










Services on Demand
Journal

Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAmeghiniana
On-line version ISSN 1851-8044
Ameghiniana vol.46 no.3 Buenos Aires July/Sept. 2009
ARTÍCULOS ORIGINALES
A new angiosperm leaf morphotype from the Early Cretaceous (Late Aptian) of San Luis Basin, Argentina
Gabriela G. Puebla1
1Unidad de Paleopalinología. Instituto Argentino de Nivología y Glaciología, Centro Científico Tecnológico. CONICET. C.C.131, 5500 Mendoza, Argentina. gpuebla@lab.cricyt.edu.ar
Abstract. A new fossil angiosperm leaf morphotype, "LC-Microphyll trifoliate", from the Late Aptian La Cantera Formation, central western Argentina, is described. The leaf remains occurred at the type section of the formation within a low diversity macrofloral association. Fossils are found mainly as isolated small size leaflets (0.3-1.5 cm long). The leaves are pinnately compound (imparipinnate, trifoliate), the leaflets have pinnately lobed margin, first vein category pinnate, secondary veins craspedodromous opposite to subopposite ending at the apex of lobes and an intramarginal vein. "LC-Microphyll trifoliate" is among the earliest record of leaf remains possibly allied to the eudicots and one of the first evidence of angiosperm with compound leaves.
Resumen. Nuevo morfotipo de hoja de angiosperma del Cretácico Temprano (Aptiano Tardío), Cuenca de San Luis, Argentina. Se describe un nuevo morfotipo de hoja de angiosperma fósil, "LC- Microfila trifoliada", procedente de la Formación La Cantera (Aptiano Tardío), centro oeste de Argentina. Los restos de hojas fueron hallados en la sección tipo de la formación formando parte de una asociación macrofloral de baja diversidad. Los fósiles se encuentran en general como folíolos aislados de tamaño pequeño (0,3- 1,5 cm. long). Las hojas son pinnaticompuestas (imparipinnadas, trifoliadas), los folíolos tienen el margen pinnatilobado, la venación primaria es pinnada, la venas secundarias son craspedódromas, opuestas a subopuestas finalizando en el ápice de los lóbulos y en una vena intramarginal. "LC-Microfila trifoliata" es uno de los más tempranos registros de hojas fósiles posiblemente vinculado con las eudicotiledóneas, y una de las primeras evidencias de angiospermas con hojas compuestas.
Key words. Angiosperm; Eudicots; Late Aptian; La Cantera Formation; Argentina.
Palabras clave. Angiospermas; Eudicotiledóneas; Aptiano tardío; Formación La Cantera; Argentina.
Introduction
The Cretaceous is considered to be crucial for understanding the evolution of the angiosperms. Angiosperm radiation is one of the major evolutionary events in Earth history, and therefore one of the ways to understand and characterize it is through the study of fossil leaves and pollen grains, that can be unequivocally assigned as belonging to the angiosperms. The oldest unequivocal angiosperm remains, mostly dispersed structures, are from Lower Cretaceous strata (Taylor and Hickey, 1990; Sun and Dilcher, 2002; Friis et al., 2006).
The Cretaceous history of the angiosperms in southern South America is based mainly on the records from Patagonian basins (Archangelsky et al. 2004, 2009). Very little is known on the fossil record of angiosperms that illustrates their evolution during the Cretaceous in central Argentina and their relationships with the rest of South America. The La Cantera Formation (Late Aptian), which crops out at the northwest of the San Luis Province, is one of the few Argentinean lithostratigrahic units that contain Cretaceous fossil plants. This formation is included within the Del Gigante Group (Flores and Criado Roque, 1972) and the sections of this group are exposed in a restricted area on the eastern side of the southern tip of the Sierra del Gigante (figure 1). It corresponds to the continental filling of an extensional basin, related to the break up of Gondwana, and represents ephemeral lagoons associated with a fluvial system (Rivarola and Spalletti, 2006). Reports on the fossil content of the La Cantera Formation at Sierra del Gigante are known since the beginning of the twentieth century and include macro and microflora, insects and fishes remains (Fossa Mancini, 1939; Flores, 1969; Mazzoni, 1985; Mazzoni and Hünicken, 1984, 1987; Arcucci et al., 2002). The presence of angiosperms in the La Cantera Formation is known from both pollen (Prámparo 1990, 1994, 1999b) and leaf records. Although some fossil plants were recovered from the San Luis Basin in the past, there is only one published contribution by Hünicken and Romero (1981) on fossil leaves of the La Cantera Formation, however these authors did not include illustrations of the fossils.
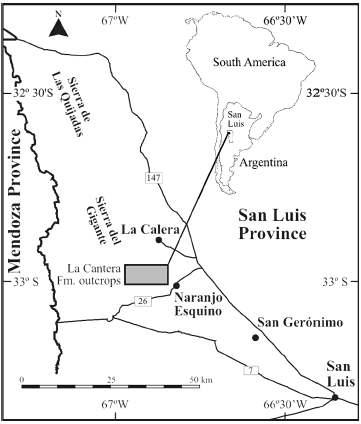
Figure 1. Location map of the La Cantera Formation, at the northwest of the San Luis Province, Argentina / mapa de ubicación de la Formación La Cantera al noroeste de la provincia de San Luis.
There are scarce records of Late Aptian fossil angiosperms worldwide and only a few reporting the occurrence of both macro and microfossils (palynomorphs) in the same deposit. From the Early Cretaceous, well-preserved floras with flowers, fruits, seeds, and stamens and pollen are known from North America, the Potomac Group sequence (Hickey and Doyle, 1977) and from Portugal (Friis et al., 2006). The fossil remains of the La Cantera Formation and the Baqueró Group, southern Deseado Massif (Del Fueyo et al., 2007) are among the most representatives with those characteristics in Argentina.
Besides Argentina, another place with both occurrences of plants and palynomorphs of Aptian age in South America is the Late Aptian or Early Albian Brazilian Crato Formation (Maisey, 1991; Martill et al., 1993; Mohr and Friis, 2000; Mohr and Rydin, 2002; Mohr and Eklund, 2003; Mohr and Bernardes-de- Oliveira, 2004; Mohr et al., 2006). The Crato Formation provides one of the most complete Early Cretaceous macrofloral associations in number of specimens and diversity, including roots, stems, leaves and reproductive structures. This Brazilian flora is important for further correlations with the La Cantera Formation.
Several outcrops with Early Cretaceous fossil floras were studied in Santa Cruz and Chubut provinces, southern Argentina (see Del Fueyo et al., 2007 for a review).
Nevertheless, the La Cantera Formation (San Luis basin) is the only one, bearing fossil plants of Early Cretaceous age in the western part of central Argentina (Hünicken and Romero, 1981; Puebla, 2004).
In this contribution, a new leaf morphotype collected at the La Cantera Formation at its type locality is described.
Geology and age
The fossil remains studied herein were collected in the La Cantera Formation that crops out in the San Luis Basin, 85 km northwest from San Luis city, San Luis Province Argentina (figure 1).
The La Cantera Formation is part of the Del Gigante Group (Flores and Criado, 1972) that consists of five formations sensu Flores (1969): Los Riscos, El Jume, La Cantera, El Toscal and La Cruz, or six formations, the five just mentioned and the Lagarcito Formation as suggested by Rivarola and Di Paola (1992).
The La Cantera Formation is mainly composed of green-grey siltstone and claystone, with red sandstone and claystone interbedded at the top (figure 2). It could represent ephemeral lagoons related to a fluvial environment with some periods of a very quiet lacustrine system (Criado et al., 1981; Prámparo, 1989a), which allowed the preservation of delicate structures such as leaves, stems and flowers. Gypsum is interbedded with clastics at the top of the unit, and corresponds to the final stage of the lacustrine deposition. Based on palynological data, the age suggested for these strata is Late Aptian (Prámparo, 1990, 1994, 1999a, 1999b, Prámparo et al., 2007).
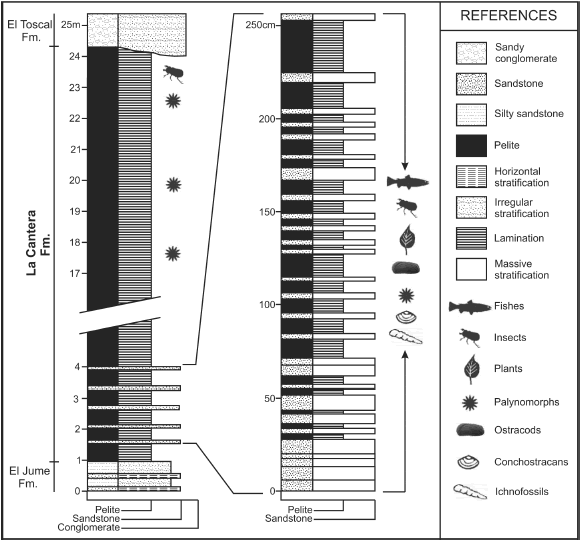
Figure 2. Columnar stratigraphic section of the La Cantera Formation in the San Luis Basin / perfil estratigráfico de la Formación La Cantera en la Cuenca de San Luis.
The fossil specimens were collected at the type section of the La Cantera Formation (at 32º 59' 25"S, 66º 52' 48"W) from the greenish brown siltstones of its lower part. The plant rich levels are developed in cyclically repeated millimetric laminated sediments (figure 2).
Materials and methods
About 250 specimens have been collected from the La Cantera Formation, and include leaves, stems and reproductive structures. These remains have been found in fine laminated shales as coalified compressions/impressions without preserved cuticles.
The leaf remains are most frequently found as isolated leaflets and are of small size, (about 3 cm long). The majority of the leaves belong to angiosperms and up to now three different morphotypes were identified (Puebla, 2003; Archangelsky et al., 2004).
In this report, 28 specimens of a single morphotype collected from the type section are studied. The fossils were studied using a stereomicroscope Leica Mz 125 and photographed with a Nikon Coolpix 990 digital camera.
The description of the macrofossils follows the Manual of Leaf Architecture (Leaf Architecture Working Group, 1999) that incorporates the terminology proposed by Hickey (1973), Dilcher (1973) and Hickey and Wolfe (1975). The description focuses on general morphology and venation patterns. Morphotypes are considered as artificial categories applied to fossils that cannot be placed with any modern taxa, and are commonly used for Cretaceous floras (e.g. Crabtree, 1987) and ranks are basically evolutionary degrees that represent adaptative advances in the evolution of the angiosperms (Hickey, 1977) and have been previously established. Emphasis has been put in determining the phylogenetic relationships of the fossils with modern taxa.
The fossil specimens are housed at the Paleontological Collection of the Universidad Nacional de San Luis, Departamento de Geología, under the initials MHIN-UNSL-GEO-P483; P484; P461-P464; P472- P477; P7; P41; P42; P46; P48; P56; P61; P62; P64; P80; P81; P89; P94; P111; P119; P209.
Systematic paleontology
LC-Microphyll trifoliate
(figures 3, 4)
Figure 3. "LC-Microphyll trifoliate" (bar= 0.5 cm). 1, 3 and 5 morphotype example (MHIN- UNSL-GEO-P483), 1, Complete specimen of "LC-Microphyll trifoliate"with three leaflets. 3, Line drawings based on the morphotype example. 5, Detail of the venation pattern: Intramarginal vein (black arrow), tertiary veins (white arrow) and intersecondary veins (discontinuous white arrow). 2, 4 and 6 (MHINUNSL- GEO-P461), 2, Isolated leaflet having the lobe apex with glandular-like tips (white arrow indicates gland). 4, Line drawings based on (MHIN-UNSL-GEO-P461). 6, Detail of the lobe apex with glandular-like tips / "LC-Microfila trifoliata" (escala= 0. 5 cm). A, C y E morfotipo ejemplo (MHIN- UNSL-GEO-P483), 1, especimen completo de "LC-Microfila trifoliata" con tres folíolos. 3, dibujo esquemático basado en el morfotipo ejemplo. 5, detalle del patrón de venación: vena intramarginal (flecha negra), venas terciarias (flecha blanca) y venas intersecundarias (flecha blanca discontinua). 2, 4 y 6 (MHIN-UNSL-GEO-P461), 2, folíolo aislado presentando el ápice del lóbulo con terminaciones de tipo glandular (flecha blanca indicando la glándula). 4, dibujo esquemático basado en (MHIN-UNSL-GEO-P461). 6, detalle del ápice del lóbulo con terminación de tipo glandular.

Morphotype example. MHIN- UNSL-GEO-P483.
Studied material. MHIN-UNSL-GEO-P483; P484; P461- P464; P472- P477; P7; P41; P42; P46; P48; P56; P61; P62; P64; P80; P81; P89; P94; P111; P119; P209. LC refers to the La Cantera Formation; Microphyll trifoliate refers to morphological characters preserved.
Description. Pinnately compound microphyll trifoliate leaf consisting of two lateral and one terminal leaflets, petiolate. The petiole is 3.4 cm long. The leaflets are nanophylls, mostly symmetrical, but their shape and size are slightly variable; they vary between 0.3-1.5 cm long and 0.3-1 cm wide. The apex is acute and the base can be acute, obtuse as well as slightly cordate. The texture seems to be membranaceous. The petiolule of the terminal leaflet widens towards the base. The lateral leaflets are sessile or petiolulate. The margin of the leaflets is pinnately lobed; having as many as four pairs of lobes that are convex- convex in shape, the margin of the lobes is mostly entire. The lobes are more developed towards the base than the apex; they are separated by angular sinuses (figures 3, 4). Some specimens (figures 3.1, 5; 4.3) show a dentate instead of lobed margin (the teeth are marginal projections with sinuses indented less than ¼ of the distance to the midvein of the leaf). The presence of organic deposits on leaflet and lobe apex is here considered as evidence of glandular-like tips (figures 3.2, 6). Smaller secondary lobes may be present as well (figures 3.1, 2, 4, 5; 4.1, 3, 5, 7). The first vein category is pinnate. The primary and secondary veins have straight course, and moderate size. The secondary veins are craspedodromous, 3-5 pairs arising at acute angles (30-60º); they are opposite to subopposite and extend into the lobes ending at their glandular apex (figures 3.1-7) veins may be present running toward the sinus lobes (figure 3.5). Within the lobes, the secondary veins enter medially and there is an intramarginal vein in which these secondary veins end (figure 3.5). The tertiary veins are apparently alternate percurrent or not well differentiated from those of lower categories and they have sinuous course. Higher category veins are not preserved. Secondary lobes, if present, are innerved by tertiary veins or branches of secondary veins entering medially that ends at the tip of the lobe.
Remarks. This foliar type is the dominant in the assemblage. "LC-Microphyll trifoliate" presents a mosaic of characters that allows its placement within the dicots. It presents the pinnate venation, character considered to a primitive feature for the dicots but it is characterized by it compound leaves, craspedodromous secondary veins, three levels of venation, and toothed margin that are considered as advance characters for the mentioned group (Hickey and Wolfe, 1975). The leaflets are usually isolated. These are nanophylls (figures 3.2, 4; 4) which are smaller than other fossils angiosperm leaves of similar age and morphology in Argentina (e.g. the leaves from the Baqueró Group, are large, micro-mesophyll size class). The leaflets lobes are slightly variable in shape and deepness (figures 3, 4). Nevertheless, they share the general features that characterize the "LCMicrophyll trifoliate" (e.g. margin type and venation). The glandular apex of the leaflet, is irrigated by a vein that enters medially (this vein is mainly a secondary vein) and by two lateral veins that correspond apparently to the intramarginal vein. Glandular apexes of Chloranthoid and Rosoid types sensu Hickey and Wolf (1975) show this type of venation, nevertheless the poor preservation of the glandular apexes of the fossil taxon studied here does not allow to determine what type of tooth they belong to (figures 3.2, 6; 4.1, 4).

Figure 4. "LC-Microphyll trifoliate" (bar= 0.5 cm, except in 2 and 6 bar= 0.25 cm). 1 and 4 (MHIN- UNSL-GEO-P463) isolated leaflet, 4, Detail of the lobe apex with glandular-like tips. 2, (MHIN- UNSL-GEO-P472). 3, (MHIN- UNSL-GEO-P476). 5, (MHIN- UNSL-GEOP484). 6, (MHIN- UNSL-GEO-P474). 7, (MHIN- UNSL-GEO-P462) / "LC-Microfila trifoliada" (escala= 0,5 cm, excepto en 2 y 6 escala= 0,25 cm). 1 y 4 (MHIN- UNSL-GEO-P463), 4, detalle del ápice del lóbulo con terminaciones de tipo glandular. 2, (MHIN- UNSL-GEO-P472). 3, (MHINUNSL- GEO-P476). 5, (MHIN- UNSL-GEO-P484). 6, (MHIN- UNSL-GEO-P474). 7, (MHIN- UNSL-GEO-P462).
"LC-Microphyll trifoliate" can be considered as belonging to the "rank II" sensu Hickey (1974). It has secondary veins of relatively regular courses, with the size and shape of the areas between secondary veins uniform, but with tertiary and higher order venation randomly in course. It is important to note, that the presence of an intramarginal vein, is a new evolutive character and constitutes a more complex morphological level. The "rank II" leaf venation shows a more uniform pattern compared to the irregular "first-rank" present in those families considered as "basal angiosperms".
The studied morphotype has no cuticle and the venation is not completely preserved, consequently it was impossible to assign it to any previously described taxa. For this reason, the assignation to "LCMicrophyll trifoliate" of the specimens studied herein is considered more appropriate.
Discussion
Comparison with other fossil taxa
The angiosperms leaflets from the La Cantera Formation are pinnately lobed with craspedodromous venation. These characteristics are shared with other angiosperm fossil remains found in floras of similar age from different places of the world, such as the fossil leaves from the Baqueró Group, Anfiteatro de Ticó Formation (Early Late Aptian of Patagonia, Argentina; Romero and Archangelsky, 1986), "Type 5" leaf from the Crato Formation (Aptian of Brazil; Mohr and Friis, 2000), and Jixia Guo and Sun 2002 from the Chengzihe Formation (Hauterivian or Hauterivian-Early Barremian of Northern China; Sun and Dilcher, 2002).
The Baqueró Group leaves are large (micro-mesophyll size class), lobate, craspedodromous, dentate and have ramified tertiary veins and random fourthorder venation. These leaves can be distinguished from the La Cantera taxon based on margin, size and venation pattern.
Mohr and Friis (2000) described eight types of angiosperm leaves from the Crato Formation Among them, they described briefly only one specimen as "Type 5 leaf", which is characterized by its deeply 5- lobed lamina, entire margin and pinnate craspedodromous venation pattern. LC-Microphyll trifoliate and the Crato "Type 5 leaf" have pinnate and craspedodromous venation. However, there are some differences in size and margin shape between them, the Crato leaf is much larger (ca. 4.7 cm long and 6.5 cm wide approximately the triple in size) than the Argentinean specimens.
"LC-Microphyll trifoliate" shows some similarities with the fossil species of the genus Jixia (J. pinnatipartita (Guo and Sun) emend. Sun and Dilcher 2002 and J. chengzihensis Sun and Dilcher 2002), from the Chinese Chengzihe Formation, such as the pinnately lobed margin and pinnate venation, but they differ in their general shape. Jixia is characterized by deeper lobes and simple leaves meanwhile the Argentinean morphotype has compound leaves.
The leaves included in Crataegites Samylina 1960 and Cissites Debey (Capellini and Heer, 1866) are the most similar to the fossil described here. The genus Crataegites was erected by Samylina (1960, 1968) and includes pinnatilobed fossil leaves from the Early Cretaceous of Kolyma Basin, Russia.
Crataegites borealis Samylina 1968 consists of small (0.1-3.5 cm long), pinnately 3-7 lobed leaves with cuneate base and irregularly serrate teeth. Its venation is pinnate craspedodromous and no third vein category was reported, and along the margin a thin intramarginal vein can be seen (Samylina, 1968). This species resembles the fossils described herein in size, presence of pinnatilobed laminae with lobes apically oriented, deep and rounded sinuses, as well as by their venation. However, the irregularly serrate teeth present in C. borealis are not found in the La Cantera fossil.
Crataegites borealis f. rotundosinuosa Samylina 1968 has well developed lower lobes and possibly actinodromous or palinactinodromous venation that distinguish it from the La Cantera specimens. Due to the fact that the original diagnosis of Crataegites is relatively poor, further comparisons with the Argentinean fossil are doubtful.
Some species of Debey have also similarities with the "LC-Microphyll trifoliate". Originally, Cissites included fossil leaves presumably allied to the extant genus Cissus L., but subsequently was used by other authors to include palmate leaves having more or less resemblance to those of extant genera such as Vitis L., Platanus L., Sassafras Nees, among others. Cissites tavardensis Teixeira 1948 is comparable to "LCMicrophyll trifoliate" because of the presence of pinnatilobed laminae and craspedodromous venation. However, the lack of an adequate diagnosis for Cissites makes difficult the inclusion of our specimens within this genus.
One of the main features of "LC-Microphyll trifoliate" is the pinnately compound leaves, but none of the fossil species mentioned above show this feature, therefore, the inclusion of the La Cantera fossils is not possible in any of them. There are very few records of compound leaves from the Early and mid Cretaceous, among them the fossil genera Asiatifolium Sun, Guo and Zheng 2002 (Sun and Dilcher, 2002) from China, Sapindopsis Fontaine 1889, from the Subzone 2B of the Potomac Group, North America (Hickey and Doyle, 1977) and Kachaikenia compuesta Cúneo and Gandolfo 2005 (a recently described fossil species from the Albian-Cenomanian Kachaike Formation of Patagonia, Argentina). However, the leaflets of Asiatifolium are oblanceolate, opposite with margin entire. Sapindopsis differs from the "LCMicrophyll trifoliate" in their elliptic leaflets and a venation pattern with defined intercostal areas, intersecondary veins, and without an intramarginal vein. Kachaikenia compuesta have unlobed entire margin and the third vein category random reticulate meanwhile in "LC-Microphyll trifoliate" the margin is pinnately lobed and the tertiary veins are very irregular or not well differentiated from the lower orders.
Botanical affinity
"LC-Microphyll trifoliate" is placed without doubts within the dicotyledonous angiosperms based on leaf morphology and venation pattern. The characters that confirm the relationship with dicots and more specifically with the eudicots are the presence of at least three orders of veins forming a reticulate network, the toothed or lobate margin and the presence of glandular teeth with at least two orders of tooth (Hickey and Wolfe, 1975).
This new morphotype also presents chlorantoid or rosoid glandular teeth. The chlorantoid tooth-type is a distinctive feature of Chloranthaceae, but is also present in several groups of angiosperms including the Ranunculales. Furthermore, according to the leaf key of Hickey and Wolfe (1975) this morphotype could be placed within the range of the Ranunculidae. The APG II system (2003) recognizes the order Ranunculales and places it among the basal eudicots. On the other hand, the rosoid tooth-type could be suggests an affinity with the subclass Rosidae, supported also, by other features present on "LC-Microphyll trifoliate" (pinnately compound leaves, leaflets pinnately lobed with glandular margin and apex and craspedodromous venation) that characterize this Subclass (Hickey and Wolf, 1975). The Subclass Rosidae, corresponds to the "core eudicots" clade sensu the APG II system, which comprises the majority of "eudicots".
Following Hickey and Wolfe (1975) Ranunculidae present leaves that are basically pinnately compound with a ternate forking rachis with toothed margin and Chloranthoid teeth, pinnate venation, secondary veins craspedodromous, all distinctive characters of "LC-Microphyll trifoliate". In summary, "LC-Microphyll trifoliate" shares features with some modern groups of Rosid clade and Ranunculales clade. In general, fossil leaves of Early and Mid-Cretaceous age are difficult to place systematically with accuracy because they probably belong to extinct lineages (Friis et al., 2006). Consequently, based on the evidence presented here, it is suggested that this new morphotype, "LC-Microphyll trifoliate" from the Cretaceous of Argentina, could be representing an extinct primitive eudicot.
Tricolpate pollen grains are recorded from the Late Barremian-Early Aptian (Doyle, 1992; Hughes, 1994) and are the earliest evidence of the eudicot angiosperms. In southern South America, the first record of eudicot pollen grains occurred later by the Early Albian (Volkheimer and Salas, 1975). Besides the pollen record, the discovery of fossil angiosperm reproductive structures (flowers, fruits and seeds) that can be placed within the eudicot group reflects their evolutionary diversification during the Cretaceous (e.g., Friis and Skarby, 1981; Eklund, 2003; Friis et al., 1988, 1992, 1995, 1999; Nixon and Crepet, 1993; Gandolfo et al., 1998). Therefore the presence of a probable eudicot of Late Aptian age in Argentina is of considerable importance for those interested in angiosperm origin, since it is the oldest record of megafossil representing this group. Although angiosperm leaves are of much more restricted occurrence in the fossil record than angiosperm pollen and reproductive structures, they are also important evidences supporting this fact.
The La Cantera Formation has provided also an important palynofloristic assemblage, including many pollen grains of primitive angiosperm affinities such as Afropollis, Clavatipollenites, Pennipollis, Stellatopollis, Liliacidites, and Asteropollis (Prámparo 1988a, b, 1989a, b, 1990, 1994, 1999b). However, there are no records of tricolpate pollen grains in the palynofloristic association, making the presence of this new morphotype more important, because it is good evidence that the eudicots were present in Argentina probably before than what it is indicated by the palynological records.
Conclusions
A new Late Aptian angiosperm leaf morphotype from the La Cantera Formation, central western Argentina, is described and it is assigned as "LC-Microphyll trifoliate". The leaf remains occurred at the type section of the formation within a low diversity macrofloral association. Fragments of sphenophytes (equisetalean) and diverse types of reproductive structures (seeds and flowers) complete the association, together with other remains of leaves and undetermined stems, although their descriptions escape the scope of this report.
"LC-Microphyll trifoliate" shows features (venation and tooth type) present on extant Ranunculales, as well as Rosids clade. Therefore, this new morphotype can be considered as a possibly allied to the eudicots, being among the earliest leaf records within this group. Also, it could be among of the earliest records of angiosperm compound leaves, since there are very few records of compound leaves from the Early and Middle Cretaceous, and they all have leaflets with features clearly different to the La Cantera fossil.
To conclude, the association of fossil leaves and pollen grains found in the La Cantera Formation constitutes one of the most ancient and complete record of the first angiosperms in South America, and its study will give important clues to understand the enigmatic origin and evolution of this group of plants in the Southern Hemisphere.
Acknowledgments
I am grateful to M.G. Passalía for his help and advice regarding angiosperm fossil leaves identification. I thank also M.B. Prámparo and M.A. Gandolfo for their critical suggestions which improved the manuscript and to the Consejo Nacional de Investigaciones Científicas y Tecnicas (CONICET) of Argentina, for the financial support. I thank especially Rafael Bottero for his help in preparing the figures.
References
1. Angiosperm Phylogeny Group (APGII). 2003. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APGII. Botanical Journal of the Linnean Society 141: 399-436. [ Links ]
2. Archangelsky, S., Barreda, V., Passalía, M., Prámparo, M.B., Romero E.J., Zamuner, A., Cúneo, R., Gandolfo, M.A., Iglesias, A., Llorens, M. and Puebla, G.G. 2004. Early angiosperm diversification in the Cretaceous of Argentina: first approach. 7º International Organization of Paleobotany Conference. Bariloche, Abstracts: 4-6. [ Links ]
3. Archangelsky, S., Barreda, V., Passalía, M., Gandolfo, M.A., Prámparo, M.B., Romero E.J., Cúneo, R., Zamuner, A., Iglesias, A., Llorens, M., Puebla, G.G., Quattrocchio, M., Volkheimer, W. 2009. Early angiosperm diversification: evidence from southern South America. Cretaceous Research (in press). [ Links ]
4. Arcucci, A., Rivarola, D., Codorniú, L., Sallenave, A.S. and Puebla, G. 2002. Diversidad de la Asociación Biótica en el Paleoambiente Lacustre de la Formación La Cantera (Aptiano) de San Luis, Argentina. Boletim do 6º Simposio sobre o Cretaceo do Brasil. 2º Simposio sobre Cretácico de América del Sur. Sao Pablo, Brasil, pp. 119-122. [ Links ]
5. Capellini, G. and Heer, O. 1866. Les phyllites cretacees du Nebraska. Soc. Helvetique Sci. Nat. Nouv. Memory 22: 1-22. [ Links ]
6. Crabtree, D.R. 1987. Angiosperms of the Northern Rocky Mountains: Albian to Campanian (Cretaceous) megafossil floras. Annals of the Missouri Botanical Garden 74: 707-747. [ Links ]
7. Criado Roque, P., Mombru, C.A. and Moreno, J. 1981. Sedimentitas mesozoicas. Geología de la Provincia de San Luis. 7º Congreso Geológico Argentino (San Luis), pp. 79-96. [ Links ]
8. Cúneo, R. and Gandolfo, M.A. 2005. Angiosperm leaves from the Kachaike Formation, Lower Cretaceous of Patagonia, Argentina. Review of Palaeobotany and Palynology 136: 29-47. [ Links ]
9. Del Fueyo, G.M., Villar de Seoane, L., Archangelsky, A., Guler, V., Llorens, M., Archangelsky, S., Gamerro, J.C., Musacchio, E.A., Passalia, M.G., Barreda, V.D. 2007. Biodiversidad de las Paleofloras de Patagonia Austral durante el Cretácico Inferior. Asociación Paleontológica Argentina Publicación Especial 11: 101-122. [ Links ]
10. Dilcher, D.L. 1973. The Eocene floras of southeaestern North America. In: A. Graham (ed.), Vegetation and vegetational history of northern Latin America. Elsevier Scientific Publishing. Amsterdam, pp. 39-59 [ Links ]
11. Doyle, J.A. 1992. Revised palynological correlations of the lower Potomac Group (USA) and the Cocobeach sequence of Gabon (Barremian-Aptian). Cretaceous Research 13: 337-349. [ Links ]
12. Eklund, H. 2003. First Cretaceous flowers from Antarctica. Review of Palaeobotany and Palynology 127: 187-217. [ Links ]
13. Flores, M. 1969. El Bolsón de Las Salinas en la Provincia de San Luis. 4º Jornadas Geológicas Argentinas (Mendoza), Actas 1:: 311-327. [ Links ]
14. Flores, M. and Criado Roque, P. 1972. Cuenca de San Luis. 1º Simposio de Geología Regional Argentina. Academia Nacional de Ciencias (Córdoba), pp. 567-580. [ Links ]
15. Fontaine, W.M. 1889. The Potomac or younger Mesozoic Flora. Monograph. United States Geological Survey 15: 1-377. [ Links ]
16. Fossa Mancini, E. 1939. Vestigios de antiguos lagos en la Provincia de San Luis. Boletín de Información Petrolera 16: 17-42. [ Links ]
17. Friis, E.M. and Skarby, A. 1981. Structurally preserved angiosperm flowers from the Upper Cretaceous of southern Sweden. Nature 291: 485-486. [ Links ]
18. Friis, E.M., Crane, P.R., Pedersen, K.R. 1988. Reproductive structure of Cretaceous Platanaceae. Biologiske Skrifter, Det Kongelige Danske Videnskabernes Selskab 31: 1-55. [ Links ]
19. Friis, E.M., Pedersen, K.R. and Crane, P.R. 1992. Esgueiria gen. nov., fossil flowers with combretaceous features from the Late Cretaceous of Portugal. Biologiske Skrifter, Det Kongelige Danske Videnskabernes Selskab 41: 1-45. [ Links ]
20. Friis, E.M., Pedersen, K.R. and Crane, P.R. 1995. Appomattoxia ancistrophora gen. et sp. nov., a new Early Cretaceous plant with similarities to Circaeaster and extant Magnoliidae. American Journal of Botany 82: 933-943. [ Links ]
21. Friis, E.M., Pedersen, K.R. and Crane, P.R. 1999. Early angiosperm diversification: the diversity of pollen associated with angiosperm reproductive structures in Early Cretaceous floras from Portugal. Annals of the Missouri Botanical Garden 86: 259-296. [ Links ]
22. Friis, E.M., Pedersen, K.R. and Crane, P.R. 2006. Cretaceous angiosperm flowers: Innovation and evolution in plant reproduction. Palaeogeography, Palaeoclimatology, Palaeoecology 232: 251-293. [ Links ]
23. Gandolfo, M.A., Nixon, K.C. and Crepet, W.L. 1998. A new fossil flower from the Turonian of New Jersey: Dressiantha bicarpelata gen. et sp. nov. (Capparales). American Journal of Botany 85: 964-974. [ Links ]
24. Hickey, L.J. 1973. Classification of the arquitecture of dicotyledonous leaves. American Journal of Botany 60: 17-33. [ Links ]
25. Hickey, L.J. 1974. A revised classification of the architecture of dicotyledonous leaves. In: Metcalfe, C.R. y Chalk, L. (eds.), Anatomy of the dicotyledons, Vol. 1, Second Edition, Clarendon Press, Oxford, pp. 25-39. [ Links ]
26. Hickey, L.J. 1977. Stratigraphy and paleobotany of the Golden Valley Formation (Early Tertiary) of western North Dakota. The Geological Society of America, Memoirs 150: 157-164. [ Links ]
27. Hickey, L.J. and Doyle, J.A. 1977. Early Cretaceous fossil evidence for angiosperm evolution. The Botanical Review 43: 2-104. [ Links ]
28. Hickey, L.J. and Wolfe, J.A. 1975. The bases of angiosperm phylogeny: vegetative morphology. Annals of the Missouri Botanical Garden 62: 538-589. [ Links ]
29. Hughes, N.F. 1994. The enigma of angiosperms origins. Cambridge University Press, Cambridge, pp. 1-303. [ Links ]
30. Hünicken, M. and Romero, E. 1981. Improntas de angiospermas de la Formación La Cantera (Cretácico Inferior) de San Luis, República Argentina. 4º Simposio Argentino de Paleobotánica y Palinología (Córdoba, 1980). Resumen en Paleobotánica Latinoamericana. Circular Informativa da ALPP, Sao Paulo, 3: 1-18. [ Links ]
31. Leaf Architecture Working Group. 1999. Manual of leaf architecture, morphological description and categorization of dicotyledoneous and net-veined monocotyledonous angiosperms. Smithsonian Institution, Washington, 65 pp. [ Links ]
32. Maisey, J.G. 1991. Santana fossils: an illustrated atlas. TFH, Neptune City, N.J., 459 pp. [ Links ]
33. Martill, D.M., Brito, P.M., Wenz, S. and Wilby, P.R. 1993. Fossils of the Santana and Crato Formations, Brazil, pp. 1-129. In: E.A. Jarzembowski (ed.), Palaeontological Association, London, 158 pp. [ Links ]
34. Mazzoni, A. 1985. Notonectidae (Hemiptera, Heteroptera) de la Formación La Cantera (Cretácico Inferior), Provincia de San Luis, Argentina. Boletín de la Academia Nacional de Ciencias (Córdoba) 56: 259-273. [ Links ]
35. Mazzoni, A. and Hünicken, M. 1984. Ontogenia de los Notonectidos (Insecta, Heteroptera) del Cretácico Inferior de San Luis, Argentina. 3º Congreso Latinoamericano de Paleontología (México), Memoria pp. 388-393. [ Links ]
36. Mazzoni, A. and Hünicken, M. 1987. Corixidae (Insecta, Heteroptera) en el Cretácico Inferior de la Sierra del Gigante, San Luis, Argentina. 4º Congreso Latinoamericano de Paleontología. Bolivia, 2: 731-738. [ Links ]
37. Mohr, B.A.R. and Bernardes-de-Oliveira, M.E.C. 2004. Endressinia brasiliana, a magnolialean angiosperm from the Lower Cretaceous Crato Formation (Brazil). International Journal of Plant Sciences 165: 1121-1133. [ Links ]
38. Mohr, B.A.R., Bernardes-de-Oliveira, M.E.C., Barale, G. and Ouaja, M. 2006. Palaeogeographic distribution and ecology of Klitzschophyllites, an early Cretaceous angiosperm in southern Laurasia and northern Gondwana. Cretaceous Research 27: 464-472. [ Links ]
39. Mohr, B.A.R and Eklund, H. 2003. Araripia florifera, a magnoliid angiosperm from the Lower Cretaceous Crato Formation (Brazil). Review of Palaeobotany and Palynology 126: 279-292. [ Links ]
40. Mohr, B.A.R. and Friis, E.M. 2000. Early angiosperms from the Lower Cretaceous Crato Formation (Brazil), a preliminary report. International Journal of Plant Sciences 161(6 Supplement): 155-167. [ Links ]
41. Mohr, B.A.R. and Rydin, C. 2002. Trifurcatia flabellata n. gen. n. sp., a putative monocotyledon angiosperm from the lower Cretaceous Crato Formation (Brazil). Mitteilungen aus dem Museum für Naturkunde der Humboldt-Universität Berlin, Geowissenschaftliche Reihe 5: 335-334. [ Links ]
42. Nixon, K.C. and Crepet, W.L. 1993. Late Cretaceous fossil flowers of ericalean affinity. American Journal of Botany 80: 616-623. [ Links ]
43. Prámparo, M.B. 1988 a. Nuevos aportes a la palinología de la Formación La Cantera, Cretácico de la Cuenca de San Luis, en su localidad tipo. 4º Congreso Argentino de Paleontología y Bioestratigrafía. Mendoza. Actas III, pp. 41-50. [ Links ]
44. Prámparo, M.B. 1988 b. Esporas triletes levigadas y apiculadas de la Formación La Cantera (Cretácico de la Cuenca de San Luis) en su localidad tipo. 4º Congreso Argentino de Paleontología y Bioestratigrafía. Mendoza. Actas III, pp. 51-62. [ Links ]
45. Prámparo, M.B. 1989 a. [Palinología estratigráfica del Cretácico de la Cuenca de San Luis. Tesis Doctoral. Universidad Nacional de Rio Cuarto, 242 pp. Unpublished. [ Links ]].
46. Prámparo, M.B. 1989b. Las esporas de Schizaeaceae (Cicatricosisporites y Appendicisporites) del Cretácico Inferior, Cuenca de San Luis, Argentina. Revista Española de Micropaleontología 21: 355-372. [ Links ]
47. Prámparo, M.B. 1990. Palinoestratigraphy of the Lower Cretaceous of the San Luis Basin, Argentina. Its place in the Lower Cretaceous floral provinces pattern. Neues Jahrbuch für Geologie und Paläontologie Abhadlungen 181: 255-266. [ Links ]
48. Prámparo, M.B. 1994. Lower Cretaceous palynoflora of the La Cantera Formation, San Luis Basin. Correlation with other Cretaceous palynofloras of Argentina. Cretaceous Research 15: 193-203. [ Links ]
49. Prámparo, M.B. 1999a. Microfitoplancton orgánico del Cretácico Inferior de la Cuenca de San Luis. Parte I: Scenedesmaceae y Chlorococcaceae. Asociación Paleontológica Argentina. Publicación Especial 6. 10º Simposio Argentino de Paleobotánica y Palinología (Buenos Aires), pp. 39-42. [ Links ]
50. Prámparo, M.B. 1999 b. Granos de polen de primitivas angiospermas en el Cretácico Inferior de la Cuenca de San Luis y su distribución en otras cuencas cretácicas de Argentina. Boletim 5º Simposio sobre o Cretaceo do Brasil, UNESP, pp. 539-543. [ Links ]
51. Prámparo, M.B., Quattrocchio, M., Gandolfo, M.A., Zamaloa, M.C. and Romero, E. 2007. Historia evolutiva de las angiospermas (Cretácico-Paleógeno)en Argentina a través de los registros paleoflorísticos. Asociación Paleontológica Argentina Publicación Especial 11: 157-172. [ Links ]
52. Puebla, G.G. 2003. Nueva angiosperma del Cretácico Inferior de la Provincia de San Luis, Argentina. 29º Jornadas Argentinas de Botánica y 15º Reunión Anual de la Sociedad Botánica de Chile. Boletín de la Sociedad Argentina de Botánica 38: pp. 299. [ Links ]
53. Puebla, G.G. 2004. [La Megaflora de la Formación La Cantera (Cretácico Temprano) Sierra del Gigante, San Luis. Argentina. Tesis de Licenciatura, Facultad de Química, Bioquímica y Farmacia, Universidad Nacional de San Luis , 49 pp. Unpublished. [ Links ]].
54. Rivarola, D. and Di Paola, E. 1992. Secuencias mesozoicas de la Sierra de Las Quijadas. Paleoambientes y paleoclimas. Provincia de San Luis. República Argentina. 1º Encontro Sobre Sedimentacao Continental Bacias Mesozoicas Brasileiras. Acta Geológica Leopoldensia 15: 143-145. [ Links ]
55. Rivarola, D. and Spalletti, L. 2006. Modelo de sedimentación continental para el rift Cretácico de la Argentina central. Ejemplo de la Sierra de las Quijadas, San Luis. Revista de la Asociación Geológica Argentina 61: 63-80. [ Links ]
56. Romero, E.J. and Archangelsky, S. 1986. Early Cretaceous angiosperms from the Southern South America. Science 234: 1580- 1582. [ Links ]
57. Samylina, V.A. 1960. Angiosperm plants from the Lower Cretaceous deposits of the Kolyma River. Botanicheskii Zhurnal 45: 335-352 [in Russian]. [ Links ]
58. Samylina, V.A. 1968. Early Cretaceous angiosperms of the Soviet Union based on leaf and fruit remains. Botanical Journal of the Linnean Society 61: 207-216. [ Links ]
59. Sun, G. and Dilcher, D.L. 2002. Early angiosperms from the Lower Cretaceous of Jixi, eastern Heilongjiang, China. Review of Palaeobotany and Palynology 121: 91-112. [ Links ]
60. Taylor, D.W.and Hickey, L.J. 1990. An Aptian Plant with Attached Leaves and Flowers: Implications for Angiosperm Origin. Science 247: 702-704. [ Links ]
61. Teixeira, C. 1948. Flora Mesozoica Portuguesa, Parte I. Serviços Geologicos de Portugal, Lisboa, 118 pp. [ Links ]
62. Volkheimer, W. and Salas, A. 1975. Die älteste Angiospermen- Palynoflora Argentiniens von der Typuslokalität der unterkretazischen Huitrín-Folge des Neuquén-Beckens. Ihre Mikrofloristische Assoziation und biostratigraphische Bedeutung. Neues Jahrbuch für Geologie Paläontologie, Monatshefte 1975: 424-436. [ Links ]
Recibido: 6 de enero de 2009.
Aceptado: 23 de junio de 2009.

