










Services on Demand
Journal

Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAmeghiniana
On-line version ISSN 1851-8044
Ameghiniana vol.46 no.2 Buenos Aires Apr./June 2009
ARTÍCULOS ORIGINALES
Nanofósiles calcáreos paleógenos del sudeste de la provincia de Santa Cruz, Patagonia, Argentina
Juan Pablo Pérez Panera1
1CONICET. División Paleozoología Invertebrados, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Paseo del Bosque s / n, 1900, La Plata, Argentina. perezpanera@ciudad.com.ar
Resumen. Se realizó un estudio cuantitativo de los nanofósiles calcáreos paleógenos del subsuelo de la localidad Cerro Redondo, al sureste de la provincia de Santa Cruz, Cuenca Austral, Argentina. Se reconoció el Paleoceno temprano y Paleoceno medio, Eoceno temprano alto y Eoceno medio. Para establecer el nivel base del Paleógeno se identificó el intervalo Cretácico - Paleógeno, que evidencia una discontinuidad por la ausencia del Maastrichtiano medio y tardío y la base del Paleoceno. Se infiere que la depositación durante el Paleoceno y el Eoceno medio ocurrió en condiciones de mar alto y aguas superficiales frías. Durante el Eoceno medio alto la depositación se produjo en condiciones de aguas someras cercanas a la línea de costa.
Abstract. Paleogene calcareous nannofossils from the southeastern Santa Cruz Province, Patagonia, Argentina. This study presents the results of a quantitative analysis of calcareous nannofossils recovered from subsurface samples in drilled wells at the locality of Cerro Redondo, southeastern Santa Cruz Province, Austral Basin of Argentina. Index species identified in the succession allow to recognize stratigraphic levels in the early and middle Paleocene, the late early Eocene and middle Eocene. The basal level of the Paleogene was established based on the Cretaceous-Paleogene interval, which includes an unconformity from the middle Maastrichtian to the lowest Paleocene. It has been inferred high sea and superficial cold water condition for the Paleocene and middle Eocene and shoreface condition during the late middle Eocene.
Key words. Calcareous nannofossils; Paleogene; Austral Basin; Argentina.
Palabras clave. Nanofósiles calcáreos; Paleógeno; Cuenca Austral; Argentina.
Introducción
La Cuenca Austral se extiende en el extremo sur del continente sudamericano. Abarca casi la totalidad de la provincia argentina de Santa Cruz, la provincia chilena de Magallanes, la Isla Grande de Tierra del Fuego y una porción de la plataforma continental argentina. Hacia el oeste está delimitada por la Cordillera Patagónica, al sur por la Cordillera Fueguina, al este por el Alto del Río Chico, en la plataforma continental y al norte, por el Macizo del Deseado. (figura 1) El relleno sedimentario de esta cuenca se inicia en el Jurásico tardío y culmina en el Neógeno.

Figura 1. Mapa del sur de Argentina mostrando la ubicación de la localidad de Cerro Redondo, provincia de Santa Cruz. / map of southern Argentina showing the studied area Cerro Redondo, Santa Cruz province.
Durante el Paléogeno y Neógeno se produjeron importantes ingresiones marinas atlánticas que, en una sucesión de ciclos regresivos - transgresivos, originaron una serie de depósitos marinos con claras diferencias faciales (Malumián, 1999; 2002). Esto permitió agrupar los depósitos en distintas unidades formacionales, tanto en subsuelo como en superficie, pero debido a la gran extensión de la cuenca y sus diversos encuadres tectónicos, se han definido numerosas formaciones con sus equivalentes en los diferentes sectores de la cuenca, lo que dificulta en gran medida una clara correlación cronoestratigráfica regional.
En el presente trabajo se estudiaron cuantitativamente las asociaciones de nanofósiles calcáreos recuperados en muestras de recorte de perforación lavados de un pozo exploratorio en la localidad de Cerro Redondo, ubicado en el sector oriental de la Cuenca Austral, al sudeste de la provincia de Santa Cruz, Argentina (figura 1). El objetivo ha sido identificar las especies de nanofósiles presentes y a partir de ellos interpretar la edad de las sedimentitas, estableciendo equivalencias con los modelos bioestratigráficos globales para este grupo. Este trabajo es un aporte nuevo para la comprensión del desarrollo histórico de la cuenca y futuros estudios basados en nanofósiles calcáreos brindarán una herramienta importante en el conocimiento de la cronoestratigrafía regional.
Los nanofósiles calcáreos son restos de organismos unicelulares fotosintetizadores, planctónicos, con un registro que abarca desde el Triásico tardío hasta la actualidad. Han tenido una historia evolutiva de grandes cambios a partir del Jurásico medio y en su mayoría las especies son cosmopolitas. Estas características los convierten en fósiles guía muy importantes en secuencias marinas.
Antecedentes
La Cuenca Austral es un área de interés petrolero en la que se vienen realizando estudios bioestratigráficos tanto en subsuelo como en superficie, por lo que existen numerosos informes inéditos de las empresas petroleras que los llevan a cabo.
En cuanto a los trabajos de consulta pública relativos a microfósiles en sedimentitas de edad paleógena, existe una vasta bibliografía, principalmente basada en el estudio de los foraminíferos. El trabajo pionero, realizado sobre muestras de subsuelo, es el de Malumián (1968) donde detalla los foraminíferos hallados para el Cretácico superior, Paléogeno y Neógeno del pozo SC - 3, ubicado en el centro sur de la provincia de Santa Cruz. Otra referencia temprana es la de Malumián y Masiuk (1973) donde se presentan las asociaciones de foraminíferos reconocidas en la cuenca para el Cretácico, Paléogeno y Neógeno. Malumián y Náñez (1996) describen las asociaciones de foraminíferos y nanofósiles calcáreos desde el Jurásico al Cuaternario en el área de la plataforma submarina, sin embargo los nanofósiles calcáreos son tratados escuetamente. Malumián y Náñez (2002) en un importante trabajo de revisión, han resumido el conocimiento de los foraminíferos para la Cuenca Austral y cuencas aledañas haciendo referencia a las condiciones paleoambientales en los diferentes sectores de la cuenca y a los detalles bioestratigráficos. Olivero et al. (2003) estudiaron la secuencia sedimentaria, los foraminíferos, palinomorfos y amonoideos en el sur de la Isla Grande de Tierra del fuego identificando el límite Maastrichtiano-Paleoceno. Proponen además un ambiente de depositación marino profundo bajo condiciones disóxicas a anóxicas para el Cretácico tardío, que se va somerizando en el Paleoceno, culminando la secuencia sedimentaria con abanicos deltáicos y aportes terrígenos.
Los trabajos referidos principalmente a los nanofósiles calcáreos para el Paleógeno son más escasos. Los cuatro más importantes son el de Mostajo (1991), Concheyro (1991), Ronchi y Angelozzi (1994), y Concheyro y Angelozzi (2002). El más próxima al área de estudio que aquí nos ocupa es el de Mostajo (1991). Ella estudia la secuencia de sedimentitas de la perforación Las Violetas 3, situada en la costa noreste de la Isla Grande de Tierra del Fuego. Allí reconoce treinta y dos especies y en base a ellas establece una edad Cretácico superior a Oligoceno para la secuencia. Además advierte la ausencia de asociaciones de edad daniana e interpreta condiciones de aguas superficialmente frías durante la depositación en una ambiente de mar alto con pulsos regresivos hacia el tope de la secuencia. Concheyro (1991) describe sistemáticamente veinticinco especies del Eoceno para la Formación Man-Aike, recuperadas en el sondeo YCF SEC 7 (ENADISMA) al sureste del lago Cardiel. Ronchi y Angelozzi (1994) correlacionan dos sondeos al oeste de la Cuenca Austral en base a las asociaciones de foraminíferos y nanofósiles calcáreos. Concheyro y Angelozzi (2002) sintetizan para el área central y occidental de la cuenca el conocimiento del grupo, basado principálmente en informes inéditos, proponiendo una bioestratigrafía generalizada y describiendo las asociaciones de nanofósiles calcáreos reconocidas desde el Cretácico al Neógeno temprano.
Marco geológico
La Cuenca Austral se origina en relación a la ruptura de Gondwana, que da inicio a la generación de una cuenca de tipo marginal durante el Jurásico superior. Comienza con una etapa de rift durante la separación de América del sur y África asociado a la apertura del mar de Wedell. Para el Cretácico inferior la apertura de la Cuenca Austral había terminado y se produce una etapa de hundimiento termal. Posteriormente y mancomunado a los movimientos patagónicos del Cretácico medio, la fase peruana del Cretácico superior y los movimientos ándicos terciarios, se instala en la cuenca una etapa de antepaís. Hacia el sector occidental de la Cuenca Austral se depositan sedimentitas de cuenca de antepaís y en el oriental, de rampas de antepaís (Robbiano et al., 1996). En este último sector, se sobreimpone una subsidencia térmica y tectónica haciendo migrar el depocentro de la cuenca al sureste. Debido a esto las secuencias terciarias están más desarrolladas en el sector oriental.
Durante el Paleógeno se registran dos transgresiones marinas, la primera que comienza en el Maastrichtiano y la segunda en el Eoceno medio (Malumián, 1999). Las sedimentitas que corresponden a estos eventos se conocen en subsuelo en el sector oriental como Formación Campo Bola (Russo et al. 1980) (pars) y Formación Man Aike (Furque 1973) de edad Paleoceno y Eoceno respectivamente.
Materiales y métodos
El material corresponde a 16 muestras de recorte de perforación lavadas que representan 25 metros de intervalo cada una. Provienen de un pozo de exploración cedido por la empresa Petrobras Energía S.A. y comprenden 375 metros, del intervalo entre los 725 y 1.100 metros bajo boca de pozo (mbbp). Éstas se procesaron según el método de decantación por gravedad (Moshkovitz y Ehrlich, 1976; Hay, 1977) y cada 3 muestras se realizaron "muestras control" con la técnica de smear slide (Bown y Young, 1999). Las muestras realizadas con la técnica de decantación por gravedad dieron mejores resultados, especialmente en la concentración de los especímenes.
Las muestras se estudiaron con un microscopio óptico con contraste de fases y luz polarizada a un aumento de 1500 X. En cada muestra se contabilizaron al menos trescientos individuos, a fin de obtener datos que pudieran ser analizados con técnicas estadísticas. Este esfuerzo de muestreo representa una confianza del 95 % de hallar un individuo con una abundancia específica del 1 % (Thierstein et al., 1977). En muestras con una alta diversidad específica se contabilizaron más individuos, a veces mediante las "muestras control". En dos de las profundidades estudiadas (725-750 mbbp y 850-875 mbbp; cuadro 1) no se logró alcanzar el mínimo de trescientos individuos. Para cuantificar la abundancia relativa se utilizó el criterio de Stoico y Baldanza (1995) y para el estado de preservación el de Roth y Thierstein (1972).
Cuadro 1. Tabla de distribución de especies e índices de diversidad / Chart of species distribution and diversity index. Referencias / References: Preservación / Preservation según / according to Roth y Thierstein, 1972; E-3: Disolución importante / important dissolution, E-2: Disolución / dissolution; E-1: Disolución moderada / moderate dissolution; X: Muy buena / very good; O-1: Sobrecrecimiento moderado / moderate overgrown; O-2: Sobrecrecimiento / overgrown; O-3: Sobrecrecimiento importante / important overgrown. N/A: No aplicable / not applicable. Abundancia relativa / relative abundance según / according to Stoico y Baldanza, 1995; A: Abundante / abundant (más de un individuo por campo / more than one individual per field); C: Común / common (un individuo en uno a diez campos / one individual in one to ten fields); P: Pocos / few (un individuo en diez a treinta campos / one individual in ten to thirty fields); R: Raros / rare (un individuo en treinta a cien campos / one individual in thirty to one hundred fields). * Indica cocósfera / indicates coccosphere.
Una vez obtenidos todos los datos correspondientes a cada profundidad, se ordenaron en un gráfico de distribución: profundidad / especie (cuadro 1), donde consta la referencia al estado de preservación, la abundancia relativa, el número total de individuos contabilizados, el número total de especies y el índice de biodiversidad de Shannon y Wiener (1963).
La interpretación bioestratigráfica está remitida al modelo zonal de Martini (1971) y se grafica en la figura 2. El tiempo en millones de años al que corresponden las zonas de nanofósiles calcáreos está calibrado según Berggren et al. (1995). Todas las especies identificadas están referidas a las citas de Wise y Mostajo (1983), Perch-Nielsen (1985a, 1985b) y Bown (1999, 2005).
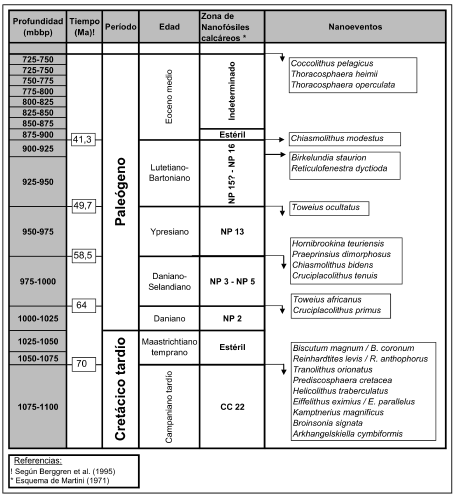
Figura 2. Bioestratigrafía propuesta en base a los nanofósiles calcáreos / proposed biostratigraphy based on calcareous nannofossils.
El material ilustrado (figura 3) queda depositado en la colección del Departamento de Ciencias Geológicas de la Facultad de Ciencias Exactas y Naturales de la Universidad de Buenos Aires, bajo la sigla BAFC-Np (nanoplancton calcáreo) y los números 2751 al 2756.
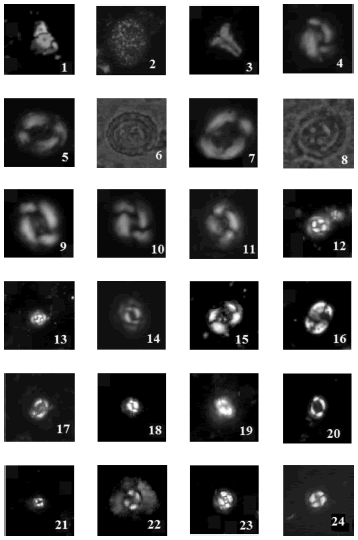
Figura 3. 1, Thoracosphaera heimii (Lohmann, 1919) Kamptner, 1941 (Luz polarizada / polarized light) 725-750 mbbp. Tamaño / dimension 7 μm. BAFC-Np 2751; 2, Thoracosphaera operculata Bramlette y Martini, 1964 (Luz polarizada / polarized light) 725-750 mbbp. Tamaño / dimension 12 μm. BAFC-Np 2751; 3, Zygrablithus bijugatus (Deflandre en Deflandre y Fert, 1954) Deflandre, 1959 (Luz polarizada / polarized light) 900-925 mbbp. Tamaño / dimension 8 μm. BAFC-Np 2752; 4, Reticulofenestra daviesii (Haq, 1968) Haq, 1971 (Luz polarizada / polarized light) 900-925 mbbp. Tamaño / dimension 8 μm. BAFC-Np 2752; 5, Chiasmolithus grandis (Bramlette y Riedel, 1954) Radomski, 1968 (Luz polarizada / polarized light) 900-925 mbbp. Tamaño / dimension 14 μm. BAFC-Np 2752; 6, Chiasmolithus grandis (Bramlette y Riedel, 1954) Radomski, 1968 (Luz transmitida / transmitted light) 900-925 mbbp. Tamaño / dimension 14 μm. BAFC-Np 2752; 7, Chiasmolithus modestus Perch-Nielsen, 1971 (Luz polarizada / polarized light) 900-925 mbbp. Tamaño / dimension 10 μm. BAFCNp 2752; 8, Chiasmolithus modestus Perch-Nielsen, 1971 (Luz transmitida / transmitted light) 900-925 mbbp. Tamaño / dimension 10 μm. BAFC-Np 2752; 9, Reticulofenestra bisecta ssp. filewiczii Wise y Wiegand en Wise, 1983 (Luz polarizada / polarized light) 900-925 mbbp. Tamaño / dimension 9 μm. BAFC-Np 2752; 10, Reticulofenestra bisecta (Hay et al., 1966) Roth, 1970 ssp. bisecta Wise y Wiegand en Wise, 1983 (Luz polarizada / polarized light) 900-925 mbbp. Tamaño / dimension 8 μm. BAFC-Np 2752; 11, Birkelundia staurion (Bramlette y Sullivan, 1961) Perch-Nielsen, 1971 (Luz polarizada / polarized light) 925-950 mbbp. Tamaño / dimension 7 μm. BAFC-Np 2753; 12, Toweius callosus Perch- Nielsen, 1971 (Luz polarizada / polarized light) 950-975 mbbp. Tamaño / dimension 6 μm. BAFC-Np 2754; 13, Toweius rotundus Perch-Nielsen en Perch-Nielsen et al., 1978 (Luz polarizada / polarized light) 950-975 mbbp. Tamaño / dimension 4 μm.BAFC-Np 2754; 14, Toweius occultatus (Locker, 1967) Perch-Nielsen, 1971 (Luz polarizada / polarized light) 950-975 mbbp. Tamaño / dimension 4 μm. BAFC-Np 2754; 15, Chiasmolithus consuetus (Bramlette y Sullivan, 1961) Hay y Mohler, 1967 (Luz polarizada / polarized light) 950-975 mbbp. Tamaño / dimension 8 μm. BAFC-Np 2754; 16, Pontosphaera pulchra (Deflandre en Deflandre y Fert, 1954) Romein, 1979 (Luz polarizada / polarized light) 950-975 mbbp. Tamaño / dimension 7 μm. BAFC-Np 2754; 17, Hornibrookina teuriensis Edwards, 1973 (Luz polarizada / polarized light) 975-1000 mbbp. Tamaño / dimension 8 μm. BAFC-Np 2755; 18, Praeprinsius dimorphosus (Perch-Nielsen, 1969) Varol y Jakubowski, 1989 (Luz polarizada / polarized light) 975-1000 mbbp. Tamaño / dimension 3 μm. BAFC-Np 2755; 19, Praeprinsius dimorphosus (Perch- Nielsen, 1969) Varol y Jakubowski, 1989 (Luz polarizada / polarized light) 975-1000 mbbp. Tamaño / dimension 3 μm. BAFC-Np 2755; 20, Chiasmolithus bidens (Bramlette y Sullivan, 1961) Hay y Molher, 1967 (Luz polarizada / polarized light) 975-1000 mbbp. Tamaño / dimension 10 μm. BAFC-Np 2755; 21, Toweius africanus (Perch-Nielsen, 1980) Perch-Nielsen, 1984 (Luz polarizada / polarized light) 1000-1025 mbbp. Tamaño / dimension 2.5 μm. BAFC-Np 2756; 22, Markalius inversus (Deflandre en Deflandre y Fert, 1954) Bramlette y Martini, 1964 (Luz polarizada / polarized light) 1000-1025 mbbp. Tamaño / dimension 8 μm. BAFCNp 2756; 23, Cruciplacolithus primus Perch-Nielsen, 1977 (Luz polarizada / polarized light) 1000-1025 mbbp. Tamaño / dimension 6 μm. BAFC-Np 2756; 24, Toweius cf. T. sp2 Bown, 2005 (Luz polarizada / polarized light) 1000-1025 mbbp. Tamaño / dimension 4 μm. BAFCNp 2756.
Interpretación bioestratigráfica
De acuerdo a los resultados obtenidos se reconocieron 5 tramos: entre 725 y 875 mbbp, entre 875 y 900 mbbp, entre 900 y 975 mbbp, entre 975 y 1025 mbbp, entre 1025 y 1150 mbbp. Se individualizan y desarrollan por separado para una mejor comprensión. El criterio de segregación de los tramos responde a la acentuación de los nanoeventos más importantes, a la identificación de posibles discontinuidades y al aislamiento de secciones nanofosilíferamente estériles.
1er. Tramo: 725 - 875 mbbp.
Las profundidades aquí comprendidas, que corresponden a las primeras muestras estudiadas en el sentido normal de la perforación, presentan una baja riqueza específica pero una muy buena preservación (cuadro 1). Toda la asociación identificada en este tramo está dominada por Thoracosphaera operculata Bramlette y Martini, 1964. Como especies acompañantes raras se han reconocido Reticulofenestra minutula (Gartner, 1967) Haq y Berggren, 1978; Reticulo-fenestra pseudoumbilica (Gartner, 1967) Gartner, 1969; Reticulofenestra haqii Backman, 1978 (que se consideran como contaminación desde rocas suprayacentes); Pontosphaera discopora Schiller, 1925; Thoracosphaera heimii (Lohmann, 1919) Kamptner, 1941; Reticulo-fenestra minuta Roth, 1970 y Coccolithus pelagicus (Wallich, 1877) Schiller, 1930.
De acuerdo con Perch-Nielsen (1985a), T. operculata no sobrevive más allá del Eoceno medio. Sin otros marcadores que precisen con mayor detalle la edad de la asociación, se la asigna al Eoceno medio alto.
2do. Tramo: 875 - 900 mbbp.
No se recuperaron nanofósiles.
3er. Tramo: 900 - 975 mbbp.
En las muestras correspondientes a este tramo se reconoció una asociación caracterizada por la presencia de abundantes reticulofenestras, junto a C. pelagicus y T. operculata, pero esta última en una relación de dominancia mucho menor que en el tramo suprayacente (725 - 875 mbbp, cuadro 1). Desde los 900 a los 950 mbbp las muestras presentan una muy buena preservación y abundancia. A partir de los 950 mbbp los individuos aparecen con menor abundancia y con signos de disolución, aunque se observó uno de los valores más altos de biodiversidad (véase cuadro 1).
En estos 75 metros se recuperaron buenos marcadores bioestratigráficos. A los 900-925 mbbp se observó la primera ocurrencia, según el sentido normal de la perforación, de Chiasmolithus modestus Perch - Nielsen, 1971; principal marcador que permite asignar este nivel a la zona NP16 (Eoceno medio, Lutetiano-Bartoniano). Integrando la asociación se recuperaron nanofósiles característicos de esta edad como Chiasmolihus grandis (Bramlette y Riedel, 1954) Radomski, 1968; Zygrhablithus bijugatus (Deflandre en Deflandre y Fert, 1954) Deflandre, 1959; Reticulofenestra bisecta (Hay et al., 1966) Roth, 1970 Reticulofenestra bisecta ssp. filewiczii Wise y Wiegand en Wise, 1983; Reticulofenestra daviesii (Haq, 1968) Haq, 1971 y Helicosphaera seminulum Bramlette y Sullivan, 1961.
A los 925-950 mbbp se reconocieron como especies marcadoras a Reticulofenestra dictyoda (Deflandre en Deflandre y Fert, 1954) Stradner en Stradner y Edwards, 1968 y Birkelundia staurion (Bramlette y Sullivan, 1961) Perch-Nielsen, 1971 que se registran por primera vez en esta profundidad. De acuerdo a sus biocrones (zonas NP13-NP16 y NP15-NP16, respectivamente), la posición estratigráfica del nivel en consideración y la ausencia de Ch. modestus, especie que es hallada en el nivel superior y sólo ocurre en la zona NP16, se asigna este nivel a la zona NP15 con dudas. Por este motivo en la figura 2 se representan estos dos últimos niveles (900-925 y 925-950) juntos y referidos a las zonas NP 15? - NP16. Si bien estos nanoeventos concuerdan con los modelos bioestratigráficos globales, R. dictyoda ha sido recuperada en otras localidades de la Cuenca Austral en niveles más jóvenes (comunicación personal Angelozzi, 2006). Esto pondría en duda su importancia como marcador para el Atlántico Sur, y la asignación bioestratigráfica de este nivel se basa en B. staurion y los nanofósiles acompañantes.
A los 950-975 mbbp se reconoce la mayor biodiversidad entre los tres niveles ahora considerados y, en base a las primeras apariciones según el sentido normal de la perforación de Toweius callosus Perch- Nielsen, 1971 y Toweius occultatus (Locker, 1967) Perch-Nielsen, 1971; se asigna este intervalo del tramo a la zona NP13.
En su conjunto, estos tres niveles definen las profundidades comprendidas entre los 900 y los 975 mbbp como Eoceno temprano alto a Eoceno medio. La zona NP14 no ha podido ser reconocida, y esta ausencia podría estar indicando una discontinuidad a los 950 mbbp.
4to. Tramo: 975 - 1025 mbbp.
En este tramo se observó una asociación dominada por especies del género Toweius Hay y Mohler, 1967. A los 975-1000 mbbp se registran las primeras apariciones, según el sentido normal de la perforación, de Hornibrookina teuriensis Edwards, 1973; Preaprinsius dimorphosus (Perch-Nielsen, 1969) Varol y Jakubowski, 1989; Chiasmolithus bidens (Bramlette y Sullivan, 1961) Hay y Mohler, 1967 y Cruciplacolithustenuis (Stradner, 1961) Hay y Mohler en Hay et al. 1967. En su conjunto todas estas especies marcadoras coinciden en una asignación para este nivel correspondiente a las zonas NP3 a NP5 (Varol, 1999).
A los 1000-1025 mbbp se identificaron los marcadores Toweius africanus (Perch - Nielsen, 1980) Perch -Nielsen, 1984; Markalius inversus (Deflandre en Deflandre y Fert, 1954) Bramlette y Martini, 1964 y Cruciplacolithus primus Perch-Nielsen, 1977. En base a ellas se asignó este nivel a la zona NP2. A pesar de los esfuerzos de muestreo no se lograron recuperar marcadores que indicaran la presencia de la zona NP1.
En consecuencia, desde los 975 hasta los 1025 mbbp la asociación nanofosilífera se incluye dentro del Daniano-Selandiano, zonas NP2-NP5, con ausencia de la base del Paleoceno, zona NP1 que involucra aproximadamente un millón de años (Berggren et al., 1995) (figura 2).
5to.Tramo: 1025 - 1150 mbbp.
Las muestras comprendidas entre los 1025 y 1075 mbbp resultaron estériles desde el punto de vista de los nanofósiles calcáreos, y entre los 1075 y 1150 mbbp se observa una asociación típicamente cretácica tardía, con muy buenos marcadores y una alta biodiversidad. Las especies más importantes desde el punto de vista bioestratigráfico son Biscutum magnum Wind y Wise en Wise y Wind, 1977; Biscutum coronum Wind y Wise en Wise y Wind, 1977; Reinhardtites levis Prins y Sissingh en Sissingh, 1977; Reinhardtites anthophorus (Deflandre, 1959) Perch- Nielsen, 1968; Tranolithus orionatus (Reinhardt, 1966a) Reinhardt, 1966b; Prediscosphaera cretacea (Arkhan-gelski, 1912) Gartner, 1968; Helicolithus trabeculatus (Górka, 1957) Verbeek, 1977; Eiffellithus eximius (Sto-ver, 1966) Perch-Nielsen, 1968; Eiffellithus parallelus Perch-Nielsen, 1973; Kamptnerius magnificus Deflan-dre, 1959; Broinsonia signata (Noël, 1969) Noël, 1970 y Arkhangelskiella cymbiformis Vekshina, 1959; esta última en baja proporción, y no de manera dominante como es común en las últimas zonas del Maastrichtiano (Burnett, 1999). De acuerdo a estos marcadores se asigna este nivel a la zona CC22 del esquema zonal de Sissingh (1977), del Campaniano tardío. Esta asociación de nanoplancton calcáreo se correspondería con la zona de Biscutum coronum que Wise (1983) propone para el Cretácico tardío de la plataforma de Malvinas.
Las muestras comprendidas entre los 1025 y 1075 mbbp, si bien resultaron estériles desde el punto de vista nanofosilífero, son portadoras de una asociación de foraminíferos (comunicación personal Ronchi, 2006) que corresponden a la "zona de foraminíferos arenáceos" de Malumián y Masiuk (1976), denominación informal de edad maastrichtiana temprana, lo que establecería a los 1025 mbbp el límite Cretácico-Paleógeno mediante una discontinuidad Maastrichtiano temprano-Daniano.
Discusión
Las transgresiones atlánticas que se produjeron en la Cuenca Austral durante el Paléogeno y Neógeno corresponden a cuatro ciclos mayores. Estos ciclos han sido identificados a partir de técnicas secuenciales (Calegari y Baldi, 1993; Calegari et al., 1993) y de discontinuidades reconocidas mediante el estudio de los foraminíferos (Malumián, 1999; Malumián y Náñez, 2002). Incluso Malumián (2002) relacionó estos ciclos transgresivos a los cambios eustáticos globales y los comparó con los datos del sur de Australia. El tramo aquí estudiado registra las dos primeras transgresiones mayores, las que se inician en el Maastrichtiano y en el Eoceno medio tardío.
El límite Cretácico-Paleógeno involucra una discordancia que Robbiano et al. (1996) atribuyeron a los movimientos ándicos iniciales. Considerando los resultados obtenidos en este trabajo, la discontinuidad que se observa a los 1025 mbbp, entre la asociación de foraminíferos arenáceos por debajo de esa profundidad y la asociación de nanofósiles danianos por encima, correspondería a un evento de no depositación y/o erosión del Maastrichtiano medio, Maastrichtiano tardío y la base del Paleoceno (zona NP1).
La asociación nanofosilífera recuperada entre los 975-1025 mbbp es de edad paleocena temprana a media, registrando el Daniano, excepto su base, y casi todo el Selandiano (zonas NP2 a NP5). Según Malumián (1999, 2002) estas edades corresponden a las sedimentitas de la Formación Cerro Dorotea en la zona de Río Turbio. Para el subsuelo de Santa Cruz el Daniano se supone condensado (Malumián et al., 1971; Malumián y Caramés, 1995; 1997) y contenido dentro de la Formación Campo Bola (pars). El espaciado muestreo de este estudio y la falta de información sedimentológica hacen imposible decidir si el Daniano aquí aparece condensado o no, pero se desarrollaría sólo en cincuenta metros (véase figura 2).
La asociación recuperada presenta una nanoflora fósil característica de altas latitudes (Chiasmolithus sp., Toweius sp., Hornibrookina teuriensis, Preaprinsius dimprphosus) y relativamente altos índices de biodiversidad, lo que supone un ambiente marino de plataforma con aguas frías. Mostajo (1991) infiere estas mismas condiciones en este sector de la cuenca y Concheyro y Angelozzi (2002) lo proponen para los sectores central y occidental durante el Paleoceno.
El índice de biodiversidad permite hacer comparaciones entre las diferentes muestras dando una idea aproximada de la relación de las diferentes asociaciones con el ambiente. Cerca de la costa, a profundidades menores de 40 metros (Perch-Nielsen, 1985b), la energía del ambiente es muy alta y los organismos planctónicos son arrastrados; en la zona oceánica de mar abierto la escasez de nutrientes limita el desarrollo de la comunidad fitoplanctónica. McIntyre y Bé (1967) y Okada y McIntyre (1977, 1979) han estudiado la distribución de especies actuales de cocolitofóridos en el océano Atlántico y encontraron que en mar abierto las asociaciones son oligoespecíficas de formas de tamaño pequeño, y que la mayor abundancia y diversidad se da en las áreas de plataforma, ya que son áreas ricas en nutrientes.
A los 975 mbbp se reconoce una discontinuidad importante que divide la asociación paleocena de una eocena temprana tardía a media. Esta discontinuidad se evidencia por la ausencia de sedimentitas que contengan asociaciones del Paleoceno tardío a Eoceno temprano y es reconocida en todo el sector medio y norte de la cuenca (Malumián y Náñez, 1996; Biddle et al., 1986; Concheyro y Angelozzi, 2002) e incluso en la Cuenca del Golfo de San Jorge (Heizler et al., 1998). Según Malumián (2002) esta discontinuidad responde a una regresión coincidente con un pico menor de magmatismo dentro de la Pa-tagonia, originando un ciclo erosivo en la plataforma.
Entre los 950 y los 975 mbbp las sedimentitas son asignadas a la zona NP 13, del Eoceno temprano tardío. La presencia de la zona NP13 se sostiene en la importante participación de Toweius occultatus, con cerca del 20 % de abundancia relativa en la asociación (cuadro 1), especie que en su rango total no supera dicha zona. A los 950 mbbp se supone una discontinuidad por ausencia de al menos la zona NP 14, ya que encima de esa profundidad y hasta los 900 mbbp se depositan sedimentitas que, de acuerdo a los nanofósiles que portan, se asignan a las zonas NP15 (con dudas) a NP16.
De acuerdo al estado de preservación y los índices de biodiversidad, se interpreta que estos niveles (900 - 975 mbbp, tercer tramo) fueron depositados en un ambiente de plataforma a cierta distancia de la línea de costa. La presencia de especies del género Chiasmolithus Hay et al. 1966 sugiere asimismo un ambiente de aguas superficiales relativamente frías (Mostajo, 1991; Concheyro y Angelozzi, 2002). Estas sedimentitas de edad Eoceno medio podrían estar indicando el inicio de la segunda transgresión atlántica.
La muestra que comprende los 875 a 900 mbbp resultó estéril y por encima de ella y hasta los 725 mbbp se desarrolla la asociación dominada por Thoracosphaera operculata. Como ya se mencionó, estos niveles se asignan al Eoceno medio sin mayor exactitud. La notoria escasez de cocolitos en este tramo supone un ambiente deposicional más restringido y cercano a la costa. Las thoracosphaeras actualmente son halladas en mar abierto, bajo condiciones marinas normales, pero se sabe que son oportunistas que alcanzan un buen desarrollo en ambientes estresantes donde no tienen competencia con otros organismos planctónicos calcáreos (Perch-Nielsen, 1985b; Höll et al., 1998). Estos organismos florecen en aguas oligotróficas de mares y océanos tropicales a subantárticos, tanto en ambientes oceánicos como en ambientes de plataforma interna (Vink et al., 2000; Zonneveld et al. 1999). A partir de las asociaciones de foraminíferos recuperadas para el Eoceno medio tardío en diferentes localidades de la Cuenca Austral, Malumián (1999) infiere también un ambiente de mar somero interno con comunicación oceánica restringida. Este evento podría corresponderse a la fase regresiva de la segunda transgresión atlántica.
Conclusiones
Entre los 1025 y 975 mbbp se reconocieron las zonas NP2 a NP5 del Paleoceno, correspondientes al Daniano y Selandiano. Desde los 975 hasta los 900 mbbp y mediante una discontinuidad se registra la zona NP 13 del Eoceno temprano tardío. A los 950 mbbp se reconoce otra discontinuidad sobre la que se desarrolla el Eoceno medio, zonas NP15 con dudas y NP16; y desde los 900 hasta los 725 mbbp la edad de las sedimentitas es eocena media tardía sin que pueda precisarse con mayor detalle bioestratigráfico.
La transgresión atlántica que se inicia en el Maastrichtiano temprano deposita sedimentitas marinas de un ambiente de plataforma normal y aguas superficiales frías. Las sedimentitas que portan la asociación de nanofósiles paleocenos (zonas NP2 a NP5) corresponden al evento de mar alto producto de esta transgresión. Por encima de ellas, y mediante una clara discontinuidad se iniciaría el segundo ciclo transgresivo depositando sedimentitas con una na-noflora fósil de edad Eocena temprana alta a Eocena media (zonas NP13, NP15 con dudas a NP16) que sugieren así mismo un ambiente de plataforma abierta y aguas superficiales frías. La fase regresiva de esta segunda transgresión atlántica estaría evidenciada por las sedimentitas suprayacentes portadoras de una asociación casi exclusiva de thoracosphaeras, sugiriendo un ambiente de depositación marino somero probablemente con mayor cercanía a la costa.
Agradecimientos
A la consultora Bioestratigráfica GEMA S.R.L. por permitir el uso de sus laboratorios y a Petrobras Energía S.A. por el material de estudio. Este trabajo fue realizado con fondos provistos por subsidios del CONICET (PIP 5613) y de la Universidad Nacional de La Plata (N 460).
Bibliografía
1. Arkhangelsky, A.D. 1912. Upper Cretaceous deposits of east European Russia. Materialien zur Geologie Russlands 25: 1-631. [ Links ]
2. Backman, J. 1978. Late Miocene-Early Pliocene nannofossil biochronology and biogeography in the Vera Basin, SE Spain. Stockholm Contributions in Geology 32 :93-114. [ Links ]
3. Berggren, W.A., Kent, D.V., Swisher III, C.C. y Aubry, M. 1995. A revised Cenozoic Geochronology and Chronostratigraphy. Geochronology Time Scales and Stratigraphic correlation SEPM Nº 54. [ Links ]
4. Biddle, K.T., Uliana, M.A., Mitchum Jr., R.M., Fitzgerald, M.G. y Wright, R.C. 1986. The Stratigraphic and structural evolution of the central and eastern Magallanes basin, southern South America. En: P.A. Allen y P. Homewoods (eds.), Foreland Basins. Special Publication International Association of Sedimentologists, Blackwell 8: 41-61. [ Links ]
5. Bown, P. 1999. Calcareous Nannofossil Biostratigraphy. British Micropaleontological Society Publication Series, Second Edition, Chapman y Hall, London, 315 pp. [ Links ]
6. Bown, P. 2005. Palaeogene calcareous nannofossils from the Kilwa and Lindi areas of coastal Tanzania (Tanzania Drilling Project 2003 - 4). Journal of Nannoplankton Research 27: 21-95. [ Links ]
7. Bown, P. y Young, J.R. 1999. Techniques. En: P. Bown (ed.), Calcareous Nannofossil Biostratigraphy. British Micro-paleontological Society Publications Series, Second Edition, Chapman y Hall, London, pp. 16-28. [ Links ]
8. Bramlette, M.N. y Martini, E. 1964. The great change in calcareous nannoplankton fossils between the Maestrichtian and Danian. Micropaleontology 10: 291-322. [ Links ]
9. Bramlette, M.N. y Riedel, W.R. 1954. Stratigraphic value of discoasters and some other microfossils related to Recent coccolit-hophores. Journal of Paleontology 28: 385-403. [ Links ]
10. Bramlette, M.N y Sullivan, F.R. 1961. Coccolithophorids and related Nannoplankton of the early Tertiary in California. Micropaleontology 7: 129-188. [ Links ]
11. Burnett, J. 1999. Upper Cretaceous. En: P. Bown (ed.), Calcareous Nannofossil Biostratigraphy. British Micropaleontological Socity Publications Series, Second Edition, Chapman y Hall, London, pp. 132-199. [ Links ]
12. Calegari, R. y Baldi, J. 1993. Análisis secuencial para el Terciario inferior de cuenca Austral mediante estudios sismoestratigráficos. 12º Congreso Geológico Argentino y 2º Congreso de Exploración de Hidrocarburos (Mendoza), Actas 1: 87-95. [ Links ]
13. Calegari, R., Baldi, M.J. y Pioli, O. 1993. Sismoestratigrafía del Terciario de cuenca Austral. Aplicación en proyectos exploratorios. Boletín de Informaciones Petroleras 10: 2-23. [ Links ]
14. Concheyro, A. 1991. Nanofósiles Calcáreos de la Formación Man Aike (Eoceno, sudeste del lago Cardiel) Santa Cruz, Argentina. Ameghiniana 28: 385-399. [ Links ]
15. Concheyro, A. y Angelozzi, G. 2002. Nanofósiles Calcáreos. En: M.J. Haller (ed.), Geología y Recursos Naturales de Santa Cruz. 15º Congreso Geológico Argentino (El Calafate, 2002), Relatorio 9: 495-517. [ Links ]
16. Deflandre, G. 1959. Sur les nannofossiles calcaires et leur systématique. Revue de Micropaléontologie 2: 127-152. [ Links ]
17. Deflandre, G. y Fert, C. 1954. Observations sur les coccolithophoridés actuels et fossiles en microscopie ordinaire et électronique. Annales de Paléontologie 40: 115-176. [ Links ]
18. Edwards, A.R. 1973. Key species of New Zealand calcareous nannofossils. New Zealand Journal of Geology and Geophysics 16: 68-89. [ Links ]
19. Furque, G. 1973. Descripción geológica de la Hoja 58b Lago Argentino. Boletín del servicio Nacional Minero y Geológico 140: 1-49. [ Links ]
20. Gartner, S. 1967. Calcareous nannofossil from Neogene of Trinidad, Jamaica and Gulf of Mexico. The University of Kansas Paleontological Contributions 29: 1-7. [ Links ]
21. Gartner, S. 1968. Coccoliths and related calcareous nannofossils from Upper Cretaceous deposits of Texas and Arkansas. University of Kansas Paleontological Contributions 48: 1-56. [ Links ]
22. Gartner, S. 1969. Correlation of Neogene planktonic foraminifera and calcareous nannofossil zones. Transactions. Gulf Coast Association of Geological Societies 19: 585-599. [ Links ]
23. Górka, H. 1957. Les Coccolithophoridés du Maestrichtien supérieur de Pologne. Acta Palaeontologica Polonica 2: 239-284. [ Links ]
24. Haq, B.U. 1968. Studies on upper Eocene calcareous nannoplankton from NW Germany. Stockholm Contributions in Geology 18: 13-74. [ Links ]
25. Haq, B.U. 1971. Paleogene calcareous nannoflora Part II: Oligocene of western Germany. Stockholm Contributions in Geology 25: 57-97. [ Links ]
26. Haq, B.U. y Berggren, W.A. 1978. Late Neogene calcareous plankton biochronology of the Rio Grande Rise (South Atlantic Ocean). Journal of Paleontology 52: 1167-1194. [ Links ]
27. Hay, W.W. 1977. Calcareous nannofossils. En: A.T.S. Ramsay (ed.), Oceanic Micropaleontology, Academic Press, London, vol. 2, pp. 1055-1200. [ Links ]
28. Hay, W.W. y Mohler, H.P. 1967. Calcareous nannoplankton from early Tertiary rocks at Pont Labau, France, and Paleocene- early Eocene correlations. Journal of Paleontology 41: 1505-1541. [ Links ]
29. Hay, W.W., Mohler, H.P. y Wade, M.E. 1966. Calcareous nannofossils from Nal`chik (northwest Caucasus). Eclogae Geologicae Helveticae 59: 379-399. [ Links ]
30. Hay, W.W., Mohler, H.P., Roth, P.H., Schmidt, R.R. y Boudreaux, J.E. 1967. Calcareous nannoplankton zonation of the Cenozoic of the Gulf Coast and Caribbean-Antillean area, and transoceanic correlation. Transactions of the Gulf Coast Association of Geological Societies 17: 428-480. [ Links ]
31. Heizler, M., Kay, R.F., Madden, R.H., Mazzoni, M.M., Ré, G.H., Sandeman, H. y Vucetich, M.G. 1998. Geocronologic age of the Casamayoran fauna at Gran Barranca, Chubut Province, Argentina. 7º Congreso Argentino de Paleontología y Bioestratigrafía (Bahía Blanca), Resúmenes: 89. [ Links ]
32. Höll, C., Zonneveld, K.A.F., Wilems, H. 1998. On the ecology of calcareous dinoflagellates: The Quaternary Eastern Equatorial Atlantic. Marine micropaleontology 33: 1-25. [ Links ]
33. Kamptner, E. 1941. Die Coccolithineen der Südwestküste von Istrien. Annalen des Naturhistorischen Museums Wien 51: 54-149. [ Links ]
34. Locker, S. 1967. Neue Coccolithophoriden (Flagellata) aus dem Alttertiär Norddeutschlands. Geologie (Berlin) 16: 361-364. [ Links ]
35. Lohmann, H. 1919. Die Bevölkerung des Ozeans mit Plankton nach den Ergebnissen der Zentrifugenfänge während der Ausreise der "Deutschland" 1911. Archive für Biontologie 4: 1-617. [ Links ]
36. Malumián, N. 1968. Foraminíferos del Cretácico superior y Terciario del subsuelo de la Provincia Santa Cruz, Argentina. Ameghiniana 5: 191-227. [ Links ]
37. Malumián, N. 1999. La Sedimentación y el Volcanismo Terciarios en la Patagonia Extraandina. En: R. Caminos (ed.), Geología Argentina, Instituto de Geología y Recursos Minerales, Anales 29: 557-612. [ Links ]
38. Malumián, N. 2002. El Terciario Marino. Sus relaciones con el eustatismo. En: M.J. Haller (ed.), Geología y Recursos Naturales de Santa Cruz. 15º Congreso Geológico Argentino (El Calafate) Relatorio 15: 237-244. [ Links ]
39. Malumián, N. y Caramés, A. 1995. El Daniano Marino de Patagonia (Argentina): Paleobiogeografía de los foraminíferos bentónicos. En: C. Náñez (ed.), Paleógeno de América del Sur. Asociación Paleontológica Argentina. Publicación Especial 3: 83-105. [ Links ]
40. Malumián, N. y Caramés, A. 1997. Upper Campanean-Paleogene from Río Turbio Coal measures area: micropaleontology and the Paleocene / Eocene boundary. Austral Basin. Journal of South American Earth Sciences 10: 189-201. [ Links ]
41. Malumián, N. y Masiuk, V. 1973. Asociaciones foraminiferológicas fósiles de la República Argentina. 5º Congreso Geológico Argentino (Carlos Paz, 1972), Actas 3: 433-453. [ Links ]
42. Malumián, N. y Masiuk, V. 1976. Foraminíferos de la Formación Cabeza de León (Cretácico Superior, Tierra del Fuego, Rep. Argentina). Revista de la Asociación Geológica Argentina 31: 180-202. [ Links ]
43. Malumián, N. y Náñez, C. 1996. Microfósiles y Nanofósiles calcáreos de la Plataforma Continental. En: V.A. Ramos y M.A. Turic (eds.), Geología y Recursos Naturales de la Plataforma Continental Argentina. 13º Congreso Geológico Argentino y 3º Congreso de Exploración de Hidrocarburos (Buenos Aires), Relatorio 5: 73-93. [ Links ]
44. Malumián, N. y Náñez, C. 2002. Los Foraminíferos. Su significado geológico y ambiental. En: M.J. Haller (ed.), Geología y Recursos Naturales de Santa Cruz. 15º Congreso Geológico Argentino (El Calafate), Relatorio 8: 481-493. [ Links ]
45. Malumián, N., Masiuk, V. y Riggi, J.C. 1971. Micropaleontología y sedimentología de la perforación SC-1, provincia de Santa Cruz, Argentina. Su importancia y correlaciones. Revista de la Asociación Geológica Argentina 46: 175-208. [ Links ]
46. Martini, E. 1971. Standard Tertiary and Quaternary calcareous nannoplankton zonation. En: A. Farinacci (ed.), Proceedings 2º Planktonic Conference (Roma, 1970), 2: 739-785. [ Links ]
47. McIntyre, A. y Bé, A.W.H. 1967. Modern Coccolithophoridae of the Atlantic Ocean. I. Placoliths and cyrtopliths. Deep Sea Research 14: 561-597. [ Links ]
48. Moshkovitz, S. y Ehrlich, A. 1976. Distribution of Middle and Upper Jurassic calcareous nannofossils in the Northeastern Negev, Israel and in Gebel Maghara, Northern Sinai. Geological Survey of Israel Bulletin 69: 1-47. [ Links ]
49. Mostajo, E. 1991. Nanofósiles calcáreos cenozóicos del pozo "Las Violetas 3". Isla Grande de Tierra del Fuego. Argentina. Ameghiniana 28: 311-315. [ Links ]
50. Noël, D. 1969. Arkhangelskiella (Coccolithes Crétacés) et formes affines du Bassin de Paris. Revue de Micropaléontologie 11: 191-204. [ Links ]
51. Nöel, D. 1970. Coccolithes Crétacés: La Craie Campanienne du Bassin de Paris. Éditions du centre National de la Recherche Scientifique, Paris, pp. 129 [ Links ]
52. Okada, H. y McIntyre, A. 1977. Modern coccolithophores of the Pacific and North Atlantic Oceans. Micropaleontology 23: 1-55. [ Links ]
53. Okada, H. y McIntyre, A. 1979. Seasonal distribution of modern coccolithophores in the Western North Atlantic Ocean. Marine Biology 54: 318-328. [ Links ]
54. Olivero, E.B., Malumián, N. y Palamarczuk, S. 2003. Estratigrafía del Cretácico superior-Paleoceno del área Bahía Thetis, Andes fueguinos, Argentina: Acontecimientos tectónicos y paleobiológicos. Revista Geológica de Chile 30: 245-263. [ Links ]
55. Perch-Nielsen, K. 1968. Der Feinbau und die Klassifikation der Coccolithen aus dem Maastrichtien von Dänemark. Kongelige Danske Videnskabernes Selskabs. Biologiske Skrifter 16: 1-96. [ Links ]
56. Perch-Nielsen, K. 1969. Die Coccolithen einiger dänischer Maastrichtien und Danienlokalitäten. Bulletin of the Geological Society of Denmark 19: 51-68. [ Links ]
57. Perch-Nielsen, K. 1971. Elektronenmikroskopische Untersu-chungen an Coccolithen und verwandten Formen aus dem Eoczän von Dänmark. Kongelige Danske Videnskabernes Selskabs. Biologiske Skrifter 18: 1-76. [ Links ]
58. Perch-Nielsen, K. 1973. Neue Coccolithen aus dem Maastrichtien von Dänemark, Madagascar und Ägypten. Bulletin of the Geological Society of Denmark 22: 306-333. [ Links ]
59. Perch-Nielsen, K. 1977. Albian to Pleistocene calcareous nanno-fossils from the Western South Atlantic, DSDP leg 39. En: P.R. Supko, Perch-Nielsen, K. et al., Initial Reports of the Deep Sea Drilling Project 39: 699-825. [ Links ]
60. Perch-Nielsen, K. 1980. New Tertiary calcareous nannofossils from the South Atlantic. Eclogae Geologicae Helvetiae 73: 1-7. [ Links ]
61. Perch-Nielsen, K., 1984. Validation of new combinations. International Nannofossil Association Newsletter 6: 42-46. [ Links ]
62. Perch-Nielsen, K., 1985a. Mesozoic Calcareous Nannofossils. En: H.H. Bolli, J.B. Saunders y K. Perch - Nielsen (eds.), Plankton Stratigraphy, Cambridge University Press, Cambridge, pp. 329-426. [ Links ]
63. Perch-Nielsen, K. 1985b. Cenozoic Calcareous Nannofossils. En: H.H. Bolli, J.B. Saunders y K. Perch-Nielsen (eds.), Plankton Stratigraphy, Cambridge University Press, Cambridge, pp. 427-554. [ Links ]
64. Perch-Nielsen, K., Sadek, A., Barakat, M.G. y Teleb, F. 1978. Late Cretaceous and Early Tertiary Calcareous nannofossil and Planktonic foraminifera zones from Egypt. Actes du 6º Colloque Africain de Micropaléontologie (Tunis 1974), Annales des Mines et de la Géologie 28: 337-403. [ Links ]
65. Radomski, A. 1968. Calcareous nannoplankton zones in Paleogene of the western Polish Carpatians. Rocznik Polskiego Towarzystwa Geologicznego 38: 545-605. [ Links ]
66. Reinhardt, P. 1966a. Zur Taxionomie und Biostratigraphie des fossilen Nannoplanktons aus dem Malm, der Kreide und dem Alttertiär Mitteleuropas. Freiberger Forschungshefte 196: 5-109. [ Links ]
67. Reinhardt, P. 1966b. Fossile vertreter coronoider und styloider Coccolithen (Familia Coccolithaceae Poche 1913). Monatsberiche der Deustchen Akademie der Wissenschaften zu Berlin 8: 513-524. [ Links ]
68. Robbiano, J.A., Arbe, H.A. y Bangui, A. 1996. Cuenca Austral Marina. En: V.A. Ramos y M.A. Turic (eds.), Geología y Recursos Naturales de la Plataforma Continental Argentina. 13º Congreso Geológico Argentino y 3º Congreso de Exploración de Hidrocarburos (Buenos Aires), Relatorio 17: 323-341. [ Links ]
69. Romein, A.J.T. 1979. Lineales in early Paleogene calcareous nan-noplankton. Utrecht Micropaleontological Bulletins 22: 1-230. [ Links ]
70. Ronchi, D. y Angelozzi, G. 1994. Bioestratigrafía del Cretácico-Terciario en dos pozos ubicados al Oeste de la Cuenca Austral. Boletín de Informaciones Petroleras Tercera Época 10: 65-76. [ Links ]
71. Roth, P.H. 1970. Oligocene calcareous nannoplankton biostratigraphy. Eclogae Geologicae Helveticae 63: 799 881. [ Links ]
72. Roth, P.H. y Thierstien, H. 1972. Calcareous Nannoplankton: Leg 14 of the Deep Sea Drilling Project. Initial Reports of the Deep Sea Drilling Project 14: 421-485. [ Links ]
73. Russo, A., Flores. M.A. y Di Benedetto, H. 1980. Patagonia austral extraandina. 2º Simposio Geología Regional Argentina. Academia Nacional de Ciencias (Córdoba), 2: 1431-1462. [ Links ]
74. Schiller, J. 1925. Die planktonischen Vegetationen des adriatischen Meeres. Archiv für Protistenkunde 51: 1-130. [ Links ]
75. Schiller, J. 1930. Coccolithineae. En: L. Rabenhorst (ed.), Kryptogamen-Flora von Deutschland, Osterreich und der Schweiz, Akademische Verlagsgesellschaft, Leipzig, pp. 89-267. [ Links ]
76. Shannon, C.E. y Wiener, W. 1963. The Mathematical Theory of Communication. University of Illinois Press, Illinois, 117 pp. [ Links ]
77. Sissingh, W. 1977. Biostratigraphy of Cretaceous calcareous nannoplankton. Geologie en Mijnbouw 56: 37-65. [ Links ]
78. Stoico, M. y Baldanza, A. 1995. Early and middle Jurassic calcareous nannofossil biozonation of the Monti Sabini area (Latium, Norhern Apennines, Central Italy). Palaeopelagos 5: 75-110. [ Links ]
79. Stover, L.E. 1966. Cretaceous coccoliths and associated nannofossils from France and the Netherlands. Micropaleontology 12: 133-167. [ Links ]
80. Stradner, H. 1961. Vorkommen von Nannofossilien im Mesozoikum und Alttertiär. Erdoël-Zeitschrift 77: 77-88. [ Links ]
81. Stradner, H. y Edwards, A.R. 1968. Electron microscopic studies on upper Eocene coccoliths from the Oamaru diatomite, New Zealand, Jahrbuch. Geologische Bundesanstalt 13: 1-66. [ Links ]
82. Thierstein, H.R., Geitzenauer, K.R., Molfino, B. y Shackleton, N. 1977. Global Synchroneity of The Late Quaternary coccolith datum levels: validation by oxygen isotopes. Geology 5: 400-404. [ Links ]
83. Varol, O. 1999. Palaeogene. En: P. Bown (ed.), Calcareous Nannofossil Biostratigraphy, British Micropaleontological Society, Publications Series, Second Edition, Chapman y Hall, London, pp. 200-224. [ Links ]
84.Varol, O. y Jakubowski, M. 1989. Some new nannofossil taxa. International Nannofossil Association Newsletter 11: 24-29. [ Links ]
85. Vekshina, V.N. 1959. Coccolithophoridea of the Maastrichtian deposits of the west Siberian lowlands. Siberian Science Research Institute of Geology, Geophysics, Mineralogy and Raw Materials 2: 56-81. [ Links ]
86. Verbeek, J.W. 1977. Calcareous nannoplankton biostratigraphy of Middle and Upper Cretaceous deposits in Tunisia, southern Spain and France. Utrecht Micropaleontological Bulletins 16: 1-157. [ Links ]
87. Vink, A., Zonneveld, K.A.F. y Willems, H. 2000. Distribution of calcareous dinoflagellate cysts in surface sediments of the western equatorial Atlantic Ocean, and their potential use in palaeoceanography. Marine Micropaleontology 38: 149-180. [ Links ]
88. Wallich, G.C. 1877. Observations on the coccosphere. Annals and Magazine of Natural History 19: 342-350. [ Links ]
89. Wise, S.W.Jr. 1983. Mesozoic and Cenozoic calcareous nannofossils recovered by Deep Sea Drilling Project Leg 71 in the Falkland Plateau Region, Southwest Atlantic Ocean. Initial Reports of the Deep Sea Drilling Project 71: 481-550. [ Links ]
90.Wise, S.W.Jr. y Mostajo, E.L. 1983. Correlation of Eocene- Oligocene Calcareous Nannofossil assemblages from piston cores taken near Deep Sea Drilling Sites 511 and 512, Southwest Atlantic Ocean. Initial Reports of the Deep Sea Drilling Project 71: 1171-1180. [ Links ]
91. Wise, S.W.Jr. y Wind, F.H. 1977. Mesozoic and Cenozoic calcareous nannofossils recovered by DSDP Leg 36 drilling on the Falkland Plateau, SW Atlantic sector of the Southern Ocean. Initial Reports of the Deep Sea Drilling Project 36: 296-309. [ Links ]
92. Zonneveld, K.A.F., Höll, C., Janofske, D., Karwath, B., Kerntopf, B., Rühlemán, C., Willems, H. 1999. Calcareous dinoflagellate cysts as palaeo-environmental tool. En: G. Fischer y G. Wefer (eds.), Use of Proxys in Paleoceanography: Examples from the south Atlantic. Springer, Berlín. pp. 145-164. [ Links ]
Recibido: 6 de diciembre de 2007.
Aceptado: 15 de diciembre de 2008.

