










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana vol.46 no.2 Buenos Aires abr./jun. 2009
ARTÍCULOS ORIGINALES
Nuevos restos fósiles del dinosaurio Lirainosaurus (Sauropoda, Titanosauria) en el Cretácico Superior (Campaniano-Maastrichtiano) de la Península Ibérica
Julio Company1, Xabier Pereda Suberbiola2 y José Ignacio Ruiz-Omeñaca3
1Departamento de Ingeniería del Terreno, Universidad Politécnica de Valencia, 46022 Valencia, España; Instituto "Cavanilles" de Biodiversidad y Biología Evolutiva, Universidad de Valencia, Apartado de Correos 22085, 46071 Valencia, España. company@uv.es
2Departamento de Estratigrafía y Paleontología, Facultad de Ciencia y Tecnología, Universidad del País Vasco/EHU, Apartado 644, 48080 Bilbao, España. xabier.pereda@ehu.es
3Museo del Jurásico de Asturias (MUJA), 33328 Colunga, España, y Grupo Aragosaurus-IUCA, Departamento de Ciencias de la Tierra (Paleontología), Universidad de Zaragoza, 50009 Zaragoza, España. jigruiz@unizar.es
Resumen. El saurópodo Lirainosaurus astibiae Sanz et al., 1999, un grácil titanosaurio de talla media (entre 8 y 10 m de longitud), fue definido a partir de un fragmento de cráneo, dientes y huesos postcraneales procedentes del Cretácico Superior (Campaniano superior a Maastrichtiano inferior) de Laño, en el norte de la Península Ibérica. En este trabajo se describen nuevos restos fósiles que constituyen el primer registro del género Lirainosaurus fuera de su localidad tipo. El nuevo material, procedente de niveles continentales lacustres de la Formación Sierra Perenchiza (Campaniano superior a Maastrichtiano inferior) de la localidad de Chera (provincia de Valencia, este de España), comprende elementos vertebrales, huesos de la cintura escapular y de las extremidades, y es asignado a Lirainosaurus cf. astibiae por la presencia de las siguientes autapomorfias: (1) centros caudales posteriores con un cóndilo articular reducido dotado de un característico surco sagital, (2) lámina escapular provista medialmente de una cresta ventral marcada, y (3) foramen coracoideo situado cerca de los márgenes escapular y dorsal del coracoides. Esta combinación de caracteres es exclusiva de Lirainosaurus. Otros caracteres propios de la especie L. astibiae, tales como la morfología de la placa esternal o de las vértebras dorsales no se han preservado y/o no se observan claramente en los materiales recuperados. Lirainosaurus ha sido previamente relacionado con los Saltasaurinae de América del Sur, aunque sólo se puedan reconocer afinidades filogenéticas claras con un clado más inclusivo, los Eutitanosauria. Esta interpretación es coherente con la amplia distribución biogeográfica de estos titanosaurios durante el Cretácico Tardío.
Abstract. New fossil remains of the dinosaur Lirainosaurus (Sauropoda, Titanosauria) from the Upper Cretaceous (Campanian-Maastrichtian) of the Iberian Peninsula. The sauropod Lirainosaurus astibiae Sanz et al., 1999, a medium-sized, gracile titanosaur (about 8-10 m in length), is known by a fragmentary skull, teeth and postcranial bones from the Late Cretaceous (late Campanian to early Maastrichtian) of Laño, northern Iberian Peninsula. The aim of this paper is to describe new fossil bones of Lirainosaurus which constitute the first record of Lirainosaurus outside its type locality. The new material, recovered from lacustrine levels of the Sierra Perenchiza Formation (late Campanian to early Maastrichtian) at Chera locality (Valencia Province, eastern Spain), consists of vertebral remains, pectoral girdle elements and limb bones. The material is referred to Lirainosaurus cf. astibiae on the basis of the presence of the following autapomorphies: (1) posterior caudal centra characterized by a restricted articular condyle with a sagittal condylar groove, (2) prominent ridge on the ventral margin of the scapular blade, and (3) coracoidal foramen located close to the scapular border and to the dorsal margin of the coracoid. This combination of features is exclusive of Lirainosaurus. Other diagnostic features of L. astibiae are not preserved or can not be recognised in the recovered material. Lirainosaurus has been previously related with the Saltasaurinae of South America, but clear phylogenetic affinities can be only recognized with a more inclusive clade, the Eutitanosauria. This interpretation is coherent with the wide biogeographical distribution of the eutitanosaurians during the Late Cretaceous.
Palabras clave. Sauropoda; Titanosauria; Lirainosaurus; Chera; Campaniano-Maastrichtiano; España.
Key words. Sauropoda; Titanosauria; Lirainosaurus; Chera; Campanian-Maastrichtian; Spain.
Introducción
Los titanosaurios son los dinosaurios saurópodos con mejor registro fósil a nivel mundial en el Cretácico Superior, y constituyen el único grupo de saurópodos conocido en Europa a finales del Cretácico (Le Loeuff, 1993; Hunt et al., 1994; Upchurch et al., 2004). Restos fósiles de titanosaurios han sido descubiertos en yacimientos del viejo continente desde hace más de un siglo, y provienen en su mayor parte de Europa Central y del Dominio Iberoarmoricano (Península Ibérica y sur de Francia) (Le Loeuff, 1992, 1993). Hasta la fecha, sólo tres taxones han sido definidos formalmente: Magyarosaurus dacus Nopcsa, 1915 en Transilvania, Rumania (Nopcsa, 1915; Huene, 1932), Ampelosaurus atacis Le Loeuff, 1995 en Languedoc, Francia (Le Loeuff, 1995, 2005a), y Lirainosaurus astibiae Sanz et al., 1999 en el norte de la Península Ibérica (Sanz et al., 1999). En la Península Ibérica se ha mencionado tanto la presencia de restos directos aislados y fragmentarios de dinosaurios titanosaurios (Lapparent y Aguirre, 1956; Casanovas et al., 1987, 1995; Sanz y Buscalioni, 1987; Masriera y Ullastre, 1988; Sanz et al., 1992; Casanovas, 1993; Casanovas y Santafé, 1993; Pereda Suberbiola et al., 1999; Canudo, 2001; Pereda Suberbiola y Ruiz-Omeñaca, 2002), como de evidencias indirectas, consistentes en icnitas halladas en el área pirenaica (Le Loeuff y Martínez, 1997; Schulp y Brokx, 1999; Vila et al., 2005), y en fragmentos de cáscaras de huevos atribuibles a dinosaurios titanosaurios (Sanz et al., 1992; Moratalla-García, 1993; López-Martínez, 2000; López-Martínez et al., 2000; Bravo et al., 2005).
Lirainosaurus astibiae es la única especie de saurópodo definida en el Cretácico Superior de la Península Ibérica (Sanz et al., 1999). Este taxón fue descrito a partir de un fragmento craneal, dientes aislados y numerosos restos postcraneales asociados, incluyendo osteodermos, procedentes del yacimiento Campano-Maastrichtiano de Laño (Región Vasco-Cantábrica). El resto del material ibérico anteriormente mencionado ha sido atribuido provisionalmente a Titanosauridae o a Titanosauria indet. (véase Royo-Torres y Canudo, 2003). Algunos de estos restos fueron asignados a Titanosaurus indicus e Hypselosaurus sp. (Lapparent y Aguirre, 1956; Casanovas et al., 1987), si bien su carácter fragmentario no permite justificar una asignación genérica válida (Le Loeuff, 1993; Upchurch et al., 2004).
En este trabajo se describen los restos fósiles de titanosaurio procedentes de diversos yacimientos paleontológicos localizados próximos al municipio de Chera (provincia de Valencia, España), consistentes en elementos vertebrales y huesos apendiculares. Se trata del primer registro de restos directos de saurópodos titanosaurios en la provincia de Valencia y la primera cita del género Lirainosaurus fuera de su localidad tipo.
Abreviaturas. CMPGC, Centro de Divulgación Cultural del Parque Geológico de Chera, Chera, España; MCNA, Museo de Ciencias Naturales de Alava/Arabako Natur Zientzien Museoa, Vitoria-Gasteiz, España; MGUV, Museo de Geología de la Universidad de Valencia, Burjassot, España; MHNM, Musée d'Histoire Naturelle, Palais Longchamp, Marsella, Francia.
Geología y estratigrafía
El material descrito en este trabajo procede de un conjunto de yacimientos paleontológicos localizados en la Fosa de Chera, unidad estructural de alineación NE-SW ubicada en el sector Suroccidental de la Cordillera Ibérica, aproximadamente a 60 Km al oeste de la ciudad de Valencia (figura 1). El área de estudio alberga importantes depósitos de carbonatos marinos del Cretácico Superior, culminados por secuencias regresivas del Campaniano superior-Maas-trichtiano inferior. Estas secuencias finales son asignadas por Vilas et al. (1982) a la formación Calizas y Margas de la Sierra Perenchiza, en cuyos afloramientos se localizan los yacimientos de procedencia de los materiales estudiados (véase tabla 1).

Figura 1. Mapa de localización de los yacimientos paleontológicos de Chera en la provincia de Valencia, Este de España / Location map of the Chera localities in Valencia province, eastern Spain.
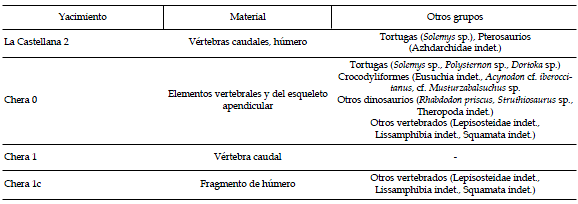
Tabla 1. Relación de yacimientos en el área de estudio que han proporcionado restos del saurópodo Lirainosaurus / list of fossiliferous horizons in the study area that have yielded remains of Lirainosaurus.
La Formación Sierra Perenchiza es una unidad continental constituida por una alternancia de calizas y margas lacustres ordenadas en secuencias regresivas de somerización. Los primeros estudios descriptivos de estos materiales fueron llevados a cabo por Gutiérrez et al. (1975a, 1975b), quienes asignaron estos depósitos al Cretácico terminal en base a la presencia de carófitas y gasterópodos continentales. La definición formal de la unidad se debe al trabajo de síntesis de Vilas et al. (1982) en el que se establece el paraestratotipo de la unidad en el Barranco del Agua de la Fosa de Chera, lugar donde se localizan los yacimientos referidos en este trabajo.
Los sedimentos que albergan los yacimientos paleontológicos de Chera son característicos de medios palustres o lacustres someros, emplazados en un medio litoral y sometidos a episodios periódicos de inundación y desecación. Estos materiales se ordenan en secuencias cíclicas de somerización, de escaso desarrollo vertical (Platt y Wright, 1991, 1992), que evolucionan desde términos basales de marcado carácter lacustre, representados por margas y calizas micríticas con abundantes gasterópodos, ostrácodos y carófitas, hacia términos palustres, constituidos por calizas margosas considerablemente alteradas por procesos derivados de una exposición subaérea, evidenciando incluso una intensa edafización (Alonso et al., 1987, 1991). Los fósiles de vertebrados se concentran preferentemente en la interfase que separa las facies palustres de calizas nodulares edafizadas (máxima retracción de las lagunas) de los depósitos arcillomargosos que marcan una nueva fase de inundación y el comienzo de la sedimentación de la secuencia suprayacente (Company, 2004).
área de estudio se han localizado numerosos afloramientos fosilíferos con restos de vertebrados fósiles, habiéndose recolectado restos del saurópodo Lirainosaurus en cuatro niveles diferentes: Chera 0, Chera 1, Chera 1c, y La Castellana 2 (véase tabla 1).
Tafonomía y paleoecología
Los restos fósiles de titanosaurios descubiertos en Chera corresponden a elementos del esqueleto axial, fundamentalmente vértebras dorsales y caudales y, en menor proporción, a huesos del esqueleto apendicular. La mayor concentración de restos fósiles procede del yacimiento denominado Chera 0. Se trata de un nivel de lutitas margosas grises, de aspecto masivo, limitado por sendos niveles lenticulares de calizas lacustres intensamente edafizadas, formando, en conjunto, una secuencia palustre de expansión y retracción. En el yacimiento Chera 0 la repetición de elementos esqueletales sugiere la presencia de, al menos, tres individuos diferentes. Otros niveles de similares características (Chera 1, Chera 1c y La Castellana 2) han proporcionado únicamente restos aislados de titanosaurios (tabla 1).
La asociación fósil procedente de Chera 0 se compone de fauna y flora límnica autóctona (carófitas, gasterópodos de agua dulce, ostrácodos, peces actinopterigios), asociada a restos de vertebrados continentales de modo de vida semiacuática (anfibios, tortugas, cocodrilos) o terrestre (dinosaurios, pterosaurios, escamosos), constituyendo una asociación mezclada multiespecífica y parautóctona (Kidwell et al., 1986) que confirma el origen palustre del medio de depósito.
El análisis tafonómico de los restos de macrovertebrados de Chera 0 muestra que la mayor parte de los restos óseos presenta escasas evidencias de meteorización por exposición subaérea o alteración por abrasión, indicando un transporte limitado y un enterramiento moderadamente rápido después de la desarticulación de los esqueletos (Company, 2004). Los restos se distribuyen en el nivel fosilífero como un conjunto de elementos asociados pero dispersos (Behrensmeyer, 1991), esparcidos sobre un área superior al tamaño de un esqueleto articulado. Este hecho no impide que determinados elementos puedan ser asignados, con cierta reserva, a un mismo individuo, en función de sus características anatómicas y de tamaño. En este sentido, elementos de la cintura escapular de un ejemplar de Lirainosaurus (escápula y coracoides) próximos a sendos húmeros, son asignados a un mismo individuo. En menor medida, ciertos elementos aparecen fracturados y presentan buzamientos considerables. Las fracturas incluyen bordes afilados, sugiriendo fracturación y enterramiento por pisoteo previo al enterramiento (Hill, 1979). Finalmente, un reducido número de restos muy fragmentarios presenta evidencias de mayor modificación tafonómica, producida probablemente por retrabajamiento de los elementos esqueléticos en la propia cuenca, durante las fases de secado y posterior inundación de las cuencas palustres (Burton, 1998). La ausencia de orientaciones preferentes en los huesos largos indica que la acción de corrientes transportadoras no jugó un papel importante en los procesos de acumulación de los restos (Company, 2004). No hay evidencias de reelaboración fosildiagenética.
La concentración de fósiles en el yacimiento se puede interpretar como resultado de una acumulación gradual de cadáveres in situ, y enterramiento moderadamente rápido, con cierto grado de desarticulación y dispersión post mortem en un medio sedimentario de baja energía. Se trataría de un paleoambiente constituido por lagunas sometidas a ciclos de desecaciones e inundaciones periódicas, que con frecuencia constituye un lugar típico de concentración de biotas de vertebrados (Behrensmeyer, 1991).
Paleontología sistemática
Sauropodomorpha Huene, 1932
Sauropoda Marsh, 1878
Titanosauria Bonaparte y Coria, 1993
Eutitanosauria Sanz, Powell, Le Loeuff, Martínez y Pereda Suberbiola, 1999
Género Lirainosaurus Sanz, Powell, Le Loeuff, Martínez y Pereda Suberbiola, 1999
Especie tipo. Lirainosaurus astibiae Sanz, Powell, Le Loeuff, Martínez y Pereda Suberbiola, 1999
Localidad y horizonte. Cantera de Laño (Condado de Treviño, Burgos); niveles continentales del Sinclinorio Subcantábrico (Unidad S1 U3) equivalentes a la Formación Sedano de la Plataforma Norcastellana (definida por Floquet, 1991; véase Lécuyer et al., 2003) (subunidad B3 según Baceta et al., 1999). Cretácico Superior, Campaniano superior a Maastrichtiano inferior.
Lirainosaurus cf. astibiae Figuras 2-4

Figura 2. Lirainosaurus cf. astibiae, Formación Sierra Perenchiza, Cretácico Superior (Campaniano-Maastrichtiano), Chera, provincia de Valencia, España / Lirainosaurus cf. astibiae, Sierra Perenchiza Formation, Late Cretaceous (Campanian-Maastrichtian), Chera, Valencia province, Spain. 1-3, vértebra dorsal (MGUV 17320) en vistas lateral derecha (1), posterior (2) y lateral izquierda (3) / 1-3, dorsal vertebra (MGUV 17320) in right lateral (1), posterior (2) and left lateral (3) views. 4-6, vértebra caudal anterior (MGUV 17169) en vistas anterior (4), lateral izquierda (5), y posterior (6) / 4-6, anterior caudal vertebra (MGUV 17169) in anterior (4), left lateral (5), and posterior (6) views. 7-10, vértebra caudal media a posterior (MGUV-17028) en vistas dorsal (7), anterior (8), lateral izquierda (9) y posterior (10) / 7-10, middle to posterior caudal vertebra (MGUV-17028) in dorsal (7), anterior (8), left lateral (9) and posterior (10) views. 11-13, vértebra caudal posterior (MGUV 17096) en vistas lateral izquierda (11), anterior (12) y posterior (13) / 11-13, posterior distal caudal vertebra (MGUV 17096) in left lateral (11), anterior (12) and posterior (13) views. 14-15, arco hemal (MGUV 17168) en vistas lateral izquierda (14) y posterior (15) / 14-15, chevron (MGUV 17168) in left lateral (14) and posterior (15) views. Escala gráfica = 5 cm / scale bar = 5 cm.
Material estudiado y procedencia. Material desarticulado procedente del yacimiento Chera 0, consistente en vértebras dorsales, vértebras caudales, arcos hemales, una placa esternal fragmentaria, elementos de la cintura escapular (escápula, coracoides) y del esqueleto apendicular (húmeros, ulnas, fémures). Material aislado procedente de otros niveles fosilíferos (Chera 1, Chera 1c, La Castellana 2), consistente en dos húmeros fragmentarios y diversas vértebras caudales. Véase anexo I y tabla I. Chera, provincia de Valencia (España); Formación Calizas y Margas de la Sierra Perenchiza, Cretácico Superior, Campaniano superior a Maastrichtiano inferior (Vilas et al., 1982; Company, 2004; Company et al., 2005).
Descripción
Vértebras dorsales. Se han recuperado cuatro vértebras dorsales incompletas (MGUV 17229, 17295, 17315, 17320) que conservan la mayor parte del centro vertebral, así como parte del arco neural (figura 2.1-3). A pesar de su carácter fragmentario, es posible reconocer en ellas algunas de las principales características de las vértebras dorsales de los dinosaurios titanosaurios, y particularmente de Lirainosaurus astibiae: centros vertebrales opistocélicos, con la cara articular anterior marcadamente convexa, considerablemente expandidos en sus extremos, que alojan, en la base del arco neural, sendos pleurocelos de tipo ocelar. El interior de los centros está constituido por tejido esponjoso de escasa densidad trabecular (estructura de tipo "panal de abeja"), recubierto por una delgada capa externa de tejido compacto (1-2 mm de grosor). Esta constitución histológica ha sido también descrita en otros saurópodos titanosauriformes (Salgado et al., 1997; Upchurch, 1998; Wilson y Sereno, 1998; Wilson, 2002; Le Loeuff, 2005a; Bonaparte et al., 2006).
El arco neural se sitúa en posición central, y ocupa aproximadamente el 50% de la longitud del centro. En los especímenes recuperados se ha conservado parte de las láminas óseas que refuerzan los diferentes elementos del arco neural, incluyendo la base de la lámina centrodiapofisial posterior, y la parte basal de la lámina centropostzigapofisial (figura 2.1)
La morfología de las vértebras dorsales de Chera es compatible con la descrita por Sanz et al. (1999) para Lirainosaurus: los pleurocelos son alargados y profundos, con los bordes anterior y posterior acuminados, como en otros titanosaurios (Salgado et al., 1997; Bonaparte et al., 2006), incluido Lirainosaurus. A diferencia de los saltasáuridos, entre otros saurópodos (Upchurch et al., 2004: apéndice), los centros dorsales de Chera y de Lirainosaurus son subcirculares en sección transversal. El material de Chera comparte con Lirainosaurus y algunos Lithostrotia (Upchurch et al., 2004) la ausencia de articulaciones de tipo hipósfeno-hipantro, aunque no puede descartarse que se trate de un efecto debido al carácter fragmentario del material.
Vértebras caudales. Se ha recuperado una veintena de vértebras caudales, incluyendo desde elementos prácticamente completos (MGUV 16438, 16448, 17016, 17027, 17028, 17094, 17096, 17105, 17169, 17314, CMPGC 49), hasta centros vertebrales en diferente estado de conservación (MGUV 17090, 17092, 17093, 17097, 17101, 17337, 17513, 17539) (figura 2.4-13). En su mayor parte proceden de Chera 0, a excepción de un conjunto de tres vértebras caudales distales (MGUV 17096, 17243, CMPGC 49), procedentes del yacimiento La Castellana 2. Estos elementos, muy semejantes en forma y dimensiones, fueron hallados asociados en un mismo nivel, por lo que podrían formar parte de una misma serie caudal.
En líneas generales, las vértebras caudales anteriores presentan centros vertebrales relativamente cortos, procélicos, con la superficie articular posterior fuertemente convexa y prominente, y provistos de caras laterales cóncavas (figura 2.4-6). A lo largo de la serie caudal, los centros se vuelven considerablemente más alargados y deprimidos dorsoventralmente, de forma que la longitud de los centros vertebrales más distales llega a ser hasta tres veces la altura o anchura máxima de las caras articulares (figura 2.7-13). La característica más notable de los centros vertebrales caudales es la presencia de un cóndilo articular posterior muy prominente, pero restringido a la superficie central de la cara articular. En vista posterior, el cóndilo presenta un surco sagital (figura 2.13), que probablemente sirvió para la inserción de un ligamento intervertebral que daría mayor rigidez a la parte distal de la cola (Sanz et al., 1999). Un surco similar también puede apreciarse en el cótilo anterior (figura 2.12). Una depresión longitudinal recorre la superficie lateral de los centros desde el borde articular anterior hasta la parte posterior de la inserción del arco neural, justo por debajo de éste.
Los arcos neurales son bajos y robustos, y se sitúan en posición anterior. Presentan unas prezigapófisis considerablemente más desarrolladas que las postzigapófisis, y que se proyectan horizontalmente hasta alcanzar la región central de la vértebra precedente. Las superficies articulares de las prezigapófisis y las postzigapófisis son ovaladas.
Internamente, los centros vertebrales caudales están constituidos por tejido trabecular de mayor densidad que el observado en los centros vertebrales dorsales. La capa de tejido compacto cortical presenta, externamente, un fino estriado longitudinal.
La presencia de arcos neurales situados en la mitad anterior de los centros vertebrales caudales posteriores es un carácter derivado de los saurópodos titanosauriformes (Salgado et al., 1997; Upchurch, 1998). Los centros vertebrales de las caudales posteriores de Chera son procélicos, con una cara articular posterior convexa, carácter propio de los titanosaurios derivados (Salgado et al., 1997; Wilson, 2002) o Lithostrotia (Upchurch et al., 2004). Además, presentan un cóndilo articular reducido, provisto de un surco sagital en posición central. Ambos caracteres han sido definidos como autapomorfías de Lirainosaurus astibiae. No obstante, la reducción del cóndilo articular está también presente en las vértebras caudales posteriores de otros titanosaurios, como Mendozasaurus neguyelap (González-Riga, 2003) y Maxakalisaurus topai (Kellner et al., 2006). En Mendozasaurus existe además una pequeña depresión circular en la base del cóndilo, pero carece del surco sagital característico de Lirainosaurus (Sanz et al., 1999). El carácter fragmentario del material de Chera no permite confirmar la presencia de una lámina en la fosa interzigapofi-siaria de las vértebras caudales anteriores, definida por Sanz et al. (1999) como autapomórfica de Lirainosaurus astibiae.
Otra autapomorfía de Lirainosaurus astibiae descrita por Sanz et al. (1999) es la escasa proyección de la estructura espinopostzigapofisiaria en las vértebras caudales posteriores. Este carácter también parece estar presente en las vértebras caudales de Chera (figura 2.7-10) y, con reservas, en Ampelosaurus (Le Loeuff, 2005a: fig. 4.9). En otros titanosaurios, la estructura formada por la espina neural y las postzigapófisis de las vértebras caudales medias y posteriores se extiende posteriormente y puede alcanzar y/o sobrepasar el cóndilo articular (Powell, 2003; Kellner et al., 2005, 2006).
Magyarosaurus dacus de Transilvania (Nopcsa, 1915; Huene, 1932; Le Loeuff, 1993) y Ampelosaurus atacis de Languedoc (Le Loeuff, 1992, 1995, 2005a) están más comprimidos lateromedialmente que los centros vertebrales caudales de Lirainosaurus astibiae y los ejemplares de Chera. Estos últimos presentan incluso un cuerpo vertebral prácticamente cilíndrico, más ancho que alto (figura 2.8-10, 12-13). La presencia de centros caudales dorsoventralmente deprimidos es un carácter que Lirainosaurus comparte con otros titanosaurios, como Alamosaurus, Opisthocoelicaudia, Pellegrinisaurus y Saltasaurus, y el titanosauriforme basal Cedarosaurus (Upchurch et al., 2004: suplemento). Las vértebras caudales del material tipo de "Hypselosaurus priscus" presentan igualmente esa morfología (Le Loeuff, 1992). Por último, ninguno de los titanosaurios europeos parece presentar centros caudales distales biconvexos, lo que los diferencia de formas como Saltasaurus, Neuquensaurus y Opisthocoelicaudia (Wilson et al., 1999).
Arcos hemales. Se han recuperado tres arcos hemales (MGUV 17168, 17192, 17332) asociados a las vértebras caudales. Se caracterizan por presentar dos gruesas ramas articulares que finalizan en facetas articulares separadas, sencillas, las cuales delimitan un canal hemal estrecho (figura 2.14-15). La espina hemal está constituida por una delgada lámina ósea de morfología subcuadrangular. Su longitud equivale a la de las ramas hemales y se arquea ligeramente en sentido caudal.
No se han descrito arcos hemales en el material original de Lirainosaurus astibiae (Sanz et al., 1999) que permitan su comparación. Tampoco han sido descritos en otros titanosaurios del Cretácico Superior de Europa.
Escápula y coracoides. El yacimiento de Chera 0 ha proporcionado una escápula (MCUV 17645) y un coracoides (MCUV 17238) asociados, ambos incompletos (figura 3.1-3). Se trata de dos elementos izquierdos que, por su relación de tamaño, podrían pertenecer al mismo individuo.

Figura 3. Lirainosaurus cf. astibiae, Formación Sierra Perenchiza, Cretácico Superior (Campaniano-Maastrichtiano), Chera, provincia de Valencia, España / Lirainosaurus cf. astibiae, Sierra Perenchiza Formation, Late Cretaceous (Campanian-Maastrichtian), Chera, Valencia province, Spain. 1, escápula izquierda (MGUV 17645) en vista lateral / 1, left scapula (MGUV 17645) in lateral view. 2-3, coracoides izquierdo (MGUV 17238) en vistas lateral (2) y distal (3) / 2-3, left coracoid (MGUV 17238) in lateral (2) and distal (3) views. 4-5, húmero izquierdo (MGUV 17166) en vistas posterior (4) y anterior (5) / 4-5, left humerus (MGUV 17166) in posterior (4) and anterior (5) views. 6-8, ulna izquierda (MGUV-17274) en vistas medial (6), lateral (7) y anterior (8) / 6-8, left ulna (MGUV-17274) in medial (6), lateral (7) and anterior (8) views. Escala gráfica = 5 cm / scale bar = 5 cm.
La escápula conserva parte de la lámina escapular, la superficie de articulación para el coracoides y la fosa supracoracoidea (figura 3.1). La extremidad dorsal de la lámina escapular y el borde anterodorsal (acromion) no se han conservado. La escápula es convexa lateralmente. La lámina escapular es estrecha. En vista medial, se observa una cresta rugosa en el margen ventral del tercio proximal de la lámina y una prominencia cerca del borde dorsal de la misma. Ambas crestas sirvieron probablemente como áreas de inserción muscular (M. serratus superficialis en la cresta ventral y M. subcoracoscapularis en la prominencia dorsal; Borsuk-Bialynicka, 1977, fig. 6.1).
El coracoides está incompleto, ya que no conserva parte del borde anterodorsal ni de la superficie de articulación para la escápula (figura 3.2-3). Presenta forma subcuadrangular y parece relativamente grande con respecto a la escápula. El contorno del borde anteroventral es anguloso. El foramen coracoideo está situado cerca de la superficie de articulación para la escápula y próximo al borde dorsal. No se observa una prominencia marcada por debajo de la cavidad glenoidea.
La forma subcuadrangular del coracoides, con un borde anteromedial recto, está presente en numerosos titanosaurios (Salgado et al., 1997; Sanz et al., 1999; Wilson, 2002; Powell, 2003). Otro carácter que Lirainosaurus comparte con algunos titanosaurios, como Aeolosaurus, Saltasaurus, Magyarosaurus y Opisthocoelicaudia (Borsuk-Bialynicka, 1977; Le Loeuff, 1992; Powell, 1992, 2003; Sanz et al., 1999), es la existencia de una prominencia medial situada próxima al borde dorsal de la lámina escapular. Esta prominencia también se observa en el material de Chera. Además, el material valenciano comparte con Lirainosaurus la presencia de una cresta ventral en la cara medial de la lámina escapular (Sanz et al., 1999). Esta cresta se observa, asimismo, en otros titanosaurios, como Opisthocoelicaudia, por lo que algunos autores (Upchurch et al., 2004: suplemento) alegan que no se trata de una autapomorfía de Lirainosaurus. Le Loeuff (1992) indica que la escápula de Ampelosaurus posee igualmente una cresta ventral, pero mucho menos marcada que en Lirainosaurus. Otro carácter compartido por el titanosaurio valenciano y Lirainosaurus es la existencia de un foramen coracoideo situado próximo al margen escapular y al borde dorsal del coracoides (Sanz et al., 1999). Este carácter también podría observarse en los titanosaurios europeos Ampelosaurus y Magyarosaurus (Sanz et al., 1999), aunque Powell (2003) señala que el foramen coracoideo está situado lejos del margen escapular en Ampelosaurus. La aparente presencia en algunos especímenes de Ampelosaurus de un foramen abierto dorsalmente es un artefacto debido a rotura (Le Loeuff, 2005a). Por su parte, en Magyarosaurus la presencia de este carácter debe aún ser confirmada. En este trabajo se adopta la interpretación de Sanz et al. (1999), según la cual la posición del foramen cerca de los márgenes escapular y dorsal del coracoides es una autapomorfía de Lirainosaurus. Por último, el coracoides de Chera no muestra una marcada prominencia infraglenoidea, a diferencia de Lirainosaurus astibiae y de los Saltasauridae (sensu Wilson, 2002).
Placa esternal. Se ha recuperado una placa esternal incompleta procedente del yacimiento Chera 0. Presenta una forma semilunar, característica de las placas esternales de titanosaurios (Salgado et al., 1997; Calvo y González-Riga, 2003). Exhibe un margen lateral engrosado, moderadamente cóncavo, y un borde medial convexo y muy adelgazado, consistiendo en una delgada lámina ósea. La superficie externa es convexa, mientras que la superficie interna es cóncava. Debido al mal estado de conservación, no es posible confirmar en el espécimen de Chera la presencia de una cresta anteroventral y de un proceso anterolateral, cuya combinación, junto con un margen lateral moderadamente cóncavo, es una autapomorfia de Lirainosaurus (Sanz et al., 1999; Kellner et al., 2006).
Húmero. El húmero de Lirainosaurus es un elemento esbelto, aunque considerablemente expandido en sus extremos. Está representado en la asociación de Chera por siete húmeros incompletos que han sido recuperados de diferentes horizontes: MGUV 17165, 17166, 17286, 16449 y 17683 en Chera 0; MGUV 17552 en La Castellana 2 y MGUV 17721 en Chera 1c. Los especímenes más completos (figura 3.4-5) conservan la mayor parte de la diáfisis, aunque han perdido parte de los extremos proximal y distal.
En líneas generales, los húmeros recuperados muestran una diáfisis recta y estrecha, considerablemente expandida en sus extremos, especialmente en el extremo proximal, y presentan una cresta deltopectoral muy marcada, proyectada medialmente. La diáfisis presenta una sección transversal ovalada.
En los húmeros de Chera se aprecia cierta variabilidad morfológica, siendo algunos ejemplares más esbeltos que otros. Entre los titanosaurios europeos, Ampelosaurus atacis y, especialmente, Magyarosaurus dacus, presentan húmeros robustos, muy expandidos en sus extremos (Le Loeuff, 1992, 2005a, 2005b), contrastando con la gracilidad y esbeltez de los húmeros de Chera y Laño. Los húmeros de Lirainosaurus astibiae se cuentan entre los más esbeltos conocidos entre los titanosaurios, comparables por su gracilidad con los de Andesaurus (Calvo y Bonaparte, 1991), Rapetosaurus (Curry Rogers y Forster, 2001), y Maxakalisaurus topai (Kellner et al., 2006).
Ulna. Se han recuperado dos ulnas prácticamente completas pertenecientes a individuos diferentes (MGUV 17045, MGUV 17274). Como el resto de los huesos apendiculares de Lirainosaurus, las ulnas recuperadas son de constitución esbelta, provistas de una delgada diáfisis, ligeramente arqueada en vista lateral, que se expande considerablemente en su extremo proximal (figura 3.6-8). En vista proximal, este extremo es trirradiado. En esta región, la ulna presenta unos procesos lateral y medial muy prominentes, que delimitan una fosa para el acomodo del radio. El proceso posterior, menos desarrollado que los anteriores, se proyecta como una arista que recorre la cara posterior de la diáfisis, hasta desvanecerse cerca de su extremo distal. El olecranon es prominente. El extremo distal está escasamente expandido y presenta en su superficie anterior una protuberancia que marca el punto de contacto con el radio.
Las ulnas de titanosaurios presentan como principal característica un olecranon proyectado por encima de la superficie articular proximal (Wilson y Sereno, 1998; Wilson, 2002). Este carácter se observa en las ulnas de L. astibiae y en los ejemplares procedentes de Chera. La ulna de L. astibiae no fue incluida en la diagnosis inicial del género (Sanz et al., 1999) pero la preparación posterior de un ejemplar (MCNA 3157) permite comprobar que sendas ulnas presentan las mismas características morfológicas. Por este motivo, las ulnas de titanosaurio de Chera son las primeras en ser asignadas a este taxón. Entre los titanosaurios europeos, las ulnas de A. atacis y M. dacus presentan un configuración más robusta.
Fémur. En Chera 0 se han recuperado dos fémures izquierdos prácticamente completos (MGUV 17235, 17294), siendo este último ligeramente más pequeño que el anterior, pero de configuración más robusta (figura 4.1-5), y dos fémures derechos (MGUV 16450, 17194), incompletos por carecer de los extremos proximal y distal.

Figura 4. Lirainosaurus cf. astibiae, Formación Sierra Perenchiza, Cretácico Superior (Campaniano-Maastrichtiano), Chera, provincia de Valencia, España / Lirainosaurus cf. astibiae, Sierra Perenchiza Formation, Late Cretaceous (Campanian-Maastrichtian), Chera, Valencia province, Spain. 1-3, fémur izquierdo (MGUV 17235) en vistas posterior (1), medial (2) y anterior (3) / 1-3, left femur (MGUV 17235) in posterior (1), medial (2) and anterior (3) views. 4-5, fémur izquierdo (MGUV 17294) en vistas posterior (4) y anterior (5) / 4-5, left femur (MGUV 17294) in posterior (4) and anterior (5) views. Escala gráfica= 10 cm / scale bar = 10 cm.
Se trata de un hueso esbelto, con una diáfisis recta, notoriamente comprimida en sentido anteroposterior. Presenta extremos relativamente expandidos, siendo la extremidad distal, en promedio, un 60 % más ancha que la anchura mínima de la diáfisis (situada hacia la mitad de la longitud del hueso). Además de su aspecto grácil, las principales características del fémur son: trocánter mayor situado por debajo de la cabeza femoral, comba lateral en el tercio proximal de la diáfisis, por debajo del trocánter mayor, y presencia de un cuarto trocánter reducido, localizado cerca del borde medial de la mitad proximal de la diáfisis, ligeramente por encima de su punto más estrecho. El índice entre la anchura mínima de la diáfisis y la longitud total es de 1,6 en MGUV 17294, mientras que sólo alcanza un valor de 1,3 en MGUV 17235.
Distalmente, los cóndilos articulares son prominentes, con un cóndilo medial (tibial) más ancho que el cóndilo lateral (fibular). La orientación de los cóndilos es ligeramente oblicua con respecto al eje longitudinal del fémur.
Los saurópodos se caracterizan por presentar fémures rectos, con una diáfisis oval en sección transversal y con un cuarto trocánter poco desarrollado, en forma de cresta (McIntosh, 1990; Salgado et al., 1997; Wilson y Sereno, 1998). La presencia de una comba lateral por debajo del trocánter mayor es un sinapomorfía de los Titanosauriformes (Salgado et al., 1997; Wilson, 2002). El grado de desarrollo de esta comba es variable según los géneros (Salgado et al., 1997). En los fémures de Chera, así como en Lirainosaurus, el desarrollo relativo de la comba lateral es moderado, inferior al observado en formas como Brachiosaurus y Opisthocoelicaudia (Salgado et al., 1997: fig. 10).
Según Sanz et al. (1999), un cuarto trocánter reducido es un carácter que Lirainosaurus comparte con otros titanosaurios. La cabeza de los fémures de Chera y de Laño está situada muy por encima del trocánter mayor, siendo éste un carácter presente en eutitanosaurios, como Rapetosaurus y Saltasaurus. Tanto los fémures de Chera como los de Lirainosaurus de Laño presentan cóndilos distales orientados oblicuamente (y no paralelamente) con respecto al eje longitudinal de la diáfisis, proyectándose hasta la cara anterior del fémur. Algunos autores consideran que esta morfología es típica de los Saltasauridae (Wilson y Carrano, 1999; Wilson, 2002). Otro carácter presente en los fémures de Chera es la alta excentricidad de la sección transversal de la diáfisis, de forma que el diámetro lateromedial llega incluso a triplicar el valor del diámetro anteroposterior de la diáfisis, aunque se trata de una característica anatómica acentuada por la compresión litostática (tabla 2; véase también la figura 4.2).
Tabla 2. Medidas (en mm) de los diámetros máximo y mínimo de la diáfisis de los fémures de Lirainosaurus de los yacimientos de Chera (Valencia, España), medidos aproximadamente en el punto medio de la diáfisis, por debajo del cuarto trocánter. (*) Espécimen con fuerte compresión diagenética. (**) Excentricidad femoral definida como el cociente entre el diámetro lateromedial y el diámetro anterioposterior del fémur (Wilson y Carrano, 1999) / Measurements (in mm) of the maximum and minimum femoral diaphyseal diameters of the femora of Lirainosaurus from Chera (Valencia province, Spain), measured beneath the 4th trochanter, approximately half way down the shaft. (*) Specimen markedly flattened by diagenetic compression. (**) Femoral eccentricity calculated as mediolateral midshaft diameter divided by anteroposterior diameter (Wilson and Carrano, 1999).

Las diferencias de robustez entre los fémures de Chera podrían ser debidas a una variación individual o, incluso, representar un caso de dimorfismo. Un fenómeno similar se observa entre los fémures de Laño asignados a Lirainosaurus astibiae y ha sido también descrito en los fémures de Neuquensaurus australis (Salgado et al., 2005), reflejando aparentemente la natural variabilidad morfológica poblacional.
Discusión
El descubrimiento de material diagnóstico (vértebras caudales posteriores, escápula y coracoides) de Lirainosaurus en los yacimientos de Chera permite extender la distribución geográfica de este taxón al este de la Península Ibérica. Hasta la fecha, los restos fósiles conocidos de Lirainosaurus procedían exclusivamente del yacimiento de Laño (Sanz et al., 1999), ubicado en el norte de la Península Ibérica. Chera y Laño son localidades probablemente coetáneas, ya que comparten diversos taxones de vertebrados continentales (Company et al., 1999b; Company, 2004; Pereda Suberbiola et al., 2004), como son los dinosaurios Rhabdodon (Ornithopoda), Struthiosaurus (Ankylosauria) y Lirainosaurus (Titanosauria), los cocodrilos aligatoroideos Musturzabalsuchus y Acynodon (Buscalioni et al., 1999), pterosaurios azhdárquidos (Buffetaut, 1999; Company et al., 1999a), y las tortugas Dortoka, Polysternon y Solemys (Lapparent de Broin y Murelaga, 1999). Las asociaciones de carófitas y palinomorfos halladas en los niveles fosilíferos de Chera indican una edad Campaniano superior a Maastrichtiano inferior (Company, 2004; Company et al., 2005). Este resultado es compatible con los estudios de correlación estratigráfica realizados en la cantera de Laño y en áreas adyacentes del Sinclinorio Subcantábrico, cuyos niveles fosilíferos se han asignado a un intervalo de edad Campaniano superior-Maastrichtiano basal (Baceta et al., 1999), y cuya asociación palinológica sugiere una edad Campaniano superior (Núñez Betelu, 1999). En consecuencia, Lirainosaurus puede ser considerado un componente primordial de la fauna de dinosaurios del Campaniano superior-Maastrichtiano basal de la Península Ibérica.
Hasta la fecha, no se han descubierto restos fósiles de Lirainosaurus fuera de la Península Ibérica. Otros titanosaurios del Cretácico final europeo son: Ampelosaurus atacis del Campaniano superior-Maastrichtiano inferior de Languedoc, en Francia (Le Loeuff, 1995, 2005a) y Magyarosaurus dacus del Maastrichtiano de Transilvania (Nopcsa, 1915; Huene, 1932; Le Loeuff, 1993). El material provenzal de "Hypselosaurus priscus", descrito por Lapparent (1947), comparte con Lirainosaurus la presencia de centros caudales deprimidos dorsoventralmente y una cresta escapular ventral (Le Loeuff, 1992). No obstante, estos caracteres también están presentes en otros titanosaurios (Upchurch et al., 2004). Por otra parte, debido al carácter fragmentario y poco diagnóstico de sus restos fósiles, "Hypselosaurus priscus" es considerado un nomen dubium por Le Loeuff (1993) y Upchurch et al. (2004).
La pertenencia de Lirainosaurus al clado Titanosauria no ofrece dudas ya que comparte con ellos varias sinapomorfías, como son la presencia de pleurocelos acuminados en las vértebras dorsales y el carácter procélico de las vértebras caudales anteriores (Salgado et al., 1997; Sanz et al., 1999). No obstante, la posición filogenética de Lirainosaurus dentro de Titanosauria es objeto de debate. En este sentido, Sanz et al. (1999) consideran que Lirainosaurus es un miembro de Eutitanosauria, junto con Saltasaurus, Argyrosaurus y el titanosaurio hallado en Peirópolis, Brasil, Trigonosaurus pricei (Campos et al., 2005). En cambio, para Upchurch et al. (2004), Lirainosaurus es un Lithostrotia incertae sedis, siendo los Lithostrotia un clado compuesto por Malawisaurus y los titanosaurios más derivados (incluidos los Saltasauridae), todos ellos caracterizados por la presencia de vértebras caudales con centros procélicos y osificaciones dérmicas. Según Wilson y Upchurch (2003), los Eutitanosauria de Sanz et al. (1999) podrían ser un sinónimo de los Saltasauridae o Saltasaurinae (tabla 3). Estos autores siguen la definición de Sereno (1998) y consideran que son Saltasaurinae todas aquellas formas más estrechamente relacionadas con Saltasaurus que con Opithocoelicaudia. Para Wilson (2002), Lirainosaurus es un representante de los Saltasaurinae, clado que reúne a Saltasaurus y Neuquensaurus, ya que comparte con ellos al menos dos sinapomorfías: presencia de vértebras caudales con estructura interna trabecular o "en panal de abeja" y de centros caudales posteriores comprimidos dorsoventralmente, siendo el cóndilo dos veces más ancho que alto. Otra posible sinapomorfía compartida por Lirainosaurus y los Saltasaurinae (sensu Wilson, 2002) es la presencia de cóndilos distales proyectados parcialmente en la cara anterior del fémur. Estos caracteres también se observan en el material valenciano asignado a Lirainosaurus, con la salvedad de que el cóndilo de las vértebras caudales posteriores es únicamente 150 % más ancho que alto. Por otra parte, el coracoides de Chera carece de una marcada prominencia infraglenoidea, lo que lo diferencia del coracoides de los saltasáuridos (sensu Wilson, 2002).
Tabla 3. Definiciones filogenéticas de Saltasauridae, Saltasaurinae y Eutitanosauria en la literatura (véase Sereno, 2005). (N), taxón basado en el nodo; (S), taxón basado en la rama / Taxonomic definitions of the Saltasauridae, Saltasaurinae and Eutitanosauria from the published literature (see Sereno, 2005). (N), node-based taxon; (S), stem-based taxon.

Tal y como ha puesto de manifiesto Salgado (2003a), el significado taxonómico de los Saltasauridae y/o Saltasaurinae varía notablemente según los autores (tabla 3). Los Saltasauridae de Sereno (1998) y de Wilson y Upchurch (2003) serían prácticamente equivalentes de los Titanosauridae de Salgado et al. (1997), mientras que los Saltasauridae de Wilson (2002) son probablemente equivalentes de los Titanosauridae de Upchurch (1998). El clado Saltasaurinae de Curry-Rogers y Forster (2001) no es equivalente a los Saltasaurinae de Powell (2003), sino equiparables filogenéticamente a los Titanosauridae de Upchurch (1998), siendo un clado genéricamente más diverso que los Saltasauridae de Sereno (1998) (véase Salgado, 2003a). A diferencia de Upchurch et al. (2004), Curry Rogers (2005) considera que Lithostrotia es un clado incluido dentro de Saltasauridae. En este trabajo, Lirainosaurus es considerado un miembro de los Eutitanosauria de Sanz et al. (1999) siguiendo la definición propuesta por Salgado (2003a): "todos aquellos titanosaurios más próximos a Saltasaurus que a Epachthosaurus". En un reciente análisis filogenético, Calvo et al. (2007) interpretan que Lirainosaurus es un miembro de Eutitanosauria y el grupo hermano del clado Opisthocoelicaudiinae y Saltasaurinae.
Las supuestas afinidades de Lirainosaurus con los Saltasaurinae (sensu Wilson, 2002) podrían tener importantes implicaciones paleobiogeográficas, ya que se trata de un grupo de titanosaurios de porte modesto cuyos componentes provienen de Sudamérica. Según diversos autores (Salgado y Azpilicueta, 2000; Salgado, 2003b; Calvo y González Riga, 2003; Salgado et al., 2005; Bonaparte et al., 2006), el registro de Saltasaurinae (Saltasaurus, Neuquensaurus y Rocasaurus) se limitaría exclusivamente al Cretácico Superior (Campaniano-Maastrichiano de Argentina). Lirainosaurus comparte aparentemente algunos caracteres con los Saltasaurinae (véase más arriba), pero no posee varias sinapomorfías del clado (sensu Salgado et al., 1997; Salgado y Azpilicueta, 2000): los centros caudales están comprimidos dorsoventralmente, pero su anchura no supera el doble de la altura, y el borde anterior de la espina neural no parece estar ubicado posteriormente con respecto al margen anterior de las postzigapófisis en las caudales medias-posteriores. Además, la espina neural de las vértebras caudales anteriores de Lirainosaurus astibiae no está comprimida anteroposteriormente, a diferencia de los saltasaurinos (Bonaparte et al., 2006). Por estas razones, en este trabajo se descarta que Lirainosaurus sea un miembro de Saltasaurinae.
álisis filogenético realizado por Upchurch et al. (2004), los Lithostrotia (clado dentro del cual se inscribiría Lirainosaurus) son titanosaurios con una distribución casi cosmopolita durante el Cretácico, ya que se conocen con certeza en el Cretácico Inferior de África y en el Cretácico Superior de África, Asia, Europa, India, Madagascar, América del Norte y América del Sur (Upchurch et al., 2004). De acuerdo con esta hipótesis, no es necesario recurrir a un fenómeno de dispersión entre Sudamérica y Europa vía África durante el Cretácico Tardío (Le Loeuff, 1992, 1993) para explicar la presencia en el continente europeo de eutitanosaurios (saltasáuridos en el sentido de Wilson y Upchuch, 2003, véase tabla 3), como Lirainosaurus y Ampelosaurus, ya que miembros de este clado pudieron estar presentes en Europa desde, al menos, el Cretácico Temprano (Canudo y Salgado, 2003; Canudo y Cuenca-Bescós, 2004) habiéndose dispersado por Eurasia antes de que tuviera lugar el desmembramiento final de Laurasia y su separación de Gondwana.
Conclusiones
En este trabajo se describen por primera vez restos fósiles de saurópodos titanosaurios del Cretácico Superior de la provincia de Valencia (España). El material procede de los niveles continentales lacustres de la Formación Sierra Perenchiza del área de Chera, que han proporcionado, asimismo, fósiles de carófitas, palinomorfos, invertebrados y de otros vertebrados, como dinosaurios terópodos, ornitópodos y anquilosaurios, pterosaurios, cocodrilos, tortugas y peces actinopterigios. A partir de la asociación de microfósiles preservados, se interpreta que la fauna fósil de Chera es de edad Campaniano superior a Maastrichtiano inferior.
Lirainosaurus, género definido por presentar la siguiente combinación de caracteres: vértebras caudales posteriores con un cóndilo articular reducido y dividido por un surco sagital, estructura espinopostzigapofisisaria de las vértebras caudales medias y posteriores con una moderada proyección posterior, lámina escapular provista de una cresta ventral marcada en vista medial, y coracoides provisto de un foramen situado cerca de los márgenes dorsal y escapular. La aparente ausencia de una prominencia infraglenoidea en el coracoides, un carácter observado en Lirainosaurus astibiae, nos lleva a asignar provisionalmente el material de Chera a Lirainosaurus cf. astibiae. Material adicional descubierto en Chera (vértebras dorsales, placa esternal, húmeros, ulnas, fémures, etc.) es asignado al mismo taxón por aparecer asociado con el material diagnóstico y presentar características morfológicas análogas a las de sus equivalentes anatómicos de Lirainosaurus astibiae de Laño.
Además de los caracteres morfológicos, el moderado tamaño de los restos fósiles de Chera, pertenecientes a individuos adultos según sus características histológicas (Company, 2004), es coherente con su asignación a un titanosaurio de talla media (longitud adulta estimada: 8-10 m), como Lirainosaurus (véase Sanz et al., 1999).
Lirainosaurus es un componente de las faunas de dinosaurios del Campaniano superior-Maastrichtiano inferior de la Península Ibérica. Sus supuestas afinidades con los saltasaurinos (Saltasaurus, Neuquensaurus) son interesantes desde un punto de vista paleobiogeográfico, ya que este clado ha sido definido a partir de titanosaurios descubiertos en el Cretácico terminal de Argentina. No obstante, sus características permiten incluirlo dentro de Eutitanosauria y no de Saltasaurinae. La interpretación de Lirainosaurus como un miembro de los Eutitanosauria es coherente con la distribución geográfica cuasicosmopolita de este grupo de Lithostrotia durante el Cretácico.
Anexo 1
Material asignado a Lirainosaurus cf. astibiae: MGUV 17229 (fragmento de arco neural de vértebra dorsal), MGUV 17295 (vértebra dorsal), MGUV 17315 (vértebra dorsal), MGUV 17320 (vértebra dorsal), MGUV 16438 (vértebra caudal anterior), MGUV 17314 (vértebra caudal anterior), MGUV 16448 (vértebra caudal), MGUV 17016 (vértebra caudal posterior), MGUV 17027 (vértebra caudal), MGUV 17028 (vértebra caudal), MGUV 17028 (vértebra caudal), MGUV 17044 (vértebra caudal), MGUV 17090 (vértebra caudal), MGUV 17092 (centro de vértebra caudal), MGUV 17093 (centro de vértebra caudal), MGUV 17094 (vértebra caudal), MGUV 17095 (vértebra caudal), MGUV 17096 (vértebra caudal), MGUV 17097 (fragmento de vértebra caudal), MGUV 17098 (fragmento de vértebra caudal), MGUV 17100 (vértebra caudal), MGUV 17101 (vértebra caudal), MGUV 17105 (vértebra caudal), MGUV 17169 (vértebra caudal), 17243 (vértebra caudal), MGUV 17272 (vértebra caudal), MGUV 17280 (vértebra caudal), MGUV 17288 (vértebra caudal), MGUV 17337 (fragmento de vértebra caudal), MGUV 17539 (vértebra caudal), MGUV 17705 (vértebra caudal), CMPGC 49 (vértebra caudal), MGUV 17168 (arco hemal), MGUV 17192 (fragmento de arco hemal), MGUV 17332 (arco hemal), MGUV 17374 (fragmento de costilla), MGUV 16446 (fragmento de placa esternal), MGUV 17645 (escápula izquierda), MGUV 17238 (coracoides), MGUV 16449 (fragmento de húmero), MGUV 16454 (fragmento de húmero?), MGUV 17165 (fragmento de húmero), MGUV 17166 (fragmento de húmero), MGUV 17286 (fragmento distal de húmero), MGUV 17552 (húmero izquierdo), MGUV 17683 (fragmento de húmero), MGUV 17045 (ulna derecha), MGUV 17274 (ulna izquierda), MGUV 16450 (fémur derecho), MGUV 17194 (fémur derecho), MGUV 17235 (fémur izquierdo), MGUV 17294 (fémur izquierdo).
Agradecimientos
El segundo autor (X.P.S.) agradece al Ministerio de Educación y Ciencia (Ciencia y Tecnología) la concesión de una ayuda del Programa Ramón y Cajal para la realización de un proyecto de investigación sobre las faunas de vertebrados continentales del Cretácico final de la Península Ibérica en la Universidad del País Vasco/EHU (Bilbao). Este trabajo ha sido financiado por los proyectos CGL2004-02338/BTE y CGL2007-64061/BTE del Ministerio de Educación y Ciencia, por el Grupo Consolidado 9/UPV00121.310-15226/2003 de la UPV/EHU y por el grupo de investigación GIC07/14-361 del Gobierno Vasco. Los trabajos de campo han sido posibles gracias a las convocatorias anuales de ayudas para la realización de actuaciones paleontológicas promovidas por la Consellería de Cultura, Educación y Deporte de la Generalitat Valenciana. Financiación adicional para la realización de los trabajos de campo fue posible gracias a una ayuda a la investigación concedida por la Jurassic Foundation a J.C. La investigación de JIR-O forma parte del Protocolo CN-04-226 (Consejería de Cultura y Turismo/Universidad de Oviedo, Principado de Asturias) y del proyecto CGL 2007-62469. Por último, nuestro agradecimiento a los sufridos voluntarios participantes en las excavaciones y al siempre presente "innombrable", por hacer más estimulante nuestro trabajo. Finalmente, los autores desean agradecer a B.J. González-Riga (C.R.I.C.Y.T., C.O.N.I.C.E.T., Mendoza, Argentina) y a J.I. Canudo-Sanagustín (Universidad de Zaragoza, España) la atenta y enriquecedora revisión de este manuscrito.
Bibliografía
1. Alonso, A., Meléndez, N. y Mas, J.R. 1991. Sedimentación lacustre durante el Cretácico en la Cordillera Ibérica, España. Acta Geológica Hispánica 26: 35-54. [ Links ]
2. Alonso, A., Floquet, M., Mas, R., Meléndez, A., Meléndez, N., Salomon, J. y Vadot, J.-P. 1987. Modalités de la Régression marine sur le détroit Ibérique (Espagne) à la fin du Crétacé. Mémoires Géologiques de l'Université de Dijon 11: 91-102. [ Links ]
3. Baceta, J.I., Pujalte, V. y Orue-Etxebarria, X. 1999. The vertebrate fossil-bearing sites of the Laño quarry (Basque-Cantabrian Region): Stratigraphical and palaeogeographical context. Estudios del Museo de Ciencias Naturales de Álava 14 (Número especial 1): 13-28. [ Links ]
4. Behrensmeyer, A.K. 1991. Terrestrial vertebrate accumulations. En: P.A. Allison, y D.E. Briggs, (eds.), Taphonomy. Plenum Press, New York. pp. 291-335. [ Links ]
5. Bonaparte, J.F. y Coria, R.A. 1993. Un nuevo y gigantesco saurópodo titanosaurio de la Formación Río Limay (Albiano-Cenomaniano) de la Provincia del Neuquén, Argentina. Ameghiniana 30: 271-282. [ Links ]
6. Bonaparte, J.F., González-Riga, B.J. y Apesteguía, S. 2006. Ligabuesaurus leanzai gen. et sp. nov. (Dinosauria, Sauropoda), a new titanosaur from the Lohan Cura Formation (Aptian, Lower Cretaceous) of Neuquén, Patagonia, Argentina. Cretaceous Research 27: 364-376. [ Links ]
7. Buffetaut, E. 1999. Pterosauria from the Upper Cretaceous of Laño (Iberian Peninsula): a preliminary comparative study. Estudios del Museo de Ciencias Naturales de Álava 14 (Número especial 11): 289-294. [ Links ]
8. Borsuk-Bialynicka, M. 1977. A new camarasaurid sauropod Opisthocoelicaudia skarynskii, gen. n. sp. n. from the Upper Cretaceous of Mongolia. Palaeontologica Polonica 37: 5-64. [ Links ]
9. Bravo, A.M., Vila, B., Galobart, A. y Oms, O. 2005. Restos de huevos de dinosaurio en el Cretácico superior del Sinclinal de Vallcebre (Berguedà, provincia de Barcelona). Revista Española de Paleontología Número extraordinario 10: 49-57. [ Links ]
10. Burton, A. 1998. [ A taphonomic study of the Wealden Beds of Southern Britain. Ph.D. thesis, Cambridge University, Unpublished.] World Wide Web: http://www.conrad41.free-serve.co.uk/Home.htm [ Links ]
11. Buscalioni, A.D., Ortega, F. y Vasse, D. 1999. The Upper Cretaceous crocodilian assemblage from Laño (northcentral Spain): Implications in the knowledge of the finicretaceous European faunas. Estudios del Museo de Ciencias Naturales de Álava (Número Especial 1) 14: 213-233. [ Links ]
12. Calvo, J.O. y Bonaparte, J.F. 1991. Andesaurus delgadoi gen et sp. nov. (Saurischia-Sauropoda) dinosaurio Titanosauridae de la Formacion Rio Limay (Albiano-Cenomaniano), Neuquen, Argentina. Ameghiniana 28: 303-310. [ Links ]
13. Calvo, J.O. y González Riga, B.J. 2003. Rinconsaurus caudamirus gen. et sp. nov., a new titanosaurid (Dinosauria, Sauropoda) from the Late Cretaceous of Patagonia, Argentina. Revista Geológica de Chile 30: 333-353. [ Links ]
14. Calvo, J.O., Porfiri J.D., González-Riga, B.J. y Kellner, A.W.A. 2007. A new Cretaceous terrestrial ecosystem from Gondwana with the description of a new sauropod dinosaur. Anais da Academia Brasileira de Ciências 79: 529-541 [ Links ]
15. Campos, D. de A., Kellner, A.W.A., Bertini, R.J. y Santucci, R.M. 2005. On a titanosaurid (Dinosauria, Sauropoda) vertebral column from the Bauru Group, Late Cretaceous of Brazil. Arquivos do Museu Nacional, Rio de Janeiro 63: 565-593. [ Links ]
16. Canudo, J.I. 2001. Descripción de un fragmento proximal de fémur de Titanosauridae (Dinosauria, Sauropoda) del Maastrichtiano superior de Serraduy (Huesca). En: G. Meléndez, Z. Herrera, G. Delvene y B. Azanza (eds.), 17º Jornadas de la Sociedad Española de Paleontología: Los fósiles y la paleogeografía. Publicaciones del Seminario de Paleontología de Zaragoza (SEPAZ) 5: 255-262. [ Links ]
17. Canudo, J.I. y Cuenca-Bescós, G. 2004. Morphometric approach to Titanosauriformes (Sauropoda, Dinosauria) femora: Implications to the paleobiogeographic analysis. En: A.M.T. Elewa (ed.), Morphometrics, Applications in Biology and Paleontology, Springer Verlag, pp. 143-156. [ Links ]
18. Canudo, J.I. y Salgado, L. 2003. Los dinosaurios del Neocomiense (Cretácico inferior) de la Península Ibérica y Gondwana occidental: implicaciones paleobiogeográficas. En: F. Pérez-Lorente (Coord.), Dinosaurios y otros reptiles mesozoicos de España. (IER, Ciencias de la Tierra, 26) Instituto de Estudios Riojanos, Logroño, pp. 251-268. [ Links ]
19. Casanovas, M.L. 1993. Novedades en el registro fósil de dinosaurios del Levante español. Zubia 10 [ correspondiente a 1992]: 139-151. [ Links ]
20. Casanovas, M.L. y Santafé, J.V. 1993. Presencia de Titanosáuridos (Dinosauria) en el Cretácico Superior de Fontllonga (Lleida, España). Treballs del Museo de Geologia de Barcelona 3: 67-80. [ Links ]
21. Casanovas, M.L., Santafé, J.V., Sanz, J.L. y Buscalioni, A.D. 1987. Arcosaurios (Crocodilia, Dinosauria) del Cretácico superior de la Conca de Tremp (Lleida, España). Estudios geológicos volumen extraordinario Galve-Tremp: 95-110. [ Links ]
22. Casanovas, M.L., Santafé, J.V., Sanz, J.L. y Powell, J.E., 1995. Nuevos restos de dinosaurios (Titanosauria y Ornithopoda) en el Cretácico superior de las Cuencas de Tremp y Dellà (Lleida, España). Estudios geológicos 51: 277-283. [ Links ]
23. Company, J. 2004. [ Vertebrados continentales del Cretácico superior (Campaniano-Maastrichtiano) de Valencia. Tesis doctoral, Universitat de València, 410 pp. Inédita. [ Links ]].
24. Company, J., Ruiz-Omeñaca, J.I., y Pereda Suberbiola, X. 1999a. A longnecked pterosaur (Pterodactyloidea, Azhdarchidae) from the Upper Cretaceous of Valencia, Spain. Geologie en Mijnbouw 78: 319-333. [ Links ]
25. Company, J., Murelaga, X., Pereda Suberbiola, X. y Ruiz-Omeñaca, J.I. 1999b. The vertebrate fauna from the new Late Cretaceous Chera locality (Valencia Province, Spain). En: J.I. Canudo y G. Cuenca-Bescós (eds.), 4º European Workshop on Vertebrate Palaeontology, Albarracín (Teruel, Spain, 1999). Programme and Abstracts, Field Guide, pp. 37-38. [ Links ]
26. Company, J., Feist, M., Peyrot, D., Barrón, E., Robles, F., Pereda Suberbiola, X. y Ruiz-Omeñaca, J.I. 2005. Stratigraphic position and palaeoenvironmental traits of the Late Cretaceous vertebrate-bearing sites of Chera (Valencia, Spain), based on micropalaeontological data. Kaupia (Darmstädter Beiträge zur Naturgeschichte) 14: 76. [ Links ]
27. Curry-Rogers, K. 2005. Titanosauria. A phylogenetic overview. pp. 50-103. En: K. Curry-Rogers y J.A. Wilson (eds.), The Sauropods. Evolution and Paleobiology, University of California Press, Berkeley, Los Angeles and London, pp. 115-137. [ Links ]
28. Curry-Rogers, K. y Forster, C.A. 2001. The last of the dinosaur titans: a new sauropod from Madagascar. Nature 412: 530-534. [ Links ]
29. Floquet, M. 1991. La plateforme nord-castillane au Crétacé supérieur (Espagne). Arrièrepays ibérique de la marge passive bascocantabrique, sédimentation et vie. Memoires Geólogiques de l'Universite de Dijon 14, 1-925. [ Links ]
30. González Riga, B.J. 2003. A new titanosaur (Dinosauria, Sauropoda) from the Upper Cretaceous of Mendoza Province, Argentina. Ameghiniana 40: 155-172. [ Links ]
31. Gutiérrez, G., Meléndez, A., Robles, F., y Usera, J. 1975a. El Cretácico Superior de la Sierra Perenchiza (Valencia). 1º Coloquio de Estratigrafía y Paleogeografía del Cretácico de España 7: 151-158. [ Links ]
32. Gutiérrez, G., Meléndez, A., Robles, F., y Usera, J. 1975b. El Cretácico Superior continental de la Sierra de Besorí (Valencia). Estudios Geológicos 31: 567-570. [ Links ]
33. Hill, A. 1979. Disarticulation and scattering of mammal skeletons. Paleobiology 5: 261-274. [ Links ]
34. Huene, F. 1932. Die fossile Reptil-Ordnung Saurischia, ihre Entwicklung und Geschichte. Monographien zur Geologie und Palaeontologie 4: 1-361. [ Links ]
35. Hunt, A.P., Lockley, M.G., Lucas, S.G. y Meyer, C.A. 1994. The global sauropod fossil record. En: M.G. Lockley, V.F. dos Santos, C.A. Meyer y A.P. Hunt (eds.), Aspects of Sauropod Paleobiology. Gaia 10: 261-279. [ Links ]
36. Kellner, A.W.A., Campos, D.A. y Trotta, M.N.F. 2005. Description of a titanosaurid caudal series from the Bauru Group, Late Cretaceous of Brazil. Arquivos do Museum Nacional, Río de Janeiro 63: 529-564. [ Links ]
37. Kellner, A.W.A., Campos, D.A., Azevedo, S.A.K., Henriques, D.D.R., Craik, M.M.T. y Silva, H.P. 2006. A new titanosaur sauropod from the Bauru Group, Late Cretaceous of Brazil. Boletim do Museu Nacional Nova Série, Geologia 74: 1-31. [ Links ]
38. Kidwell, S.M., Fürsich, F.T. y Aigner, T. 1986. Conceptual framework for the analysis and classification of fossil concentrations. Palaios 1: 228-238. [ Links ]
39. Lapparent, A.F. 1947. Les dinosauriens du Crétacé supérieur du Midi de la France. Mémoires de la Société géologique de France 56: 1-54. [ Links ]
40. Lapparent, A.F. y Aguirre, E. 1956. Algunos yacimientos de dinosaurios en el Cretácico superior de la Cuenca de Tremp. Estudios Geológicos 12: 31-32. [ Links ]
41. Lapparent de Broin, F. y Murelaga, X. 1999. Turtles from the Upper Cretaceous of Laño (Iberian Peninsula). Estudios del Museo de Ciencias Naturales de Álava 14 (Número especial 1): 135-211. [ Links ]
42. Lécuyer, C., Bogey, C., Garcia, J.-P., Grandjean, P., Barrat, J.-A., Floquet, M., Bardet, N. y Pereda Suberbiola, X. 2003. Stable isotope composition and rare earth element content of vertebrate remains from the Late Cretaceous of northern Spain (Laño): did the environmental record survive? Palaeogeography Palaeoclimatology Palaeoecology 193: 457-471 (Corrigendum: Palaeogeography Palaeoclimatology Palaeoecology 196: 427-428). [ Links ]
43. Le Loeuff, J. 1992. [ Les vertébrés continentaux du Crétacé supérieur d'Europe: Paléoécologie, Biostratigraphie et Paléobiogéographie. Mémoires des Sciences de la Terre, Tesis doctoral, Université Pierre et Marie Curie, nº 92-3, Paris, 273 pp. Inédita. [ Links ]].
44. Le Loeuff, J. 1993. European titanosaurids. Revue de Paléobiologie volume spécial 7: 105-117. [ Links ]
45. Le Loeuff, J. 1995. Ampelosaurus atacis (nov. gen., nov. sp.), un nouveau Titanosauridae (Dinosauria, Sauropoda) du Crétacé supérieur de la haute vallée de l'Aude (France). Comptes Rendus de l'Académie des Sciences Paris 321: 693-696. [ Links ]
46. Le Loeuff, J. 2005a. Osteology of Ampelosaurus atacis (Titanosauria) from southern France. En: V. Tidwell y K. Carpenter (eds.), Thunder-Lizards, The Sauropodomorph Dinosaurs, Indiana University Press, Bloomington and Indianapolis, pp. 115-137. [ Links ]
47. Le Loeuff, J. 2005b. Romanian Late Cretaceous dinosaurs: big dwarfs or small giants? Historical Biology 17: 15-17. [ Links ]
48. Le Loeuff, J. y Martínez, A. 1997. Afloramiento de icnitas de Titanosauridae en la zona de Fumanya (Maastrichtiense, Pirineo oriental): Estudio preliminar. Geogaceta 21: 151-153. [ Links ]
49. López-Martínez, N. 2000. Eggshell sites from the Cretaceous-Tertiary transition in South-Central Pyrenees (Spain). En: A.M Bravo y T. Reyes (eds.), 1º International Symposium on Dinosaur Eggs and Babies. Extendend Abstracts, 95-115. [ Links ]
50. López-Martínez, N., Moratalla, J.J. y Sanz, J.L. 2000. Dinosaur nesting on tidal flats. Palaeogeography, Palaeoclimatology, Palaeoecology 160: 153-163. [ Links ]
51. McIntosh, J.S. 1990. Sauropoda. En: D.B. Weishampel, P. Dodson y H. Osmólska (eds.), The Dinosauria, 1st ed., University of California Press, Berkeley and Los Angeles, pp. 345-401. [ Links ]
52. Marsh, O.C. 1878. Principal characters of American Jurassic dinosaurs. Part I. American Journal of Science 16: 411-416. [ Links ]
53. Masriera, A. y Ullastre, J. 1988. Nuevos datos sobre las capas maestrichtienses con Septorella: su presencia al norte del Montsec (Pirineo catalán). Acta Geológica Hispana 23: 71-77. [ Links ]
54. Moratalla-García, J.J. 1993. [ Restos indirectos de dinosaurios del registro español. Paleoicnología de la Cuenca de Cameros (Jurásico superior-Cretácico inferior) y Paleoología del Cretácico superior. Tesis doctoral, Universidad Autónoma de Madrid, 727 pp. Inédita. [ Links ]].
55. Nopcsa, F. 1915. Die Dinosaurier der Siebenbürgischen Landesteile Ungarns. Mitteilungen aus dem Jahrbuch der königlichen ungarischen geologischen Reichsanstalt 23: 1-26. [ Links ]
56. Núñez Betelu, K. 1999. Preliminary palynological assessment of the vertebrate-rich Laño beds: Age and paleoenvironment. Estudios del Museo de Ciencias Naturales de Álava 14 (Número especial 1): 37-42. [ Links ]
57. Pereda Suberbiola, X. y Ruiz-Omeñaca, J.I., 2002. Un dinosaurio saurópodo (Titanosauria) en el Cretácico superior de Cubilla, Soria (España). Geogaceta 30 [ correspondiente a 2001]: 175-178. [ Links ]
58. Pereda Suberbiola, X., Company, J. y Ruiz-Omeñaca, J.I. 2004. Dinosaurios y otros vertebrados continentales del Cretácico final (Campaniense-Maastrichtiense) de la Península Ibérica: Composición y sucesiones faunísticas. Geo-Temas 6: 55-58. [ Links ]
59. Pereda Suberbiola, X., Murelaga, X., Baceta, J.I., Corral, J.C., Badiola, A. y Astibia, H. 1999. Nuevos restos fósiles de vertebrados continentales en el Cretácico Superior de Álava (Región Vasco-Cantábrica): sistemática y posición estratigráfica. Geogaceta 26: 79-82. [ Links ]
60. Platt, N.H. y Wright, V.P. 1991. Lacustrine carbonates, facies distributions and hydrocarbon aspects, pp. 55-73. En: P. Anadón, L. Cabrera y K. Kelts (eds.), Lacustrine Facies Analysis (International Association of Sedimentologists, Special Publication 13). Blackwell Scientific, Oxford. [ Links ]
61. Platt, N.H. y Wright, V.P. 1992. Palustrine Carbonates and the Florida Everglades: towards an exposure index for the fresh-water environment? Journal of Sedimentary Petrology 62: 1058-1071. [ Links ]
62. Powell, J.E. 1992. Osteología de Saltasaurus loricatus (Sauropoda, Titanosauridae) del Cretácico superior del noroeste argentino. En: J.L. Sanz y A.D. Buscalioni (eds.), Los dinosaurios y su entorno biótico, Actas del 2º Curso de Paleontología en Cuenca, Instituto Juan de Valdés, Cuenca, España, pp. 165-230. [ Links ]
63. Powell, J.E. 2003. Revision of South American titanosaurid dinosaurs: palaeobiological, palaeobiogeographical and phylogenetic aspects. Records of the Queen Victoria Museum 111: 1-173. [ Links ]
64. Royo-Torres, R. y Canudo, J.I. 2003. Restos directos de dinosaurios saurópodos en España (Jurásico Superior-Cretácico Superior). En: F. Pérez-Lorente (coord.), Dinosaurios y otros reptiles mesozoicos en España, (IER, Ciencias de la Tierra, 26). Instituto de Estudios Riojanos, Logroño, pp. 313-334. [ Links ]
65. Salgado L. 2003a. Should we abandon the name Titanosauridae? Some comments on the taxonomy of titanosaurian sauropods (Dinosauria). Revista Española de Paleontología 18: 15-21. [ Links ]
66. Salgado, L. 2003b. Los saurópodos de Patagonia: Sistemática, evolución y paleobiología. En: Colectivo Arqueológico-Paleontológico de Salas (eds.), Actas de las 2º Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno, Salas de los Infantes (Burgos), pp. 139-168. [ Links ]
67. Salgado, L. y Azpilicueta, C. 2000. Un nuevo saltasaurino (Sauropoda, Titanosauridae) de la Provincia de Río Negro (Formación Allen, Cretácico superior), Patagonia, Argentina. Ameghiniana 37: 259-264. [ Links ]
68. Salgado, L., Apesteguía, S. y Heredia, S.E. 2005. A new specimen of Neuquensaurus australis, a Late Cretaceous saltasaurine titanosaur from North Patagonia. Journal of Vertebrate Paleontology 25: 623-634. [ Links ]
69. Salgado, L., Coria, R.A. y Calvo, J.O. 1997. Evolution of titanosaurid sauropods. I: Phylogenetic analysis based on the postcranial evidence. Ameghiniana 34: 3-32. [ Links ]
70. Sanz, J.L. y Buscalioni, A.D. 1987. New evidence of armoured dinosaurs in the Upper Cretaceous of Spain. En: P.J. Currie y E.H. Koster (eds.), 4º Symposium on Mesozoic Terrestrial Ecosystems, Occasional Papers of the Tyrrell Museum of Paleontology 3: 199-204. [ Links ]
71. Sanz, J.L., Buscalioni, A.D., Pérez-Moreno, B.P., Moratalla, J.J. y Jiménez García, S. 1992. Los dinosaurios de Castilla y León. En: E. Jiménez Fuentes (ed.), Vertebrados fósiles de Castilla y León, Museo Provincial de Salamanca, Salamanca, pp. 47-57. [ Links ]
72. Sanz, J.L., Powell, J.E., Le Loeuff, J., Martínez, R. y Pereda Suberbiola, X. 1999. Sauropod remains from the Upper Cretaceous of Laño (northcentral Spain). Titanosaur phylogenetic relationships. Estudios del Museo de Ciencias Naturales de Álava 14 (Número especial 1): 235-255. [ Links ]
73. Schulp, A.S. y Brokx, A.W. 1999. Maastrichtian sauropod foot-prints from the Fumanya site, Bergueda, Spain. Ichnos 6: 239-250. [ Links ]
74. Sereno, P.C. 1998. A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria. Neues Jarhbuch für Geologie und Paläontologie Abhandlungen 210: 41-83. [ Links ]
75. Sereno, P.C. 2005. TaxonSearch. Stem Archosauria [ version 1.0, November 07, 2005. World Wide Web: http://www.taxonse-arch.org/Archive/stem-archosauria-1.0.php] [ Links ]
76. Upchurch, P. 1998. The phylogenetic relationships of sauropod dinosaurs. Zoological Journal of the Linnean Society of London 124: 43-103. [ Links ]
77. Upchurch, P., Barrett, P.M. y Dodson, P. 2004. Sauropoda. En: D.B. Weishampel, P. Dodson y H. Osmólska (eds.), The Dinosauria, 2º ed., University of California Press, Berkeley, Los Angeles, London, pp. 259-322 [ suplemento: http://dinosau-ria.ucpress.edu] [ Links ]
78. Vila, B., Oms, O. y Galobart, A. 2005. Manusonly titanosaurid trackway from Fumanya (Maastrichtian, Pyrenees): further evidence for an underprint origin. Lethaia 38: 211-218. [ Links ]
79. Vilas, L., Mas, R., García, A., Arias, C., Alonso, A., Meléndez, N. y Rincón, R. 1982. Ibérica Suroccidental. En: A. Alonso, C. Arias, A. García, R. Mas, R. Rincón y L. Vilas (eds.), El Cretácico de España, Universidad Complutense de Madrid, pp. 457-513 [ Links ]
80. Wilson, J.A. 2002. Sauropod dinosaur phylogeny: critique and cladistic analysis. Zoological Journal of the Linnean Society 136: 217-276. [ Links ]
81. Wilson, J.A. 2005. Overview of Sauropod Phylogeny and Evolution. En: K.A. Curry Rogers y J.A. Wilson (eds.), The Sauropods: Evolution and Paleobiology, University of California Press, 15-49. [ Links ]
82. Wilson, J.A. y Carrano, M.T. 1999. Titanosaurs and the origin of "wide-gauge" trackways: a biomechanical and systematic perspective on sauropod locomotion. Paleobiology 25: 252-267. [ Links ]
83. Wilson, J.A. y Sereno, P.C. 1998. Early evolution and higher-level phylogeny of sauropod dinosaurs. Journal of Vertebrate Paleontology 18: 1-68. [ Links ]
84. Wilson, J.A. y Upchurch, P. 2003. A revision of Titanosaurus Lydekker (Dinosauria-Sauropoda), the first dinosaur genus with a "Gondwanan" distribution. Journal of Systematic Palaeontology 1: 125-160. [ Links ]
85. Wilson, J.A., Martínez, R.N. y Alcober, O. 1999. Distal tail segment of a titanosaur (Dinosauria: Sauropoda) from the Upper Cretaceous of Mendoza, Argentina. Journal of Vertebrate Paleontology 19: 591-594. [ Links ]
Recibido: 27 de diciembre de 2007.
Aceptado: 25 de marzo de 2009.

