










Services on Demand
Journal

Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAmeghiniana
On-line version ISSN 1851-8044
Ameghiniana vol.47 no.2 Buenos Aires June 2010
ARTÍCULOS ORIGINALES
Estudio tafonómico de los mamíferos pleistocenos del yacimiento de Playa del Barco (Pehuen Co), provincia de Buenos Aires, Argentina
Rodrigo L. Tomassini1-2, Claudia I. Montalvo2, Teresa Manera3-4 y Cristian Oliva4
1CONICET. rodher23@yahoo.com.ar
2Facultad de Ciencias Exactas y Naturales, UNLPam. Avenida Uruguay 151, 6300 Santa Rosa, La Pampa, Argentina. cmontalvo@exactas.unlpam.edu.ar
3Departamento de Geología, Universidad Nacional del Sur, San Juan 670, 8000 Bahía Blanca, Buenos Aires, Argentina. tmanera@criba.edu.ar
4Museo Municipal de Ciencias Naturales "Carlos Darwin", Urquiza 123, 8109 Punta Alta, Buenos Aires, Argentina. cristianoliva78@yahoo.com.ar
Resumen. Se dan a conocer los resultados del estudio tafonómico comparativo realizado sobre restos de mamíferos fósiles de Playa del Barco, provincia de Buenos Aires, Argentina. En este yacimiento se reconocieron dos niveles portadores que forman parte de un ambiente fluvial, uno constituido fundamentalmente por conglomerados y el otro por areniscas limosas. Los restos de mamíferos recuperados se incluyen en los órdenes Xenarthra, Notoungulata, Litopterna, Rodentia, Carnivora, Perissodactyla, Artiodactyla y Proboscidea, todos ellos constituyentes de la típica megafauna del Pleistoceno. Teniendo en cuenta el nivel de procedencia, se observaron variaciones en los atributos tafonómicos de los materiales analizados. Los materiales provenientes del conglomerado presentan evidencias que indican su enterramiento rápido y que luego fueron sometidos a procesos de reelaboración antes de su depositación final. Los fósiles procedentes de las areniscas limosas presentan indicios de una exposición prolongada a la intemperie, sometidos con mayor intensidad a procesos ocurridos antes del enterramiento. Por sus características se interpretó que se trataría de restos resedimentados. Si bien en ambos casos se infieren procesos relacionados con el transporte en masa, la diversidad en el estado de conservación y en los atributos tafonómicos surgirían como resultado de procesos originados en cada uno de los niveles portadores. Las evidencias litoestratigráficas, taxonómicas y tafonómicas no permiten relacionar de manera directa estos dos niveles como parte de un único evento, por lo que los fósiles que contienen se interpretan como integrantes de dos asociaciones distintas.
Abstract. .Taphonomic study of pleistocene mammals from Playa del Barco site (Pehuen Co) Buenos Aires Province, Argentina Results are presented of a comparative taphonomic study conducted on mammal remains from Playa del Barco, Buenos Aires Province, Argentina. Two bearing levels, belonging to a fluvial environment were recognized: one constituted mainly by conglomerates and the other by silty sandstones. The mammal remains collected belong to the orders Xenarthra, Notoungulata, Litopterna, Rodentia, Carnivora, Perissodactyla, Artiodactyla, and Proboscidea, all of them components of the typical Pleistocene megafauna. When taking into account the source level, variations in the taphonomic attributes of the analysed materials were observed. Remains from the conglomerate show evidence of having been rapidly buried and then reelaborated before their final deposition. Fossils from silty sandstones show signs of longer exposure to weather, more intensely subject to processes that took place before burial. Due to their characteristics they are interpreted as re-deposited remains. In both cases, processes related to mass transport are inferred. Nevertheless, the diversity in conservation status and taphonomic attributes would result from processes originated at each of the source levels. The lithostratigraphic, taxonomic, and taphonomic evidences do not allow relating these two levels to a unique event. Therefore, the fossils that they contain are interpreted as members of two different associations.
Key words. Taphonomy; Mammals; Pleistocene; Playa del Barco; Buenos Aires.
Palabras clave. Tafonomía; Mamíferos; Pleistoceno; Playa del Barco; Buenos Aires.
Introducción
En el marco del estudio de asociaciones de vertebrados del Neógeno de Argentina, se ha ido incrementando paulatinamente el número de trabajos que incluyen análisis tafonómicos, lo que ha permitido ampliar las interpretaciones sobre las mismas (Tauber, 1997a, 1997b; Montalvo, 2002, 2004; Cladera et al., 2004; Montalvo et al., 2008; Pomi, 2009).
Los estudios tafonómicos de asociaciones sudamericanas que incluyen en su composición megamamíferos pleistocenos son aún escasos y restringidos generalmente al análisis de restos asociados al registro humano (Fariña, 2002; Pomi, 2006; Politis y Messineo, 2008). En particular, en los casos en que los restos de vertebrados y principalmente de mamíferos están asociados a sistemas fluviales, resulta interesante este tipo de análisis, ya que las características de los restos fósiles recuperados en cada uno de los subambientes presentan gran variabilidad, como resultado de diferentes procesos que los afectaron tanto antes como después del enterramiento (Behrensmeyer, 1982). También resulta de interés la problemática relacionada con los fenómenos de mezcla temporal (time averaging) y espacial (spatial averaging), habituales en este tipo de ambiente (Behrensmeyer, 1982, 1988; Aslan y Behrensmeyer, 1996; Trapani, 1998; Straight y Eberth, 2002).
En el presente trabajo se analizan las características tafonómicas de los restos de mamíferos fósiles provenientes del yacimiento de Playa del Barco, incluyendo las geoquímicas y mineralógicas. Este yacimiento pleistoceno, incluido en un sistema fluvial (Vega et al., 1989; Zavala y Quattrocchio, 2001; Aramayo et al., 2002), está ubicado al sudoeste de la provincia de Buenos Aires. Desde comienzos del siglo XX ha brindado abundantes materiales de vertebrados, en especial mamíferos asignados al Pleistoceno (Ameghino, 1908; Cabrera, 1929; Frenguelli, 1928; Kraglievich, 1934, 1946; Aramayo y Manera de Bianco, 1989; Manera de Bianco y Aramayo, 1998) e icnitas de mamíferos de la misma edad (Aramayo y Manera de Bianco, 1987; Aramayo et al., 2002; Aramayo et al., 2005).
El material analizado está constituido por restos de mamíferos fósiles provenientes de dos niveles portadores, en los que se observaron variaciones en los estados de conservación que se pueden relacionar con los diferentes procesos tafonómicos que los afectaron. Con los resultados obtenidos se realiza un análisis comparativo que, sumado a los datos taxonómicos y estratigráficos, contribuye a mejorar el conocimiento de las condiciones de formación del yacimiento.
Ubicación y marco geológico
Playa del Barco es un yacimiento pleistoceno ubicado sobre la costa sudoeste de la provincia de Buenos Aires (61º 34' 52''O y 39º 00' 09''S), en inmediaciones de la localidad balnearia de Pehuen Co y a unos 70 km al sudeste de la ciudad de Bahía Blanca (figura 1.1).
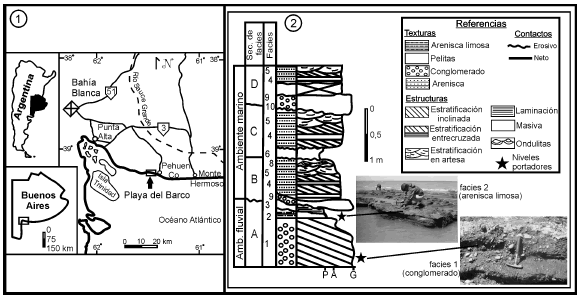
Figura 1. 1, Mapa de ubicación geográfica de Playa del Barco / location map of Playa del Barco. 2, Sección columnar integrada. Modificada de Vega et al. (1989) / integrated columnar section. Modified from Vega et al. (1989).
Los afloramientos ubicados sobre la línea de bajamar son discontinuos y su observación está supeditada a la disponibilidad de arena en la playa, que los descubre en forma esporádica. Existen variaciones sedimentológicas a lo largo del perfil, de modo tal que Vega et al. (1989) definieron diez facies distintas, que incluyen depósitos conglomerádicos, areniscas con variaciones en las estructuras, areniscas arcillosas y pelitas, asociadas a ambientes fluviales y marinos someros. Los niveles portadores corresponden a la secuencia de facies de barra longitudinal y llanura de inundación del sistema fluvial (secuencia A), de acuerdo a la clasificación de Vega et al. (1989) (figura 1.2). Por su parte, Zavala y Quattrocchio (2001) reconocieron, dentro de los conglomerados, facies de tipo clasto-sostén, interpretadas como depósitos residuales y otros niveles matriz-sostén que evidencian debris- flow cohesivos, flujos hiperconcentrados y corrientes de turbidez de alta densidad. Estos conglomerados corresponden a la facies 1 de Vega et al. (1989), son portadores de una gran variedad de restos de vertebrados pleistocenos y, además, presentan grandes bloques de limolitas terciarias correspondientes a la Formación Monte Hermoso, que en algunos casos incluyen restos fósiles asignables a la edad Montehermosense (Mioceno tardío-Plioceno temprano; e.g. Pseudotypotherium sp.). Sin embargo, la mayor riqueza fosilífera, en cuanto a cantidad, proviene de un pequeño banco de areniscas limosas color pardo claro, probablemente asignables a la facies 2 de Vega et al. (1989) y que solo aflora en bajamares extremas (Aramayo y Manera de Bianco, 1989).
Más allá de las asociaciones faciales propuestas por Vega et al. (1989), esta dependencia de las mareas es la que ha impedido hasta el momento el reconocimiento de una relación estratigráfica clara entre estos dos niveles fosilíferos. Zavala y Quattrocchio (2001) no describieron los niveles de areniscas limosas y, por lo tanto, no los relacionaron con las facies conglomerádicas.
Frenguelli (1928) consideró que tanto los conglomerados como las areniscas limosas fueron originados por paleocursos del arroyo Las Mostazas o del río Sauce Grande, asignando los materiales provenientes de las areniscas limosas al Lujanense (Pleistoceno tardío- Holoceno temprano). Kraglievich (1930, 1934) ubicó esta asociación en la cúspide del Ensenadense (Pleistoceno temprano), nominándola como fauna Barcoense.
Zavala y Quattrocchio (2001) correlacionaron estratigráficamente los niveles conglomerádicos con los de la secuencia San José, interpretada primeramente como Ensenadense (Deschamps y Borromei, 1992; Pardiñas y Deschamps, 1996; Deschamps, 1998), pero que estudios biocronoestratigráficos recientes (Verzi et al., 2004; Deschamps, 2005) la ubican en el Bonaerense (Pleistoceno medio). Un fechado cronológico radiocarbónico realizado sobre un cuerpo vertebral de un ejemplar de Lestodon sp. proveniente de las areniscas limosas, arrojó una edad de 16.440 ± 320 años AP, ubicándolo en el Lujanense (Aramayo, 1997).
Materiales y métodos
La muestra analizada consta de 468 piezas y corresponde únicamente a materiales de mamíferos hallados in situ, incluyendo restos óseos determinables y fragmentos indeterminados. Los mismos se encuentran depositados en la colección del Museo Municipal "Carlos Darwin" de Punta Alta y en el laboratorio de paleontología de la Universidad Nacional del Sur, Bahía Blanca.
Para el análisis de las variables tafonómicas se utilizó principalmente la metodología y las categorías propuestas por Alcalá (1994). Sobre cada uno de los restos se consideraron las siguientes características: determinación anatómica y taxonómica; grados de meteorización, abrasión, articulación y empaquetamiento; integridad y tipo de fracturas; color e impregnaciones; bioerosión y encostramiento. Estos dos últimos atributos no se registraron en ningún resto. Teniendo en cuenta que algunos materiales con los cuales se trabajó pertenecen a colecciones antiguas, los datos de campo -orientación, buzamiento y posición estratigráfica- están basados en la información disponible en las fichas de colección. El análisis de los tipos de fracturas se realizó en los distintos huesos largos de las extremidades y para ello se consideró el ángulo formado entre el eje de mayor dimensión y la fractura, reconociéndose fracturas longitudinales (ángulos entre 0° y 29°), oblicuas (ángulos entre 30° y 59°) y transversales (ángulos entre 60° y 90°). Para evaluar las modificaciones en el color de los restos, vinculadas al grado de impregnación que presentan, se proponen cinco categorías (modificado de López- González et al., 2005) que varían entre ausencia total de impregnación (categoría 0) e impregnación completa (categoría 4).
También se realizaron estudios histológicos, mineralógicos y geoquímicos que incluyen análisis de difractometría de rayos X, EDX (Energy Dispersive Xray), microscopía electrónica de barrido (SEM) y observación de secciones delgadas bajo lupa binocular y microscopio petrográfico. Las secciones delgadas, transversales y longitudinales, se realizaron sobre fragmentos de costillas y fragmentos indeterminados provenientes tanto de las areniscas limosas como del conglomerado, utilizando la metodología descripta por Cuezva y Élez (2000). Para los estudios de rayos X se utilizó un Difractómetro Rigaku D-Max III-C con radiación de Cu Ka y monocromador de grafito, en condiciones de 35 Kv y 15mA, perteneciente a la Universidad Nacional del Sur (UNS). La base de datos utilizada fue la ICDD (1993). Los estudios de composición elemental se realizaron con un sistema EDX, marca EDAX, modelo DX-4, con capacidad de detección desde Boro hasta Uranio, y algunos de los materiales se fotografiaron mediante un JEOL 35 CF de la Unidad de Administración Territorial (UAT) del Centro Científico y Tecnológico CONICET Bahía Blanca (CCT-CONICET-BB), Argentina.
Resultados
Análisis taxonómico y anatómico
Los materiales recuperados del yacimiento de Playa del Barco (Anexo 1) provienen tanto de los niveles conglomerádicos (n=137) como de las areniscas limosas (n=331) y están integrados principalmente por mamíferos de los órdenes Xenarthra, Notoungulata, Litopterna, Rodentia, Carnivora, Perissodactyla, Proboscidea y Artiodactyla. En menor proporción y provenientes exclusivamente del conglomerado, se encuentran restos de aves, reptiles y peces. La mayoría de los elementos esqueléticos recuperados de ambos niveles provienen de individuos cuya masa corporal estimada supera los 1.000 kg (Fariña et al., 1998) y no se han hallado restos de mamíferos de menos de 5 kg de masa corporal estimada.
Entre los restos de los depósitos conglomerádicos, el 89,79% fue determinado taxonómicamente; hay una alta diversidad de taxones, con predominio de Artiodactyla (40,88%), principalmente Cervidae, y Xenarthra (27,01%). En cambio, en las areniscas limosas, sólo el 58,91% pudo ser determinado taxonómicamente y se observó un marcado predominio de Xenarthra (49,85%), en particular de Lestodon sp., mientras que los representantes de otros órdenes de mamíferos son escasos (figura 2.1).

Figura 2. 1, Distribución de frecuencia de los órdenes de mamíferos y restos indeterminados registrados en ambas facies / frequency distribution of recorded mammal orders and indeterminate remains from both facies. 2, Distribución de frecuencias de los elementos anatómicos y fragmentos indeterminados registrados en ambas facies / frequency distribution of recorded anatomical elements and indeterminate fragments from both facies.
El 3,64% del total de restos del conglomerado y el 13,59% de las areniscas consistían en astillas y fragmentos indeterminables anatómicamente. En ambos niveles están representados la mayoría de los elementos esqueléticos, pero hay importantes variaciones en cuanto a su frecuencia (figura 2.2). Sólo las vértebras y las costillas predominan en las areniscas.
Grado de meteorización
Alcalá (1994), basado en los estadíos propuestos previamente por Behrensmeyer (1978) para macromamíferos actuales, propuso tres categorías de meteorización para el análisis de este atributo en restos fósiles de grandes vertebrados. De acuerdo a este criterio, el 85,40% de los materiales del conglomerado se encuentran intactos (categoría 1, sin ningún tipo de alteración), mientras que los demás ejemplares presentan pérdida superficial de materia ósea (categoría 2) (figura 3.1). En las areniscas limosas, el porcentaje de restos intactos es menor (57,40%) y aumenta considerablemente el porcentaje de restos en las categorías 2 (35,95%) y 3 (6,65%), con pérdida de materia ósea profunda y exposición de tejido esponjoso (tabla 1).

Figura 3. 1, Metatarso de Lestodon sp. con evidencias de meteorización en las epífisis / metatarsus of Lestodon sp. with weathering evidences on the epiphyses. 2, Fragmento de metápodo de Artiodactyla indet. con fractura transversal / metapodial fragment of Artiodactyla indet. with transverse fracture. 3, Epífisis distal de húmero de Artiodactyla indet. con fractura longitudinal / humeral distal epiphysis of Artiodactyla indet. with longitudinal fracture. 4, Osteodermo de Glyptodontidae indet. con evidencias de abrasión / osteoderm of Glyptodontidae indet. with evidences of abrasion. 5, Fragmento craneal de Morenelaphus sp. con las astas articuladas / skull fragment of Morenelaphus sp., with articulated horns. 6, Fragmento de mandíbula de Cervidae indet. proveniente de las areniscas, color pardo claro sin evidencias de impregnaciones / jaw fragment of Cervidae indet. from sandstones, light brownish colored, with no evidence of impregnations. 7, Fragmento de mandíbula de Cervidae indet. proveniente del conglomerado, de color negro debido a las impregnaciones de manganeso / jaw fragment of Cervidae indet. from the conglomerate, black colored due to manganese impregnations. 8, Fragmento de mandíbula de Cervidae indet. en posición vertical dentro de las areniscas / jaw fragment of Cervidae indet. in vertical position within the sandstones. Escala / scale bar = 3 cm.
Tabla 1. Análisis de los atributos tafonómicos presentes en los restos de ambas facies. Los valores indican número de restos / analysis of taphonomic attributes present in the remains of both facies. Values indicate the number of remains.

Integridad y tipos de fracturas
Se analizó la integridad de cada resto, considerando si los mismos se encontraban enteros o rotos. En el conglomerado, el porcentaje de elementos esqueléticos enteros (25,55%) es mayor que en las areniscas limosas (16,31%).
El 45,32% de los restos provenientes de las areniscas limosas presenta dos o más fracturas, mientras que para el conglomerado el valor es sólo del 28,47%, evidenciando que los primeros han sido afectados más intensamente por procesos de rotura (tabla 1).
El análisis de los distintos tipos de fracturas (longitudinales, transversales y oblicuas) en huesos largos mostró que el porcentaje de fracturas combinadas es elevado en ambos niveles, aunque existen marcadas diferencias en las proporciones entre las fracturas longitudinales y transversales (tabla 1). En el conglomerado predominan las fracturas transversales (27,59%) (figura 3.2) sobre las longitudinales (13,79%), mientras que en las areniscas limosas las transversales son muy escasas (6,67%) y se incrementa notablemente el porcentaje de fracturas longitudinales (43,33%) (figura 3.3).
Grado de abrasión
La abrasión es el proceso de desgaste que genera la suavización de las aristas y vértices de los restos (Fernández López, 2000). De acuerdo a las categorías propuestas por Alcalá (1994), la mayoría de los restos provenientes de ambos niveles no evidenció indicios de abrasión (categoría 1) y ninguno mostró signos de abrasión extrema (categoría 3). El 23,36% de los ejemplares provenientes del conglomerado presenta la categoría 2 (redondeado) (figura 3.4), mientras que entre los de las areniscas limosas solo el 6,34% alcanza este nivel (tabla 1). Se destaca que algunos de los restos del conglomerado que tienen fracturas con sus cavidades rellenas de sedimento consolidado presentan redondeamiento también en el relleno.
Grado de articulación
De acuerdo a los distintos estados de articulación definidos por Behrensmeyer (1991) los materiales articulados son aquellos que están representados por elementos esqueléticos en conexión, manteniendo sus posiciones anatómicas relativas.
Sólo en los niveles conglomerádicos se hallaron elementos esqueléticos con algún grado de articulación. Se trata de unos pocos restos craneanos de distintos taxones, en el caso de algunos cérvidos articulados con las astas (figura 3.5), fragmentos de caparazón de gliptodontes y dos vértebras dorsales de un proboscídeo.
Empaquetamiento
Este atributo hace referencia al modo en el que los elementos conservados se encuentran ordenados o distribuidos dentro del cuerpo rocoso que los contiene. Uno de los conceptos utilizados para determinar el empaquetamiento es la densidad, es decir, el número de restos por unidad de volumen (Fernández López, 2000; Behrensmeyer, 1991). Considerando, por un lado, las dimensiones estimadas para los depósitos aflorantes de las areniscas limosas (400 m2 aproximadamente según Aramayo y Manera de Bianco, 1989) y, por otro, la totalidad de elementos analizados (n=331) que provienen de ese área, se determinó que la densidad por unidad de superficie para esta facies es de 0,82 restos/m2, aproximadamente.
Color e impregnaciones
Una de las características macroscópicas más evidente que distingue a los restos de ambos niveles es la variación en el color que presentan. En las areniscas limosas son principalmente pardo claros (figura 3.6), mientras que en el conglomerado son negros (figura 3.7) y en menor medida pardo oscuros. El análisis de los cortes delgados realizados sobre restos de ambos niveles evidenció la presencia de óxidos de manganeso, sugiriendo que esas impregnaciones son las que provocan el color negro y las manchas oscuras (Tomassini et al., 2006). Además, macroscópicamente, estas impregnaciones pueden ser reconocidas fácilmente por su característico hábito dendrítico.
No se observaron evidencias de impregnación en el 8,03% de los restos del conglomerado ni en el 30,21% de los de las areniscas limosas (tabla 1). El contraste resulta evidente si se tiene en cuenta que más de la mitad de los restos del conglomerado están totalmente impregnados (categoría 4) o muy impregnados (categoría 3), en tanto que en las areniscas limosas sólo el 9,96% de los restos se ubica en esas categorías y aquéllos que se incluyen en las categorías 1 y 2 presentan impregnaciones limitadas a pequeños manchones dendríticos que ocupan áreas superficiales de diferente tamaño.
Análisis histológico, geoquímico y mineralógico
La mayoría de los cortes delgados realizados muestran la microestructura ósea original perfectamente conservada (figura 4.1) y sólo en algunos se registraron alteraciones tafonómicas que la modifican. Las estructuras que corresponden a las osteonas presentan los canales centrales (de Havers) rellenos, en algunos casos, con apatito recristalizado (figura 4.2), óxidos de manganeso (figura 4.3) y en menor medida por carbonatos como la calcita (figura 4.4). Se distinguen también los canales oblicuos o transversales de Volkmann (De Ricqlés et al., 1991; Fawcett, 1995; Paniagua, 1996), en la mayoría de los casos rellenos por óxidos de manganeso (figura 4.5). La observación de los cortes longitudinales con nicoles cruzados reveló la presencia de cristales alargados de apatito con hábito fibroso (figura 4.6). Estos se ubican según la estructura original del fosfato del hueso, orientados de acuerdo a la disposición de las fibras de colágeno (según Lucas y Prévôt, 1991).

Figura 4. 1, Sección delgada transversal de una costilla con la microestructura ósea perfectamente conservada / transversal thin section of a rib with perfectly preserved bone microstructure. 2, Osteona con su canal central (de Havers) relleno con apatito recristalizado / osteon with central canal (Haversian) filled with recrystallized apatite. 3, Óxidos de manganeso rellenando parte del canal central de una osteona / manganese oxides filling part of the central canal of an osteon. 4, Canal de Havers con un cristal de calcita en el centro y óxidos de manganeso en los bordes / haversian canal with a calcite crystal in the centre and manganese oxides on the edges. 5, Canales de Havers y Volkmann rellenos de óxidos de manganeso / Haversian and Volkmann's canals filled with manganese oxides. 6, Cristales de apatito con hábito fibroso / apatite crystals with fibrous habit.
El análisis de difractometría de rayos X muestra que los restos de ambos niveles conservan su composición mineralógica original prácticamente sin variaciones, si bien cuando se los compara con un gráfico de referencia de hidroxilapatito (ICDD, 1993, diagrama número 9-432) se observan leves desplazamientos en algunos picos (figura 5.1-2).

Figura 5. 1, Análisis de difracción de rayos X en un fragmento de costilla del conglomerado / X-ray diffraction analysis of a rib fragment from the conglomerate. 2, Análisis de difracción de rayos X en un fragmento de costilla de las areniscas / X-ray diffraction analysis of a rib fragment from sandstones.
Los estudios de EDX confirmaron la presencia de manganeso en restos del conglomerado y en menor medida también en los de las areniscas. El manganeso, como se mencionó, se relaciona directamente con las impregnaciones y la coloración oscura de los fósiles (figura 6.1-2).

Figura 6. 1, Resultado del análisis de EDX en una muestra del conglomerado / EDX analysis results of a conglomerate sample. 2, Resultado del análisis de EDX en una muestra de las areniscas / EDX analysis results of a sandstone sample.
Discusión
Los resultados obtenidos sobre la base de los restos recuperados de ambos niveles del yacimiento de Playa del Barco muestran algunas diferencias con respecto a su representación taxonómica. En las areniscas limosas no se registraron representantes de Rodentia y Carnivora, así como tampoco de peces, reptiles y aves. Hay un predominio muy marcado de Xenarthra, particularmente de Lestodon sp. (61,76% del total de Xenarthra). Este último taxón es escaso en el conglomerado, donde predominan los Artiodactyla, en especial la familia Cervidae, y los Xenarthra acorazados (figura 2.1).
En ambos niveles, los mamíferos registrados corresponden a la clásica megafauna pleistocena pampeana y se registran taxones de todas las edades-mamífero incluidas en el Pleistoceno (Anexo 1). Cabe destacar la presencia de Equus (Amerhippus) neogeus Lund, 1840 en las areniscas limosas, el cual, de acuerdo al esquema bioestratigráfico propuesto por Cione y Tonni (1999, 2005), caracteriza una biozona que representa la base bioestratigráfica del Lujanense. De este mismo nivel también proceden restos de Macrauchenia patachonica Owen, 1838. De acuerdo a Cione y Tonni (2005), este taxón abarca el lapso Bonaerense-Lujanense. Sin embargo, Deschamps (2005) lo utiliza para definir, en el sur de la provincia de Buenos Aires, la zona de Equus (Amerhippus) neogeus-Macrauchenia patachonica, ubicándola en el Lujanense y que según Cione y Tonni (2005: 195) "corresponde conceptualmente con la Biozona de Equus (Amerhippus) neogeus".
Hasta el momento, no se ha reconocido en el conglomerado la presencia de algún taxón que permita asignar una edad precisa a los sedimentos portadores.
El predominio de elementos esqueléticos correspondientes a megamamíferos (la mayoría de ellos con masas corporales estimadas superiores a los 1.000 kg) y la ausencia de restos de micromamíferos (con masas corporales estimadas menores a 5 kg) es un carácter que evidencia un sesgo en ese sentido, que podría estar vinculado con las características del mecanismo de transporte.
La frecuencia de hallazgo de los distintos elementos esqueléticos presentó diferencias en ambos niveles. En las areniscas limosas, los más abundantes son las costillas y las vértebras (25,08% y 22,96% del total respectivamente), mientras que del total de restos recuperados del conglomerado, el 21,16% corresponde a huesos largos en general, 14,60% a elementos del autopodio, el 11,68% son vértebras, el 10,95% elementos mandibulares y el 9,49% elementos craneanos (figura 2.2).
En función del comportamiento de los distintos tipos de restos ante el transporte hidráulico, Voorhies (1969) caracterizó los distintos depósitos sedimentarios. El grupo I involucra a aquellos elementos que se mueven con facilidad y por flotación con corrientes muy débiles (costillas, vértebras y escápulas); los del grupo II requieren un poco más de energía que los anteriores para ser transportados (huesos largos y metápodos) y, finalmente, los del grupo III tienden a resistirse al movimiento (cráneos y mandíbulas) por su mayor densidad. Se destaca que es marcada la diferencia de tamaño de los restos aquí analizados con respecto a los utilizados por Voorhies (1969). En ese sentido, Todd y Frison (1986), en su experimentación con huesos de elefante en un canal fluvial natural, obtuvieron resultados similares a los descriptos por Voorhies (1969) experimentando con huesos de animales de menor tamaño en un medio artificial.
Tanto en el conglomerado como en las areniscas limosas están representados juntos todos los elementos esqueléticos. Sin embargo, como se indicó, costillas y vértebras (grupo I) predominan en las areniscas limosas, por lo que se podría interpretar que una corriente débil los movilizó hasta el área. En el conglomerado predominan los huesos largos, elementos del autopodio, elementos craneanos y elementos mandibulares (grupo IIIII), lo cual podría estar indicando el desarrollo de una corriente de mayor energía que los movilizara.
El índice dientes aislados / vértebras (Behrensmeyer, 1975) evalúa la proporción de estos elementos de acuerdo a su propia densidad y permite determinar el grado de selección hidrodinámica, obteniéndose relaciones cercanas a 1:1 cuando hay ausencia de selección. En las areniscas limosas, este índice da un valor muy bajo (1 / 5,84), debido a la gran cantidad de vértebras presentes, lo que podría indicar una cierta selección hidrodinámica hacia elementos de baja densidad por parte, como se mencionó previamente, de alguna corriente de baja energía. En el conglomerado, el valor de este índice es algo mayor (1 / 2,66), más cercano a la relación 1:1, sugiriendo una menor selección hidrodinámica, probablemente por la mayor intensidad del flujo que permite la incorporación de elementos con mayor densidad.
Si bien los niveles conglomerádicos y de areniscas limosas forman parte de una secuencia fluvial, la representación anatómica no permitió identificar un claro proceso de selección de restos que se pueda vincular con el transporte hidráulico.
Los valores obtenidos en la evaluación del grado de meteorización difieren en los dos niveles. Sobre los materiales de las areniscas limosas fue importante el porcentaje de restos en la categoría 2 y hay algunos incluidos en la 3. Estos caracteres indicarían que los procesos de meteorización actuaron con mayor intensidad en este nivel. Sumado a eso, no se recuperaron elementos articulados, es alto el porcentaje de elementos esqueléticos muy fracturados y también el de restos indeterminables (tabla 1). Estas características sugieren que muchos de estos materiales habrían estado expuestos un tiempo considerable en superficie, sometidos a la intemperie.
Los materiales provenientes del conglomerado presentan sólo leves indicios de meteorización, lo que sugiere un lapso menor de exposición subaérea. Planteado así, estos restos se habrían enterrado rápidamente, lo que habría facilitado además la conservación de muchos elementos esqueléticos enteros y de los elementos articulados que se registraron en este nivel.
La alta proporción de fracturas combinadas en ambas facies evidencia la acción de procesos destructivos intensos sobre los restos e indica un elevado grado de alteración tafonómica. En las areniscas limosas es alto el porcentaje de elementos con dos o más roturas (tabla 1) y, en los huesos largos, son frecuentes las fracturas longitudinales, las que se interpretan como ocurridas en la etapa previa al enterramiento cuando se produce la contracción del tejido óseo, en particular de las fibras de colágeno (Haynes, 1980). Además, son muy escasas las fracturas transversales, con la rotura producida en ángulo recto con respecto a la dirección principal de las fibras de colágeno (Alcalá y Escorza, 1998), las cuales se habrían originado después del enterramiento. En el conglomerado, por el contrario, es más elevado el porcentaje de restos enteros y disminuye notablemente la cantidad de materiales con dos o más roturas. Entre los huesos largos de esta facies predominan las fracturas transversales.
En cuanto al grado de abrasión, se encontró un mayor porcentaje de elementos redondeados entre los provenientes del conglomerado (tabla 1). Muchos de los huesos largos de esta facies que tienen fracturas transversales (producidas después del enterramiento) presentan redondeamiento en la superficie de fractura, lo cual indica que la abrasión por transporte fue posterior a la rotura.
El análisis de la composición de los restos surgido a partir de la difractometría de rayos X, evidencia variaciones en la posición de algunos picos respecto de los diagramas de referencia del hidroxilapatito, que probablemente se pueden explicar debido al incremento en el contenido de calcio en los restos fósiles.
Como se indicó, el color oscuro de los restos está vinculado a las impregnaciones por óxidos de manganeso. La presencia de manganeso fue confirmada mediante los análisis de EDX para los restos provenientes de ambos niveles; sin embargo, en los diagramas obtenidos para los materiales de las areniscas limosas la proporción de manganeso es muy baja. Este resultado es coherente con las observaciones macroscópicas de estos restos, ya que las impregnaciones, cuando están presentes, se limitan a manchones dendríticos superficiales y pequeños. Si bien las impregnaciones pueden aportar información acerca de diferentes procesos tafonómicos que afectaron a los restos y sus secuencias de formación (López-González et al., 2005), por sus características en los restos de Playa del Barco no fue posible determinar el momento en el que se produjeron. Sólo se pudo relacionar este atributo con las fracturas. La impregnación que presentan las superficies de fractura de algunos restos indicaría que la rotura ocurrió primero y facilitó el ingreso de los minerales al interior del hueso.
La presencia de óxidos de manganeso cubriendo superficies y rellenando cavidades en los restos es indicativa de un ambiente con condiciones alcalinas (valores elevados de pH) con intensificación de las condiciones oxidantes (González-Bonorino, 1972, fig. 15; Marín Arroyo et al., 2008).
De lo expuesto surge que la mayoría de los elementos provenientes de las areniscas limosas muestran elevada meteorización, poca abrasión y fracturas que pueden relacionarse con procesos ocurridos tempranamente en la historia tafonómica. Por otro lado, la mayoría de los materiales del conglomerado presentan porcentajes altos de abrasión y predominio de fracturas transversales en huesos largos, características que se habrían producido en etapas posteriores al enterramiento.
En los restos del conglomerado se observa una importante variabilidad en el estado de conservación, de modo tal que coexisten materiales que, en parte, se podrían incluir en el modelo I (elementos rápidamente enterrados) junto con otros del modelo II (restos con diferentes grados de meteorización y moderada fragmentación) y del modelo III (baja proporción de restos articulados y materiales con evidencia de abrasión), siguiendo la propuesta de Johnson (1960) para caracterizar los distintos tipos de formación de concentraciones fósiles. Por otro lado, la diversidad en los estados de conservación es notablemente menor en las areniscas limosas y la mayoría de los restos provenientes de las mismas responde al modelo II.
Considerando el contexto sedimentológico de ambos niveles y los caracteres tafonómicos antes expuestos, un mecanismo que podría explicar estos tipos de concentraciones de fósiles (con variabilidad en los estados de conservación y caracteres tafonómicos) es el de transporte en masa.
En Playa del Barco, la presencia de un sistema de debris flow cohesivos o flujos de alta densidad (Zavala y Quattrocchio, 2001) implicó la posibilidad de una fuerte erosión, arranque y transporte en masa de todos los materiales que encontró a su paso. En los debrisflow cohesivos, el mecanismo de sustentación es la cohesión interna de la matriz, por lo cual los materiales viajan "flotando" dentro de esa masa de sedimento viscoso, casi sin contacto unos con otros e incluso, en algunos casos, es común la presencia y movilización de bloques de grandes dimensiones (Iverson, 1997; Reineck y Singh, 1980).
En los niveles conglomerádicos, estas características están evidenciadas a partir de algunos atributos tafonómicos de los restos (elementos articulados, elementos en posición vertical, elementos enteros, bajo porcentaje de elementos con más de dos fracturas) y por la presencia de grandes bloques con fósiles de edad Montehermosense dentro de estos depósitos pleistocenos (Zavala y Quattrocchio, 2001).
Esta capacidad de los debris flow de arrastrar todo lo que encuentran a su paso también es coherente con los datos de frecuencia de distribución anatómica observada en el conglomerado, en el cual están representados todos los elementos esqueléticos (figura 2.2). El predominio de restos de densidad elevada ( huesos largos, elementos craneanos y mandibulares) avalaría el desarrollo de un flujo de energía considerable y con bajo grado de selección hidrodinámica (de acuerdo también al valor obtenido del índice dientes aislados / vértebras).
Además, en esta facies se identificaron elementos reelaborados (sensu Fernández López, 2000). Se trata de los restos articulados, algunos de ellos con evidencias de abrasión (fragmentos de caparazón de gliptodontes), otros que además presentan abrasión en los sedimentos que rellenan fracturas y cavidades y, entre los huesos largos, los que presentan fracturas transversales. La presencia de este tipo de restos avalaría también el transporte en masa, ya que los mismos habrían sido desenterrados y desplazados hacia otra área para su depositación y enterramiento final. La abundancia de elementos enteros y otros articulados entre sí, dentro de los niveles conglomerádicos, se puede explicar considerando que la integridad de los restos y las articulaciones se mantuvieron debido únicamente a este tipo de transporte en masa tan particular, en el cual los elementos viajan "flotando" y casi sin contacto entre ellos.
Sumado a lo anterior, en estos ambientes es común que algunos elementos queden atrapados en posiciones más o menos inclinadas o poco estables (Fernández López, 2000). Este carácter de "congelamiento cohesivo", propio de este tipo de depósitos, en el cual los materiales quedan en la misma posición en la que fueron transportados (Iverson, 1997; Reineck y Singh, 1980), fue identificado en el campo por uno de los autores (T. M.), tanto en el conglomerado (clastos y restos fósiles) como en las areniscas limosas (restos fósiles) (figura 3.8).
Los atributos tafonómicos que se observan en la mayoría de los restos de las areniscas limosas son alto grado de rotura, predominancia de fracturas longitudinales, baja abrasión e importante meteorización. Es común que algunas de estas características estén presentes en restos recuperados en ambientes de llanura de inundación, ya que generalmente permanecen más tiempo expuestos en superficie sometidos a la intemperie (Behrensmeyer, 1975, 1982). Sin embargo, en la asociación proveniente de las areniscas limosas no se identificaron otras evidencias que permitan avalar ese origen.
En su análisis sedimentológico, Vega et al. (1989) mencionaron la presencia de laminación paralela, grietas de desecación e icnitas de aves; sin embargo, se referían a los depósitos pelíticos correspondientes a la facies 3 (figura 1.2), la que interpretaron como ambientes de llanura de inundación. En este tipo de ambiente, la inclinación de los restos muchas veces está relacionada con fenómenos de pisoteo o "trampling" (Behrensmeyer, 1988; Olsen y Shipman, 1988). Sin embargo, si bien en las areniscas limosas se encontraron restos inclinados, no se identificaron caracteres relacionados a ese proceso como raspaduras, tipo de fracturas y marcas (macro y microscópicas) en la superficie cortical, en forma de líneas muy finas, poco profundas y orientadas en forma azarosa, que permitan confirmarlo (Behrensmeyer et al., 1986; Fiorillo, 1989; Olsen y Shipman, 1988).
La inclinación de algunos restos en las areniscas limosas avalaría la interpretación de que esos niveles corresponderían a las facies más distales dentro del flujo turbidítico conglomerádico o, por otro lado, a depósitos de transporte en masa más finos de tipo mud-flow, con génesis y caracteres distintos a los de los niveles conglomerádicos. En cualquiera de los dos casos, el flujo de sedimentos areno-limosos presentaría menor capacidad de abrasión, menor fuerza y poder de arranque que el conglomerádico, por lo que el potencial de transporte sería más limitado. Teniendo en cuenta esto y de acuerdo a los atributos tafonómicos previamente mencionados, el flujo contendría sólo aquellos materiales que se encontraban en superficie al momento de ser movilizados y que, por sus características habrían estado expuestos previamente durante un tiempo prolongado a la intemperie (probablemente en una llanura de inundación) indicando, de esta manera, que se trataría de restos resedimentados (sensu Fernández López, 2000).
De acuerdo a lo expuesto por Hill (1979), las costillas son los elementos esqueléticos que más demoran en ser arrastrados por ser los que más tarde se desarticulan. Si se tiene en cuenta que en las areniscas limosas las costillas constituyen el elemento predominante y en ningún caso se hallaban articuladas, esto significaría que, previo a la movilización por parte de este flujo areno-limoso, los restos debieron haber estado un tiempo prolongado en superficie. La presencia de restos con meteorización extrema y la ausencia de otros elementos articulados en esta facies avalarían esta idea.
El marcado predominio de costillas y vértebras, que indicaría la presencia de corrientes muy débiles, y el bajo valor del índice dientes aislados / vértebras, que sugiere cierta selección hidrodinámica hacia elementos de baja densidad, serían características coherentes para un flujo denso de baja energía, con bajo potencial de transporte, que sólo contiene materiales que han estado expuestos durante un tiempo prolongado en superficie sobre alguna llanura de inundación. Esta menor intensidad del flujo areno-limoso, respecto del conglomerádico, sería coherente con la interpretación planteada por Vega et al. (1989), quienes consideraron que estos depósitos de carga más finos se habrían originado por una desaceleración del flujo del río. Sumado a esto, los análisis de la morfoscopía de granos de cuarzo de las areniscas limosas evidencian cierto grado de desgaste fluvial (ver Aramayo y Manera de Bianco, 1989).
La densidad por unidad de superficie determinada para las areniscas limosas (0,82 restos/m2) es mucho mayor que las estimadas en llanuras actuales (Berhensmeyer, 1982) y que las obtenidas para depósitos de llanura de inundación proximales (1 resto/ 67m2) y distales (1 resto/160m2), del Pérmico tardío de la cuenca de Karoo, en Sudáfrica (Smith, 1993). Por el contrario, este resultado se asemeja más al obtenido en depósitos asociados a fenómenos de transporte en masa de tipo mud-flow (4,22 restos/m2) del Mioceno medio de Somosaguas, España (Polonio y López Martínez, 2000). Si bien en las areniscas limosas de Playa del Barco el valor es menor que en Somosaguas, esto podría deberse a que la granulometría de los niveles es más fina, por lo cual probablemente el potencial de captación y transporte de materiales haya sido menor.
De todas maneras, más allá de las distintas características tafonómicas presentes y de las asociaciones faciales previamente propuestas por Vega et al. (1989), al no existir una clara relación estratigráfica entre las areniscas y el conglomerado no es posible determinar con precisión el origen de cada depósito, el momento en el que ocurrió cada uno de los eventos y la relación que existe entre ambos.
Conclusiones
La presencia en las areniscas limosas de restos asignados Equus (Amerhippus) neogeus y Macrauchenia patachonica sugeriría para estos depósitos una edad Lujanense (Pleistoceno tardío-Holoceno temprano). Estos datos son coherentes con la edad de 16.440± 320 años AP obtenida previamente por métodos radiocarbónicos (Aramayo, 1997).
La ausencia de dataciones y fósiles guía impide mejorar la asignación temporal de la asociación recuperada del conglomerado. Sin embargo, teniendo en cuenta los biocrones de los taxones presentes, se sugiere una antigüedad máxima Bonaerense (de acuerdo al esquema bioestratigráfico de Cione y Tonni, 2005) para la ocurrencia del evento y la formación de ese depósito.
Los atributos tafonómicos observados en la mayoría de los ejemplares provenientes del conglomerado indicarían su reelaboración por acción de un flujo denso de alta energía. En tanto los de las areniscas limosas sugieren que los mismos habrían sido resedimentados, movilizados por un flujo denso de baja energía luego de haber estado un tiempo prolongado expuestos a la intemperie en una llanura de inundación.
Las características sedimentológicas, sumadas a la diversidad en el estado de conservación y en los atributos tafonómicos presentes en los restos de ambos niveles, serían el resultado de procesos originados por fenómenos de transporte en masa.
Los datos obtenidos permitirían plantear que los dos niveles y los restos que contienen se habrían originado a partir de diferentes eventos. Los atributos tafonómicos detectados serían el resultado de procesos ocurridos en cada nivel, indicando, por lo tanto, historias tafonómicas distintas para los restos de cada uno. Por el momento, las evidencias tafonómicas, taxonómicas y litoestratigráficas no permiten relacionar de manera directa estos dos niveles como parte de un único evento, por lo que los fósiles que contienen se interpretan como integrantes de dos asociaciones distintas.
Anexo 1. Listado sistemático, facies portadoras y biocrones de los taxones de mamíferos registrados / systematic list, bearing facies, and biochrons of recorded mammal taxa.


Agradecimientos
A los revisores (L. Alcalá y G. Cladera), por las correcciones y sugerencias que enriquecieron el trabajo. Los autores desean expresar su agradecimiento al Museo Carlos Darwin y a la Universidad Nacional del Sur por permitir el acceso a las colecciones, a H. Ortiz y a M.J. Yáñez por su colaboración en la preparación de las secciones delgadas y los análisis de EDX respectivamente. A E. Navarro por sus comentarios sobre procesos sedimentarios y a C. Deschamps por la lectura crítica del manuscrito.
Bibliografía
1. Alcalá, L. 1994. Macromamíferos neógenos de la fosa de Alfambra- Teruel. Instituto de estudios Turolenses y Museo Nacional de Ciencias Naturales, Teruel, 554 pp. [ Links ]
2. Alcalá, L. y Escorza, C. 1998. Modelling diagenetic bone fractures. Bulletin de la Société Géologique de France 169: 101-108. [ Links ]
3. Ameghino, F. 1902. Notas sobre algunos mamíferos fósiles nuevos o poco conocidos del valle de Tarija. Anales del Museo Nacional de Buenos Aires 3: 225-261. [ Links ]
4. Ameghino, F. 1904. Nuevas especies de mamíferos cretáceos y terciarios de la República Argentina. Anales de la Sociedad Científica Argentina 58: 225-291. [ Links ]
5. Ameghino, F. 1908. Las formaciones sedimentarias de la región litoral de Mar del Plata y Chapadmalal. Anales del Museo Nacional de Historia Natural, Serie 3: 343-428. [ Links ]
6. Aramayo, S. 1997. Cronología radiocarbónica de localidades fosilíferas pleistocenas y holocenas de la costa sud-sudeste de la provincia de Buenos Aires, Argentina. 6° Congresso da Associação Brasileira de Estudos do Quaternário e Reunido sobre o Quaternário da América do Sul (Curitiba, Paraná), Resúmenes expandidos: 305-308. [ Links ]
7. Aramayo, S. y Manera de Bianco, T. 1987. Hallazgo de una icnofauna continental (Pleistoceno tardío) en la localidad de Pehuen Co (partido de Coronel Rosales) provincia de Buenos Aires, Argentina. Parte I: Edentata, Litopterna, Proboscidea. Parte II: Carnivora, Artiodactyla y Aves. 4° Congreso Latinoamericano de Paleontología (Santa Cruz de la Sierra), Actas 1: 532-547. [ Links ]
8. Aramayo, S. y Manera de Bianco, T. 1989. Nuevos hallazgos de mamíferos pleistocénicos en el yacimiento de Playa del Barco, provincia de Buenos Aires. 1° Jornadas Geológicas Bonaerenses (Tandil, 1985), Actas: 701-712. [ Links ]
9. Aramayo, S.A., Schillizzi, R.A., Gutiérrez Téllez, B.M. 2002. Evolución paleoambiental del Cuaternario en la costa atlántica del sur de la provincia de Buenos Aires, Argentina. Revista de la Sociedad Geológica de España 15: 95-104. [ Links ]
10. Aramayo, S.A., Schillizzi, R.A., Gutiérrez Téllez, B.M. 2005. Sedimentologic and paleontologic study of the southeast coast of Buenos Aires province, Argentina. A late Pleistocene- Holocene paleoenvironmental reconstruction. Journal of South American Earth Sciences 20: 65-71. [ Links ]
11. Aslan, A. y Behrensmeyer, A. 1996. Taphonomy and time resolution of bone assemblages in a contemporary fluvial system: the East Fork River, Wyoming. Palaios 11: 411-421. [ Links ]
12. Behrensmeyer, A. 1975. The taphonomy and paleoecology of Plio- Pleistocene vertebrate assemblages of Lake Rudolf, Kenya. Bulletin of the Museum of Comparative Zoology 146: 473-578. [ Links ]
13. Behrensmeyer, A. 1978. Taphonomic and ecologic information from bone weathering. Paleobiology 4: 150-162. [ Links ]
14. Behrensmeyer, A. 1982. Time resolution in fluvial vertebrate assemblages. Paleobiology 8: 211-227. [ Links ]
15. Behrensmeyer, A. 1988. Vertebrate preservation in fluvial channels. Palaeogeography, Palaeoclimatology, Palaeoecology 63: 183- 189. [ Links ]
16. Behrensmeyer, A.K. 1991. Terrestrial vertebrate accumulations. En: P.A. Allison y D. Briggs (eds.), Taphonomy: Releasing the Data Locked in the Fossil Record. Plenum Press, New York: 291- 335. [ Links ]
17. Behrensmeyer, A. K., Gordon, K. D. y Yanagi, G. T. 1986. Trampling as a cause of bone surface damage and pseudo-cutmark. Nature 319: 768-771. [ Links ]
18. Burmeister, G. 1866. Lista de los mamíferos fósiles del terreno diluviano. Anales del Museo Público de Buenos Aires 1: 121-232. [ Links ]
19. Burmeister, G. 1874. Monografía de los Glyptodontes en el Museo Público de Buenos Aires. Anales del Museo Público de Buenos Aires 2: 367-377. [ Links ]
20. Cabrera, A. 1929. Revisión de los mastodontes argentinos. Revista del Museo de La Plata 32: 61-144. [ Links ]
21. Carette, E. 1922. Cérvidos actuales y fósiles de Sudamérica. Revisión de las formas extinguidas pampeanas. Revista del Museo de La Plata 26: 393-472. [ Links ]
22. Cione, A.L. y Tonni, E.P. 1999. Biostratigraphy and chronological scale of uppermost Cenozoic in the Pampean area, Argentina. En: E.P. Tonni y A.L. Cione (eds.), Quaternary vertebrate paleontology in South America. Quaternary of South America and Antarctic Peninsula 12: 23-51. [ Links ]
23. Cione, A.L. y Tonni, E.P. 2005. Bioestratigrafía basada en mamíferos del Cenozoico superior de la provincia de Buenos Aires, Argentina. En: R.E. de Barrio, R.O. Etcheverry, M.F. Caballé y E. Llambías (eds.), 16º Congreso Geológico Argentino (La Plata), Relatorio, 11: 183-200. [ Links ]
24. Cladera, G., Ruigomez, E., Ortiz Jaureguizar, E., Bond, M. y López, G. 2004. Tafonomía de la Gran Hondonada (Formación Sarmiento, Edad-mamífero Mustersense, Eoceno Medio) Chubut, Argentina. Ameghiniana 41: 315-330. [ Links ]
25. Cuezva, S. y Élez, J. 2000. Estudio preliminar de la microestructura de los huesos fósiles de mamíferos de Somosaguas (Mioceno medio, Madrid). Coloquios de Paleontología 51: 137- 157. [ Links ]
26. Cuvier, G. 1796. Notice sur de squelette d´une tres grande espéce de quadrupéde inconnue jusqu´ á présent, trouvé au Paraguay, et déposé au Cabinet daHistoire Naturelle de Madrid. Magazín encyclopédique, ou Journal des Sciences, les lettres et des Arts 2: 303-310. [ Links ]
27. De Ricqlés, A., Meunier, F., Castanet, J. y Francillon-Vieillot, H. 1991. Comparative microstructure of bone. En B.K. Hall (ed.), Bone, vol. 3: Bone matrix and bone specific products. CRC Press, Boca Raton, Florida, 78 pp. [ Links ]
28. Deschamps, C. 1998. The presence of Neochoerus Hay (Rodentia, Hydrochoeridae) in Pleistocenic sediments of southwestern Buenos Aires Province. Quaternary of South America and Antarctic Peninsula 11: 1-14. [ Links ]
29. Deschamps, C. 2005. Late Cenozoic mammal bio-chronostratigraphy in southwestern Buenos Aires province, Argentina. Ameghiniana 42: 733-750. [ Links ]
30. Deschamps, C. y Borromei, A. 1992. La fauna de vertebrados pleistocénicos del Bajo San José (provincia de Buenos Aires, Argentina). Aspectos paleoambientales. Ameghiniana 29: 177- 183. [ Links ]
31. Fariña, R.A. 2002. Taphonomy and palaeoecology of the South American giant mammals. En: M. De Renzi, M. Pardo Alonso, M. Belinchón, E. Peñalver, P. Montoya y A. Márquez-Arriaga (eds.), Current Topics on Taphonomy and Fossilization, Valencia, pp. 97-113. [ Links ]
32. Fariña, R.A., Vizcaíno, S.F. y Bargo, M.S. 1998. Body mass estimations in Lujanian (late Pleistocene- early Holocene of South America) mammal megafauna. Mastozoologia Neotropical 5: 87- 108. [ Links ]
33. Fawcett, D. 1995. Tratado de histología. Interamericana, McGraw- Hill, Madrid, 1044 pp. [ Links ]
34. Fernández López, S. 2000. Temas de Tafonomía. Departamento de Paleontología, Universidad Complutense de Madrid, 167 pp. [ Links ]
35. Fiorillo, A. R. 1989. An experimental study of trampling: implication for the fossil record. En: R. Bonnichsen y M. H. Sorg (eds.), Bone Modification. Center for the Study of the First Americans, University of Maine, Orono: 61-72. [ Links ]
36. Frenguelli, J. 1928. Observaciones geológicas en la región costanera sur de la Provincia de Buenos Aires. Anales de la Facultad de Ciencias Educacionales de Paraná 2: 1-145. [ Links ]
37. Gervais, P. 1855. Recherches sur les mammifères fossiles de l´Amérique méridionale. Annales des Sciences Naturelles, Zoologie 4:: 330-338. [ Links ]
38. Gervais, P. 1867. Sur une nouvelle collection d´ossements fossils de mammifères recueillies par M. F. Seguin dans la Conféderation Argentine. Comptes Rendus de l´Académie des Sciences 65: 279-282. [ Links ]
39. Gervais, H. y Ameghino, F. 1880. Los mamíferos de la América del Sur. F. Savy-Igon Hermanos, Buenos Aires, 225 pp. [ Links ]
40. González-Bonorino, F. 1972. Introducción a la geoquímica. Programa Regional de Desarrollo Científico y Tecnológico, Departamento de Asuntos Científicos. Secretaría General de la Organización de los Estados Americanos, Washington D.C., 140 pp. [ Links ]
41. Haynes, G. 1980. Evidence of carnivore gnawing on Pleistocene and Recent mammalian bones. Paleobiology 6: 341-351. [ Links ]
42. Hill, A. 1979. Disarticulation and scattering of mammal skeletons. Paleobiology 5: 261-274. [ Links ]
43. Hoffstetter, R. 1950. Algunas observaciones sobre los caballos fósiles de América del Sur. Amerhippus gen. nov. Boletín Informaciones Científicas Nacionales 3: 426-454. [ Links ]
44. ICDD (1993). International Centre for Diffraction Data. Mineral powder diffraction file. Databook. [ Links ]
45. Iverson, R. M. 1997. The physics of debris flows. Reviews of Geophysics 35: 245-296. [ Links ]
46. Johnson, R. G. 1960. Models and methods for analysis of the mode of formation of fossil assemblages. Bulletin of the Geological Society of America 71: 1075-1086. [ Links ]
47. Kraglievich, L. 1930. La Formación Friasense del río Frías, río Fénix, Laguna Blanca, etcétera y su fauna de mamíferos. Physis 10: 127-161. [ Links ]
48. Kraglievich, L. 1934. La antigüedad pliocena de las faunas de Monte Hermoso y Chapadmalal, deducidas de su comparación con las que le precedieron y sucedieron. Imprenta "El Siglo Ilustrado", Montevideo, 136 pp. [ Links ]
49. Kraglievich, L. 1946. Resultados de una excursión a Monte Hermoso y zonas vecinas. Holmbergia, Notas geopaleontológicas 4: 197-213. [ Links ]
50. López-González, F., Grandal-d'Anglade, A. y Vidal-Romaní, J. 2005. Deciphering bone depositional sequences in caves through the study of manganese coatings. Journal of Archaeological Science 20: 1-11. [ Links ]
51. Lucas, J. y Prévôt, L. 1991. Phosphates and fossil preservation. En: P. Allison y D. Briggs (eds.), Taphonomy: Releasing the Data Locked in the Fossil Record, Plenum Press, New York, pp. 389- 409. [ Links ]
52. Lund, P.W. 1839. Blik paa Brasiliens dyreverden för sidste jordomvaeltning. Anden afhandling: Pattedyrene (Lagoa Santa d. 16 de novbr. 1837). Det kongelige Danske Videnskabernes Selskabs naturvidenskabelige og mathematiske Afhandlinger 8: 61-144. [ Links ]
53. Lund, P.W. 1840. Nouvelles recherches sur la faune fossile du Brésil. Annales des Sciences Naturelles 13: 310-319. [ Links ]
54. Lund, P.W. 1842. Blik paa Brasiliens dyreverden för sidste Jordomvaeltning. Fjerde Afhandling, Fortsaettelse af Pattedryene 9: 137-208. [ Links ]
55. Manera de Bianco, T. y Aramayo, S. 1998. Presencia de Neochoerus sp. en el Pleistoceno tardío de Pehuen Co, provincia de Buenos Aires, Argentina. 7° Congreso Argentino de Paleontología y Bioestratigrafía (Bahía Blanca), Actas: 68. [ Links ]
56. Marín Arroyo, A.B., Landete Ruiz, M.D., Vidal Bernabeu, G., Seva Román, R., González Morales, M.R. y Straus, L.G. 2008. Archaeological implications of human-derived manganese coatings: a study of blackened bones in El Mirón Cave, Cantabrian Spain. Journal of Archaeological Science 35: 801-813. [ Links ]
57. Montalvo, C. I. 2002. Taphonomic analysis of the Mio-Pliocene micromammal assemblage (Cerro Azul Formation), Caleufú, La Pampa, Argentina. En: De Renzi, Pardo Alonso, Belinchón, Peñalver, Montoya y Márquez-Arriaga (eds.), Current Topics on Taphonomy and Fossilization, Valencia: pp. 353-359. [ Links ]
58. Montalvo, C.I. 2004. [Paleobiología de la asociación faunística de Caleufú (La Pampa, Formación Cerro Azul, Mioceno superior- Plioceno inferior) a través de análisis tafonómicos. Tesis doctoral N° 830, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, 251 pp. Inédita. [ Links ]].
59. Montalvo, C., Melchor, R., Visconti, G. y Cerdeño, E. 2008. Vertebrate taphonomy in loess-palaeosol deposits: A case study from the late Miocene of central Argentina. Geobios 41: 133-143. [ Links ]
60. Olsen, S.L. y Shipman, P. 1988. Surface modification on bone: trampling versus butchery. Journal of Archaeological Science 15: 535-553. [ Links ]
61. Owen, R. 1837. A description of the cranium of the Toxodon platensis, a gigantic extinct mammiferous species, referable by its dentition to the Rodentia, but with affinities to the Pachydermata and the herbivorous Cetacea. Geological Society of London, Proceedings 2: 541-542. [ Links ]
62. Owen, R. 1838a. Note of the Glyptodon. En: W. Parrish (ed.), Buenos Aires and the provinces of Río de la Plata, 1-178 pp. [ Links ]
63. Owen, R. 1838b. Fossil Mammalia (1). En: C.R. Darwin (ed.), Zoology of the Voyage of H.M.S Beagle, under the command of Captain Fitz-roy, during the years 1832 to 1836, 1: 13-40. [ Links ]
64. Owen, R. 1839. Fossil Mammalia (2). En: C.R. Darwin (ed.), Zoology of the Voyage of H.M.S Beagle, under the command of Captain Fitz-roy, during the years 1832 to 1836, 1: 41-64. [ Links ]
65. Owen, R. 1845. Account of various portions of the Glyptodon, an extinct quadruped, allied to the armadillo, and recently obtained from the Tertiary deposits in the neighbourhood of Buenos Aires. Quaterly Journal of the Geological Society of London 1: 257-262. [ Links ]
66. Owen, R. 1847. Notice of some fossil Mammalia of South America. Report of the 16º Meeting of the British Association for the Advancement of Science (Southampton): 65-67. [ Links ]
67. Owen, R. 1869. On fossil remains of Equines from Central and South America referable to Equus conversidens, Ow., Equus tau, Ow., and Equus arcidens, Ow. Philosophical Transactions of the Royal Society of London 159: 559-573. [ Links ]
68. Paniagua, R. 1996. Citología e histología vegetal y animal. Biología de las células y tejidos animales y vegetales. Interamericana McGraw-Hill, Madrid, 807 pp. [ Links ]
69. Pardiñas, U. y Deschamps, C. 1996. Sigmodontinos (Mammalia, Rodentia) pleistocénicos del sudoeste de la provincia de Buenos Aires (Argentina): aspectos sistemáticos, paleozoogeográficos y paleoambientales. Estudios Geológicos 52: 367-379. [ Links ]
70. Paula Couto, C. 1957. Sôbre um gliptodonte do Brasil. Boletin Divisão de Geologia y Mineralogia 165: 1-37. [ Links ]
71. Pohlig, H. 1912. Sur une vieille mandibule de "Tetracaulodon ohioticum" Blum., avec défense in situ. Bulletin de la Société Belge de Géologie 26: 187-193. [ Links ]
72. Politis, G.G. y Messineo, P.G. 2008. The Campo Laborde site: new evidence for the Holocene survival of Pleistocene megafauna in the Argentine Pampas. Quaternary Internacional 191: 98-114. [ Links ]
73. Polonio, I. y López-Martínez, N. 2000. Análisis tafonómico de los yacimientos de Somosaguas (Mioceno medio, Madrid). Coloquios de Paleontología 51: 235-265. [ Links ]
74. Pomi, L. 2006. Tafonomía en el Miembro Guerrero de la Formación Luján (Pleistoceno tardío). Meteorización ósea. Ameghiniana, Suplemento Resúmenes 43: 53R. [ Links ]
75. Pomi, L.H. 2009. Entrampamientos en la Formación Luján (Pleistoceno tardío). Análisis tafonómico de un espécimen de Hippidion Owen de la cuenca del río Salado (provincia de Buenos Aires, Argentina. Ameghiniana 46: 17-25. [ Links ]
76. Reineck, H. y Singh, I. 1980. Depositional sedimentary environments, with reference to terrigenous clastics. Springer-Verlag, Berlin, Heidelberg, New York, 549 pp. [ Links ]
77. Smith, R. 1993. Vertebrate taphonomy of Late Permian floodplain deposits in the Southwestern Karoo Basin of South Africa. Palaios 8: 45-67. [ Links ]
78. Straight, W. y Eberth, D. 2002. Testing the utility of vertebrate remains in recognizing patterns in fluvial deposits: an example from the Lower Horseshoe Canyon Formation, Alberta. Palaios 17: 472-490. [ Links ]
79. Tauber, A. A. 1997a. Paleoecología de la Formación Santa Cruz (Mioceno Inferior) en el extremo sudeste de la Patagonia. Ameghiniana 34: 517-529. [ Links ]
80. Tauber, A. A. 1997b. Bioestratigrafía de la Formación Santa Cruz (Mioceno Inferior) en el extremo sudeste de la Patagonia. Ameghiniana 34: 413-426. [ Links ]
81. Todd, L. C. y Frison, G. C. 1986. Taphonomic Study of the Colby Site Mammoth Bones. En: G.C. Frison y L.C. Todd (eds.), The Colby Mammoth Site. Taphonomy and Archaeology of a Clovis Kill in Northern Wyoming, University of New Mexico Press, Albuquerque, pp. 27-90. [ Links ]
82. Tomassini, R., Manera de Bianco, T. y Oliva, C. 2006. Estudio tafonómico de los vertebrados del Pleistoceno de Playa del Barco, Pehuen Co, provincia de Buenos Aires, Argentina. 9º Congreso Argentino de Paleontología y Bioestratigrafía (Córdoba), Resúmenes: 272. [ Links ]
83. Trapani, J. 1998. Hydrodynamic sorting of avian skeletal remains. Journal of Archaeological Science 25: 477-487. [ Links ]
84. Vega, V., Rodríguez, S. y Valente, M. 1989. Shallow marine and fluvial enviroments of Quaternary deposits in Pehuén Co Beach, Buenos Aires, Argentina. Quaternary of South America and Antarctic Peninsula 7: 51-80. [ Links ]
85. Verzi, D., Deschamps, C. y Tonni, E. 2004. Biostratigraphic and palaeoclimatic meaning of the Middle Pleistocene South American rodent Ctenomys kraglievichi (Caviomorpha, Octodontidae). Palaeogeography, Palaeoclimatology, Palaeoecology 212: 315- 329. [ Links ]
86. Voorhies, M. 1969. Taphonomy and population dynamics of an Early Pliocene vertebrate fauna, Knox County, Nebraska. Contributions to geology, Special Papers 1: 1-69 pp. [ Links ]
87. Zavala, C. y Quattrocchio, M. 2001. Estratigrafía y evolución geológica del río Sauce Grande (Cuaternario), provincia de Buenos Aires, Argentina. Revista de la Asociación Geológica Argentina 56: 25-37. [ Links ]
Recibido: 2 de marzo de 2009.
Aceptado: 22 de septiembre de 2009.

