










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana vol.47 no.1 Buenos Aires ene./mar. 2010
NOTA PALEONTOLÓGICA
West Antarctic Rift system: a possible New Zealand-Patagonia Oligocene paleobiogeographic link
Silvio Casadío1, Campbell Nelson2, Paul Taylor3, Miguel Griffin4 and Dennis Gordon5
1Universidad Nacional de La Pampa, Uruguay 151, 6300 Santa Rosa, Argentina. scasadio@cpenet.com.ar
2University of Waikato, Private Bag 3105, Hamilton 3240, New Zealand. c.nelson@waikato.ac.nz
3Natural History Museum, Cromwell Road, London SW7 5BD, United Kingdom. p.taylor@nhm.ac.uk
4Museo de La Plata, Paseo del Bosque s/n, 1900 La Plata, Argentina. miguelgriffin@aol.com
5NIWA, PO Box 14901, Kilbirnie, Wellington, New Zealand. d.gordon@niwa.co.nz
The extant marine benthic faunas in the New Zealand region and those belonging to the Magallanes and Antarctic biogeographic provinces may share a common origin dating back as far as the Early Cretaceous (Zinsmeister, 1982; Crame, 1999). These faunas have diverged ever since but the tempo and mode of this divergence remains imperfectly understood. Studies on the biogeographical affinities among the marine benthic faunas from the Southern Hemisphere point to close paleobiogeographical relationships between New Zealand, West Antarctica and southern South America after the final break-up of Gondwana (Griffin and Hünicken, 1994; Zinsmeister and Griffin, 1995; Stilwell, 2003).
The paleobiogeographical evolution of these faunas occurred in the context of the dramatic and continuous geodynamic events that led from rela-tively warm, ice-free "greenhouse" conditions to an "icehouse" state (Francis et al., 2009). Crucial oceanographic modifications took place in the Southern Hemisphere during the period between 35 and 15 Ma. These included the establishment of broad zonal belts of surface-water masses separated by oceanic fronts (Nelson and Cooke, 2001; Lagabrielle et al., 2009). These belts had a paramount role in the isolation and evolution of the marine faunas in the Southern Hemisphere (Arntz, 1999). However, paleontological evidence demonstrates that during this period paleobiogeographic connections between New Zealand and Patagonia were maintained, as shown by the presence of numerous shared taxa. Twenty two mollusk genera appearing in South American Late Oligocene-Miocene rocks are recorded also in New Zealand, but making their first appearance there in older rocks (Deminucula, Notolimopsis, Glycymeris (Glycymerula), Cyclochlamys, Gonimyrtea, Spissatella, Scalpomactra, Turia, Crosseola, Sigapatella, Trichosirius, Sassia zealta group, Ataxocerithium, Penion, Nassarius (Hima), Xymenella, Austromitra, Zeacuminia, Eoturris, Neoguraleus (Fusiguraleus), Oamaruia (Oamaruia), and Opimilda. At the same time, 16 taxa appear in South America in rocks older than those containing these taxa in New Zealand (Zygochlamys, Tiostrea, Aulacomya, Lucinoma, Antisolarium (from Valdesia), Xymene, ?Trophon, Provocator, Antimelatoma, Solemyarina, Anadara, Mytilus, Protothaca, Diloma, Argobuccinum, Fusitriton). At the same time (Late Oligocene-Early Miocene), eight genera appear in both areas (Neopanis, Austrovenus (from Ameghinomya), Cirsotrema (large species), Oamaruia (Zeadmete), Puyseguria, Cosa, Notolimea, Lamprodomina (Beu et al., 1997).
Bryozoans recorded from around the Oligocene-Miocene boundary in Patagonia are typically pan-Gondwanan. However, a recent record of a species belonging to Cinctiporidae (figure 1) -a distinctive family previously regarded as a New Zealand endemic (Boardman et al., 1992) - within the Monte León Formation (Early Miocene) cropping out in the Puerto Deseado area (47º 32' 52'S, 66º 30' 57''W), Argentina, reinforces the idea of connections between the western South Atlantic and New Zealand as late as the Early Miocene. With the exception of a doubtful occurrence in the Maastrichtian of South Africa (Boardman et al., 1992), cinctiporids were known only from the Late Paleocene-Early Eocene to Recent of New Zealand (Gordon and Taylor, 1999) before the surprising discovery of the Argentinean material.

Figure 1. Specimen GHUNLPam 28.000 from Puerto Deseado area (Argentina) belonging to the cyclostome bryozoan family Cinctiporidae hitherto considered to be endemic to New Zealand. The specimen is housed in the Departamento de Ciencias Naturales of the Universidad Nacional de La Pampa / especímen GHUNLPam 28.000 del área de Puerto Deseado (Argentina) perteneciente a un briozoo ciclostomado de la familia Cinctiporidae hasta ahora considerada endémica de Nueva Zelanda. El ejemplar se encuentra depositado en la colección del Departamento de Ciencias Naturales de la Universidad Nacional de La Pampa.
Despite the enormous distance separating New Zealand and South America during the Oligocene (15,000 km eastward from South America to New Zealand and 9,000 km from New Zealand to South America), these affinities among benthic faunas with mainly short-lived larvae have been attributed by Beu et al. (1997) largely to the action of the Antarctic Circumpolar Current, established as a consequence of the opening of Drake Passage. However, if this were true it would be difficult to explain the low dispersion rate after the Early Miocene.
Here we postulate that the paleobiogeographic connection may have been along the West Antarctic Rift system. This system underwent an episode of significant extension during the Eocene-Oligocene (Cande et al., 2000), especially evident from seismic imaging in the McMurdo Sound (Fielding et al., 2008), and at the same time the full opening (ca. 31 Ma) of the Tasmanian gateway possibly elicited the onset of a protocircum-Antarctic flow of cold waters, perhaps through a West Antarctic seaway connecting the southern Pacific and Atlantic Oceans (figure 2), a scenario advocated by Nelson and Cooke (2001). While conclusive support for this hypothesis is presently limited, information assembled during the last decade after research in McMurdo Sound is very enlightening. Boreholes CRP-2/2a and CRP-3 of the Cape Roberts Project cored approximately 1,300 m of Oligocene sediments in the Victoria Land Basin of the Ross Sea (CRST, 1999a, 1999b, 2000a, 2000b), yielding mollusks (Taviani and Beu, 2001), corals (Stolarski and Taviani, 2001), and ostracods (Dingle and Majoran, 2001) that show affinities with New Zealand and Patagonian taxa alike. Erratic boulders with giant oysters found in McMurdo Sound (albeit with no precise dating) strongly suggest the existence of great oyster reefs along the Antarctic shelf (Stilwell, 2000) similar to those described from Late Oligocene-Early Miocene successions in Patagonia (Parras and Casadío, 2005, 2006) and New Zealand (Nelson et al., 1983). These oysters built framework reefs in shallow-shelf environments, forming large biogenic concentrations exposed at many localities in Patagonia and New Zealand (figure 3).
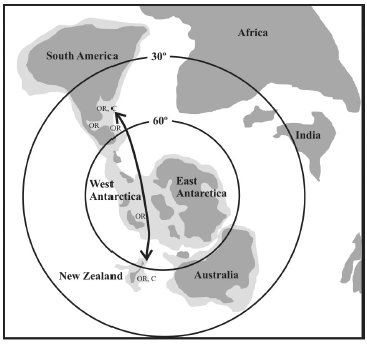
Figure 2. Schematic paleogeographic map showing the suggested marine connections during the Oligocene. OR = oyster reefs, C = Cinctiporidae. Base map modified from Nelson and Cooke (2001), Ghidella et al. (2002) and Ghiglione et al. (2008) / mapa paleogeográfico esquemático mostrando las conexiones marinas sugeridas para el Oligoceno. OR = arrecifes ostreros, C = Cinctiporidae. Mapa base modificado de Nelson y Cooke (2001), Ghidella et al. (2002) y Ghiglione et al. (2008).

Figure 3. Oyster reefs. 1, oyster reef in the Late Oligocene-Early Miocene Centinela Formation, La Siberia, Santa Cruz, Argentina; 2, Oyster reef in the Late Oligocene-Early Miocene Te Kuiti Group, Piripiri Cave, Waitomo, New Zealand / arrecifes ostreros. 1, arrecife ostrero en la Formación Centinela (Oligoceno Tardío-Mioceno Temprano), La Siberia, Santa Cruz, Argentina; 2, Arrecife ostrero en el Grupo Te Kuiti (Oligoceno Tardío-Mioceno Temprano), Cueva Piripiri, Waitomo, Nueva Zelanda.
In conclusion, there is an accumulating body of tectonic, sedimentary and paleontological information that the West Antarctic Rift system could have afforded a principal paleobiogeographic seaway linkage between New Zealand and Patagonia in the Oligocene.
References
1. Arntz, W.E. 1999. Magellan - Antarctic: Ecosystems that drifted apart. Summary review. Scientia Marina 63 (Supl. 1): 503-511. [ Links ]
2. Beu, A.G., Griffin, M. and Maxwell, P.A. 1997. Opening of Drake Passage gateway and Late Miocene-Pleistocene cooling reflected in Southern Ocean molluscan dispersal: evidence from New Zealand and Argentina. Tectonophysics 281: 83-97. [ Links ]
3. Boardman, R.S., McKinney, F.K. and Taylor, P.D. 1992. Morphology, anatomy, and taxonomy of the Cinctiporidae, new family (Bryozoa: Stenolaemata). Smithsonian Contributions in Paleobiology 70: 1-81. [ Links ]
4. Cande, S.C., Stock, J.M., Müller, R.D. and Ishihara, T. 2000. Cenozoic motion between East and West Antarctica. Nature 404: 145-150. [ Links ]
5. Crame, J.A. 1999. An evolutionary perspective on marine faunal connections between southernmost South America and Antarctica. Scientia Marina 63 (Supl. 1): 1-14. [ Links ]
6. Cape Roberts Science Team. 1999a. Summary of results. Terra Antarctica 6: 156-173. [ Links ]
7. Cape Roberts Science Team. 1999b. Palaeontology. Terra Antarctica 6: 105-144. [ Links ]
8. Cape Roberts Science Team. 2000a. Summary of results. Terra Antarctica 7: 185-209. [ Links ]
9. Cape Roberts Science Team. 2000b. Palaeontology. Terra Antarctica 7: 133-170. [ Links ]
10. Dingle, R.V. and Majoran, S. 2001. Palaeoclimatic and -biogeographical implications of Oligocene ostracoda from CRP-2/2A and CRP-3 drillholes, Victoria Land Basin, Antarctica. Terra Antarctica 8: 369-382. [ Links ]
11. Fielding, C.R., Whittaker, J., Henrys, S.A., Wilson, T.J. and Naish, T.R. 2008. Seismic facies and stratigraphy of the Cenozoic succession in McMurdo Sound, Antarctica: Implications for tec-tonic, climatic and glacial history. Palaeogeography, Palaeoclimatology, Palaeoecology 260: 8-29. [ Links ]
12. Francis, J.E., Marenssi, S., Levy, R., Hambrey, M., Thorn, V.C., Mohr, B., Brinkhuis, H., Warnaar, J., Zachos, J., Bohaty, S. and DeConto, R. 2009. From Greenhouse to Icehouse-The Eocene/Oligocene in Antarctica. In: F. Florindo and M. Siegert (eds.), Antarctic Climate Evolution. Developments in Earth & Environmental Sciences 8: 309-368. [ Links ]
13. Ghidella, M.E., Yáñez, G. and LaBrecque, J.L. 2002. Revised tectonic implications for the magnetic anomalies of the western Weddell Sea. Tectonophysics 347: 65-86. [ Links ]
14. Ghiglione, M.C., Yagupsky, D., Ghidella, M. and Ramos, V.A. 2008. Continental stretching preceding the opening of the Drake Passage: Evidence from Tierra del Fuego. Geology 36: 643-646. [ Links ]
15. Gordon, D.P. and Taylor, P.D. 1999. Latest Paleocene to earliest Eocene bryozoans from Chatham Island, New Zealand. Bulletin of the Natural History Museum, London, Geology Series 55: 1-45. [ Links ]
16. Griffin, M. and Hünicken, M.A. 1994. Late Cretaceous-early Tertiary gastropods from southwestern Patagonia. Journal of Paleontology 68: 257-274. [ Links ]
17. Lagabrielle, Y., Goddéris, Y., Donnadieu, Y., Malavieille, J. and Suárez, M. 2009. The tectonic history of Drake Passage and its possible impacts on global climate. Earth and Planetary Science Letters 279: 197-211. [ Links ]
18. Nelson, C.S. and Cooke, P.J. 2001. History of oceanic front development in the New Zealand sector of the Southern Ocean dur-ing the Cenozoic - a synthesis. New Zealand Journal of Geology and Geophysics 44: 535-553. [ Links ]
19. Nelson, C.S., Burns, D.A. and Rodgers, K.A. 1983. The taxonomic status, and isotopic evidence for paleoenvironments, of giant oysters from Oligocene Te Kuiti Group, South Auckland, New Zealand. New Zealand Journal of Geology and Geophysics 26: 289-299. [ Links ]
20. Parras, A. and Casadío, S. 2005. Stratigraphy, taphonomy and sequence stratigraphic significance of oyster-dominated concentrations from the San Julián Formation, Oligocene of Patagonia, Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology 217: 47-66. [ Links ]
21. Parras, A. and Casadío, S. 2006. The oyster Crassostrea? hatcheri (Ortmann, 1897), a physical ecosystem engineer from the upper Oligocene-lower Miocene of Patagonia, southern Argentina. Palaios 21: 168-186. [ Links ]
22. Stilwell, J.D. 2000. Eocene mollusca (Bivalvia, Gastropoda and Scaphopoda) from McMurdo Sound: systematics and paleoecologic significance. In: J.D. Stilwell and R. Feldmann (eds.), Paleobiology and paleoenvironments of Eocene rocks, McMurdo Sound, East Antarctica. Antarctic Research Series 76: 261-320. [ Links ]
23. Stilwell, J.D. 2003. Patterns of biodiversity and faunal rebound following the K/T boundary extinction event in Austral Palaeocene molluscan faunas. Palaeogeography, Palaeoclimatology, Palaeoecology 195: 319-356. [ Links ]
24. Stolarski, J. and Taviani, M. 2001. Oligocene scleractinian corals from CRP-3 drillhole, McMurdo Sound (Victoria Land Basin, Antarctica). Terra Antarctica 8: 1-4. [ Links ]
25. Taviani, M. y Beu, A.G. 2001. Palaeogene macrofossils from CRP-3 drillhole, Victoria Land Basin, Antarctica. Terra Antarctica 8: 423-434. [ Links ]
26. Zinsmeister, W.J. 1982. Late Cretaceous-early Tertiary molluscan biogeography of the southern circun-Pacific. Journal of Paleontology 56: 84-102. [ Links ]
27. Zinsmeister, W.J., Griffin, M. 1995. Late Cretaceous and Tertiary aporrhaid gastropods from the southern rim of the Pacific Ocean. Journal of Paleontology 69: 692-702. [ Links ]
Recibido: 19 de junio de 2009.
Aceptado: 18 de setiembre 2009.

