










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana vol.46 no.4 Buenos Aires oct./dic. 2009
ARTÍCULOS ORIGINALES
Late Cretaceous Megaloolithid eggs from Salitral de Santa Rosa (Río Negro, Patagonia, Argentina): inferences on the titanosaurian reproductive biology
Leonardo Salgado1, Claudia Magalhaes Ribeiro2, Rodolfo A. García1, and Mariela S. Fernández1
1Conicet-Inibioma, Museo de Geología y Paleontología, Universidad Nacional del Comahue, Buenos Aires 1400, 8300 Neuquén, Argentina lsalgado@uncoma.edu.ar rgarcia@uncoma.edu.ar mariu_soledad_fernandez@hotmail.com
2Departamento de Geociências, Instituto de Agronomia, Universidade Federal Rural do Rio de Janeiro, BR-465, km 7, 23890-000 Seropédica/RJ, Brazil claudiammr@yahoo.com.br
Abstract. Late Cretaceous eggs and eggshell fragments assigned to the oofamily Megaloolithidae from Salitral de Santa Rosa (Río Negro Province, northern Patagonia, Argentina) are studied and compared with other Patagonian specimens assigned to the same oofamily. Megaloolithids from Santa Rosa are nearly spherical (mean diameter 10-12 cm) and their eggshell surface is ornamented with hemispheric nodes 0.3-0.8 mm in diameter; pore openings are subcircular to polygonal, and their diameter varies between 0.1 and 0.32 mm. Eggshell thickness ranges from 1 to 2.6 mm. The eggshell is composed of fanshaped units that normally display diverging borders and arched accretionary lines. The megaloolithid eggs from Santa Rosa are associated, at least in some sites of the area, to eggs of different kinds, more usually, thick-shelled faveoloolithids.
Resumen. Huevos Megaloolitidos del Cretácico tardío del Salitral de Santa Rosa (Río Negro, Patagonia, Argentina): inferencias sobre la biología reproductiva de los Titanosaurios. Cáscaras y huevos del Cretácico Tardío asignados a la oofamilia Megaloolithidae, provenientes del Salitral de Santa Rosa (Provincia de Río Negro, norte de Patagonia, Argentina) son estudiados y comparados con otros especímenes asignados a la misma oofamilia. Los huevos megaloolítidos de Santa Rosa son subesféricos (diámetro promedio entre 10-12 cm.) y la superficie de su cáscara está ornamentada con nódulos hemiesféricos de 0.3-0.8 mm de diámetro, las aperturas de poro son subesféricas a poligonales, y sus diámetros varían entre 0.1 y 0.32 mm. El rango de grosor de las cáscaras varía entre 1 y 2.6 mm. Las cáscaras están formadas por unidades en forma de abanico, las cuales normalmente se disponen con bordes divergentes y con líneas de crecimiento arqueadas. Los huevos megaloolítidos de Santa Rosa están asociados, por lo menos en algunos sitios del área, a huevos de diferentes tipos, usualmente a faveoloolítidos de cáscara gruesa.
Key words. Fossil eggs; Megaloolithidae; Late Cretaceous; Río Negro province; Patagonia; Argentina.
Palabras clave. Huevos fósiles; Megaloolithidae; Cretácico Tardío; Río Negro; Patagonia; Argentina.
Introduction
The discovery of dinosaur skeletons in ovo at the locality of Auca Mahuevo (northern Neuquén Province, Patagonia, Argentina), made possible to link, by first time, a variety of fossil egg identified as Megaloolithidae of the traditional parataxonomic classification (Zhao, 1979) to titanosaur dinosaurs (Chiappe et al., 1998, 2001).
Megaloolithid eggs are characterized by their spheroidal form (although some eggs from France attributed to this oofamily seem to have a rather oval shape [Vianey-Liaud et al., 1994]), and their eggshell formed by fan-shaped shell units (that show, in a thin section, arched lines of accretion), whose tops constitute superficial nodes (Carpenter, 1999).
The material ascribed to this oofamily shows minor differences in a number of features (size, eggshell thickness, pore diameter, shell unit proportions, etc.); precisely, this variation have allowed to a number of authors to distinguish numerous ooespecies (e.g., Mohabey, 1996; Vianey-Liaud et al., 2003), although not all researchers agree entirely with this practice (e.g., Peitz, 2000; Grellet-Tinner et al., 2006).
Neuquenian megaloolithids are also represented by fragmentary pieces from the suburbs of Neuquén city, described by Calvo et al. (1997), and collected from the Río Colorado Subgroup (possibly from the Bajo de la Carpa Formation, lower Campanian). In Salitral de Santa Rosa (Allen Formation, middle Campanian-lower Maastrichtian), in the center of Río Negro Province, a field trip conducted in 2003 by the Universidad Nacional del Comahue, the Museo "Carmen Funes" (Plaza Huincul, Neuquén), and the Museo de Lamarque (Río Negro), resulted in the discovery of hundreds of dinosaur eggs and eggshells, including megaloolithids. These ones, briefly described elsewhere (Salgado et al., 2007b), are the main focus of this paper.
Although it is absolutely reasonable that different groups of dinosaurs have laid eggs of a same type (that is, of a same ootaxon), this possibility does not seem to have been seriously taken into account, at least in the case of megaloolithid eggs. Furthermore, the Auca Mahuevo discovery seems to have persuaded most researchers that all megaloolithids belong to sauropods (García et al., 2006), specifically to titanosaurs. An exemption to this is the suspicion raised by Vianey-Liaud et al. (2003, p. 583), that different species of Megaloolithus may have been laid by different dinosaur taxa, not only sauropods, and the studies of Grigorescu et al. (1994) and Grigorescu (2006), which report an association of hatchling hadrosaurs with megaloolithids.
In this work we describe in depth the megaloolithid eggs and eggshells from Salitral de Santa Rosa, and compare them with other Patagonian eggs ascribed to titanosaurian sauropods, specially with the megaloolithids from Auca Mahuevo, Neuquén city (Neuquén Province), and Salitral Moreno (Río Negro Province). Specially in the first locality, an exhaustive work has been carried out, both on the own eggs/eggshells (Grellet-Tinner et al., 2004; Jackson et al., 2004), as well as on the embryos (Chiappe et al., 2001; Salgado et al., 2005; García, 2007). Finally, we hypothesize on some subjects concerning to the reproductive biology of the sauropods, in the light of the evidence recovered in the area.
Material and methodology
The eggshells studied (see Material examined in Systematic Paleontology) were collected with precise stratigraphic control, in the area of Salitral de Santa Rosa-Salinas de Trapalcó, Río Negro, Argentina (figure 1, tables 1 and 2).
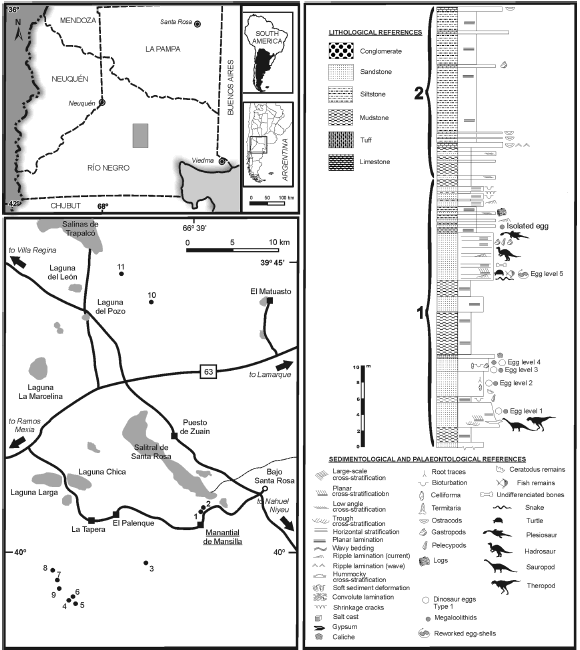
Figure 1. Location map of the sites mentioned in the text that yielded dinosaur eggs, and stratigraphical section of the Allen Formation in Salitral de Santa Rosa-Salinas de Trapalcó (Río Negro Province), showing the five egg-bearing levels / mapa de ubicación de los sitios mencionados en el texto que han aportado huevos de dinosaurios y sección estratigráfica de la Formación Allen en Salitral de Santa Rosa-Salinas de Trapalcó (provincia de Río Negro), mostrando los cinco niveles portadores de huevos. Sites / sitios: 1, Mansilla I; 2, Mansilla II; 3, García I; 4, Cerro Bonaparte; 5, Berthe II; 6, Berthe III; 7, Berthe IV; 8, Berthe V ("Loma Puntuda"); 9, Berthe VI; 10, Cerro Tortugas; 11, Cerro Laguna Trapalcó.
Table 1. GPS coordinates for the 19 sites with dinosaur eggs in Salinas de Trapalcó-Salitral de Santa Rosa / coordenadas de GPS para los 19 sitios con huevos de dinosaurio en Salinas de Trapalcó- Salitral de Santa Rosa.

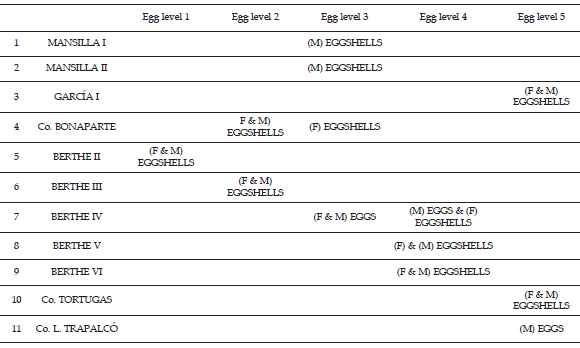
Table 2. Egg levels and eggshell types distribution in the sites included in the study. F, faveoloolithids; / megaloolithids / distribución de los niveles y tipos de cáscara de huevo en los sitios incluidos en este estudio. F, faveoloolítidos; M, megaloolítidos.
In order to remove the particles adhered to the eggshells, we employed an ultrasound equipment. The eggshell thicknesses were measured using a digital caliber Mitutoyo model CD-6" C, and in some cases, directly from the thin sections. The specimens were first analysed with a binocular lupa Stemi SV6 Zeiss, and the thin sections observed with a polarized microscope Zeiss Axioplan Image 2. They were photographied with a camera Sony-shot DSC S-85. Smaller fragments were gold metalized and observed with a Scanning Electronic Microscope Phillips 515 in the Facultad de Ingeniería de la Universidad Nacional del Comahue, and with a Phillips SEM XL3 in the Museo Argentino de Ciencias Naturales of Buenos Aires "Bernardino Rivadavia".
Geology
The unit that yielded the fossil eggs is the Allen Formation, whose age has been estimated by Ballent (1980), based on ostracods, as middle Campanianlower Maastrichtian. This formation, but in the area of Lago Pellegrini, 260 km to the northwest of Salitral de Santa Rosa, has been divided by Andreis et al. (1974) in three members. With respect to its paleoenvironment, Leanza and Hugo (2001) suggested that the formation is characterized by fluvial deposits of moderate energy (the lower member) followed by lacustrine and fluvial deposits of low energy (the middle member), and, finally, shallow lacustrine deposits with evaporitic facies (the upper member).
In the area of Salitral de Santa Rosa-Salinas de Trapalcó, the Allen Formation is widely exposed (Hugo and Leanza, 2001), more than in other places of the province. Here, Salgado et al. (2007b) differentiated two subunits of the Allen Formation: (1) a lower one, comprised of fine-grained sandy deposits with subordinate muddy layers and thin evaporitic levels, and (2), an upper thick succession of siltstone and mudstone deposits with thin intercalations of ostracod- rich limestones and sandy levels (figure 1). On the basis of sedimentological and lithofacial features, the lower subunit (which bears vertebrate fossils, including dinosaur eggs) is interpreted as a brackish lagoonal and supratidal environment, associated with aeolian sands (dunes) and deposits of ephemeral rivers. Abundant rizoturbation and caliche in the egg-bearing deposits indicate the presence of palaeosols, which presumably developed in a coastal, more probably supratidal environment. The facies and environmental conditions of these deposits suggest that the lower subunit is related to the middle member of the Allen Formation sensu Andreis et al. (1974).
In the area of Salitral de Santa Rosa-Salinas de Trapalcó, five egg-bearing levels have been recognized in the middle and upper parts of the lower subunit (figure 1). In addition to the megaloolithid eggs, the focus of this paper, these layers brought other varieties of dinosaur eggs, for instance, material assignable to the oofamily Faveoloolithidae (Zhao and Ding, 1976), which is comparable to Sphaerovum erbeni Mones, 1980 (Simón, 1999, 2006) (Salgado et al.'s"type 1", 2007b).
The lowermost egg-bearing layer, the egg level 1, clearly exposed in the site Berthe II (figure 1, table 2), is continuous towards Cerro Bonaparte, where the holotype of the titanosaur Bonatitan reigi Martinelli and Forasiepi, 2004 and the partial skeleton of the dromaeosaurid theropod Austroraptor cabazai Novas et al., 2008 were found. Egg level 2 is well exposed in Cerro Bonaparte and Berthe III; egg level 3, the best exposed egg-bearing layer, is found in Mansilla I, Mansilla II, Cerro Bonaparte, and Berthe IV; egg level 4 is exposed in Berthe IV and V and Berthe VI; finally, egg level 5 exposures are in García I, Cerro Laguna Trapalcó, and Cerro Tortugas (figure 1; table 2). The few, reworked eggshells recorded in the egg level 5 were invariably associated with gastropods of the genus Paleoanculosa Parodiz, 1969, bivalves of the genus Diplodon Spix and Wagner, 1827 and numerous, fragmentary bones of fishes, turtles, snakes, plesiosaurs, dinosaurs, and mammals (Rougier et al., 2009).
Close to the site Berthe VI, on the flanks of a hill locally named as "Loma Puntuda", a partial skeleton of a polycotylid plesiosaur was found, 3.5 m above the egg level 5 (Salgado et al., 2007a). The discovery of this reptile, together with the presence of ichnofascies of Psilonichnus Frey and Pemberton, 1987 and heterolithic and evaporithic structures, indicates marginal marine conditions of low to moderate energy, as those recorded in intertidal and supratidal plains. These conditions would have prevailed in the first five transgressive-regressive cycles recorded in the Neuquén Basin, during the lapse late Campanian- Danian, corresponding to the depositional "sequence I", identified by Parras et al. (1998) and Parras and Casadío (1999) in other places of the basin.
Technical Abbreviations. EDAX, Energy Dispersive X-ray analysis; SEM, scanning electron microscope; MT, Membrana Testacea.
Institutional Abbreviations. MML, Museo Municipal de Lamarque, Río Negro, Argentina; UFRRJ, Universidade Federal Rural de Rio de Janeiro, Brazil; UNC-CIP, Centro de Investigaciones Paleontológicas, Universidad Nacional del Comahue.
Systematic paleontology
Morphotype DISCRETISPHERULITIC Mikhailov, Bray and Hirsch, 1996 Oofamily MEGALOOLITHIDAE Zhao, 1975 (emended, 1979)
Oogenus Megaloolithus Vianey-Liaud, Mallan, Buscail and Montgellard, 1994
Megaloolithus sp. Figures 2, 4,10
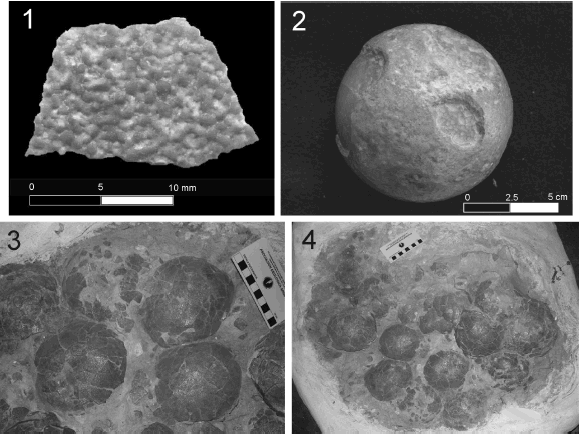
Figure 2. 1, megaloolithid eggshell surface showing the compactituberculate ornamentation / superficie de la cáscara de megaloolítidomostrando la ornamentación compactituberculada; 2, megaloolithid egg 434/P/96 / huevo megaloolítido 434/P/96; 3-4, egg clutch MML-Pv 41 recovered from Berthe IV, egg level 3 / agrupación de huevos MML-Pv 41 colectada de Berthe IV, nivel de huevo 3.
Material examined. 933 eggshell fragments and a clutch of 14 eggs, which come from different sites (and different levels) within the area of Salitral de Santa Rosa: MML-Pv-8, two eggshell fragments from Cerro Bonaparte (egg level 2); MML-Pv-11, one eggshell fragment from Berthe II (egg level 1); MML-Pv-21, eight eggshell fragments, and MML-Pv-41, clutch containing 14 eggs from Berthe IV (egg level 3); MML-Pv-17, three eggshell fragments from Berthe VI (egg level 4); MML-Pv-19, one eggshell fragment from Berthe III (egg level 2); MML-Pv-25, 622 eggshell fragments from Mansilla I (egg level 3); MML-Pv-37, 260 eggshell fragments from Mansilla II (egg level 3); MML-Pv-26, three eggshell fragments from Mansilla I (egg level 3); MML-Pv-30, 14 eggshell fragments from Berthe V (egg level 4); MML-Pv-35, six eggshell fragments from García I (egg level 5); UNC-CIP 4-7, and UNC-CIP 11 from Mansilla I (egg level 3); UNC-CIP 2, UNC-CIP 3, UNC-CIP 8- 10 from Mansilla II (egg-level 3). Three thin sections: UFRRJ-1, from Mansilla I, and UFRRJ-2 and 3, from Mansilla II.
Occurrence. Salitral de Santa Rosa and Salinas de Trapalcó, Río Negro, Argentina. The samples were collected from 11 sites (figure 1, table 1); however, more than 95 % of the collected eggshells procedes from the sites Mansilla I and II. Mid Member of the Allen Formation. Middle Campanian-lower Maastrichtian (Ballent, 1980) (figure 1).
Remarks. The following analysis of the megaloolithids from Santa Rosa will be centered on eggshells collected in the sites Mansilla I and II, which are close to a puesto (ranch) named as "Manantial de Mansilla", 8 km to the west of the little town of Bajo Santa Rosa, on the southeastern margin of Salitral de Santa Rosa (figure 1). The eggshells found in these sites do not show signs of long transport, and basically present the same degree of diagenetic alteration. For this reason, the 892 fragments collected in these two sites are presumed to come all from the same nesting area, which was possibly not so far from there.
Comparative description. Megaloolithid eggshells from Salitral de Santa Rosa are relativelly well, but not exceptionally preserved (exceptional preservation implies preservation of the original mineralogy, according to Schweitzer et al., 2004). In this regard, the elemental analysis (EDAX), performed on both surfaces of the eggshell UNC-CIP 2 (collected in Mansilla II), gave the following results:
Outer surface (taken at the center of a node)= 8.05% of Al; 11.03% of Si; 80.92% of Ca. Inner surface (taken at the borders of a mammillary structure) = 36.48% of Al; 6.55% of P; 56.10% of Ca.
Although no complete eggs were found in Mansilla I and II, entire fragments including partial clutches were discovered in some neighbor sites (Berthe IV and Cerro Laguna Trapalcó, figure 1) (table 2). The eggs are almost perfectly spherical. The specimen 434/P/96, housed in the Museo Regional de Valcheta (Río Negro Province) (figure 2.2), and collected in the area of Salitral de Santa Rosa, although from an imprecise location (M. Kopp personal communication, 2006), has a diameter of 9.42 cm; this is the smallest megaloolithid recorded in the region. The clutch MML-Pv 41, unearthed in Berthe IV, contains 14 eggs whose diameters vary between 10 and 12 cm (figures 2.3-4). As we see, the megaloolithids from Río Negro are rather small with relation to the eggs from Auca Mahuevo (13 to 15 cm, after Chiappe et al. (1998); 12.5 to 14 cm, after Grellet- Tinner et al. (2004), although some eggs from the same locality have diameters lesser than 12.5 cm (Jackson, personal communication, 2006), and Neuquén city (16 cm, according to Calvo et al., 1997), although in this last case, the important degree of deformation of the eggs inhibits a true estimation of their diameters. In addition, all megaloolithid species recorded in India and France are larger in overall terms than those from Río Negro: M. cylindricus Khosla and Sahni, 1995, 12-20 cm; M. mohabeyi Khosla and Sahni, 1995, 16-19 cm; M. khempurensis Mohabey, 1998, 17-20 cm; M. matleyi Mohabey, 1996, 16-18 cm; M. mammillare Vianey-Liaud, Mallan, Buscail and Montgelard, 1994, 19-23 cm (Vianey- Liaud et al., 2003).
The eggs from Salitral de Santa Rosa differ from those from Auca Mahuevo in that they do not have two different (lesser and greater) diameters (Grellet- Tinner et al., 2004); but this could be only indicating that the eggs from Río Negro have a lesser degree of compaction.
The eggshell of the megaloolithid eggs from Santa Rosa is relatively thin, with mean thicknesses of 1.23 mm in Berthe IV (egg level 4), and 1.8 mm and 2,01 mm in Mansilla I and II respectively (the mode is 1.54 in Mansilla I, and 2.16 in Mansilla II); considering both sites together, the mean thickness is 1.86 (while the mode lies in 2.25 mm) (figure 3). These last dimensions match well with the megaloolithid eggshells recorded in Neuquén city (range of eggshell thickness variation is 1.7-2.1 mm, although Calvo et al. (1997) did not provide other statistical data relative to eggshell thickness, such as mean or mode), whereas the thinest eggshells from Santa Rosa match best with the thickest eggshells from Auca Mahuevo (1.31 mm, Grellet -Tinner et al. (2004); although Chiappe et al. (1998) report eggshell thicknesses of up to 1.78 mm). The mean thickness recorded in Auca Mahuevo is 1.4 mm (Chiappe et al., 1998) and in Patagoolithus Simón, 2006 (taken from a total of six eggshells), 1.56 (Simón, 2006).

Figure 3. Histograms of eggshell fragment thickness assigned to Megaloolithus from Manantial de Mansilla / histogramas de espesores de fragmentos de cáscaras de huevo asignados a Megaloolithus de Manantial de Mansilla.
The thickness of the megaloolithid eggshells collected in Mansilla I and II has an unusually high range: 1.72 in Mansilla I, 1.59 in Mansilla II; 1.72, if both sites are considered together (figure 3). In the megaloolithids from Auca Mahuevo (at least in those eggs preserving embryos), the range observed is 0.66 (Grellet- Tinner et al., 2004); in Patagoolithus, 0.56 (Simón, 2006).
The external ornamentation of the megaloolithid eggshells from Salitral de Santa Rosa consists basically in hemispheric nodes (figures 2.1, 4.1-4); some eggshells, however, present relatively low nodes, which is probably due to diagenesis (Grellet-Tinner et al., 2004). These nodes are not but the tops of the shell units (figures 8.1, 8.3-4). In the literature, this type of ornamentation is known as "compactituberculate", according to the terminology adopted by Mikhailov (1991), and it is actually considered as a sauropod/titanosaurian synapomorphy (Grellet- Tinner, 2006a).
The nodes of the megaloolithid eggs from Santa Rosa have diameters ranging from 0.3 to 0.8 mm, which virtually coincide with the megaloolithids from Auca Mahuevo (0.35-0.65 mm, Chiappe et al., 1998). In UNC-CIP 11 (Mansilla I), the eggshell thickness is 1.4 mm, and the superficial nodes have a diameter that vary between 0.5 and 0.8 mm (figure 4.2). In this specimen, the nodes are apparently somewhat separated, perhaps as result of diagenesis. For instance, between two nodes of, respectively, 0.8 and 0.59 mm, there is a distance of 0.35 mm. The internodular spaces reported by Grellet-Tinner et al. (2004, table 1) in some eggshells from Auca Mahuevo, are somewhat greater (0.52 to 0.87 mm). Also, in UNCCIP 11, some coalescent nodes are observed. The coalescence is normally between two nodes, more rarely between three or more; in this regard, there is a difference with the megaloolithids from Neuquén city, where up to five coalescent nodes have been reported (Calvo et al., 1997). In other samples (UNCCIP 6 [Mansilla I], UNC-CIP 10 [Mansilla II]), whose eggshells are relatively thick (2 and 2.2 mm, respectively), sub-polygonal nodes are compacted (figure 4.1). Basically, the nodes observed on the outer surface of the megaloolithids from Santa Rosa are nearly identical to those of the faveoloolithids, although in these last, the nodes are smaller, high in respect to the nodular width, and rarely coalescent (Salgado et al., 2007b).

Figure 4. External ornamentation of the megaloolithid eggshell / Ornamentación externa de cáscara de megaloolítido. 1, UNC-CIP 6 (eggshell thickness 2.00 mm) / UNC-CIP 6 (espesor de cáscara 2.00 mm); 2, UNC-CIP 11 (eggshell thickness 1.4 mm) / UNC-CIP 11 (espesor de cáscara 1.4 mm); 3-4, UNC-CIP 2 / UNC-CIP 2. In 3-4, sub-triangular pore openings are observed / En 3-4 se observan aberturas de poro subtriangulares.
The megaloolithids from Santa Rosa display external pore apertures varying between 0.1 and 0.32 mm in diameter, although this feature may be merely reflecting the effects of diagenesis. Nevertheless, the external pore opening diameter is basically similar to the megaloolithids from Auca Mahuevo (0.15 to 0.29 mm, Grellet-Tinner et al., 2004), and greater than the specimens from Neuquén city (0.07-0.1 mm, Calvo et al., 1997). Patagoolithus salitralensis Simón, 2006 in turn, has a range of pore opening diameter inusually high (0.045-0.375 mm) (Simón, 2006). In UNC-CIP 11, pore apertures are predominantly subrounded (figures 4.2-4.4), as in UNC-CIP 10 (figure 5.3). In UNC-CIP 11 (figure 4.2), these apertures are frequently coalescent, surely due to diagenesis. In some cases, external pore openings have stepped borders; resembling those observed in croccodilians, where pore opening alteration occurs by acidic disolution of calcium (Ferguson, 1985). Apparently, in the same specimen (UNC-CIP 11), disolution has also affected the internodal areas (figure 4.2). In UNC-CIP 10 (figure 5.3), it is possible to observe, throughout the pore openings, calcite filling the pore canals, possibly as result of dissolution and subsequent recrystalization.

Figure 5. 1-2, UNC-CIP 6, external polygonal pore / UNC-CIP 6, poro externo poligonal; 3, UNC-CIP 10, external subcircular pore / UNCCIP 10, poro externo subcircular; 4, UNC-CIP 5, internal subcircular pore / UNC-CIP 5, poro interno subcircular.
In UNC-CIP 6, pentagonal pore openings have diameters of approximately 155 mm (figures 5.1-2). As in other samples (UNC-CIP 2 [Mansilla II], figures 4.3-4), the polygonal contour of these openings seems to be determined by the proximity of contiguous nodes; however, as said above, it is possible that calcium dissolution has affected the rims of the pores, and modified their original contours. Yet, size pore normally varies within the same egg clutch, as observed in the Snare's penguin (Eudyptes robustus Oliver, 1953) (Massaro and Davis, 2005).
In megaloolithid eggs, pore canals are generally long, narrow and straight with funnel-like orifices on both inner and outer surfaces. Pore canals are normally placed between two eggshell units, as observed in UNC-CIP 9 (Mansilla II) (eggshell thickness 1.68 mm). In this case, the pore canals vary between 45 and 50µm (figure 8.6). The pore canals of the megaloolithids from Santa Rosa seem to be somewhat narrower than those from Auca Mahuevo (80-200 mm, Grellet-Tinner et al., 2004, fig. 4A), and decidedly wider than in Patagoolithus (5-10 mm, Simón, 2006, table 2).
Pore canals open on the internal surface of the eggshell, between the eggshell units (the intermammillary space) (figures 8.3-4). In UNC-CIP 5 (Mansilla I, figure 5.4) is clearly observed a subcircular internal pore opening with a diameter of 57 mm, which opens at the bottom of the irregular area between the truncated bases of the shell units. Unlike the external pore openings, internal pores are always subcircular to oval.
The megaloolithid eggshell is normally composed of a single layer of radiating shell units, as is characteristic in the so-called dinosauroid spherulitic "basic type" (Mikhailov et al., 1996). The existence of only one structural layer is shared by ornithischians, crocodilians and chelonians (Grellet-Tinner and Chiappe, 2004), which means that, from a phylogenetic perspective, it should be considered a dinosaur plesiomorphy (Grellet-Tinner, 2006b).
The eggshell units of the megaloolithids from Santa Rosa are fan-shaped, and the accretionary lines are normally arched, as typically occurs in the "structural morphotype" Discretispherulitic (Tubospherulitic), of the traditional classification of Mikhailov (1992). Arched accretionary lines are observed in UNC-CIP 7 (Mansilla I, figures 8.3-4) and, more clearly, in UFRRJ-1-3 (figures 9.1, 10); in this regard, megaloolithids from Salitral de Santa Rosa match best with the oogenus Megaloolithus (and in part with Patagoolithus) than with Cairanoolithus Vianey-Liaud et al., 1994 where the accretionary lines are rather subparallel to the surface (Vianey-Liaud et al., 1994).
The maximum width of the eggshell units ranges from 0.32 to 0.8 mm. In UNC-CIP 7 (eggshell thickness 1.2 mm), the borders of the eggshell units diverge with an angle of nearly 60º, whereas in UNCCIP 9 (eggshell thickness 1.68 mm) and UNC-CIP 3 (eggshell thickness 2.2 mm), this angle is usually somewhat lesser (of nearly 45º); in other words, the borders of the shell units are more parallel in UNCCIP 9 and UNC-CIP 3 than in other samples from Mansilla I and II, but not to the extent observed in Neuquén city (Calvo et al., 1997), Auca Mahuevo (Grellet-Tinner et al., 2004), and some european megaloolithids, such as Megaloolithus mammillare (Vianey- Liaud et al., 1994).
The fan-shaped eggshell units of the specimens from Santa Rosa have their greater diameter at their outer tips; this measurement ranges from 0.7-0.8 mm in UNC-CIP 7 (figures 8.3-4), 0.9 mm in UFRRJ-1 (figure 9.1) to 1.15 mm in UNC-CIP 3 (figure 8.1) (heigth/ width estimated relation of the shell units =1.71 in UNC-CIP 7, 2.15 in UFRRJ-1, and 1.82 in UNC-CIP 3). These values are congruent with those recorded by Calvo et al. (1997) in megaloolithids from Neuquén city (1.5 to 2.5), and in part with Patagoolithus salitralensis (heigth/width estimated relation 2.28) (Simón, 2006).
In UNC-CIP 3 (figure 8.2), UNC-CIP 7 (figure 8.3- 4), and in other samples, each eggshell unit extends in a basal, pillar-like structure, whose base is slightly expanded. These pillars contain the mammilary structures that range from 0.32 to 0.8 mm, a value partially coincident with the observed in Patagoolithus (0.09 to 0.81 mm, Simón, 2006).
In a radial section (figure 8.4), UNC-CIP 7 displays successive mammillary structures separated by a series of oval fissures. According to Grellet-Tinner et al. (2004), these fissures, as wide as the own basal pillars, would correspond to a network of horizontal pore canals with fitting vertical canals. Other authors (for instance, Elez and López-Martinez, 2000), however, identify these gaps as mere intermammillary spaces.
The diameter of the pore canals ranges from 35 to 40 mm in UNC-CIP 3, with intermammillary spaces of 70-75µm (figure 8.2). However, in UNC-CIP 7 (eggshell thickness 1.20 mm) the intermammillary spaces range from 80 (figure 8.4) to 150 (figure 8.5)µm.
In most examined eggshells, a severe disolution of calcium affected the internal surfaces; this process has practically eliminated the totality of the membrana testacea (MT), as well as part of the mammillary layer. In UNC-CIP 8 (figure 7.2), between the mammillary structures, there is a large amount of recrystalized calcite.
The mammillary structures present a shallow central pit topographically coincident with the organic central core (figures 6, 7.1-3, 8.5), possily linked to calcite dissolution during embriogenesis.

Figura 6. UNC-CIP 5, internal surface showing the coalescent mammillary structures (the truncated bases of the shell units) / UNC-CIP 5, superficie interna mostrando las estructuras mamilares coalescentes (las bases truncadas de las unidades de cáscara); lower arrows: reabsortion crates; upper arrows: pore openings / flechas inferiores: cráteres de reabsorción; flechas superiores, aberturas de poro.

Figure 7. 1-2, UNC-CIP 8, internal surface of the eggshell showing the mammillary structures and rebsortion crates (white arrow) / UNC-CIP 8, superficie interna de la cáscara mostrando las estructuras mamilares y cráteres de reabsorción (flecha blanca). 3, UNC-CIP 5, detail of the mammillary structure and rebsortion crater (white arrow) / UNC-CIP 5, detalle de la estructura mamilar y cráter de reabsorción (flecha blanca). 4, UNC-CIP 3, internal surface of the eggshell showing a severe diagenized surface / UNC-CIP 3, superficie interna de la cáscara mostrando una superficie severamente diagenizada.

Figure 8. Radial section of a megaloolithid / sección radial de un megaloolítido. 1, UNC-CIP 3, shell units and superficial nodes / UNC-CIP 3, unidades de cáscara y nodos superficiales. 2, UNC-CIP 3, oblique pore canal and basal pillars. The white arrow indicates the intermammillary spaces / UNC-CIP 3, canal de poro oblicuo y pilares basales. La flecha blanca muestra los espacios intermamilares. 3-4, UNC-CIP 7, fan-shaped eggshell units / UNC-CIP 7, unidades de cáscara en forma de abanico. 5, UNC-CIP 7, the pillars and the rebsortion crates (arrows) at their centres / UNC-CIP 7, los pilares y los cráteres de reabsorción (flecha) en sus centros. 6, UNC-CIP 9, the base of the shell units, showing a radial pore canal, and its end in a transversal pore canal / UNC-CIP 9, la base de las unidades de cáscara, mostrando un canal de poro radial, y su terminación en un canal de poro transversal.
In most specimens (e.g., UNC-CIP 7), the MT is clearly not preserved, whereas in others, it may be still partially preserved. In such instances, the internal surface of the eggshells is continuous, as observed in UNC-CIP 3 (figures 7.4, 8.2). Nevertheless, the solid calcite deposited on the inner surface of the eggshell could also be the result of diagenesis. In this regard, we must take into account that much of the original composition of the eggshell has been replaced by SiO2 (figure 9, white crystals).
The MT in the Auca Mahuevo megaloolithid eggs has been preserved in two different states: a fibrous one, in the form of multiple lines parallel to the surface (Grellet-Tinner et al., 2006, fig. 2B), and other in form of solid calcite (Grellet-Tinner et al., 2006, fig. 4b). The fact that the EDAX analysis of both surfaces of the eggshell UNC-CIP 2 had revealed a different elemental composition may be indicative of a partial preservation of the MT, but we are still unable to ensure that this is not the result of diagenetic alteration of the eggshell.

Figure 9. Radial sections of megaloolithid eggshell collected in Manantial de Mansilla (transmitted polarized light) / secciones radiales de cáscaras de megaloolítido colectadas en manantial de Mansilla (luz polarizada transmitida). 1-3, UFRRJ-1 (Mansilla I) / UFRRJ-1 (Mansilla I); 4, UFRRJ-2 (Mansilla II) / UFRRJ-2 (Mansilla II). The white crystals (arrow) are of diagenetic calcedonia / los cristales blancos (flecha) son de calcedonia diagenética.

Figure 10. UFRRJ-3 (Mansilla II). Arrow: Radial wedge line crossing sucessive arched accretionary lines / UFRRJ-3 (Mansilla II). Flecha: Cuña radial atravesando líneas de acreción arqueadas.
Discussion and conclusions
Identity of the thin-shelled fossil eggs from Salitral de Santa Rosa. The spherical, thin-shelled eggs found in Salitral de Santa Rosa are attributed to the oofamily Megaloolithidae (Zhao, 1979). Within Megaloolithidae, megaloolithids from Santa Rosa are assignable to Megaloolithus, according to Mohabley's (1996) diagnosis of this oogenus: "Dinosauroidspherulitic basic type; tubospherulitic morphotype; tubocanaliculate pore system; compact tuberculate ornamentation; spherical to subspherical eggs; eggshell thickness 1.5-3 mm". Moreover, the range of egghsell thickness observed in Mansilla (1.72), is not so far from the value proposed by Mohabey (1996) for the oogenus Megaloolithus (1.5). In fact, some individual species of Megaloolithus may present similarly high thickness ranges: M. jabalpurensis Khosla and Sahni, 1995, 1.3; M. siruguei Vianey-Liaud et al., 1994, 1.5; and M. cylindricus 1.8 (although Vianey- Liaud et al. [2003] suspect that the materials assigned to the last form may belong to two different oospecies).
We did not identify oospecies within Megaloolithus; at present, we only can state that some evidence (such as the wide range in eggshell thickness, which is normally not observed at oospecific level), suggests that more than one single oospecies could be distinguished in a future.
Relationships between the megaloolithids from Santa Rosa and other Patagonian thin-shelled fossil eggs. Megaloolithids from Santa Rosa are basically similar to those from Auca Mahuevo (which, in turn, were not ascribed to any oogenus in particular), Neuquén city (Megaloolithus patagonicus Calvo et al., 1997), and, in part, Salitral Moreno (Patagoolithus salitralensis). All Patagonian megaloolithid eggs are comparable in their relatively small size, surperficial ornamentation and microstructure. However, with respect to their size, the eggs from Salitral de Santa Rosa (at least those which are complete and well preserved) are seemingly somewhat smaller. On the other hand, the fossil eggs from Neuquén city and Auca Mahuevo have eggshell units whose margins are rather parallel, unlike those from Mansilla I and II, where the eggshell units are flared, especially those with a relatively thin eggshell (the "subtype 2A" of Salgado et al., 2007b). Patagoolithus, in turn, differs from all other Patagonian megaloolithids in their unusually narrow pore canals (diameter varying from 5 to 10 mm, Simón, 2006). Up to date, pathological megaloolithid eggshells have been only recorded in Auca Mahuevo (Jackson et al., 2004), but not in Neuquén city nor in Río Negro.
The Salitral de Santa Rosa eggs come from the Allen Formation of the Malargüe Group, which is stratigraphically higher than the unit that yielded the eggs of Neuquén city and the Anacleto Formation, whose age is estimated as Campanian (Dingus et al., 2000). Because they are recorded in different units, Patagonian megaloolithids, unlike their European relatives (García and Vianey-Liaud, 2001; Vianey- Liaud and López-Martínez, 1997), are actually not useful biochronological markers.
The overall morphological similarity among the diverse Patagonian megaloolithids does not mean that the specific sauropods that laid them are necessarily phylogenetically related. The relation existing between oological and biological diversity is still cause of debates; on this subject, it is possible to find different conjectures, specially in the parataxonomical literature. For instance, Vianey-Liaud et al. (2003) stated that one oogenus would correspond to one biological family or suborder of dinosaur (thus, they embrace Mikhailov's [1997] criterion, that many related biological genera may lay eggs of a same oogenus). This is a valid possibility in the case of the suborder Sauropoda, because the oogenus Megaloolithus has a biochron and geographic distribution (Upper Cretaceous of India, France, and Patagonia) that is unknown among the known sauropod genera. In short, we can not determine yet whether the Late Cretaceous megaloolithids from northern Patagonia were laid by one or many titanosaurian species.
Significance of variation in eggshell thickness. With reference to the biological meaning of the variation in eggshell thickness (the most palpable varying character in Santa Rosa), there are still unclear aspects. In the case of the eggs from Auca Mahuevo, it has been proposed that variation in thickness is mainly due to the different degree of diagenesis to which the eggs/eggshells have been submitted (Grellet-Tinner et al., 2004). With respect to the eggshells from Mansilla I and II, the fact that the relatively thick eggshells (Salgado et al.'s subtype "2B") consist of fan-shaped shell units, whose margins form a low angle with relation to the relatively thin eggshells (Salgado et al.'s subtype "2A"), suggests that the last ones are not mere diagenized thicker eggshells. On the other hand, and ignoring a possible (but improbable) taphonomic bias, the fact that the mean thickness varies from one locality to another, at least in the areas studied, as well as the existence of "clusters" within the subtype "2B" in the site Mansilla I (Salgado et al., 2007b), suggest that the eggshell thickness is not a random variable, but an attribute associate to taxonomic diversity (eggs produced by different species), or relative maturity of the female (produced by individuals of the same species but of different ages). Vianey-Liaud et al. (1994), Sahni et al. (1994), and Panadés I Blas (2005), favored the first explanation in the case of the megaloolithids from France, India, and Catalonia.
Nevertheless, the possibility that many or all of the ooforms from Santa Rosa belong to the same sauropod species, is equally probable. After all, the variation in thickness admited in some European oospecies matches well with the variation reported in the megaloolithids from Salitral de Santa Rosa. Here, the matter is to explain the reasons (to count with a plausible causal explanation) by which a single sauropod species lays eggs so different (according to the parataxonomical standards). Below, we suggest some possibilities for this.
It is well known that egg size is positively correlated to female body size (Hendry and Day, 2003); we also know that exists a general positive correlation between egg size and eggshell thickness, with larger eggs having thicker egghshells (Hirsch and Zelenitzky, 1997; Carpenter, 1999). Therefore, is reasonable that large females (the older ones?) lay relatively large eggs, with relatively thick eggshells. In this way, the variation in thickness recorded in the megaloolithid eggshells from Santa Rosa may be reflecting nothing but the populational structure of a single producer.
Other possibility (that is not incompatible with the previous one) emerges from the avian reproductive biology. In the case of the penguins, it has been demonstrated that the oldest females (not neccesarly the largest) lay larger eggs, whose eggshells are thicker, possibly because they utilize more efficiently the extracalcium (Massaro and Davis, 2004). Many authors affirmed that in some birds, eggs laid lately in the reproductive season have thinner eggshells, compared with those laid in earlier stages (Hipfner et al., 2001; but see Massaro and Davis, 2004, for a criticism). In other penguin species (e.g., Eudyptes robustus), some authors demonstrated that the egg laid in second term by a single female (and within a same reproductive season), has a thicker eggshell (also it is a bit larger) (Massaro and Davis, 2005). Analogous situations may be applied to the titanosaurs from Santa Rosa.
Also, crocodilian reproductive biology offers some clues to understand the heterogeneity of the eggshell thickness reported in the megaloolithids from Salitral de Santa Rosa. In these reptiles, eggshell thickness decreases during the incubation process (Peucker et al., 2005); therefore, theoretically, the eggshell of unfertilized and hatched eggs (including within a same clutch) will have different thickness. A comparable situation has been reported in dinosaur eggs (Horner, 1999), and it is not unlikely to have occurred among titanosaurs.
In sum, we can see that the heterogeneity in eggshell thickness documented in the megaloolithids from Río Negro, may not be due to diagenesis alone, but to multiple causes, such as age and size of the female, order in the sequence of successively laid eggs, etc., as well as to random variation. Of course (in Río Negro and elsewhere), the expected variation is not only in eggshell thickness, but also in egg-size, ornamentation, pore density, diameter of the pore canal, morphology and proportions of the eggshell units, etc. But we still do not know whether or not (and, if it is the case, to what extention) these variables are biologically correlated to eggshell thickness.
Ecology and reproductive behavior of titanosaurs. The environmental conditions under which the sauropods from Santa Rosa reproduced were undoubtedly different to those prevailing in Neuquén city and Auca Mahuevo; this clearly gives an idea on the versatility of these dinosaurs with respect to their reproductive behaviour. The paleoenvironment of the Allen Formation is usually interpreted as brackish lagoons in a supratidal environment, associated with aeolian sands (dunes) and deposits of ephemeral rivers. In Salitral de Santa Rosa-Salinas de Trapalcó, and Salitral Ojo de Agua, abundant rizoturbation, termite structures, and caliches in the egg-bearing deposits, indicate the presence of palaeosols, which presumably developed in an environment in close proximity to the coast. In Auca Mahuevo, on the contrary, the paleoenvironment is a fluvial system, with the largest concentration of eggs in the areas closer to the rivers. In fact, a few nests are found in the bed of the river (Garrido, 2005). In Neuquén city, megaloolithids are in fluvial sandstones deposited in braid plains, which show some pedogenic features that suggest soil development (Calvo et al., 1997).
In Neuquén city and Auca Mahuevo, megaloolithids are the only recorded type of fossil egg; probably, in these places, reproducing dinosaurs aggregated in monospecific colonies. In some sites of Salitral de Santa Rosa and Salitral Ojo de Agua, on the contrary, megaloolithids are intimately associated to eggs of "type I" (faveoloolithids, Coria et al., 2006; Salgado et al., 2007b), and, at least in Arriagada III (Salitral Ojo de Agua), to eggs (and bones) of nonavian and avian theropods (Salgado et al., 2007b; Salgado et al., 2009). However, the association of different types of fossil eggs is not universal in Salitral de Santa Rosa; in some sites there are only megaloolithids, whereas in others, only faveoloolithids (Salgado et al., 2007b). We provisionally interpret that the colonies were in fact monospecific (as proposed in Neuquén city and Auca Mahuevo), and that adjacent colonies of different species (megaloolithid and faveoloolithid-laying dinosaurs) were partially superimposed. Hypothetically, the area that surrounds the town of Bajo de Santa Rosa (sites Santa Rosa, Santos, etc., Salgado et al., 2007b), which have yielded only in situ eggs of "type I", was close to the center of the colony of the faveoloolithid-laying dinosaurs; in turn, Mansilla I and II were close to the center of the colony of the megaloolithid-laying dinosaurs (presumably titanosaurian sauropods) (although in this last case, we must remember that there are not in situ eggs). In turn, García I, Cerro Bonaparte, and Berthe II-V, in sum, all those sites where both eggshell morphotypes are found together, were probably common to more than one colony. Perhaps, territory overlapping was possible because the egglaying dinosaurs would not have brood their clutches, which is inferred from phylogenetic oological characters (Grellet-Tinner et al., 2006). Although all these ideas are not but mere speculations, these are illustrative of the wide range of possibilities that reproductive scenarios from Río Negro offer to understand diverse aspects of the dinosaurian reproductive behavior, particularly, of titanosaurs.
Acknowledgements
The authors are deeply in debt with D. Cabaza (Director of the Museum of Lamarque, Río Negro) and collaborators L. Zuain, M. Villablanca, A. Villablanca, N. Alarcón, E. Pincheira, R. Passadore. Our thanks also to L. Berthe, M. García, J. Rosales, "Beto" Arriagada, A. Santos, the Mansilla family for providing information and logistical support during field work; M. I. Kopp (Director of the Museo Regional de Valcheta, Río Negro) and the Municipality of Valcheta, for their valuable assistance during the fieldworks; R. Coria and his crew, for their scientific contribution to the development of this project; G. Grellet-Tinner and F. Jackson for reading an early version of the manuscript; the reviewers R. Coria and Á. Galobart. This study was supported by National Geographic (grant 7396-03), CONICET (PIP 6455), ANPCT (PICT-2006-00357), Municipalities of Lamarque and Valcheta (Río Negro), and Plaza Huincul (Neuquén). This was accomplished by means of an agreement subscribed with the Agencia Cultura Río Negro.
References
1. Andreis, R.A., Iñiguez, A.M., Rodríguez Lluch, J.J. and Sabio, D.A. 1974. Estudio sedimentológico de las formaciones del Cretácico Superior del área del Lago Pellegrini (Prov. de Río Negro, Rep. Argentina). Revista de la Asociación Geológica Argentina 29: 85-104. [ Links ]
2. Ballent, S.C. 1980. Ostrácodos de ambiente salobre de la Formación Allen (Cretácico Superior) en la Provincia de Río Negro (República Argentina). Ameghiniana 17: 67-82. [ Links ]
3. Calvo, J.O., Engelland, S., Heredia, S.E. and Salgado, L. 1997. First record of dinosaur eggshells (?Sauropoda-Megaloolithidae) from Neuquén, Patagonia. Gaia 14:23-32. [ Links ]
4. Carpenter, K. 1999. Eggs, Nests, and Baby Dinosaurs. Indiana University Press, Bloomington, 336 p. [ Links ]
5. Chiappe, L.M., Salgado, L. and Coria, R.A. 2001. Embryonic skulls of titanosaur sauropod dinosaurs. Science 293: 2444-2446. [ Links ]
6. Chiappe, L.M., Coria, R.A., Dingus, L., Jackson, F., Chinsamy, A. and Fox, M. 1998. Sauropod dinosaur embryos from the Late Cretaceous of Patagonia. Nature 396: 258-261. [ Links ]
7. Coria, R.A., Salgado, L. and Chiappe, L.M. 2006. Multiple dinosaur egg-shell occurrence in an Upper Cretaceous nesting site from Patagonia. 3º International Symposium On Dinosaur Eggs, Babies, and Developmental Biology (Plaza Huincul, Neuquén, Argentina), Abstracts, p. 1. [ Links ]
8. Dingus, L., Clarke, J., Scott, G.R., Swisher, C.C., Chiappe, L.M. and Coria, R.A. 2000. Stratigraphy and magnetostratigraphic/ faunal constraints for the age of sauropod embryo-bearing rocks in the Neuquén Group (Late Cretaceous, Neuquén Province, Argentina). American Museum Novitates 3290: 1-11. [ Links ]
9. Elez, J. and López-Martínez, N. 2000. Interrelationships between growth of mineral phase and pore system in dinosaur eggshells. 1º International Symposium on Dinosaur Eggs and Babies (Isona i Conca Della, Catalonia, Spain), Extended Abstracts, pp. 43-49. [ Links ]
10. Ferguson, M.W.J. 1985. Reproductive biology and embryology of the crocodilians. In: C. Gans, F. Billet and P. T. A. Maderson (eds.), Biology of the Reptilia, vol. 14, John Wiley and Sons, New York, pp. 329-491. [ Links ]
11. Frey, R.W. and Pemberton, S.G. 1987. The Psilonichnus ichnocoenose, and its relationships to adjacent marine and nonmarine ichnocoenoses along the Georgia coast. Bulletin of Canadian Petroleum Geology 35: 333-357. [ Links ]
12. García, R.A. 2007. An "egg-tooth"-like structure in titanosaurian sauropod embryos. Journal of Vertebrate Paleontology 27: 247-252 [ Links ]
13. García, G. and Vianey-Liaud, M. 2001. Dinosaur eggshells as biochronological markers in Upper Cretaceous continental deposits. Palaeogeography, Palaeoclimatology, Palaeoecology 169: 153-164. [ Links ]
14. García, G., Marivaux, L., Pelissié, T. and Vianey-Liaud, M. 2006. Earliest Laurasian sauropod eggshells. Acta Palaeontologica Polonica 51: 99-104. [ Links ]
15. Garrido, A.C. 2005. Auca Mahuevo nesting site: paleoenvironmental reconstruction and evolution. Ameghiniana 42: 69R. [ Links ]
16. Grellet-Tinner, G. 2006a. Oology and the Evolution of thermophysiology in saurischian dinosaurs: homeotherm and entotherm deinonychosaurians? Papéis Avulsos de Zoologia 46: 1- 10. [ Links ]
17. Grellet-Tinner, G. 2006b. Phylogenetic interpretation of eggs and eggshells: Implications for Phylogeny of Palaeognathae. Alcheringa 30: 141-182. [ Links ]
18. Grellet-Tinner, G. and Chiappe, L.M. 2004. Dinosaur eggs and nestings: implications for understanding the origin of birds. In: P.J. Currie, E.B. Koppelhus, M.A. Shugar and J.L. Wright (eds.), Feathered Dragons: Studies on the Transition from Dinosaurs to Birds. Indiana University Press, Bloomington, pp. 184-214. [ Links ]
19. Grellet-Tinner, G., Chiappe, L.M. and Coria, R.A. 2004. Eggs of titanosaurid sauropods from the Upper Cretaceous of Auca Mahuevo (Argentina). Canadian Journal of Earth Sciences 41: 949-960. [ Links ]
20. Grellet-Tinner, G., Chiappe, L.M., Norell, M. and Bottjer, D. 2006. Dinosaur eggs and nesting behaviors: a paleobiological investigation. Palaeogeography, Palaeoclimatology, Palaeoecology 232: 294-321. [ Links ]
21. Grigorescu, D. 2006. Hatchlings of Telmatosaurus transsylvanicus (Ornitischia, Hadrosauridae) associated with megaloolithid eggs in the Tustea nesting site (Hateg Basin, Romania). 3º International Symposium on Dinosaur Eggs, Babies, And Developmental Biology (Plaza Huincul, Neuquén, Argentina), Abstracts, pp. 6-7. [ Links ]
22. Grigorescu, D., Weishampel, D.B., Norman, D., Seclamen, M., Rusu, M., Baltres, A. and Teodorescu, V. 1994. Late Maastrichtian dinosaur eggs from the Hateg Basin (Romania). In: K. Carpenter, K.F. Hirsch and J.R. Horner (eds.), Dinosaur Eggs and Babies. Cambridge University Press, Cambridge, pp. 75-87. [ Links ]
23. Hendry, A. P. and Day, T. 2003. Revisiting the positive correlation between female size and egg size. Evolutionary Ecology Research 5: 421-429. [ Links ]
24. Hipfner, J.M., Gaston, A.J., Martin, D.L. and Jones, I.L. 2001. Seasonal declines in incubation periods of Brünnich's Guillemots Uria lomvia: testing proximate causes. Ibis 143: 92-98. [ Links ]
25. Hirsch, K.F. and Zelenitzky, D.K. 1997. Dinosaur eggs. In: J.O. Farlow and M.K. Brett Surman (eds.), The Complete Dinosaur. Indiana University Press, Bloomington, pp. 394-402. [ Links ]
26. Horner, J.R. 1999. Egg clutches and embryos of two hadrosaurian dinosaurs. Journal of Vertebrate Paleontology 19: 607-611. [ Links ]
27. Hugo, C.A. and Leanza, H.A. 2001. Hoja Geológica 3966-III Villa Regina. Boletín Segemar 309: 1-58. [ Links ]
28. Jackson, F.D., Garrido, A.C., Schmitt, J.G., Chiappe, L.M., Dingus, L. and Loope, D.B. 2004. Abnormal Multilayered titanosaur (Dinosauria: Sauropoda) eggs from in situ clutches at the Auca Mahuevo Locality, Neuquén Province, Argentina. Journal of Vertebrate Paleontology 24: 913-922. [ Links ]
29. Khosla, A. and Sahni, A. 1995. Parataxonomic classification of Late Cretaceous dinosaur eggshells from India. Journal Palaeontological Society of India 40: 87-102. [ Links ]
30. Leanza, H.A. and Hugo, C.A. 2001. Cretaceous red beds from southern Neuquén Basis (Argentina): age, distribution and stratigraphic discontinuities. 7º International Symposium on Mesozoic Terrestrial Ecosystem, Asociación Paleontológica Argentina, Publicación Especial 7: 117-122. [ Links ]
31. Martinelli, A.G. and Forasiepi, A.M. 2004. Late Cretaceous vertebrates from Bajo de Santa Rosa (Allen Formation), Río Negro province, Argentina, with the description of a new sauropod dinosaur (Titanosauridae). Revista del Museo Argentino de Ciencias Naturales 6: 257-305. [ Links ]
32. Massaro, M. and Davis, L.S. 2004. The influence of laying date and maternal age on eggshell thickness and pore density in yellow- eyed penguins. The Condor 106: 496-505. [ Links ]
33. Massaro, M. and Davis, L.S. 2005. Differences in egg size, shell thickness, pore density, pore diameter and water vapour conductance between first and second eggs of Snare's Penguins Eudyptes robustus and their influence on hatching asynchrony. Ibis 147: 251-258. [ Links ]
34. Mikhailov, K.E. 1991. Classification of fossil eggshells of amniotic vertebrates. Acta Palaeontologica Polonica 36: 193-238. [ Links ]
35. Mikhailov, K.E. 1992. The microstructure of avian and dinosaurian eggshell: phylogenetic implications. In K. Campbell (ed.), Papers in Avian Physiology Honoring Pierce Brodkord. Los Angeles, Natural History Museum of Los Angeles County, pp. 361-373. [ Links ]
36. Mikhailov, K.E. 1997. Fossil and recent eggshell in amniotic vertebrates: fine structure, comparative morphology and classification. Special Papers in Palaeontology 56: 1-80. [ Links ]
37. Mikhailov, K.E., Bray, E.S. and Hirsch, K.F. 1996. Parataxonomy of fossil egg remains (Veterovata): Principles and applications. Journal of Vertebrate Paleontology 16: 763-769. [ Links ]
38. Mohabey, D.M. 1996. A new oospecies, Megaloolithus matleyi, from the Lameta Formation (Upper Cretaceous) of Chandrapur district, Maharashtra, India, and general remarks on the palaeoenvironment and nesting behaviour of dinosaurs. Cretaceous Research 17: 183-196. [ Links ]
39. Mohabey, D.M. 1998. Systematics of Indian Upper Cretaceous dinosaur and chelonian eggshells. Journal of Vertebrate Paleontology 18: 342-362. [ Links ]
40. Mones A. 1980. Nuevos elementos de la paleoherpetofauna del Uruguay (Cocodrilia y Dinosauria). 2º Congreso Argentino de Paleontología y Bioestratigrafía, y 1º Latinoamericano de Paleontología (Buenos Aires), Actas 1: 265-274. [ Links ]
41. Novas, F.E., Pol, D., Canale, J.I., Porfiri, J.D. and Calvo, J.O. 2008. A bizarre Cretaceous theropod dinosaur from Patagonia and the evolution of Gondwann dromaeosaurids. Proceedings of the Royal Society B, doi: 10.1098/rspb.2008.1554. [ Links ]
42. Oliver, W.R.B. 1953. The crested penguins of New Zealand. Emu 53: 185-187. [ Links ]
43. Panadés I Blas, X. 2005. Diversity versus variability in Megaloolithid dinosaur eggshells. www.PalArch.nl, vertebrate palaeontology 2: 1-13. [ Links ]
44. Parodiz, J.J. 1969. The Tertiary non-marine mollusca of South America. Annals of the Carnegie Museum 40: 1-242. [ Links ]
45. Parras, A. and Casadío, S. 1999. Paleogeografía del sector septentrional de la cuenca Neuquina durante el intervalo Campaniano- Daniano. 7º Jornadas Pampeanas de Ciencias Naturales (Santa Rosa, La Pampa), Actas, pp. 261-268. [ Links ]
46. Parras, A., Casadío, S. and Pires, M. 1998. Secuencias depositacionales del Grupo Malargüe y el límite Cretácico-Paleógeno, en el sur de la provincia de Mendoza, Argentina. In S. Casadío (ed.), Paleógeno de América del Sur y de la Península Antártica. Publicación Especial de la Asociación Paleontológica Argentina, Actas 5: 61-69. [ Links ]
47. Peitz, C. 2000. Megaloolithid dinosaur eggs from the Maastrichtian of Catalunya (NE-Spain)- Parataxonomic implications and stratigraphic utility. In A. M. Bravo and T. Reyes (eds.), 1º International Symposium on Dinosaur Eggs and Babies (Isona i Conca Dellà, Catalonia, Spain), Extended Abstracts, pp. 155-159. [ Links ]
48. Peucker, S.K.J., Davis, B.M. and Van Barneveld, R.J. 2005. Crocodile Farming Research: Hatching to Harvest. Australian Government, Rural Industries Research and Development Corporation, Special Publication Nro. 05/152. 34 pp. [ Links ]
49. Rougier, G.W., Chornogubsky, L., Casadío, S., Paéz Arango, N. and Giallombardo, A. 2009. Mammals from the Allen Formation, Late Cretaceous, Argentina. Cretaceous Research 30: 223-238. [ Links ]
50. Sahni, A., Tandon, S.K., Jolly, A., Bajpai, S., Sood, A. and Srinivasan, S. 1994. Upper Cretaceous dinosaur eggs and nesting sites from the Deccan volcano-sedimentary province of peninsular India. In: K. Carpenter, K.F. Hirsch and J.R. Horner (eds.), Dinosaur Eggs and Babies. Cambridge University Press, Cambridge, pp. 204-226. [ Links ]
51. Salgado, L., Coria, R.A. and Chiappe, L.M. 2005. Anatomy of the sauropod embryos from Auca Mahuevo (Upper Cretaceous), Neuquén, Argentina. Acta Palaeontologica Polonica 50: 79-92. [ Links ]
52. Salgado, L., Parras, A. and Gasparini, Z. 2007a. Un plesiosaurio de cuello corto (Plesiosauroidea, Polycotylidae) del Cretácico Superior del norte de Patagonia. Ameghiniana 44: 349-358. [ Links ]
53. Salgado, L., Coria, R.A., Arcucci, A.B. and Chiappe, L.M. 2009. Restos de Alvarezsauridae (Theropoda, Coelurosauria) en la Formación Allen (Campaniano-Maastrichtiano), en Salitral Ojo de Agua, Provincia de Río Negro, Argentina. Andean Geology 36: 67-80. [ Links ]
54. Salgado, L., Coria, R.A., Magalhães Ribeiro, C.M., Garrido, C.A., Rogers, R., Simón, M.E., Arcucci, A.B., Curry Rogers, K., Paulina Carabajal, A., Apesteguía, S., Fernández, M.S., García, R.A. and Talevi, M. 2007b. Upper Cretaceous dinosaur nesting sites of Río Negro (Salitral Ojo de Agua and Salinas de Trapalcó-Salitral de Santa Rosa), Northern Patagonia, Argentina. Cretaceous Research 28: 392-404. [ Links ]
55. Schweitzer, M.H., Chiappe, L.M., Garrido, A.C., Lowenstein, J.M. and Pincus, S.H.. 2004. Molecular preservation in late Cretaceous sauropod dinosaur eggshells. Proceedings of the Royal Society 2876: 1-10. [ Links ]
56. Simón, M.E. 1999. Estudio De Fragmentos De Cáscaras De Huevos De La Formación Allen (Campaniano-Maastrichtiano), Provincia De Río Negro, Argentina. Graduation Dissertation, Universidad Nacional de Córdoba, Córdoba, 249 pp. Unpublished. [ Links ]
57. Simón, M.E. 2006. Cáscaras de huevos de dinosaurios de la Formación Allen (Campaniano-Maastrichtiano), en Salitral Moreno, Provincia de Río Negro, Argentina. Ameghiniana 43: 513-528. [ Links ]
58. Spix, J.B. and Wagner, J.A. 1827. Testacea Fluviatilia Brasiliam, 36 pp. Leipzig. [ Links ]
59. Vianey-Liaud, M., Mallan, P., Buscail, O. and Montgelard, C. 1994. Review of French dinosaur eggshells: Morphology, structure, mineral, and organic composition. In: K. Carpenter, K. F. Hirsch and J. R. Horner (eds.), Dinosaur Eggs and Babies. Cambridge University Press, New York, pp. 151-183. [ Links ]
60. Vianey-Liaud, M. and López-Martínez, N. 1997. Late Cretaceous dinosaur eggshells from the Tremp Basin, Southern Pyrenees, Lleida, Spain. Journal of Paleontology 71: 1157-1171. [ Links ]
61. Vianey-Liaud, M., Kholasla, A. and García, G. 2003. Relationships between European and Indian dinosaur eggs and eggshells of the oofamily Megaloolithidae. Journal of Vertebrate Paleontology 23: 575-585. [ Links ]
62. Zhao, Z. 1975. The microstructure of dinosaurian eggshells of Nanshsiung Basin, Guangdong Province. Vertebrata PalAsiatica 17: 131-136. [ Links ]
63. Zhao, Z. 1979. Progress in the research of dinosaur eggs. In:: Mesozoic and Cenozoic Red Beds of South China. Beijing: Science Press, pp. 330-340. [ Links ]
64. Zhao, Z. and S. Ding. 1976. Discovery of the dinosaurian eggshells from Alxa, Ningxia and its stratigraphic significance. Vertebrata PalAsiatica 14: 42-44. [ Links ]
Recibido: 7 de abril de 2009.
Aceptado: 15 de julio de 2009.

