










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.45 n.4 Buenos Aires dic. 2008
ARTICULOS ORIGINALES
Primera evidencia de dientes aislados atribuidos a Spinosauridae (Theropoda) en el Aptiano inferior (Cretácico Inferior) de Europa: Formación Arcillas de Morella (España)
José I. Canudo1, José M. Gasulla2, D. Gómez-Fernández1, Francisco Ortega3, José L. Sanz2 y Pilar Yagüe2,3
1Grupo Aragosaurus http://www.aragosaurus.com. Paleontología. Facultad de Ciencias, Pedro Cerbuna 12, Universidad de Zaragoza, 50009 Zaragoza, España. jicanudo@unizar.es
2Unidad de Paleontología, Departamento de Biología, Universidad Autónoma de Madrid, Campus de Cantoblanco, 28049 Madrid, España.
3Universidad Nacional de Educación a Distancia, Senda del Rey 9, 28040 Madrid, España.
Resumen. Se estudian 20 dientes de dinosaurios terópodos espinosáuridos del Aptiano inferior (Cretácico Inferior) de Morella (Castellón, España). Los dientes exhiben un mosaico de caracteres derivados para terópodos como son una ornamentación en crestas longitudinales junto con dentículos muy pequeños y de tamaño uniforme que los relaciona con los espinosáuridos barinonícinos como Baryonyx. Sin embargo presentan otros caracteres derivados ausentes en los espinosáuridos del Cretácico Inferior de Europa. Además los dientes comparten algunos caracteres con taxones gondwánicos como Suchomimus e Irritator.
Abstract. First evidence of isolated teeth referred to Spinosauridae (Theropoda) from the lower Aptian (Lower Cretaceous from Europe: Arcillas de Morella Formation (Spain). In this work we study twenty teeth of spinosaurid theropod dinosaurs from the Early Aptian (Early Cretaceous) of Morella (Castellón, Spain). The teeth exhibit a mosaic of derived theropod characters such as ornamentation in longitudinal crests together with very small, uniformly sized denticles, which resemble baryonychine spinosaurids such as Baryonyx. However, they also display other derived characters not found in European Lower Cretaceous spinosaurids. Furthermore, the teeth share certain characters with Gondwanan taxa such as Suchomimus and Irritator.
Palabras clave. Theropoda; Spinosauridae; Aptiano inferior; Morella; Península Ibérica.
Key words. Theropoda; Spinosauridae; Lower Aptian; Morella; Iberian Peninsula.
Introducción
Los espinosáuridos son un grupo de terópodos tetanuros basales que se caracterizan por tener un cráneo alargado y dientes subcónicos, con crestas longitudinales más o menos desarrolladas y dentículos diminutos que permiten diferenciarlos del resto de los terópodos (Buffetaut, 1992; Charig y Milner, 1997; Sereno et al., 1998). Los caracteres derivados que presentan los dientes de los espinosáuridos permiten cierto grado de confianza para identificarlos como tales, aunque no se conozcan otros restos craneales o postcraneales asociados.
Los espinosáuridos están presentes en el Cretácico Inferior y en la base del Cretácico Superior de África y Sudamérica (Sereno et al., 1998; Kellner y Mader, 1997), siendo recientemente citados en Asia (Buffetaut y Suteethorn, 1999; Hasegawa et al., 2003), aunque hay algunas dudas sobre este material, recientemente se insiste sobre la presencia de espinosáuridos en Asia (Milner et al., 2007). En Europa son un componente habitual en sedimentos del Hauteriviano y Barremiano (Martill y Hutt, 1996; Charig y Milner, 1997; Torcida et al., 2003; Ruiz-Omeñaca et al., 2005), sin embargo no habían sido citados en sedimentos más modernos. El objetivo de este trabajo es la descripción por primera vez de dientes aislados atribuidos a espinosáuridos del Aptiano de Europa (Península Ibérica) y compararlos con los taxones conocidos en Gondwana y Laurasia.
Contexto geológico y geográfico
Los restos estudiados en este trabajo provienen de varios yacimientos encontrados en la cantera "Mas de la Parreta" (CMP), situada en el término municipal de Morella (Castellón, España), al noroeste de la Península Ibérica (figura 1). En esta cantera se extraen arcillas de la Formación Morella para la producción de cerámica.
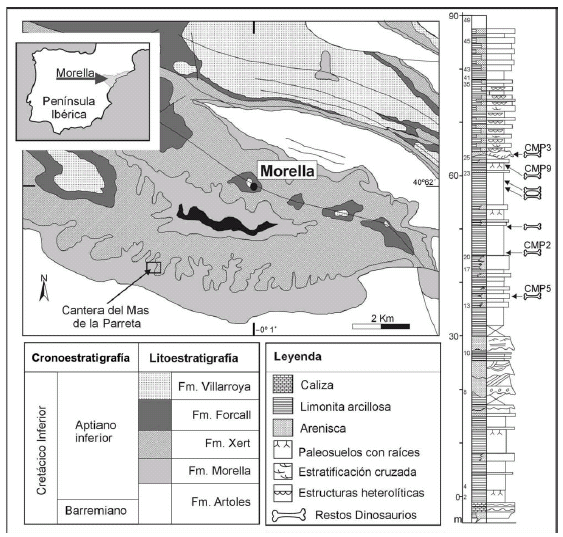
Figura 1. Situación geográfica de la Cantera del Mas de la Parreta (CMP) (Morella, España). Columna estratigráfica de la explotación de arcillas de dicha cantera con la posición de los niveles fosilíferos estudiados en este trabajo / geographical location of the Mas de la Parreta Quarry (CMP) (Morella, Spain). Stratigraphic column showing the exploitation of clays from this quarry with the position of the fossiliferous levels studied in this paper.
Los yacimientos de La Parreta se encuentran en la parte noroeste de la cuenca del Maestrazgo (Salas et al., 2001). El principal componente sedimentario de esta cuenca son carbonatos marinos depositados en aguas poco profundas. También hay materiales siliciclásticos en facies transicionales y continentales, como la Formación Morella (Gámez et al., 2003). Esta formación representa un episodio regresivo en el Aptiano inferior intercalado entre los depósitos carbonatados marinos de la Formación Artoles (Barremiano superior) en la base y la Formación Xert (Aptiano inferior) en el techo (figura 1).
El máximo desarrollo de la Formación Morella se encuentra al norte de la Cuenca del Maestrazgo (subcuenca de Morella), donde llega a alcanzar un espesor de 100 metros y es donde se encuentra la Parreta. Esta formación se depositó en llanuras fangosas y fluviales con alguna influencia marina y mareal, sobre todo hacia el techo de la formación (figura 1). De la base al techo se suceden varias asociaciones de facies que se interpretan como llanuras aluviales intercaladas con otras de relleno de canal, pertenecientes a un sistema fluvial efímero con momentos de influencia mareal (figura 1). La llanura aluvial deja paso a un canal fluvial con abundantes estructuras mareales, finalizando con una asociación típica de llanura mareal siliciclástica (Salas et al., 2003; Gámez et al., 2003). Además, la presencia de un rico y diverso registro de tetrápodos continentales y marinos a lo largo de toda la formación, que incluye dinosaurios, pterosaurios, cocodrilos, escamosos, plesiosaurios, quelonios y "peces" óseos y cartilaginosos (Santafé et al., 1982; Yagüe et al., 2003; Salas et al., 2003), refuerza la interpretación de medios continentales y transicionales para esta unidad geológica.
La edad de la Formación Morella está bien establecida a partir de correlación a nivel de cuenca (Salas et al., 2001). Esta datación se ha realizado integrando los datos de carófitas en los niveles continentales, con los foraminíferos, nanoplancton calcáreo y ammonites en las facies de plataforma marina. Esto permite situar a la Formación Morella en la parte inferior de la secuencia de depósito K.18 datada como Aptiano inferior (Santafé et al., 1982; Salas et al., 2001). La Formación Morella tiene un gran interés histórico al ser el primer lugar donde se describieron restos de dinosaurios en España (Pereda Suberbiola y Ruiz-Omeñaca, 2005; Gasulla, 2005). Entre 1978 y 1980, el Instituto Miquel Crusafont de Sabadell realizó campañas de excavación que permitieron el descubrimiento de abundante y diverso material de dinosaurios ornitópodos, tireóforos, saurópodos y terópodos (Santafé et al., 1982). Posteriormente, se han excavado nuevos yacimientos, pero la mayoría de estos fósiles se encuentran sin publicar. Una de estas localidades son los yacimientos de la cantera del Mas de la Parreta.
La cantera del Mas de la Parreta
Los restos fósiles del Mas de la Parreta provienen del seguimiento y control paleontológico de las labores de extracción de las arcillas de esta cantera. Como consecuencia de este seguimiento, hasta el año 2006, se han recuperado y documentado cientos de restos desarticulados de vertebrados, además de plantas e invertebrados, distribuidos a lo largo de la serie estratigráfica de la Formación Morella que aflora en la cantera (Salas et al., 2003; Gasulla et al., 2003; Yagüe et al., 2003; Gasulla et al., 2006). Cada punto fosilífero se ha tratado como un yacimiento individual nombrándolos con su sigla correspondiente (figura 1). El lugar donde se ubicaban estos yacimientos una vez excavados, ha sido destruido por el avance del frente de explotación. Los dientes descritos en este estudio se han localizado en los yacimientos denominados CMP-2, CMP-3, CMP-5 y CMP-9 (tablas 1 y 2). La posición estratigráfica relativa de estos yacimientos en la Formación Morella en la cantera de la Parreta se expresa en la figura 1. Los números de los yacimientos no expresan una secuencia estratigráfica, ya que los yacimientos se numeran conforme la explotación los va descubriendo.
Tabla 1. Medidas en mm de los dientes aislados de espinosáuridos de la Cantera del Mas de la Parreta (Aptiano inferior, Morella, Castellón, España). FABL: Longitud mesiodistal de la base de la corona en mm. LMW: Anchura lateromedial de la base de la corona en mm. ALT: Altura del diente (conservada). La relación FABL/ALT se ha obtenido a partir de la altura estimada, por lo que es un valor aproximado. de/mm mesial: Número de dentículos por mm en la carena mesial. de/mm distal: Número de dentículos por mm en la carena distal. Cr/lab: Número de crestas labiales. Cr/li: Número de crestas labiales *: medidas estimadas / measurements in mm of the individual spinosaurid teeth from the Mas de la Parreta Quarry (Lower Aptian, Morella, Castellón, Spain). FABL: Mesiodistal length of the base of the crown in mm LMW: Lateromedial width of the base of the crown in mm. ALT: Height of the tooth (preserved). The ratio FABL/ALT has been calculated on the basis of the estimated height and so is an approximate value. de/mm mesial: Number of denticles per mm on the mesial carena. de/mm distal: Number of denticles per mm on the distal carena. Cr/lab: Number of labial ridges. Cr/li: Number of lingual ridges *: estimated measurements.
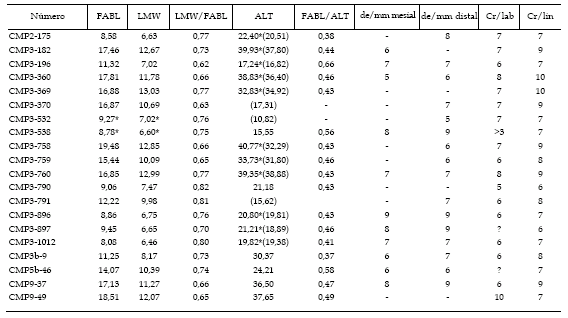
Tabla 2. Conservación y posición anatómica sugerida de los dientes de espinosáuridos de la Cantera del Mas de la Parreta (Aptiano inferior, Morella, Castellón, España). Abrasión: incluimos tanto la que se ha producido por el transporte, como la alteración superficial del esmalte por la posible ingestión del mismo terópodo. Presente: Se observa algo de abrasión en el esmalte o en las crestas longitudinales pero se diferencia la microornamentación. Moderado: Pérdida de la mayor parte de la microornamentación. Fuerte: Pérdida de parte de las crestas longitudinales / preservation and suggested anatomical position of the spinosaurid teeth from the Mas de la Parreta Quarry (Lower Aptian, Morella, Castellón, Spain). Abrasion: we include both what has been produced by transportation and the possible superficial alteration of the enamel produced by possible ingestion on the part of the theropod itself. Present: Some abrasion is observed in the enamel or in the longitudinal crests, but the micro-ornamentation can be distinguished. Moderate: Loss of most of the micro-ornamentation. Strong: Loss of part of the longitudinal crests.
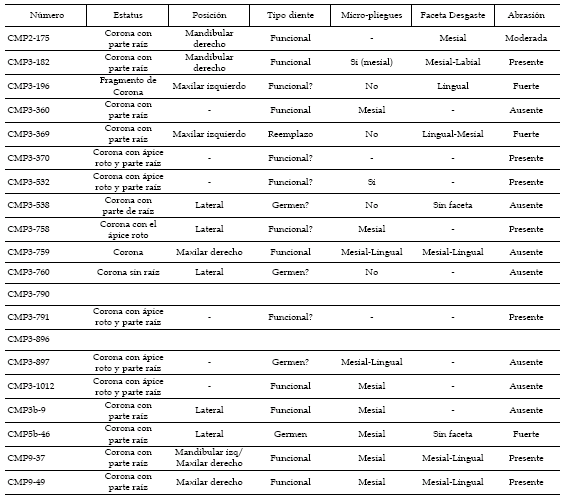
El yacimiento CMP-2 se ha destruido por las labores de explotación minera. Litológicamente eran arcillas rojas, verdosas y abigarradas de la parte media de la Formación Morella (figura 1). Se interpreta como parte de un canal somero. Se encontraron más doscientos restos óseos desarticulados, la mayoría de pequeño tamaño, y dientes de terópodos (Spinosauridae indet., Allosauroidea? indet., Theropoda indet.), cocodrilos (Bernissartia y Goniopholis), Quelonia indet., Squamata indet. y Anura indet. El material más completo era una concentración de restos de Iguanodon.
El yacimiento CMP-9 está excavado por completo y el sustrato eliminado por la actividad minera. Se encontraba en un nivel de limolitas arcillosas rojas y ligeramente abigarradas, de granulometría muy fina, situado en la parte media de la Formación Morella, estratigráficamente muy cercano a CMP-2 (figura 1). Estaba formado en un suelo hidromorfo de baja energía en un contexto de llanura fangosa. Se recuperaron 152 restos fósiles pertenecientes mayoritariamente a la carcasa desarticulada de un solo individuo de Iguanodon. Asociados se encontraron dientes de espinosáuridos, cocodrilos (Goniopholis?) y coprolitos.
El yacimiento CMP-5 está explotado por completo. Se trataba de un nivel de limonitas arcillosas y limonitas arenosas interpretado como un canal fluvial. Se recuperaron 178 fósiles pertenecientes en su mayoría a un ornitópodo similar a Iguanodon. Además se recuperaron escamas de osteíctios, postcraneano de cocodrilos, diente de terópodo y moldes de invertebrados.
El yacimiento CMP-3 se localiza en un nivel de gran extensión horizontal del cual se siguen recuperando fósiles conforme la explotación minera avanza. Litológicamente son conglomerados heterométricos grises y marrones distribuidos en depósitos lenticulares. Además de los clastos, están formados por una gran cantidad de restos fósiles vegetales y de vertebrados tanto continentales como marinos (Gámez et al., 2003). Se interpretan como una superficie transgresiva donde se han desarrollado canales fluviales con influencia mareal (figura 1). En CMP-3 hay una gran abundancia de frondas de Frenelopsis, una conífera que caracterizaba las comunidades de tipo manglar y los pantanos límnicos en el Cretácico Inferior (Gómez et al., 2002). Abundan los troncos del género Brachyoxylon, muchos de ellos presentan bioerosión de bivalvos atribuible a Teredolites (Salas et al., 2003). Este icnogénero es típicamente marino, su abundancia en troncos en el nivel CMP-3 indica que está asociado a un nivel transgresivo. De esta manera, los troncos con Teredolites podrían venir de un mayor aporte de substratos xílicos en ambientes marinos marginales debido a la inundación de llanuras costeras con bosques (Ferrer y Gibert, 2005). En cuanto a la fauna hay restos atribuibles a varios taxones de osteíctios (semionotiformes), condrictios (hibodóntidos), quelonios, plesiosaurios, cocodrilos (Thalattosuchia?, Goniopholis, Bernissartia y Theriosuchus), pterosaurios, dinosaurios saurópodos (Brachiosauridae indet.), terópodos (Dromaeosauridae indet., Spinosauridae indet., Theropoda indet. varios), ornitópodos (Iguanodontidae e Hypsilophodontidae) y del tireóforo Polacanthus sp. (Salas et al., 2003; este trabajo).
Paleontología sistemática
Orden SAURISCHIA Seeley, 1887-1888
Suborden THEROPODA Marsh, 1881
Infraorden TETANURAE Gauthier, 1986
Familia SPINOSAURIDAE Stromer, 1915
SPINOSAURIDAE indet.
Material. Se estudian 20 dientes aislados (listado en tablas 1 y 2) depositados provisionalmente en el Museo de la Valltorta (Castellón, España) con las siglas CMP-2 (1 diente), CMP-3 (16 dientes), CMP-5 (1 diente) y CMP-9 (2 dientes).
Descripción. Los dientes están bien conservados, pero presentan numerosas fracturas debido al proceso de extracción. La muestra incluye coronas funcionales con faceta de desgaste (figuras 2 y 8) y piezas con la superficie del esmalte pulida, bien porque haya sufrido algo de transporte (figura 3) o porque posiblemente esté reabsorbida (figura 2). El grado de abrasión de los dientes se encuentra en la tabla 2. Siete de los dientes carecen de abrasión. En los trece restantes hay una abrasión que varía de suave a moderada, que podría reflejar transporte. Algunos dientes presentan grietas longitudinales (figura 3) posiblemente producidas por alteración subaérea antes del enterramiento.

Figura 2. CMP9-37. 1, labial o lingual / labial or lingual. 2, ápice / apex. 3, base / base. 4, labial o lingual / labial or lingual. 5, distal / distal. 6, mesial / mesial. 7, detalle del borde con el desarrollo en abanico de las microgranulaciones del esmalte en el borde / close-up of the edge with the fan-shaped development of the microgranulations of the enamel on the edge.

Figura 3. CMP3-369. 1, labial / labial. 2, lingual / lingual. 3, distal / distal. 4, mesial / mesial.
El tamaño de los dientes es variable (FABL entre 7 y 20 mm, tabla 1). La diferenciación entre superiores e inferiores se ha realizado por la posición de las facetas de desgaste (tabla 1), mesial-labial (inferiores) y medial-lingual (superiores). En los casos que carecen de faceta de desgaste o ésta se sitúa en una posición imprecisa (CMP9-49) no se ha distinguido entre mandíbulas superiores e inferiores. Ninguno de los ejemplares conserva la raíz completa, por lo que no se ha podido medir en ninguno la longitud total del diente. Hay coronas con pliegues muy suaves (figura 3), perpendiculares al crecimiento del diente (bandas en la terminología de Brusatte et al., 2007), que interpretamos como problemas en el crecimiento llamadas hipoplasias y por tanto sin interés sistemático, aunque si paleoecológico.
Las coronas están curvadas distalmente y presentan una forma general cónica, pero con un ligero aplastamiento labiolingual (figuras 3 y 4), aunque en menor medida que en la mayoría de los dientes de terópodos. La relación entre la longitud y el ancho de la base de las coronas oscila entre 0,62 y 0,82. Las coronas son en general altas (figura 4), con una relación entre la altura y la longitud de la base oscilando entre 0,38 y 0,66. La diferencia de altura posiblemente refleje distintas posiciones a lo largo de la hilera dental (figura 5). Las carenas mesial y distal están bien desarrolladas. El diente no está más aplastado en los bordes para adaptarse a la carena, sino que ésta se presenta como una estructura superpuesta a la sección subcircular. Las carenas son prácticamente rectas. La carena distal se encuentra situada en el plano medial del diente. La parte apical se inclina labialmente respecto a la parte basal de la carena. Carece de la forma de S, característica de otros dientes de terópodos, o está muy ligeramente marcada (figura 6.3). Ambas carenas presentan dentículos extremadamente pequeños rectos, estrechos y subparalelos, prácticamente sin separación entre ellos, lo que da un borde continuo y afilado (figuras 5 y 6). La forma recta del borde cortante posiblemente se deba al desgaste. Los dentículos varían de tamaño (figuras 5.3 y 5.6), pero se mantienen en un número bastante constante de 8 dentículos/mm en ambas carenas. Los dentículos de la carena distal se distribuyen desde la base del diente hasta la corona, cuyo ápice carece de dentículos. El desarrollo longitudinal de la carena mesial es variable. La mayoría de ejemplares posee una carena que no llega hasta la base de la corona (figuras 3.3 y 6.4), mientras que sí ocurre en unos pocos (figura 4.5).

Figura 4. CMP3b-9. 1, labial o lingual / labial or lingual. 2, apical / apical. 3, labial o lingual / labial or lingual. 4, distal / distal. 5, mesial / mesial.

Figura 5. CMP3-1012. 1, labial o lingual / labial or lingual. 2, distal / distal. 3, detalle de los dentículos de la carena mesial en vista lingual / close-up of the denticles of the mesial carena in lingual view. 4, labial o lingual / labial or lingual. 5, mesial / mesial. 6, detalle de los dentículos de la carena distal / close-up of the denticles of the distal carena.

Figura 6. CMP3-760. 1, labial o lingual / labial or lingual. 2, labial o lingual / labial or lingual. 3, distal / distal. 4, mesial / mesial. 5, detalle de las microganulaciones en las crestas longitudinales / close-up of the microgranulations in the longitudinal crests. 6, detalle de la final de la carena mesial / close-up of the end of the mesial carena. 7, detalle de la carena distal y las microgranulaciones en abanico / close-up of the distal carena and the fan-shaped microgranulations.
El esmalte está distintivamente ornamentado por varias estructuras. Todos los dientes presentan unas conspicuas crestas longitudinales (figura 6) y unos pequeños abultamientos discontinuos, visibles ambos a simple vista. Estos últimos le dan al esmalte un aspecto finamente granuloso en toda la superficie de la corona (figuras 6.5-6.7), a diferencia de lo que sucede en la mayoría de los dientes de terópodos que es liso. Estos abultamientos del esmalte se encuentran de manera uniforme por todo el esmalte y, vistos en microscopio se distribuyen paralelamente a las crestas longitudinales (figura 6.6). Hacia los bordes mesial y distal estos abultamientos se abren en forma de un distintivo abanico hacia las carenas (figura 6.7). Las crestas longitudinales son el carácter que más resalta de los dientes de la Parreta. En todos los ejemplares es mayor el número de crestas en el lado lingual que en el labial, con excepción de CMP9-49, donde hay un anormal número de crestas en el labial (10). El número generalmente varía de 6 a 7 en el lado labial y de 7 a 9 en el lingual (tabla 1), aunque en algún caso se presentan más debido a que se subdividen. Estas crestas presentan un desarrollo variable, soliendo estar menos marcadas las crestas más cercanas a los bordes mesial y distal (figuras 6.1 y 6.2). Suelen ser continuas en todo su desarrollo y sólo en algunos ejemplares son discontinuas (figura 6.2). Las crestas no alcanzan hasta el ápice y tampoco llegan hasta el contacto de la raíz con la corona. En tres ejemplares, que aparentemente representan dientes germinales, el desarrollo de las crestas en el lado labial es escaso (tabla 1), por lo que esta ausencia de las crestas labiales podría ser ontogenética.
Unos micropliegues apretados ("enamel wrinkles" de Brusatte et al., 2007), rectos o ligeramente curvos y bien diferenciados de las crestas longitudinales se presentan en contacto con la carena mesial (figuras 5.3 y 8.5) y en algunos ejemplares también se presentan en la distal (tabla 2). Es un carácter bastante variable, tanto en su desarrollo longitudinal como en la profundidad de dichos pliegues. Hay ejemplares en los que se pueden observar a simple vista (figura 8.5). Sin embargo en otros solamente se insinúan, y sólo son visibles al microscopio (figura 5.3). En cuatro ejemplares no se han podido reconocer (tabla 2), aunque dos de ellos los hemos interpretado como gérmenes (CMP3-538 y CMP3-760), por lo que este carácter podría tener también un desarrollo ontogenético. Los otros dos ejemplares, CMP3-196 y CMP3-369 presentan un importante deterioro del esmalte (figura 3) y posiblemente se han perdido por erosión.

Figura 8. CMP3-759. 1, labial o lingual / labial or lingual. 2, labial o lingual / labial or lingual. 3, distal / distal. 4, mesial / mesial. 5, detalle de los micropliegues apretados del borde mesial / close-up of the tightly packed microfolds of the mesial edge.
Discusión
Los dientes de la Parreta presentan algunos caracteres derivados que permiten precisar su posición sistemática. La ornamentación con crestas longitudinales que presentan estos dientes es convergente con la de algunos taxones de cocodrilos como Goniopholis, bien representados en el Cretácico Inferior de la Península Ibérica (Buscalioni y Sanz, 1984). Se diferencian bien porque los dientes de Goniopholis presentan una corona recta y más cónica, careciendo de carenas con dentículos. Hay también una cierta semejanza con los dientes de plesiosaurios por su ornamentación de estrías longitudinales. De hecho, alguno de los dientes estudiados en este trabajo fueron inicialmente asignados a plesiosaurios (Yagüe et al., 2003). Sin embargo, en plesiosaurios la curvatura hacia el lado lingual de la corona está distintivamente más marcada y las crestas son más numerosas, finas y más irregulares, anastomosándose en muchas ocasiones. Además, la cavidad pulpar amplia, como la que presentan los dientes de la Parreta, es más típica de un arcosaurio que de un sauropterigio (Buffetaut e Ingavat, 1986). Los dientes de la Parreta presentan una sección mucho más redondeada que la presente en terópodos con dientes de similar tamaño como pueden ser alosaurios o abelisaurios. La forma cónica es un carácter derivado en Archosauria que aparece, convergentemente, en cocodrilos y en los terópodos Spinosauridae (Charig y Milner, 1997; Sereno et al., 1998), pudiendo ser una sinapomorfía de estos últimos (Kellner y Campos, 1996).
Los dientes de la Parreta presentan una ornamentación constituida por crestas longitudinales distribuidas del ápice a la base. La mayoría de los terópodos presentan dientes con esmalte liso, sin ornamentación, pero hay algunos ejemplos de dientes aislados con ornamentación en terópodos del Jurásico Superior sin clara posición taxonómica, como por ejemplo Megalosaurus meriani Greppin 1870 de Suiza (Greppin, 1870), "Labrosaurus sulcatus" Marsh 1896 de Norteamérica y Labrosaurus stechowi Janensch 1925 de Tanzania. El taxón "Labrosaurus sulcatus" es nomen nudum, teniendo en cuenta las ilustraciones de los dientes premaxilares de Ceratosaurus figurados por Marsh (1896, véase Madsen y Welles, 2000). Este tipo de ornamentación también está presente en dientes aislados relacionados con Ceratosaurus (Madsen y Welles 2000; Rauhut, 2005). Estrías longitudinales con patrones más irregulares que los encontrados en el material de la Parreta, también se han citado en dientes de terópodos enigmáticos como Paronychodon del Cretácico Superior de Norteamérica (Currie et al., 1990) y formas afines del Cretácico Inferior de la Península Ibérica (Zinke y Rauhut, 1994).
Por otro lado, las formas con ornamentación consistente en crestas longitudinales del Cretácico Inferior son asignadas a Spinosauridae (Kellner y Campos, 1996; Martill y Hutt, 1996; Ruiz-Omeñaca et al., 2005), que está compuesto por dos clados hermanos: los Baryonychinae (los espinosáuridos más cercanos a Baryonyx que a Spinosaurus), y los Spinosaurinae (los espinosáuridos más cercanos a Spinosaurus que a Baryonyx). Los miembros de estos clados presentan dos tipos distintos de dientes. Apomórficamente, Spinosaurus tiene dientes cónicos, más parecidos a los de un cocodrilo y sin ornamentación, ya que Sereno et al. (1998) consideran que la presencia de crestas es la condición primitiva para los espinosáuridos. Los dientes de los espinosaurinos son menos curvados; mientras que los barionícinos están más cercanos a la curvatura que, en vista lateral, presentan mayoritariamente los terópodos. Para algunos autores, los barionícinos tienen una ornamentación compuesta por crestas longitudinales bien desarrolladas. Mientras que los espinosaurinos han perdido esta ornamentación, o la tienen poco desarrollada (Martill y Hutt, 1996). Sin embargo, el espinosaurino Irritator presenta unos dientes con unas crestas bien desarrolladas (Sues et al., 2002: fig. 5). Esto dificulta situar a los dientes de la Parreta en una de las dos subfamilias, teniendo en cuenta el criterio de las crestas longitudinales.
Además de las crestas longitudinales, los dientes de la Parreta presentan pequeñas granulaciones en la superficie del esmalte, lo que los diferencia de la mayoría de los terópodos que presentan un esmalte totalmente liso. Este esmalte con pequeñas granulaciones está presente en el ejemplar holotipo de Baryonyx walkeri Charig y Milner 1986 (Charig y Milner, 1997), en dientes aislados del Weald inglés (Martill y Hutt, 1996), del Cretácico Inferior de la Península Ibérica (Ruiz-Omeñaca et al., 2005), en Suchomimus tenerensis Sereno, Beck, Dutheil, Gado, Larsson, Lyon, Marcot, Rauhut, Sadleir, Sidor, Varricchio, Wilson y Wilson, 1998 y en Irritator (Rauhut, com. pers.). Esto parece indicar que es una sinapomorfía de los espinosáuridos.
Otra de las sinapomorfías presente en los dientes de la Parreta es el pequeño tamaño de los dentículos de las carenas mesial y distal. La mayoría de los dientes de los terópodos presentan bordes aserrados con desarrollo de dentículos, cuyo tamaño se ha utilizado tradicionalmente en la identificación de dientes aislados (Farlow et al., 1991). El número de dentículos en los terópodos de tamaño mediano-grande suele oscilar entre 2 y 3 por mm (10-15 cada 5 mm). Sin embargo, en los barionícinos se observa una miniaturización de los dentículos (Martill y Hutt, 1996). El aumento del número de dentículos se puede interpretar como un paso anterior a su desaparición, produciéndose esta evolución de manera independiente en diferentes grupos de tetanuros. Así, dinosaurios con un elevado número de dentículos se observan en Maniraptoriformes como Saurornitholestes langstoni Sues 1978, que en la carena anterior presenta 7 por mm. De manera independiente, en los espinosaurios barionícinos hay una miniaturización de los dentículos: 7 dentículos por mm en Baryonyx (Charig y Milner, 1997), o 7-8 en Suchomimus (Sereno et al., 1998) o en los dientes de la Parreta. En formas más derivadas se presenta una pérdida de los mismos, como sucede en los espinosaurinos (Martill y Hutt, 1996).
Los dientes de la Parreta presentan variabilidad con respecto al desarrollo de la carena mesial. El valor sistemático de este carácter está en discusión, ya que la longitud de esta carena podría variar a lo largo de la mandíbula y, especialmente, con el estadio de erupción del diente (Rauhut, com. pers.). En otros grupos de Tetanurae basales, la reducción de la carena mesial podría tener valor sistemático como estadio intermedio de este carácter con algunos coelurosaurios que pierden por completo la carena mesial (Canudo et al., 2006). En los dientes de la Parreta hay varios ejemplares que se han considerado como gérmenes (tabla 2) al carecer de facetas de desgaste. Estos dientes presentan la carena mesial hasta la base de la corona (figuras 5 y 7), sin embargo algunos de los dientes funcionales presentan la reducción de la carena (figura 3). Esta observación preliminar podría indicar tal vez que en los espinosáuridos de la Parreta este carácter podría estar relacionado con el estadio de erupción del diente. Hay que tener en cuenta que el material de la Parreta son dientes aislados y provenientes de diferentes niveles estratigráficos (Gámez et al., 2003), y no tenemos la certeza que representen a un solo taxón, como se ha considerado en nuestro trabajo
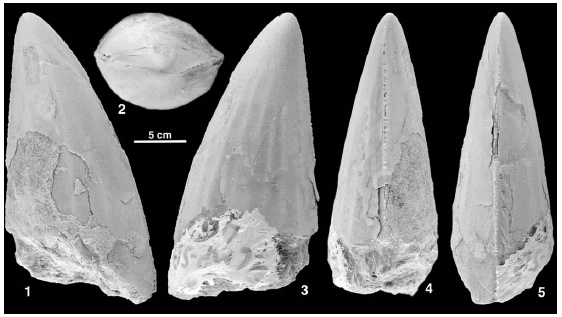
Figura 7. CMP5b-46. 1, labial o lingual / labial or lingual. 2, apical / apical. 3, labial o lingual / labial or lingual. 4, distal /distal. 5, mesial / mesial.
Los dientes de la Parreta presentan una apreciable variabilidad en el tamaño, forma y ornamentación. Esta variabilidad podría considerarse como ontogenética y/o de heterodoncia. Si lo comparamos con material craneal de otros espinosáuridos podemos contestar la cuestión. La reconstrucción de la mandíbula de Baryonyx walkeri presenta los dientes mesiales distintivamente más grandes que el resto (figura 12 en Charig y Milner, 1997). Esto también se observa en los premaxilares (figura 2 en Charig y Milner, 1997). La forma general también varía al presentar los premaxilares una sección más circular (figura 2 en Charig y Milner, 1997), mientras que los maxilares están más aplastados lateromedialmente (figura 3 en Charig y Milner, 1997). Además, la significativa diferencia de tamaño de los dientes de la Parreta indica que están representados diferentes estadios ontogenéticos. Sin embargo, los caracteres más diagnósticos, como número y forma de los dentículos o los patrones de la ornamentación, se mantienen independientemente del tamaño del diente. Por lo que interpretamos que la morfología es constante durante la ontogenia y las variaciones que se detectan entre los ejemplares de la Parreta son debidas a las diferencias en su posición en la serie dental.
Comparación de los dientes de la Parreta con otros espinosáuridos
Los dientes aislados de Formación Arcillas de Morella son difíciles de asignar taxonómicamente, ya que el registro conocido de los terópodos en esta formación es fragmentario. Únicamente se han citado restos postcraneanos y dientes aislados de grandes y medianos tetanuros (Santafé et al., 1982; Gasulla et al., 2006). Sin embargo, la derivada morfología que presentan los dientes de la Parreta nos permite, por lo menos, descartar su inclusión en algunos taxones.
El material de la Parreta se diferencia de los dientes de Spinosaurus por la ausencia de ornamentación en el género africano (Stromer, 1915; Martill y Hutt, 1996; Charig y Milner, 1997; Sereno et al., 1998). Uno de los taxones mejor conocidos es el barionicino Baryonyx walkeri del Weald de la Isla de Wight (Reino Unido, Inglaterra). Es un ejemplar del que se conoce material craneano y postcraneano (Charig y Milner, 1997), y varios dientes aislados asociados con el holotipo. En el dentario no se encontró ningún diente (con excepción de uno muy pequeño, de reemplazo), seis en el premaxilar y uno en el maxilar (Charig y Milner, 1997). Estos autores figuran cuatro dientes con unas distintivas raíces muy largas, que no son posibles de comparar con el material de la Parreta. Las carenas mesial y distal poseen dentículos de pequeño tamaño (7 en Baryonyx, 7-8 en los de la Parreta), un número similar de crestas en el lado lingual (6-8) y el esmalte porta una ligera rugosidad visible a simple vista. Estos caracteres están presentes en el holotipo de Baryonyx del Barremiano de Inglaterra (Charig y Milner, 1997). Sin embargo los dientes de Baryonyx presentan la ornamentación de crestas sólo en el lado lingual, a diferencia de los dientes del Mas de la Parreta que la tienen en los dos lados. También se diferencian por la presencia de los micropliegues del esmalte en los dientes de la Parreta, no citados en Baryonyx. Por tanto, los dientes de la Parreta son de un taxón distinto de Baryonyx walkeri, aunque podría estar relacionado.
El único taxón de espinosaurios descrito a partir de dientes aislados es Siamosaurus suteethorni Buffetaut e Ingavat, 1986 de la Formación Sao Khua (Tailandia). Esta Formación fue datada en el Jurásico Superior, pero posteriormente ha sido correctamente incluida en el Cretácico Inferior (Buffetaut y Suteethorn, 1999). Los dientes asignados a este taxón son frecuentes en dicha formación, pero no se conocen restos postcraneanos. Lo peculiar de estos dientes hizo que Buffetaut e Ingavat (1986) inicialmente los consideraran, con dudas, como de un Spinosauridae, posteriormente confirmado (Hasegawa et al., 2003). Siamosaurus presenta un mayor número (15) de crestas longitudinales (Buffetaut e Ingavat, 1986) que los dientes de la Parreta. Las formas asiáticas de espinosáuridos parecen tener un mayor número de crestas que las formas europeas y africanas, ya que también otros dientes aislados de Japón identificados como Siamosaurus? presentan 12 crestas longitudinales (Hasegawa et al., 2003).
Los dientes de la Parreta comparten algunos caracteres con taxones del final del Cretácico Inferior de Gondwana. Por una parte, tienen un gran parecido con los de Suchomimus tenerensis del Cretácico Inferior de Níger (Sereno et al., 1998), tanto en el número de crestas y número de dentículos como en la ornamentación microgranular en forma de abanico en los bordes (vease figura 2e en Sereno et al., 1998). Este último carácter sólo compartido por Suchomimus y el espinosáurido de la Parreta. Por otra parte, comparte con Irritator challengeri Martill, Cruickshank, Frey, Small y Clarke, 1996 del Aptiano de Brasil la presencia de micropliegues apretados en el esmalte del borde de los dientes. Sues et al. (2002) figuran una corona maxilar del holotipo de Irritator en vista labial donde se observan claramente los micropliegues en contacto con la carena mesial (ver figura 5 de Sues et al., 2002). La presencia de esmalte arrugado ("enamel wrinkles") se ha propuesto recientemente que pueda tratarse como una sinapomorfía de Tetanurae y que la forma, disposición de este esmalte arrugado pueda ser característico de un clado en particular (Brusatte et al., 2007).
Se han descrito dientes aislados de barionícinos en varias localidades del Cretácico Inferior de Europa (Península Ibérica y Reino Unido). Martill y Hutt (1996) describen dientes aislados de un posible barionícino en la Formación Wessex (Wealden Group) del Barremiano inferior de la isla de Wight (Reino Unido). Estudian cinco dientes morfológicamente similares al holotipo de Baryonyx walkeri, con unas carenas compuestas por 5 dentículos por mm. En el entorno de Salas de los Infantes (Burgos, España) aflora la Formación Castrillo de la Reina donde se han descrito dientes aislados de barionícidos del Barremiano superior-Aptiano (Torcida et al., 2003). Estos dientes presentan una morfología similar a los de Mas de la Parreta, con similar número de dentículos por milímetro y de crestas. Sin embargo la ausencia de la ornamentación de pequeños pliegues y de la ornamentación en forma de abanico en estos dientes nos hace pensar que son taxones distintos. Los dientes de la Parreta también presentan caracteres diferentes a los dientes de espinosáuridos recuperados en niveles más antiguos, del Hauteriviano superior y del Barremiano de España (Ruiz-Omeñaca et al., 2005). Éstos tampoco presentan la ornamentación de los espinosáuridos de Mas de la Parreta. Además, presentan una sola carena y poseen más dentículos por milímetro, por lo que son considerados taxones distintos.
Conclusiones
Se describen por primera vez dientes de espinosáuridos en el Aptiano inferior de Europa. Se trata de 20 dientes aislados de la Formación Arcillas de Morella recuperados en la explotación de arcillas de la Parreta. Provienen de tres yacimientos distintos situados en la parte media y superior de esta formación. Teniendo en cuenta el pequeño intervalo temporal que cubre la Formación Morella, estos yacimientos se pueden considerar casi coetáneos. La acumulación de los dientes en los yacimientos de la Parreta tiene un origen complejo, ya que hay una mezcla de dientes funcionales, germinales, con y sin evidencias de transporte (tabla 2), con y sin evidencias de exposición subaérea. Algunos yacimientos como CMP-3 son claramente de acumulación, en el que uno de los elementos son los dientes acumulados de los espinosáuridos. En otros, como CMP-9, la acumulación fosilífera esta formada mayoritariamente por la carcasa de un ornitópodo; en este caso la presencia de los dientes de los espinosáuridos podría ser el resultado de la rotura durante el consumo de dicha presa por parte de los terópodos. La relación de carcasas de dinosaurios fitófagos y dientes aislados de terópodos está bien documentada en la literatura y se relaciona con la alimentación (Buffetaut y Suteethorn, 1989).
Los dientes de los espinosáuridos de los yacimientos de la Parreta presentan una variabilidad morfológica y biométrica que es coherente con que puedan pertenecer al mismo taxón. Morfológicamente estos dientes son diferentes de los descritos en el Hauteriviano-Barremiano del resto de Europa. Además presentan algunos caracteres derivados, como es la presencia de pliegues apretados en los bordes, o el desarrollo de microgranulaciones en abanico presente en taxones gondwánicos (Irritator y Suchomimus, respectivamente). Esto indicaría una conexión entre las faunas ibéricas y gondwánicas apuntada por otros autores en el Cretácico Inferior (Dalla-Vecchia, 2005). Sin embargo, para demostrar esta posible cercana conexión entre el espinosáurido de la Parreta y los espinosáuridos gondwánicas sería necesario la comparación de material más completo que únicamente dientes aislados.
Agradecimientos
Especialmente gracias a la empresa Vega del Moll, S.A que sin su sensibilidad por el patrimonio paleontológico nunca hubieran existido los fósiles estudiados en este trabajo. El Grupo Aragosaurus (http://www.aragosaurus.com/) está subvencionado por el Ministerio de Educación y Ciencia, por los fondos Feder (CGL2004-03393 y CGL2007-62469) y Gobierno de Aragón (Grupos Consolidados 2008). Las fotografías han sido realizadas por la Dra. Zarela Herrera e Isabel Pérez Urresti del Servicio de Fotografía Paleontológica de la Universidad de Zaragoza. Las correcciones de los revisores Oliver Rauhut y Rodolfo Coria han mejorado notablemente el texto original.
Bibliografía
1. Brusatte, S.L., Benson, R.B., Carr, T.D., Williamson, T.E. y Sereno, P. 2007. The systematic utility of theropod enamel wrinkles. Journal of Vertebrate Paleontology 27: 1052-1056. [ Links ]
2. Buscalioni, A.D. y Sanz, J.L. 1984. Los Arcosaurios (Reptilia) del Jurásico Superior - Cretácico Inferior de Galve (Teruel, España). Teruel 71: 9-28. [ Links ]
3. Buffetaut, E. 1992. Remarks on the Cretaceous theropod dinosaurs Spinosaurus and Baryonyx. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 2: 88-96. [ Links ]
4. Buffetaut, E. e Ingavat, R. 1986. Unusual theropod dinosaur teeth from the Upper Jurassic of Phu Wiang, northeastern Thailand. Revue de Paléobiologie 5: 217-220. [ Links ]
5. Buffetaut, E. y Suteethorn, V. 1989. A sauropod skeleton associated with theropod teeth in the Upper Jurassic of Thailand: Remarks on the taphonomic and palaeocological significance of such associations. Palaeogeography, Palaeoclimatology, Palaeoecology 73: 77-83. [ Links ]
6. Buffetaut, E. y Suteethorn, V. 1999. The dinosaur fauna of the Sao Khua Formation of Thailand and the beginning of the Cretaceous radiation of dinosaurs in Asia. Palaeogeography, Palaeoclimatology, Palaeoecology 150: 13-23. [ Links ]
7. Canudo J.I., Ruiz-Omeñaca, J.I., Aurell, M., Barco, J.L. y Cuenca-Bescos, G. 2006. A megatheropod tooth from the late Tithonian-lower Berriasian (Jurassic-Cretaceous transition) of Galve (Aragon, NE Spain). Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 239: 77-99. [ Links ]
8. Charig, A.J. y Milner, A.C. 1986. Baryonyx, a remarkable new theropod dinosaur. Nature 324: 359-361. [ Links ]
9. Charig, A.J. y Milner, A.C. 1997. Baryonyx walkeri, a fish-eating dinosaur from the Wealden of Surrey. Bulletin of the Natural History Museum of London (Geology Series) 53: 11-70. [ Links ]
10. Currie, P.J., Rigby, J.K. y Sloan, R.E. 1990. Theropod teeth from the Judith River Formation of southern Alberta, Canada. En: K. Carpenter y P.J. Currie (eds.), Dinosaurs systematics. Approaches and Perspectives. Cambridge University Press, Cambridge, pp. 282-298. [ Links ]
11. Dalla-Vecchia, F.M. 2005. Between Gondwana and Laurasia: Cretaceous Sauropods in an intraoceanic carbonate platform. En: V. Tidwell y K. Carpenter (eds.), Thunder-Lizards. The Sauropodomorph Dinosaurs. Indiana University Press, Bloomington y Indianopolis, pp. 395-429. [ Links ]
12. Farlow, J.O., Brinkman, D.L., Abler, D.L. y Currie, P.J. 1991. Size, shape, and serration density of theropod dinosaur lateral teeth. Modern Geology 16: 161-198. [ Links ]
13. Ferrer, O. y Gibert, J.M. de. 2005. Presencia de Teredolites en la Formación Arcillas de Morella (Cretácico Inferior, Castellón). Revista Española de Paleontología, Número Especial 10: 39-47. [ Links ]
14. Gámez, D., Paciotti, P., Colombo, F. y Salas, R. 2003. La Formación Arcillas de Morella (Aptiense inferior), Cadena Ibérica oriental (España): caracterización sedimentológica. Geogaceta 34: 191-194. [ Links ]
15. Gasulla, J.M. 2005. Los dinosaurios de Morella (Castellón, España): historia de su investigación. Revista Española de Paleontología, Número Extraordinario 10: 29-38. [ Links ]
16. Gasulla, J.M., Ortega, F., Pereda-Suberbiola, X. y Sanz, J.L. 2003. Elementos de la armadura dérmica del dinosaurio anquilosaurio Polacanthus (Cretácico inferior, Morella, Castellón, España). 19º Jornadas de la Sociedad Española de Paleontología (Morella): 83. [ Links ]
17. Gasulla, J.M., Ortega, F., Escaso, F. y Sanz, J.L. 2006. Diversidad de terópodos del Cretácico Inferior (Fm. Arcillas de Morella, Aptiense) en los yacimientos del Mas de la Parreta (Morella, Castellón). 22º Jornadas de la Sociedad Española de Paleontología (León): 124-125. [ Links ]
18. Gómez, B., Martín-Closas, C., Barale, G., Solé de Porta, N., Thévenard, F. y Guignard, G. 2002. Frenelopsis (Coniferales: Cheirolepidiaceae) and related male organ genera from the Lower Cretaceous of Spain. Palaeontology 45: 997-1036. [ Links ]
19. Greppin, J.B. 1870. Description géologique du Jura bernois et de quelques districts adjacents. Beitrage Geologische Karte Schweiz 8: 1-357. [ Links ]
20. Hasegawa, Y., Buffetaut, E., Manabe, M. y Takakuwa, Y. 2003. A possible spinosaurid tooth from the Sebayashi Formation (Lower Cretaceous), Gunma, Japan. Bulletin of Gunma Museum of Natural History 7: 1-5. [ Links ]
21. Janensch, W. 1925. Die Coelurosaurier und Theropoden der Tendaguru-Schichten Deutsch-Ostafrikas. Palaeontographica, Supplement 7: 1-99. [ Links ]
22. Kellner, A.W.A. y Campos, D.A. 1996. First early Cretaceous theropod dinosaur from Brazil, with comments on Spinosauridae. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 199: 151-166. [ Links ]
23. Kellner, A.W.A. y Mader, B.J. 1997. Archosaur teeth from the Cretaceous of Morocco. Journal of Paleontology 71: 525-527. [ Links ]
24. Madsen, J.H.Jr. y Welles, S.P. 2000. Ceratosaurus (Dinosauria, Theropoda); a revised osteology. Miscellaneous Publication, Utah Geological Survey 2: 1-80. [ Links ]
25. Marsh, O. 1896. The Dinosaurs of North America. United States Geological Survey 16º Annual Report 1894-95: 133-414. [ Links ]
26. Martill, D.M., Cruickshank, A.R.I., Frey, E., Small, P.G. y Clarke, M. 1996. A new crested maniraptoran dinosaur from the Santana Formation (Lower Cretaceous) of Brazil. Journal of the Geological Society 153: 5-8. [ Links ]
27. Martill, D.M. y Hutt, S. 1996. Possible baryonychid dinosaur teeth from the Wessex Formation (Lower Cretaceous, Barremian) of the isle of Wight, England. Proceedings of the Geologists' Association 107: 81-84. [ Links ]
28. Milner, A., Buffetaut, E. y Suteethorn, V. 2007. A tall-spined spinosaurid theropod from Thailand and the biogeography of spinosaurs. 67º Annual Meeting. Society of Vertebrate Paleontology. Journal of Vertebrate Paleontology 27: 118A. [ Links ]
29. Pereda Suberbiola, X. y Ruiz-Omeñaca, J.I. 2005. Los primeros descubrimientos de dinosaurios en España. Revista Española de Paleontología. Número extraordinario 10: 15-28. [ Links ]
30. Rauhut, O. 2005. Post-cranial remains of 'coelurosaurs' (Dinosauria, Theropoda) from the Late Jurassic of Tanzania. Geological Magazine 142: 97-107. [ Links ]
31. Ruiz-Omeñaca, J.I., Canudo, J.I., Cruzado-Caballero, P., Infante, P. y Moreno-Azanza, M. 2005. Baryonychine teeth (Theropoda: Spinosauridae) from the Lower Cretaceous of La Cantalera (Josa, NE Spain). Kaupia. Darmstädter Beiträge zur Naturgeschichte 14: 59-63. [ Links ]
32. Salas, R., Guimerá, J., Mas, R., Martín-Closas, C., Meléndez, A. y Alonso, A. 2001. Evolution of the Mesozoic Central Iberian Rift System and its Cainozoic inversion (Iberian chain). Mémoires du Museum national d'Histoire naturelle 186: 145-185. [ Links ]
33. Salas, R., Colombo, F., Gámez, D., Gasulla, J.M., Martín Closas, C., Moratalla, J., Paciotti, P. y Querol, X. 2003. Guía de la Excursión. 19º Jornadas de la Sociedad Española de Paleontología, Sociedad Española de Paleontología (Morella): 31 pp. [ Links ]
34. Santafé, J.V., Casanovas, M.L., Sanz, J.L. y Calzada, S. 1982. Geología y paleontología (Dinosaurios) de las Capas Rojas de Morella (Castellón, España). Diputación Provincial de Castellón y Diputación de Barcelona, Barcelona,169 pp. [ Links ]
35. Sereno, P.C., Beck, A.L., Dutheil, D.B., Gado, B., Larsson, C.E., Lyon, G.H., Marcot, J.D., Rauhut, O.W., Sadleir, R.W., Sidor, C.A., Varricchio, D.D., Wilson, G.P. y Wilson, J.A. 1998. A long-snouted predatory dinosaur from Africa and the evolution of spinosaurids. Science 282: 1298-1302. [ Links ]
36. Stromer, E. 1915. Wirbeltier-reste der Baharije-Sufe (unterstes Cenoman). 3. Das Original des Theropoden Spinosaurus aegyptiacus nov. gen. nov. spec. Abhandlungen der Mathemat.- Physikalischen Classe der Koeniglich Bayerischen Akademie der Wissenschaften 28: 1-32. [ Links ]
37. Sues, H.-D., 1978. A new small theropod dinosaur from the Judith River Formation (Campanian) of Alberta Canada. Zoological Journal of the Linnean Society 62: 381-400. [ Links ]
38. Sues, H.-D., Frey, E., Martill, D.M. y Scott, D.M. 2002. Irritator challengeri, a spinosaurid (Dinosauria: Theropoda) from the Lower Cretaceous of Brasil. Journal of Vertebrate Paleontology 22: 535-547. [ Links ]
39. Torcida Fernández, F., Izquierdo Montero, L.A., Huerta Hurtado, P., Montero Huerta, D. y Pérez Martínez, G. 2003. Dientes de dinosaurios (Theropoda, Sauropoda), en el Cretácico inferior de Burgos (España). Ciencias de la Tierra 26: 335-346. [ Links ]
40. Yagüe, P., Ortega, F., Noé, L., Gasulla, J.M. y García, M.D. 2003. Reptiles marinos (Plesiosauria) del Aptiense inferior de Morella (Castellón). Ciencias de la Tierra 26: 399-404. [ Links ]
41. Zinke, J. y Rauhut, O.W.M. 1994. Small theropods (Dinosauria, Saurischia) from the Upper Jurassic and Lower Cretaceous of the Iberian Peninsula. Berliner geowissenschftliche Abhandlungen E 13: 163-177. [ Links ]
Recibido: 29 de marzo de 2007.
Aceptado: 15 de julio de 2008.

