










Serviços Personalizados
Journal

Artigo
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAmeghiniana
versão On-line ISSN 1851-8044
Ameghiniana vol.46 no.1 Buenos Aires jan./mar. 2009
ARTÍCULOS ORIGINALES
Un nuevo Oldfieldthomasiidae (Mammalia: Notoungulata) del Paleógeno de la provincia de Salta, Argentina
Daniel A. García López1,2 y Jaime E. Powell1,2
1Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán. Miguel Lillo 205, 4000 San Miguel de Tucumán, Tucumán, Argentina.
2Consejo Nacional de Investigaciones Científicas y Técnicas. dgarcialopez@csnat.unt.edu.ar, jpowell@csnat.unt.edu.ar
Resumen. Se da a conocer un nuevo representante de la familia Oldfieldthomasiidae del Noroeste Argentino, Dolichostylodon saltensis gen. et sp. nov. Sus restos provienen de afloramientos de la Formación Lumbrera, situados por encima la "Faja Verde II", en El Simbolar, Departamento Guachipas, provincia de Salta. Dolichostylodon saltensis está definido por una serie de caracteres tales como: dentición superior decidua y permanente similar a Ultrapithecus rutilans Ameghino, 1901, pero dP más triangulares, con paracono más prominente, sin foseta mesiolabial y metalofo del dP3 bien desarrollado. Molares superiores con parastilo y metas-tilo prominentes. Columna del metacono más amplia que en Colbertia lumbrerense Bond, 1981. Foseta mesio-labial ausente, foseta distolabial relativamente amplia y somera. Crochet bien desarrollado y curvado, rodeando a la foseta distolabial. Cíngulo distal amplio. Hipocono pequeño en relación al protocono. Surco labial poco marcado. Molares inferiores con entocónido expandido y poco diferenciado del hipoconúlido, y con un surco reducido entre ambos. Valle del talónido más profundo que en C. lumbrerense. Este hallazgo constituye la segunda especie de Oldfieldthomasiidae para esta formación y la tercera para el Noroeste Argentino. Otros restos asignados a Oldfieldthomasiidae han sido reportados para Antofagasta de la Sierra pero fueron referidos al género Colbertia Paula Couto, 1952. La especie muestra afinidades morfológicas con Ultrapithecus ruti-lans, del "Barranquense" (Casamayorense) de Patagonia. Dolichostylodon saltensis parece tener mayor relación con las formas patagónicas que con las especies extrapatagónicas de Oldfieldthomasiidae, a pesar de su grado de braquiodoncia relativamente mayor.
Abstract. A new Oldfieldthomasiidae (Mammalia, Notoungulata) from the Paleogene of Salta Province, Argentina. A new representative of the family Oldfieldthomasiidae from Northwestern Argentina, Doli-chostylodon saltensis gen. et sp. nov., is described. The material was found in the Lumbrera Formation, above the "Faja Verde II", at El Simbolar, Guachipas Department, Salta Province, Argentina. Dolichostylodon salten-sis is defined by the following features: deciduous and permanent upper teeth similar to Ultrapithecus rutilans Ameghino, 1901, but with more triangular dP, more prominent paracone, without mesiolabial fossette and with dP3 presenting well developed metaloph. Upper molars with prominent parastyle and metastile. Column of metacone wider than in Colbertia lumbrerense Bond, 1981. Mesiolabial fossette absent, distolabial fossette relatively wide and shallow. Well developed and curved crochet, surrounding the distolabial fosset-te. Wide distal cingulum. Hypocone reduced in relation to the protocone. Labial sulcus not very pronounced. Lower molars with expanded entoconid, located close to the hypoconulid and presenting a reduced sulcus between both. Talonid valley deeper than in C. lumbrerense. This discovery constitutes the second species of Oldfieldthomasiidae for this Formation and the third for Northwest Argentina. Other materials assigned to Oldfieldthomasiidae have been reported from Antofagasta de la Sierra (Catamarca Province) but they were referred to the genus Colbertia Paula Couto, 1952. This species shows morphological similarities with Ultrapithecus rutilans, of the "Barrancan" (Casamayoran) of Patagonia. Dolichostylodon saltensis seems to have closer relationship with the patagonic forms than with the non-Patagonian species of Oldfieldthomasiidae, despite its brachydont teeth.
Key words. Notoungulata; Oldfieldthomasiidae; Paleogene; Argentina.
Palabras clave. Notoungulata; Oldfieldthomasiidae; Paleógeno; Argentina
Introducción
La familia Oldfieldthomasiidae constituye un grupo de notoungulados paleógenos de pequeña a mediana talla registrados probablemente desde el Tiu-pampense, si bien este registro de la localidad boliviana de Tiupampa está basado en un único molar aislado cuya identificación es dudosa (Muizón, 1991; López, 1995). Según otros criterios el registro más antiguo de la familia correspondería al Itaboraiense (López, 1995) o al Riochiquense (Bond, 1981). Por otra parte, los restos considerados como más tardíos fueron reportados para el "Divisaderense" (Montalvo y Bond, 1998). El grupo comprende formas consideradas como ramoneadoras, pero con cierta tendencia al aumento de la altura de las coronas dentarias.
Esta familia es basal en el Suborden Typotheria y representa una radiación temprana de los Notoungulata (Cifelli, 1993). En el citado trabajo también se considera que la familia es muy probablemente parafilética o polifilética.
érica del Sur, Tiupampa en Bolivia (si bien la asignación no es precisa, como se señaló anteriormente) y San José de Itaboraí, Río de Janeiro, en Brasil. Hasta el momento, la mayor diversidad está documentada en Patagonia, donde se registraron géneros tales como Maxschlosseria Ameghino, 1901 y Ultrapithecus Ameghino, 1901 (Simpson, 1967). En afloramientos extrapatagónicos, la familia es parte importante de la fauna de Divisadero Largo, en la Provincia de Mendoza (Simpson et al., 1962; Bond, 1981; López, 1995) sobre la cual se estableció la "Edad Mamífero Divisaderense", cuya identidad ha sido puesta en duda recientemente (López y Manassero, 2006).
En Brasil se ha registrado el género Colbertia Paula Couto, 1952, con la especie Colbertia magellanica (Price y Paula Couto, 1950). La inclusión en la familia de Itaboraitherium atavum (Paula Couto, 1954) ha sido puesta en duda (Bond, 1981).
En el noroeste argentino, la familia se encontraba representada hasta el momento por dos taxones: Colbertia lumbrerense Bond, 1981, que proviene de la Estancia Pampa Grande en la provincia de Salta, donde afloran sedimentos de la Formación Lumbrera (Casamayorense) y Suniodon catamarcensis López, 1995, que ha sido registrado en afloramientos de la Formación Geste (Mustersense) de Antofagasta de la Sierra, provincia de Catamarca. Además, para la Formación Geste se ha referido un molar aislado a Colbertia (aunque con dudas) y dos fragmentos mandibulares fueron asignados tentativamente a Oldfieldthomasiidae (López, 1995).
En este trabajo se da a conocer un nuevo representante de la familia reconocido en base al análisis de la morfología craneodentaria. Además, se compara este taxón con los Oldfieldthomasiidae ya conocidos y se discuten sus relaciones, así como algunos aspectos de su proveniencia estratigráfica.
El material fue exhumado de afloramientos de la Formación Lumbrera, por arriba de la "Faja Verde II", en El Simbolar, Departamento Guachipas, provincia de Salta. Estos niveles han sido identificados por algunos autores como "Lumbrera Superior" (Gómez Omil et al., 1989; del Papa, 2006). Este hallazgo constituye el segundo representante de Oldfieldthomasiidae para esta formación.
Materiales y métodos
El material corresponde a elementos craneales separados en cuatro fragmentos principales (cráneo, basicráneo, mandíbula incompleta y fragmento de hemimandíbula derecha) correspondientes a un mismo individuo juvenil. Las medidas dentarias del ejemplar se muestran en las tablas 1 y 2,
Tabla 1. Dolichostylodon saltensis gen. et sp. nov. Medidas (en mm) de la dentición superior / measurements (in mm) of the upper dentition. L, longitud mesiodistal / mesiodistal length. A, ancho / width. I, izquierdo / left. D, derecho / right. * Aproximación / approximation. ? Parcialmente emergido / partially erupted
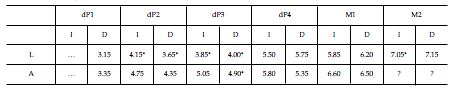
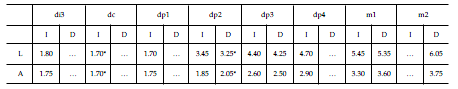
Tabla 2. Dolichostylodon saltensis gen. et sp. nov. Medidas (en mm) de la dentición inferior / measurements (in mm) of the lower dentition. L, longitud mesiodistal / mesiodistal length. A, ancho / width. I, izquierdo / left. D, derecho / right. * AAproximación / approximation.
El estudio del ejemplar se llevó a cabo analizando principalmente la morfología craneal superficial y las características dentarias. Se comparó el espécimen con representantes de diferentes familias de Notoungulata basales de varias zonas del país contenidos en la Colección Paleontología Vertebrados Lillo del Instituto Miguel Lillo (Tucumán, Argentina), la Colección de Paleontología de Vertebrados del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" (Ciudad Autónoma de Buenos Aires, Argentina) y la Colección de Paleontología de Vertebrados del Museo de La Plata (La Plata, Argentina). En este último museo se accedió también a calcos de materiales depositados en colecciones del extranjero. Además los restos se compararon con datos bibliográficos de varios autores. El material presentado en este trabajo está depositado en la Colección de Paleontología de Vertebrados Lillo del Instituto Miguel Lillo, Tucumán, Argentina.
Paleontología sistemática
Orden Notoungulata Roth, 1903
Suborden Typotheria Zittel, 1893
Familia Oldfieldthomasiidae Simpson, 1945
Dolichostylodon gen. nov. Figuras 1-3

Figura 1. PVL 6219, Dolichostylodon saltensis gen. et sp. nov. 1, vista dorsal del cráneo / skull in dorsal view; 2, vista ventral del cráneo / skull in ventral view; 3, diagrama de los dP1-M2 derechos / diagram of right dP1-M2. Escala / scale = 1 cm.
Especie tipo. Dolichostylodon saltensis sp. nov.
Derivatio nominis. "Dolichos" del griego: largo, alargado; "stylos" del griego: pilar, columna y "odontos" del griego: diente.
Diagnosis. La misma de la especie por monotipia / as for the species due to monotypy.
Dolichostylodon saltensis sp. nov. Figuras 1-3
Holotipo. PVL 6219. Cráneo incompleto con dP2-4 y M1-2 izquierdos y dP1-4 y M1-2 derechos (figura 1); fragmento de basicráneo (figura 2); mandíbula incompleta con di3, raíz de dc, dp1-4 y m1 izquierdos y dp2-3 derechos (figura 3.1,2); fragmento de hemimandíbula derecha con m1-2 (figura 3.3) y fragmentos aislados indeterminables.
Derivatio nominis. De la provincia de Salta, República Argentina y "ensis" sufijo latino que significa "de" o "perteneciente a".
Procedencia geográfica y estratigráfica. 25° 43.564' S, 65° 24.252' O, El Simbolar, Departamento Guachipas, provincia de Salta. Miembro superior de la Formación Lumbrera, por arriba de la "Faja Verde II", reconocido como Lumbrera Superior (del Papa, 2006; Gómez Omil et al., 1989).
Repositorio. Colección Paleontología Vertebrados Lillo, Instituto Miguel Lillo, San Miguel de Tucumán, Tucumán, Argentina.
Diagnosis. Dentición superior decidua y permanente similar a Ultrapithecus rutilans Ameghino, 1901, pero con dP más triangulares, con paracono más prominente, sin foseta mesiolabial y con dP3 presentando metalofo bien desarrollado. Molares superiores con parastilo y metastilo prominentes. Columna del metacono más amplia que en Colbertia lumbrerense. Foseta mesiolabial ausente, foseta distolabial relativamente amplia y somera. Crochet bien desarrollado y curvado rodeando a la foseta distolabial. Cíngulo distal amplio. Hipocono más pequeño en relación al protocono. Surco lingual poco marcado. Molares inferiores con entocónido expandido y situado más distalmente en relación al hipoconúlido que en C. lumbrerense. Valle del talónido más agudo y cerrado que en C. lumbrerense.
Diagnosis. Deciduous and permanent dentition similar to Ultrapithecus rutilans Ameghino, 1901, but with more triangular dP, paracono prominent, without mesiolabial fossette and with dP3 showing a well developed metalof. Upper molars with prominent parastyle and metastyle. Column of metacone wider than in Colbertia lumbrerense. Mesiolabial fossette absent, distolabial fossette relatively wide and shallow. Crochet well developed and curved surrounding the distolabial fossette. Wide distal cingulum. Reduced hypocone in proportion to the protocone. Lingual sulcus not well developed. Lower molars with expanded entononid located more distally to the hipoconulid than in C. lumbrerense. Valley of talonid sharper and more closed than in C. lumbrerense.
Descripción
Cráneo (figura 1)
El cráneo presenta el rostro corto. Las órbitas son relativamente grandes y la cresta sagital conspicua pero poco desarrollada. Estas características, conjuntamente con la alta proporción de dientes deciduos preservados, sugieren un estadio juvenil.
Los nasales son largos y se expanden en el extremo posterior. Su unión con el frontal forma una sutura recta a ambos lados del plano sagital que forma un ángulo abierto hacia atrás.
Los frontales tienen forma aproximadamente hexagonal. Se observan procesos postorbitales bien definidos y dirigidos laterodistalmente.
El foramen infraorbital es de contorno bilobulado (claramente visible en el lado derecho). Debido a la deformación, el foramen no se observa en el lado izquierdo.
La raíz anterior del arco zigomático está ligeramente expandida y se extiende desde el extremo mesial del dP4 hasta la parte media del M2. La morfología de la raíz anterior del arco se observa mejor del lado derecho. El maxilar compone la mayor parte de la raíz y en este elemento se destaca la zona de inserción del masetero. El yugal ocupa la parte dorsal del extremo anterior del arco y, aparentemente, llega hasta el lacrimal, aunque esto no puede observarse con claridad. El cuerpo del arco zigomático es delgado y no está muy expandido lateralmente.
La bóveda craneana se ha conservado parcialmente del lado izquierdo. Está presente también la parte dorsal anterior, donde se observa la cresta sagital, poco desarrollada.
Si bien la cresta lambdoidea no se ha conservado, habría sido prominente, a juzgar por el reborde óseo ubicado justo por detrás de la raíz posterior del arco zigomático. No pueden observarse las suturas en el paladar.
Basicráneo (figura 2)

Figura 2. PVL 6219 Dolichostylodon saltensis gen. et sp. nov. 1, basicráneo en vista ventral / ventral view of basicranium; 2, diagrama donde se muestran las partes que pudieron ser identificadas / diagram illustrating identifiable parts; 3, diagrama de la vista ventral del cráneo en un notoungulado generalizado. La parte coloreada en gris corresponde aproximadamente al fragmento de basicráneo conservado / diagram of the ventral view of the skull in a generalized notoungulate. The gray coloured part corresponds approximately to the preserved basicranium fragment. b, bulla timpánica / tympanic bulla; fra, foramen retroarticular / retroarticular foramen; mae, meato auditivo externo / external auditory meatus; p, petroso / petrosal; pra, proceso retroarticular / retroarticular process; rh, receso hioideo / hyoid recess. Escala / scale = 1 cm.
Se trata de un fragmento perteneciente al mismo individuo, separado durante la preparación. Comprende parte del basiesfenoides, de los pterigoides y de los escamosales (incluyendo secciones de las fosas glenoideas y los senos epitimpánicos), además de las bullas timpánicas y los petrosos. Se ha perdido toda la zona occipital y el material presenta un alto grado de deformación, por lo cual algunos elementos (particularmente los petrosos) seguramente no conservan su verdadera orientación.
La zona de los pterigoides y del basiesfenoides está particularmente deteriorada y no se ha podido identificar ningún elemento.
Las fosas glenoideas tienen una posición algo elevada respecto al poro acústico externo. Distalmente a las mismas se observa el proceso retroarticular (postglenoideo), en el cual se observa claramente el foramen retroarticular. En el lado derecho la deformación ha causado el cierre de este foramen.
En el extremo medial de la fosa glenoidea se observa un pequeño foramen rodeado por el borde de la bulla. Se trataría del foramen petrotimpánico (fisura glaseriana).
Distalmente a las estructuras ya mencionadas se observa el poro acústico externo. Es relativamente grande y está claramente definido. El meato acústico es muy corto y su dirección es posterolateral. La cresta meatal no está diferenciada. Inmediatamente adyacente al meato, en el lado distal, se encuentra el foramen estilomastoideo, el cual es muy pequeño.
En el borde posterior del poro acústico se observa parte de las bases de los procesos postimpánico y paraoccipital. Entre estos elementos y la bulla se encuentra el receso hioideo, que está muy desarrollado.
Como se expresó anteriormente, la zona del occipucio no se ha conservado. Las porciones mastoideas de los petrosos han quedado expuestas, pero dado que se desconocen varios elementos, no puede determinarse si los petrosos presentaban o no una exposición occipital en el cráneo completo.
Las bullas se encuentran muy deformadas y aplanadas. Aún así parecen haber sido prominentes.
El petroso derecho se encuentra completo. La cara cerebelar (que contiene la fosa subarcuata y el meato acústico interno) está orientada medialmente, enfrentada, por deformación, a la misma cara del petroso izquierdo. La fosa subarcuata está bien definida y es profunda. En el meato acústico interno se observan los foramenes que conducen los nervios facial y vestibulococlear. Estos forámenes están separados por un septo bien definido. El ventral, o sea el foramen vestibulococlear (salida del nervio del mismo nombre), es más grande que el dorsal. La cresta petrosa es llamativamente prominente, separa a la cara cerebelar de una sección orientada dorsalmente en la cual se distingue un surco y un pequeño foramen no identificado.
El petroso izquierdo presenta una rotura a nivel de la fosa subarcuata. En la sección resultante se observan pequeños orificios que podrían corresponder a cortes transversales de alguno de los canales semicirculares.
Mandíbula (figura 3)

Figura 3. PVL 6219 Dolichostylodon saltensis gen. et sp. nov. 1, mandíbula en vista dorsal / mandible in dorsal view; 2, mandíbula en vista lateral izquierda / mandible in left lateral view; 3, fragmento de hemimandíbula derecha con m1-2 / fragment of right hemimandible with m1-2; 4, diagrama de m1-2 derechos / diagram of right m1-2; 5, diagrama de dp1-4 izquierdos / diagram of left dp1-4. Escala / scale = 1 cm.
Las ramas mandibulares son delgadas y bajas. El ángulo entre la rama ascendente y la rama horizontal es levemente agudo.
No se ha preservado la zona de la sínfisis y la zona mesial de la rama horizontal está muy deteriorada, por lo que no pueden observarse los forámene mentonianos. El borde ventral de la mandíbula se curva dorsalmente justo por detrás de la fila de dientes, para continuar casi horizontal en la zona ventra de la rama ascendente. La fosa masetérica es poco profunda. La cresta coronoidea presenta un desarrollo moderado.
El cóndilo articular es globoso y ligerament transverso. Esta ubicado en una posición elevada bien por arriba de la fila de dientes. La cara media está más expandida dorso-ventralmente. La fenestr mandibular es bastante somera.
El proceso coronoides es delgado y ligerament curvado (cóncavo distalmente). Su altura ocupa aproximadamente un tercio de la altura total de la rama ascendente.
En la cara interna de la mandíbula se observa qu la fosa pterigoidea está poco desarrollada. El foramen mandibular está ubicado aproximadamente e el centro de la rama ascendente, un poco desplazad mesialmente. Desde este foramen parte mesialmente un surco horizontal amplio que parece extenderse hasta los alvéolos de los últimos molares. Evidentemente se trata de una deformación y no de una es tructura natural.
Dentición superior (figura 1)
Dientes braquiodontes. Presenta dP2-4 y M1-2 en e maxilar izquierdo y dP1-4 y M1-2 en el maxilar derecho.
Dentición superior decidua. Los premolares deciduos, a excepción del dP1, presentan parastilo bien desarrollado y prominente.
El dP1 es pequeño y de contorno subcircular. Se observa un parastilo definido pero más bajo que de los demás premolares deciduos y algo desplazad lingualmente. El protocono y el paracono son las cús pides más desarrolladas. El paracono es la cúspide más prominente en el ectolofo. El protocono es pequeño y bunoide. Sobre sus paredes mesial y distal se observan pequeños cíngulos de desarrollo casi simétrico. El cíngulo mesial se continúa con un reborde de esmalte que presenta una pequeña cúspide lingual a parastilo. Esta pequeña cúspide correspondería topo lógicamente a un paracónulo. El cíngulo distal se continúa en un reborde de esmalte de trayectoria curva (convexa distalmente) que se conecta con el borde distal del ectolofo (donde se observa el metastilo). La foseta central está ausente. El diente presenta la zona distal expandida en una pequeña cuenca somera carente de esmalte por detrás del protocono.
En el dP2, el parastilo está bien desarrollado. Est separado del paracono por un surco muy profundo El paracono presenta una columna amplia, por detrás el ectolofo presenta una leve convexidad pero no hay un metacono definido. El metastilo está poco desarrollado. El protocono es pequeño y bunoide. Se observa una pequeña foseta central; ésta presenta forma reniforme, con la concavidad orientada labialmente. No se observan indicios de foseta mesiolabial. Los cíngulos mesial y distal son pequeños formando sólo un leve reborde de esmalte. En este diente también está presente la faceta plana por detrás del protocono, pero en este caso está inclinada distalmente y su amplitud relativa es menor.
El dP3 presenta un parastilo muy desarrollado, ubicado en el borde lingual del diente y separado del paracono por un surco amplio y muy profundo sobre la pared labial. El paracono presenta una columna delgada y bien delimitada. Distalmente en el ectolofo se observa un metacono levemente insinuado. El metastilo es pequeño pero está claramente definido. El protocono es también pequeño. En este diente ya se observan protolofo y metalofo bien desarrollados pero delgados, de manera semejante a Colbertia y Brachystephanus Simpson, Minoprio y Patterson, 1962. El protolofo está alineado con el parastilo; el metalofo es transverso y está menos desarrollado que el protolofo. La fosa central está alineada en sentido oblicuo (distolingual - mesiolabial) y se observan pequeñas cristas en su borde labial. El cíngulo mesial está poco desarrollado, el distal es más prominente y está ubicado en una posición más elevada en la corona.
El dP4 presenta la morfología de un molar definitivo. El parastilo está bien desarrollado y proyectado mesiolabialmente. La columna del paracono esta bien marcada. Esta cúspide está separada del metacono por un surco amplio y profundo de la pared labial. El metacono es más amplio y menos definido que el paracono. El metastilo está proyectado, aunque menos que el parastilo, y se orienta en sentido distal, mostrando una ubicación más lingual en relación a la alineación de paracono y metacono. El protocono es amplio y bunoide. El hipocono es más pequeño. Ambas cúspides están separadas por una escotadura bien marcada en vista oclusal, pero no se observa un surco lingual bien marcado. La fosa central está orientada en sentido oblicuo (distolingual - mesiolabial) y es casi recta. Se observa un vestigio de una foseta distolabial pequeña y somera. El protolofo está bien desarrollado, se encuentra ligeramente curvado labialmente (cóncavo mesialmente). El metalofo es transverso y más corto que el protolofo. El crochet, casi recto, parte desde el metalofo y se une al ectolofo aproximadamente en el punto en el que el surco labial es más profundo. El cíngulo mesial está bien desarrollado, el distal es más pequeño.
Molares superiores definitivos. El M1 presenta muy poco desgaste. El parastilo es prominente, aunque es tá menos desarrollado que en el dP4 en vista oclusal Se orienta en sentido ánterolabial de manera similar a Ultrapithecus rutilans. Está separado del paracono por un surco muy profundo, que les da a ambas cúspides un aspecto columnar. El paracono es alto, presentando una columna muy prominente. Se observa un surco labial bien marcado entre paracono y metacono. El metacono está menos desarrollado que el paracono La columna del metacono presenta la particularidad de mostrar un reborde vertical estrecho que correspondería a la columna del metacono sensu stricto, la cual se asienta sobre una amplia convexidad del metalofo que ocupa toda la zona del metacono (a ambos lados de la columna estrecha mencionada anteriormente) y que está separada de la columna del paracono por un surco estrecho aunque no muy profundo El metastilo está proyectado distalmente y es prominente, sobrepasando el borde del cíngulo distal. El protocono es bunoide. El hipocono es marcadamente menor y su ápice es más labial que el anterior. Protocono e hipocono están separados por una escotadura pero el surco de la pared lingual es apenas notorio. La fosa central es profunda y se extiende oblicuamente (distolingual - mesiolabial), curvándose en sentido labial (convexa mesialmente). Su extremo distolingua es más amplio y redondeado. Se observa una foseta distolabial relativamente amplia pero somera. El protolofo es convexo hacia la cara mesiolingual del diente, el metalofo es mucho más corto y transverso. El crochet está bien desarrollado y contornea a la foseta disto labial. Los cíngulos mesial y distal son prominentes, particularmente el distal, que presenta una ubicación más elevada en la corona.
El M2 es muy similar al diente anterior. Se encuentra en erupción y no presenta desgaste. En este punto cabe destacar que, aún sin desgaste, la dentina está expuesta en la corona, principalmente sobre la pared lingual del ectolofo y el crochet (figura 1.2). El parastilo y particularmente el metastilo están relati vamente más proyectados. El surco labial entre para cono y metacono es también marcado. La foseta central es más profunda, al igual que la foseta distola bial. Debido a que el diente no presenta desgaste, se observa que el crochet presenta una pequeña cúspide bien diferenciada a mitad de su trayecto. El surco que separa al protocono del hipocono en la pared lingua es mucho más marcado que en el M1. El cíngulo distal está bien desarrollado.
Dentición inferior (figura 3)
El ejemplar presenta en la rama mandibular izquierda el di3, la raíz del dc, dp1-4 y m1 mientras que en la rama mandibular derecha conserva los dp2-3 y los m1-2 (estos últimos en un fragmento separado).
Dentición inferior decidua. El di3 no presenta desgaste y su forma es espatulada. El borde mesial es más delgado que el distal y presenta en su base, sobre la cara lingual, un cíngulo apenas insinuado. Hay una ligera concavidad lingual. La capa de esmalte que cubre la cara lingual del diente es aparentemente más delgada que en la cara labial.
La raíz del dc presenta una sección de contorno subcircular. Evidentemente este diente era más pequeño que el di3.
El dp1 es pequeño, unirradiculado y levemente comprimido labiolingualmente. Sólo se observa una cúspide bien desarrollada de la que parte una paracrístida proyectada mesialmente. Lingualmente puede apreciarse un pequeño cíngulo, expresado sólo como un ligero reborde de esmalte. Sobre la base del borde distal del diente se observa un talónido incipiente y muy pequeño.
El dp2 presenta ya la morfología típica de un premolar. Protocónido y metacónido están bien diferenciados y corresponden a la parte más alta del diente. A partir del protocónido parte una paracrístida recta y proyectada mesialmente. A partir del metacónido surge la crístida oblicua, que forma un ángulo en el hipocónido y llega hasta el entocónido, el cual es muy pequeño. El valle del talónido está poco desarrollado. El surco labial está bien marcado. En la zona mesiolingual se observa un cíngulo levemente marcado.
El dp3 es más molariforme y más alargado mesiodistalmente que el anterior. La paracrístida corre recta mesialmente, formando una cresta prominente. El metacónido es la cúspide más alta del diente y muestra una cresta distal de esmalte que cierra parcialmente el pequeño valle del talónido. El ápice del metacónido se conecta con una corta crístida oblicua que se curva bruscamente en el hipocónido, formando una "V" y se conecta transversalmente con el entocónido, que es relativamente prominente. Se observa un fuerte surco labial y un cíngulo mesiolingual más desarrollado que en el dp2.
El dp4 presenta la morfología de un molar definitivo. Se observa un trigónido muy corto, de aproximadamente la mitad de la longitud mesiodistal del talónido. Desde el protocónido surge el paralófido, de poco desarrollo y curvado lingualmente (rodeando el valle del trigónido). El metacónido es la cúspide más alta del diente, adosada a su borde mesial se observa una pequeña cúspide coalescente correspondiente al paracónido. El diente presenta un pequeño cíngulo mesiolingual. La crístida oblicua contacta la parte media del protolófido. El entocónido se conecta transversalmente con la crístida oblicua, sin embargo, no puede observarse su morfología debido a que la zona se encuentra muy deteriorada. El surco labial es profundo, está inclinado mesialmente y no llega hasta la base del diente. El valle del talónido es amplio y poco profundo.
Molares inferiores definitivos. El trigónido del m1 es corto mesiodistalmente pero está proporcionalmente más desarrollado respecto a lo observado en el dp4. El paralófido es muy corto y curvado lingualmente y no se conecta con el cíngulo mesiolingual, que es muy pequeño. El metacónido es prominente y presenta una pequeña cúspide coalescente (paracónido) en su pared mesial. Tanto en este diente como en el dp4 y el m2, esta última cúspide está algo desplazada lingualmente. La crístida oblicua contacta la mitad del protolófido y es arqueada. E hipocónido no está diferenciado y el hipoconúlido es prominente. El entocónido es bunoide y presenta su base expandida. Entre entocónido e hipoconúlido se observa una pequeña cresta de esmalte. El surco labial es profundo. El valle del talónido está bien desarrollado.
El m2 no presenta desgaste y, al igual que el M2 presenta dentina claramente expuesta tanto en e trigónido como en el talónido (figura 3.3). El trigónido es relativamente más grande respecto a los casos anteriores. El paralófido está bien desarrollado pero no se conecta con el cíngulo mesiolingual, que es pequeño. El metacónido está bien desarrollado y el paracónido es relativamente más prominente en este diente. La crístida oblicua forma una curva bien pronunciada hasta el pequeño hipoconúlido que se observa en la pared distal. El entocónido es bunoide y su base está expandida y forma parte del borde distal del diente. El surco entre el entocónido y el hipoconúlido está poco desarrollado. La cresta que se forma en la base de este surco está menos desarrollada que en el m1. El surco labial y particularmente el valle del talónido son muy profundos. Este último presenta paredes abruptas, que confluyen en e fondo, por lo que no forman una superficie horizontal.
Discusión
Sobre el estadío ontogenético del espécimen PVL 6219
La interpretación de este material como un individuo juvenil se realizó sobre la base de algunos caracteres craneales y dentarios. Los caracteres craneales incluyen órbitas relativamente grandes, hocico corto, caja craneana prominente y la cresta sagita poco desarrollada. Con respecto a los caracteres dentarios, las piezas anteriores muestran indicios que permiten establecer que se trata de dientes deciduos. La morfología del último premolar deciduo (tanto superior como inferior) suele ser muy similar a la del M1 (o m1), pero la altura de la corona es más baja (Simpson, 1948; 1967). Esto se observa claramente en los dientes identificados como dP4 y dp4. Además, todas las piezas anteriores hasta el último molar deciduo superior e inferior presentan una coloración más clara con respecto a las dos últimas piezas, lo que puede indicar diferencias en el espesor de la capa de esmalte. Esto se ha observado en otras series deciduas.
Otro aspecto destacable es el desgaste diferencial o relativo en cada diente. Este se considera uno de los criterios principales para identificar dientes deciduos (Hitz et al., 2006). La diferencia en el grado de desgaste se observa más claramente en las piezas dentarias superiores anteriores al M1. El dP1 es el menos desgastado de los dientes deciduos, mientras que los restantes muestran un desgaste muy conspicuo. Por su parte, los molares superiores permanentes casi no presentan desgaste (muy leve en el M1 y ninguno en el M2). Este patrón está relacionado con la secuencia de erupción de los primeros dientes (dP2, dP3, dP4, dP1, M1, M2).
Simpson (1948) realizó un estudio comparativo de tres cráneos de diferentes edades pertenecientes a la especie Notostylops murinus Ameghino, 1897. Este autor clasifica a los cráneos como A (juvenil), B (juvenil avanzado) y C (adulto). En el juvenil están presentes todos los dientes deciduos y ya ha emergido el M1, pero este último no presenta desgaste. En el juvenil avanzado están presentes todos los dientes de leche, el M1 (levemente desgastado) y el M2 (sin desgaste). Por último, en el adulto se han reemplazado los dientes deciduos, los M1 y 2 presentan desgaste y ya ha emergido el M3.
El holotipo de Dolichostylodon saltensis presenta las características correspondientes al estadío B, o sea, juvenil avanzado. Los rasgos craneales descriptos por Simpson (1948) coinciden también con lo observado en el PVL 6219 (e.g. ancho del hocico, tamaño relativo de las órbitas, etc.). Las únicas diferencias notables de D. saltensis con respecto al estadío B son el mayor desarrollo de la caja craneana y la cresta sagital menos prominente. Sin embargo, debe tenerse en cuenta que Notostylops murinus y D. saltensis son especies alejadas filogenéticamente y que estas diferencias morfológicas seguramente reflejan este factor.
En resumen, los caracteres craneales y dentarios del PVL 6219 indican que se trata de un juvenil avanzado, probablemente algo más inmaduro que el estadío B de Simpson (1948). En esta etapa de desarrollo el cráneo no ha alcanzado su tamaño definitivo. El cráneo del holotipo de Dolichostylodon saltensis es un poco más pequeño que el de los ejemplares adultos de Colbertia lumbrerense analizados (incluyendo al tipo). Por lo tanto, probablemente el cráneo de los adultos de D. saltensis haya sido ligeramente mayor a los de C. lumbrerense.
Comparación con otros representantes de Oldfieldthomasiidae
De la comparación de Dolichostylodon saltensis con las otras formas patagónicas y extrapatagónicas de Oldfielthomasiidae se desprenden varias observaciones.
Los premolares deciduos son, en conjunto, semejantes a los observados en ejemplares juveniles de Ultrapithecus rutilans. En todas las piezas (a excepción del dP1) se observa un parastilo bien desarrollado y prominente, el cual también se observa en la dentición decidua de U. rutilans.
Sin embargo, los premolares deciduos del nuevo taxón presentan claras diferencias con esta especie patagónica: menor altura coronal, forma más triangular de los dientes y ausencia de fosetas mesiolabiales.
Los molares definitivos superiores también denotan similitudes con Ultrapithecus tanto en la morfología general como en caracteres más puntuales (e.g. forma de la fosa central, el desarrollo del parastilo, forma y extensión del crochet, proporción entre protocono y metacono).
Por otra parte, se observan algunas diferencias entre el nuevo taxón y Colbertia lumbrerense, como el contorno más trapezoidal de las piezas dentarias, el parastilo más desarrollado (éste último carácter también constituye una diferencia con Suniodon catamarcensis), la morfología del surco labial entre las columnas del paracono y metacono (más definido y estrecho en Dolichostylodon saltensis) y la diferencia de tamaño entre protocono e hipocono (más acusada en el nuevo taxón).
Los molares inferiores definitivos muestran características claramente diferentes a las del género Colbertia, como una altura coronal aparentemente mayor y una mayor expansión del entocónido. Este último carácter y la reducción del surco entre entocónido e hipoconúlido representan similitudes con las formas "Divisaderenses" como Brachystephanus.
Sobre la posición de Dolichostylodon saltensis dentro de los Oldfieldthomasiidae
La familia Oldfieldthomasiidae ha sido dividida en dos "grupos morfológicos" (Bond, 1981; López, 1995; Montalvo y Bond, 1998). La división se basa en características dentarias como el grado de lofodoncia y de hipsodoncia (Bond, 1981; López, 1995). El primer grupo está representado por géneros exclusivamente patagónicos, entre los cuales se incluye a Kibenikhoria Simpson, 1935 del Itaboraiense (Montalvo y Bond, 1998), Oldfieldthomasia Ameghino, 1901 del Riochiquense (Reguero et al., 2002) al Barranquense (Casamayorense tardío), Ultrapithecus del Barranquense y Tsamnichoria del Mustersense. El segundo grupo está constituido por el género Colbertia (Itaboraiense-Casamayorense), las formas del "Divisaderense", Brachystephanus, Xenostephanus y Allalmeia Rusconi, 1946 y un género patagónico, Maxschlosseria, del Riochiquense (Reguero et al., 2002) al Vaquense (Casamayorense temprano), que guardan "semejanzas de tipo no sólo estructural, sino muy probablemente filogenéticas" (Bond, 1981). Recientemente López y Bond (2003) propusieron el reconocimiento de una nueva familia de Notoungulata, conformada por este segundo grupo morfológico (incluyendo sólo algunos representantes del género Maxschlosseria), sobre la base de caracteres dentarios y craneales.
En la provincia de Catamarca se registró a Suniodon catamarcensis, asignado a niveles del Mustersense, primera forma extrapatagónica perteneciente al primer grupo morfológico de Oldfieldthomasiidae (López, 1995). La asignación de S. catamarcensis a este grupo es plausible dadas sus semejanzas con el género Kibenikhoria del Itaboraiense y por su grado de hipsodoncia.
La dentición decidua y definitiva de Dolichostylodon saltensis es semejante a la de Ultrapithecus rutilans. Sin embargo, la altura relativa de las coronas dentarias es menor que la de los géneros patagónicos, pero mayor que la de Colbertia lumbrerense, proveniente de niveles más bajos de la Formación Lumbrera. Si se toman las características morfológicas como argumento suficiente, el nuevo taxón presentado en este trabajo sería un nuevo representante norteño del grupo patagónico. Sin embargo, se trataría del representante más braquiodonte, dado que la altura relativa de las coronas dentarias de D. saltensis (reflejada en el índice de hipsodoncia) es menor a la de S. catamarcensis (en el M2) y a la de los géneros patagónicos (en los M1 y M2). Usualmente el índice de hipsodoncia se calcula en molares superiores sin desgaste dividiendo la altura máxima del ectolofo en su longitud mesiodistal máxima (Croft et al., 2003; Hitz et al., 2006). En D. saltensis este índice tiene un valor de 0,68 en el M1 (con desgaste mínimo) y aproximadamente 0,70 para el M2 (parcialmente emergido y sin desgaste). En los ejemplares medidos de C. lumbrerense, el valor promedio del índice es de 0,61 para ambos molares, en todos los casos con poco desgaste. En S. catamarcensis el valor del índice es menor a D. saltensis para el M1 (0,61), pero debe tenerse en cuenta que este molar muestra un fuerte desgaste. En el M2 el índice es de 0,73, es decir que es mayor a D. saltensis, aún con fuerte desgaste. Por otra parte, en el ejemplar AMNH 28583 de U. rutilans el índice de hipsodoncia del M1 es igual a 0,90 con poco desgaste y en el ejemplar AMNH 28691 de Oldfieldthomasia debilitata el valor del índice es de 0,73 en el M1 y 0,82 en el M2, ambos escasamente desgastados.
La situación de Suniodon catamarcensis y Dolicho tylodon saltensis con respecto al resto de los Oldfield thomasiidae extrapatagónicos es llamativa. Los estudios realizados hasta el momento en el Noroeste, tanto en geología como en paleontología (de mamífero y otros vertebrados terrestres), indican que durant el Eoceno las condiciones climáticas eran más cálida y húmedas que las que se registraban en Patagoni contemporáneamente (Pascual et al., 1981; Pascual al., 1996; Goin et al., 1998; Ortiz Jaureguizar Cladera, 2006), lo cual explicaría el predominio de formas más ramoneadoras en los ambientes continentales extrapatagónicos, incluyendo la fauna d Divisadero Largo, en la provincia de Mendoza. Esto implicaría, en términos de edad, que fuera de Patagonia persistieron condiciones relativamente más cál das y húmedas hasta por lo menos el Eoceno tardí las cuales estaban dominadas por formas más braquiodontes correspondientes al segundo grupo morfológico de Oldfieldthomasiidae (Bond, 1981; Pascu et al., 1996). Es entonces interesante la presencia de representantes de Oldfieldthomasiidae emparenta dos a géneros comúnmente asociados a climas más fríos y secos. Las implicancias paleobiogeográficas de estas observaciones deben ser aún analizadas.
Comentarios sobre la edad de los niveles portadores
Los mamíferos provenientes de la Formación Lumbrera han sido asignados a la edad mamífero Casamayorense (Vucetich, 1980; Bond, 1981; Pascual et al., 1981). Convencionalmente, esta edad mamífero ha sido referida al Eoceno inferior por diferentes autores (Cifelli, 1985; Bond y López, 1993; López, 1995 Sin embargo, Kay et al. (1999), sobre la base de data ciones isotópicas (Ar/Ar), expandieron el lapso d esta edad desde el Eoceno temprano hasta el Eocen tardío. Las "subedades" Vaquense y Barranquens del Casamayorense, propuestas por Cifelli (1985 quedaron restringidas respectivamente a ambos extremos del Eoceno, separadas por un amplio laps entre ambas. Posteriormente, Bellosi y Madde (2005) obtuvieron para el Miembro Gran Barranca de la Formación Sarmiento una antigüedad Ar-Ar comprendida entre 42,1-38,3 millones de años, acotando considerablemente el lapso intermedio inferido por Kay et al. (1999) para las subedades del Casamayorense y ubicando al Barranquense en el Eoceno medio.
Colbertia lumbrerense proviene de niveles por de bajo de la "Faja Verde I". El género Colbertia se registra desde el Itaboraiense de Brasil (Paleoceno me dio) y ha sido relacionado al género Maxschlosseri (Riochiquense-Vaquense) de Patagonia. Por su parte, Dolichostylodon saltensis proviene de niveles ubicados por arriba de la "Faja Verde II", en asociación con el leontínido basal Coquenia bondi Deraco, Powell y López, 2008 y se relaciona morfológicamente con el género Ultrapithecus, del Barranquense de Patagonia.
Como resultado de la comparación entre Dolichostylodon saltensis y Colbertia lumbrerense se observan caracteres que permiten inferir un estado más derivado para el nuevo género y especie, como ser la mayor altura coronal, la reducción del número de fosetas y la reducción de cíngulos (Bond, 1981; Montalvo y Bond, 1998). Estas evidencias permiten inferir que los niveles portadores de Dolichostylodon saltensis pueden estar separados de los niveles que infrayacen a la "Faja Verde I" por un lapso considerable. Sin embargo, debido a ciertos caracteres de esta especie (como el bajo índice de hipsodoncia respecto a formas patagónicas y extrapatagónicas del Eoceno tardío), no puede afirmarse que pueda ser asignada a una edad Barranquense, aunque debe tenerse aún en cuenta lo expuesto acerca de la posible persistencia de condiciones más cálidas en el noroeste argentino hacia el Eoceno superior. Es probable, en definitiva, que los niveles correspondan a una edad anterior inmediata al Barranquense.
Conclusiones
Dolichostylodon saltensis representa el segundo Oldfieldthomasiidae registrado para la Formación Lumbrera y el tercero para todo el noroeste argentino. Otros restos asignados a Oldfieldthomasiidae han sido reportados para Antofagasta de la Sierra (López 1995) pero fueron referidos tentativamente al género Colbertia.
Este nuevo taxón muestra afinidades morfológicas con Ultrapithecus rutilans, del Barranquense (Casamayorense tardío) de Patagonia, por lo que se encuadra en el grupo morfológico de las formas en su mayoría patagónicas. A su vez presenta caracteres que lo diferencian marcadamente de Colbertia, otro taxón registrado en Formación Lumbrera.
Al menos en el caso de los Oldfieldthomasiidae, la evidencia morfológica sugiere que los niveles portadores de Colbertia lumbrerense y Dolichostylodon saltensis están separados por un lapso temporal relativamente amplio. Sin embargo, la evidencia es insuficiente para determinar la correlación con la subedad Barranquense reconocida en Patagonia.
Abreviaturas. AMNH, American Museum of Natural History, Nueva York, Estados Unidos; PVL, Colección Paleontología Vertebrados Lillo, Instituto Miguel Lillo, Tucumán, Argentina; dP, premolares deciduos superiores; M, molares permanentes superiores; di, incisivos deciduos inferiores; dc, caninos deciduos inferiores; dp, premolares deciduos inferiores; m, molares permanentes inferiores.
Agradecimientos
Los autores desean agradecer a A. Kramarz y M. Reguero por su autorización para analizar material a su cargo. G. López, P. Ortiz y J. Babot aportaron útiles comentarios y sugerencias acerca de la redacción y compaginación del trabajo. P. Ortiz también brindó valiosa ayuda para la obtención de las fotografías. D. Croft y J. Gelfo realizaron valiosas críticas y aportaron sugerencias sumamente útiles para la publicación final del manuscrito. Este trabajo fue solventado con fondos del Consejo Nacional de Investigaciones Científicas y Técnicas (PIP5255-CONICET), el Consejo de Investigaciones de la Universidad Nacional de Tucumán (G319-CIUNT) Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT-FONCyT) y la Fundación Miguel Lillo (Tucumán).
Bibliografía
1. Ameghino, F. 1897. Mammifères crétacés de l'Argentine. (Duxième contribution à la connaissance de la faune mammalogique des couches à Pyrotherium). Boletín del Instituto Geográfico Argentino 18: 406-429, 431-521. [ Links ]
2. Ameghino, F. 1901. Notices préliminaires sur des ongulés nouve-aux des terrains Crétacés de Patagonie. Boletín de la Academia Nacional de Ciencias (Córdoba) 16: 349-426. [ Links ]
3. Bellosi, E.S. y Madden, R.H. 2005. Estratigrafía física preliminar de las secuencias piroclásticas terrestres de la Formación Sarmiento (Eoceno-Mioceno) en la Gran Barranca, Chubut. 16° Congreso Geológico Argentino (La Plata, 2005), Actas 4: 427-432. [ Links ]
4. Bond, M. 1981. Un nuevo Oldfieldthomasiidae (Mammalia, Noto-ungulata) del Eoceno Inferior (Fm. Lumbrera, Grupo Salta) del NW argentino. 2° Congreso Latinoamericano de Paleontología (Porto Alegre), Anais 2: 521-536. [ Links ]
5. Bond, M. y López, G. 1993. El primer Notohippidae (Mammalia, Notoungulata) de la Formación Lumbrera (Grupo Salta) del Noroeste Argentino. Consideraciones sobre la sistemática de la Familia Notohippidae. Ameghiniana 30: 59-68. [ Links ]
6. Cifelli, R.L. 1985. Biostratigraphy of the Casamayoran, Early Eocene, of Patagonia. American Museum Novitates 2820: 1-26. [ Links ]
7. Cifelli, R.L. 1993. The Phylogeny of the Native South American Ungulates. En: F.S. Szalay, M.J. Novacek y M.C. McKenna (eds.). Mammal Phylogeny, vol 2: Placentals. Springer-Verlag, New York, pp. 195-216. [ Links ]
8. Croft, D.A., Bond, M., Flynn, J.J., Reguero, M. y Wyss, A.R. 2003. Large Archaeohyracids (Typotheria, Notoungulata) from Central Chile and Patagonia, including a revision of Archaeotypotherium. Fieldiana: Geology, New Series 49: 1-38. [ Links ]
9. del Papa, C.E. 2006. Estratigrafía y paleoambientes de la Formación Lumbrera, Grupo Salta, Noroeste Argentino. Revista de la Asociación Geológica Argentina 61: 313-327. [ Links ]
10. Deraco, M.V., Powell, J.E. y López, G. 2008. Primer leontínido (Mammalia, Notoungulata) de la Formación Lumbrera (Subgrupo Santa Bárbara, Grupo Salta-Paleógeno) del noroeste argentino. Ameghiniana 45: 83-91. [ Links ]
11. Goin, F.J., Candela, A. y López, G. 1998. Middle Eocene marsupials from Antofagasta de la Sierra, Northwestern Argentina. Geobios 31: 75-85. [ Links ]
12. Gómez Omil, R.J., Boll, A. y Hernández, R.M. 1989. Cuenca Cretácico-Terciaria del Noroeste Argentino (Grupo Salta). En: G.A. Chebli y L.A. Spalletti (eds.). Cuencas Sedimentarias Argentinas. Universidad Nacional de Tucumán, Serie de Correlación Geológica 6: 43-64. [ Links ]
13. Hitz, R.B., Flynn, J.J. y Wyss, A.R. 2006. New basal Interatheriidae (Typotheria, Notoungulata, Mammalia) from the Paleogene of Central Chile. American Museum Novitates 3520: 1-32. [ Links ]
14. Kay, R.F., Madden, R.H., Vucetich, M.G., Carlini, A.A., Mazzoni, M.M., Re, G.H., Heizler, M. y Sandeman, H. 1999. Revised geochronology of the Casamayoran South American Land Mammal Age: Climatic and biotic implications. Proceedings of the National Academy of Sciences 96: 13235-13240. [ Links ]
15. López, G.M. 1995. Suniodon catamarcensis gen. et sp. nov. y otros Oldfieldthomasiidae (Notoungulata, Typotheria) del Eoceno de Antofagasta de la Sierra, Catamarca, Argentina. 4° Congreso Argentino de Paleontología y Bioestratigrafía (Trelew), Actas: 167-172. [ Links ]
16. López, G. y Bond, M. 2003. Una nueva familia de ungulados (Mammalia, Notoungulata) del Paleógeno sudamericano. Ameghiniana Suplemento Resúmenes 40: 60R. [ Links ]
17. López, G. y Manassero, M. 2006. Determinación de la procedencia estratigráfica de Ethegotherium carettei Minoprio, 1947 (Notoungulata, Hegetotheriidae) a partir de un análisis de petrografía sedimentaria. Ameghiniana Suplemento Resúmenes 43: 44R-45R. [ Links ]
18. Montalvo, C.I., y Bond, M. 1998. Un Notoungulata de la Formación Vaca Mahuida (Eoceno) de la Provincia de La Pampa. Asociación Paleontológica Argentina. Publicación Especial 5. Paleógeno de América del Sur y de la Península Antártica: 55-60. [ Links ]
19. Muizón, C. de. 1991. La Fauna de Mamíferos de Tiupampa (Paleoceno inferior, Formación Santa Lucía), Bolivia. En: R. Suárez-Soruco (ed.) Fósiles y facies de Bolivia. I Vertebrados, Revista Técnica YPFB 12: 575-624. [ Links ]
20. Ortiz Jaureguizar, E. y Cladera, G.A. 2006. Paleoenvironmental evolution of suthern South America during the Cenozoic. Journal of Arid Environments 66: 498-532. [ Links ]
21. Pascual, R., Bond, M. y Vucetich, M.G. 1981. El Subgrupo Santa Bárbara (Grupo Salta) y sus vertebrados, cronología, paleoambientes y paleobiogeografía. 8° Congreso Geológico Argentino (San Luis), Actas 3: 743-758. [ Links ]
22. Pascual, R., Ortiz Jaureguizar, E. y Prado, J.L. 1996. Land Mammals: Paradigm for Cenozoic South American Geobiotic Evolution. Münchner Geowissenschaftliche Abhandlungen (A) 30: 265-319. [ Links ]
23. Paula Couto, C. de. 1952. Fossil Mammals of the Beginning of the Cenozoic in Brazil. Notoungulata. American Museum Novitates 1568:1-16. [ Links ]
24. Paula Couto, C. de. 1954. On a notostylopid from the Paleocene of Itaboraí, Brazil. American Museum Novitates 3: 77-86. [ Links ]
25. Price, Ll. I. y Paula Couto, C. de. 1950. Vertebrados terrestres do Eoceno na bacia calcárea de Itaborai. 2° Congresso Panamericano de Engenharia de Minas e Geología (Río de Janeiro), Anais 3: 149-173. [ Links ]
26. Reguero, A.M., Marenssi, S.A. y Santillana, S.N. 2002. Antarctic Peninsula and South America (Patagonia) Paleogene terrestrial faunas and environments: biogeographic relationships. Paleogeography, Paleoclimatology, Paleoecology 179: 189-210. [ Links ]
27. Rusconi, C. 1946. Nuevo mamífero fósil de Mendoza. Boletín Paleontológico de Buenos Aires 20: 1-2. [ Links ]
28. Simpson, G.G. 1935. Descriptions of the oldest known South American mammals, from the Río Chico formation. American Museum Novitates 793: 1-25. [ Links ]
29. Simpson, G.G. 1948. The Beginning of the Age of Mammals in South América. Part 1. Introduction. Systematics: Marsupialia, Edentata, Condylarthra, Litopterna and Notioprogonia. Bulletin of the American Museum of Natural History 91: 1-232. [ Links ]
30. Simpson, G.G. 1967. The Beginning of the Age of Mammals in South América. Part 2. Systematics: Notoungulata, concluded (Typotheria; Hegetotheria, Toxodonta, Notoungulata insertae-sedis); Astrapotheria Trigonostylopoidea; Pyrotheria; Xenungulata; Mammalia insertaesedis. Bulletin of the American Museum of Natural History 137: 1-259. [ Links ]
31. Simpson, G.G., Minoprio, J.L. y Patterson, B. 1962. The mammalian fauna of the Divisadero Largo Formation, Mendoza, Argentina. Bulletin of the Museum of Comparative Zoology at Harvard College 127: 237-303 [ Links ]
32. Vucetich, M.G. 1980. Un nuevo Notostylopidae (Mammalia, Notoungulata) proveniente de la Formación Lumbrera (Grupo Salta) del Noroeste Argentino. Ameghiniana 17: 363-372. [ Links ]
Recibido: 10 de marzo 2008.
Aceptado: 15 de octubre de 2008.

