










Services on Demand
Journal

Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAmeghiniana
On-line version ISSN 1851-8044
Ameghiniana vol.45 no.3 Buenos Aires July/Sept. 2008
NOTA PALEONTOLÓGICA
La traza Dactyloidites ottoi (Geinitz) en la Formación La Manga, Oxfordiano de Mendoza
Darío G. Lazo1, Ricardo M. Palma1 y Ricardo D. Piethé1
1Departamento de Ciencias Geológicas, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Pabellón 2, Ciudad Universitaria, 1428 Buenos Aires, Argentina - CONICET; <dlazo@gl.fcen.uba.ar>
Introducción
La Formación La Manga ha sido recientemente objeto de estudios estratigráficos y sedimentológicos de detalle en las cercanías de Bardas Blancas, Mendoza. Un trabajo basado en cinco secciones completas reveló la existencia de seis grandes asociaciones de facies sedimentarias (A-F en Palma et al. 2007). Dichas facies corresponden a ambientes de rampa externa (A), rampa media (B), rampa interna-barrera oolítica (C), arrecifes en parche de margen de rampa interna (D), lagoon (E) y ambiente de karst (F).
En particular, los depósitos de rampa media (Facies B) están compuestos por packstones-grainstones, wackestones y subordinadas calciruditas con una tendencia vertical estrato creciente. Estos depósitos son sumamente interesantes debido a que presentan un conjunto de trazas fósiles de organismos excavadores en un contexto carbonático. Dentro de esta asociación de trazas fósiles se registraron abundantes especímenes asignados a Dactyloidites ottoi (Geinitz, 1849). Esta es una traza fósil relativamente poco frecuente en el Mesozoico y Cenozoico que se registra típicamente en facies clásticas (véase Aguirrezabala y Gibert, 2004). Por lo tanto su presencia en facies carbonáticas de la Formación La Manga, con un excelente estado de preservación, amerita un análisis particular. Sobre la base de lo expuesto, el objetivo de la presente nota es la documentación e interpretación de los ejemplares asignados a D. ottoi.
Área de estudio y metodología
El área de estudio se encuentra localizada en las inmediaciones de Bardas Blancas, en el departamento Malargüe, Mendoza (figura 1.1). En este sector aparecen una serie de pliegues de eje norte-sur que contienen una sucesión sedimentaria jurásica sobrepuesta a volcanitas permotriásicas. El presente trabajo está basado en descripciones de campo de cinco secciones completas de la Formación La Manga de alrededor de 60 m de espesor y en estudios de microfacies basados en Palma et al. (2007). La edad de estos depósitos es asignada al Oxfordiano sobre la base de la fauna de amonites (Riccardi, 1984). Las trazas fósiles analizadas fueron observadas principalmente en el campo (figura 1.2). Dos de los ejemplares analizados de Dactyloidites ottoi y uno de Gyrochorte isp. pertenecen a la colección de Paleontología de la Universidad de Buenos Aires y están catalogados bajo los códigos CPBA 20123.1, 20123.2 y 20124.
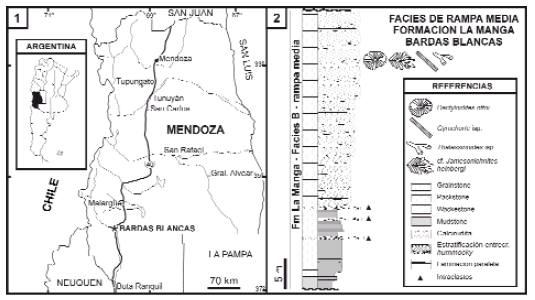
Figura 1. 1, Mapa de localización de la sección analizada; 2, Perfil litológico y ubicación de las trazas fósiles analizadas, Formación La Manga, Bardas Blancas, Mendoza. Modificado de Palma et al. (2007) / 1, Location map of the studied section; 2, Stratigraphic section and trace fossils of La Manga Formation, Bardas Blancas, Mendoza. Modified from Palma et al. (2007).
Icnología
Dactyloidites ottoi (Geinitz, 1849) Figura 2.1-7
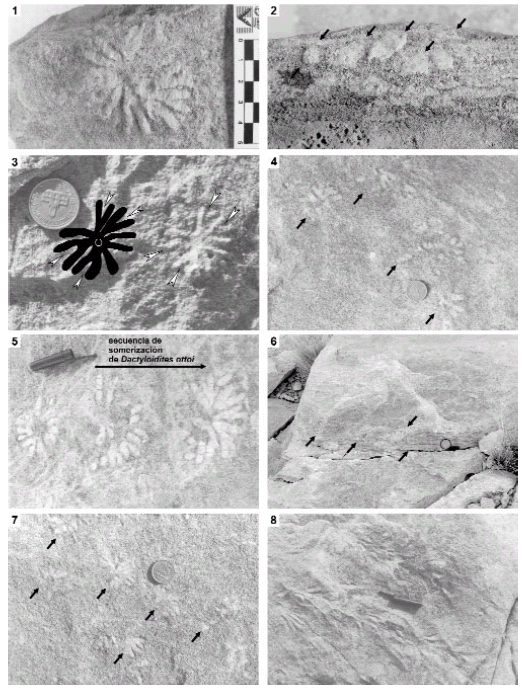
Figura 2. Dactyloidites ottoi y otras trazas fósiles registradas en el tope de packstones-grainstones con HCS, Formación La Manga, Oxfordiano de Mendoza / Dactyloidites ottoi and associated trace fossils on top of packstones-grainstones with HCS, La Manga Formation, Oxfordian, Mendoza. 1-2, D. ottoi en vistas superior y lateral mostrando el spreite protrusivo (flechas), CPBA 20123.1, x 0,75 / 1-2, D. ottoi in upper and lateral views showing protrusive spreite (arrows), x 0,75; 3, Bifurcaciones primarias isótomas (flechas) de D. ottoi, x 0,7; 3, Primary isotomous branching (arrows) in D. ottoi, x 0.7; 4, 7, D. ottoi en forma de roseta y de abanico indicados por flechas, x 0,3 / 4, 7, Rosette and fan-shaped D. ottoi (arrows), x 0.3; 5, secuencia de somerización de D. ottoi, x 0,5 / 5, Shallowing trend in traces of D. ottoi, x 0,5; 6, D. ottoi en vista superior y lateral indicados por flechas, x 0,1; 6, D. ottoi in upper and lateral views (arrows), x 0.1; 8, estructura comparable con Jamesonichnites heinbergi, x 0,4 / 8, upper view of trace fossils comparable to Jamesonichnites heinbergi, x 0.4.
Descripción. Traza de relieve completo con forma de abanico a roseta conformada por elementos radiales y un tubo central vertical. Los elementos radiales poseen un spreite protrusivo y no superan los 40 mm de largo cuando su preservación es completa. Cada elemento radial generalmente presenta una dicotomización isótoma en la base adyacente al tubo vertical (figura 3). Los elementos radiales tienen un largo promedio de 27 mm y un ancho promedio de 5 mm. A partir de dicha dicotomización puede aparecer una segunda ramificación, pero en éste caso es heterótoma ya que se produce hacia un solo lado de la estructura generando una rama secundaria más corta y menos gruesa que la rama primaria resultando un patrón de ramificación asimétrico. De cada elemento radial pueden generarse hasta cuatro terminales como máximo, sin embargo hay casos de tres o dos terminales. El número de terminales no supera los 20 en ninguno de los ejemplares observados. Los elementos radiales pueden presentarse subhorizontales o inclinados con respecto al plano de estratificación. En el caso de baja o nula inclinación de los elementos radiales, la estructura resultante tiende a una forma de roseta casi completa de 360º. En estos casos suele quedar preservado el tubo vertical. Hay ejemplares donde los elementos radiales de una misma traza poseen inclinaciones variables con respecto al plano. Los más inclinados tienen preservadas sólo sus terminales, mientras que los menos inclinados tienden a preservar una mayor proporción de su longitud. En estos casos la traza resultante posee forma de abanico y el tubo vertical se preserva sólo en la porción de la traza que presenta una menor inclinación. Los elementos radiales poseen un relleno mejor seleccionado y de coloración más clara que la roca hospedante. En el plano de estratificación los abanicos y rosetas aparecen aislados o agrupados pero nunca solapados. Las rosetas y abanicos aparecen en diversos niveles.
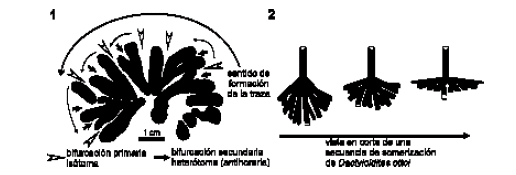
Figura 3. 1, Esquema en planta de Dactyloidites ottoi (Geintiz) mostrando el patrón de ramificación asimétrico hacia la izquierda. Tomado de la Figura 2.5 / 1, Plane view of D. ottoi (Geinitz) showing its asymmetric branching pattern to the left side. Redrawn from Figure 2.5; 2, Esquema en corte de una secuencia de somerización de D. ottoi, fuera de escala / 2, Sketch of a shallowing trend in traces of D. ottoi, not in scale.
Distribución facial. Se registra abundantemente en los topes de packstones-grainstones oopelbiointra-clásticos con estratificación entrecruzada de tipo hummocky (HCS) correspondiendo a un ambiente de shoreface inferior ubicado en el sector de rampa media (véase Facies B en Palma et al., 2007). Los períodos de baja energía entre la depositación de las capas de tormenta permitió la explotación del sustrato por organismos depositívoros del tipo oportunista como Dactyloidites. Una situación similar fue registrada por Aguirrezabala y Gibert (2004), pero en un contexto sedimentario diferente: delta de tipo Gilbert. La presencia de capas con HCS refleja la frecuente acción de tormentas evidenciadas a su vez por una alta desarticulación y fragmentación de bioclastos y abundancia de intraclastos que sugieren un intenso retrabajo y una depositación por encima del tren de base de olas de tormentas (Aigner, 1982; Brenchley, 1985). La presencia de HCS isótropas y anisótropas indica la acción de flujos combinados y profundidades someras (Myrow y Southard, 1991). La frecuente amalgamación de capas con HCS se genera como consecuencia de repetidos eventos de tormentas y es una característica propia de áreas someras (Kreisa, 1981).
Icnocenosis. D. ottoi aparece asociada a otras trazas fósiles, entre las que se reconocen Gyrochorte isp., Thalassinoides isp. y estructuras similares a Jamesoichnites heinbergi Dam, 1990 (figura 4). Gyrochorte isp. se preserva sobre el plano de estratificación como un traza horizontal bilobada elongada y de curso sinuoso. Dentro del sedimento esta traza se presenta como un arco doble convexo que se repite verticalmente formando un spreite protrusivo (Heinberg, 1973). En la Formación La Manga se han observado especímenes de hasta 15 cm de largo. Gyrochorte es generalmente interpretada como una estructura de alimentación de organismos depositívoros producida por especies oportunistas que colonizan el sustrato luego de un evento de depositación de alta energía (Gibert y Benner, 2002). Thalassinoides isp. corresponde a sistemas de galerías tridimensionales excavadas dentro del sedimento. Los tubos son esencialmente cilíndricos, poseen paredes lisas y bifurcaciones en forma de T o Y (Howard y Frey, 1984). Estas galerías son interpretadas como trazas de habitación y alimentación de crustáceos decápodos. Jamesonichnites heinbergi es una traza fósil descripta para el Jurásico Temprano de Groenlandia cuyo material tipo fue refigurado y discutido por Bromley et al. (2003, p. 181, fig. 17). Esta traza se caracteriza por un tubo axial horizontal y tubos secundarios que divergen desde el axial en forma alternada hacia la derecha e izquierda. La traza comparable con J. heinbergi registrada en la Formación La Manga es similar en cuanto al patrón general de ramificación, sin embargo los tubos no presentan paredes (figura 2.8). El tubo axial y los extremos proximales de las ramificaciones se encuentran sobre el plano de estratificación, mientras que los extremos distales son verticales. Esta traza fue originalmente identificada como Macaronichnus isp. (véase Palma et al., 2007). Según Bromley et al. (2003) este tipo de estructuras resultan de las actividades de locomoción y alimentación de bivalvos depositívoros tellinoideos.

Figura 4. Icnocenosis de Dactyloidites ottoi en el tope de packstones-grainstones con HCS interpretados como depósitos de shoreface inferior / Dactyloidites ottoi ichnocoenosis occuring on top of packstones-grainstones with HCS deposited in a lower shoreface.
La icnocenosis de D. ottoi está indicando una consistencia blanda del sustrato carbonático, oxigenación normal, períodos de estabilidad energética y una amplia disponibilidad de materia orgánica. Sin embargo, no hay evidencias claras de escalonamiento dado que no se han encontrado entrecruzamientos. Esto probablemente indica que el tiempo disponible para la colonización del fondo fue breve.
Discusión. Fürsich y Bromley (1985) han planteado un modelo de construcción endogénico para D. ottoi que aún sigue vigente. Este modelo plantea que los elementos radiales surgen a partir del tubo vertical, y presentan spreite protrusivo generado por el movimiento hacia abajo del tubo causativo. Cuando el elemento radial es abandonado, el organismo productor retrorellena el tubo causativo, generando una estructura meniscada. Este modelo se basa en que algunos poliquetos actuales, como Arenicola, podrían cambiar de posición la porción cefálica del tubo manteniendo constante la porción caudal del mismo (Bromley, 1990, p. 44). De esta manera, la rotación alrededor de la porción vertical del tubo produciría un patrón radial o en roseta semejante al de D. ottoi. Sin embargo, el desplazamiento horizontal del organismo constructor que genera rosetas contiguas no solapadas no ha sido explicado satisfactoriamente (véase fig. 3, Fürsich y Bromley, 1985).
Teniendo en cuenta los ejemplares analizados en este trabajo, el modelo de construcción parece ser algo más complejo que lo planteado por Fürsich y Bromley (1985). Es claro que D. ottoi presenta una amplia variedad en cuanto al número de sus elementos radiales y, por lo tanto, no necesariamente presenta forma de roseta dado que la inclinación de los elementos radiales es variable y no siempre es horizontal. En muchos casos éstos son construidos oblicuamente y al ser cortados a diferentes alturas generan distintas figuras sobre el plano de estratificación (figuras 2.4, 2.7). Además, el patrón de ramificación parece ser variable. En el caso de los ejemplares de la Formación La Manga, las ramificaciones aparecen sólo en un sentido, por ejemplo de derecha a izquierda en sentido antihorario (figura 3). En cambio, según lo muestran Fürsich y Bromley (1985, fig. 3), el patrón de ramificación puede tener simetría bilateral. Esta diferencia en la simetría podría reflejar un patrón corporal diferente o simplemente una diferente estrategia de incursión dentro del sustrato. Por otro lado, el material estudiado presenta similitudes morfológicas con los ejemplares descriptos por Aguirrezabala y Gibert (2004) del Aptiano-Albiano de España. Ambos materiales presentan radios de menos de 40 mm de longitud y menos de 20 ramas terminales. Además, los ejemplares de La Manga poseen un menor número de ramas terminales y una menor longitud que los ejemplares descriptos por López-Martínez y Ardévol (1999) del Cretácico Superior de España, Gibert et al. (1995) del Mioceno de España y Pickerill et al. (1993) del Neógeno de Jamaica. Es interesante destacar que, desde el Cretácico Tardío hasta el Neógeno, D. ottoi presenta hasta tres dicotomizaciones heterótomas y por lo tanto los especimenes poseen un mayor número de ramas terminales hasta un máximo de 32 (ver Pickerril et al., 1993, López-Martínez y Ardévol, 1999). Asimismo, cada elemento radial puede tener hasta 70 mm de longitud y por lo tanto la roseta puede alcanzar 14 cm de diámetro, lo cual es considerablemente mayor que las registradas en la Formación La Manga. Sin bien la relación entre el tamaño/número de ramas terminales y la escala temporal es interesante, es importante también considerar los parámetros paleoambientales ya que los especímenes desarrollados en ambientes con restricción en la salinidad presentan en general tamaños menores (véase discusión en Aguirrezabala y Gibert, 2004).
Finalmente, es destacable que los especímenes comparables con J. heinbergi y D. ottoi se registran en los mismos planos de estratificación. Esto, sumado a que ambas trazas poseen dimensiones y rellenos similares, sugiere que ambos tipos de trazas estarían hechas por los mismos organismos reflejando comportamientos diferentes. En el primer caso se refleja un desplazamiento lateral más pronunciado formando una traza espigada, mientras que en el segundo se refleja una búsqueda de alimento más estacionaria tendiendo a una forma en roseta o abanico. Siguiendo este razonamiento se podría pensar que ambas estructuras, tanto D. ottoi como J. heinbergi, fueron realizadas por poliquetos o bivalvos depositívoros sifonados como ya fuera propuesto por Bromley et al. (2003).
Agradecimientos
Los autores desean agradecer a L.A. Buatois, M.G. Mángano, P.J. Pazos, y N. Carmona por su colaboración en la identificación e interpretación de las trazas y por la bibliografía sugerida. N. Carmona y J. de Gibert revisaron y mejoraron sustancialmente el manuscrito. En los trabajos de campo se contó con la invalorable ayuda de D. Kietzmann. La financiación provino de los subsidios UBACyT X-133 y CONICET PIP 5152 otorgados a RMP.
Bibliografía
1. Aigner, T. 1982. Calcareous tempestites: Storm-dominated stratification in Upper Muschelkalk limestones (Middle Trias, SW Germany). En: G. Einsele y A. Seilacher (eds.), Cyclic and Event Stratification, Springer Verlag, Berlin, pp. 180-198. [ Links ]
2. Aguirrezabala, L.M. y Gibert, J.M. de. 2004. Paleodepth and paleoenvironment of Dactyloidites ottoi (Geinitz, 1849) from Lower Cretaceous deltaic deposits (Basque-Cantabrian Basin, west Pyrenees). Palaios 19: 276-291. [ Links ]
3. Brenchley, P.J. 1985. Storm influenced sandstone beds. Modern Geology 9: 369-396. [ Links ]
4. Bromley, R.G. 1990. Trace fossils. Biology and taphnomy. Special Topics in Palaeontology. Unwin Hyman. Londres, 280 pp. [ Links ]
5. Bromley, R.G., Uchman, A., Gregory, M.R. y Martin, A.J. 2003. Hillichnus lobosensis igen. et isp. nov., a complex trace fossil produced by tellinacean bivalves, Paleocene, Monterey, California, USA. Palaeogeography, Palaeoclimatology, Palaeoecology 192: 157-186. [ Links ]
6. Dam, G. 1990. Systematics of trace fossils from the marine Lower Jurassic Neill Klinter Formation, central East Greenland. Bulletin Geological Society Denmark 38: 119-144. [ Links ]
7. Fürsich, F.T. y Bromley R.G. 1985. Behavioural interpretation of a rosetted spreite trace fossil: Dactyloidites ottoi (Geinitz). Lethaia 18: 199-207. [ Links ]
8. Geinitz, H.B. 1849-1850. Das Quadersandsteingebirge oder Kreidegebirge in Deutschland. Craz und Gerlach. Freiberg, 292 pp. [ Links ]
9. Gibert, J.M. de y Benner, J.S. 2002. The trace fossil Gyrochorte: ethology and paleoecology. Revista Española de Paleontología 17: 1-12. [ Links ]
10. Gibert, J.M. de, Martinell, J. y Doménech, R. 1995. The rossetted feeding trace fósil Dactyloidites ottoi (Geinitz) from the Miocene of Catalonia. Geobios 28: 769-776. [ Links ]
11. Heinberg, C. 1973. The internal structure of the trace fossils Gyrochorte and Curvolithus. Lethaia 6: 227-238. [ Links ]
12. Howard, J.D. y Frey, R.W. 1984. Characteristic trace fossils in nearshore to offshore sequences, Upper Cretaceous of east-central Utah. Canadian Journal of Earth Sciences 21: 200-219. [ Links ]
13. Kreisa, R.D. 1981. Storm-generated sedimentary structures in subtidal marine facies with examples from the middle and upper Ordovician of southwestern Virginia. Journal of Sedimentary Petrology 51: 832-848 [ Links ]
14. López-Martínez, N. y Ardévol L. 1999. Hallazgo de Dactyloidites ottoi (Geinitz, 1849) en la arenisca de Arén (Cretácico Superior Surpirenaico). Revista Española de Paleontología, Número Extraordinario Homenaje al Profesor J. Truyols, pp. 129-133. [ Links ]
15. Myrow, P.M. y Southard, J.B. 1991. Combined-flow model for vertical stratification sequences in shallow marine storm-domina-ted beds. Journal of Sedimentary Research 61: 202-210. [ Links ]
16. Palma, R.M., López Gómez, J. y Piethé, R.D. 2007. Oxfordian ramp system (La Manga Formation) in the Bardas Blancas area (Mendoza Province) Neuquén Basin, Argentina: Facies and depositional sequences. Sedimentary Geology 195: 113-134. [ Links ]
17. Pickerril, R.K., Donovan S.K. y Dixon, H.L. 1993. The trace fossil Dactyloidites ottoi (Geinitz, 1849) from the Neogene August Town Formation of south-central Jamaica. Journal of Paleontology 67: 1070-1074. [ Links ]
18. Riccardi, A.C. 1984. Asociación de amonites del Jurásico y Cretácico de la Argentina. 9º Congreso Geológico Argentino (Bariloche), Actas 4: 559-595. [ Links ]
Recibido: 2 de agosto de 2007.
Aceptado: 10 de abril de 2008.

