










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana vol.47 no.1 Buenos Aires ene./mar. 2010
ARTÍCULOS ORIGINALES
Palinologia das formações Rio Bonito e Palermo (Permiano Inferior, Bacia do Paraná) em Candiota, Rio Grande do Sul, Brasil: novos dados e implicações bioestratigráficas
Ana L.O. Mori1 e Paulo A. Souza2
1Departamento de Paleontologia e Estratigrafia, Instituto de Geociências, Programa de Pós-Graduação em Geociências, Universidade Federal do Rio Grande do Sul. Avenida Bento Gonçalves, 9500, CEP 91.509-000, Porto Alegre, RS, Brasil. luisaouta@yahoo,com.br; paulo.alves.souza@ufrgs.br
Resumo. Novos conjuntos palinológicos foram recuperados de um afloramento localizado às margens da BR 293, situado entre os municípios de Bagé e Candiota, sul do Estado do Rio Grande do Sul, correspondentes às formações Rio Bonito e Palermo, Permiano Inferior da Bacia do Paraná. Um total de oito amostras foi estudado, revelando palinofloras ricas e diversificadas, com a identificação de 76 espécies, dentre as quais 42 são relativas a esporos, 24 a grãos de pólen, e 10 relacionadas a algas e fungos. Em termos bioestratigráficos, palinomorfos documentados na seqüência sedimentar analisada permitiram identificar duas unidades bioestratigráficas na seção, as zonas Vittatina costabilis e Lueckisporites virrkiae. Algumas espécies de distribuição estratigráfica limitada a determinados níveis do Subgrupo Itararé e da Formação Rio Bonito foram registrados em posição estratigráfica mais superior, podendo sugerir, à primeira vista, aumento na amplitude estratigráfica destes táxons. Entretanto, outras implicações estratigráficas são também admitidas, e constituem parte da discussão da presente contribuição.
Abstract. Palynology of the Rio Bonito and Palermo formations (Lower Permian, Paraná Basin) in Candiota, Rio Grande do Sul, Brazil: new data and biostratigraphic implications. New palynologic associations were recovered from an outcrop located at BR 293 Highway, between Bagé and Candiota municipalities, Rio Grande do Sul state, Brazil, related to the Rio Bonito and Palermo formations, Lower Permian of Paraná Basin. Eight samples were colected and studied, showing rich and diversified palynofloras. A total of 76 species were recognized, 42 corresponding to spores, 24 to pollen grains and 10 related to algae and fungi. Palynomorphs registered in the section here analysed indicate the recognition of the Vittatina costabilis and Lueckisporites virrkiae zones. Species with stratigraphic ranges limitated to Itararé Subgroup and Rio Bonito Formation were identified in a higher stratigrapical levels, meaning for instance, increasing on the stratigraphic range of these taxa. On the other hand, other stratigraphic interpretations are possible, constituting the scope of the present contribution.
Palavras-chave. Palinologia; Permiano Inferior; Bacia do Paraná; Bioestratigrafia.
Key words. Palynology; Lower Permian; Paraná Basin; Biostratigraphy.
Introdução
A sucessão sedimentar pensilvaniana e permiana da Bacia do Paraná é constituída por depósitos vinculados ao final da glaciação que atingiu grande parte do Gondwana, com representação nos estratos do Subgrupo Itararé, base do Grupo Tubarão, e pelos depósitos pós-glaciais relacionados à progressiva melhoria climática do Permiano, materializados nas demais unidades constituintes dos grupos Tubarão e Passa Dois. No contexto pós-glacial, a abundante paleoflora registrada no Grupo Tubarão, destacadamente na da Formação Rio Bonito, permitiu o desenvolvimento de extensos depósitos de carvão, concentrados principalmente nos estados do Rio Grande do Sul e de Santa Catarina, porção sul da Bacia do Paraná.
No município de Candiota, sudoeste do Rio Grande do Sul ocorrem as mais espessas camadas de carvão do país, o que motivou uma grande produção de trabalhos de cunho estratigráfico, seja para um melhor reconhecimento e identificação das camadas economicamente viáveis à exploração (e.g. Fontes e Cava, 1980; Menezes-Filho e Brito, 1982; Wildner, 1983; Lopes et al., 1986; Picolli et al., 1986; Lavina e Lopes, 1987) ou para o melhor entendimento do comportamento e evolução da bacia na sua seção sul, incluindo a aplicação de conceitos da Estratigrafia de Seqüências (Alves e Ade, 1996; Holz, 1998; Holz et al., 2006).
Macrofósseis vegetais e megásporos são conhecidos na área desde o século XIX. Entretanto, palinomorfos são relativamente mais abundantes, constituindo a ferramenta bioestratigráfica mais importante, uma vez que permitem correlações locais e regionais dos depósitos da bacia.
Embora datações radiométricas sejam disponíveis para determinados níveis da Formação Rio Bonito na Mina de Candiota (Matos et al., 2001; Guerra-Sommer et al., 2005, 2008a, 2008b), as informações palinoestratigráficas associadas são carentes de maior aprofundamento. Portanto, a análise é bastante limitada, seja pela ausência de amarração bioestratigráfica dos níveis onde os palinomorfos ocorrem, seja por falta de ilustrações e ou descrições originais, impedindo comparações com o zoneamento vigente para a bacia.
Desta forma, este trabalho objetiva apresentar o conteúdo palinológico de oito amostras coletadas em um afloramento inédito, localizado às margens da BR 293, entre os municípios de Bagé e Candiota, correspondentes às formações Rio Bonito e Palermo, discutindo as implicações bioestratigráficas das associações palinológicas recuperadas, à luz dos zoneamentos palinobioestratigráficos disponíveis para a bacia (Daemon e Quadros, 1970; Souza e Marques-Toigo, 2003, 2005; Souza, 2006). Para uma melhor resolução e entendimento das questões bioestratigráficas, os táxons identificados foram apresentados sob a forma de listagem taxonômica, com ilustrações e respectiva distribuição estratigráfica, fornecendo uma base de dados mais segura para o detalhamento dos limites das palinozonas paleozóicas para a bacia. Além disso, o nível de tonstein presente no afloramento permitiu a obtenção de datações radiométricas, dados que estão sendo tratados em trabalho à parte, e que, incorporados às novas informações aqui expostas, permitirão melhorar o posicionamento das unidades palinobioestratigráfias em termos biocronoestratigráficos, ampliando a integração das informações intra e extrabacinais.
Contexto geológico e paleontológico
Geologia local
A Bacia do Paraná é uma grande bacia intracratônica sul-americana, cobrindo cerca de 1.700.000 km2 em área, distribuída em porções territoriais do Brasil, Uruguai, Argentina e Paraguai (figura 1.1), envolvendo uma grande seqüência sedimentar, que excede 6.000 metros em seu depocentro. De acordo com Milani (1997), seis grandes unidades aloestratigráficas são identificadas para a bacia, nomeadas como Supersequência Rio Ivaí (litoestratigraficamente relacionada ao Grupo Rio Ivaí, de idade Ordoviciano/Siluriano), Paraná (Grupo Paraná, Devoniano), Gondwana I (grupos Tubarão e Passa Dois, Pensilvaniano/Permiano), Gondwana II (unidades triássicas), Gondwana III (Grupo São Bento, Jurássico/Cretáceo) e Bauru (Grupo Bauru, Cretáceo).
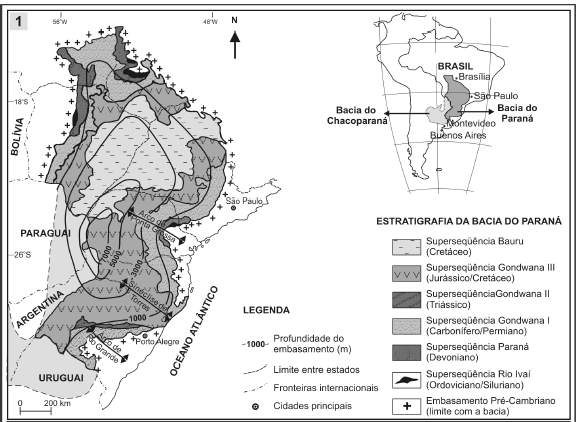
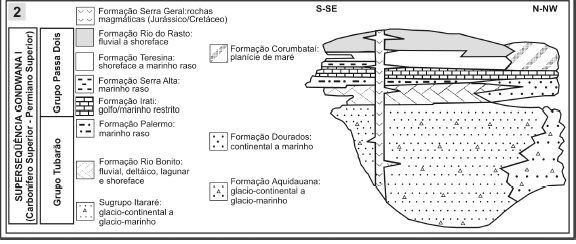
Figura 1. 1, Localização e subdivisão aloestratigráfica da Bacia do Paraná / location and alostratigraphic subdivision of Paraná Basin; 2, Detalhe das unidades constituintes da Superseqüência Gondwana I (todas as figuras modificadas de Milani, 1997) / details of components units of Gondwana I Supersequence (all figures modified from Milani, 1997).
A Supersequência Gondwana I (figura 1.2) corresponde a um grande ciclo transgressivo-regressivo, sendo referente, em termos litoestratigráficos, às unidades do Grupo Tubarão (Subgrupo Itararé, formações Aquidauana, Rio Bonito, Palermo e Tatuí), de idade entre o Pensilvanino (Bashkiriano/Moskoviano) ao Cisuraliano (Artinskiano), e ao Grupo Passa Dois, onde estão incluídas as formações Irati, Serra Alta, Teresina, Corumbataí e Rio do Rasto, de idade entre o Cisuraliano (Artinskiano) e o Lopingiano (Wuchiapingiano?). Esta supersequência compreende um espesso pacote sedimentar, litologicamente heterogêneo, relacionado a processos e paleoambientes de natureza distinta, resultado das expressivas mudanças ambientais ocorridas no setor ocidental do Gondwana, tal como sintetizado em Milani e Zalán (1999).
Segundo Milani (1997), os depósitos das formações Rio Bonito e Palermo estão associados às condições pós-glaciais do Permiano Inferior. A primeira é predominantemente constituída por arenitos fluviais e associações de pelitos, originados a partir de ambientes lagunares e deltaicos, incluindo espessas camadas de carvão. Por outro lado, a Formação Palermo é constituída por siltitos e argilitos, interpretados como de origem marinha plataformal, representando os depósitos de máxima inundação da Supersequência Gondwana I. Na porção nordeste da bacia, estas unidades são parcialmente correlatas à Formação Tatuí Estado de São Paulo, composta por arenitos, siltitos e calcários, interpretados também como de origem marinha, ou à Formação Dourados ao norte, com predomínio de arenitos (figura 1.2).
Síntese dos dados paleontológicos prévios
A listagem de trabalhos de cunho paleobotânico e palinológico desenvolvidos nos depósitos pensilvanianos e permianos da Bacia do Paraná é relativamente extensa. Particularmente sobre as formações Rio Bonito e Palermo, a maior parte é procedente de diversas localidades do Estado do Rio Grande do Sul (e.g., Cauduro, 1970; Dias-Fabrício, 1981; Picarelli et al., 1986), focando, principalmente, os carvões e rochas associadas.
Os primeiros registros fósseis para a região de Candiota resultam das publicações sobre achados paleobotânicos (incluindose megásporos e miósporos associados), a partir dos trabalhos de Carruthers (1869), Plant (1869), Hartt (1870), Liais (1872), Zeiller (1895), Lundquist (1920) e posteriormente Trindade (1954) e Dijkstra (1955).
Estudos palinológicos dos carvões e rochas associadas das formações Rio Bonito e Palermo na área foram desenvolvidos por vários autores a partir da década de 60 do século passado. As listas de táxons são relativamente extensas, incluindo diversas proposições taxonômicas. Ao longo dos últimos anos, diversos desses táxons foram revistos e sinonimizados por outros autores. Considerando os objetivos do presente trabalho, a seguir é apresentada uma síntese sobre o significado bioestratigráfico de cada uma dessas contribuições, de modo a ressaltar sua importância e aspectos problemáticos.
As primeiras análises essencialmente palinológicas para a região de Candiota foram realizadas por Pant e Srivastava (1965), relativas a uma amostra procedente de Candiota além de uma segunda localidade não especificada do Estado de Santa Catarina. O material foi reestudado por Tiwari e Navale (1967), segundo os quais o conteúdo das duas amostras seria distinto do ponto de vista quantitativo, sugerindo que poderiam representar diferentes fácies deposicionais. Em termos de idade, a comparação entre as duas associações palinológicas permitiu inferir idade mais antiga para o material analisado do Estado do Rio Grande do Sul do que aquele coletado em Santa Catarina.
A amostragem do material publicado por Nahuys et al. (1968) é oriunda de diversas localidades da Formação Rio Bonito dos estados do sul do Brasil, incluindo amostras procedentes de Candiota. Segundo os autores, os depósitos de carvão da região seriam de idade permiana, contrariando a atribuição de idade carbonífera proposta por autores prévios (e.g., Beurlen, 1953; Dolianiti, 1953). Embora a amostragem tenha sido relativamente ampla e exaustiva, a carência de informações sobre as procedências geográfica ou estratigráfica para cada táxon, dentro da Formação Rio Bonito, dificulta uma melhor análise bioestratigráfica e paleoambiental.
O trabalho de Ybert (1975) é baseado em amostras de testemunho de sondagem e afloramentos exclusivamente da Formação Rio Bonito, associadas aos depósitos de carvão da região dos municípios de Candiota e Hulha Negra, incluindo perfis estratigráficos e dados sobre a localização estratigráfica. Entretanto, notase que somente os níveis de carvão foram amostrados, o que era muito comum na época, uma vez que o foco principal dos estudos era voltado para o estudo desse recurso, visando seu aproveitamento econômico. Esta tendência perdurou por muito tempo depois, resultando em um conjunto de dados que reflete muito mais o contexto local - da palinoflora formadora do carvão - do que as associações faciológicas como um todo. Além disso, tal como em Nahuys et al. (1968), o autor não detalhou o local de ocorrência das espécies documentadas, informação que é fornecida somente para as novas espécies propostas ou para os espécimes fotografados.
Na mesma época, Marques-Toigo et al. (1975) apresentaram gêneros de megásporos e micrósporos identificados do carvão de Candiota, cujo estudo foi aprofundado por Corrêa da Silva e Marques-Toigo (1975). Além dos níveis de carvão, as autoras amostraram níveis pelíticos e de tonstein, com análises quantitativas por nível estratigráfico, com considerações do ponto de vista paleoecológico. Além de palinomorfos de origem continental (esporomorfos, zignematáceas), já conhecidos para os depósitos de carvão da região de Candiota, microfósseis de origem marinha (Navifusa e Cymatiosphaera) foram documentados por Meyer e Marques-Toigo (2000), permitindo corroborar esquemas estratigráficos que propunham ingressões marinhas no sistema formador de carvões (Alves e Ade, 1996; Holz, 1998).
As contribuições realizadas por Cazzulo-Klepzig et al. (2002, 2005) e Guerra-Sommer et al. (2008a, 2008b) apresentam listas de táxons ocorrentes na região com base em trabalhos prévios, alguns dos quais ilustrados, embora parte deles seja proveniente de outras localidades do Estado (vide Santos et al., 2006, p. 461). Em Guerra-Sommer et al. (2008a), citase que Protohaploxypinus goraiensis e espécies do gênero Stritatopodocarpites teriam sido documentadas na Mina de Candiota (Cazzulo-Klepzig et al., 2005). O registro destes palinomorfos permitiu aos autores o posicionamento da seção analisada na Subzona Protohaploxypinus goraiensis e a atribuição da datação radiométrica obtida por eles à unidade bioestratigráfica identificada. Entretanto, apenas o segundo gênero é citado em Cazzulo-Klepzig et al. (2005), sem ilustrações. Dessa forma, a análise sobre o significado bioestratigráfico tornase prejudicada.
Palinoestratigrafia da Bacia do Paraná
Diversos esquemas bioestratigráficos foram propostos para o intervalo Pensilvaniano a Permiano da Bacia do Paraná, dentre os quais o de Daemon e Quadros (1970) é o mais abrangente, seja em termos estratigráficos e geográficos (vide Souza e Milani, 2007). Entretanto, atualizações contínuas no conhecimento da palinoflora e da distribuição estratigráfica de seus táxons constituintes permitiram aprimoramentos nas unidades bioestratigráficas da bacia, tornandoas mais operacionais. Desta forma, a partir de trabalhos de detalhe, Souza e Marques-Toigo (2003, 2005) e Souza (2006) apresentaram esquemas baseados em zonas de intervalo, caracterizados por limites bem definidos de aparecimento e desaparecimento de táxons selecionados.
De acordo com Souza e Marques-Toigo (2003, 2005) e Souza (2006), a sucessão neopaleozóica da Bacia do Paraná compreende por quatro zonas de intervalo, nomeadas como: zonas Ahrensisporites cristatus (ZAc), Crucisaccites monoletus (ZCm), Vittatina costabilis (ZVc) e Lueckisporites virkkiae (ZLv). As ZAc e ZCm caracterizam o intervalo Pensilvaniano da bacia, sendo relativas às porções inferior e média do Subgrupo Itararé, registradas na porção nordeste da Bacia. Nos estados do Rio Grande do Sul e parte de Santa Catarina, somente as duas últimas unidades são reconhecidas, podendo ser identificadas a partir do topo do Subgrupo Itararé e estendidas às unidades mais superiores do Grupo Passa Dois.
A partir do topo do Subgrupo Itararé podem ser verificados os níveis basais de ocorrência da ZVc, a qual estendese até o topo da Formação Rio Bonito. Esta zona é dividida em duas unidades, Subzona Protohaploxypinus goraiensis, a qual é reconhecida também nas camadas de carvão, e na Subzona Hamiapollenites karrooensis, definida principalmente pela restrição da espécie que nomeia a unidade e pelo primeiro aparecimento das espécies Striatopodocarpites fusus e Staurosaccites cordubensis. A ZLv ocorre estratigraficamente acima da ZVc, sendo relacionada ao topo da Formação Rio Bonito, estendendose até a base da Formação Rio do Rasto (Membro Serrinha), conforme dados recentes de Neregato (2007) e Neregato et al. (2008), os quais registraram assembléias relativamente diversificadas nos depósitos desta última unidade, em níveis anteriormente considerados estéreis.
Materiais e métodos
As amostras analisadas foram coletadas em um afloramento situado às margens da Rodovia BR 293, na altura do km 152, entre os municípios de Bagé e Candiota, Estado do Rio Grande do Sul, região sul do Brasil (figura 2). O afloramento é constituído por camadas de carvão e níveis de siltitos carbonosos relacionadas à Formação Rio Bonito, com um nível de tonstein intercalado. Níveis basais da Formação Palermo ocorrem em contato erosivo com a unidade subjacente, os quais também foram analisados. Oito amostras foram selecionadas para análise palinológica, nomeadas, em ordem estratigráfica ascendente: C1, C2, C3, C4, C5, C6 (correspondentes à Formação Rio Bonito), C7 e C8 (Formação Palermo). O perfil litoestratigráfico com a indicação dos níveis analisados é apresentado na figura 3.
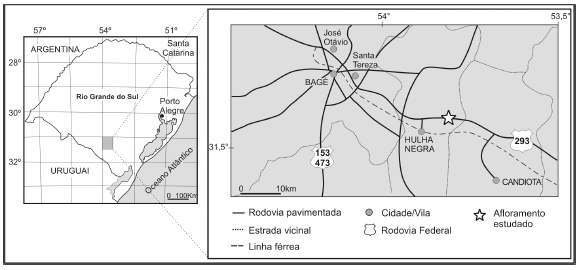
Figura 2. Localização do afloramento estudado / location map of the studied outcrop.
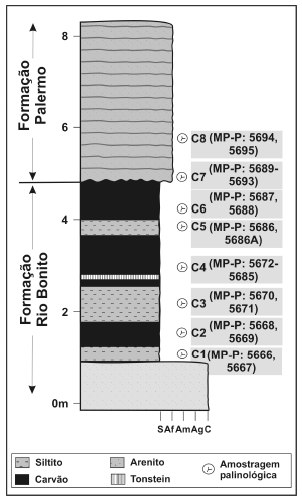
Figura 3. Perfil litoestratigráfico do afloramento, com indicação dos níveis amostrados e lâminas palinológicas correspondentes (MP-P: Museu de Paleontologia - Palinologia) / litostratigraphic section of the outcrop, with indication of sampled levels and their respective palynological slides.
As amostras foram processadas no Laboratório de Palinologia "Marleni Marques-Toigo" do Instituto de Geociências da Universidade Federal do Rio Grande do Sul, seguindo o método padrão para amostras paleozóicas (Quadros e Melo, 1987). Após a desagregação física do material, foram adicionados ácidos (HCl e HF) para dissolução dos minerais carbonáticos e silicáticos, permitindo a recuperação de matéria orgânica incluída na matriz mineral. Após sucessivas lavagens para neutralização do material, obteve-se um resíduo a partir de peneiramento entre as frações de 20 a 250 micrometros. Parte do resíduo foi tratado com solução de KOH 10% para remoção de matéria orgânica amorfa, de modo a facilitar a identificação taxonômica. Um total de 29 lâminas foi confeccionado para estudo, depositadas na Palinoteca do Museu de Paleontologia do Departamento de Paleontologia e Estratigrafia do Instituto de Geociências da Universidade Federal do Rio Grande do Sul, sob os códigos MP-P: 5666 a 5695.
As análises palinológicas constituíramse na identificação taxonômica e caracterização quantitativa do material sob microscopia óptica em aumentos de 200 a 1000 vezes, com contagem mínima de 200 espécimes por nível estratigráfico. Para a interpretação bio-estratigráfica, foram selecionadas espécies incluídas nas biozonas propostas a partir de Souza e Marques-Toigo (2003, 2005) e refinadas em Souza (2006).
Resultados
Composição dos conjuntos palinológicos
As amostras analisadas apresentaram conjuntos palinológicos relativamente abundantes, diversificados e bem preservados. Um total de 76 esporomorfos foi identificado, dentre os quais 42 espécies são relativas a esporos, 24 grãos de pólen, enquanto 10 formas são relacionadas a elementos constituintes do microplâncton. Todos os táxons registrados estão listados a seguir. Palinomorfos considerados como comuns nas associações assim como aqueles diagnosticados como espécies índices foram preferencialmente ilustrados. Nas tabelas 1 e 2 são apresentadas as abundâncias relativas e as distribuições de cada espécie por nível estratigráfico na seção estudada.
Tabela 1. Quadro de distribuição estratigráfica e abundância relativa de esporos identificados na seção analisada/ vertical stratigraphic distribution and relative abundance of the recovered spores species. As freqüências são interpretadas de acordo com a legenda: x - raro (freqüência< 2%) ; xx - comum (freqüência entre 2-5%); xxx - abundante (freqüência >5%) / frequences are interpreted as: x - rare (frequence< 2%) ; xx - commom (frequence between 2-5%); xxx - abundant (frequence >5%). Na coluna "figura", os números são correspondentes às ilustrações das espécies/ on "figura", numbers are related to ilustration of species.

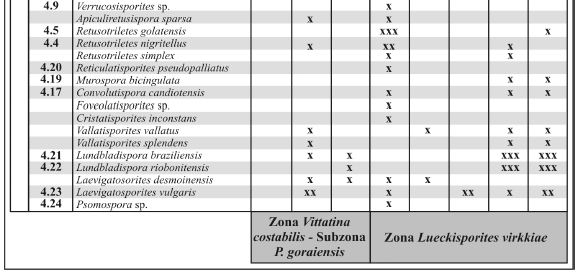
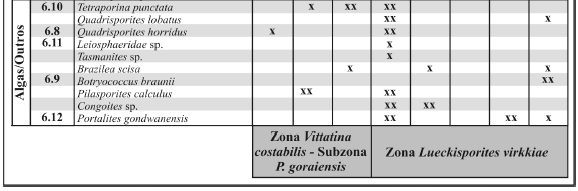
Tabela 2. Quadro de distribuição estratigráfica e abundância relativa dos grãos de pólen, algas e fungos identificados na seção analisada / vertical stratigraphic distribution and relative abundance of the recovered pollen grains, algae and fungi species. As freqüências são interpretadas de acordo com a legenda: x - raro (freqüência < 2%) ; xx - comum (freqüência entre 2-5%); xxx - abundante (freqüência >5%) / frequences are interpreted as: x - rare (frequence < 2%) ; xx - commom (frequence between 2-5%); xxx - abundant (frequence >5%). Na coluna"figura", os números são correspondentes às ilustrações das espécies / on "figura", numbers are related to ilustration of species.

Esporos triletes acavados
Apiculatisporis levis Balme e Hennelly 1956
Brevitriletes cornutus (Balme e Hennelly) Backhouse 1991 (figura 4.11)
Brevitriletes irregularis (Nahuys, Alpern e Ybert)
Césari, Archangelsky e Seoane 1995 (figura 4.10)
Calamospora exigua Staplin 1960
Calamospora inderjensis (Malayavkina ex Luber e Valts) Hart 1965
Calamospora liquida Kosanke 1950
Calamospora plicata (Luber e Valts) Hart 1965
Calamospora sinuosa Leschik 1955 (figura 4.1)
Converrucosisporites micronodosus (Balme e Hennelly) Playford e Dino 2002
Converrucosispoeires confluens (Archangelsky e Gamerro) Playford e Dino 2002 (figura 4.8)
Convolutispora candiotensis Ybert 1975 (figura 4.17)
Cyclogranisporites gondwanensis Bharadwaj e Salujha 1964 (figura 4.6)
Cyclogranisporites parvigranulosus (Leschik) Ybert 1975
Foveosporites sp. (figura 4.16)
Foveolatisporites sp.
Granulatisporites austroamericanus Archangelsky e Gamerro 1979 (figura 4.7)
Horriditriletes curvibaculosus Bharadwaj e Salujha 1964 (figura 4.13)
Horriditriletes gondwanensis (Tiwari e Moiz) Foster 1975
Horriditriletes pathakheraensis Anand-Prakash 1970 (figura 4.14)
Horriditriletes ramosus (Balme e Hennelly) Bharadwaj e Salujha 1964
Horriditriletes superbus (Foster) Césari, Archangelsky e Seoane 1995
Horriditrieletes uuguaiensis (Marques-Toigo) Archangelsky e Gamerro 1979 (figura 4.15)
Leiotriletes virkkii Tiwari 1965 (figura 4.2)
Lophotriletes pseudoaculeatus Potonié e Kremp 1955 (figura 4.12)
Murospora bicingulata Ybert 1975 (figura 4.19)
Psomospora sp. (figura 4.24)
Punctatisporites gretensis Balme e Hennelly 1956 (figura 4.3)
Apiculatisporis levis Menendez e Azcuy 1979
Retusotriletes golatensis Staplin 1960 (figura 4.5)
Retusotriletes nigritellus (Luber) Foster 1979 (figura 4.4)
Retusotriletes simplex Naumova 1953
Verrucosisporites pseudoreticulatus forma minor Ybert 1975
Verrucosisporites sp. (figura 4.9)

Figura 4. Todas as figuras x1000 / all specimens x1000. (MP-P: Museu de Paleontologia - Palinologia). 1, Calamospora sinuosa Leschik 1955 (MP-P 5671, O43). 2, Leiotriletes virkkii Tiwari 1965 (MP-P 5677, G52-4). 3, Punctatisporites gretensis Balme e Hennelly 1956 (MPP 5668, M33-1). 4, Retusotriletes nigritellus (Luber) Foster 1979 (MP-P 5684, M53-4). 5, Retusotriletes golatensis Naumova 1953 (MP-P 5682, L47-3). 6, Cyclogranisporites gondwanensis Bharadwaj e Salujha 1964 (MP-P 5686, H31-1). 7, Granulatisporites austroamericanus Archangelsky e Gamerro 1979 (MP-P 5677, J54-3). 8, Converrucosisporites confluens (Archangelsky e Gamerro) Playford e Dino 2000 (MP-P 5674, S50). 9, Verrucosisporites sp. (MP-P 5688, Q28-4). 10, Brevitriletes irregularis (Nahuys, Alpern e Ybert) Césari, Archangelsky e Seoane 1995 (MP-P 5682, E51-1). 11, Brevitriletes cornutus (Balme e Hennelly) Backhouse 1991 (MP-P 5677, B55). 12, Lophotriletes pseudoaculetus Potonié e Kremp 1955 (MP-P 5688, E38). 13, Horriditriletes curvibaculosus Bharadwaj e Salujha 1964 (MP-P 5684, H63-1. 14, Horriditriletes pathakeraensis Anand-Prakash 1970 (MP-P 5684, O56). 15, Horriditriletes uruguaiensis (Marques-Toigo) Archangelsky e Gamerro 1979 (MP-P 5686A, K38-3). 16, Foveosporites sp. (MP-P 5676, S42). 17, Convolustispora candiotensis Ybert 1975 (MP-P 5692, H48-2). 18, Diatomonozonotriletes subbaculiferus (Nahuys, Alpern e Ybert) Césari, Archangelsky e Gamerro 1995 (MP-P 5692, K43-1). 19, Murospora bicingulata Ybert 1975 (MP-P 5695, F36-1). 20, Reticulatisporites pseudopalliatus Staplin 1960 (MP-P 5682, O47). 21, Lundbladispora braziliensis (Marques-Toigo e Pons) Marques-Toigo e Picarelli 1984 (MP-P 5692, C53). 22, Lundbladispora riobonitensis Marques-Toigo e Picarelli 1984 (MP-P 5693, Q56-1). 23, Laevigatosporites vulgaris Ibrahim 1933 (MP-P 5693, P42). 24, Psomospora sp. (MP-P 5676, E57-1). Escala gráfica: 10 μm/ scale bar: 10 μm.
Esporos triletes cavados cingulizonados
Cristatisporites inconstans Archangelsky e Gamerro 1979
Diatomonozonotriletes subbaculiferus (Nahuys, Alpern e Ybert) Césari, Archangelsky e Gamerro 1995 (figura 4.18)
Reticulatisporites pseudopalliatus Staplin 1960 (figura 4.20)
Vallatisporites splendens Staplin e Jansonius 1962
Vallatisporites vallatus Hacquebard 1957
Lundbladispora braziliensis (Marques-Toigo e Pons) Marques-Toigo e Picarelli 1984 (figura 4.21)
Lundbladispora riobonitensis Marques-Toigo e Picarelli 1984 (figura 4.22)
Esporos monoletes
Laevigatosporites desmoinensis (Wilson e Coe) Shopf, Wilson e Bentall 1944
Laevigatosporites vulgaris Ibrahim 1933 (figura 4.23)
Grãos de pólen monossacados
Caheniasaccites elongatus Bose e Kar 1966 (figura 5.1)
Potonieisporites lelei Maheshwari 1967 (figura 5.2)
Distriomonosaccites crucistriatus (Ybert) Césari, Archangelsky e Seoane 1995 (figura 5.3)
Striomonosaccites sp.cf.
Striomonosaccites ovatus Bharadwaj 1962 (figura 5.4)

Figura 5. Todas as figuras x1000 / all specimens x1000. (MP-P: Museu de Paleontologia - Palinologia). 1, Caheniasaccites elongatus Bose e Kar 1966 (MP-P 5695, H45-1). 2, Potonieisporites lelei Maheshwari 1967 (MP-P 5674, O48-2). 3, Striomonosaccites cf S. ovatus Bharadwaj 1962. (MP-P 5692, Q50-4). 4, Distriomonosaccites crucistriatus (Ybert) Césari, Archangeslky e Seoane (MP-P 5676, G54). 5, Illinites unicus (Kosanke) Jansonius e Hills 1976 (MP-P 5677, R59-3). 6, Protohaploxypinus goraiensis (Potonié e Lele) Hart 1964 (MP-P 5676, F43-2). 7, Protohaploxypinus limpidus (Balme e Hennelly) Balme e Playford 1967 (MP-P 5685, N49-3). 8, Protohaploxypinus perfectus (Nauomova) Samoilovich 1953 (MP-P 5682, P61-2). 9, Protohaploxypinus samoilovichii (Jansonius) Hart 1964 (MP-P 5676, X59-2). 10, Lunatisporites variesectus Archangeslky e Gamerro 1979 (MP-P 5692, F46-2). 11, Striatopodocarpites cancelattus (Balme e Hennelly) Hart 1963 (MP-P 5692, U40-2). 12, Striatopodocarpites fusus (Balme e Hennelly) Potonié 1958 (MP-P 5693, Q49). Escala gráfica: 10 μm / scale bar: 10 μm.
Grãos de pólen bissacados não teniados
Limitisporites rectus Leschik 1955
Grãos de pólen bissacados teniados
Hamiapollenites fusiformis Marques-Toigo 1974 (figura 6.1)
Illinites unicus (Kosanke) Jansonius e Hills 1976 (figura 5.5)
Lueckisporites virkkiae (Potonié e Klaus) Clarke 1965 (figura 6.3)
Lunatisporites variesectus Archangeslky e Gamerro 1979 (figura 5.10)
Protohaploxypinus goraiensis (Potonié e Lele) Hart 1964 (figura 5.6)
Protohaploxypinus limpidus (Balme e Hennelly) Balme e Playford 1967 (figura 5.7)
Protohaploxypinus perfectus (Nauomova) Samoilovich 1953 (figura 5.8)
Protohaphoxypinus samoilovichii (Jansonius) Hart 1964 (figura 5.9)
Striatoabieites multistriatus (Balme e Hennelly) Hart 1964 (figura 6.2)
Striatopodocarpites cancellatus (Balme e Hennelly) Hart 1963 (figura 5.11)
Striatopodocarpites fusus (Balme e Hennelly) Potonié 1958 (figura 5.12)

Figura 6. Todas as figuras x1000 / all specimens x1000. (MP-P: Museu de Paleontologia - Palinologia). 1, Hamiapollenites fusiformis Marques-Toigo 1974 (MP-P 5693, W65). 2, Striatoabieites multistriatus (Balme e Hennelly) Hart 1964 (MP-P 5692, L53). 3, Lueckisporites virkkiae (Potonié e Klaus) Clarke 1965 (MP-P 5692, R43-2). 4, Fusacolpites fusus Bose e Kar 1966 (MP-P 5695, E52-4). 5, Marsupipollenites cf. M. striatus (Balme e Hennelly) Foster 1975 (MP-P 5676, T53-3). 6, Vittatina costabilis Wilson 1962 (MP-P 5692, J47). 7, Weylandites lucifer (Bharadwaj e Salujha) Foster 1975 (MP-P 5694, H37-3). 8, Quadrisporites horridus (Hennelly) Potonie e Lele 1961 (MP-P 5676, Y43-1). 9, Brazilea scisa (Balme e Hennelly) Foster 1975 (MP-P 5695, X50-2). 10, Tetraporina punctata (Tiwari e Navale) Kar e Bose 1976 (MP-P 5676, G46-2). 11, Leiosphaeridia sp. (MP-P 5676, P43). 12, Portalites gondwanensis Nahuys, Alpern e Ybert 1968 (MP-P 5695, E52-3). Escala gráfica: 10 μm/ scale bar: 10 μm.
Grãos de pólen praeocolpados
Pakhapites fusus Bose e Kar 1966 Menéndez 1971 (figura 6.4)
Pakhapites ovatus (Bose e Kar) Playford e Dino 2000
Marsupipollenites sp. cf. Marsupipollenites striatus (Balme e Hennelly) Foster 1975 (figura 6.5)
Grãos de pólen monocolpados
Cycadopites sp.
Grãos de pólen poliplicados
Vittatina costabilis Wilson 1962 (figura 6.6)
Vittatina saccata (Hart) Playford e Dino 2000
Vittatina subsaccata Samoilovich 1953
Weylandites lucifer (Bharadwaj e Salujha) Foster 1975 (figura 6.7)
Algas clorofíceas
Brazilea scisa (Balme e Hennelly) Foster 1975 (figura 6.9)
Botryococcus braunii Kutzing 1849
Congoites sp.
Leiosphaeridae sp. (figura 6.11)
Pilasporites calculus (Balme e Hennelly) Tiwari e Navale 1967
Quadrisporites lobatus (Tiwari e Navale) Ybert 1975
Quadrisporites horridus (Hennelly) Potonie e Lele 1961 (figura 6.8)
Tasmanites sp.
Tetraporina punctata (Tiwari e Navale) Kar e Bose 1976 (figura 6.10)
Fungos
Portalites gondwanensis Nahuys, Alpern e Ybert 1968 (figura 6.12)
Características gerais dos conjuntos palinológicos
Diferenças quantitativas são observadas nos conjuntos palinológicos registrados para as duas unidades, com maior abundância de esporos triletes nas amostras da Formação Rio Bonito. Embora as amostras da Formação Palermo apresentem conteúdo semelhante, diferenciamse por maior freqüência relativa de grãos de pólen bissacados e teniados. De maneira geral, grãos de pólen monossacados são raros nas duas unidades, e freqüentemente apresentam mau estado de preservação, por vezes dificultando sua classificação em nível específico.
Esporos associados a diferentes grupos vegetais foram documentados, tais como representantes de Filicophyta (Apiculatisporites, Brevitriletes, Convolutispora, Converrucosisporites, Cyclogranisporites, Granulatisporites, Horriditriletes, Leiotriletes, Lophotriletes, Punctatisporites, Reticulatisporites, Retusotriletes, Verrucosisporites), Sphenophyta (Calamospora, Laevigatosporites e Lycophyta (Cristatisporites, Lundbladispora, Vallatisporites). Grãos de pólen relacionados às Gimnospermas, tais como Coniferophyta (Caheniasaccites, Hamiapollenites, Limitisporites, Lueckisporites, Lunatisporites, Potonieisporites, Striatoabieites, Staurosaccites), Glossopteridophyta (Illinites, Fusacolpites, Marsupipollenites, Protohaploxypinus, Striatopodocarpites, Vittatina, Weylandites) e Cycadales (Cycadopites) (Balme, 1995, também foram documentados).
Nas associações da Formação Rio Bonito predominam esporos dos gêneros Punctatisporites, Granulatisporites, Horritiditriletes, Lophotriletes, e de maneira subordinada, os gêneros Cyclogranisporites e Verrucosisporites. Grãos de pólen monossacados ocorrem em baixas proporções (Caheniasaccites, Potonieisporites), frequentemente em mau estado de preservação. Por outro lado, nas palinofloras da Formação Palermo dominam esporos do gênero Punctatisporites e formas cingulizonadas como Lundbladispora (incluindo tétrades), registrandose também aumento na freqüência de grãos de pólen, principalmente de teniados (Protohaploxypinus, Lunatisporites, Striatopodocarpites) e poliplicados (Vittatina, Weylandites). Por sua vez, em termos de diversidade, a amostra C4, a qual é incluída na Formação Rio Bonito, é constituída por uma palinoflora "transicional" entre duas unidades litoestratigráficas. Esta associação é caracterizada por uma grande variedade e abundância de esporos (e.g. Punctatisporites, Leiotriletes, Brevitriletes, Horriditriletes, Granulatisporites, Cyclogranisporites, Retusotriletes, Murospora, Laevigatosporites), grãos de pólen monossacados e estriados (e.g. Caheniasaccites, Protohaploxypinus, Hamiapollenites, Striatoabieites), grãos de pólen poliplicados (Vittatina, Weylandites) e elementos relativos a algas e fungos (Tetraporina, Quadrisporites, Brazilea, Portalites).
Formas associadas a algas zignematáceas, entre elas Brazilea, Tetraporina e Quadrisporites, são frequentemente documentadas nas associações da Formação Rio Bonito. Esses táxons também ocorrem na Formação Palermo, porém em baixas proporções. Além disso, Portalites gondwanensis, considerado por alguns autores como fungo (Elsik, 1996), foi identificado em ambas unidades.
Posicionamento bioestratigráfico
O posicionamento bioestratigráfico apresentado a seguir é realizado com base na ocorrência dos táxons por nível estratigráfico, considerando o zoneamento vigente para o intervalo na bacia (Souza e Marques-Toigo, 2003, 2005; Souza, 2006). Maiores considerações sobre esse tema fazem parte das discussões, tendo em vista a detecção de que os conjuntos palinológicos não se enquadram peritamente nas zonas disponíveis.
A presença de G. austroamericanus e V. subsaccata indicam o posicionamento das amostras dos níveis C1, C2 e C3 na ZVc, provavelmente Subzona Protohaploxypinus goraiensis, o que também é confirmado pela ausência de táxons diagnósticos das unidades suprajacentes. Na amostra do nível C4 ocorrem G. austroamericanus, C. confluens, I. unicus, H. fusiformis, P. goraiensis, V. costabilis e V. subsaccata, táxons diagnósticos da Subzona P. goraiensis. Contudo, são registrados associados L. virkkiae e W. lucifer, os quais têm aparecimento a partir da Zona L. virkkiae. Tendo em vista que o zoneamento vigente é baseado em limites de aparecimento e desaparecimento, o nível C4 deveria ser posicionado nesta última zona, o que resulta em problemas do ponto de vista das caracterizações das zonas, conforme será discutido mais adiante. O conteúdo das amostras C5 e C6, ambas da Formação Rio Bonito, é constituído por esporos e grãos de pólen caracterizados por amplo alcance estratigráfico, dificultando qualquer determinação de posicionamento bioestratigráfico particular. Dessa forma, o posicionamento do intervalo C5-C6 é realizado com base nas amostras supra e subjacentes.
As amostras C7 e C8, da Formação Palermo, possuem táxons diagnósticos da Zona L. virkkiae, tais como S. fusus, S. cancellatus, L. variesectus, S. multistriatus, W. lucifer e L. virkkiae. Contudo, tal como ocorre na amostra C4, há presença de táxons estratigraficamente restritos a outras zonas, como é o caso de G. austroamericanus, C. confluens e H. fusiformis, considerados restringidos à Subzona P. goraiensis.
Dessa forma, considerando o exposto e tendo como base o zoneamento vigente, duas zonas ocorrem nas amostras estudadas: Subzona P. goraiensis, base da Zona V. costabilis, identificada nas amostras C1 a C3, da Formação Rio Bonito; Zona Lueckisporites virkkiae, amostras C4 a C8, abrangendo desde o topo da Formação Rio Bonito no afloramento até a base da Formação Palermo. A distribuição estratigráfica de espécies selecionadas, assim como o posicionamento bioestratigráfico dos níveis amostrados é sumarizado na figura 7. No entanto, as distribuições estratigráficas registradas nesse trabalho não modificam, a priori, as amplitudes dos táxons previamente descritos em cada zona, embora sugiram aprimoramentos futuros.
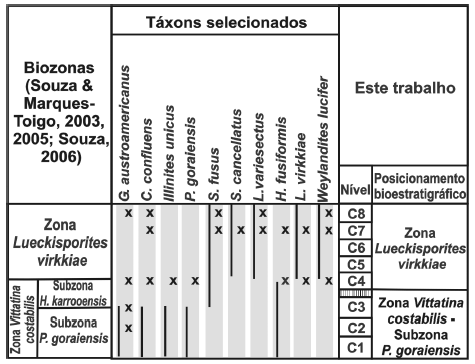
Figura 7. Quadro síntese ilustrando a distribuição estratigráfica de espécies selecionadas, correlacionando os níveis amostrados e biozonas identificadas neste trabalho / synthesis of the stratigraphic distribution of selected species, with indication of sampled levels and identified biozones in this paper. As linhas contínuas indicam a distribuição estratigráfica de espécies selecionadas conforme o esquema de Souza & Marques-Toigo (2003, 2005) e Souza (2006); marcações em X referem-se à distribuição das espécies neste trabalho / full lines are relative to stratigraphic distribution of selected species stablished by Souza & Marques-Toigo (2003, 2005) and Souza (2006); the X marks the stratigraphic distribution of species registered in this contribution.
Discussão dos resultados
Análise palinoestratigráfica
Como relatado anteriormente, alguns problemas com relação à distribuição de determinados táxons foram verificados, podendo implicar em diferentes interpretações sobre o posicionamento bioestratigráfico da seção analisada, bem como em indicações de mudanças no zoneamento vigente.
O reconhecimento da ZLv na seção analisada é baseado no primeiro registro estratigráfico de L. virkkiae e W. lucifer (Souza e Marques-Toigo, 2005), no nível C4. Embora raros nesta amostra, as espécies definem o limite inferior da citada zona, que foi erigida com base em datum de primeiro aparecimento de táxons selecionados. A ZLv é correlata aos intervalos K e L de Daemon e Quadros (1970). Segundo esses autores, grande parte da Formação Rio Bonito foi depositada durante o intervalo K, constatação observada principalmente nos poços mais a norte da área estudada (no Paraná, São Paulo e Mato Grosso do Sul). Geralmente, os níveis estudados da unidade no Estado do Rio Grande do Sul, correspondem aos intervalos mais inferiores (I a J), equivalentes à ZVc, onde está posicionada a maioria dos níveis de carvão, ou seja, a maior parte das camadas expostas na região de Candiota deve corresponder à ZVc, uma vez que nenhum dos táxons da ZLv havia sido detectado associados aos carvões, diferentemente do aqui observado.
Por outro lado, espécies anteriormente conhecidas como confinadas à ZVc, tais como C. confluens e G. austroamericanus e H. fusiformis foram detectadas na zona suprajacente (ZLv), e ainda que representadas por poucas espécies, denotam maior alcance estratigráfico das mesmas. Deve ser destacado que a Subzona Hamiapollenites karrooensis não foi reconhecida. Dentre as três espécies que definem o limite basal da unidade (Hamiapollenites karooensis, Staurosaccites cordubensis, Striatopodocarpites fusus) somente S. fusus foi registrada. Contudo, tratase de uma espécie com alcance estendido até à ZLv.
A julgar de que se trata dos níveis mais superiores da Formação Rio Bonito, sujeitos à fases contínuas de erosão, como decorrência de eventos transgressivos, até o estabelecimento das fácies plataformais que constituem a Formação Palermo (vide Holz, 2003; Holz et al., 2006), supõese que a ZVc não esteja totalmente representada na seção analisada.
Baseados na análise de dados de superfície e subsuperfície da área, Holz et al. (2006) registraram três seqüências deposicionais, dentre as quais, a segunda (S2) e a terceira (S3) são correlatas, respectivamente, aos depósitos das formações Rio Bonito e Palermo, e cujo limite de seqüências foi nomeado como SB3. Segundo esses autores, durante a geração do SB3, reconhecido na seção analisada (limite entre as duas unidades litoestratigráfica), a bacia teria sofrido movimentos tectônicos localizados, o que teria influenciado na modificação da linha de costa e resultado em subsidência diferencial na região de Candiota. Durante a transgressão marinha, parte dos depósitos de carvão do topo da Formação Rio Bonito teria sido retrabalhada e erodida, e posteriormente sobreposta por depósitos de shoreface da Formação Palermo. Esta interpretação estratigráfica poderia justificar a ausência de elementos marcantes da subzona Hamiapollenites karrooensis.
A constatação de esporos de Lundbladispora, por vezes em tétrades, associados a outros elementos higrófilos-hidrófilos (Punctatisporites, Leiotriletes) nos níveis basais da Formação Palermo indica a permanência da vegetação formadora dos carvões ainda no tempo Palermo e, conseqüentemente, de condições ambientais não tão distintas do que aquelas reinantes durante a deposição da Formação Rio Bonito. Entretanto, os dados relativos à macroflora da base da Formação Palermo são escassos, representados apenas por restos de troncos silicificados identificados como Dadoxylon sp. (Schneider, 1974), o que não permite maiores considerações sobre a manutenção da paleoflora correspondente à base da zona L. virkkiae.
Por outro lado, uma vez que espécies consideradas restritas à Subzona P. goraiensis, tais como Granulatisporites austroamericanus, Converrucosis-porites confluens, Illinites unicus foram reconhecidas associadas à zona suprajacente, podem ser sugeridas modificações no zoneamento atual, em termos de ampliação no alcance estratigráfico destes táxons. A julgar somente pelo estado de preservação, não há indícios de retrabalhamento a partir de depósitos mais antigos.
Comparações com outras assembléias
A comparação das associações apresentadas nesta contribuição é realizada com base no conteúdo qualitativo, ou seja, na listagem de espécies, uma vez que diferenças composicionais são esperadas, resultado das variações paleoecológicas intrínsecas aos contextos paleoambientais de cada unidade estratigráfica e de cada amostra.
As associações palinológicas documentadas neste trabalho exibem conteúdo distinto daqueles publicados para a região de Candiota por outros autores (e.g., Ybert, 1975; Meyer e Marques-Toigo, 2000; Cazzulo-Klepzig et al., 2005; Guerra-Sommer et al., 2008a, 2008b). Os resultados bioestratigráficos apresentados aqui sugerem associações palinológicas mais jovens do que as anteriormente descritas, as quais haviam posicionado parte dos níveis da Mina de Candiota na Subzona Protohaploxypinus goraiensis (Guerra-Sommer et al., 2008b). Esta datação mais "jovem" pode também resultar do fato de que a seção analisada neste trabalho encontrase numa posição estratigráfica ligeiramente acima daqueles aflorantes na Mina de Candiota.
Guerra-Sommer et al. (2008a) discutiram a validade da Subzona Protohaploxypinus goraiensis, considerada estratigraficamente muito extensa, incluindo diferentes contextos paleoambientais, em um intervalo de tempo relativamente muito longo (aproximadamente 15 Ma). No entanto, biozonas propostas para o mesmo intervalo de tempo em outras partes do Gondwana são semelhantes em seus conjuntos palinológicos e distribuição estratigráfica, abrangendo espessos pacotes sedimentares (Césari, 2007).
Quando comparadas com palinozonas documentadas na porção oeste do Gondwana, as ZVc e ZLv são correlacionáveis às zonas Fusacolpites-fusus- Vittatina subsaccata (FS) e Lueckisporites-Weylandites (LW), erigidas por Césari e Gutiérrez (2001) para depósitos da porção centro-oeste da Argentina. Estas biozonas apresentam como características gerais de seus conjuntos palinológicos o primeiro aparecimento de espécies de Vittatina e abundância de grãos de pólen estriados. Com relação à biozonas erigidas na Bacia do Chacoparaná, as ZVc e ZLv encontram, respectivamente, correspondência nas zonas Cristatisporites e Striatites (Russo et al., 1980; Vergel, 1993), as quais são caracterizadas pelo primeiro aparecimento de Converrucosisporites micronodosus e C. confluens (Zona Cristatisporites) e pelo predomínio de grãos de pólen estriados na zona Stritatites. Por fim, de acordo com os dados de Béri et al. (2004) obtidos na porção nordeste do Uruguai em depósitos também pertencentes à Bacia do Chacoparaná, as ZVc e ZLV podem ser correlacionáveis às zonas Cristatisporites inconstans-Vittatina subsaccata e Striatitoabietites anaverrucosus-Staurosaccites corbubensis, definidas, respectivamente, pelo predomínio de esporos triletes associados a grãos de pólen bissacados, e pela abundância de grãos de pólen estriados e poliplicados.
Conjuntos palinológicos semelhantes aos constituintes das ZVc e ZLv são documentados em outras regiões do Gondwana, tal como África (Falcon, 1975), Antártica (Lindoström, 1995), Austrália (Jones e Truswell, 1992). Entretanto, diferenças de distribuição fitogeográfica existentes entre as regiões podem refletir numa ampla variação na amplitude vertical de um mesmo táxon, o que por sua vez, limita uma correlação mais precisa entre estas biozonas (vide Souza e Marques-Toigo, 2005).
Biocronoestratigrafia
Embora a quantidade de datações absolutas tenha aumentado significativamente na última década (vide Césari, 2007; Rocha-Campos et al., 2007; Guerra-Sommer et al., 2008a, 2008b), o tema ainda é muito controverso, em razão das diferenças entre os métodos e dos resultados que muitas vezes ainda são conflitantes.
Para a Mina de Candiota, diversas datações radiométricas foram obtidas, tal como Matos et al. (2001), Guerra-Sommer et al. (2005, 2008a, 2008b), Rocha-Campos et al. (2006, 2007) ou em camadas correlacionáveis a esta (Marques et al., 2007). Estes dados radiométricos apresentam idades que variam entre 267.1 ± 3.4 Ma (Matos et al., 2001) a 299.1 ± 2,6 Ma (Guerra-Sommer et al., 2005), o que denota a necessidade de estudos mais detalhados nesse tema. Além disso, devese ressaltar que tais trabalhos frequentemente exibem escassas informações palinológicas, limitando o posicionamento e a análise bioestratigráfica adequada das unidades em questão.
O nível de tonstein aflorante na seção, localizado entre os níveis C3 e C4, foi amostrado para datação através de U-Pb em zircões, método LA-ICP-MS. Análises iniciais do material reveleram idade aproximada de 278.2 ± 1.8 M.a. (vide Marques et al, 2007). Entretanto, tratam-se de resultados preliminares, os quais serão apresentados em trabalho à parte, como continuidade desta publicação, tendo em vista que fornece uma nova abordagem geocronológica, balizada pelos dados palinoestratigráficos aqui apresentados
Considerações finais
O reconhecimento e a análise de conjuntos de esporomorfos relativamente diversificados e abundantes em oito níveis distintos da seção analisada, entre as formações Rio Bonito e Palermo, permitiram a obtenção dos seguintes resultados conclusivos e das considerações finais abaixo listadas.
Diferenças tanto quantitativas quanto qualitativas foram verificadas entre cada um dos níveis analisados, com implicações de caráter bioestratigráfico. Duas unidades palinoestratigráficas foram observadas na seção, quais sejam, a Zona Vittatina costabilis (níveis C1 a C3) e a Zona Lueckisporites virkkiae (C4 a C8), sendo a primeira ocorrente na Formação Rio Bonito, enquanto a segunda ocorre desde o topo da Formação Rio Bonito na seção até a base da Formação Palermo.
Algumas espécies conhecidas como confinadas à Zona Vittatina costabilis, tais como C. confluens, G. autroamericanus e H. fusiformis, foram detectadas em níveis mais superiores, da Zona Lueckisporites virkkiae. Algumas hipóteses podem ser levantadas para explicar tal incongruência: (i) possibilidade de extensão da amplitude vertical dessas espécies, para níveis mais superiores; (ii) espécies que delimitam a base da Zona Lueckisporites virkkiae poderiam ter registro mais baixo; (iii) esse comportamento pode significar uma "transição" entre as duas unidades palinoestratigráficas, não detectada em outras localidades, como reflexo do estabelecimento das fácies costeiras da Formação Palermo, que obstruem o registro da unidade imediatamente abaixo (Formação Rio Bonito).
A problemática aqui levantada é advinda do estudo detalhado de oito amostras de uma seção aflorante de cerca de 8 m de espessura. Um menor número de amostras poderia "mascarar" alguns desses aspectos, pelo qual fica explícita a necessidade de estudos mais detalhados em outras localidades da bacia. Por outro lado, o apoio de dados de subsuperfície é fundamental para a resolução mais definitiva da problemática bioestratigráfica. Dessa forma, poços perfurados no entorno da área já foram selecionados e estão em estudo a fim de ampliar os dados e complementar a discussão, fornecendo maiores subsídios para uma melhor caracterização de cada uma das palinozonas citadas.
Uma idade absoluta foi obtida para o nível de tonstein ocorrente entre os níveis carbonosos da Formação Rio Bonito no afloramento, sendo objeto de publicação a parte pelos autores, na qual os aspectos biocronoestratigráficos envolvidos serão melhor discutidos. Considerando o conhecimento atual sobre o tema e as observações já apontadas em Santos et al. (2006), os níveis estudados são posicionados no Cisuraliano (Artinskiano), correspondente ao Permiano Inferior.
Agradecimentos
Esta contribuição foi realizada a partir de recursos advindos do Conselho Nacional de Desenvolvimento Científico (CNPq) Projeto 474153/2004-5. Os autores prestam agradecimentos à Companhia de Recursos Minerais (CPRM), através do trabalho de campo com R.C. Lopes e relevantes discussões com J.C. Marques. Agradecimentos também são feitos ao CNPq pela concessão de bolsa de doutorado (primeira autora) e produção científica dos dois autores. Este trabalho constitui parte da Tese do Dautoramento da primeira autora junto ao Programa de Pos Graduação em Geocências da UFRGS.
Referências
1. Alves, R.G. e Ade, M.V.B. 1996. Sequence stratigraphy and coal petrography applied to the Candiota Coal Filed, Rio Grande do Sul, Brazil: a depositional model. International Journal of Coal Geology 30: 231-248. [ Links ]
2. Balme, B.E. 1995. Fossil in situ spores and pollen grains: an annotated catalogue. Review of Palaeobotany and Palynology 87: 81-323. [ Links ]
3. Béri, A., Gutiérrez, P.R., Cernuschi, F. e Balarino, L. 2004. Palinoestratigrafia del Pérmico de la Cuenca Paraná, Uruguay. 11º Reunião de Paleobotânicos e Palinológos (Gramado), Boletim de Resumos, p. 29. [ Links ]
4. Beurlen, K. 1953. Análise paleogeográfica da Série Tubarão e possibilidade de novas bacias carboníferas. Notas Preliminares e Estudos (DNPM) 74: 1-10. [ Links ]
5. Bharadwaj, D.C., Kar, R.K. e Navale, G.K.B. 1976. Palynostratigraphy of Lower Gondwana deposits in Paraná and Maranhão Basins, Brazil. Biological Memoirs 1: 56-103. [ Links ]
6. Carruthers, W. 1869. On the plant remains from the brazilian coal beds with remarks on the genus Flemingites. Geological Magazine 6: 151-155. [ Links ]
7. Cauduro, A.D. 1970. Lower Gondwana miospores from São Sepé outcrop (Rio Grande do Sul, Brasil). Escola de Geologia da UFRGS, Publicação Especial 17, 34 p. [ Links ]
8. Cazzulo-Klepzig, M., Guerra-Sommer, M., Formoso, N.L. e Calarge, L.M. 2002. Geochemical and palynological evidence for the age determination of Permian coals, southern Brazil. Journal of South American Earth Sciences 25: 375-380. [ Links ]
9. Cazzulo-Klepzig, M., Menegat, R. e Guerra-Sommer, M. 2005. Palynology and paleobotany in the reconstruction of landscape units from the Candiota Coalfield, Permian of Paraná Basin, Brazil. Revista Brasileira de Paleontologia 8: 83-98. [ Links ]
10. Césari, S.N. 2007. Palynological biozones and radiometric data at the Carboniferous-Permian boundary in western Gondwana. Gondwana Research 11: 529-536. [ Links ]
11. Césari, S.N. e Gutiérrez, P.R., 2001. Palynostratigraphy of Upper Paleozoic sequences in central-western Argentina. Palynology 24: 113-146. [ Links ]
12. Corrêa da Silva, Z. e Marques-Toigo, M. 1975. Carvão no Brasil: Mina de Candiota - Rio Grande do Sul. 2º Congresso IberoAmericano de Geologia Econômica (Buenos Aires), 1: 263-286. [ Links ]
13. Corrêa da Silva, Z. e Marques-Toigo, M. 1985. Considerações petrológicas e palinológicas sobre a Camada Candiota, Jazida Carbonífera de Candiota, Rio Grande do Sul. 2º Simpósio Sul-Brasileiro de Geologia (Florianópolis), Anais 432- 447. [ Links ]
14. Daemon, R.F. e Quadros, L.P. 1970. Bioestratigrafia no Neopaleozóico da Bacia do Paraná. 24º Congresso Brasileiro de Geologia (Brasília), Anais 359-412. [ Links ]
15. Dias-Fabrício, M.E. 1981. Palinologia da Formação Rio Bonito na área de Gravataí-Morungava, Rio Grande do Sul. Pesquisas 14: 69-130. [ Links ]
16. Dijkstra, S.J. 1955. Some brazilian megaspores, Lower Permian in age and their comparison with lower gondwana spores from India. Medelingen van de Geologishe Stichting 10: 5-10. [ Links ]
17. Dolianiti, E. 1953. A flora Gondwana Inferior em Santa Catarina. I. Notas Preliminares e Estudos (DNPM) 60: 1-7. [ Links ]
18. Elsik, W.C. 1996. Fungi. En: J. Jansonius e D.C. McGregor (eds.), Palynology: Principles and Applications, Vol. 1. American Association of Stratigraphic Palynologists Foundation, Dallas, pp. 293-305. [ Links ]
19. Falcon, R.M.S. 1975. Palyno-stratigraphy of the Lower Karoo sequence in the central Sebungwe District, Mid-Zambezi Basin, Rhodesia. Paleontologia Africana 18: 1-29. [ Links ]
20. Fontes, L.C. e Cava, L. T. 1980. Análise ambiental preliminar do Gonduana na região de Candiota, Rio Grande do Sul. 31º Congresso Brasileiro de Geologia (Balneário Camboriú), Anais 2: 755-765. [ Links ]
21. Guerra-Sommer, M., Cazzulo-Klepzig, M., Formozo, M. L., Menegat, R. e Basei, M.A.S. 2005. New radiometric data from ash fall rocks in Candiota coal-bearing strata and the palynostratigraphic framework in southern Paraná Basin (Brazil). Gondwana 12 (Mendoza), Abstracts p. 89. [ Links ]
22. Guerra-Sommer, M., Cazzulo-Klepzig, M., Santos, J.O.S., Hartman, L.A., Ketzer, J.M. e Formoso, M.L.L. 2008a. Radiometric age determination of tonstein and stratigraphic constraints for the Lower Permian coal sucession in southern Paraná Basin, Brazil. International Journal of Coal Geology 74: 13-27. [ Links ]
23. Guerra-Sommer, M., Cazzulo-Klepzig, M., Formoso, M.L.L., Menegat, R. e Mendonça, J.G. 2008b. U-Pb dating of tonstein layers from a coal sucession of the soutrhen Parana Basin (Brazil): A new geochonological aproach. Gondwana Research 14: 474-482. [ Links ]
24. Hartt, C.F. 1870. Geology and physical geography of Brazil. Boston, 620pp. [ Links ]
25. Holz, M. 1998. The Eo-Permian coal seams of the Paraná basin in southernmost Brazil: An anlysis of the depositional conditions using sequence stratigraphy concepts. Internatinal Journal of Coal Geology 36: 141-163. [ Links ]
26. Holz, M. 2003. Sequence stratigraphy of a lagoonal estuarine system-an example from the lower Permian Rio Bonito Formation, Paraná Basin, Brazil. Sedimentary Geology 162: 305-331. [ Links ]
27. Holz, M., Küchle, J., Philipp, R.P, Bischoff, A.P. e Arima, N. 2006. Hierarchy of tectonic control and stratigraphic signatures: Base-level changes during the Early Permian in the Paraná Basin, soutrhernmost Brazil. Journal of South American Earth Sciences 22: 185-204. [ Links ]
28. Jones, M.J. e Truswell, E.M. 1992. Late Carboniferous and Early Permian palynostratigraphy of the Joe Joe Group, southern Galilee Basin, Queensland, and implications for Gondwanan stratigraphy. Journal of Australian Geology and Geophysics 13: 143-185. [ Links ]
29. Lavina, E.L. e Lopes, R.C. 1987. A transgressão marinha do Permiano Inferior e a evolução paleogegráfica do Supergrupo Tubarão no Estado do Rio Grande do Sul. Paula-Coutiana 1: 51-103. [ Links ]
30. Liais, E. 1872. Climats, géologie, faune et geographie du Brésil. Paris, 640 pp. [ Links ]
31. Lindström, S. 1995. Early Permian palynostratigraphy of the northern Heimefrontfjella mountain-range, Dronning Maud Land, Antarctica. Review of Palaeobotany and Palynology 89: 359-415. [ Links ]
32. Lopes, R.C., Lavina, E.L. e Signorelli, N. 1986. Fácies sedimentares e evolução paleoambiental do Supergrupo Tubarão na borda leste da Bacia do Paraná. Uma seção regional nos Estados do Rio Grande do Sul e Santa Catarina. 34º Congresso Brasileiro de Geologia (Goiânia), Anais 1: 206-218. [ Links ]
33. Lundquist, G. 1920. Fossile pflanzen der Glossopteris Flora aus Brasilien. Kungl Svenska Vetenskaps Akademis Handlingear 60: 1-36. [ Links ]
34. Marques, J. C., Chemale, F. J., Lopes, R. C., Souza, P.A., Justino, D. e Dussin, I. A. 2007. LA-ICP-MS U-Pb Zircon age form an ash fall rock of the uppermost Rio Bonito Formation, southern Paraná Basin. 4º Simpósio sobre Cronoestratigrafia da Bacia do Paraná (Búzios), Boletim de Resumos p. 32. [ Links ]
35. Marques-Toigo, M., Corrêa da Silva, Z. e Herter, G.G. 1975. Geology and palynology of Candiota Coal Mine, RS, Brazil. 1º Congresso Argentino de Paleontologia y Bioestratigrafia (Tucuman), Actas 1: 401-427. [ Links ]
36. Matos, S.L.F., Yamamoto, J.K., Riccomini, C., Hachiro, J. e Tassinari, C.C.G. 2001. Absolute dating of Permian ash-fall in the Rio Bonito Formation, Paraná Basin, Brazil. Gondwana Research 4: 421-426. [ Links ]
37. Menezes-Filho, N.R. e Brito, P.C. 1982. Estratigrafia da seqüência Gonduânica na região de Candiota, RS. 32º Congresso Brasileiro de Geologia (Salvador), Anais 4: 1392-1405. [ Links ]
38. Meyer, K.E. e Marques-Toigo, M. 2000. O Siginificado paleoambiental da microflora dos carvões da Malha IV, Mina de Candiota, RS, Permiano da Bacia do Paraná, Brasil. Revista de Geociências 5: 17-20. [ Links ]
39. Milani, E.J. 1997. Evolucão tectono-estratigráfica da Bacia do Paraná e seu relacionamento com a geodinâmica fanerozóica do Gondwana sul-oriental. Programa de Pós-graduação em Geociências, Universidade Federal do Rio Grande do Sul, Tese de Doutorado, 255p. [ Links ]
40. Milani, E.J. e Zalán, P.V. 1999. An outline of the geology and petroleum systems of the Paleozoic interior basins of South America. Episodes 22: 199-205. [ Links ]
41. Nahuys, J., Alpern, B. e Ybert, J.P. 1968. Estudo palinológico e petrográfico de alguns carvões do sul do Brasil. Boletim Instituto Tecnológico do Rio Grande do Sul 46: 1-61. [ Links ]
42. Neregato, R. 2007. Estudo palinológico das Formações Serra Alta, Teresina e Rio do Rasto nos furos de sondagem SP-23-PR e SP-58-PR, centro-norte do Paraná (Permiano, Bacia do Paraná). Programa de Pos-Graduaçao em Geologia Regional, Universidade Estadual Paulista, Dissertação de Mestrado, 107p. [ Links ]
43. Neregato, R., Souza, P.A. e Rohn, R. 2008. Registros palinológicos inéditos nas formações Teresina e Rio do Rasto (Permiano, Grupo Passa Dois, Bacia do Paraná): implicações biocronoestratigráficas e paleoambientais. Pesquisas em Geociências 35: 9-21. [ Links ]
44. Pant, D.D. e Srivastava, G.K. 1965. Some Lower Gondwana miospores from Brazil. Micropaleontology 11: 468-478. [ Links ]
45. Picarelli, A.T., Dias-Fabrício, M.E. e Cazzulo-Klepzig, M. 1987. Considerações sobre a paleocologia e a palinologia da jazida carbonífera de Santa Terezinha, RS, Brasil - Permiano da Bacia do Paraná. 3º Simpósio Sul-Brasileiro de Geologia (Curitiba), Atas 1: 351-372. [ Links ]
46. Piccoli, A.E., Lopes, R.C. e Camozzato, E. 1986. Análise paleoambiental das jazidas de carvão do Estado do Rio Grande do Sul. Acta Geologiga Leopoldensia 22: 35-54. [ Links ]
47. Plant, N. 1869. The brazilian coal fields. Geological Magazine 6: 147-150. [ Links ]
48. Quadros, L.P. e Melo, J.H.G. 1987. Método prático de preparação palinológica em sedimentos prémesozóicos. Boletim de Geociências da Petrobrás 1: 205-214. [ Links ]
49. Rocha-Campos,A.C., Basei, M.A.S., Nutman, A.P. e Santos, P.R. 2006. Shrimp U-Pb Zircon geochronological calibration of the Late Paleozoic Supersequence, Paraná Basin, Brazil. 5º South American Symposium on Isotopic Geology (Punta del Este), Short Papers pp. 298-301. [ Links ]
50. Rocha-Campos, A.C., Basei, M.A.S., Nutman, A.P. e Santos, P.R. dos. 2007. SHRIMP U-PB zircons ages of the late Paleozoic sedimentary sequence, Paraná Basin, Brazil. 4º Simpósio sobre Cronoestratigrafia da Bacia do Paraná (Búzios), Boletim de Resumos p. 33. [ Links ]
51. Russo, A., Archangelsky, S. e Gamerro, J.C. 1980. Los depósitos suprapaleozóicos em el subsuelo de la Llanura Chaco-Pampeana, Argentina. 2º Congresso Argentino de Paleontologia e Bioestratigrafía y 1º Congresso Latinoamericano de Paleontogia (Buenos Aires), Actas 4: 157-173. [ Links ]
52. Santos, R.V., Souza, P.A., Alvarenga, C.J.S. de, Dantas, E.L., Pimentel, M.M., Oliveira, C.G. de. e Araújo, L.M. de. 2006. Shrimp U-Pb zircon dating and palynology of bentonitic layers from the Permian Irati Formation, Paraná Basin, Brazil. Gondwana Research 9: 456-463. [ Links ]
53. Souza, P.A. 2006. Late Carboniferous palynostratigrafphy of the Itararé Subgroup, northeastern Paraná Basin, Brazil. Review of Palaeobotany and Palynology 138: 9-29 [ Links ]
54. Souza, P.A. e Marques-Toigo, M. 2003. An overview in the Palynostratigraphy of the Upper Paleozoic strata of the Brazilian Paraná Basin. Revista del Museo Argentino de Ciencias Naturais 5: 205-214. [ Links ]
55. Souza, P.A. e Marques-Toigo, M. 2005. Progress on the palynostratigraphy of the Permian strata in Rio Grande do Sul State, Paraná Basin, Brazil. Anais da Academia Brasileira de Ciências 77: 353-365. [ Links ]
56. Souza, P.A. e Milani, E.J. 2007. O pioneirismo do zoneamento palinoestratigráfico de Daemon e Quadros (1970): um marco no conhecimento estratigráfico da Bacia do Paraná. Boletim de Geociências da Petrobrás 15: 118-130. [ Links ]
57. Tiwari, R.S. e Navale, G.K.B. 1967. Pollen and spore assemblage in some coals of Brazil. Pollen et spores 9: 583-605. [ Links ]
58. Trindade, N.M. 1954. Megásporos do carvão gondwânico do Rio Grande do Sul e sua aplicação em correlações estratigráficas. Notas preliminares e estudos 78: 1-6. [ Links ]
59. Vergel, M.M. 1993. Palinoestratigrafia de la secuencia neopaleozoica en la Cuenca Chacoparanaense, Argentina. 12º Congrès International de la Stratigraphie et Géologie du Carbonifère et Permien (Buenos Aires), Comptes Rendus 1: 201-212. [ Links ]
60. Wildner, W. 1983. Análise faciológica de uma porção do Grupo Guatá, na região de Candiota, RS: I. Acta Geológica Leopoldensia 8: 145-168. [ Links ]
61. Ybert, J.P. 1975. Etude des miospores du bassin houiller de Candiota-Hulha Negra, Rio Grande do Sul, Bresil. Pesquisas 5: 181-226. [ Links ]
62. Zeiller, R. 1895. Note sur l aflore fossile des gisements houillers de Rio Grande do Sul. Bulletin de la Societe Geologique de France 23: 601-623. [ Links ]
Recibido: 13 de abril de 2009.
Aceptado: 21 de octubre de 2009.

