










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.45 n.4 Buenos Aires dic. 2008
ARTICULOS ORIGINALES
Maderas gimnospérmicas de la Formación Solca (Pérmico Inferior), provincia de La Rioja, Argentina
Alexandra Crisafulli1 y Rafael Herbst2
1Facultad de Ciencias Exactas y Naturales y Agrimensura. Universidad Nacional del Nordeste. Casilla de Correo 128, 3400 Corrientes. alexandracrisafulli@hotmail.com
2Instituto Superior de Correlación Geológica. Las Piedras 201, 7º B, 4000 Tucumán. rafaherbst@uolsinectis.com.ar
Resumen. Por primera vez se describen leños gimnospérmicos procedentes de las localidades Olta y Parque Guasamayo, en la provincia de La Rioja, donde aflora la Formación Solca, datada como Pérmico Inferior. Se establecen nuevas combinaciones referidas a las especies de Araucarioxylon descriptas y mencionadas en este trabajo. Se determinaron las siguientes especies de Coniferales: Agathoxylon kumarpurensis (Bajpai y Singh) Crisafulli y Herbst nov comb. Agathoxylon ningahense (Maheshwari) Crisafulli y Herbst nov. comb., Podocarpoxylon indicum (Bhardwaj) Bose y Maheshwari, Chapmanoxylon jamuriense Pant y Singh y una nueva especie Chapmanoxylon oltaense. Chapmanoxylon conserva la médula homogénea, el xilema primario y secundario; los restantes géneros sólo preservan el xilema secundario que en todos los taxones presenta interrupciones en el crecimiento y distorsión en sección transversal. Estos leños picnoxílicos, en base a sus características traqueidales se ordenan en tres grupos: con presencia de punteaduras araucarioides, con punteaduras araucarioides-abietinoides y el último con punteaduras araucarioides, mixtas y con tendencia abietinoide. Chapmanoxylon jamuriense está citado en la Formación Raniganj en India donde también se encontró Agathoxylon ningahense y A. kumarpurensis. Además Agathoxylon ningahense se registró en las Formaciones Monte Glossopteris (Antártida) y Melo (Uruguay) y Agathoxylon kumarpurensis y Podocarpoxylon indicum en Springfontein (Sud África), todas formaciones del Pérmico. Sin embargo, la última especie también fue hallada en el Triásico del Grupo El Tranquilo (Argentina) y en el Jurásico de Rajmahal (India).
Abstract. Gymnospermous woods from Solca Formation (Lower Permian), La Rioja Province, Argentina. For the first time gymnospermous woods from the Solca Formation (Lower Permian) from the Olta and Parque Guasamayo localities are described. The conifers Agathoxylon kumarpurensis (Bajpai and Singh) Crisafulli and Herbst nov.comb. and Agathoxylon ningahense (Maheshwari) Crisafulli and Herbst nov. comb. have been transferred from Araucarioxylon; also Podocarpoxylon indicum (Bhardwaj) Bose and Maheshwari, Chapmanoxylon jamuriense Pant and Singh and the new species Chapmanoxylon oltaense Crisafulli and Herbst are described. Chapmanoxylon preserves homogeneous pith, primary and secondary xylem; the other taxa only have secondary xylem all of which present interruptions in the growth and distorsion in transverse sections. These picnoxylic woods based on tracheidal characters are clasified in three groups: with araucarioid pits, with araucarioid-abietinoid pits and finally with araucarioid, mixed and tending to abietinoid pits. Chapmanoxylon jamuriense, Agathoxylon kumarpurensis and Agathoxylon ningahense were found in the Raniganj Formation in India. The latter species was also recorded in the Mount Glossopteris (Antarctica) and Melo (Uruguay) Formations while Agathoxylon kumarpurensis together with Podocarpoxylon indicum were found in Springfontain (South Africa) , all from the Permian. Genus Podocarpoxylon is also present in the Triassic El Tranquilo Group (Patagonia) and Jurassic of Rajmahal (India).
Palabras clave. Anatomía; Maderas; Gimnospermas; Pérmico; Argentina.
Key words. Anatomy; Woods; Gymnosperms; Permian; Argentina.
Introducción
Con este aporte se inicia el estudio anatómico y sistemático de maderas gimnospérmicas procedentes de la Formación Solca que aflora en la provincia de La Rioja (Argentina) en las localidades de Olta y Parque Guasamayo, entre otras (figura 1). En dichos sitios se han podido coleccionar numerosos fragmentos leñosos provenientes del localmente conocido como "Bosque petrificado de Olta". Los materiales se encontraron, algunos rodados y otros incluidos en un conglomerado de areniscas blanquecinas y amarillentas. La Formación Solca ha sido estudiada desde el punto de vista geológico, por diversos autores, entre ellos: Ciano (1989), Andreis et al. (1986), Sterren y Martinez (1996), Net y Limarino (1999), para citar los trabajos más recientes. De acuerdo a la opinión de Andreis et al. (1986), y Net y Limarino (1999), estratigráficamente la Formación Solca se intercala entre las Formaciones Loma Larga (abajo) y La Colina (arriba) y ha sido datada como Pérmico Inferior (figura 2).

Figura 1. Mapa de ubicación de las localidades con maderas fósiles / location map of fossil localities.

Figura 2. Localización estratigráfica de la Formación Solca / stratigraphic position of the Solca Formation.
Hasta el presente, no se conocen estudios previos de la paleoxiloflora hallada en esta Formación. Cabe señalar que en la presente contribución se realiza una serie de nuevas combinaciones, del género Araucarioxylon Kraus a Agathoxylon Hartig. Ello se efectúa por cuanto ya se adoptó este criterio propuesto por Phillipe (1993) y aceptado por Bamford y Phillipe (2001) y Zamuner y Falaschi (2005) y Gnaedinger y Herbst (enviado) por citar sólo algunos, para ir saneando la nomenclatura y la taxonomía, a medida que se van presentando los casos en que estas nuevas combinaciones se justifican, por el análisis de su anatomía. En la Addenda de este trabajo se formaliza esta situación.
Materiales y métodos
Los materiales son fragmentos silicificados, decorticados, de diversos tamaños, principalmente de color marrón, que en la mayoría de los casos han sufrido diversos grados de distorsión. Por ello, de la gran cantidad de materiales coleccionados, sólo un 20 % es apto para un estudio anatómico detallado. Esto ha sucedido principalmente con los leños colectados en el Parque Guasamayo. Para su estudio se ha aplicado la metodología tradicional, por medio de cortes petrográficos, montados y pulidos al punto óptimo de delgadez en los tres planos: corte transversal (CT), corte longitudinal radial (CLRd) y corte longitudinal tangencial (CLTg). Las observaciones y fotografías se hicieron con microscopio Leitz (Ortholux-Orthomax) y lupa estereoscópica Leitz.
La medida de los distintos elementos anatómicos se obtuvo sobre la base de un mínimo de 15 mediciones; en todos los casos se cita primero la media y entre paréntesis los valores mínimos y máximos. La terminología utilizada es de la lista de caracteres microscópicos para la identificación de maderas blandas de la I.A.W.A. (2004), del trabajo de Greguss (1955) y de las medidas estándar establecidas por Chattaway (1932).
Los materiales estudiados se encuentran depositados en la Colección Paleobotánica-Corrientes (CTES-PB) y Preparados Micropaleontológicos-Corrientes (CTES-PMP) de la Facultad de Ciencias Exactas y Naturales y Agrimensura de la Universidad Nacional del Nordeste.
Paleontología sistemática
División TRACHEOPHYTA Stewart y Rothwell 1993
Clase CONIFEROPSIDA Gifford y Foster 1989
Género Chapmanoxylon Pant y Singh, 1987
Especie Tipo. Chapmanoxylon raniganjensis Pant y Singh, 1987.
Discusión. Este género presenta médula homogénea parenquimática, compacta, no tabicada. Estos caracteres permiten compararlo con Bageopitys Dohms (1976), Abietopitys Kräusel (en Kräusel y Range, 1928), Phyllocladopitys Kräusel (en Kräusel y Range (1928), Arauspiropitys Pant y Singh (1987), Chapmanoxylon Pant y Singh (1987) y Gingkgophytoxylon Tidwell y Munzing (1995). La particularidad de presentar este tipo de médula, no es muy común en el conjunto de las maderas gondwánicas pérmicas, ya que la mayoría posee médulas heterogéneas con canales y células secretoras y esclerenquimáticas. De Bageopitys se diferencia principalmente por el tipo de radios leñosos que no son articulados y por la ausencia de punteaduras tangenciales en las paredes traqueidales, rasgos que presentan sus dos especies B. articulata Dohms (1976) y B. herbstii Crisafulli (2001). De Abietopitys se diferencia principalmente por el xilema primario, que es mesarco, el tipo de punteaduras en las paredes radiales no tan abietinoides y los radios leñosos. Arauspiropitys se distingue de este género por presentar xilema secundario con engrosamientos helicoidales y barras de Sanio en las paredes de las traqueidas. Finalmente, Gingkgophytoxylon es diferente por poseer radios uniseriados y biseriados, parénquima axial y el xilema secundario característico de las ginkgophyta presentando variabilidad en el tamaño de las traqueidas e irregular apariencia en sección transversal (Greguss, 1961, 1967). El género más afín para albergar algunos leños de la Formación Solca es Chapmanoxylon que ha sido creado para reunir a aquellos con médula homogénea asignados a Dadoxylon; de acuerdo con el Código Internacional de Nomenclatura Botánica de 1978 este último género se considera inválido.
Chapmanoxylon jamuriense (Maheshwari) Pant y Singh, 1987 Figuras 3.1-4, 4.1-2

Figura 3. 1-4. Chapmanoxylon jamuriense Pant y Singh. 1, CTES-PMP 2632. CT. del xilema secundario mostrando las traqueidas / transverse section of xylem showing tracheids. 2 y 4, CTES-PMP 2634. CLTg. 2, Aspecto de la médula de una traza rameal, rodeada de los radios leñosos / pith of a branch trace surrounded by rays. 4. Radios leñosos uniseriados/ uniseriate rays. 3, CTES-PMP 2633. CLR. Punteaduras uniseriadas y biseriadas en las paredes de las traqueidas/ uniseriate and biseriate pits on the tracheidal walls. 5-6, Chapmanoxylon oltaense nov. sp . CTES-PMP 2629. CT. 5, Células parenquimáticas de la médula/ pith parenchyma cells. 6, Xilema secundario: traqueidas y radios / secondary xylem: tracheids and rays. Escalas / scale bars: 1: 16 μm, 2: 70 μm, 3 y 4: 34 μm, 5 y 6: 60 μm.

Figura 4. Chapmanoxylon jamuriense Pant y Singh. 1, CTES-PMP 2633. CLR. Esquema de las células parenquimáticas de la médula (m), engrosamientos anulares en las traqueidas del xilema primario (x1) y punteaduras variadas en las paredes de las traqueidas del xilema secundario y en los campos de cruzamiento (x2) / sketch of pith parenchyma cells (m), annular thickenings on primary xylem (x1) and varied pits on tracheidal walls of xylem on cross fields(x2) 2, CTES-PMP 2634. CLTg. Xilema Secundario/Secondary xylem.. Tipo y frecuencia de los radios leñosos / type and frecuency of rays. Escalas / scale bars: 1: 80 μm, 2: 60 μm.
1964. Dadoxylon jamuriense Maheshwari, 148-152, lám.1, figs 1-5, lám. 2, fig. 6.
1972. Damudoxylon jamuriense (Maheshwari) Maheshwari, 11.
1987. Chapmanoxylon jamuriense (Maheshwari) Pant y Singh, 4- 21.
Descripción. Fragmento de leño silicificado, decorticado que mide 8 cm de longitud, 5 cm de diámetro mayor y 4,5 cm de diámetro menor; de color marrón rojizo oscuro. Ha preservado la médula, el xilema primario y el xilema secundario. No se ha preservado la corteza. La médula tiene contorno lobado. Mide 1,5 cm de diámetro. Es homogénea, compacta, no tabicada, compuesta de células parenquimáticas de contorno redondeado en corte transversal. Su diámetro medio tangencial y radial respectivamente es de 35 x 57 (30 x 40; 45 x 75) µm. En sección longitudinal radial son de contorno rectangular (figura 4.1). En corte transversal entre la médula y los fascículos del xilema primario se ven espacios producto de la destrucción de las células en el proceso de fosilización. El xilema primario forma proyecciones cuneiformes. El protoxilema es endarco. En las traqueidas del xilema primario se ven engrosamientos espiralados en sección longitudinal radial (figura 4.1). El xilema secundario es picnoxílico, con anillos de crecimiento marcados. En sección transversal tiene traqueidas de contorno rectangular; su diámetro promedio radial y tangencial respectivamente es de 32 x 22 (23 x 15; 38 x 23) µm (figura 3.1) .Un promedio de 4 traqueidas (3-5) separan los radios entre sí (figura 4.1). En las paredes radiales de las traqueidas del xilema secundario se reconocen punteaduras uniseriadas y biseriadas circulares contiguas y espaciadas. Las punteaduras biseriadas se disponen en forma alterna. Los campos de cruzamiento tienen forma cuadrangular, con punteaduras circulares, areoladas con la apertura de la aréola circular. La mayoría de los campos tienen 1 punteadura (2-3) central que mide 7,5 µm de diámetro transversal (figuras 3. 3 y 4.1). En sección longitudinal tangencial el sistema radial es homogéneo. Los radios son homocelulares uniseriados y bajos. La altura promedio es de 7 células (3-9). Las células centrales son de contorno cuadrangular y triangularelipsoidal las de los extremos, vistas en esta sección (figuras 3.2-3 y 4.2).
Material. CTES-PB 12332, CTES-PMP 2632-2633-2634.
Localidad. "Bosque petrificado de Olta" cerca de Olta, La Rioja, Argentina
Horizonte. Formación Solca
Edad. Pérmico Inferior
Discusión. De las especies conocidas hasta ahora de este morfogénero el material riojano puede ser asimilado a Chapmanoxylon jamuriense, citado por primera vez para el Pérmico de la Formación Raniganj (India) (Pant y Singh, 1987) y también para la Formación Tres Islas (Uruguay) (Crisafulli, 2003); la única diferencia con el material indio es que el aquí estudiado presenta radios leñosos más bajos, dado que este carácter, en el holotipo, alcanza a tener hasta 45 células de altura, aunque su promedio es igual al ejemplar de la Formación Solca. Tampoco presenta diferencias significativas con el material de Uruguay, en cuanto a los elementos celulares solo variaciones en el número de células. Esta especie y las siguientes de Chapmanoxylon: Ch. daintreei (Chapman), Ch. kräuselii (Sahni), Ch. parenchymosum (Surange y Maithy), Ch. rangei (Kräusel), Ch. porosum (Kräusel) son nuevas combinaciones de Pant y Singh (1987), antes incluidas en el género Dadoxylon. La presencia de una médula lobada como la de este material es relacionada por Zeiller (en Maithy, 1968) con la disposición de las trazas en el leño. Este es un carácter frecuente en leños paleozoicos del Hemisferio Sur presente también en Taxopitys Kräusel (en Kräusel y Range, 1928); Barakaroxylon (Surange y Maithy) Kulkarni et al (1970), Polysolenoxylon Kräusel y Dolianiti (1958) y Lobatoxylon Kräusel (1956) a diferencia de la mayoría de las médulas del Hemisferio Norte que son discoidales.
Chapmanoxylon oltaense Crisafulli y Herbst nov. sp. Figuras 3. 5-6, 5.1-2, 6.1-3, 7

Figura 5. Chapmanoxylon oltaense nov. sp 1. CTES-PMP 2630 CLR. Esquema de las punteaduras sobre las traqueidas del xilema secundario y detalle de los campos de cruzamiento/ sketch of pits on tracheidal walls of secondary xylem and details of cross-fields. 2. CTES-PMP. 2631. CLTg. Xilema secundario: detalle de los radios y punteaduras tangenciales / secondary xylem: detail of rays and tangential pits. Escalas / Scale bars: 1: 57 μm, 2: 36 μm.

Figura 6. Chapmanoxylon oltaense nov. sp. 1-3. 1, 2, CTES-PMP 2630. CLR. Punteaduras biseriadas alternas y triseriadas en las paredes de las traqueidas del xilema secundario / biseriate, alternate and triseriate pits on the tracheidal walls of secondary xylem. 3, CTES-PMP. 2631. CLTg. Xilema secundario: radios leñosos uniseriados y parcialmente biseriados/secondary xylem: uniseriate, partially biseriate rays. 4-5, Agathoxylon kumarpurensis (Bajpai y Singh ) nov. comb. Xilema secundario /secondary xylem. 4, CT. CTES- PMP: 2635. Traqueidas / tracheids. 5, CTES- PMP: 2637. CLTg. Radios leñosos / rays. 6, Agathoxylon ningahense (Maheshwari) nov. comb. CTES- PMP: 2638. CT. Xilema secundario: traqueidas y "shearing-zones" / secondary xylem: tracheids and shearing-zones. Escalas / scale bars: 1: 50, 2: 40 μm, 3.: 75μm, 4: 60 μm, 5: 100 μm y 6.: 120 μm.
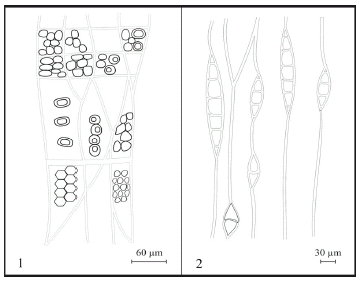
Figura 7. Agathoxylon kumarpurensis (Bajpai y Singh ) nov. comb. Xilema secundario / secondary xylem. 1, CTES- PMP: 2636. CLR. Esquema de las punteaduras en las paredes de las traqueidas y campos de cruzamiento / sketch of pits on the tracheidal walls and cross field. 2, CTES- PMP: 2637. CLTg. Radios leñosos / rays. Escalas / scale bars: 1. 60 µm. 2. 30 µm.
Diagnosis. Pith homogeneous, oval, with circular or irregular parenchymatic cells. Primary xylem endarch with tracheids with scalariform thickenings. Picnoxylic secondary wood, without marked growth rings. Medullary rays in cross section placed at intervals of 4-7 tracheids; rays homogeneous, uniseriate, 3-8 cells in height. Tangential wall of tracheids with, uniseriate, separate, circular pits. Radial wall pitting, 1- 3 seriate, circular, bordered, alternate or opposite, contiguous or separate pits. Cross fields with 1-2, oculipore, separate pits.
Diagnosis. Médula homogénea, ovalada, con células parenquimáticas de contorno circular o irregular. Xilema primario endarco con traqueidas con engrosamientos escalariformes. Xilema secundario picnoxílico sin anillos de crecimiento marcados. Radios leñosos separados por un intervalo de 4-7 traqueidas; radios homogéneos, uniseriados, 3-8 células de altura. Pared tangencial de las traqueidas punteadas, punteaduras uniseriadas, espaciadas, circulares. Pared radial con punteaduras 1-3 seriadas, circulares, areoladas, alternas u opuestas, contiguas o separadas. Campos de cruzamiento con 1-2 punteaduras oculíporas, espaciadas.
Descripción. Fragmento de leño silicificado, aplastado tangencialmente, de color marrón rojizo, decorticado. Mide 18 cm de longitud, 11 cm de diámetro mayor y 5 cm de diámetro menor. Ha preservado la médula, el xilema primario y el xilema secundario con falsos anillos de crecimiento a causa de la distorsión sufrida. La médula es homogénea, compuesta de células parenquimáticas de contorno circular o irregular, en corte transversal miden aproximadamente 90 (87-95) µm de diámetro radial y 75 (68-80) µm de diámetro tangencial (figura 3. 5). El xilema primario forma proyecciones cuneiformes. El protoxilema es endarco con traqueidas de contorno poligonal al igual que las traqueidas del metaxilema. En sección transversal, las traqueidas del xilema secundario son cuadrangulares a ovoidales; su diámetro promedio radial y tangencial respectivamente es de 28 x 24 (35 x 36; 21 x 14) µm. Un promedio de 5 traqueidas (4-7) separan los radios entre sí (figura 3. 6). En corte longitudinal radial el leño secundario es del tipo Araucarioxylon de acuerdo a Lepekhina y Yatsenko-Khimelevsky (1966). Se observan en las paredes traqueidales punteaduras uniseriadas circulares, areoladas, contiguas y espaciadas. También punteaduras biseriadas circulares alternas, en menor proporción opuestas y punteaduras triseriadas subopuestas a opuestas. Los extremos de las traqueidas son romos o en bisel. Los campos de cruzamiento tienen punteaduras areoladas en número de 1 a 2, dispuestas en forma espaciada. La apertura de la aréola es de tipo oculíporo (cupresoide), en aquellas punteaduras que las han preservado. Las punteaduras miden 7,5 um de diámetro (figuras 5. 1, 6. 1-2). El sistema radial es homogéneo. Los radios leñosos son homogéneos, uniseriados y bajos en sección longitudinal tangencial. Tienen una altura promedio de 5 células (4-7). Las células de los radios son cuadrangulares a ovoidales las centrales y triangulares a elipsoidales las de los extremos. Se observan punteaduras uniseriadas circulares areoladas espaciadas en las paredes tangenciales de las traqueidas (figuras 5. 2, 6.3).
Holotipo. CTES- PB12331, CTES- PMP 2629- 2630-2631.
Localidad. Bosque Petrificado de Olta, La Rioja, Argentina Horizonte. Formación Solca
Edad. Pérmico Inferior.
Derivatio nominis. La especie hace referencia a la localidad donde fue hallada.
Discusión. Siguiendo el criterio sistemático de Lepekhina (1972) de determinar los leños Paleozoicos teniendo en cuenta los caracteres de la médula y el xilema secundario cuando ambos están preservados, este material se adjudica al género Chapmanoxylon ya tratado, por las características homogéneas de la médula y la presencia de xilema primario endarco. Si se consideran los rasgos anatómicos del xilema secundario es muy parecido a Agathoxylon lathiense (Agashe, Prasad y Suresh) Crisafulli y Herbst nov. comb. (este trabajo) por la disposición de las punteaduras. No obstante esta especie no tiene punteaduras uniseriadas espaciadas y la altura de sus radios es mayor (1-27). Conforme a lo antes mencionado, es prudente albergarlo dentro de Chapmanoxylon.
No obstante, de las 9 especies reconocidas y señaladas en el cuadro 1, ninguna comparte el conjunto de las características xilemáticas del material y los rasgos específicos de la médula, por lo que es asignado a una nueva especie: Chapmanoxylon oltaense cuyos caracteres son comparados en la figura ya mencionada.
De acuerdo con Alves y Guerra Sommer (2000) maderas con formas elipsoides en sección transversal como Chapmanoxylon oltaense asociadas a la distorsión en el xilema secundario permite inferir compactación inicial con alguna presión vertical. La mala preservación de las punteaduras radiales de las traqueidas (Alves, 1994) es indicio que en el xilema secundario el proceso de degradación no fue acompañado de una evolución rápida de permineralización.
Familia ARAUCARIACEAE Henkel y Hochstetter 1865
Género: Agathoxylon Hartig 1848
Especie tipo. Agathoxylon cordaianum Hartig 1848.
Agathoxylon kurmarpurensis (Bajpai y Singh ) Crisafulli y Herbst nov. comb. Figuras 6.4-5, 7.1-2
Basónimo. 1986. Araucarioxylon kurmarpurensis Bajpai y Singh, 53-56, lám. 1, figs. 1-5, text. fig.1 A-E.
1997. Araucarioxylon kurmarpurensis Bajpai y Singh en Roberts, Bamford y Millsteed, 157-168 lám. 6 A-D.
Diagnosis original. Véase Bajpai y Singh (1986: 53-56, lám. 1, figs. 1-5, text. fig.1 A-E).
Descripción. Fragmento de leño secundario picnoxílico con anillos de crecimiento marcados, silicificado, de color marrón, veteado de marrón claro. Mide aproximadamente 12 cm de longitud, 7 cm de diámetro mayor y 6 cm de diámetro menor. En sección transversal las traqueidas del xilema secundario tienen contorno cuadrangular a rectangular, miden en promedio 60 x 40 µm de diámetro radial y tangencial respectivamente. Un promedio de 6 traqueidas separan los radios entre sí (figura 6.4). En sección longitudinal radial hay punteaduras circulares uniseriadas, contiguas y espaciadas, uniseriadas parcialmente biseriadas, contiguas y triseriadas contiguas. Las punteaduras se disponen en forma opuesta a subopuesta y alterna. Hay algunas punteaduras hexagonales biseriadas en las traqueidas aguzadas. Los campos de cruzamiento tienen punteaduras areoladas, circulares a ovoidales, en número de 4 a 8 (5-7), dispuestas sin orden particular Las punteaduras de estos campos miden 9 µm (figura 7.1).
El sistema radial es homogéneo. Los radios son homocelulares, uniseriados y bajos. En sección longitudinal tangencial, las células radiales son ovoidales preferentemente. El rango de altura es de 1-12 células, siendo el valor promedio 5 células (figuras 6.5, 7.2).
Material. CTES-PB 1233; CTES-PMP 2635, 2636, 2637.
Localidad. Parque Guasamayo. La Rioja, Argentina.
Horizonte. Formación Solca.
Edad. Pérmico Inferior.
Discusión. La disposición de punteaduras que muestra este material riojano, permite asimilarlo a Araucarioxylon kumarpurensis Bajpai y Singh (1986), quienes lo encontraron en la región de Bengala Occidental (India) en las sedimentitas del Pérmico Superior de la Formación Raniganj. Este mismo taxon fue encontrado en la localidad de Springfontein en East London en Sudáfrica donde Roberts et al. (1997) lo registraron en depósitos de finales del Pérmico y también en la Formación Tres Islas en el Pérmico Inferior de Uruguay, Crisafulli (2003).
En cuanto a comparaciones es parecido a Agathoxylon loharense (Agashe y Gowda) Crisafulli y Herbst nov. comb. (este trabajo), pero esta especie tiene punteaduras tangenciales. Se comparó con Agathoxylon ningahense (Maheshwari) Crisafulli y Herbst nov. comb. (éste trabajo), sin embargo Agathoxylon kumarpurensis presenta punteaduras uniseriadas parcialmente biseriadas, mayor número de punteaduras en los campos de cruzamiento y ausencia de punteaduras mixtas que son observables en A. ningahense.
Cuadro 1. Cuadro comparativo de las especies de Chapmanoxylon / comparative table of Chapmanoxylon species.

Agathoxylon ningahense (Maheshwari) Crisafulli y Herbst nov. comb. Figuras 6.6, 8.1-3, 9

Figura 8. Agathoxylon ningahense (Maheshwari) nov. comb. Xilema secundario / secondary xylem 1-3. 1-2, CTES- PMP: 2639. CLR. Punteaduras uniseriadas, biseriadas y mixtas/ uniseriate, biseriate and mixed pits. 3, CTES- PMP: 2640. CLTg. Tipo, distribución y frecuencia de los radios leñosos / type, distribution and frecuency of rays. 4-6, Podocarpoxylon indicum Bose y Maheshwari. 4 y 5. CT. 4, CTESPMP: 2641. Traqueidas del xilema secundario / Tracheids of secondary xylem. 5, CTES-PB: 12335. Aspecto de un fragmento del leño secundario / aspect of a part of secondary wood. 6, CTES-PMP: 2642. CLR. Punteaduras uniseriadas, espaciadas en las paredes de las traqueidas del xilema secundario y en los campos de cruzamiento/ uniseriate, spaciate pits on the tracheidal walls and on the cross-fields of secondary xylem. Escalas / scale bars: 1-3 y 6: 70 μm, 4: 80 μm, 5: 1 cm.

Figura 9. Agathoxylon ningahense (Maheshwari) nov. comb. 1, CTES- PMP: 2639. CLR. Esquema de las punteaduras uniseriadas, biseriadas hexagonales y mixtas en las paredes de las traqueidas y campos de cruzamiento / sketch of uniseriate, biseriate and mixed pits on the tracheidal walls and cross-fields. 2, CTES- PMP: 2640. CLTg. Radios leñosos uniseriados y bajos/ uniseriate and low rays. Escalas/ scale bars: 1: 40 µm, 2: 35 µm.
Basónimo. 1964. Dadoxylon ningahense Maheshwari, 149-152, lámina II, figuras 7-12.
1972. Araucarioxylon cf. ningahense Maheshwari, 21-23, lámina II, figuras 2-6; lámina III, figura 4; Text figura 8.
1982. Australoxylon ningahense (Maheshwari) Prasad, 142.
1997 Araucarioxylon ningahense (Maheshwari) Crisafulli y Lutz, 437-445, lám 2, figs 1-5, fig. 2.
Diagnosis original. Véase Maheshwari (1964:149-152, lámina II, figuras 7-12).
Descripción. Fragmento de leño secundario picnoxílico decorticado silicificado. Mide 16 cm de longitud, 8 cm de diámetro mayor y 6 cm de diámetro menor. Presenta anillos de crecimiento y "shearing zones" sensu Erasmus (1976: 128). Las traqueidas del xilema secundario son rectangulares, de paredes engrosadas, con un lúmen subcircular-ovoidal. Su diámetro promedio radial es de 75 (80-62) µm y el tangencial es de 70 (75-57) µm. Un promedio de 7 traqueidas (2-6) separan los radios entre sí. (figura 6.6). En la pared radial de la traqueidas hay punteaduras areoladas circulares uniseriadas, contiguas y espaciadas;uniseriadas opuestas y alternas y punteaduras mixtas. En menor proporción hay punteaduras triseriadas hexagonales. Algunos campos de cruzamiento son simples y otros con punteaduras de tipo oculíporo con el poro elongado en sentido oblícuo (figuras 8.1-2, 9.1). El sistema radial es homogéneo los radios son homocelulares y bajos (1-8) siendo el valor promedio de 4 células de altura. Las células centrales de los radios son ovales y elípticas las de ambos extremos. Se han visto punteaduras uniseriadas, con el poro dispuesto en forma oblícua en las paredes tangenciales de las traqueidas (figuras 8.3, 9. 2).
Material. CTES-PB 12334; CTES-PMP 2638-2639-2640.
Localidad. Bosque Petrificado de Olta, cerca de Olta, La Rioja, Argentina.
Horizonte. Formación Solca.
Edad. Pérmico Inferior.
Discusión. Los caracteres anatómicos señalados avalan la asignación de estos ejemplares a Agathoxylon ningahense Crisafulli y Herbst nov. comb. Este taxón fue registrado en el Pérmico de la Formación Raniganj (India) por Maheshwari (1964), y en la Formación Monte Glossopteris (Antártica) Maheshwari (1972). Crisafulli y Lutz (1997) encontraron esta especie en la Formación Melo (Uruguay) en los estratos del Pérmico Inferior manteniéndola dentro de este morfogénero. Este criterio difiere al de Prasad (1982) quien pasa algunas especies de Araucarioxylon (entre ellas a A. ningahense), arbitrariamente, al género Australoxylon Marguerier (1973), porque presentan campos de cruzamiento con punteaduras areoladas o simples, considerándolos como carácter definitorio del último género; empero, retiene dentro de Araucarioxylon a A. loharense Agashe y Gowda (1978), A. manieroi (Kräusel y Dolianiti) Maheshwari (1972) y A. gondwanense (Maithy) Maheshwari (1972), que muestran ese tipo de campos. Por consiguiente, en esta contribución se alberga a esta especie dentro de Agathoxylon respetando el criterio de prioridad nomenclatural con respecto a Araucarioxylon y al mismo tiempo se establecen las nuevas combinaciones para cada una de las especies mencionadas en esta discusión. (Véase Addenda, al final del trabajo).
Género Podocarpoxylon Gothan, 1905
Especie tipo. Podocarpoxylon juniperoides Gothan, 1905.
Podocarpoxylon indicum (Bhardwaj) Bose y Maheshwari, 1974 Figuras 8.4-6, 10.1-2, 11.1-2

Figura 10. Podocarpoxylon indicum Bose y Maheshwari. 1, CTES-PMP: 2642. CLR. Detalle de los campos de cruzamiento / detail of crossfields. 2, CTES-PMP: 2643. CLTg. Las flechas muestran los radios leñosos uniseriados y parcialmente biseriados / arrows showing uniseriate and parcially biseriate rays. Escalas / scale bars: 1: 60 μm, 2: 80 μm.
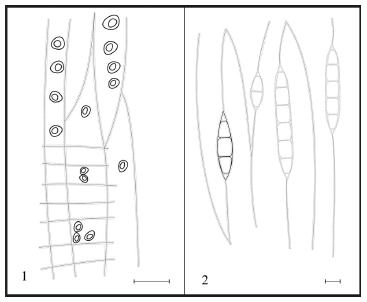
Figura 11. Podocarpoxylon indicum Bose y Maheshwari. 1, CTESPMP: 2642. CLR. Punteaduras uniseriadas en las paredes traqueidales y en los campos de cruzamiento / uniseriate pits on the tracheidal walls and on the cross-fields. 2, CTES-PMP: 2643. CLTg. Radios leñosos uniseriados/ uniseriate rays Escalas / scale bars: 1: 80 µm, 2: 60 µm.
1953. Mesembrioxylon indicum Bhardwaj, 59-70, lám. 1, figs 1-6.
1974. Podocarpoxylon indicum (Bhardwaj) Bose y Maheshwari, 212-223.
1997. Podocarpoxylon indicum (Bhardwaj) Bose y Maheshwari en Roberts, Bamford y Millsteed, 157-168, fig. 9 A-D.
Descripción. Porción de leño silicificado, decorticado que ha sufrido un achatamiento tangencial de sus tejidos. Ha conservado el xilema secundario picnoxílico con shearing zones, y una parte central, en corte transversal, donde parecería hubiera estado la médula y el xilema primario (figura 8.5). En sección transversal, las traqueidas tienen contorno cuadrangular a rectangular. El diámetro medio radial es de 46 (38 -68) µm y el diámetro medio tangencial es de 36 (23 -45) µm. Un promedio de 4 traqueidas (3-15) separan los radios entre sí (figura 8.4). En sección longitudinal radial en las traqueidas hay punteaduras uniseriadas, circulares areoladas, espaciadas (figuras 8.6, 11.1). Las traqueidas son muy angostas, con los extremos aguzados. Presentan placas de resina bicóncavas, tanto en esta sección y como en el corte transversal cercanas a los radios. En los campos de cruzamiento hay punteaduras areoladas en número de 2-3 en hileras verticales (figuras 10.1, 11.1). El sistema radial es homogéneo. Los radios son homocelulares, uniseriados, sólo unos pocos, parcialmente biseriados. En sección longitudinal tangencial las células de los extremos de los radios se ven triangulares y cuadrangulares las centrales. Son bajos, la altura promedio de los mismos es de 7 células (2-10 células) (figuras 10. 2, 11.2).
Material. CTES- PB12335, CTES- PMP 2641-2642-2643.
Localidad. Parque Guasamayo, La Rioja, Argentina.
Horizonte. Formación Solca.
Edad. Pérmico Inferior.
Discusión. Este tipo de punteaduras uniseriadas con la disposición que se ha detallado es característica del género Podocarpoxylon. Desde que fue creado por Gothan en 1905, ha sido objeto de numerosos cuestionamientos sistemáticos, discutiéndose su validéz, ya que aparentemente se lo relacionaría muy estrechamente con las Podocarpaceae actuales. Por otra parte, queda poco claro los criterios relativos a su posición taxonómica frente a morfogéneros afines como Phyllocladoxylon (Gothan, 1905) y Mesembrioxylon (Seward, 1919). Este último aceptado también porque este nombre no implica una afinidad directa del leño fósil con plantas actuales.
Este material caracterizado principalmente por la presencia de traqueidas con punteaduras areoladas, circulares, uniseriadas, espaciadas y campos de cruzamiento de tipo podocarpoide, con pocas punteaduras es asimilable a la especie P. indicum ( Roberts et al., 1997) citada anteriormente en los estratos pérmicos de Springfontein en Sud África y triásicos de la Formación Laguna Colorada en el Grupo El Tranquilo (Santa Cruz, Argentina) (Crisafulli, 2006) y jurásicos de Rajmahall en India, (Bose y Maheshwari ,1974).
Comentarios
Siendo éstos los primeros cinco taxones de maderas fósiles que se han identificado para la Formación Solca, los resultados a los que se ha podido arribar aún son preliminares.
No obstante, es posible resaltar algunos aspectos:
1- Se han reconocido maderas del tipo I, II, de Pant y Singh (1987) por las punteaduras que presentan las traqueidas en sus paredes radiales. Por sus caracteres evolutivos éstas se ajustan más adecuadamente a sedimentos de edad Pérmica que Carbonífera. Además, al igual que en otros yacimientos del Pémico Inferior de La Pampa (Argentina) (Crisafulli et al., 2000) y Uruguay (Crisafulli y Lutz 1997, 2000) que se han estudiado, no se ven punteaduras 4 y 5 seriadas que de acuerdo con Agashe y Prasad (1989) es considerado como un rasgo más primitivo.
2- Si bien las especies: Agathoxylon kumarpurensis, A. ninghaense y A. bengalense poseen punteaduras mixtas en sus paredes, aunque el porcentaje es menor que las de tipo araucarioide, se prefiere apoyar el criterio de Bajpai y Singh (1986) y dejarlas en la familia Araucariaceae antes que Protopinaceae ya que además son especies incluidas en sedimentos Pérmicos y las Protopinaceae, una familia que principalmente agrupa maderas del Mesozoico. No obstante, hay otros leños pérmicos asignados a especies de Taxopitys, Agathoxylon =Araucarioxylon y Baieroxylon que también tienen estas punteaduras, lo que de asignarlas a esta familia obligaría a trasladar a varios de los géneros citados a las Protopinaceae, habiendo sido definidos principalmente por otros caracteres que no son todos compartidos por los miembros de ese grupo a que se hace referencia. Bamford y Philipe (2001) también sostienen este criterio.
3- De acuerdo con Agashe y Prasad (1989), los grupos de plantas fósiles afines a las familias de coníferas como Araucariaceae, Taxaceae y Podocarpaceae, como las especies aquí citadas, indicarían un registro de clima cálido y húmedo durante el período Pérmico en que crecieron en el Gondwana.
4- Hay alto porcentaje de maderas abietíneas (con punteaduras espaciadas) como las observadas en el Pérmico Superior de otras formaciones de las cuenca de Paraná, (Crisafulli y Lutz, 1997 y 2000) y en el Pérmico Inferior de la Formación Carapacha, en la provincia de La Pampa, Argentina (Crisafulli et al., 2000).
5- En este trabajo se registra una nueva localidad pérmica en el Hemisferio Sur con la presencia de leños afines a las Podocarpaceae. Hasta el presente, de este periodo paleozoico, se conocen registros de Podocarpoxylon en Brasil, Paraguay, Australia, Sudáfrica y Nueva Zelanda y recientemente en el Triásico de Argentina (Crisafulli, 2006). Citas del Jurásico, lo muestran con una distribución generalizada en ambos hemisferios por ejemplo en Libia (Philippe et al., 2004) y en Marruecos (Attims y Cremier, 1969) que en ese tiempo pudieran haber estado en Gondwana, aunque hoy son parte del Hemisferio Norte.
6- La edad Pérmico Inferior asignada a la Formación Solca por Net y Limarino (1999 ) se ve apoyada tanto por las características anatómicas de estas especies así como por su presencia en otras Formaciones de la misma edad.
Addenda taxonómica
Familia ARAUCARIACEAE Henkel y Hochstetter 1865
Género Agathoxylon Hartig 1848
Especie tipo. Agathoxylon cordaianum Hartig 1848.
Agathoxylon kurmarpurensis (Bajpai y Singh ) Crisafulli y Herbst nov. comb.
Basónimo. 1986. Araucarioxylon kurmarpurensis Bajpai y Singh Bajpai y Singh, 53-56, lám. 1, figs. 1-5, text. fig.1 A-E.
1997. Araucarioxylon kurmarpurensis Bajpai y Singh en Roberts, Bamford y Millsteed, 157-168 lám. 6 A-D.
Diagnosis. Como en Bajpai y Singh 1986: 55.
Agathoxylon ningahense (Maheshwari) Crisafulli y Herbst nov. comb.
Basónimo. 1964. Dadoxylon ningahense Maheshwari, 149-152, lámina II, figuras 7-12.
1972. Araucarioxylon cf. ningahense Maheshwari, 21-23, lámina II, figuras 2-6; lámina III, figura 4; Text figura 8.
1982. Australoxylon ningahense (Maheshwari) Prasad, 142.
1997 Araucarioxylon ningahense (Maheshwari) Crisafulli y Lutz, 437-445, lám 2, figs 1-5, fig. 2
Diagnosis. Como en Maheshwari 1964: 150
Agathoxylon loharense (Agashe y Gowda) Crisafulli y Herbst nov. comb.
Basónimo. 1978. Araucarioxylon loharense Agashe y Gowda, 269-274.
Diagnosis. Como en Agashe y Gowda, 1978: 270
Agathoxylon lathiense (Agashe, Prasad y Suresh) Crisafulli y Herbst nov. comb.
Basónimo. 1981. Araucarioxylon lathiense Agashe, Prasad, y Suresh, 122-127.
Diagnosis. Como en Agashe, Prasad, y Suresh 1981: 125.
Agathoxylon manieroi (Kräusel y Dolianiti) Crisafulli y Herbst nov comb.
Basónimo. 1958. Dadoxylon manieroi en Kräusel y Dolianiti, 115-137, pls 16-26, 7 tables.
1972. Araucarioxylon manieroi Maheshwari, 1-82.
Diagnosis. Como en Kräusel y Dolianiti 1958: 119.
Agathoxylon gondwanense (Maithy) Crisafulli y Herbst nov comb.
Basónimo. 1964 Dadoxylon gondwanense Maithy, 89-93, 1 pl., 4 figs.
1972. Araucarioxylon gondwanense Maheshwari, 1-82.
Diagnosis. Como en Maithy: 90.
Agathoxylon bengalense (Maheshwari) Crisafulli y Herbst nov.comb.
Basónimo. 1917. Dadoxylon bengalense Holden, 320-322, pl. 20, figs 19-22.
1972. Araucarioxylon bengalense (Holden) Maheshwari, pl.1 figs. 1-5; pl 2, fig. 1; text fig. 7.
1973. Australoxylon bengalense (Holden) Marguerier, 37-58: tab. 1.
Diagnosis. Como en Holden, 1917: 20.
Agradecimientos
Queremos agradecer muy especialmente a A. Lutz, a la Técnica de Laboratorio E. Acevedo y a M. Vince quienes nos acompañaron al lugar de colección. Agradecemos a A. Arcucci por indicarnos y acompañarnos al sitio paleontológico del "bosque de Olta", a O. Revuelta por la digitalización y confección de las ilustraciones. Hacemos extensivo nuestro agradecimiento a los señores árbitros M. Brea y R. Pujana por sus valiosas sugerencias las que contribuyeron a mejorar la calidad de nuestro trabajo.
Bibliografía
1. Agashe, S. y Gowda, P. 1978. Anatomical study of a gymnospermous wood from Lower Gondwana of Maharashtra. Phytomorphology 28: 269-274. [ Links ]
2. Agashe, S., Prasad, K. y Suresh, F. 1981. Two new species, Araucarioxylon surangei and Araucarioxylon lathiense, of petrified woods from lower Gondwana strata. The Palaeobotanist 28-29: 122-127. [ Links ]
3. Agashe, S. y Prasad, K., 1989. Studies in fossil gymnospermous woods, part 7: Six new species of Lower Gondwana (Permian) gymnospermous woods from Chandrapur district, Maharashtra state, India. Palaeontographica 138 B: 71-102. [ Links ]
4. Alves, L. 1994. Integraçao entre a Lignoflora e a Palinología no Afloramento Passo Sao Borja (Fm. Iratí- Bc. Do Parana), (Disertacao de Mestrado, Instituto de Geociencias, Universaidade Federal di Rio Grande do Sul) 174. p [ Links ]
5. Alves,L. y Guerra Somer, M. 2000. Inférencias tafonómicas para un fragmento de lenho da Formacao Iratí, Permiano Superior , da Bacia do Parana, RS Brasil. X Reuniao de Paleobotanicos e Palinologos . Revista Universidade Guarulhos, Geociencias 5 (nº especial ). 44: 21-30. [ Links ]
6. Andreis, R., Leguizamón, R., y Archangelsky. S, 1986. El paleovalle de Malanzán: nuevos criterios para la estratigrafía del Neopaleozoico de la Sierra de Los Llanos, La Rioja, República Argentina. Academia Nacional de Ciencias 57: 3-119. [ Links ]
7. Attims, J. y Cremier, F. 1969. Etude de quelques bois jurassiques du Mésozoique du Maroc. Notes Serv. Geol. Maroc, 210: 19-92. [ Links ]
8. Bajpai, U. y Singh, V. 1986. Araucarioxylon kumarpurensis, a new gymnospermous wood from the Upper Permian of West Bengal. The Palaeobotanist 35: 53-56. [ Links ]
9. Bamford, M. y Phillipe, M. 2001. Jurassic-Early Cretaceous Gondwanan homoxylous woods: a nomenclatural revision of the genera with taxonomic notes. Review of Palaeobotany and Palynology 113: 287-297. [ Links ]
10. Bhardwaj, D. 1953. Jurassic woods from the Rajmahal Hills, Bihar. The Palaeobotanist 2: 59-70. [ Links ]
11. Bose, M. y Maheshwari, H. 1974. Mesozoic conifers. In Surange K., Lakhanpal R. y Bhardwaj, D. (Editors) Birbal Sahni Institute of Palaeobotany. Aspects and Appraisal of Indian Palaeobotany: 212-223. [ Links ]
12. Chattaway, M. 1932. Proposed standart for numerical values used in describing woods. Tropical woods 29: 20-28. [ Links ]
13. Ciano, C. 1989. [Estratigrafía y paleoambientes de las Formaciones neopaleozoicas (Solca y La Colina) de la cuenca intermontana de la Sierra de Los Llanos, Provincia de La Rioja. Universidad de Buenos Aires. Facultad de Ciencias Exactas y Naturales. Departamento de Ciencias Geológicas. Trabajo Final de Licenciatura: 110 pp. Inédito.]. [ Links ]
14. Crisafulli, A. 2001. Leños pérmicos de la Formación Yaguarí (Pérmico Superior), Uruguay. Ameghiniana 38: 61-72. [ Links ]
15. Crisafulli, A. 2003. Maderas Gimnospérmicas de la Formación Tres Islas. (Pérmico Inferior de Uruguay) Resúmenes del Simposio Argentino del Paleozoico Superior: p. 12. [ Links ]
16. Crisafulli, A. 2006. Maderas Gimnospérmicas del Grupo El Tranquilo (Triásico Superior) Santa Cruz Argentina. Resúmenes del Simposio de Paleobotánica y Palinología: p. 35. [ Links ]
17. Crisafulli, A. y Lutz, A. 1997. Leños gimnospérmicos de la Formación Melo (Pérmico Inferior) Uruguay. Parte I. Barakaroxylon jhariense Surange y Maithy, 1962 y Araucarioxylon Kraus, 1870. Ameghiniana 34: 437- 445. [ Links ]
18. Crisafulli, A. y Lutz, A. 2000 . Xiloflora de la Formación Melo (Pérmico Inferior), Uruguay. Ameghiniana 37: 73-80. [ Links ]
19. Crisafulli, A., Lutz, A. y Melchor, R. 2000 . Maderas gimnospérmicas de la Formación Carapacha (Pérmico), provincia de La Pampa, Argentina. Ameghiniana: 37: 180-191. [ Links ]
20. Dohms, M. 1976. Nova madeira de Gimnosperma do Permiano do Rio Grande Do Sul. Revista Brasileira de Geociencias 6: 164-181. [ Links ]
21. Erasmus, T. 1976. On the anatomy of Dadoxylon arberi Seward, with some remarks on thephylogenetical tendencies of its tracheid pits. Paleontologia Africana 19: 127-133. [ Links ]
22. Gifford, E.M. y Foster, A.S. 1989. Morphology and Evolution of Vascular Plants. W.H. Freeman (ed.), 626 pp. [ Links ]
23. Gnaedinger, S. y Herbst, R. (en prensa). Primer registro de maderas fósiles de la Formación Roca Blanca (Jurásico Inferior), provincia de Santa Cruz, Argentina. Ameghiniana 46. [ Links ]
24. Gothan, W.1905. Zur anatomie lebender und fossiler Gymnospermenhölzer, Abahandlungen der Köninglich. Preuss. Geol. Landesantst., N. F. (Berlín) 44: 1- 108. [ Links ]
25. Greguss, P. 1955. Identification of living gymnosperms on the basis of xylotomy. Akadémiai Kiadó, 508 pp. [ Links ]
26. Greguss, P. 1961. Permische fossile Hölzer aus Ungan. Palaeontographica 109: 131-146. [ Links ]
27. Greguss, P. 1967. Fossil gymnosperm woods in Hungary from the Permian to the Pliocene, Akademiai Kiadó, Budapest, 136 pp. [ Links ]
28. Hartig, A. 1848. Beiträge zur Geschichte der Pflanzen und zur Kenntnis der norddeutschen Braunkohlen - Flora. Botanische Zeitung 6: 185-190. [ Links ]
29. Henkel, A. y Hochstetter, W. 1865. Synopsis der Nadelhölzer 17. [ Links ]
30. Holden, R.1917. On the anatomy of two Palaeozoic stems from India. Annals of Botany 31: 315-326. [ Links ]
31. I.A.W.A. Comité, 2004. List of microscopic features for softwood identification. International Association Wood Anatomist Journal 25: 1-70. [ Links ]
32. Kräusel R. 1956. Der "Versteinerte Wald" im Kaokoveld, Südwest- Afrika Senckerbergiana lethaea 37: 411-456. [ Links ]
33. Kräusel, R. y Dolianiti E. 1958. Gymnospermenhölzer aus dem Paläozoikum Brasiliens. Palaeontographica, 104: 115-137. [ Links ]
34. Kräusel, R. y Range, P. 1928. Beitrage zur Kenntnis der Karru- formation Deutsch Sudwest Afrikas, Beitrage geologie Erforschungen 20: 1-54. [ Links ]
35. Kulkarni, S., Maithy, P. y Surange, K. 1970. On Barakaroxylon jhariense. The Palaeobotanist 18: 305-308. [ Links ]
36. Lepekhina, V.1972. Woods of Palaeozoic pycnoxilic gimnosperms with special reference to North Eurasia representatives. Palaeontographica 138: 44-106 [ Links ]
37. Lepekhina, V. y Yatzenko-Khimelevsky, A. 1966. Clasification and nomenclature of woods of Palaeozoic pycnoxylic plants. Taxon 15: 66-70. [ Links ]
38. Maheshwari, H., 1964. Studies in the Glossopteris flora of India. 24. On two new species of fossil wood from the Raniganj stage of Raniganj Coaldfield, Bengal. The Palaeobotanist 13: 148 - 154. [ Links ]
39. Maheshwari, H. 1972. Permian woods from Antarctica and revision of some Lower Gondwana woods taxa. Palaeontographica B 203:1-82. [ Links ]
40. Maithy, P. 1964.Two new species of Dadoxylon from the Lower Gondwana of India, pt 19 in Studies in the Glossopteris flora of India. The Paleobotanist 13: 89-93. [ Links ]
41. Maithy, P. 1968. Some further observations and remarks on Araucarioxylon daintreeii Chapman. The Palaeobotanist 16: 145-147. [ Links ]
42. Marguerier, J. 1973. Paleoxylologie du Gondwana africain: etude et affinites du genre Australoxylon Paleontologie. Africane 16: 37-58 [ Links ]
43. Net, L. y Limarino, C. 1999. Paleogeografía y correlación estratigráfica del Paleozoico Tardío de la Sierra de Los Llanos, Provincia de La Rioja, Argentina. Revista de la Asociación Geológica Argentina 54: 229-239. [ Links ]
44. Pant, D. y Singh, V. 1987. Xylotomy of some woods from Raniganj Formation (Permian), Raniganj Coalfield, India, Palaeontographica B 203: 1-82. [ Links ]
45. Philippe, M. 1993. Nomenclature generique des trachéidoxyles mésozöiques á champs araucarioides. Taxon 42: 74-80. [ Links ]
46. Philippe, M, Bamford, M., Mc Loughlin, S. Alves, L., Falcon-Lang, H., Gnaedinger, S. Ottone, E, Pole, M., Rajanikanth, A. Shoemaker, R. Torres, T. y Zamuner, A. 2004. Biogeographic analysis of Jurassic- Early Cretaceous wood assemblages from Gondwana. Review of Palaeobotany and Palynology 129: 141-173. [ Links ]
47. Prasad, M.1982. An annotated synopsis of Indian Palaeozoic gymnospermous woods. Review of Paleobotany and Palynology 38: 119-156. [ Links ]
48. Roberts, D., Bamford, M y Millsteed, B. 1997. Permo-Triassic macro-plant fossils in the Fort Grey silcrete, East London South Africain. Journal Geology 100: 157-168. [ Links ]
49. Seward, A. 1919. réédité en 1963. Fossil plants, IV, Cambridge University Press Biological, 543pp. [ Links ]
50. Sterren, A. y Martínez, M. 1996. El Paleovalle de Olta (Carbonífero): paleoambiente y paleogeografía. 12º Congreso Geológico Argentino y 3º Congreso de Exploración de Hidrocarburos, Actas 2: 89-103. [ Links ]
51. Steward, W. y Rothwell, G. 1993. Paleobotany and the evolution of plants . Cambridge University Press, 521 pp. [ Links ]
52. Tidwell, W. y Munzing, G. 1995. Gymnospermous woods from the Lower Permian Huaco Formation of South-Central New Mexico. New Mexico Museum of Natural History and Science Bulletin: 91-100. [ Links ]
53. Zamuner, A. y Falaschi, P. 2005. Aghatoxylon matildense n. sp., leño araucariáceo del Bosque petrificado del cerro Madre e Hija, Formación La Matilde (Jurásico medio), provincia de Santa Cruz, Argentina. Ameghiniana 42: 339-346. [ Links ]
54. Zhang, W., Wang, Y., Zheng, S., Yang, X., Li, Y. y Fu, X y Li, N. 2007. Taxonomic investigations on permineralized conifer woods from the Late Paleozoic Angaran deposits of northeastrn Inner Mongolia, China and their paleoclimatic significance. Review of Paleobotany and Palynology 114: 261. [ Links ]
Recibido: 3 de diciembre de 2007.
Aceptado: 15 de septiembre de 2008.

