










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana vol.46 no.1 Buenos Aires ene./mar. 2009
NOTA PALEONTOLÓGICA
Tympanoctomys (Mammalia: Rodentia) en el Holoceno de Patagonia, Argentina
Daniel E. Udrizar Sauthier1, Ulyses F.J. Pardiñas1 y Eduardo P. Tonni2
1Unidad de Investigación Diversidad, Sistemática y Evolución, Centro Nacional Patagónico, CC 128, 9120 Puerto Madryn, Chubut, Argentina. dsauthier@cenpat.edu.ar; ulyses@cenpat.edu.ar
2División Paleontología Vertebrados, Museo de La Plata, Paseo del Bosque s/n, B1900FWA, La Plata, Argentina. eptonni@fcnym.unlp.edu.ar
Introducción
Tympanoctomys es un género de roedores octodóntidos, endémico de Argentina, cuyos representantes muestran adaptaciones extremas a la vida en ambientes áridos (e.g., Mares et al., 1997; Ojeda et al., 1999; Diaz et al., 2000). Con una única especie viviente reconocida, T. barrerae (Lawrence, 1941), su distribución actual se verifica en localidades aisladas -básicamente salares- insertas en el bioma del Monte y ecotono Monte Patagonia (Ojeda et al., 2007). Durante la última década este roedor ha sido objeto de intensos estudios y debate en relación a su condición de tetraploide (e.g., Gallardo et al., 1999; Svartman et al., 2005; Gallardo et al., 2006).
El registro fósil de Tympanoctomys es pobremente conocido. En una reciente revisión, Verzi et al. (2002) han discutido en detalle los restos que conforman el mismo y sus procedencias geográficas y cronológicas. Estos autores registran T. cordubensis (Ameghino, 1889), una especie apenas diferenciable de T. barrerae, en el Pleistoceno de Buenos Aires y Córdoba. Para la especie viviente no existen menciones en la literatura paleontológica (Diaz et al., 2000).
En este trabajo se da a conocer, por primera vez para depósitos holocénicos, el hallazgo de numerosos materiales craneanos asignables a T. barrerae. Los mismos han sido exhumados en tres secuencias estratigráficas emplazadas en cercanías del río Chubut (Chubut, Argentina). En este contexto, se discuten aspectos de la paleodistribución de este roedor y sus posibles causas de extinción local.
Los tres yacimientos aquí mencionados fueron excavados mediante la aplicación de técnicas arqueológicas y control estratigráfico riguroso. Todo el sedimento obtenido fue tamizado en seco (con malla de 2 mm de paso) y el remanente, concentrado en el campo y revisado bajo lupa. Los restos fueron determinados anatómica y taxonómicamente hasta el grado posible, se cuantificaron mediante el recuento de elementos homólogos únicos (número mínimo de individuos MNI) y están depositados en la Colección de Material de Egagrópilas y Afines "Elio Massoia" del Centro Nacional Patagónico (CNP-E), Puerto Madryn, Chubut. Las medidas fueron tomadas con calibre manual (precisión 2 centésimos) y se expresan en mm. Para la determinación taxonómica y descripción anatómica del material estudiado se emplearon las contribuciones De Santis et al. (1991), Diaz et al. (2000) y Verzi et al. (2002). Las dataciones radiocarbónicas informadas en este trabajo se expresan en años radiocarbónicos antes del presente (ar AP) y fueron realizadas en el Laboratorio de Tritio y Radiocarbono (LATYR) del Museo de La Plata.
Paleontología sistemática
Orden RODENTIA Bowdich, 1821
Familia OCTODONTIDAE Waterhouse, 1840Género Tympanoctomys Yepes, 1942
Especie tipo. Octomys barrerae Lawrence, 1941.
Tympanoctomys barrerae (Lawrence, 1941)
Figuras 2.1, 2.2
Material referido. CNP-E 259-19 A, hemimandíbula izquierda; CNP-E 259-19 B, hemimandíbula derecha; CNP-E 260-19 A, hemimandíbula derecha; CNP-E 260-19 B, hemimandíbula izquierda; CNP-E 261-19 A, paladar; CNP-E 261-19 B, hemimandíbula izquierda; CNP-E 262-19 A, hemimandíbula izquierda; CNP-E 262- 19 B, hemimandíbula izquierda; CNP-E 262-19 C, hemimandíbula izquierda; CNP-E 262-19 D, hemimandíbula derecha; CNP-E 262- 19 E, fragmento de hemimaxilar derecho; CNP-E 263-20 A, hemimandíbula izquierda; CNP-E 263-20 B, hemimandíbula izquierda; CNP-E 263-20 C, hemimandíbula derecha; CNP-E 264-19 A, hemimandíbula izquierda; CNP-E 264-19 B, hemimandíbula izquierda; CNP-E 264-19 C, hemimandíbula izquierda; CNP-E 265-19 A, hemimandíbula derecha; CNP-E 265-19 B, hemimandíbula derecha; CNP-E 265-19 C, hemimandíbula derecha; CNP-E 265-19 D, hemimaxilar derecho; CNP-E 265-19 E, paladar; CNP-E 265-19 F, hemimandíbula izquierda; CNP-E 266-19 A, hemimandíbula derecha; CNP-E 266-19 B, hemimandíbula izquierda; CNP-E 292-1 A, hemimandíbula izquierda; CNP-E 293-1 A, paladar; CNP-E 324-2 A, hemimandíbula derecha; CNP-E 325-2 A, hemimandíbula derecha; CNP-E 326-2 A, fragmento de hemimaxilar derecho.
Procedencia estratigráfica y geográfica. Los fósiles fueron recuperados a partir de la excavación de tres depósitos sedimentarios en localidades emplazadas en cercanías del río Chubut, provincia del Chubut, Argentina (figura 1), cuyos rasgos principales y cronologías se sintetizan seguidamente.

Figura 1. Ubicación geográfica y perfiles estratigráficos esquemáticos de los yacimientos portadores de restos de Tympanoctomys barrerae (Holoceno medio-tardío, Chubut, Argentina). 1, Porción austral de América del Sur; se indican con círculos los registros actuales más australes para T. barrerae (basado en Ojeda et al., 2007); 2, Mapa del valle medio e inferior del río Chubut con la ubicación de las localidades fosilíferas discutidas en este trabajo (1 = "Perfil Los Altares", 2 = "Cueva de la Virgen" y 3 = "Cueva Caolinera Dique Ameghino"); 3, Perfiles estratigráficos esquemáticos para las secuencias sedimentarias de los yacimientos portadores de T. barrerae / Geographical sitting and schematic stratigraphical profiles of the localities with remains of Tympanoctomys barrerae (Middle-Late Holocene, Chubut, Argentina). 1, Austral portion of South America; circles indicate the southernmost recent recording localities for T. barrerae (after Ojeda et al., 2007); 2, Map of the medium and lower valley of Chubut River with location of the fossiliferous localities discussed in this work (1 = "Perfil Los Altares", 2 = "Cueva de la Virgen" and 3 = "Cueva Caolinera Dique Ameghino"); 3, Schematic stratigraphical profiles for the sedimentary sequences of the deposits with T. barrerae.
El denominado "Perfil Los Altares" (43° 53' 35,2" S, 68° 23' 21,3" O, 219 m), ubicado sobre la banquina sur de la ruta nacional 25 a 1,5 km al sudeste de Los Altares, corresponde a una acumulación sedimentaria remanente del relleno de una antigua cueva eliminada por trabajos viales (figura 1.3). La secuencia, de 120 cm de espesor, está conformada básicamente por limos arenosos de origen eólico con participación variable de material clástico procedente de la meteorización de la roca de caja, camadas de residuos vegetales y fragmentos de carbón. Fue excavado mediante 12 niveles artificiales de 10 cm de espesor cada uno. Dataciones radiocarbónicas sobre carbón vegetal brindaron, para el nivel basal (artificial N° 12), 2210 ± 70 años ar AP (LP-1837) y para un nivel intermedio (artificial N° 5), 1280 ± 90 años ar AP (LP- 1864). En los niveles superiores (artificiales N° 1 y N° 2), la presencia de huesos y heces de Ovis aries indican su referencia cronológica al segmento final del siglo XIX y principios del XX. Los restos de T. barrerae (números de lote CNP-E 259 a CNP-E 266) están presentes en los artificiales N° 1 a N° 8 inclusive.
Los otros dos yacimientos se encuentran en las proximidades de la villa Dique Florentino Ameghino y corresponden a rellenos de cuevas generadas por desplazamientos de grandes bloques en las ignimbritas jurásicas de la Formación Marifil. La denominada"Cueva de la Virgen" (43° 42' 10" S, 66° 27' 44" O, 89 m) se ubica sobre la banquina sur de la ruta provincial 31 en las afueras de la villa, mientras que la"Cueva Caolinera Dique Ameghino" (43° 40' 48" S, 66° 25' 57" O, 23 m) se emplaza sobre la margen derecha del río Chubut, unos 5 km aguas abajo de la presa. La "Cueva de la Virgen" tenía un importante relleno sedimentario que fue parcialmente alterado para el emplazamiento de un altar religioso. Allí se excavó una cuadrícula de 1 m de lado que alcanzó roca de caja luego de 11 niveles artificiales (aprox. 90 cm de espesor total). La secuencia comporta un término inferior limoso franco (aprox. 30 cm basales), casi desprovisto de clastos, que contrasta con una porción superior que descansa discordante, más pulverulent a y con abundantes fragmentos angulosos de la roca de caja. Una datación radiocarbónica del nivel basal, efectuada sobre coprolitos de roedores (artificial -170-180 cm) brindó 5470 ± 110 años ar AP (LP-1865); no se han efectuado fechados para los niveles suprayacentes pero existen claras evidencias de materiales post-conquista (restos de Ovis aries, papel periódico con fecha impresa 1962, objetos de metal). Seexhumaron restos referibles a T. barrerae (CNP-E 293- 1 A; figura 2.1) en el nivel acotado por la datación radiocarbónica; los niveles suprayacentes aún no han sido estudiados. En la "Cueva Caolinera Dique Ameghino", se excavó asimismo una cuadrícula de 1 m de lado de la que se extrajeron 15 niveles artificiales de 10 cm de espesor cada uno, alcanzando el basal una profundidad de 165 cm sobre la roca de caja. La secuencia está constituida por limos con intercalaciones de lentes carbonosas. Sendas dataciones radiocarbónicas sobre carbón vegetal brindaron, para la parte inferior (artificial N° 12), 3050 ± 70 años ar AP (LP-1780) y para el sector medio (artificial N° 6), 1230 ± 60 ar AP (LP-1817). Restos referibles a T. barrerae (CNP-E 292, CNP-E 324, CNP-E 325 y CNP-E 326) aparecen en los niveles artificiales N° 4, N° 6, N° 7 y N° 14.

Figura 2. Tympanoctomys barrerae (Holoceno medio-tardío, Chubut, Argentina) 1, CNP-E 293-1 A, paladar incompleto en vista ventral ("Cueva de la Virgen", cercanías del Dique Ameghino, artificial -170-180); 2, CNP-E 264-19 A, hemimandíbula izquierda en vista labial ("Perfil Los Altares", cercanías de Los Altares, artificial N°6). Escala = 5 mm / Tympanoctomys barrerae (Middle- Late Holocene, Chubut, Argentina) 1, CNP-E 293-1 A, incomplete palate in ventral view ("Cueva de la Virgen", near Dique Ameghino, artificial level -170-180); 2, CNP-E 264-19 A, left mandible in labial view ("Perfil Los Altares", near Los Altares, artificial N°6). Scale = 5 mm.
En los tres depósitos, los restos de T. barrerae se registran acompañados de otros muy abundantes referibles a roedores sigmodontinos y otros microvertebrados. Tafonómicamente, estas acumulaciones indican un origen mayoritario en la acción depredadora de búhos y lechuzas y pueden, de esta manera, ser consideradas como indicadores de las asociaciones de micromamíferos presentes localmente en un radio de 5 km en torno al sitio de hallazgo (Pardiñas, 1999).
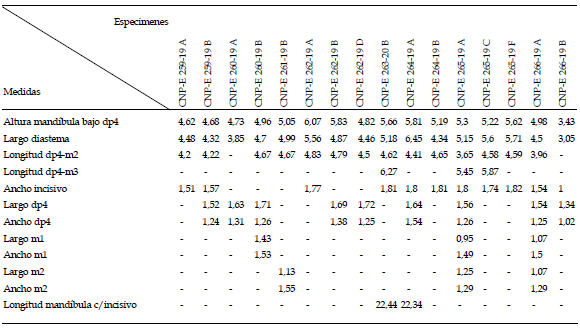
Descripción. Los especimenes CNP-E 293-1 A y CNP-E 264-19 A son aquellos más completos (figura 2), entre los varios recuperados y se emplean aquí como representativos de los rasgos reseñados. El CNPE 293-1 A (figura 2.1) es un paladar, roto anteriormente a nivel de la porción posterior de los forámenes incisivos, que conserva los PM4 pero no los molares. El espacio interdentario a nivel del paladar es ligeramente mayor entre los M3, aspecto vinculado con la divergencia posterior de las series dentarias; el borde anterior de la fosa mesopterigoidea se ubica a la altura de la cara anterior del M2. Sólo se han preservado las raíces maxilares de los arcos cigomáticos, robustos y expandidos en sentido dorso-ventral, que ostentan sendos tubérculos masetéricos bien definidos. Los PM4 son bilobulados y comprimidos anteroposteriormente, con una leve inclinación labial. Los alvéolos del M1 y M2 son cuadrangulares; el del M3 está comparativamente reducido en tamaño y ostenta una morfología de perímetro subtriangular. El CNP-E 264-19 A (figura 2.2) es una hemimandíbula izquierda que conserva el incisivo y el pm4 pero carece de los m1-3. El ramus es robusto y corto, con una región sinfisiaria anteriormente expandida y un proceso genal bien desarrollado y visible en norma labial. El incisivo es ancho en sentido transverso, con borde cortante ortogonal y esmalte de color ocráceo; su recorrido es semicircular y nace sobre la base del proceso coronoides, conformando una proyección capsular poco conspicua. El diastema está medianamente excavado. La muesca para la inserción del tendón del músculo masetero medial anterior está bien marcada y comienza a la altura de la mitad del pm4. La cresta masetérica, bien desarrollada, presenta un borde filoso y se continua en un proceso angular filiforme, más posteriormente saliente que el cóndilo. La escotadura lunar es conspicua y con forma de C. El cóndilo es claramente superior respecto al plano alveolar y posee un destacado proceso postcondiloideo. El pm4 es bilobulado con su extremo anterior subtriangular. Los alvéolos de los m2-m3 son cuadrangulares; aquel del m3 está fuertemente reducido. Las principales medidas calculadas se brindan en la tabla 1.
Tabla 1. Medidas de mandíbulas seleccionadas de Tympanoctomys barrerae del Holoceno de la provincia del Chubut, Argentina / measurements of selected mandibles of Tympanoctomys barrerae from the Holocene of the Chubut province, Argentina.
Extinción local de Tympanoctomys
Los datos paleontológicos aquí brindados indican la existencia de poblaciones de T. barrerae establecidas en el valle medio e inferior del río Chubut desde, al menos, 5 ka ("Cueva de la Virgen") hasta muy cercanos ("Cueva Caolinera Dique Ameghino") o directamente post-introducción del ganado ovino (finales del siglo XIX y principios del XX; "Perfil Los Altares"). Si bien no existen registros sobre poblaciones actuales de T. barrerae en la región del río Chubut, no menos cierto es que el conocimiento de las asociaciones de micromamíferos patagónicos es aún incipiente (Pardiñas et al., 2003). Con el objetivo de subsanar, al menos parcialmente, esta situación, se estudiaron 209 muestras actuales de egagrópilas generadas por aves rapaces para la región de estudio, que involucran cerca de 30000 individuos referibles a 28 especies de mamíferos pequeños (D. Udrizar Sauthier, datos no publicados). Los resultados alcanzados indican la inexistencia de poblaciones vivientes de T. barrerae sobre el curso del río Chubut y sectores colindantes. Adicionalmente, se estima que la extinción local de este roedor -involucrando unos 150 km en línea recta entre las localidades fosilíferas más extremas- se habría producido en momentos muy recientes (< 100 años).
Establecer las causas y mecanismos del proceso de extinción local de T. barrerae en Patagonia central requiere de una mayor cantidad de datos y de un análisis más profundo. Ciertos aspectos de T. barrerae tales como su modo de vida fosorial, distribución saltuaria, bajos números poblacionales y restricción de hábitat, hacen de sus poblaciones y ambientes ocupados, sistemas frágiles y altamente susceptibles a procesos exógenos (Ojeda et al., 1996; Mares et al., 1997). Existe amplia información -aunque científicamente poco documentada- sobre los efectos adversos generados por el pisoteo ovino/caprino en los roedores cavadores (eliminación de poblaciones de Ctenomys; cf. Osgood, 1943). Las cargas de ganado que se impusieron durante la primera mitad del siglo XX en muchos sectores patagónicos, aprovechando lo prístino del ambiente (cf. Aguado, 2005[M1]), derivaron en situaciones catastróficas de compactación del suelo y desertificación severa (e.g., Del Valle et al., 1998; Aagesen, 2000). Estos elementos, aún considerados en forma tentativa, señalan al factor antrópico como el principal responsable indirecto de la extinción de Tympanoctomys en la zona de estudio.
La presencia de poblaciones de T. barrerae en Patagonia central durante el Holoceno medio-tardío, a 630 km en línea recta del registro actual más cercano (38° 10' S, 68° 53' O, Salar de Añelo, Neuquén, Ojeda et al., 2007; figura 1), indica una paleodistribución extendida para este roedor. Justamente, Verzi et al. (2002: 156) destacaron que "...the present patchy distribution of Tympanoctomys may be considered a relict, and the peculiar habitats occupied by this rodents probably represent interglacial refuges...". Existen numerosas cuencas salinas en Patagonia, como los salares que diagonalmente jalonan el norte de la provincia de Río Negro, completamente inexploradas desde un punto de vista mastozoológico y que podrían albergar poblaciones relictuales de este particular roedor.
Agradecimientos
W. Udrizar Sauthier, M. Nabte, A. Andrade, S. Zamora, F. Cremonte, C. Iglesias y la Comuna del Dique Florentino Ameghino colaboraron en los trabajos de campo y aspectos logísticos. La Secretaría de Cultura del Chubut, especialmente en la figura de P. Castro, posibilitó la obtención de permisos. A. Ojeda envió fotografías de T. barrerae y bibliografía. Esta contribución fue solventada con fondos del PICT 32405 (Agencia Nacional de Promoción Científica y Tecnológica) y Consejo Nacional de Investigaciones Científicas y Técnicas. M. Gallardo y un revisor anónimo aportaron valiosos comentarios. A las personas e instituciones mencionadas, el reconocimiento de los autores.
Bibliografía
1. Aagesen, D. 2000. Crisis and conservation at the end of the world: sheep ranching in Argentine Patagonia. Enviromental Conservation 27: 208-215. [ Links ]
2. Aguado, A. 2005[M2]. La colonización del oeste de la Patagonia central. Departamento Río Senguer, Chubut. 1890-1919. Fondo Editorial provincial, Gobierno del Chubut. 175 pp. [ Links ]
3. Ameghino, F. 1889. Contribución al conocimiento de los mamíferos fósiles de la República Argentina. Actas de la Academia Nacional de Ciencias (Córdoba) 6: 1-1027. [ Links ]
4. Del Valle, H.F., Elissalde, N.O., Gagliardini, D.A. y Milovich, J. 1998. Status of desertification in the Patagonian Region: Assessment and mapping from satellite imagery. Arid Soil Research and Rehabilitation 12: 95-122. [ Links ]
5. De Santis, L.J.M., Roig, V.G. y Justo, E.R. 1991. La anatomía cráneo- dentaria de Tympanoctomys barrerae (Lawrence). Comparación con Octomys mimax y consideraciones acerca de su estado taxonómico (Rodentia: Octodontidade). Neotrópica 37: 113-122. [ Links ]
6. Diaz, G.B., Ojeda, R.A., Gallardo, M.H. y Giannoni, S.M. 2000. Tympanoctomys barrerae. Mammalian Species 646: 1-4. American Society of Mammalogists. [ Links ]
7. Gallardo, M.H., González, C.A. y Cebrián, I. 2006. Molecular cytogenetics and allotetraploidy in the red vizcacha rat, Tympanoctomys barrerae. Genomics 88: 214-221. [ Links ]
8. Gallardo, M.H., Bickham, J.W., Honeycutt, R.L., Ojeda, R.A. y Kohler, N. 1999. Discovery of tetraploid in a mammal. Nature 401: 341. [ Links ]
9. Lawrence, B. 1941. A new species of Octomys from Argentina. Proceedings of the New England Zoology Club 18: 43-46. [ Links ]
10. Mares, M.A., Braun, J.K. y Channell, R. 1997. Ecological observations on the octodontid rodent, Tympanoctomys barrerae, in Argentina. The Southwestern Naturalist 42: 488-504. [ Links ]
11. Ojeda, A.A., Gallardo, M.H., Mondaca, F. y Ojeda, R.A. 2007. Nuevos registros de Tympanoctomys barrerae (Rodentia, Octodontidae). Mastozoología Neotropical 14: 267-270. [ Links ]
12. Ojeda, R.A., Gonnet, J., Borghi, C.E., Giannoni, S., Diaz, G.B. y Braun, J.K. 1996. Ecological observations of the red vizcacha rat Tympanoctomys barrerae in two desert habitats of Argentina. Mastozoología Neotropical 3: 183-191. [ Links ]
13. Ojeda, R.A., Borghi, C.E., Diaz, G.B., Giannoni, S.M., Mares, M.A. y Braun, J.K. 1999. Evolutionary convergence of the highly adapted desert rodent Tympanoctomys barrerae (Octodontidae). Journal of Arid Environments 41: 443-452. [ Links ]
14. Osgood, W.H. 1943. The mammals of Chile. Field Museum of Natural History, Zoological Series 30: 1-268. [ Links ]
15. Pardiñas, U.F.J. 1999. [Los roedores muroideos del Pleistoceno Tardío- Holoceno en la región pampeana (sector este) y Patagonia (República Argentina): aspectos taxonómicos, importancia bioestratigráfica y significación paleoambiental. Tesis doctoral, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Argentina, 283 pp. Inédita. [ Links ]].
16. Pardiñas, U.F.J., Teta, P., Cirignoli, S. y Podestá, D.H. 2003. Micromamíferos (Didelphimorphia y Rodentia) de norpatagonia extra andina, Argentina: taxonomía alfa y biogeografía. Mastozoología Neotropical 10: 69-113. [ Links ]
17. Svartman, M., Stone, G. y Stanyon, R. 2005. Molecular cytogenetics discards polyploidy in mammals. Genomics 85: 425-430. [ Links ]
18. Verzi, D.H., Tonni, E.P., Scaglia, O.A. y San Cristóbal, J.O. 2002. The fossil record of the desert-adapted South American rodent Tympanoctomys (Rodentia, Octodontidae). Paleoenvironmental and biogeographic significance. Palaeogeography, Palaeoclimatology, Palaeoecology 179: 149-158. [ Links ]
19. Yepes, J. 1942. Zoogeografía de los roedores octodóntidos de Argentina y descripción de un nuevo género. Revista Argentina de Zoogeografía 2: 69-81. [ Links ]
Recibido: 19 de marzo de 2008.
Aceptado: 1 de octubre de 2008.

