










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana vol.46 no.1 Buenos Aires ene./mar. 2009
ARTÍCULOS ORIGINALES
Taxaceoxylon katuatenkum sp. nov. en la Formación Koluel-Kaike (Eoceno inferior - medio), Chubut, Argentina: un componente de los bosques subtropicales paleógenos de Patagonia
Mariana Brea1,4, Eduardo Bellosi2,4 y Marcelo Krause3,4
1Laboratorio de Paleobotánica, Centro de Investigaciones Científicas, Diamante. CICYTTP-Diamante, Dr. Materi y España SN, E3105BWA Diamante, Entre Ríos, Argentina, cidmbrea@infoaire.com.ar
2Museo Argentino de Ciencias Naturales. Av. Ángel Gallardo 470, C1405DJR Buenos Aires, Argentina. ebellosi@sei.com.ar
3Museo Paleontológico Egidio Feruglio. Av. Fontana 140, 9100 Trelew, Chubut, Argentina. krausemarcelo@yahoo.com
4Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET).
Resumen. Se describe en este trabajo un fragmento de tallo silicificado, Taxaceoxylon katuatenkum sp. nov., coleccionado en la localidad Kilómetro 170, al SE de Chubut (Argentina). Este leño corresponde al segundo registro en Argentina del género Taxaceoxylon y proviene de sedimentos piroclásticos del Eoceno inferior - medio de la Formación Koluel-Kaike en Patagonia central. Tal registro fósil indica que las familias Taxaceae y/o Cephalotaxaceae tuvieron una distribución más amplía en el pasado y que estas familias no estuvieron confinadas al Hemisferio Norte. De acuerdo a la información paleobotánica y paleopedológica de la Formación Koluel-Kaike, esta nueva especie de Taxaceae y/o Cephalotaxaceae habría sido un componente de los bosques desarrollados en ambientes cálidos y húmedos de Patagonia central durante el Eoceno temprano-medio. Esta flora de carácter subtropical, expandida en América del Sur hasta latitudes medias (47° S), representa la transición entre la vegetación de bosques lluviosos y bosques esclerófilos megatérmicos durante el Óptimo climático del Eoceno temprano (EECO), y las sabanas con palmeras y pastizales desarrollados en el Eoceno medio a tardío.
Abstract. Taxaceoxylon Katuatenkum Sp. Nov. From The Koluel-Kaike Formation (Lower - Middle Eocene), Chubut, Argentina: A Component Of Paleogene Subtropical Forests Of Patagonia. This paper describes a silicified trunk fragment, Taxaceoxylon katuatenkum sp. nov., collected from Kilómetro 170 locality, SE of Sarmiento city (Chubut, Argentina). This fossil stem is the second record in Argentina of the Taxaceoxylon genus, and comes from pyroclastic sediments of the Lower - Middle Eocene Koluel-Kaike Formation of central Patagonia. This fossil record indicates that the Taxaceae and/or Cephalotaxaceae families were more widespread in the past than today and that these families were not confined to the Northern Hemisphere. According to the paleobotanical and paleopedological information of the Koluel- Kaike Formation, this new taxon of Taxaceae and/or Cephalotaxaceae would have been a component of the forests developed in humid and warm environments of central Patagonia during the Lower - Middle Eocene. This subtropical flora expanded in South America at mid latitude (47° S), represents the transition between the rainforests and dry forests (megathermal palaeocommunities) during the Early Eocene Climatic Optimum (EECO), and the savannas with palms and grasslands developed by the Middle - Late Eocene.
Key words. Patagonia; Eocene; Fossil wood; Taxaceae or Cephalotaxaceae; Paleoclimate; Koluel-Kaike Formation.
Palabras clave. Patagonia; Eoceno; Leño fósil; Taxaceae o Cephalotaxaceae; Paleoclima; Formación Koluel-Kaike.
Introducción
La transición Cretácico-Terciario y el Paleógeno temprano corresponden a un momento significativo donde ocurren marcados cambios de la composición florística global, que van a producir el conjunto ancestral que dará origen posteriormente a la vegetación moderna. Este rango de tiempo geológico coincide con la mayor actividad tectónica y volcánica asociada con la ruptura del supercontinente de Gondwana (Poole, 2000).
En la Cuenca San Jorge (Patagonia central) se produce durante el Mastrichtiano-Daniano la primera transgresión marina atlántica, cuyo registro sedimentario se conoce con el nombre de Formación Salamanca (Ihering, 1907). A continuación de este evento marino, se instaló un sistema de sedimentación continental entre el Paleoceno tardío y el Eoceno medio, representado por el Grupo Río Chico, constituido por tres formaciones, Formación Peñas Coloradas, Las Flores y Koluel-Kaike (Legarreta y Uliana, 1994; Flynn y Swisher, 1995; Marshall et al., 1997; Krause y Bellosi, 2006).
El Grupo Río Chico es portador de una paleoflora preservada en forma de palinomorfos, fitolitos y troncos permineralizados (Romero 1973; Lema et al., 1999; Ruiz et al., 1999; Brea y Zucol 2006; Brea et al., 2004, 2008; Zucol et al., 2004; Andreis, en prensa; Raigemborn et al., 2009; Zucol et al., en prensa).
Las maderas fósiles del Paleoceno-Eoceno de la Patagonia central son aún muy poco conocidas, pero unos pocos registros sugieren que las coníferas eran componentes abundantes y habrían conformado paleocomunidades arbóreas durante este período (Brea et al., 2007; Raigemborn et al., 2009).
En la presente contribución se describe una nueva conífera, representante de la Familia Taxaceae y/o Cephalotaxaceae asignada al morfogénero Taxaceoxylon Kräusel et Jain 1964. El material corresponde a un leño permineralizado hallado en la Formación Koluel-Kaike, en la localidad Kilómetro 170, Chubut, Argentina (figura 1).
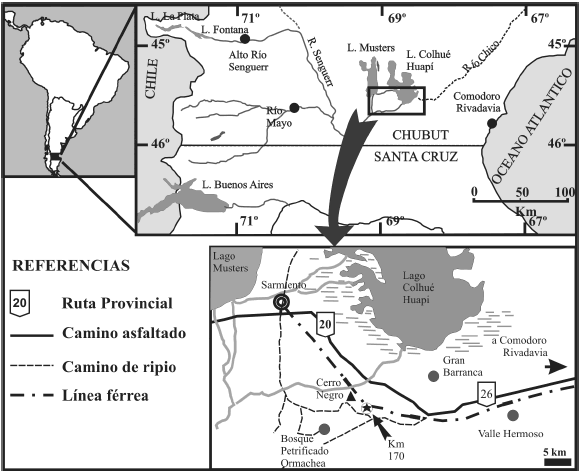
Figura 1. Localización del área de estudio (Kilómetro 170) en la provincia del Chubut (Patagonia central), donde se expone la Formación Koluel-Kaike portadora de leños fósiles/ studied area (Kilometro 170) in Chubut province (Central Patagonia), where the fossil wood bearing Koluel-Kaike Formation is exposed.
En la actualidad la Familia Taxaceae tiene una gran distribución en el Hemisferio Norte, particularmente en Europa, África del Norte, Asía y América del Norte. Austrotaxus Compton es el único género actual con distribución en el Hemisferio Sur y restringido a Nueva Caledonia (Cheng et al., 2000). La Familia Cephalotaxaceae es monotípica integrada por 5 especies del género Cephalotaxus Sieb. et Succ. ex Endl. y tiene un distribución restringida a los bosques subtropicales lluviosos y templados-fríos de China, Corea y Japón (Page, 1990).
El registro fósil de las familias Taxaceae y/o Cephalotaxaceae en el Hemisferio Sur es también muy escaso. En América del Sur los ejemplares más antiguos podrían corresponder a Sommerxylon un leño permineralizado del Triásico superior de la cuenca del Paraná en Brasil, que tiene una gran afinidad con las Taxaceae (Pires y Guerra-Sommer, 2004) y a Taxaceoxylon sp. del Triásico superior de la Precordillera riojana en Argentina (Caminos et al., 1995). También se conocen posibles leños en el Jurásico de la India (Kräusel y Jain, 1964; Sharma, 1970), en tanto que la primera evidencia en Nueva Zelanda de esta familia se registra durante el Mioceno (Pole, 2007).
El nuevo taxón descripto en este trabajo, Taxaceoxylon katuatenkum sp. nov., del Eoceno de la Patagonia central correspondería al segundo registro de esta familia en Argentina.
Marco geológico y paleontológico
Las maderas fósiles analizadas provienen de la sección media de la Formación Koluel-Kaike en la localidad Kilómetro 170 (S 45° 46´02,2´´, O 68° 52´44,3´´), al sudeste de la ciudad de Sarmiento, provincia del Chubut (figura 1). Este sitio se ubica 3,5 km al SE del Cerro Negro, próximo a las vías abandonadas del antiguo circuito férreo que unía las ciudades de Comodoro Rivadavia y Sarmiento. El acceso es posible a través de las rutas provinciales N° 26 y 20.
El área donde se estudió la sucesión paleógena se emplaza sobre el borde oriental de la Faja plegada San Bernardo. En este sector de la Cuenca San Jorge se expone un conjunto de secuencias continentales y marinas del Cretácico superior a Mioceno medio (figura 2). Originalmente denominada "Argiles Fissilaires" por Ameghino (1906), la Formación Koluel- Kaike es la unidad superior del Grupo Río Chico, constituido además por las Formaciones Peñas Coloradas y Las Flores (Legarreta y Uliana, 1994).

Figura 2. Estratigrafía del Cenozoico en la región centro-sur del Chubut. Cronología del Paleógeno-Neógeno según Gradstein et al. (2004) / Cenozoic stratigraphy in south-central Chubut. Paleogene- Neogene chronology after Gradstein et al. (2004).
Los afloramientos analizados forman parte de una extensa barranca que hacia el NE se continúa en la clásica localidad paleontológica de Gran Barranca- Las Flores, al sur del lago Colhue-Huapi; en tanto que hacia el SO se prolongan hasta el Bosque Petrificado José Ormaechea (figura 1). En Kilómetro 170 aflora también la Formación Sarmiento (Eoceno medio-Mioceno inferior), cubierta en su techo por depósitos cuaternarios de grava (figura 3). Aquí la sucesión paleógena se encuentra suavemente plegada, con buzamientos de menos de 3° al sur. La Formación Koluel-Kaike exhibe límites concordantes y transicionales tanto en su base con la Formación Las Flores, como en su techo con el Miembro Gran Barranca de la Formación Sarmiento (McCartney, 1934; Feruglio, 1949). Este último contacto se reconoce por el aumento en la frecuencia de chonitas blanquecinas relativamente friables, en detrimento de las chonitas amarillo-anaranjadas silicificadas.

Figura 3. Chonitas y bentonitas de la Formación Koluel-Kaike (aproximadamente 20 metros de espesor) en la localidad Kilómetro 170. Por encima yace la Formación Sarmiento/ pyroclastic mudstones and bentonites from the Koluel-Kaike Formation (approximately 20 meters thick) at Kilometro 170 locality. The Sarmiento Formation is lying on top.
La edad del Grupo Río Chico es poco precisa debido a la ausencia de dataciones isotópicas y de información paleontológica útil. La asignación cronológica más amplia, sustentada exclusivamente en sus relaciones con unidades colindantes, comprende el lapso Paleoceno tardío - Eoceno medio (Legarreta y Uliana, 1994; Flynn y Swisher, 1995; Marshall et al., 1997; Krause y Bellosi, 2006). La existencia de discontinuidades estratigráficas de valor hiatal incierto dentro de la sucesión paleógena continental de Patagonia central (Legarreta y Uliana, 1994) incorpora aún mayor incertidumbre a este problema. Particularmente, las relaciones estratigráficas de la Formación Koluel-Kaike sugieren una edad máxima Paleoceno cuspidal (?) - Eoceno temprano. Por otra parte, su antigüedad mínima puede ser estimada con menor grado de incertidumbre. La misma alcanzaría el Eoceno medio considerando el contacto transicional con el Miembro Gran Barranca de la Formación Sarmiento, cuyas numerosas dataciones radimétricas y perfiles de polaridad geomagnética (Madden et al., 2005) asignan su base a los Crones 19-20 (~ 42 Ma). En consecuencia, la Formación Koluel-Kaike es aquí referida al Eoceno inferior - medio (figura 2).
En la localidad Kilómetro 170, la Formación Koluel- Kaike está integrada por 33 metros de fangolitas piroclásticas o chonitas, bentonitas y tobas finas (figura 4). Las maderas estudiadas se encuentran silicificadas y ocurren en chonitas macizas en forma de fragmentos con evidencias de haber soportado cierto transporte. Las chonitas son dominantemente vítreas, cementadas con sílice y de color gris claro, amarillo, naranja y rojo, con frecuentes nódulos de hierro y manganeso (McCartney, 1934; Feruglio 1949).
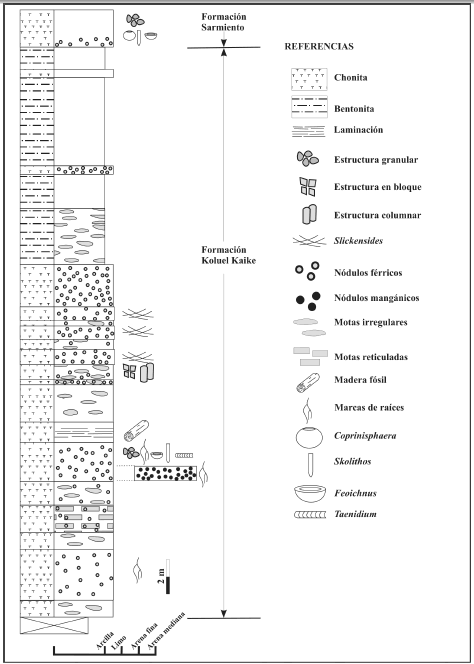
Figura 4. Perfil sedimentológico de la Formación Koluel-Kaike en la localidad Kilómetro 170 / sedimentologic profile of the Koluel-Kaike Formation at Kilometro 170 locality.
Los estratos choníticos y bentoníticos presentan marcada tabularidad y continuidad lateral, con fuerte modificación pedogénica (Krause y Bellosi 2006).
Los paleosuelos exhiben horizontes bien definidos, moteados y con estructuras variables, desde maciza a bloque subangular, laminar y granular. Ocasionalmente, se preservan relictos de laminación o estratificación primaria. Los principales elementos estructurales pedogénicos consisten en varios tipos de glébulas de color naranja-amarillento a castaño rojizo, de composición férrica y de color gris oscuro a negro, de composición mangánica, inmersas en una matriz gris clara, amarillenta o gris oliva claro. En los paleosuelos con abundantes glébulas férricas resultan evidentes los rasgos de iluviación de arcillas.
La relación alúmina/bases varía de 3 a 16, en tanto que la arcilla predominante es caolinita. La asociación de atributos estructurales junto con la composición geoquímica y mineralógica ha permitido interpretar estos paleosuelos como Ultisoles de fuerte a moderado grado de desarrollo, formados bajo intenso hidromorfismo y vegetación dominantemente arbórea (Krause y Bellosi, 2006). Un rasgo de interés de estos paleosuelos es su asociación con niveles lenticulares portadores de abundantes glébulas de óxido de Mn (pirolusita y psilomelano), y también ricos enóxidos de Fe, ya sea como nódulos férricos o formando horizontes plintíticos. El estudio de la distribución del par Mn-Fe sugeriría alternancia de condiciones reductoras-oxidantes debidas a períodos de saturación en agua y desecación del suelo (Krause et al., 2008a). En menor proporción, hacia la sección media y superior de la unidad, se reconocen Andisoles gris claros a blanquecinos, con menor grado de modificación pedogénica, escasez de glébulas férricas, espesores menores al metro y estructura maciza o en bloque pobremente desarrollada, o bien con estratificación relíctica.
A diferencia de lo reconocido en paleosuelos de la Formación Sarmiento, en la Formación Koluel-Kaike los icnofósiles son infrecuentes (Krause et al., 2007; Bellosi et al., en prensa), restringiéndose a rizolitos de variadas dimensiones, algunos de más de un metro de largo, y cámaras de alimentación de ninfas de cigarras denominadas Feoichnus challa (Krause et al., 2008b). Las bentonitas, subordinadas respecto de las chonitas silicificadas, son de color gris amarillento a gris oliva y afloran en típico paisaje de mal país. Por lo común, presentan aspecto macizo, junto a frecuentes slickensides y rizolitos muy finos. Las tobas son escasas, de granulometría fina a muy fina, con bajo grado de silicificación. Por lo general se muestran macizas, aunque algunas exhiben laminación paralela o estratificación cruzada en artesa de escala mediana.
El conjunto de rasgos sedimentarios observados en la Formación Koluel-Kaike sugiere un ambiente depositacional de planicies donde prevalecieron las caídas de polvo y cenizas volcánicas (i.e. depósitos loéssicos). En modo subordinado se desarrollaron cursos fluviales poco jerarquizados, los cuales retrabajaron los materiales téfricos. Repetidas y persistentes interrupciones en la sedimentación subaérea, y en menor medida fluvial, favorecieron la edafización de los sedimentos en condiciones climáticas subtropicales estacionales (Krause y Bellosi, 2006).
La Formación Koluel-Kaike es portadora de la fauna de Ernestokokenia (Simpson, 1935) que define la Edad Mamífero Riochiquense (Bond et al., 1995; Flynn y Swisher, 1995; Marshall et al., 1997). Esta asociación, registrada principalmente en las localidades de Bajo Palangana, Cerro Redondo y Punta Peligro (SE del Chubut), incluye tres familias de marsupiales y siete de notoungulados (Reguero et al., 2002). Algunos de sus géneros son compartidos tanto con la antecedente fauna de Kibenikhoria (Edad Itaboraiense) perteneciente a la Formación Las Flores, como con la subsiguiente fauna de Subedad Barranquense recuperada del Miembro Gran Barranca de la Formación Sarmiento (Bond et al., 1995). Los ungulados del Riochiquense presentan en su mayoría coronas bajas, aunque ocurren también algunos tipos mesodontes (Pascual y O. Jaureguízar, 1990). Reguero et al. (2002) consideran que estos mamíferos ocuparon áreas arboladas de clima cálido y húmedo.
Los estudios sobre maderas fósiles de la Formación Peñas Coloradas y de la sección inferior del Grupo Río Chico s.l. en las localidades Punta Peligro Norte, Puerto Visser y Bosque Petrificado José Ormaechea respectivamente, avalan la presencia de las familias Podocarpaceae, Styracaceae, Araliaceae, Cunoniaceae y Boraginaceae (Brea et al., 2004; Brea y Zucol, 2006, Raigemborn et al., 2009). La paleoflora registrada en la Formación Las Flores esta basada en los estudios de las asociaciones fitolíticas, caracterizadas por la abundancia de angiospermas y palmeras. Se reconocieron diversas familias: Arecaceae, Chrysobalanaceae, Mimosaceae, Magnoliaceae, Annonaceae, Burseraceae, Poaceae, Cyperaceae, Podostemaceae y el orden Zingiberales. También han sido encontradas espículas de esponjas, quistes de Chrysostomataceae y diatomeas. Estas asociaciones conformarían bosques tropicales a subtropicales húmedos, expuestos a condiciones climáticas cálidas y húmedas (Brea et al., 2008; Raigemborn, et al., 2009).
Las asociaciones fitolíticas de la Formación Koluel- Kaike en la localidad fosilífera de Gran Barranca, ubicada al sur del lago Colhue-Huapi indican una paleovegetación compuesta por bosques subtropicales a tropicales mixtos, con una gran abundancia de palmeras, así como también se registran comunidades con una mayor proporción de componentes herbáceos (Zucol et al., 2004). En cercanías de la laguna Payahilé, 130 km al norte de Gran Barranca, Andreis (en prensa) describe también para esta unidad tobácea ("Unidad A, Formación Sarmiento") una asociación fitolítica compuesta por palmeras, gramíneas y ciperáceas.
Materiales y métodos
El material corresponde a un leño permineralizado por silicificación, que no muestra procesos de aplastamiento y tiene buena preservación de la anatomía del xilema secundario. Para su estudio se realizaron secciones delgadas en sus tres cortes característicos (corte transversal, longitudinal tangencial y longitudinal radial).
El ejemplar fósil fue hallado en la localidad fosilífera Kilómetro 170, provincia del Chubut (figura 1) en los sedimentos de la Formación Koluel-Kaike (figura 4).
La terminología utilizada corresponde al Glosario de Términos de la Asociación Internacional de Anatomistas de Maderas (IAWA, 2004) y se utilizaron las clasificaciones estándar de Chattaway (1932). En este trabajo, el leño fósil fue identificado a nivel de morfogénero utilizando las publicaciones sobre identificación y descripción de maderas actuales y fósiles (Greguss, 1955; Tortorelli, 1956; Roy, 1972; Bamford y Philippe, 2001; García Esteban et al., 2002; 2004, Pires y Guerra-Sommer, 2004; Harland, et al., 2007; Philippe y Bamford, 2008).
Las mediciones que se dan en las descripciones constituyen promedios de 20 medidas, en todos los casos se cita primero la media y entre paréntesis las medidas mínimas y máximas del rango de distribución.
Los ejemplares se encuentran depositados en la Universidad Nacional de la Patagonia San Juan Bosco (sede Comodoro Rivadavia), bajo el número UNPSJB-PB 194.
El material fue estudiado con un microscopio Nikon Labophot y Nikon Eclipse E200. Las microfotografías fueron tomadas con una cámara digital Nikon Coolpix S4.
Paleontología sistemática
Orden PINALES Dumortier 1829
Familia TAXACEAE Gray 1821 o
CEPHALOTAXACEAE Neger 1907
Morfogénero Taxaceoxylon Kräusel et Jain 1964
Morfoespecie tipo. Taxaceoxylon rajmahalense Kräusel et Jain 1964
Taxaceoxylon katuatenkum sp. nov.
Figuras 5-6
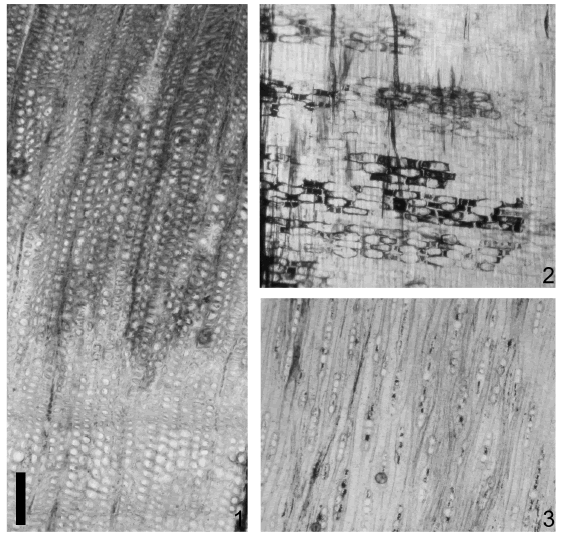
Figura 5. Taxaceoxylon katuatenkum sp. nov. UNPSJB-PB 194. 1. Traqueidas del xilema secundario en corte transversal y anillo de crecimiento poco demarcados / tracheids of secondary xylem in cross section and subtle growth ring. 2. Radios homocelulares en corte longitudinal radial / Rays homocellular in radial section. 3. Radios uniseriados y raramente biseriados en corte longitudinal tangencial / Rays uniseriate and rarely biseriate in tangential section. Escala gráfica / bar scale: 1,2,3 = 200 μm.
Derivatio nominis. El epíteto específico esta conformado por las palabras de la lengua Tehuelche (pobladores originales de la Patagonia extrandina) "kat": madera y "uatenk": quebrado, roto.
Diagnosis. Secondary picnoxylic wood with subtle growth rings. Tracheids circular to irregular in outline. Tracheids with abietoid rounded pits. Tracheid pitting in radial walls uniseriate and isolated. Cross fields with cupressoid pits, small, 5-6 pits per cross-field, irregularly disposed. Radial system homogeneous, rays homocellular, uniseriate, rarely biseriate. The height average is 6 cells (2 - 18 cells). Radial helical thickenings, simple or double helical thickenings, some bifurcated, inclination against the wall of tracheids 45-70°.
Diagnosis. Xilema secundarios picnoxilíco con anillos de crecimiento levemente demarcados. Traqueidas de contorno circular a irregular. Traqueidas con punteaduras tipo abietinoide. Punteaduras uniseriadas y aisladas en las paredes radiales de las traqueidas. Campos de cruzamiento con punteaduras del tipo cupressoides, pequeñas, de 5-6 por campo, dispuestas irregularmente. Sistema radial homogéneo, radios homocelulares, uniseriados, raramente biseriados. La altura promedio es de 6 células (2 - 18 células). Engrosamientos espiralados en las paredes radiales, simples y dobles, algunos bifurcados, con un ángulo de disposición entre 45-70°.
Descripción. Leño con xilema picnoxilico. Anillos de crecimiento levemente demarcados (figura 5.1). En corte transversal las traqueidas son circulares a irregulares (figura 6.1). El diámetro promedio radial de las traqueidas es de 32 μm (20 μm - 50 μm) y el diámetro promedio tangencial de 37 μm (22 μm - 60 μm). El número promedio de traqueidas que separan los radios es de 4 (2 - 8) hileras. En corte longitudinal radial las paredes de las traqueidas presentan punteaduras areoladas, de contorno y aperturas circulares, aisladas, del tipo o plan abietinoide (figuras 6.3, 6.8-9). Las punteaduras presentan disposición uniseriadas (figuras 6.3, 6.9) y tienen un diámetro promedio de 11 μm (10μm - 13 μm). En los campos de cruzamiento se observan de 5 a 6 punteaduras pequeñas del tipo cupressoide irregularmente dispuestas (figura 6.5). Las paredes radiales de las traqueidas presentan engrosamientos espiralados, tanto levógiros y dextrógiros, simples y dobles, siendo algunos bifurcados (figuras 6.6, 6.8). El espesor de los engrosamientos espiralados varía desde 1,27 μm a 2,54 μm y su ángulo de disposición es entre 45-70°. Se observan "spiral grooves" (figuras 6.7 y probablemente 6.10) que son el producto de la desorganización de las paredes secundarias de las traqueidas. Estas estructuras pueden confundirse con los engrosamientos espiralados que son estructuras producidas en la cara interna de las traqueidas (IAWA, Committee, 2004). En corte longitudinal tangencial los radios son homocelulares (figuras 5.2; 6.2), uniseriados siendo muy escasos los biseriados (figuras 5.3; 6.4). Sistema radial homogéneo. La altura media en número de células es de 6 (2 - 18). La altura media de los radios es de 211μm (67 μm - 559 μm) y el ancho promedio es de 28 μm (20 μm - 39 μm).

Figura 6. Taxaceoxylon katuatenkum sp. nov. UNPSJB-PB 194. 1. Detalle de traqueidas en corte transversal mostrando contorno circular a irregular / detail of tracheids in cross section showing circular to irregular outline. 2. Radios homocelulares y traqueidas con engrosamientos espiraladados en corte longitudinal radial / rays homocellular and tracheids with radial spiral thickenings in radial section. 3 y 9. Traqueidas en corte longitudinal radial mostrando las punteaduras tipo abietinoide / tracheids in radial section showing abietoide rounded pits. 4. Radios uniseriados y escasos biseriados en corte longitudinal tangencial / rays uniseriate and scarcely biseriate in tangential section. 5. Campos de cruzamiento con punteaduras cupressoides, pequeñas, 5-6 punteaduras por campo e irregularmente dispuestas / cross fields with 5-6 small irregularly disposed cupressoid pits. 6 y 8. Traqueidas mostrando los engrosamientos espiralados / tracheids with radial spiral thickenings. 7. Traqueidas mostrando "spiral grooves"/ tracheids showing spiral grooves. 10. Corte longitudinal radial mostrando punteaduras en campos de cruzamiento y probables "spiral grooves" / radial section showing pits in cross fields and probable spiral grooves. Escala gráfica / bar scale: 1,2, 3,4, = 100 μm; 5,6,7,8,9,10 = 50 μm.
Procedencia geográfica. Kilómetro 170, Chubut Argentina.
Procedencia estratigráfica. Formación Koluel-Kaike.
Edad. Eoceno inferior - medio.
Holotipo. UNPSJB-PB 194.
Comparaciones con material fósil. El material fue asignado al género Taxaceoxylon debido a que comparte caracteres diagnósticos de importancia xilológica con este morfotaxón. El carácter más relevante para cotejar el material fósil fue la presencia de engrosamientos espiralados en el xilema secundario (Pires y Guerra-Somer, 2004; Philippe y Bamford, 2008). De acuerdo a Barefoot y Hankins (1982), los engrosamientos espiralados tienen carácter genético e importancia diagnóstica.
El morfogénero Taxaceoxylon Kräusel (1949) fue propuesto para sustituir a Taxoxylon Kraus (1870), originalmente descripto como Taxites scalariformes (Goeppert) {= Taxoxylon goepperti Unger = Taxaceoxylon scalariforme (Goeppert) Seward (1919)} (Pires y Guerra-Sommer, 2004). Aplicando el Código Internacional de Nomenclatura Botánica (Greuter et al., 1999), Banford y Phillippe (2001), proponen que Taxoxylon debe considerarse inválido y que se debe utilizar el epíteto Taxaceoxylon.
Este morfotaxón incluye ejemplares con xilema secundario homoxilíco con verdaderos engrosamientos espiralados y punteaduras de tipo abietinoide en las paredes radiales de sus traqueidas, radios medulares homocelulares, ausencia de células o canales resiníferos y ausencia de parénquima axial (Kräusel, 1949; Pires y Guerra-Sommer, 2004; Harland et al., 2007; Philippe y Bamford, 2008).
Las morfoespecies reconocidas dentro de Taxaceoxylon son: T. rajmahalense (Bardwaj) Kräusel et Jain 1964, T. cupressoides Sharma 1970, T. mcmurrayensis Roy 1972, T. japonomesozoicum Nishida 1973, T. antiquum (Boeshore y Gray) Kräusel 1949 y T. torreyanum Shimakura 1936. El cuadro 1 muestra los caracteres más relevantes del morfogénero y su comparación con el material estudiado en este trabajo. Las mayores similitudes fueron halladas con T. rajmahalense registrado para el Jurásico de la India. Sin embargo, el ejemplar hallado en la Formación Koluel-Kaike (Patagonia central, Argentina) se diferencia por presentar engrosamientos espiralados sólo en las paredes radiales de las traqueidas, y por presentar engrosamientos tanto simples, dobles y bifurcados, levógiros y dextrógiros. Por este motivo se asigna una nueva entidad morfoespecífica por el conjunto de caracteres diagnósticos distintivos de las restantes morfoespecies descriptas. Al mismo tiempo, soportan la creación de un nuevo taxón las diferencias en la localidad geográfica y estratigráfica de ambos taxa.
Cuadro 1. Comparación de las características diagnósticas de Taxaceoxylon katuatenkum sp. nov. con las especies asignadas a Taxaceoxylon Kräusel et Jain 1964 / comparison of diagnostic features of Taxaceoxylon katuatenkum sp. nov. with the species assigned to Taxaceoxylon Kräusel et Jain 1964.

Comparaciones con material actual. El material fósil descripto esta estrechamente ligado a los representantes de las familias Taxaceae y/o Cephalotaxaceae. Estos están caracterizados por presentar anillos de crecimiento anuales demarcados o indistinguibles, traqueidas de contorno redondeadas o angulares en sección transversal, conductos resiníferos ausentes, en algunos géneros presentan ocasionalmente parénquima axial, las traqueidas presentan engrosamientos espiralados, campo de cruzamiento con punteaduras del tipo podocarpoide o cupressoide en número de 1-3 (4- 6), las paredes tangenciales de las células radiales son siempre lisas, mientras que las paredes horizontales esporádicamente presentan engrosamientos, la altura de los radios es de 1-28 células de alto, los radios son uniseriados ocasionalmente biseriados (Greguss, 1955; García Esteban et al., 2002; 2004). Taxus L., Pseudotaxus Cheng y Austrotaxus Compton son los taxones actuales que comparten más caracteres diagnósticos con el material fósil descripto en este trabajo.
Discusión
El Orden Coniferales o Pinales comprende a las verdaderas coníferas y esta integrado por 9 familias: Pinaceae, Phyllocladaceae, Podocarpaceae, Araucariaceae, Sciadopityaceae, Cupressaceae, Taxodiaceae, Cephalotaxaceae y Taxaceae (Chaw et al., 1997). Históricamente, la familia de los tejos era excluida de las Coniferales y conformaba un Orden propio -las Taxales- (Sahni, 1920; Florin, 1948, 1951), debido a que sus taxones comparten características propias que los diferencian de las otras coníferas. Sin embargo, las evidencias genéticas indica que los tejos son monofiléticos junto a otras coníferas y que conforman una familia dentro del orden Coniferales o Pinales (Cheng et al., 2000).
Las relaciones entre las familias actuales de las Pinophyta han sido estudiadas a través de secuencias génicas de clorosplastos, mitocondrias y núcleos (Chaw et al., 1997, 2000; Cheng et al., 2000), de estos análisis se infiere que las Taxaceae están próximas a las Cephalotaxaceae y Taxodiaceae, y en menor grado a las Sciadopityaceae y Cupressaceae. Las Phyllocladaceae- Podocarpaceae y las Araucariaceae formarían un clado independiente, mientras que las Pinaceae constituirían un clado segregado de todas las anteriores (Chaw et al., 1997).
La filogenia de los géneros de la familia Taxaceae y la monotípica familia Cephalotaxaceae ha sido extraordinariamente controversial. El género Cephalataxus primeramente estuvo incluido dentro de las Taxaceae y luego fue elevado a nivel de familia, Cephalotaxaceae por Neger (1907). Las relaciones entre los géneros de las familias Taxaceae y Cephalotaxaceae han sido debatidas por largos años (véase Cheng et al., 2000).
La Familia Taxaceae está integrada por árboles o arbustos perennifolios, monoicos o diocos, no resinosos, con hojas aciculares, opuestas o en verticilos trímeros. Sus estructuras reproductivas femeninas no están dispuestas en estróbilos y las semillas están parcial o totalmente cubiertas por una masa carnosa. Esta familia esta integrada por cinco géneros, Taxus L., Torreya Arnott, Amentotaxus Pilger, Austrotaxus Compton, Pseudotaxus Cheng y 17 especies. Los miembros de esta familia están distribuidos principalmente en el Hemisferio Norte y en Nueva Caledonia (Hils, 1993).
La Familia Cephalotaxaceae comprende pequeñosárboles y arbustos perennifolios, que se caracterizan por no poseer conductos secretores en sus órganos, son monoicos o dioicos. Las hojas son lineares a lanceoladas y tienen disposición espiralada. Los conos femeninos están reducidos, con una o pocas escamas ovulíferas, llevando una semilla por cada escama. Las semillas están cubiertas totalmente por un arilo carnoso. Está conformada por un género, Cephalotaxus Sieb. et Succ. ex Endl., y siete especies originarias de la región del Himalaya en Asia Oriental (García Esteban et al., 2002).
Cheng et al. (2000) utilizaron las secuencias del gen cloroplasto matK y secuencias nucleares ITS, para analizar seis géneros de las familias Taxaceae y Cephalotaxaceae junto a representantes de otras familias de coníferas. A partir de este estudio determinaron que los géneros de las Taxaceae y el único género de las Cephalotaxaceae son monofiléticos y el clado Taxodiaceae/Cupressaceae es su grupo hermano. Cephalotaxus es basal a los géneros de las taxáceas, de los que surgen dos clados, Torreya/Amentotaxus y Taxus/ Pseudotaxus/Austrotaxus. El género Austrotaxus es el primero en bifurcarse.
Se estima que la divergencia entre los clados Taxodiaceae/ Cupressaceae y Cephalotaxaceae/Taxaceae fue hace 192-230 Ma y la divergencia entre las taxáceas y Cephalotaxus ocurrió hace 149-179 Ma (Cheng et al., 2000).
En el Hemisferio Sur los registros fósiles más antiguos de la Familia Taxaceae se remontan al Triásico superior de la Cuenca Paraná (Estado de Rio Grande do Sul, Brasil) con la presencia de Sommerxylon spiralosus Pires et Guerra-Sommer un leño permineralizado con afinidad a las taxáceas. Esta morfoespecie preservó anatómicamente la médula, el xilema primario, el xilema secundario, el floema y la corteza.
En Argentina, el morfogénero Taxaceoxylon fue recolectado en sedimentos del Triásico superior, en el tramo medio del río del Peñon y a lo largo de la Quebrada de Santo Domingo, Precordillera de la provincia de La Rioja (Caminos et al., 1995). Este ejemplar fue hallado en asociación con leños permineralizados de Rhexoxylon sp.
Durante el período Jurásico se registran Taxaceoxylon rajmahalense (Bardwaj) Kräusel et Jain 1964 y Taxaceoxylon cupressoides Sharma 1970, dos leños fósiles de Taxaceae provenientes de la India (Kräusel y Jain, 1964; Sharma, 1970).
Los taxones anteriormente mencionados avalan la hipótesis que las Taxaceae mesozoicas no estaban confinadas al Hemisferio Norte (Kräusel y Jain, 1964; Florin, 1940; Pires y Guerra-Sommer, 2004).
Recientemente, se reportó la primera evidencia de esta familia en el Mioceno temprano del sur de Nueva Zelanda con el hallazgo de compresiones foliares (Pole, 2007).
El taxón descripto en el presente trabajo, Taxaceoxylon katuatenkum sp. nov. , del Eoceno de la Patagonia central correspondería al segundo registro de esta familia en Argentina. Este nuevo registro refuerza la hipótesis de que las Taxaceae o Cephalotaxaceae también ocuparon el Hemisferio austral desde tiempos mesozoicos. Junto a los ejemplares de Nueva Zelanda se reafirma y amplía la distribución de esta familia durante el Cenozoico en el Hemisferio Sur.
La asociación del nuevo leño de Taxaceae o Cephalotaxaceae, junto con otros fósiles y las propiedades sedimentológicas y paleopedológicas que exhibe la Formación Koluel-Kaike, permiten efectuar algunas inferencias paleoambientales para el Eoceno temprano- medio de la región central de la Cuenca San Jorge. Según lo atestiguan los datos paleobotánicos y palinológicos, la transición del Eoceno temprano a medio fue un lapso de modificación en la estructura de la flora de Patagonia (Barreda y Palazzesi, 2007; Prámparo et al., 2007). La vegetación de bosques lluviosos y bosques esclerófilos, dominados por palmeras, Podocarpaceae, Araucariaceae, Casuarinaceae, Juglandaceae, y Proteaceae, fue substituida por bosques de mayor diversidad, conformados por Podocarpaceae, Aquifoliaceae, Bombacaceae y especies meso y microtérmicas de Nothofagus Blume (Melendi et al., 2003; Wilf et al., 2005; Palazzesi y Barreda, 2007). Esta tendencia en la transformación florística se profundizó en el Eoceno medio a tardío, con la instalación y posterior expansión de pastizales arbolados, palmares y pastizales abiertos meso-megatérmicos (Pascual y O. Jaureguízar, 1990; Bellosi y González, 2005; Zucol y Brea, 2005; Prámparo et al., 2007; Bellosi et al., en prensa). Recientes estudios de trazas fósiles (i.e. bolas de cría) de escarabajos estercoleros (Coprinisphaera issp.) en la Formación Sarmiento confirman la existencia de ecosistemas de pastizales abiertos a partir del Eoceno medio alto (Sánchez et al., 2008).
La asociación fitolítica de la Formación Koluel- Kaike, caracterizada por la abundancia de angiospermas y palmeras, asociadas a Poaceae, Cyperaceae y Podostemaceae, sugiere la existencia de bosques subtropicales a tropicales mixtos (Zucol et al., 2004) y ambientes subordinados dominados por plantas herbáceas (gramíneas), conformando sabanas (Zucol, 2005). Un paleoclima similar, templado a cálido con largos períodos de alta humedad alternando con estaciones secas más breves, fue interpretado por Andreis (en prensa) para la región de laguna Payahilé. De acuerdo con Reguero et al. (2002), los vertebrados fósiles de esta unidad, integrantes de la Fauna de Ernestokokenia (Edad Mamífero Riochiquense) constituida por ramoneadores, insectívoros y en menor medida frugívoros (Pascual et al., 1996) habrían ocupado áreas boscosas bajo condiciones cálidas y húmedas. Por otro lado, los frecuentes y potentes paleosuelos rojizos y moteados, con marcas de raíces profundas y avanzado grado de desarrollo, son indicativos de un ecosistema maduro y húmedo-subtropical. La vegetación asociada a estos paleosuelos habría sido de carácter arbóreo y de altura considerable, posiblemente dominada por coníferas o especies de madera dura (Krause y Bellosi, 2006). Por su parte, los bancos lenticulares mangánicos permiten definir una significativa meteorización del manganeso, normalmente favorecida en climas cálidos y húmedos y abundancia de ácidos orgánicos (Li y Vasconcelos, 2002). Tales condiciones resultanconcordantes con las interpretadas por el estudio de los paleo-Ultisoles.
Algunos rasgos pedogénicos evidencian estacionalidad climática, la cual no parece ser del todo conciliable con la leve demarcación de los anillos anuales de crecimiento observada en T. katuatenkum sp. nov. Esta diferencia podría indicar que dicha estacionalidad estaría más vinculada al régimen pluvial o hídrico, que a la temperatura.
Conclusiones
Leños fósiles con afinidad a las Taxaceae fueron descriptos anteriormente en Argentina en sedimentos del Triásico superior de La Rioja (Caminos et al., 1995). De esta manera, Taxaceoxylon katuatenkum sp. nov. correspondería al segundo registro de esta familia para el sector más austral de América del Sur. El ejemplar proviene de la Formación Koluel- Kaike del Eoceno inferior-medio de la Patagonia central.
Se conocen sólo seis morfoespecies de leños fósiles taxodiodes, los cuales presentan verdaderos engrosamientos espiralados en sus traqueidas. Estas especies anteriormente descriptas en otras regiones del mundo difieren de Taxaceoxylon katuatenkum sp. nov., motivo por el cual se crea esta nueva entidad específica. Finalmente, Taxaceoxylon katuatenkum sp. nov. amplía la distribución geográfica y estratigráfica del morfogénero Taxaceoxylon.
De acuerdo a la información fosilífera y paleopedológica de la Formación Koluel-Kaike, esta nueva especie de Taxaceae y/o Cephalotaxaceae habría sido un componente de los bosques desarrollados en Patagonia central durante el Eoceno temprano-medio. Esta flora de carácter subtropical, expandida en América del Sur hasta latitudes medias (47° S), representa la transición entre la vegetación de bosques lluviosos y bosques esclerófilos megatérmicos (Barreda y Palazzesi, 2007) correspondientes al Óptimo climático del Eoceno temprano (EECO), y las sabanas con palmeras y pastizales desarrollados en el Eoceno medio a tardío (Pascual y O. Jaureguízar, 1990; Prámparo et al., 2007).
Agradecimientos
Los autores desean expresar su agradecimiento a J. Genise y M.V. Sánchez por su colaboración en las tareas de campo. Agradecemos a dos árbitros anónimos por la revisión de este manuscrito. La ayuda financiera fue provista por la Agencia Nacional de Promoción Científica y Tecnológica (PICT 07-13864 y PICT 03-13286).
Bibliografía
1. Andreis, R.R. en prensa. Silicofitolitos en las formaciones Río Chico (Paleoceno superior) y Sarmiento (Eoceno inferior-Oligoceno superior), Laguna Payahilé, Provincia del Chubut, Argentina. En: A.F. Zucol, A. F., M. Osterrieth y M. Brea (eds.), Fitolitos en suelos, sedimentos y sitios arqueológicos: Estado actual de sus conocimientos en América del Sur. Universidad de Mar del Plata, Mar del Plata. [ Links ]
2. Bamford, M.K. y Phillippe, M. 2001. Jurassic-Early Cretaceous Gondwanan homoxylous woods: a nomenclatural revision of the genera with taxonomic notes. Review of Palaeobotany and Palynology 113: 287-297. [ Links ]
3. Barefoot, A.C. y Hankins, F.W. 1982. Identification of modern and Tertiary woods. Oxford University Press, Oxford, 189 pp. [ Links ]
4. Barreda, V. y Palazzesi. L. 2007. Patagonian vegetation turnovers during the Paleogene-Early Neogene: origin of arid-adapted floras. The Botanical Review 73: 31-50. [ Links ]
5. Bellosi, E.S. y González. M.G. 2005. Paleosuelos de la Formación Sarmiento en Gran Barranca (Chubut, Argentina). 16° Congreso Geológico Argentino (La Plata), 4: 433- 438. [ Links ]
6. Bellosi, E.S., Laza, J. Sánchez, M.V. y Genise, J. En prensa. Ichnofacial analysis of the Sarmiento Formation (Middle Eocene-Lower Miocene) at Gran Barranca, Central Patagonia. En: Madden, R. A. Carlini, M. G. Vucetich y R. Kay (eds.), The Paleontology of Gran Barranca: Evolution and Environmental Change through the Middle Cenozoic of Patagonia. Cambridge University Press. [ Links ]
7. Bond, M., Carlini, A., Goin, F., Legarreta, L., Ortiz J. E., Pascual, R. y Uliana, M. 1995. Episodes in South American Land Mammal evolution and sedimentation: testing their apparent concurrence in a Paleocene succession from Central Patagonia. 6° Congreso Argentino de Paleontología y Bioestratigrafía (Trelew) Actas, pp. 47-58. [ Links ]
8. Brea, M. y Zucol, A.F. 2006. Leños fósiles de Boraginaceae de la Formación Peñas Coloradas (Paleoceno superior), Puerto Visser, Chubut, Argentina. Ameghiniana 43: 139-146. [ Links ]
9. Brea, M, Zucol, A.F., Raigemborn, M.S. y Matheos, S.D. 2004. Leños fósiles del Paleoceno superior (Grupo Río Chico), provincia del Chubut, Argentina. Ameghiniana Suplemento Resúmenes 41: 7R-8R. [ Links ]
10. Brea, M, Zucol, A.F., Raigemborn, M.S. y Matheos, S.D. 2008. Reconstrucción de paleocomunidades arbóreas mediante análisis fitolíticos en sedimentos del Paleoceno superior-Eoceno? (Formación Las Flores), Chubut, Argentina. En: A. Korstange y P. Babot (eds.), Matices Interdisciplinarios en Estudios Fitolíticos y de otros Microfósiles/ Interdisciplinary Nuances in Phytolith and other Microfossil Studies. BAR Internacional Series S1870, Capítulo 9: 91-108.(Oxford, Inglaterra). [ Links ]
11. Brea, M., Matheos, S.D., Raigemborn, M.S. Iglesias, A., Zucol A.F. y Prámparo M. 2007. Aspectos paleoecológicos y paleoambientales del bosque petrificado Ameghino (Daniano), Chubut, Argentina. 4° European Meeting on the Palaeontology and Stratigraphy of Latin America. En: E. Díaz- Martínez e I. Rábano (eds.), Tres Cantos, Madrid, España. Cuadernos del Museo Geominero, n° 8: 45-50. Instituto Geológico y Minero de España, Madrid. [ Links ]
12. Caminos, R., Zamuner, A.B., Limarino, C. y Fauqué, L. 1995. Triásico superior fosilífero en la Precordillera riojana. Revista de la Asociación Geológica Argentina 50: 262-265. [ Links ]
13. Chattaway, M. 1932. Proposed standards for numerical values used in describing woods. Tropical Woods 29: 20-28. [ Links ]
14. Chaw, S.M., Zharkikh, A., Sung, H.M., Lau, T.C. y Li, W.H. 1997. Molecular phylogeny of extant Gymnosperms and seed plant evolution: Analysis of nuclear 18S rRNA sequences. Molecular Biology and Evolution 14: 56-68. [ Links ]
15. Chaw, S.M., Parkinson, C.L., Cheng, Y., Vicent, T.M. y Palmer, J.D. 2000. Seed plant phylogeny inferred form all three plant genomes: monophyly of extant gymnosperms and origin of Gnetales from conifers. Procceedings of the National Academy of Sciences 97: 4086-4091. [ Links ]
16. Cheng, Y., Nicolson, R.G., Tripp, K. y Chaw, S. 2000. Phylogeny of Taxaceae and Cephalotaxaceae Genera Inferred from Chloroplast matK Gene and Nuclear rDNA ITS Region. Molecular Phylogenetics and Evolution 14: 353-365. [ Links ]
17. Dumortier, B.C. 1829. Analyse des Familles des Plantes, avec líndication des proncipaux genres qui s´y rattachent. Imprimeries de J. Casterman, aïné, 105 pp. [ Links ]
18. Feruglio, E., 1949. Descripción geológica de la Patagonia, Tomo 2. Buenos Aires, Dirección General de Yacimientos Petrolíferos Fiscales, 349 pp. [ Links ]
19. Florin, R. 1948. On the morphology and relationships of the Taxaceae. Botanical Gazette 110: 31-39. [ Links ]
20. Florin, R. 1951. Evolution of cordaites and conifers. Acta Horti Bergiani 15: 285-388. [ Links ]
21. Flynn, F. J. y Swisher, C. C. 1995. Cenozoic South American Land mammal ages: correlation to global geochronologies. En: Berggren, W., D. Kent, M-P. Aubry y J. Hardenbol (eds.), Geochronolgy time scales and Global stratigraphic correlations. SEPM, Special Publication 54: 317-333. [ Links ]
22. García Esteban, L., Palacios de Palacios, P., Guindeo Casasús, A., García Esteban, Ly., Lázaro Durán, I., González Fernández, L., Rodríguez Labrador, Y., García Fernández, F., Bobadilla Maldonado, I. y Camacho Atalaya, A. 2002. Anatomía e identificación de coníferas a nivel de especie. Fundación Conde del Valle de Salazar, Ediciones Mundi Prensa, 412 pp. Madrid. [ Links ]
23. García Esteban, L. Palacios de Palacios, L., Guindeo Casasús, A. y García Fernández, F. 2004. Characterisation of the xylem of 352 conifers. Investigación agraria: Sistemas y recursos forestales 13: 452-478. [ Links ]
24. Gray, S.F. 1821. Nat. Arr. Brit. Pl. 2: 222- 226. [ Links ]
25. Greguss, P. 1955. Identification of living Gymnosperms on the basis of xylotomy. Akademiai Kiado, Budapest, 263 pp. [ Links ]
26. Greuter, W., McNeill, J., Barrie, F.R., Burdet, H.M., Demoulin, P.C., Skog, J.E., Trehane, P., Turland, N.J. y Hawaksworth, D.L. 1999. International Code of Botanical Nomenclature. Tokyo Code, 131 pp. [ Links ]
27. Harlang, M., Francis, J.E., Brentnall, S.J. y Beerling, D.J. 2007. Cretaceous (Albian-Aptian) conifer wood from Northern Hemisphere high latitudes: Forest composition and palaeoclimate. Review of Palaeobotany and Palynology 143: 167-196. [ Links ]
28. Hils, M.H. 1993. Taxaceae Gray - Yew Family. En: Flora of North America Editorial Committee. Flora of North America North of Mexico, Volume 2, Pteridophytes and Gymnosperms, Oxford University Press, New York, pp. 423-427. [ Links ]
29. IAWA, Committee. 2004. List of microscopic features for softwood identification. En: H.G. Richter, D. Grosser, I. Heinz y P.E. Gasson (eds.), International Association Wood Anatomist Journal 25: 1-70. [ Links ]
30. Ihering, H. von. 1907. Les mollusques fosiles du Tertiaire et du Crétacé supérieur de l'Argentine. Anales Museo Nacional de Buenos Aires 3: 1-611. [ Links ]
31. Kraus, G. 1870. Bois fossils de conifers. En: Schimper W.P. (ed.),Traité de Paleontologie Végetale. Strasbourg 2: 363-385. [ Links ]
32. Krause, J.M y Bellosi, E. S. 2006. Paleosols from the Koluel-Kaike Formation (Lower-Middle Eocene) in south-central Chubut, Argentina: A preliminary analysis. 4° Congreso Latinoamericano de Sedimentología y 11° Reunión Argentina de Sedimentología (Bariloche), Resúmenes, p. 125. [ Links ]
33. Krause, J.M., Bellosi, E.S. y Genise, J.F. 2007. Evolución de la icnofauna terrestre en la Patagonia central II: el Grupo Río Chico desde el K-T hasta la explosión del Eoceno medio. Reunión Argentina de Icnología (Córdoba), Resúmenes, p. 38. [ Links ]
34. Krause J.M., Bellosi, E.S. e Iglesias M. 2008 a. Niveles mangánicos en la Formación Koluel-Kaike (Eoceno, centro-sur de Chubut): una perspectiva paleopedológica. 12° Reunión Argentina de Sedimentología (Buenos Aires). Resúmenes, p. 90. [ Links ]
35. Krause, J.M., Bown, T.M., Bellosi, E.S. y Genise, J.F. 2008 b. Trace fossils of cicadas from the Cenozoic of Central Patagonia, Argentina. Palaeontology 51: 405-418. [ Links ]
36. Kräusel, R. 1949. Die fossilen Koniferen-Hölaer (unter Ausschluss von Araucarioxylon Kraus) II. Kritische Untersuchungen zur Diagnostic lebender und fossiler Koniferen-Hoölzer. Palaeontographica B 89: 83-203. [ Links ]
37. Kräusel, R. y Jain, K.P. 1964. New fossil coniferous woods from Rajmahal Hills, Bihar, India. Palaeobotanist 12: 59-67. [ Links ]
38. Legarreta, L. y Uliana, M., 1994. Asociación de fósiles y hiatos en el supracretácico Neógeno de Patagonia: una perspectiva estratigráfico- secuencial. Ameghiniana 31: 257-281. [ Links ]
39. Lema, H., Busteros, H., Papú, O. y Sepúlveda, E. 1999. Litología y palinoestratigrafía de la Formación Río Chico en la Localidad de Estancia El Sauce, provincia de Chubut, Argentina. Simposio Paleógeno de América del Sur, Actas. Servicio Geológico Minero Argentino, Anales 33: 33: 51-58. [ Links ]
40. Li, J.W. y Vasconcelos, P.M. 2002. Cenozoic continental wheathering and its implications for paleoclimate: evidence from 40Ar/39Ar geochronology of supergene K-Mn oxides in Mt Tabor, central Queensland, Australia. Earth and Planetary Science Letters 200: 223-239. [ Links ]
41. Madden, R., Bellosi, E., Carlini, A., Heizler, M., Vilas, J., Re, G., Kay, R. y Vucetich, M.G., 2005. Geochronology of the Sarmiento Formation at Gran Barranca and elsewhere in Patagonia: calibrating Middle Cenozoic mammal evolution in South America. 16° Congreso Geológico Argentino (La Plata) 4: 411- 412. [ Links ]
42. Marshall, L., Sempere, T. y Butler, R. 1997. Chronostratigraphy of the mammal-bearing Palaeocene of South America. Journal of South American Earth Sciences 10: 49-70. [ Links ]
43. McCartney, G.C. 1934. The bentonites and closely related rocks of Patagonia. American Museum Novitates 630: 1-16. [ Links ]
44. Melendi, D.L., Scafati, L.H. y Volkheimer, W. 2003. Palynostratigraphy of the Paleogene Huitrera Formation in NW Patagonia, Argentina. Neues Jahrbuch für Geologie und Paläontologie. 228: 205-273. Abhandlungen. [ Links ]
45. Neger, F.W. 1907. Die Nadelhozer (Koniferen) und Ubrigen Gymnospermen. Leipzig. [ Links ]
46. Nishida, M. 1973. On some petrified plants from the Cretaceous of Choshi, Chiba Perfecture 6. Botanical Magazine 86: 189-202. [ Links ]
47. Page, C.N. 1990. Coniferophytina. En: K.U. Kramer y P.S. Green, (eds.), The Families and Genera of Vascular Plants. Vol. 1, "Pteridophytes and Gymnosperms". Stringer-Verlag, Berlin, pp. 282-361. [ Links ]
48. Palazzesi, L. y Barreda, V. 2007. Major vegetation trends in the Tertiary of Patagonia (Argentina): A qualitative paleoclimatic approach based on palynological evidence. Flora - Morphology, Distribution, Functional Ecology of Plants 202: 328-337. [ Links ]
49. Pascual, R. y O. Jaureguízar, E. 1990. Evolving climates and mammal faunas in Cenozoic South America. Journal of Human Evolution 19: 23-60. [ Links ]
50. Pascual, R.,.O. Jaureguízar, E. y Prado, J.L. 1996. Land mammals: paradigm for Cenozoic South America geobiotic evolution. Münchner Geowissenschaftlich Abhandlungen 30: 255-319. [ Links ]
51. Philippe, M. y Bamford, M.K. 2008. A key to morphogenera used for Mesozoic conifer-like woods. Review of Palaeobotany and Palynology 148: 184-207. [ Links ]
52. Pires, E.F. y Guerra-Sommer, M. 2004. Sommerxylon spiralosus from Upper Triassic in southernmost Paraná Basin (Brazil): A new taxon with Taxacean affinity. Anais da Academia Brasileira de Ciências 76: 595-609. [ Links ]
53. Pole, M. 2007. Conifer and cycad distribution in the Miocene of southern New Zealand. Australian Journal of Botany 55: 143-164 [ Links ]
54. Poole, I. 2000. Fossil angiosperm Wood: its role in the reconstruction of biodiversity and palaeoenvironment. Botanical Journal of the Linnean Society 134: 361-381. [ Links ]
55. Prámparo, M.B., Quattrocchio, M., Gandolfo, M.A., Zamaloa, M.C. y Romero, E. 2007. Historia evolutiva de las angiospermas (Cretácico-Paleógeno) en Argentina a través de los registros paleoflorísticos. En: S. Archangelsky, T. Sánchez y E.P. Tonni (eds.), Asociación Paleontológica Argentina. Publicación Especial 11, Ameghiniana 50° aniversario: 157-172. [ Links ]
56. Raigemborn, M.S, Brea, M., Zucol, A.F y Matheos, S.D. 2009. Early Paleogene climate at mid latitude in South America: Mineralogical and paleobotanical proxies from continental sequences in Golfo San Jorge basin (Patagonia, Argentina). Geologica Acta 7: 125-145. [ Links ]
57. Reguero, M., Marenssi, S. y Santillana, S. 2002. Antartic Peninsula and South America (Patagonia) Paleogene terrestrial faunas and environments: biogeographic relationships. Palaeogeography Palaeoclimatology Palaeoecology 179: 189-210. [ Links ]
58. Romero, E.J. 1973. Polen fósil de "Nothofagus" (Nothofagidites) del Cretácico y Paleoceno de Patagonia. Revista del Museo de La Plata (Nueva Serie), Paleontología 6: 290-305 [ Links ]
59. Roy, S.K. 1972. Fossil wood of Taxaceae from McMurray Formation (Lower Cretaceous) of Alberta. Canadian Journal of Botany 50: 349-352. [ Links ]
60. Ruiz, L., Quattrocchio, M., Guerstein, R.G. y Volkheimer, W. 1999. Rangos estratigráficos de palinomorfos del Paleógeno de Argentina, Antártica y Chile. Simposio Paleógeno de América del Sur, Actas. Servicio Geológico Minero Argentino, Anales 33: 89-103. [ Links ]
61. Sánchez, M.V., Laza, J., Bellosi, E.S. y Genise, J.F. 2008. Icnoestratigrafía y características de trazas fósiles de escarabajos coprófagos (Coprinisphaera ispp.) de la Formación Sarmiento (Cenozoico medio): aparición y evolución de ecosistemas de pastizales abiertos. 12ª Reunión Argentina de Sedimentología (Buenos Aires), Resúmenes p. 162. [ Links ] .
62. Seward, A.C. 1919. Fossil plants. New York, Cambridge 4, 514 pp. [ Links ]
63. Shani, B. 1920. On certain archaid features in the seed of Taxus baccata, with remarks on yhe antiquity of the Taxineae. Annals of Botany 34: 117-132. [ Links ]
64. Sharma, B.D. 1970. Taxaceoxylon cupressoides sp. nov. from Dhokoti in the Rajmahal Hills, India. Ameghiniana 3: 257-278. [ Links ]
65. Shimakura, M. 1936. On the fossil wood of Torreya nucifera Sieb. et Zucc. from the Pleistocene of Kanagawaken. Journal of the Geological Society of Japan 612: 297-303. [ Links ]
66. Tortorelli, L.A. 1956. Maderas y bosques Argentinos. Ed. ACME, S.A.C.I. Buenos Aires, 910 pp. [ Links ]
67. Wilf, P., Johnson, K.R., Cúneo, N.R., Smith, M.E., Singer B.S. y Gandolfo. M.A. 2005. Eocene plant diversity at Laguna del Hunco and Río Pichileufú, Patagonia, Argentina. American Naturalist 165: 634-650. [ Links ]
68. Zucol, A.F. 2005. Estudios paleoagrostológicos en el Cenozoico de Patagonia, su importancia en el conocimiento de la historia evolutiva de los paleopastizales. 16° Congreso Geológico Argentino (La Plata), 4: 403-408. [ Links ]
69. Zucol, A.F y Brea. M. 2005. El registro fitolítico de los sedimentos cenozoicos de la localidad de Gran Barranca: su aporte a la reconstrucción paleoecológica. 16° Congreso Geológico Argentino (La Plata), 4: 395-402. [ Links ]
70. Zucol, A.F., Brea, M., Raigemborn, M.S y Matheos, S.D. 2004. Asociación fitolítica en las formaciones Las Flores, Koluél Kaike y Sarmiento en Gran Barranca, lago Colhué Huapi (Chubut, Argentina). Ameghiniana Suplemento Resúmenes 41: 25R-26R. [ Links ]
Recibido: 31 de marzo de 2008.
Aceptado: 1 de octubre de 2008.

