techo del canal neural y la base de la espina neural, lo cual provoca que prácticamente ambas estructuras estén unidas. La espina neural es corta, se encuentra algo inclinada hacia atrás, superando el borde posterior del centro vertebral en la 2da? dorsal; mientras en la 1ra dorsal la espina neural se encuentra ubicada en posición vertical. Las láminas prespinales y postespinales de la 2da? dorsal son robustas y bien definidas, extendiéndose hasta la base de la espina neural; mientras que en la 1ra dorsal la lámina prespinal culmina antes de la base de la espina, no habiéndose preservado la lámina postespinal. Otro carácter observado en la 1ra dorsal (figura 6.2) y compartido con Mendozasaurus neguyelap, es la presencia de una lámina postzigapostespinal ubicada paralelamente a las facetas de las postzigapófisis, la cual conecta la superficie de éstas con la base de la lámina postespinal (González Riga, 2003). En la 2da? dorsal (figura 6.5) el ángulo de inclinación de dichas láminas postzigapostespinales varía levemente respecto de las facetas de las postzigapófisis. Entre las postzigapófisis y el techo del arco neural, se observa la presencia de dos fosas infrapostzigapofiseales de contorno ovalado, similares a las descriptas en Mendozasaurus neguyelap.
Vértebra dorsal posterior. Se ha recuperado una vértebra dorsal posterior incompleta MAU-Pv-AG-446/30 (figura 6.7), interpretada como la 8va? dorsal de acuerdo a la morfología observada en el ejemplar de Titanosauridae indeterminado Serie A de Peirópolis (Powell, 1986). Este material preserva únicamente su lado derecho, mostrando un cuerpo vertebral marcadamente opistocélico. La cara lateral y ventral son cóncavas en sentido anteroposterior, observándose una profunda depresión donde se ubica el pleurocelo. La superficie articular posterior es levemente más alta que la anterior. Sobre el pleurocelo y entre las láminas centroparapofiseal anterior y centrodiapofiseal posterior, en la base del arco neural, se presenta una amplia superficie someramente cóncava en sentido anteroposterior, similar a lo observado en Malawisaurus dixeyi (Gonami, 2005). Entre las láminas centrodiapofiseal posterior y centropostzigapofiseal, se observa una profunda fosa de contorno triangular. La postzigapófisis preservada se encuentra orientada lateroposteriormente, existiendo sobre ella una profunda fosa denominada aquí como suprapostzigapofiseal. Esta fosa se encuentra delimitada por dos ramas de la base de la lámina espinopostzigapofiseal (Wilson, 1999), las cuales conectan a la postzigapófisis con la espina neural. Al igual que en las dorsales anteriores, esta dorsal posterior exhibe el desarrollo de una lámina postespinal.
Vértebra caudal anterior. Se ha recuperado una vértebra caudal anterior bien preservada MAU-Pv-AG-446/25 (figura 7.3), cuyo centro vertebral procélico posee sus caras lateral y ventral suavemente cóncavas en sentido anteroposterior. El alto y ancho del centro vertebral es proporcional. Las carillas articulares para las hemapófisis están bien desarrolladas. Los procesos transversos son delgados, posterolateralmente orientados. El arco neural es corto, ubicándose en la porción anterior del centro e inclinado ligeramente hacia adelante. Las prezigapófisis están levemente proyectadas dorsoanteriormente, aunque de manera menos marcada que en Rinconsaurus caudamirus (Calvo y González Riga, 2003). Las carillas articulares de las prezigapófisis y postzigapófisis son subcirculares, y se orientan paralelas al plano axial. Las postzigapófisis, ubicadas en la mitad posterior del centro vertebral, se encuentran unidas a la base de la espina neural por procesos postzigapofiseales similares a los presentes en Rinconsaurus caudamirus (Calvo y González Riga, 2003), aunque menos desarrollados. La espina neural es corta, sus bordes anterior y posterior son rectos, encontrándose suavemente inclinada anteriormente. Junto a esta caudal se hallaron fragmentos de hemapófisis, los cuales indicarían que los arcos hemales se encontraban abiertos proximalmente como sucede en todos los titanosaurios.
Vértebra caudal posterior. Se han recuperado dos vértebras caudales posteriores en buen estado de preservación, MAU-Pv-AG-446/10 y MAU-Pv-AG-446/11 (figura 7.1-2). Ambas poseen las caras laterales y ventrales del centro vertebral cóncavas en sentido anteroposterior. La vértebra caudal MAU-Pv-AG-446/10 es biconvexa, con ambos cóndilos prominentes; mientras que la caudal MAU-Pv-AG-446/11 es anficélica, presentando un cóndilo articular posterior reducido y ventralmente desplazado. Este último cóndilo podría ser interpretado como un vestigio de anillo cartilaginoso intervertebral osificado o como un desarrollo patológico. En ambos casos, el arco neural es mas bajo que en Rinconsaurus caudamirus (Calvo y González Riga, 2003), disponiéndose sobre la porción anterior del centro vertebral. Las prezigapófisis son estructuras subcilíndricas mas bien cortas, anteriormente proyectadas y levemente orientadas dorsalmente. Las facetas articulares de las prezigapófisis son subcirculares y se orientan casi paralelamente al plano axial. La espina neural es baja, proyectada posteriormente y levemente inclinada dorso-distalmente. Sobre esta última se ubican las postzigapófisis, cuyas facetas articulares son apenas notorias. Un tercer material (MAU-Pv-AG-446/26) (figura 7.4), corresponde a una pequeña caudal distal, fuertemente procélica, exhibe un cóndilo posterior prominente. El centro vertebral es más largo que alto, con las caras lateral y ventral notoriamente cóncavas en sentido anteroposterior. El arco neural es bajo, ubicado en la mitad anterior del centro. Las prezigapófisis son extremadamente cortas, y se encuentran proyectadas anteriormente paralelas al centro vertebral, apenas superando la cara articular. La espina neural no se ha preservado, pero la evidencia sugiere que ha sido baja, orientándose posteriormente en forma paralela al centro. Pese a que se han recuperado caudales aisladas, es posible advertir un cambio poco usual en el tipo de articulación a lo largo de la secuencia, cuya presencia de centros anficélicos y biconvexos en las caudales posteriores, es un carácter compartido con Rinconsaurus caudamirus (Calvo y González Riga, 2003).
Ulna. La pieza recuperada MAU-Pv-AG-446/27 (figura 8.2) posee un buen estado de preservación, con una longitud total de 48 cm. Se trata de un hueso recto y robusto, con un mayor desarrollo en su extremo proximal. La superficie proximal del cóndilo posee una forma triradiada, con un buen desarrollo del olécranon y una profunda fosa radial. En la cara radial se observa una cresta longitudinal de aproximadamente unos 20 cm de longitud. La sección del extremo distal posee una forma subredondeada.
Escápula. El material MAU-Pv-AG-446/28 corresponde a una escápula derecha (figura 8.3), con una longitud total de 69 cm. El extremo distal de la hoja escapular muestra una expansión anteroposterior moderada, insinuándose una morfología más bien redondeada. Los bordes de la hoja escapular son prácticamente rectos, con una leve concavidad longitudinal más marcada en el borde ventral, similar a lo observado en el ejemplar MCNA-7459 de Lirainosaurus astibiae (Sanz, et al., 1999) y diferente de lo observado en Mendozasaurus neguyelap (González Riga, 2003) donde el borde anterodorsal de la hoja, es cóncavo en su sección proximal y recto en su sección distal. La hoja escapular mantiene longitudinalmente el mismo ancho, haciéndose sutilmente más estrecha hacia su parte media. Sobre el margen dorsal de la hoja escapular se observa una prominencia medial poco desarrollada, similar a la presente en Lirainosaurus astibiae (Sanz, et al., 1999). El proceso supraglenoide es prominente, mostrándose muy ancho transversalmente en relación con la longitud total de la escápula.
Fémur. Se ha recuperado la porción proximal del fémur izquierdo MAU-Pv-AG-446/1 (figura 8.1), cuya longitud preservada alcanza los 57 cm. El cuarto trocánter, en forma de lomo, se ubica pocos centímetros por debajo y levemente desplazado respecto de la protuberancia lateral. Dicha protuberancia lateral, ubicada debajo del trocánter mayor, se desarrolla desde el nivel de la unión entre la epífisis y la diáfisis, siendo menos extensa longitudinalmente que en Rinconsaurus caudamirus (Calvo y González Riga, 2003). La cabeza femoral manifiesta una marcada curvatura lateral.
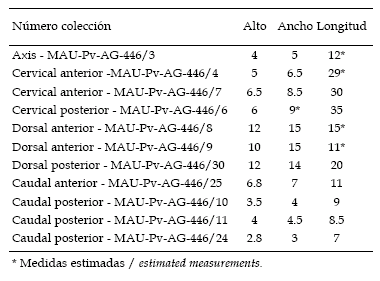
Cuadro 2. Medidas de las vértebras de Pitekunsaurus macayai (cm) / measurements of vertebrae of Pitekunsaurus macayai (cm).
Discusión y conclusiones
Si bien a lo largo de los últimos años han sido recuperados en la Cuenca Neuquina numerosos restos de titanosaurios, son relativamente escasos los ejemplares descriptos correspondientes al lapso Santoniano - Campaniano Medio (Grupo Neuquén, Subgrupo Río Colorado). Hasta el momento se contaba únicamente con los registros de Laplatasaurus araukanikus (Huene, 1929), Pellegrinisaurus powelli (Salgado, 1996), Neuquensaurus australis (Salgado et al., 2005) y Bonitasaura salgadoi (Apesteguía, 2004). En este contexto, Pitekunsaurus macayai gen. et sp. nov., aporta información sistemática novedosa para el clado Titanosauria.
Pitekunsaurus macayai gen. et sp. nov., presenta los siguientes caracteres diagnósticos de Titanosauriformes (grupo de saurópodos que incluye a Brachiosaurus, Saltasaurus, su ancestro común y todos sus descendientes, sensu Salgado et al., 1997): centros cervicales muy elongados (Wilson, 2002), arcos neurales posicionados anteriormente en centros caudales medios y posteriores, presencia de lámina prespinal en vértebras dorsales (Salgado et al., 1997), costillas dorsales con cavidades neumáticas y tercio superior de la diáfisis del fémur desviada medialmente y presencia de una protuberancia lateral (Salgado et al., 1997; Wilson y Sereno, 1998).
Asimismo, se identifican los siguientes caracteres diagnósticos de Titanosauria (Bonaparte y Coria, 1993), grupo definido como el clado que incluye al más reciente ancestro común de Andesaurus delgadoi (Calvo y Bonaparte, 1991) y Saltasaurus loricatus (Bonaparte y Powell, 1980), así como todos sus descendientes: centros cervicales con diapófisis y parapófisis proyectadas lateralmente y vértebras dorsales con láminas prespinal y postespinal robustas (Upchurch, 1999).
La ausencia de algunos caracteres sinapomórficos derivados tales como centros caudales bajos con cavidades ventrales separadas en algunos casos por un septo, como los señalados en Saltasaurus loricatus (Bonaparte y Powell, 1980; Powell, 1986; 1992) y Rocasaurus muniozi (Salgado y Azpilicueta, 2000); permite excluir a Pitekunsaurus macayai gen. et sp. nov. del clado Saltasaurinae (sensu Bonaparte y Powell, 1980; Powell, 1992).
Los procesos basipterigoides en Pitekunsaurus macayai gen. et sp. nov. están notoriamente separados entre sí y proyectados paralelamente, algo que difiere notablemente de los materiales disponibles de otros titanosauriformes conocidos (donde existe un ángulo de divergencia que puede variar entre los 20º y 45º). La condición de este carácter, sobre todo la amplia separación entre los procesos basipterigoides, podría representar en Pitekunsaurus, una condición derivada dentro de Titanosauria.
Otra característica importante en Pitekunsaurus macayai gen. et sp. nov., es la presencia en vértebras cervicales anteriores, de pequeñas depresiones o fosas longitudinales en el sector espinal de lámina espinopostzigapofiseal. Aun cuando se desconoce la función de estas estructuras, no se registra nada similar en otros titanosaurios que han preservado secuencias o elementos cervicales completos.
La lámina centrodiapofiseal posterior en la 2da? vértebra dorsal MAU-Pv-AG-446/8 de Pitekunsaurus macayai gen. et sp. nov. presenta una lámina accesoria. Esta se proyecta desde la mitad de lámina centrodiapofiseal posterior hasta conectar distalmente en el sector anterior a la diapófisis, sobre el cual se ubica la lámina paradiapofiseal. En Rinconsaurus caudamirus (Calvo y González Riga, 2003) la lámina accesoria esta presente, pero se ubica paralelamente a la lámina centrodiapofiseal posterior. Es de destacar además que en Pitekunsaurus la lámina centropostzigapofiseal posterior se encuentra bifurcada proximalmente, característica no registrada en otros titanosaurios conocidos, razón por la cual es considerada como un carácter autapomórfico.
Pese a que no se han recuperado elementos caudales articulados, Pitekunsaurus macayai gen. et sp. nov., manifiesta una notable variación morfológica de la articulación entre los centros vertebrales a lo largo de la secuencia caudal, representada por una vértebra caudal anterior procélica MAU-Pv-AG-446/25, una caudal posterior anficélica MAU-Pv-AG-446/11, otra biconvexa MAU-Pv-AG-446/10 y una caudal posterior mas terminal procélica MAUPv-AG-446/24. Dichos cambios, se observan en Rinconsaurus caudamirus (Calvo y González Riga, 2003), en el cual se registran mejor documentadas, porciones de la secuencia caudal con centros procélicos-anficélicos-biconvexos y centros opistocélicos-biconvexos-procélicos, variaciones que han sido considerados por Calvo y González Riga (2003) como caracteres autapomórficos para dicha especie. La presencia de este carácter, podría estar indicando la existencia de un vinculo entre Pitekunsaurus macayai gen. et sp. nov. y Rinconsaurus caudamirus (Calvo y González Riga, 2003), especies que pudieron haber integrando dentro de Titanosauria, un grupo particular poseedor de una morfología caudal más compleja. La ejecución de un análisis cladístico detallado, podrá arrojar conclusiones más precisas en la corroboración de dicha hipótesis.
Agradecimientos
Deseamos expresar nuestro agradecimiento a J.I. Canudo, a L. Salgado, a R.A. Coria, a A. Arcucci, a A.P. Carabajal, a A. Cambiaso y a B. González Riga, por la lectura, comentarios y observaciones realizadas en el manuscrito. A L. Macaya, por denunciar el hallazgo; a A. Hernández por recibirnos cordialmente y permitirnos ingresar en su campo durante los trabajos de rescate, como así también a los técnicos del Museo Municipal "Argentino Urquiza" (Rincón de los Sauces), a C. Fuentes, A. Paillán, S. Palomo y A. Valenzuela (┼2005), por su asistencia durante los trabajos de campo y preparación de los materiales. Finalmente nuestro agradecimiento a la Municipalidad de Rincón de los Sauces (provincia del Neuquén) por el apoyo logístico brindado.
Bibliografía
1. Apesteguía, S. 2004. Bonitasaura salgadoi gen. et sp. nov. : A break sauropod from the Late Cretaceous of Patagonia. Natur-wissenchaften 91: 493-497. [ Links ]
2. Bentham, P.A., Talling, P.J. y Burbank, D.W. 1993. Braided stream flood-plain deposition in a rapidly aggrading basin: The Escanilla Formation, Spanish Pyrenees. En: J.L. Bests y C.S. Bristow (eds.), Braided Rivers. Geological Society, Special Publication 75: 177-194. [ Links ]
3. Bonaparte, J.F. y Coria, R.A. 1993. Un nuevo y gigantesco saurópodo titanosaurio de la Formación Río Limay (Albiano-Cenomaniano) de la Provincia del Neuquén, Argentina. Ameghiniana 30: 271-282. [ Links ]
4. Bonaparte, J.F. y Powell, J.E. 1980. A continental assemblage of tetrapods from the upper cretaceous beds of El Brete, North-western Argentina (Sauropoda-Coelurosauria-Carnosauria-Aves). Mémoires de le Société Géologique de France, N. S. 59: 18-28. [ Links ]
5. Calvo, J.O. 1994. Jaw mechanics in Sauropod dinosaurs. En: M.G. Lockley, V.F. Santos y C.A. Meyer (eds.), Aspects of Sauropod Paleobiology. GAIA 10: 183-193. [ Links ]
6. Calvo, J.O. 2002. Un gigantesco Titanosauridae (Cretácico Superior) del norte de la provincia de Neuquén. Ameghiniana Suplemento Resúmenes 39: 7R. [ Links ]
7. Calvo, J.O. y Bonaparte, J.F. 1991. Andesaurus delgadoi gen et sp. nov. (Saurischia-Sauropoda), dinosaurio Titanosauridae de la Formación Río Limay (Albiano-Cenomaniano), Neuquén, Argentina. Ameghiniana 28: 303-310. [ Links ]
8. Calvo, J.O. y González Riga, B.J. 2003. Rinconsaurus caudamirus gen. et sp. nov., A new titanosaurid (Dinosauria, Sauropoda) from the Late Cretaceous of Patagonia, Argentina. Revista Geológica de Chile 30: 333-353. [ Links ]
9. Calvo, J.O. y González Riga, B.J. 2004. A new braincase from Neuquén province, northern Patagonia, Argentina. Ameghiniana Suplemento Resúmenes 41: 38R. [ Links ]
10. Calvo, J.O. y Kellner A.W. 2006. Description of a sauropod dinosaur braincase (Titanosauridae) from the Late Cretaceous Rio Colorado Subgroup, Patagonia. Anais da Academia Brasileira de Ciencias 78: 175-182. [ Links ]
11. Calvo, J.O., Coria, R.A. y Salgado, L. 1997. Uno de los más completos titanosáuridos (Dinosauria-Sauropoda) registrados en el mundo. Ameghiniana 34: 534. [ Links ]
12. Campos, D., Kellner, A.W., Bertini, R.J. y Santucci, R.M. 2005. On a titanosaurid (Dinosauria-Sauropoda) vertebral column from the Bauru Group, Late Cretaceous of Brazil. Arquivos do Museu Nacional (Rio de Janeiro) 63: 565-593. [ Links ]
13. Chatterjee, S. y Rudra, D.K. 1996. K/T events in India: Impact, rifting, volcanism and dinosaur extinction. Memoirs of the Queensland Museum 39: 489-532. [ Links ]
14. Collinson, J.D. 1986. Alluvial sediments. En: H.G. Reading (ed.), Sedimentary environments: processes, facies and stratigraphy. 3º Edition. Blackwell Science, Oxford, pp. 37-82. [ Links ]
15. Coria, R.A. y Salgado, L. 1999. Nuevos aportes a la anatomía craneana de los saurópodos titanosáuridos. Ameghiniana 36: 98. [ Links ]
16. Curry Rogers, K. y Forster, C.A. 2001. The last of the dinosaur titans: a new sauropod from Madagascar. Nature 412: 530-534. [ Links ]
17. Curry Rogers, K. y Forster, C.A. 2004. The skull of Rapetosaurus krausei (Sauropoda: Titanosauria) from the Late Cretaceous of Madagascar. Journal of Vertebrate Paleontology 24: 121-144. [ Links ]
18. Espósito, M. 2003. Diccionario Mapuche. Editorial Guadal S.A., Buenos Aires, 350 pp. [ Links ]
19. Filippi, L.S. y Garrido, A.C. 2006. Nuevos restos de titanosaurios (Saurischia, Sauropoda) en el Cretácico Superior del norte de Patagonia, Rincón de los Sauces, Neuquén, Argentina. Ameghiniana Suplemento Resúmenes 43: 39R. [ Links ]
20. Garrido, A.C. 2000. [Estudio estratigráfico y reconstrucción paleoambiental de las secuencias fosilíferas continentales del Cretácico Superior en las inmediaciones de Plaza Huincul, provincia del Neuquén. Trabajo Final para el Título de Grado en Ciencias Geológicas. Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba, Córdoba, Argentina, 78 pp. Inédita.]. [ Links ]
21. Gomani, E. 2005. Sauropod dinosaur from the Early Cretaceous of Malawi, África. Palaeontologia Electronica 8 (1) 27A: 37p. Web: http://paleoelectronica.org/palaeo/2005_1/gomani27/is-sue1_05.htm. [ Links ]
22. González Riga, B.J. 2003. A new titanosaur (Dinosauria, Sauropoda) from the Upper Cretaceous of Mendoza Province, Argentina. Ameghiniana 40: 155-172. [ Links ]
23. González Riga, B.J. 2005. Nuevos restos fósiles de Mendozasaurus neguyelap (Sauropoda, Titanosauria) del Cretácico tardío de Mendoza, Argentina. Ameghiniana 42: 535-548. [ Links ]
24. González Riga, B.J. y Calvo, J.O. 2001. A new genus and species of Titanosaurid Sauropod from the Upper Cretaceous of Rincón de los Sauces, Neuquén, Argentina. Journal of Vertebrate Paleontology 21 (Supl.): 55A. [ Links ]
25. Haszeldine, R.S. 1983. Descending tabular crossbed sets and bounding surfaces from a fluvial channel in the Upper Carboniferous coalfield of north-east England. En: J.D. Collinson y J. Lewin (eds.), Modern and Ancient Fluvial Systems. International Association of Sedimentologists, Special Publication 6: 405-420. [ Links ]
26. Huene, F. 1929. Los Saurisquios y Ornithisquios del Cretácico Argentino. Anales del Museo de La Plata, 2° Serie 3: 1-196. [ Links ]
27. Janesch, W. 1935-36. Die Schädel der Sauropoden Bachiosaurus, Barosaurus und Dicraeosaurus, aus den Tendanguru-Schichten Deutsch-Ostafrikas. Palaeontographica part 2: 147-298. [ Links ]
28. Jain, S.L. y Bandyopadhyay, S. 1997: New Titanosaurid (Dinosauria: Sauropoda) from the Late Cretaceous of Central India. Journal of Vertebrate Paleontology 17: 114-136. [ Links ]
29. Kurzanov, S.M. y Bannikov, A.F. 1983. A new sauropod from the Upper Cretaceous of Mongolia. Paleontologicheskyy Zhurnal 2: 91-97. [ Links ]
30. Legarreta, L. y Gulisano, C.A. 1989. Análisis estratigráfico secuencial de la Cuenca Neuquina (Triásico superior - Terciario inferior). En: G. Chebli y L. Spalletti (eds.), Cuencas Sedimentarias Argentinas. Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán. Serie Correlación Geológica N° 6: 221-243. [ Links ]
31. Martinelli, A.G. y Forasiepi, A.M. 2004. Late Cretaceous vertebrates from Bajo de Santa Rosa (Allen Formation), Río Negro province, Argentina, with the description of a new sauropod dinosaur (Titanosauridae). Revista Museo Argentino de Ciencias Naturales, Nueva Serie 6: 257-305. [ Links ]
32. Miall, A.D. 1996. The geology of fluvial deposits. Sedimentary facies, basin analysis, and petroleum geology. Springer, New York, 582 pp. [ Links ]
33. Nowinski, A. 1971. Nemegtosaurus mongoliensis nov. gen., nov. sp. (Sauropoda) from the Uppermost Cretaceous of Mongolia. Acta Palaeontologica Polonica 25: 57-81. [ Links ]
34. Powell, J.E. 1986. [Revisión de los Titanosaurios de América del Sur. Tesis Doctoral, Universidad Nacional de Tucumán, Argentina, 340 pp. Inédita.]. [ Links ]
35. Powell, J.E. 1992. Osteología de Saltasaurus loricatus (Saurópoda-Titanosauridae) del Cretácico Superior del Noroeste argentino. En: J.L. Sanz, y A.D. Buscalioni (ed.), Los Dinosaurios y su entorno Biótico. Insituto "Juan de Valdes", Cuenca, España, pp. 165-230. [ Links ]
36. Powell, J.E. 2003. Revision of South American Titanosaurid dinosaur: palaeobiological, palaeobiogeoraphical and phylogenetic aspects. Records of the Queen Victoria Museum 111: 1-173. [ Links ]
37. Salgado, L. 1996. Pellegrinisaurus powelli nov. gen. et sp. (Saurópoda, Titanosauridae) from the Upper Cretaceous of Lago Pellegrini, Northwersten Patagonia, Argentina. Ameghiniana 33: 335-365. [ Links ]
38. Salgado, L. 2003. Los saurópodos de Patagonia: Sistemática, Evolución y Paleobiogeografía. En: C.A.S. (ed.), Colectivo Arqueológico-Paleontológico de Sala. 2º Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno. Salas de los Infantes (Burgos, España), Actas, pp. 139-168. [ Links ]
39. Salgado, L. y Azpilicueta, C. 2000: Un nuevo saltasaurino (Sauropoda, Titanosauridae) de la provincia de Río Negro (Formación Allen, Cretácico Superior), Patagonia, Argentina. Ameghiniana 37: 259-264. [ Links ]
40. Salgado, L., Apesteguía, S., y Heredia, S.E. 2005. A new especimen of Neuquensaurus australis, a Late Cretaceous Saltasaurine Titanosaur from North Patagonia. Journal of Vertebrate Paleontology 25: 623-634. [ Links ]
41. Salgado, L., Coria, R.A. y Calvo, J.O. 1997. Evolution of titanosaurid sauropods I: Phylogenetic análisis based on the postcraneal evidence. Ameghiniana 34: 3-32. [ Links ]
42. Sanz, J.L., Powell, J., Le Loeuff, J., Martinez, R. y Pereda Suberbiola, X. 1999. Sauropod remains from the upper cretaceous of Laño (Northcentral Spain) titanosaur phylogenetic relationships. Estudios del Museo de Ciencias Naturales de Alava 14 (Número Especial) 1: 235-255. [ Links ]
43. Spalletti, L.A. 1987. Características y significado sedimentológico de las formas, cuerpos sedimentarios y estructuras originadas por agentes fluidos en sustratos incoherentes. Revista del Museo de La Plata (Nueva Serie) 10, Geología 82: 1-65. [ Links ]
44. Tidwell, V. y Carpenter, K. 2003. Braincase of an early Cretaceous Titanosauriform Sauropod from Texas. Journal of Vertebrate Paleontology 23: 176-180. [ Links ]
45. Upchruch, P. 1999. The phylogenetcis relationships of the Nemegtosauridae (Saurischia: Sauropoda). Journal of Vertebrate Paleontology 19: 106-125. [ Links ]
46. Walker, R.G. y Cant, D.J. 1984. Sandy Fluvial Systems. En: R. Walker (ed.), Facies Models. Geosciencie. Canada. Reprint Series 1: 71-89. [ Links ]
47. Wilson, J.A. y Sereno, P.C. 1998. Early Evolution and Higher-level Phylogeny of Sauropod Dinosaurs. Journal of Vertebrate Paleontology 18, Supplement 2: 1-68 [ Links ]
48. Wilson, J.A. 1999. A nomenclature for vertebral laminae in sauropods and other saurischian dinosaurs. Journal of Vertebrate Paleontology 19: 639-653. [ Links ]
49. Wilson, J.A. 2002. Sauropod dinosaur phylogeny: critique and cladistic analysis. Zoological. Journal of the Linnean Society 136: 217-276. [ Links ]
Recibido: 7 de diciembre de 2006.
Aceptado: 10 de julio de 2008.

