










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana vol.46 no.1 Buenos Aires ene./mar. 2009
ARTÍCULOS ORIGINALES
Anterior caudal transverse processes in sauropod dinosaurs: morphological, phylogenetic and functional aspects
Pablo Ariel Gallina1 and Alejandro Otero2
1Área de Paleontología, Fundación de Historia Natural Félix de Azara, CEBBAD, Universidad Maimónides, Valentín Virasoro 732, 1405 Ciudad Autónoma de Buenos Aires, Argentina. pablogallina@gmail.com
2Museo de Geología y Paleontología (Universidad Nacional del Comahue) INIBIOMA-CONICET, Buenos Aires 1400, 8300 Neuquén, Argentina. alexandros.otero@gmail.com
Abstract. A great amount of the sauropod record is based on caudal vertebrae. Morphological analyses of tail bones until recently were essentially focused on centrum shape, with other anatomical features being poorly studied. A detailed description of anterior caudal transverse processes (ACTP) is presented here in order to improve the scarce knowledge of this tail part. Within Sauropoda there are four different ACTP morphological types; one simple (single element projected on the vertebral side) and three complex (lateral bony laminar plates supported by bony bars). The identity of the ACTP elements includes a rib, a synapophysis and laminar components, which are renamed here. The ventral laminae include the anterior centroparapophyseal lamina (acpl) and the posterior centrodiapophyseal lamina (pcdl), whereas the dorsal ones are the prezygodiapophyseal lamina (prdl) and the postzygodiapophyseal lamina (podl) when present. ACTP morphology is not informative for higher-level phylogeny, but at lower levels the in-group relationships of Diplodocoidea can be improved through analyzing these structures. ACTP morphological variation between the first and consecutive caudal vertebrae should presumably be interpreted as a consequence of the relative development of the M. caudofemoralis brevis. The "sacralization" of the ACTP is related to the attachment of soft tissues associated with the tail, among which the Mm. caudofemorales should be the most important muscle group.
Resumen. Procesos Transversos De Vértebras Caudales En Dinosaurios Saurópodos: Aspectos Morfológicos, Filogenéticos y Funcionales. Gran parte del registro de dinosaurios saurópodos está basado en vértebras caudales. Los análisis morfológicos que incluyen a estos huesos de la cola se basan en la forma del cuerpo, dejando de lado otras características anatómicas. Se realizó una descripción detallada de los procesos transversos de las caudales anteriores (ACTP) con el objeto de ampliar el poco conocimiento que se tiene de esta parte de la cola. Entre los saurópodos se reconocen cuatro tipos de ACTP; uno simple (elemento único proyectado sobre el lado vertebral) y tres complejos (placas de hueso laminares laterales soportadas por barras óseas). La identidad de los elementos de los ACTP incluyen un componente costal, una sinapófisis y componentes laminares; por lo tanto las láminas de los ACTP son renominadas. Las láminas ventrales incluyen a la lámina centroparapofisial anterior (acpl) y a la lámina centrodiapofisial posterior (pcdl), mientras que las dorsales son la lámina prezigodiapofisial (prdl) y la lámina postzigodiapofisial (podl) cuando está presente. El análisis de la morfología de los ACTP no es informativo en la filogenia de grupos mayores, pero dentro de los grupos puede ser útil, por ejemplo en los Diplodocoidea. La variación morfológica de los ACTP entre las primeras caudales y las sucesivas puede ser interpretada como la consecuencia del desarrollo relativo del M. caudofemoralis brevis. La "sacralización" de los ACTP se relaciona con el sitio de origen de los tejidos blandos asociados de la cola. En este sentido los Mm. caudofemorales serían el grupo muscular mas importante.
Key words. Sauropoda; Caudal anatomy; Transverse processes; Laminae; Soft tissues.
Palabras claves. Sauropoda; Anatomía caudal; Procesos transversos; Láminas; Tejidos blandos.
Introduction
Sauropod vertebrae are one of the most useful skeletal elements for systematic identification (McIntosh, 1990; Bonaparte, 1999). The complex morphology of these bones underpins this significance, as portrayed by Bonaparte (1999) and Wilson (1999). Broadly speaking, presacral elements are the most informative due to the presence of identifiable and comparable elements on neural arch and centrum (i.e. neural spine, laminae, zygapophyses, diapophyses, parapophyses). Nevertheless, a great amount of the sauropod record is only based on caudal vertebrae (McIntosh, 1990; Upchurch et al., 2004).
Morphological analyses of tail bones have essentially focused on centrum shape (Wilson et al., 1999; Salgado and García, 2002), with the arches being poorly studied.
A large variation in form is observed in sauropod caudal vertebrae (McIntosh, 1990). A good example of this is the morphological variation observed on transverse processes (Gallina, 2006). Although several authors have suggested the presence of different components in the anterior caudal transverse processes (Hatcher, 1903; Calvo and Salgado, 1995; Upchurch et al., 2004; Salgado et al., 2006a), and hence a morphological diversity, no comprehensive description clarifies this subject and its implications. Moreover, no studies have analyzed if this variability is the result of contributions from different constituent elements, or from different development of ubiquitous elements. Likewise, the poor knowledge of this tail part has led to nomenclatural ambiguities; thus a review of transverse process nomenclature is necessary (see Nomenclatural aspects in Methods below). The aim of this contribution is to describe and analyze sauropod anterior caudal transverse process (hereafter = ACTP) morphological variation and to ascertain the identity of their constituent elements. The results will provide information about the taxonomic importance of ACTP morphology, as well as its relationships with caudal musculature and functional aspects.
Materials and methods
Examined specimens include Patagosaurus fariasi Bonaparte, 1979 (PVL-4170); Limaysaurus tessonei (Calvo and Salgado, 1995) (MUCPv-205); Cathartesaura anaerobica Gallina and Apesteguía, 2005 (MPCA- 232); Diplodocus carnegiei Hatcher, 1901 (cast housed in Museo de La Plata; Osborn, 1899); Andesaurus delgadoi Calvo and Bonaparte, 1991 (MUCPv-132); Aeolosaurus rionegrinus Powell, 1987 (MJG-R-1); Bonitasaura salgadoi Apesteguía, 2004 (MPCA-460); Rinconsaurus caudamirus Calvo and González Riga, 2003 (MRS-Pv26); Magyarosaurus dacus (Nopcsa, 1915) (MAFI-U-10344); Saltasaurus loricatus Bonaparte and Powell, 1980 (PVL-4017-92); Neuquensaurus australis (Lydekker, 1893) (MLP-Ly1/2/3/4/5/6). The other specimens analyzed in this work were examined from literature and/or photographies.
Two specimens of Caiman latirostris were dissected (one subadult and one juvenile) in order to identify the site of attachment of the caudal musculature in crocodilians as an exemplar for extant archosaurs.
The description will focus on discriminating simple (single elements projected on the vertebral side) or complex (lateral bony laminae supported by bony bars) ACTP, their lateral extent and dorso-ventral position on the side of the vertebra. That is, ACTPs structural types (i.e. various combinations of primary homology assessments), not phylogenetic categories.
When laminar ACTPs are described, we use the term "Fan-like" for transverse processes with narrow bases that are distally expanded (base length approximately½ distal length, figure 1.2), and "Wing-like" for transverse processes with wide bases, that expand distally (base length more than ½ distal length, figure 1.3). We will specify the number of the caudal segment along the caudal series when possible. If not, we will refer to them as "anterior caudals".

Figure 1. Lateral elements of sauropod vertebrae in anterior view. 1, dorsal vertebra 9 of Apatosaurus (modified from Gilmore, 1936); 2, anterior caudal vertebra of Omeisaurus (modified from Young, 1937); 3, anterior caudal vertebra of Barosaurus (modified from Lull, 1919). di, diapophysis; pa, parapophysis; tp, transverse processes. Reconstructed areas in grey. Scale bar: 10 cm / Elementos laterales de las vértebras de los saurópodos en vista anterior. 1, Vértebra dorsal 9 de Apatosaurus (modificado de Gilmore, 1936); 2, Vértebra caudal anterior de Omeisaurus (modificado de Young, 1937); 3, Vértebra caudal anterior de Barosaurus (modificado de Lull, 1919). di, diapófisis; pa, parapófisis; tp, proceso transverso. Áreas reconstruidas en gris. Escala gráfica: 10 cm.
For muscular comparisons we follow the traditional axial musculature subdivisions: dorsal -epaxial- and ventral -hypaxial- (Romer, 1923; Vanden Berge, 1982). Homology hypotheses are made by means of comparisons with extant archosaurs (crocodiles and birds, see Witmer, 1995, 1997).
Nomenclatural aspects. In vertebrates, the term transverse processes has been used for laterally projected structures on the neural arches (i.e. diapophyses) or the centra (i.e. parapophyses; basiapophyses; pleurapophyses) in presacral vertebrae (Romer, 1956, 1962; Gavrilov, 1959; Weichert, 1959; Grassé, 1975; Hildebrand, 1988; Kardong, 1999). In sacral and caudal vertebrae, the union of different elements of the neural arch, centrum and ribs also falls under this generalized term. As a consequence, structures of similar position but different compositional origin have been covered by the same terminology resulting in nomenclatorial confusion (Romer, 1956, p. 225).
The only vertebrates that have a standardized anatomical nomenclature are humans, some domesticated mammals and birds (NA, Nomina Anatomica; NAV, Nomina Anatomica Veterinaria; NAA, Nomina Anatomica Avium); in the rest of the tetrapods the use of anatomical terms has been confusing (Harris, 2004; Wilson, 2006).
Regarding archosaurs, the processus transverses appears as a synonymy of diapophysis in avian standardized terminology (NAA) (Baumel et al., 1993, p. 87). In crocodilians, the other extant archosaurs, some authors (Hofftetter and Gasc, 1969; Rieppel, 1993) argue that caudal transverse processes are simple structures homologous to dorsal ribs, since complete ontogenetic series and ossification patterns are well known.
On the other hand, scant data about ontogenetic series of extinct archosaurs (e.g. sauropods) leads to weak and subjective comparisons as the basis for homology statements.
In sauropod dinosaurs, most investigators apply traditional anatomical terminology ("Romerian" sensu Wilson, 2006). However the avian nomenclature has been used in several recent contributions, based on the fact that birds are their closest living relatives (Wedel and Sanders, 1999, 2002; Sanders et al. 2000).
In order to standardize sauropod vertebrae nomenclature, the following use of traditional terms is suggested for lateral vertebral processes.
Diapophyses, Parapophyses. These words were coined by Owen (in Romer, 1956) for lateral processes where rib-heads tuberculum and capitulum articulated respectively. In presacral vertebrae they are simple and easily distinguishable elements, thus we maintain this original use only for the presacral vertebrae (figure 1.1).
Transverse processes. This term is used for lateral bony projections in presacral vertebrae (e.g. diapophyses) (Riggs, 1904; Ikejiri et al., 2005) as well as for the lateral structure on caudal vertebrae, composed of more than one element (Hatcher, 1903). Here we consider it prudent to avoid use of transverse processes for the lateral projection in presacral vertebrae, and instead use the traditional terms (diapophyses and parapophyses). It is suitable to preserve transverse processes for caudal vertebrae, in which the lateral process seems to have a more complex developmental origin from multiple ossifications (Wilson, 1999) (figures 1.2, 1.3).
"Caudal ribs". This term is used as a synonym of caudal transverse processes (McIntosh, 1990; Up-church et al., 2004; McIntosh, 2005). We suggest discarding the term "caudal rib" in reference to the entire caudal transverse processes, because the ACTP in several sauropods comprises other elements besides ribs (i.e. ribs, ribs articulations, laminae), as will be discussed below.
Institutional abbreviations. CM, Carnegie Museum of Natural History, Pittsburgh, Pennsylvania, USA; FMNH PR, Field Museum of Natural History, Chicago, USA; MAFI, Magyar Allami Foldtani Intezet, Budapest, Hungary; Mal, Malawi Department of Antiquities Collection, Liongwe and Nguludi, Malawi; MCT, Museu de Ciências da Terra, Departamento Nacional de Produção Mineral, Río de Janeiro, Brasil; MJG, Museo "Jorge Gerhold", Jacobacci, Río Negro Province, Argentina; MLP-Ly, Museo de La Plata,
Buenos Aires Province, Argentina. Lydekker´s collection; MPCA, Museo Provincial "Carlos Ameghino", Cipolletti, Río Negro Province, Argentina; MRS-Pv, Museo "Argentino Urquiza", Rincón de los Sauces, Neuquén province, Argentina; MUCPv, Museo de Geología y Paleontología de la Universidad Nacional del Comahue, Neuquén Province, Argentina; PVL, Instituto-Fundación"Miguel Lillo", Tucumán Province, Argentina; YPM, Yale Peabody Museum, New Haven, Connecticut, USA.
Anatomical abbreviations. acdl, anterior centrodiapophyseal lamina; pcdl, posterior centrodiapophyseal lamina; prdl, prezygodiapophyseal lamina; podl, postzygodiapophyseal lamina; acpl, anterior centroparapophyseal lamina; sprl, spinoprezygapophyseallamina.
Anatomical description of ACTP
In this section we describe the ACTP in several sauropods. We list the sauropod groups in a phylogenetic context and pick representative taxa for each group and/or those with anterior caudal vertebrae preserved. Assignment of taxa to each group is based on the following recent phylogenetic schemes (e.g. Wilson, 2002; Upchurch et al., 2004; Harris, 2006)
Non-neosauropod eusauropods. The ACTPs are complex, placed on the junction between neural arch and centrum, but bony bars cannot be recognized. In Patagosaurus, Bonaparte (1986, fig.45a) described the ACTP as short and dorsoventrally compressed. However, in PVL-4170 the ACTP is a well developed, laminar, fan-like structure, that is antero-posteriorly compressed. Its base extends from the middle of the centrum up to the level of the neural canal. The distal end is wide with the dorsal edge expanded upwards. The first caudal transverse process in Omeisaurus junghsiensis is fan-shaped and anteriorly concave (figure 2.1), but unlike Patagosaurus, the ventral margin of the process is oriented downwards. The second caudal is similar to the first one, but with a less developed ACTP. The following ones have less developed transverse processes that appear as small lateral bony projections (Young, 1937, fig. 8). Mamenchisaurus hochuanensis has a well-developed ACTP in the first caudal, antero-posteriorly expanded and relatively thin and flat dorsally and ventrally (Young and Zhao, 1972). In the turiasaur Losillasaurus giganteus, the transverse processes of anterior caudals consist of fan-like structures, with the ventral portion more developed than the dorsal one. Mid-anterior caudal vertebrae show the same pattern as the most anterior caudals, although the dorsal portion of the process is very poor developed (Casanovas et al., 2001, pl.4; Royo-Torres et al., 2006).

Figure 2. Schematics anterior caudal vertebrae of sauropod dinosaurs in anterior view. 1, Omeisaurus (modified from Young, 1937); 2, Limaysaurus (modified from Calvo and Salgado, 1995); 3, Dicraeosaurus (modified from Janensch, 1914); 4, Barosaurus (modified from Lull, 1919); 5, Camarasaurus (modified from Osborn and Mook, 1921); 6, Epachthosaurus (modified from Martinez et al., 2004); 7, Baurutitan (modified from Kellner et al., 2005); 8, Saltasaurus (modified from Powell, 1986). Reconstructed areas in grey. Scale bar: 10 cm / vértebras caudales anteriores esquemáticas de dinosaurios saurópodos en vista anterior. 1, Omeisaurus (modificado de Young, 1937); 2, Limaysaurus (modificado de Calvo y Salgado, 1995); 3, Dicraeosaurus (modificado de Janensch, 1914); 4, Barosaurus (modificado de Lull, 1919); 5, Camarasaurus (modificado de Osborn y Mook, 1921); 6, Epachthosaurus (modificado de Martinez et al., 2004); 7, Baurutitan (modificado de Kellner et al., 2005); 8, Saltasaurus (modificado de Powell, 1986). Areas reconstruidas en gris. Escala gráfica: 10 cm.
Diplodocoid neosauropods. Diplodocoid neosauropods have complex ACTPs, at least in adult stages (see Diplodocids below). They are composed of bony plates supported by dorsal and ventral bars. Several authors describe this structure as wing-like transverse processes (McIntosh, 1990; Upchurch, 1998; Wilson, 2002).
Basal diplodocoids. In Zapalasaurus, the ACTP of the fourth caudal are laterally and dorsally projected, although it is not possible to discern if they are triangular or wing-like (Salgado et al., 2006a ).
Rebbachisaurids. In rebbachisaurids the ACTPs have one horizontal dorsal bar and an oblique ventral one. A thin lamina of bone is developed between them. Calvo and Salgado (1995) described this condition in Caudal 3 of Limaysaurus (="Rebbachisaurus") tessonei (figure 2.2). Neither the holotype of Limaysaurus tessonei (Calvo and Salgado, 1995) nor the recent specimens assigned to it (Salgado et al., 2004) show evidence of wing-like transverse processes. The condition in Rebbachisaurus garasbae Lavocat 1954 is problematic. Calvo and Salgado (1995) mentioned the presence of a true wing-like transverse process in R. garasbae; however, this condition is based on a field observation done by Lavocat (Lavocat, 1954). In addition, Salgado et al. (2006a) considered a winglike condition (character 25) in R. garasbae for their cladistic analyses. In the anterior caudal of Cathartesaura anaerobica, the ACTP is composed by a thin, horizontal dorsal bar and a reinforced ventral one (Gallina and Apesteguía, 2005, fig. 3). It is completely fused to arch and centrum. This dorsal aspect of the ACTP is a true laminar, laterally extended plate. The oblique ventral bar is a strong cylindrical projection of bone that is both laterally and anteriorly expanded. Both dorsal and ventral bars border a deep triangular fossa between them. (figures 3.1, 3.2)

Figure 3. Two anterior caudal vertebrae of sauropods with diferent ACTPs morphologies. 1, Anterior caudal of Cathartesaura anaerobica in anterior view (right half detail); 2, Anterior caudal of Cathartesaura anaerobica in posterior view (right half detail); 3, Caudal 2 of Bonitasaura salgadoi in anterior view (left half detail); 4, Caudal 2 of Bonitasaura salgadoi in posterior view (left half detail). Scale bar: 10 cm. / dos vértebras caudales anteriores de saurópodos con diferentes morfologías de ACTP. 1, Caudal anterior de Cathartesaura anaerobica en vista anterior (detalle de la mitad derecha); 2, Caudal anterior de Cathartesaura anaerobica en vista posterior (detalle de la mitad derecha); 3, Caudal 2 de Bonitasaura salgadoi en vista anterior (detalle de la mitad izquierda); 4, Caudal 2 de Bonitasaura salgadoi en vista posterior (detalle de la mitad izquierda). Escala gráfica: 10 cm.
Dicraeosaurids. The ACTP morphology can only be checked in the genus Dicraeosaurus. The dorsal and ventral bars are parallel, dorso-laterally projected and similar in length. This condition is present in Caudal 1 through 8 of Dicraeosaurus hansemanni (figure 2.3).
Diplodocids. Diplodocids have complex ACTPs. The dorsal and ventral bars are parallel as in dicraoesaurids, but laterally projected and variable in length. In Caudal 3 of Barosaurus lentus both bars are similar in size and well developed (figure 2.4). This condition is also observed in Caudal 1 of Apatosaurus louiseae. However, in Caudals 2 and 3, the dorsal and ventral bars do not have the same length, the last one being the most developed. They are connected by a perforated lamina (Gilmore, 1936, pl. 26). In Diplodocus carnegiei, the ACTPs are extremely complex with multiple bars and laminae. They are winglike in general aspect and central bony bars are radially arranged. This ACTP morphology extends from Caudal 1 to 7.
Macronarian neosauropods. In macronarians the ACTPs exhibit several modifications, being either complex or simple structures.
Basal macronarians. In Camarasaurus the ACTP are complex structure in the first one or two caudal vertebrae. It is composed of bony plate mainly supported by horizontal ventral bar and reduced dorsal one (figure 2.5) (Osborn and Mook, 1921, pl. 76). In Brachiosaurus altithorax the ACTPs of the first two caudals are simple structures, which project laterally from the uppermost side of the centrum (Riggs, 1904, pl. 55). Haplocanthosaurus priscus has a simple ACTP, projected laterally from the first caudals (Hatcher, 1903, pl. 3). Surprisingly, McIntosh and Williams (1988, p.14) described the condition in Haplocanthosaurus delfsi, as "in the first two vertebrae the diapophyses bear a hint of the wing-like processes seen in anterior caudals of the diplodocids", but this condition cannot be checked from figures.
Basal titanosaurs. There are two ACTP patterns in basal titanosaurs: simple and complex. In Epachthosaurus sciuttoi the first ACTP is a complex structure with the dorsal bar formed as a knob, and the ventral one is enlarged and laterally projected (figure 2.6) (Martínez et al., 2004). Malawisaurus dixeyi has a similar condition in Caudal 2 producing a triangular projection in anterior view (Jacobs et al., 1993). In other materials referred to Malawisaurus (Mal-200, Mal- 225, Gomani, 1999, 2005), the ACTP presents a single element laterally projected with the tips oriented downwards. Andesaurus delgadoi also possesses a simple structure, which is projected backwards and situated on the superior margin of the centrum in the midline of the antero-posterior length (Calvo and Bonaparte, 1991).
Derived titanosaurs (excluding Saltasaurinae). There are two ACTP patterns in non-saltasaurine, derived titanosaurs. Aeolosaurus rionegrinus has a complex ACTP in the anterior caudals, with the dorsal element poorly developed and placed above the dorsal margin line of the neural canal (Salgado and Coria, 1993). A similar morphology is observable in Caudal 1 of Baurutitan britoi (Kellner et al., 2005) (figure 2.7) and Alamosaurus sanjuanensis (Lucas and Hunt, 1989, fig. 4I). On the other hand, in Opisthocoelicaudia the simple transverse processes are placed along the dorsal margin of the centrum in the midline of the anterior- posterior length (Borsuk-Bialynicka, 1977). In the first or second caudal of Mendozasaurus neguyelap it is placed on the posterior margin of the centrum and is postero-laterally projected; furthermore there is a prezygodiapophyseal lamina that joins the ACTP to the prezygapophysis (González Riga, 2003). Isisaurus colberti has a simple element constituting the laterally projected transverse process placed at the midline of antero-posterior length (Jain and Bandyopadhyay, 1997, figs. 7, 11). The first caudal vertebrae of Paralititan have the same condition as Isisaurus. The other proximal caudals of the former taxon have well developed transverse processes that are projected downwards and backwards (Smith et al., 2001, fig 2B, C). In Rinconsaurus caudamirus the first caudal has laterally directed transverse processes. The other anterior caudals have transverse processes directed laterally and posteriorly projected (Calvo and González Riga, 2003). A similar condition is observed in Magyarosaurus dacus where the ACTP are laterally and posteriorly projected. In Bonitasaura salgadoi, the ACTPs of Caudal 2 are composed of a latero-posteriorly projected element. This single element is fused to the centrum, but a suture can be recognize beetween them. Moreover, several independent elements with similar aspect were recovered as isolated elements and later recognized as the unfused ACTPs (figures 3.3, 3.4). This juvenile condition is also observable in anterior caudal elements of Rapetosaurus, including several vertebrae without lateral elements in the transverse processes position.
Saltasaurinae. The ACTP is a complex structure at least in the first caudal of Saltasaurus loricatus where the dorsal and ventral elements are well developed and contact each other along their length (figure 2.8). In the second caudal of Saltasaurus loricatus and the anterior caudals of Neuquensaurus australis, the ACTP is a simple, latero-posteriorly projected element. In the former taxon, the ACTP is anteriorly concave.
Discussion
Morphological aspects: ACTP component identity
At this point, we can recognize four ACTP morphological types: three different types of complex ACTP and one type of simple ACTP. The complex type present in non-neosauropod eusauropods (fanlike, without recognizable bony bars) (figure 2.1) differs from those present in diplodocoid sauropods (with dorsal element well developed) (figures 2.2, 2.3, 2.4) and some macronarians (with dorsal element poorly developed) (figures 2.5, 2.6). The simple ACTP is a single element, which is laterally and posteriorly projected in most cases (figure 3.3). This morphological variation (without a phylogenetic correlate) clearly shows that different elements participate in ACTP construction. We will analyze the possible contribution of several elements related to axial skeleton formation to the ACTP.
Ribs. The absence of complete ontogenetic series in sauropods dinosaur obscures direct recognition of homologous structures. However, several juvenile and sub-adult specimens contribute toward clarifying this matter. Recently published diplodocid material (SMA 0009) from the Late Jurassic of Wyoming (Morrison Formation) shows an independent lateral element in the ACTP without any laminar connection to either the arch or centrum (figure 4.1; Schwarz et al., 2007, fig 6F). This condition is also observable in sub-adult specimens of Camarasaurus (CM 11338, YPM 1910), Bonitasaura salgadoi, as well as in Rapetosaurus (FMNH PR 2209). As can be seen in several extant reptiles (squamates and crocodilians), the rib elements ossify separately from the neural arch and centrum in caudal vertebrae (Rieppel, 1992, 1993). Thus, the lateral element present in SMA 0009 may represent a caudal rib, as was already suggested by Schwarz et al. (2007).
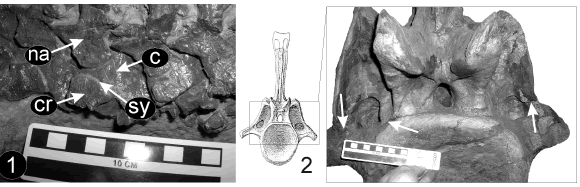
Figure 4. ACTP complex identity in diplodocids. 1, Caudals 1-4 of a juvenile diplodocid (SMA 0009) in left lateral view; 2, Caudal 2 of Apatosaurus louisae (CM 3018) in anterior view. Note suture line indicated with white arrows. c, centrum; cr, caudal rib; na, neural arch; sy, synapophysis. Scale bar: 10 cm / identidad de los ACTP complejos en diplodócidos. 1, Caudales 1-4 de un diplodócido juvenil (SMA 0009) en vista lateral izquierda; 2, Caudal 2 de Apatosaurus louisae (CM 3018) en vista anterior. Nótese la línea de sutura indicada con flechas blancas. c, centro; cr, costilla caudal; na, arco neural; sy, sinapofisis. Escala gráfica: 10 cm.
As mentioned, adult diplodocoids have complex ACTPs with dorsal and ventral elements. Compared to juvenile specimens, the ventral one would represent the rib component both in location and apparent independence from the centrum with sutural differences indicating separate ossifications (see adult of Apatosaurus louisae CM 3018, figure 4.2). On the other hand, the remarkable ACTP dorsal component can be explained by means of laminar development (see below).
Rib articulations. Double-headed and single-headed ribs are distributed along the axial skeleton. Romer (1956, p. 225) remarked "(a)n articular process or surface to which attaches a rib whose single head is known or assumed to be formed by union of capitular and tubercular areas is sometimes termed a synapophysis, particularly if it formed jointly by arch and centrum...". In this way, a synapophysis would be present in the caudal region since caudal ribs are single-headed. This is not in agreement with Calvo and Salgado (1995) who recognized a diapophyseal and parapophyseal contribution in the Limaysaurus tessonei ACTP.
Laminae. Laminae are clearly present in the ACTPs of several sauropods. Wilson (1999) suggests that anterior caudal transverse process laminae correspond to the presacral diapophyseal laminae (the ventral acdl, pcdl and the dorsal prdl, podl) as a conservative measure taking into account that the diapophysis is the dominant rib articulation both in terms of size and number of laminar supports (Wilson, 1999, fig. 4). Nonetheless, as the single-headed rib element articulates with a synapophysis ventrally in caudal region, the identity of the laminae remains uncertain. Thus, the laminae of ACTPs could be named according to the presacral laminae arrangement.
Ventral laminae. As the paparapophyses migrate and meet the diapophyses along the dorsal series, the acdl disappear. Concomitantly, the acpl become the main anterior laminae connecting to the centra. This condition is maintained in the anterior caudal vertebrae. Posteriorly, the pcdl are present along the dorsal series, and it seems that the same condition holds for the caudal series (Wilson, 1999).
Dorsal laminae. Dorsally, laminae are present in ACTPs of several sauropods although their number and homologies are variable. The following discussion is based on Apatosaurus louisae (CM 3018) and Trigonosaurus pricei (MCT 1488-R, Campos et al., 2005), because of the presence of complete vertebral series as well as representatives of two well-known sauropod groups. In diplodocoid sauropods the prdl join the prezygapophyses to the diapophyses in dorsal and caudal vertebrae as in titanosaurs (e.g. Mendozasaurus, Bonitasaura). On the other hand, the posterior connection between ACTPs and postzygapophyses shows significant differences within sauropod groups. In diplodocoids, the podl remain uniform along dorsal and caudal series (figure 5.1). Conversely, in the titanosaur Trigonosaurus the podl migrates to the spine along the dorsal series as was recognized by Salgado et al. (2006b). In the caudal region, this lamina becomes disconnected from the diapophysis and postzygapophysis. Thus, no laminar structure unites the ACTPs with the postzygapophysis. As a consequence of this new arrangement, the sprl of titanosaur caudals would be homologous to podl, not to cervical sprl (figure 5.2).

Figure 5. Laminar contribution in ACTP. 1, Dorsal 2,5,9 and caudal 2 of Apatosaurus louisae in lateral view (modified from Gilmore, 1936); 2, Dorsal 4,5,9,10 and caudal 2 of Trigonosaurus pricei in lateral view (modified from Campos et al., 2005). acdl, anterior centrodiapophyseal lamina; acpl, anterior centroparapophyseal lamina; pcdl, posterior centrodiapophyseal lamina; podl, postzygodiapophyseal lamina; poz, postzygapophysis; prdl, prezygodiapophyseal lamina; prz, prezygapophysis; sprl, spinoprezygapophyseal lamina. Not to scale / contribución de láminas en los ACTP. 1, Dorsales 2,5,9 y caudal 2 de Apatosaurus louisae en vista lateral (modificado de Gilmore, 1936); 2, Dorsales 4,5,9,10 y caudal 2 de Trigonosaurus pricei en vista lateral (modificado de Campos et al., 2005). acdl, lámina centrodiapofisial anterior; acpl, lámina centroparapofisial anterior; pcdl, lámina centrodiapofisial posterior; podl, lámina postzigodiapofisial; poz, postzigapófisis;a prdl, lámina prezigodiapofisial; prz, prezigapófisis; sprl, lámina espinoprezigapofisial. Sin escala.
In summary, in complex ACTPs (with an extreme development in diplodocoids) the ventral element is a synapophysis with rib fusion, while the dorsal element is composed by lateral growth of laminae. Since podl are present in this ACTP type, this lamina seems to be the most important dorsal constituent. By contrast, the simple ACTPs are composed of synapophyses with rib fusion.
Phylogenetic aspects
Here we have revealed that there is widespread morphological variation in the ACTP. However, this is not strictly related to the subdivisions of the major groups within Sauropoda.
In recent years, several contributions on sauropod phylogeny include an ACTP morphological treatment. Calvo and Salgado (1995) recognized three different states for their character 32 ("wing like processes: absent (0), present, well developed (1), formed by a dorsal and ventral bar (2)"). In our judgment, a well developed wing-like process is actually composed of a dorsal and ventral bar, and thus character states (1) and (2) describe the same condition. Furthermore, the condition described in Limaysaurus tessonei (state 2) is not a wing-like transverse process; consequently this cannot be treated as a wing-like type.
Upchurch (1998) used the ACTP morphology to codify his character 143 ("wing or fan-like caudal ribs: absent (0), present on first 3-7 caudals (1)"). According to our proposed nomenclature, the term "caudal ribs" shoul be avoided regarding ACTPs as a whole. In addition, wing-like ACTPs are different from fan-like ACTPs since the former has a wide base and the latter has a narrow base. As a consequence, Patagosaurus (fan-like ACTP) would be miscoded in Upchurch's scheme.
In Wilson's (2002) analysis, the character 128 ("anterior caudal transverse processes, shape: triangular, tapering distally (0); wing-like, not tapering distally (1)") was used to discriminate Diplodocoidea from others sauropods. However, he did not consider the relative development of the ACTP dorsal component as a phylogenetically informative trait (unlike Salgado et al., 2004; Gallina and Apesteguía, 2005; Salgado et al., 2006a).
In this way we suggest the presence/absence of a well development dorsal bony bar in ACTP as valid character states for discerning Diplodocoids from others sauropods instead presence/absence of"wing-like" transverse processes (see above).
At lower-level phylogeny, Diplodocoidea ingroup relationships may be improved by means of analyzing the morphology of the anterior caudal transverse processes. Particular ACTP morphologies are recognized in the well-known families of Diplodocoidea (Rebbachisauridae, Dicraeosauridae, Diplodocidae) (Upchurch, 1998; Wilson, 2002; Upchurch et al., 2004; Harris, 2006). In a phylogenetic sense, such morphology should be analyzed in termsof relative development and orientation of the dorsal and ventral components (figure 6). Potential character states should be presence/absence of dorsal and ventral bony bars with parallel arrangement, adding support to Flagellicaudata.

Figure 6. Morphology of the anterior caudal transverse processes within Diplodocoidea. Phylogenetic scheme based on Wilson (2002). Vertebrae from figure 2. Scale bar: 10 cm / morfología de los procesos transversos de las vértebras caudales anteriores dentro de Diplodocoidea. Esquema filogenético basado en Wilson (2002). Vértebras tomadas de la figura 2. Escala: 10 cm.
Musculoskeletal and functional aspects
The ACTPs are sites of muscle attachment. Hence, variations in ACTP morphology should reflect modifications in musculature. Moreover, because of the epigenetic nature of the bones, the different development of the soft tissues are probably responsible for some of the observed variation in bone morphology (Banks, 1996).
Historical interpretations of sauropod caudal musculature have been predominantly focused on muscles related to the appendicular skeleton that are attached on caudal vertebrae, such as the Mm. caudofemorales (Borsuk-Bialynicka, 1977; Wilhite, 2003; Otero and Vizcaíno, 2006, 2008). However, the sauropod caudal musculature itself is poorly known.
The caudal transverse processes delimit the epaxial and hypaxial musculature. In crocodilians, the dorsal musculature (M. dorsalis trunci) -which runs posteriorly above the lumbar lateral projections- continues posteriorly as the M. dorsalis caudae between the spines and caudal ribs of the caudal vertebrae (Romer, 1923; Frey, 1988). In Neornithes there are two muscles that originate from the dorsal surface of caudal transverse processes: M. levator caudae and M. lateralis caudae (Baumel et al., 1993). Such musculature provides dorsal and lateral movements of the tail. Because several sauropods (e.g., Diplodocoids) have a well-developed ACTP dorsal element, the presence of a similar musculature with the same position and function is inferred.
In the ventral region of the tail there are caudal ventral musculature origins. In crocodilians the M. ilio-ischio-caudalis is a muscle that extends along the ventral half of the tail on either side between the caudal ribs and the midline (Romer, 1923; Frey, 1988). The most important muscle that has an origin site on proximal caudal vertebrae is the M. caudofemoralis longus (=CFL). In crocodilians, it originates from the sides of the centrum and ventral surface of the caudal ribs of caudal vertebrae 3-15 (Romer, 1923; Galton, 1969). Wilhite (2003) also places the origin of the CFL from the lateral surface of the first 13 chevrons.
In sauropod dinosaurs there is no consensus on the precise origin of M. caudofemoralis longus. Borsuk- Bialynicka (1977) proposes an origin from the lateral and ventral surfaces of proximal caudal vertebrae (see also Salgado and García, 2002; Salgado et al., 2005). Other authors (Wilhite, 2003; Otero and Vizcaíno, 2006, 2008) propose an additional origin on the lateral sides of proximal haemal arches. As can be seen, no attention has been paid to the transverse processes as an origin site for this muscle.
The M. caudofemoralis longus has a two-component line of action: anterior-posterior (retractor) and lateral- medial (adductor). In titanosaurs (e.g., Laplatasaurus MPCA 1501, Neuquensaurus) the adductor component is very important due to their wide-gauge posture. This condition may be related to the particular lateral surface morphology of the anterior caudal centra (Salgado and García, 2002) and the antero-posterior development of the haemal archs (Wilhite, 2003; Otero and Vizcaíno, 2006, 2008) (figure 7.1). On the other hand, in non-titanosaurs sauropods the retractor component is significant for a narrow-gauge posture, although caution would be warranted with some non-titanosaurs (i.e. Apatosaurus), as potential wide gauge trackmakers (Henderson, 2006). Diplodocidae do not possess primary and secondary lateral surfaces in the anterior caudal centra, but the ACTP ventral component has a well developed concave tip oriented downwards. We propose a ventral site origin of the M. caudofemorales on the ACTP (figure 7.2).

Figure 7. Probable development of the caudofemoralis muscular group in sauropod dinosaurs. 1, left, "wide-gauge" posture (modified from Wilson and Carrano, 1999), and right, caudal 5 of Baurutitan in posterior view (modified from Kellner et al., 2005); 2, left, "narrowgauge" posture (modified from Wilson and Carrano, 1999), and right, caudal 7 of Diplodocus in posterior view (modified from Osborn, 1899); 3, Relative development of the caudofemoralis group in titanosaurs. cfb, caudofemoralis brevis; cfl, caudofemoralis longus; ct, concave tip. Muscular areas in grey. Scale bar: 10 cm / probable desarrollo del grupo muscular caudofemoralis en dinosaurios saurópodos. 1, izquierda, postura "wide-gauge" (modificado de Wilson y Carrano, 1999), y derecha, caudal 5 de Baurutitan en vista posterior (modificado de Kellner et al., 2005); 2, izquierda, postura "narrow-gauge" (modificado de Wilson y Carrano, 1999), y derecha, caudal 7 de Diplodocus en vista posterior (modificado de Osborn, 1899); 3, Desarrollo relativo del grupo muscular caudofemoralis en titanosaurios. cfb, caudofemoralis brevis; cfl, caudofemoralis longus; ct, punta cóncava. Areas musculares en gris. Escala: 10 cm.
In Caiman latirostris the M. caudofemoralis brevis origin site not only involves the medial and lateral surfaces of the postacetabular ilium, but also in the first caudal vertebra (pers. obs.). This situation is also present in Alligator mississippiensis as was recognized only by Romer (1923). In some titanosaurs (e.g. Saltasaurus) there are noteworthy morphological variations in the ACTPs (dorso-ventral development and lateral extension) of the first caudal vertebra compared to the following ones. This variation should presumably be interpreted as a consequence of the relative development of the M. caudofemoralis brevis taking into account soft tissues have morphogenetic dominance over osseous tissues (Witmer, 1997). This relative development could be related to the procoelus condition of titanosaurian caudal vertebrae given that procoely has advantages from a biomechanical point of view (Powell, 2003). That is because a "ball and socket" articulation allows more degrees of freedom than an amphyplatic condition, so the proximal tail needs a muscular arrangement to enclose those mobile joints. In addition, this morphological variation agrees with the anterior extent of the M. caudofemoralis longus (figure 7.3).
As a result, we suggest that the relative development of the Mm. caudofemorales group occurs in intimate relation with the anterior caudal transverse processes morphological variation along the tail. This caudal morphological variation is expressed as a "sacralization" (general aspect of caudal vertebrae resembling general aspect of sacral ones) and extends up to the first fifteen caudal vertebrae in Crocodilus and Caiman among extant archosaurs (pers. obs.) as well as in Diplodocus (related with M. caudofemoralis longus development ) whereas in Saltasaurus (related with M. caudofemoralis brevis development) involves only the first caudal.
Conclusions
There are four different kinds of ACTP. One simple composed of a single element projected on the vertebral side, and three complex composed of lateral bony laminar plates supported by bony bars in most cases.
The simple ACTPs are composed of a synapophysis with rib fusion. In the complex ACTP (with an extreme developed in diplodocoids) the ventral element is a synapophysis with rib fusion, while the dorsal element is composed by lateral growth of laminae. Since podl are present in this ACTP type, this lamina seems to be the most important dorsal constituent.
The ACTP laminae include the acpl and the pcdl ventrally, and the prdl and the podl (when present) dorsally. In-group relationships among Diplodocoidea can be improved by means of analyzing the morphology of the anterior caudal transverse processes.
The "sacralization" of the ACTPs could be related to the attachment of the soft tissues associated to the tail. In this respect the Mm. caudofemorales should be the most important muscle group. The morphological ACTP variation between the first and consecutive caudal vertebrae in some macronarians can be interpreted as a consequence of the relative development of the M. caudofemoralis brevis. On the other hand, the extensive "sacralization" in diplodocoids can be explain with the relative development of the M. caudofemoralis longus.
Acknowledgements
We would like to thank L. Salgado, J. Wilson, S.Apesteguía and J. Bonaparte for comments and discussions at different stages of this paper. We also thank the revision made by R. Royo-Torres and P. Makovicky with several and productive suggestions. J. Bodnar and P. Makovicky improved the English version of the original manuscript. P. Chiarelli and M. Taglioretti are acknowledged for assistance with figure 2. We thank the authorities of the CICYTTP-CONICET (Diamante, Entre Ríos) where the dissections were made and C. Piña for providing the specimens of Caiman. J. González is thanked for helping on dissections. We also thank T. Ikejiri for sending photographs and information of juvenile materials of Camarasaurus and diplodocids. The first author thanks Fundación de Historia Natural "Félix de Azara".
References
1. Apesteguía, S. 2004. Bonitasaura salgadoi gen. et sp. nov.: a beaked sauropod from the Late Cretaceous of Patagonia. Naturwissenschaften 91: 493-497. [ Links ]
2. Banks, W.J. 1996. Histología Veterinaria Aplicada. Second Edition. El Manual Moderno. México D.F., 750 pp. [ Links ]
3. Baumel, J.J., King, A.S., Breazile, J.E., Evans, H.E. and Vanden Berge, J.C. 1993. Handbook of avian anatomy: Nomina Anatomica Avium, 2° ed. Nuttal Ornithological Club. Cambridge, 779 pp. [ Links ]
4. Bonaparte, J.F. 1979. Dinosaurs: a Jurassic assemblage from Patagonia. Science 205: 1377-1379. [ Links ]
5. Bonaparte, J.F. 1986. Les Dinosaures (Carnosaures, Allosauridés, Sauropodes, Cétiosauridés) du Jurassique moyen de Cerro Cóndor (Chubut, Argentine). Annales de Paléontologie 72: 325- 386. [ Links ]
6. Bonaparte, J.F. 1999. Evolución de las vértebras presacras en Sauropodomorpha. Ameghiniana 36: 115-187. [ Links ]
7. Bonaparte, J.F. and Powell, J.E. 1980. A continental assemblage of tetrapods from the Upper Cretaceous beds of El Brete, Northwestern Argentina (Sauropoda-Coelurosauria-Carnosauria- Aves). Mémoirs de la Societé Géologique de France 139: 19- 28. [ Links ]
8. Borsuk-Bialynicka, M. 1977. A new camarasaurid sauropod Opisthocoelicaudia skarzynskii, gen. n., sp. n. from the Upper Cretaceous of Mongolia. Palaeontologia Polonica 37: 1-64. [ Links ]
9. Calvo, J.O. and Bonaparte, J.F. 1991. Andesaurus delgadoi gen. et sp. nov. (Saurischia-Sauropoda), dinosaurio Titanosauridae de la Formación Río Limay (Albiano-Cenomaniano), Neuquén, Argentina. Ameghiniana 28: 303-310. [ Links ]
10. Calvo, J.O. and González Riga, B.J. 2003. Rinconsaurus caudamirus gen. et sp. nov., a new titanosaurid (Dinosaurio, Sauropoda) from the Late Cretaceous of Patagonia, Argentina. Revista Geológica de Chile 30: 333-353. [ Links ]
11. Calvo, J.O. and Salgado, L. 1995. Rebbachisaurus tessonei sp. nov. a new Sauropoda from the Albian-Cenomanian of Argentina; new evidence on the origin of Diplodocidae. Gaia 11: 13-33. [ Links ]
12. Campos, D.A, Kellner, A.W.A., Bertini, R.J. and Santucci, R.M. 2005. On a titanosaurid (Dinosauria, Sauropoda) vertebral column from the Bauru group, Late Cretaceous of Brazil. Arquivos do Museu Nacional 63: 565-593. [ Links ]
13. Casanovas, M.L., Santafé, J.V. and Sanz, J.L. 2001. Losillasaurus giganteus, un nuevo saurópodo del tránsito Jurásico - Cretácico de la cuenca "Los Serranos" (Valencia, España). Paleontología i Evolució 32-33: 99-122. [ Links ]
14. Frey, E. 1988. Anatomie des Korperstammes von Alligator mississippiensis Daudin. Stuttgarter Beitrage zur Naturkunde, Series A 424: 1-106. [ Links ]
15. Gallina, P.A. 2006. Morfología de los procesos transversos en las vértebras caudales anteriores de los saurópodos (Dinosauria). 9° Congreso Argentino de Paleontología y Bioestratigrafía (Córdoba). Academia Nacional de Ciencias, Resúmenes p. 127. [ Links ]
16. Gallina, P.A. and Apesteguía, S. 2005. Cathartesaura anaerobica gen. et sp. nov., a new rebbachisaurid (Dinosauria, Sauropoda) from the Huincul Formation (Upper Cretaceous), Río Negro, Argentina. Revista del Museo Argentina de Ciencias Naturales, n.s. 7: 153-166. [ Links ]
17. Galton, P.M. 1969. The pelvic musculature of the dinosaur Hypsilophodon (Reptilia: Ornithischia). Postilla 131: 64. [ Links ]
18. Gavrilov, K. 1959. Curso de Anatomía y Fisiología Comparada. IV: Esqueleto. Primera Parte. Universidad Nacional de Tucumán, Escuela Universitaria de Ciencias Naturales. Tucumán, 119 pp. [ Links ]
19. Gilmore, C.W. 1936. Osteology of Apatosaurus, with special referente to specimens in the Carnegie Museum. Memoirs of The Carnegie Museum 11: 175-300. [ Links ]
20. Gomani, E.M. 1999. Sauropod caudal vertebrae from Malawi, Africa. Proceedings of the Second Gondwanan Dinosaur Symposium, National Science Museum Monographs 15: 235-248. [ Links ]
21. Gomani, E.M. 2005. Sauropod dinosaurs from the early Cretaceous of Malawi, Africa. Palaeontologia Electronica 8: 27A: 37p. http://palaeo-electronica.org/2005_1/gomani27/issue1_ 05.htm [ Links ]
22. González Riga, B.J. 2003. A new titanosaur (Dinosauria, Sauropoda) from the Upper Cretaceous of Mendoza province, Argentina. Ameghiniana 40: 155-172. [ Links ]
23. Grassé, P. 1975. Zoología II. Vertebrados. Anatomía Comparada. Editorial Torai-Masson S.A. 438 pp. [ Links ]
24. Harris, J.D. 2004. Confusing dinosaurs with mammals: tetrapod phylogenetics and anatomical terminology in the world of homology. The Anatomical Record 281A: 1240-1246. [ Links ]
25. Harris, J.D. 2006. The significance of Suuwassea emilieae (Dinosauria: Sauropoda) for flagellicaudatan intrarelationships and evolution. Journal of Systematic Paleontology 4: 185-198. [ Links ]
26. Hatcher, J.B. 1901. Diplodocus (Marsh): Its osteology, taxonomy, and probable habits, with a restoration of the skeleton. Memoirs of the Carnegie Museum 1: 1-63. [ Links ]
27. Hatcher, J.B. 1903. Osteology of Haplocanthosaurus, with description of a new species, and remaks on the probable habits of the Sauropoda and the age and origin of the Atlantosaurus Beds. Memoirs of the Carnegie Museum 2: 1-72. [ Links ]
28. Henderson, D.M. 2006. Burly gaits: centers of mass, stability, and the trackways of sauropod dinosaurs. Journal of Vertebrate Paleontology 26: 907-921. [ Links ]
29. Hildebrand, M. 1988. Analysis of Vertebrate Structure. John Wiley& Sons, Inc. 701 pp. [ Links ]
30. Hofftetter, R. and Gasc, J. 1969. Vertebrae and Ribs of Modern Reptiles. In: C. Gans (ed.), Biology of the Reptilia, Volume 1, Academic Press, London, pp. 201-310. [ Links ]
31. Ikejiri, T., Tidwell, V. and Trexler, D.L. 2005. New adult Specimens of Camarasaurus lentus highlight ontogenetic variation within the species. In: V. Tidwell and K. Carpenter (eds.), Thunder-lizards: the Sauropodomorph dinosaurs. Indiana University Press, Bloomington, pp. 154-179. [ Links ]
32. Jacobs, L.L., Winkler, D.A., Downs, W.R. and Gomani, E.M. 1993. New material of an Early Cretaceous titanosaurid sauropod dinosaur from Malawi. Palaeontology 36: 523-534. [ Links ]
33. Jain, S.L, and Bandyopadhyay, S. 1997. New titanosaurid (Dinosauria: Sauropoda) from the Late Cretaceous of Central India. Journal of Vertebrate Paleontology 17: 114-136. [ Links ]
34. Janensch, W. 1914. Übersicht über die Wirbeltierfauna der Tendaguruschichten, nebst einer kurzen Charakterisierung der neu aufgeführten Arten von Sauropoden. Archiv für Biontologie 3: 81-110. [ Links ]
35. Kardong, K.V. 1999. Vertebrates: Comparative Anatomy, Function, Evolution. McGraw-Hill. 896 pp. [ Links ]
36. Kellner, A.W.A., Campos, D.A. and Trotta, M.N.F. 2005. Description of a titanosaurid caudal series from The Bauru Group, The Late Cretaceous of Brazil. Arquivos do Museu Nacional 63: 529-564. [ Links ]
37. Lavocat, R. 1954. Sur les Dinosauriens du continental intercalaire des Kem Kem de la Daoura. Comptes Rendus 19° Session Congres Geologique International (1952) 3: 65-68. [ Links ]
38. Lucas, S.G. and Hunt, A.P. 1989. Alamosaurus and the sauropod hiatus in the Cretaceous of the North American Western Interior. Geological Society of America Special Paper 238: 75-85. [ Links ]
39. Lull, R.S. 1919. The sauropod dinosaur Barosaurus Marsh. Memoirs of the Connecticut Academy of Arts and Science 6: 1-42 [ Links ]
40. Lydekker, R. 1893. The dinosaurs of Patagonia. Anales del Museo de La Plata 2: 1-14. [ Links ]
41. Martínez, R.D., Giménez, O., Rodríguez, J., Luna, M. and Lamanna, M.C. 2004. An articulated specimen of the basal titanosaurian (Dinosauria: Sauropoda) Epachthosaurus sciuttoi from the Early Late Cretaceous Bajo Barreal Formation of Chubut province, Argentina. Journal of Vertebrate Paleontology 24: 107-120. [ Links ]
42. McIntosh, J.S. 1990. Sauropoda. In: D.B. Weishampel, P. Dodson and H. Osmólska (eds.), The Dinosauria. University of California Press, Berkeley, pp. 345-401. [ Links ]
43. McIntosh, J.S. 2005. The genus Barosaurus Marsh (Sauropoda, Diplodocidae) In: V. Tidwell and K. Carpenter (eds.), Thunderlizards: the Sauropodomorph dinosaurs. Indiana University Press, Bloomington, pp. 38-77. [ Links ]
44. McIntosh, J.S. and Williams, M.E. 1988. A new species of sauropod dinosaur, Haplocanthosaurus delfsi sp. nov., from the Upper Jurassic Morrison Fm. of Colorado. Kirtlandia 43: 3-26. [ Links ]
45. Nopcsa, F. 1915. Die Dinosaurier der Siebenbürgischen landesteile Ungarns. Mitteilungen aus dem Jahrbuch der Ungarischen Geologischen Reichsanstalt 23: 3-24. [ Links ]
46. Osborn, H.F.1899. A skeleton of Diplodocus. Memories of the American Museum of Natural History 1: 191-214. [ Links ]
47. Osborn, H.F. and Mook, C. 1921. Camarasaurus, Amphicoelias, and other sauropods of Cope. Memoirs of the American Museum of Natural History 3: 247-387. [ Links ]
48. Otero, A. and Vizcaíno, S.F. 2006. Hindlimb musculature of Neuquensaurus australis Lydekker (Sauropoda, Titanosauria). 9° Congreso Argentino de Paleontología y Bioestratigrafía (Córdoba). Academia Nacional de Ciencias, Resúmenes p. 135. [ Links ]
49. Otero, A. and Vizcaíno, S.F. 2008. Hindlimb musculature and function of Neuquensaurus australis Lydekker (Sauropoda: Titanosauria). Ameghiniana 45: 333-348. [ Links ]
50. Powell, J.E. 1986. [Revisión de los Titanosáuridos de América del Sur. Ph. D. Thesis, Universidad Nacional de Tucumán, 493 pp. Unpublished. [ Links ]].
51. Powell, J.E. 1987. The Late Cretaceous Fauna of Los Alamitos, Patagonia, Argentina. Part IV - The Titanosaurids. Revista del Museo Argentino de Ciencias Naturales 3: 147-153. [ Links ]
52. Powell, J.E. 2003. Revision of South American Titanosaurid dinosaurs: palaeobiological, palaeobiogeographical and phylogenetic aspects. Records of the Queen Victoria Museum. N°111. Launceston, 173 pp. [ Links ]
53. Rieppel, O. 1992. Studies on skeleton formation in reptiles. III. Patterns of ossification in the skeleton of Lacerta vivipara Jacquin (Reptilia, Squamata). Fieldiana: Zoology n.s. 68: 1-25 [ Links ]
54. Rieppel, O. 1993. Studies on skeleton formation in reptiles. V. Patterns of ossification in the skeleton of Alligator mississippiensis Daudin (Reptilia, Crocodylia). Zoological Journal of the Linnean Society 109: 301-325. [ Links ]
55. Riggs, E.S. 1904. Structure and relationships of opisthocoelian dinosaurs. Part II: The Brachiosauridae. Field Columbian Museum of Geology 2: 229-248. [ Links ]
56. Romer, A.S. 1923. Crocodilian pelvic muscles and their avian and reptilian homologues. Bulletin of the American Museum of Natural History 48: 533-552. [ Links ]
57. Romer, A.S. 1956. Osteology of reptiles. The University of Chicago Press. Chicago, 770pp. [ Links ]
58. Romer, A.S. 1962. The vertebrate Body (shorter version). W. B. Saunders Company. Philadelphia, 425pp. [ Links ]
59. Royo-Torres, R., Cobos, A. and Alcalá, L. 2006. A giant European dinosaur and a new sauropod clade. Science 314: 1925-1927. [ Links ]
60. Salgado, L. and Coria, R.A. 1993. El género Aeolosaurus (Sauropoda, Titanosauridae) en la Formación Allen (Campaniano-Maastrichtiano) de la provincia de Río Negro. Ameghiniana 30: 119-128. [ Links ]
61. Salgado, L. y García, R. 2002. Variación morfológica en la secuencia de vértebras caudales de algunos saurópodos titanosaurios. Revista Española de Paleontología 17: 211-216. [ Links ]
62. Salgado L, Apesteguía S, Heredia S.E. 2005. A new specimen of Neuquensaurus australis, a Late Cretaceous Saltasaurinae titanosaur from North Patagonia. Journal of Vertebrate Paleontology 25: 623-634. [ Links ]
63. Salgado, L., Carvalho, I.S. and Garrido, A. 2006a. Zapalasaurus bonapartei, un nuevo dinosaurio saurópodo de La Formación La Amarga (Cretácico Inferior), noroeste de Patagonia, Provincia de Neuquén, Argentina. Geobios 39: 695-707. [ Links ]
64. Salgado, L., García, R.A. and Daza, J.D. 2006b. Consideraciones sobre las láminas neurales de los dinosaurios saurópodos y su significado morfofuncional. Revista del Museo Argentina de Ciencias Naturales, n.s. 8: 69-79. [ Links ]
65. Salgado, L., Garrido, A., Cocca, S.E. and Cocca, J.R. 2004. Lower Cretaceous rebbachisaurid sauropods from Aguada del león (Lohan Cura Formation), Neuquén Province, Northwestern Patagonia, Argentina. Journal of Vertebrate Paleontology 24: 903- 912. [ Links ]
66. Sanders, R.K., Wedel, M.J., Sereno, P.C. and Staab, G. 2000. A restoration of the cranio-cervical system in Jobaria. Journal of Vertebrate Paleontology 20: 67A. [ Links ]
67. Schwarz, D., Ikejiri, T., Breithaupt, B.H., Sander, P.M. and Klein, N. 2007. A nearly complete skeleton of an early juvenile diplodocid (Dinosauria: Sauropoda) from the Lower Morrison Formation (Late Jurassic) of north central Wyoming and its implications for early ontogeny and pneumaticity in sauropods. Historical Biology 19: 225-253. [ Links ]
68. Smith, J.B., Lamanna, M.C., Lacovara, K.J., Dodson, P., Smith, J.R., Poole, J.C., Giegengack, R. and Attia, Y. 2001. A giant sauropod dinosaur from an Upper Cretaceous mangroove deposit in Egypt. Science 292: 1704-1706. [ Links ]
69. Upchurch, P. 1998. The phylogenetic relationships of sauropod dinosaurs. Zoological Journal of the Linnean Society 124: 43-103. [ Links ]
70. Upchurch, P., Barret, P.M. and Dodson, P. 2004. Sauropoda. In: D.B. Weishampel, P. Dodson and H. Osmólska (eds.), The Dinosauria, 2° ed. University of California Press, Berkeley pp. 259-322. [ Links ]
71. Vanden Berge, J.C. 1982. Miología de las aves. In: S. Sisson and J.D. Grossman (eds.), Anatomía de los animales domésticos, Vol. 2, Salvat, Barcelona, pp. 1973-2022. [ Links ]
72. Wedel, M.J. and Sanders, K. 1999. Comparative anatomy and functional morphology of the cervical series in Aves and Sauropoda. Journal of Vertebrate Paleontology 19: 83A. [ Links ]
73. Wedel, M.J. and Sanders, K. 2002. Osteological correlates of cervical musculature in Aves and Sauropoda (Dinosauria: Saurischia), with comments on the cervical ribs of Apatosaurus. PaleoBios 22: 1-16. [ Links ]
74. Weichert, C.K. 1959. Elements of Chordate Anatomy. McGraw-Hill Book Company, Inc. 509 pp. [ Links ]
75. Wilhite, R. 2003. [Biomechanical reconstruction of the appendicular skeleton in three North American Jurassic Sauropods. Ph. D. dissertation. Louisiana State University, Baton Rouge, 198 pp. Unpublished. [ Links ]].
76. Wilson, J.A. 1999. A nomenclature for vertebral laminae in sauropods and other saurischian dinosaurs. Journal of Vertebrate Paleontology 19: 639-653. [ Links ]
77. Wilson, J.A. 2002. Sauropod dinosaur phylogeny: critique and cladistic analysis. Zoological Journal of the Linnean Society 136: 217-276. [ Links ]
78. Wilson, J.A. 2006. Anatomical nomenclature of fossil vertebrates: standardized terms or ´lingua franca'? Journal of Vertebrate Paleontology 26: 511-518. [ Links ]
79. Wilson, J.A. and Carrano, M.T. 1999. Titanosaurs and the origin of "wide-gauge" trackways: a biomechanical and systematic perspective on sauropod locomotion. Paleobiology 25: 252-267. [ Links ]
80. Wilson, J.A., Martinez, R.N. and Alcober, A. 1999. Distal tail segment of a titanosaur (Dinosauria: Sauropoda) from the Upper Cretaceous of Mendoza, Argentina. Journal of Vertebrate Paleontology 19: 591-594. [ Links ]
81. Witmer, L.M. 1995. The extant phylogenetic bracket and the importance of reconstructing soft tissues in fossils. In: J.J Thomason (ed.), Functional morphology in vertebrate paleontology. Cambridge University Press, Cambridge, pp. 19-33. [ Links ]
82. Witmer, L.M. 1997. The evolution of the antorbital cavity in archosaurs: a study in soft-tissue reconstruction in the fossil record with an analysis of the function of pneumaticity. Society of Vertebrate Paleontology Memoir 3: 1-73. [ Links ]
83. Young, C.C. 1937. New Triassic and Cretaceous Reptiles in China. Bulletin of The Geological Society of China 17: 113-118. [ Links ]
84. Young, C.C. and Zhao, X.-J. 1972. Mamenchisaurus hochuanensis sp. nov. Institute of Vertebrate Paleontology and paleoanthropology. Monographs, A, 8: 1-30. [ Links ]
Recibido: 8 de mayo de 2008.
Aceptado: 15 de setiembre de 2008.

