










Serviços Personalizados
Journal

Artigo
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAmeghiniana
versão On-line ISSN 1851-8044
Ameghiniana vol.47 no.3 Buenos Aires set. 2010
ARTÍCULOS ORIGINALES
Tooth Root Size, Chewing Muscle Leverage, and the Biology of Homunculus patagonicus (Primates) from the late early Miocene of Patagonia
Jonathan M. G. Perry1, Richard F. Kay2, Sergio F. Vizcaíno3, and M. Susana Bargo3
1Department of Anatomy, Midwestern University, 555 31st St., Downers Grove, IL, 60515, ph: (630) 515-7215, jperry@midwestern.edu
2Department of Evolutionary Anthropology, Box 90383, Biological Sciences Bldg., Science Drive, Duke University, Durham, NC, U.S.A, 27708-0383, Richard. kay@duke.edu
3División Paleontología de Vertebrados, Museo de La Plata, Paseo del Bosque s/n, B1900FWA La Plata, Argentina. CONICET-CIC. vizcaino@fcnym.unlp.edu.ar, msbargo@fcnym.unlp.edu.ar
Abstract. Inferences about the diet of Miocene platyrrhine monkeys have relied upon the morphology of the molar teeth, specifically the crests on the molars. Using a library of Micro-CT images of a broad comparative sample of living platyrrhines (callitrichines, cebines, pitheciids and atelids), late early Miocene Homunculus, and the early Miocene Tremacebus and Dolichocebus, we extend these inferences by examining the surface areas of the tooth roots, anchor points for the periodontal ligaments. From muscle scars on the skull, we estimate the mechanical leverage of the chewing muscles at bite points from the canine to the last molar. Extant platyrrhines that gouge bark to obtain exudates do not have especially large canine roots or anterior premolar roots compared with their less specialized close relatives. Extant platyrrhines that have more folivorous diets have much larger molar roots than do similar-sized more frugivorous species. Homunculus patagonicus has large postcanine roots relative to body size and poor masticatory leverage compared to the extant platyrrhines in our sample. The large postcanine roots, heavy tooth wear, and moderately-long shearing crests suggests a diet of abrasive, resistant foods. However, relatively poor jaw adductor leverage would have put the masticatory apparatus of Homunculus at a mechanical disadvantage for producing high bite forces compared to the condition in extant platyrrhines. Tremacebus and Dolichocebus, like Homunculus, have larger tooth root surfaces than comparable-sized living platyrrhines. They also resemble Homunculus in being more prognathic and having posteriorly-located temporalis origins - all features of a relatively poor leverage system.
Resumen. Tamaño de las raíces dentarias, brazos de palanca de la musculatura masticatoria y biología de Homunculus patagonicus (Primates) del Mioceno temprano de Patagonia. Las inferencias sobre dieta de monos platirrinos miocenos se han basado en la morfología de los molares, específicamente las crestas. Aquí se estudia la superficie de las raíces de los dientes, área de anclaje del ligamento periodontal. Se utilizó una base de datos de imágenes Micro-CT de platirrinos actuales (calitriquinos, cebinos, pitécidos y atélidos), Tremacebus y Dolichocebus (Mioceno temprano) y Homunculus patagonicus (Mioceno temprano tardío). Basándose en las cicatrices musculares preservadas en los cráneos, se estimó el brazo de palanca de los músculos masticatorios en puntos de mordida desde el canino hasta el último molar. En los platirrinos actuales que se alimentan de exudados, las raíces de los caninos o premolares anteriores no son especialmente grandes; en los folívoros, las raíces de los molares son mucho más grandes que en los frugívoros de tamaño similar. Las raíces de los postcaninos de H. patagonicus son más grandes en relación a su tamaño corporal que en cualquier otro platirrino viviente analizado; las raíces postcaninas grandes, el excesivo desgaste de los dientes y las crestas cortantes moderadamente largas sugieren un consumo de alimentos abrasivos y resistentes. Sin embargo, la palanca aductora relativamente pobre de la mandíbula indica una desventaja mecánica para producir fuerzas de mordida elevadas en comparación con platirrinos actuales. Tremacebus y Dolichocebus también poseen superficies de las raíces más grandes que los platirrinos vivientes de tamaño equivalente y se asemejan a Homunculus por ser más prognatos y por el origen del músculo temporal ubicado posteriormente, rasgos que indican un sistema de palancas relativamente pobre.
Key words. Primates; Platyrrhini; Homunculus; Miocene; Teeth; Diet.
Palabras clave. Primates; Platyrrhini; Homunculus; Mioceno; Dientes; Dieta.
Introduction
Platyrrhines, or New World monkeys, are a diverse radiation of primates that includes a wide range of body size and dietary preferences (Ford and Davis, 1992). Today, more than 70 platyrrhine species are recognized, belonging to three families (Schneider, 2000). In contrast, fossil platyrrhines are rare and fragmentary (MacFadden, 1990; Fleagle and Kay, 1997) and inferences about their paleobiology are largely limited to dental remains (Kay et al., 2002), although a modest record of postcrania has allowed for locomotor inference in some species (Ford, 1990; Meldrum, 1993).
Primates first appear in the fossil record of South America in the late Oligocene (Deseadan), represented by teeth and jaws of the species Branisella boliviana Hoffstetter, 1969 from a single stratigraphic level at Salla, Bolivia (Hoffstetter, 1969; Wolff, 1984; Rosenberger et al., 1991; Takai and Anaya, 1996; Kay et al., 2002). All the other Tertiary records of platyrrhines are from the Miocene. These are from sites in Argentina, Colombia, Chile, and Brazil, with fossil primates from five Land Mammal Ages-Colhue-huapian, Santacrucian, Colloncuran, Laventan, and Huayquerian (Rusconi, 1935; Hershkovitz, 1970; Fleagle and Kay, 1989; Kay and Frailey, 1993; Flynn et al., 1995; Fleagle et al., 1997; Fleagle and Tejedor, 2002; Tejedor, 2005a, 2005b; Kay and Cozzuol, 2006; Tejedor et al., 2006; Kay et al., 2008a; Tejedor, 2008).
Homunculus patagonicus Ameghino, 1891 is the best-known and best-preserved fossil primate from the Tertiary of South America. It comes from the late early Miocene Santa Cruz Formation of Patagonia. Homunculus fossils come from a series of localities along the Atlantic coast of Santa Cruz Province and along the estuary of the Río Gallegos, Argentina (Tauber, 1991; Kay et al., 2008b). The extraordinary completeness of the cranial specimens allows us an exceptional opportunity to reconstruct aspects of cranial functional morphology that are unavailable for other Tertiary platyrrhines.
Florentino Ameghino described several specimens now considered to belong to Homunculus patagonicus, including relatively complete mandibles, fragments of long bones, and a partial cranium (Ameghino, 1891; Bluntschli, 1931; Rusconi, 1935). Several decades later, another partial cranium (CORD-PZ 1130) was discovered and described by Tauber (1991). Since 2003, a yearly joint U.S.-Argentina field expedition has recovered four additional partial crania - including two that are nearly complete, a partial mandible, and additional long bone fragments from localities along the coast (Estancia La Costa, Puesto Estancia La Costa = Corrigüen Aike; Marshall, 1976) and at Killik Aike Norte on the north shore of the estuary of the Río Gallegos. An additional monkey cranium preserving the face and orbits from Killik Aike Norte was described in 2006 as a new genus and species, Killikaike blakei Tejedor et al., 2006. This specimen is insufficiently complete for inclusion in our biomechanical analysis.
To date, most of the work on Homunculus has focused on its phylogeny and taxonomy (Bluntschli, 1931; Stirton, 1951; Hershkovitz, 1970, 1974; Rosenberger, 1978; Hershkovitz, 1981, 1984; Rosenberger et al., 1990; Zhang et al., 2000; Kay et al., 2005; Tejedor and Rosenberger, 2008). Studies of cranial functional morphology are limited to an analysis of the molar structure (Fleagle et al., 1997) and remarks on masticatory function in two papers by Tauber (1991, 1999) and one by Tejedor and Rosenberger (2008).
Molar crown morphology is correlated with diet in primates (Butler, 1973; Kay, 1975, 1977; Corruccini and Beecher, 1982; Gordon, 1982; Janis, 1984; Teaford and Walker, 1984; Covert, 1986; Kinzey, 1992; Rosenberger, 1992; Strait, 1993; Kay et al., 2004; Lucas, 2004). Dental morphology, especially the small incisors and moderately developed shearing crests on the cheek teeth suggest that Homunculus was a mixed frugivore-folivore (Fleagle and Kay, 1997; Fleagle et al., 1997). An abrasive diet is inferred because most specimens of Homunculus have very high degrees of molar wear (figure 1) (Tauber, 1999). Extant frugivorous platyrrhines seldom show such a high degree of molar wear (Kay et al., 2002).
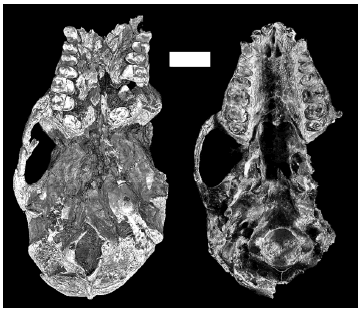
Figure 1. Ventral views of Homunculus patagonicus specimens MPM-PV 3501 (left) and MPM-PV 3502 (right) / vistas ventrales de los especímenes de Homunculus patagonicus MPM-PV 3501 (izquierda) y MPM-PV 3502 (derecha). The picture at left is a volume reconstructed from CT scans / la imagen de la izquierda está reconstruida a partir de tomografías computadas. The picture at right is a photograph of the original specimen / la imagen de la derecha es una foto del espécimen original. Notice the heavy tooth wear on both specimens / nótese el excesivo desgaste dentario de ambos especímenes. Scale bar = 1 cm / escala = 1 cm.
Tauber (1991) suggested, based on the surface rugosity and sharp delineation of attachments for the temporalis and buccinator muscles, that this extinct platyrrhine had a very powerful masticatory system - more so than in extant pitheciines and more so than in early Miocene Dolichocebus gaimanensis (Kraglievich, 1951) and Tremacebus harringtoni (Rusconi, 1933).
Tejedor and Rosenberger (2008) compared Homunculus with pitheciines. They suggested that the morphology of the anterior dentition and the shape of the mandibular corpus in the former could be a precursor to the derived condition seen in modern pitheciines. These authors did not venture ecological inferences for Homunculus beyond the comparison to pitheciines. Rosenberger et al. (2009) summarized a view of the dietary adaptations of Homunculus based on the teeth. They suggested that the prognathic incisors and premolar morphology indicate hard-fruit husking. They further note that Homunculus has large cheek teeth with heavy wear and well-developed molar shearing crests, suggesting some folivory, at least seasonally.
New cranial material for Homunculus permits us to refine the observations of Tauber (1991, 1999) and those of Tejedor and Rosenberger (2008) and Rosenberger et al. (2009). While it is impossible to evaluate the forcefulness of the chewing muscles directly in a fossil, it is possible to evaluate whether or not its skull geometry was especially well-suited to bite-force generation and absorption. Because several crania and mandibles of Homunculus are preserved, we can analyze muscle leverage in a composite specimen. Unfortunately, it is not possible to perform a detailed analysis on Tremacebus or Dolichocebus because known material is insufficient: an intact lower jaw has not been recovered for either species. However, we can offer some observations, based on the available cranial morphology.
Tooth root size may be a useful indicator of chewing forces (Wood et al., 1988; Spencer, 2003; Kupczik and Dean, 2007). Therefore, it may be a valuable adjunct in reconstructing the diets of extinct species, particularly when tooth crowns are heavily worn or broken away. Bite force is transmitted through the tooth roots. The periodontal ligament is composed of Sharpie's fibers that anchor the root to the surrounding alveolar bone and act to resist root displacement (Beertsen et al., 1997). A root that has a greater surface area can potentially accommodate more Sharpie's fibers, is likely to be more stable within its alveolus, and is better equipped to resist heavy or repetitive incisive or masticatory loads.
The goal of this study is to evaluate the hypothesis that the masticatory system of Homunculus was adapted for producing considerable bite force (Tauber, 1991, 1999) and to offer inferences about the properties of the foods in the diet of Homunculus based on masticatory biomechanics and tooth root size. We measured tooth root areas in Homunculus to test the hypothesis that this Miocene primate was eating extremely resistant foods that may have contributed to its high degree of molar wear. We also offer observations on Tremacebus and Dolichocebus where pertinent. These analyses are comparative, using a sample of extant platyrrhines with known diets.
Based on the anatomy of living platyrrhines we can evaluate several functional possibilities relating diet and tooth root areas. We assume that repeated loading cycles should favor large tooth root areas where the load occurs. There should also be systematic variation in the distribution of root area in relation to where chewing forces are generated. Canine roots are expected to be larger in taxa that process primarily fruits and nuts as these species often perform a lot of forceful food processing with the canines. The hypothesized relationship between canine size and fruit consumption is potentially confounded by several factors: 1) Many species of platyrrhines exhibit canine sexual dimorphism but there is little evidence that diet or food acquisition behavior differs between males and females. 2) Some platyrrhines (e.g., Chiropotes) use their canines to pry open the husks of hard-shelled fruits to gain access to seeds; others (e.g., Saguinus) ingest soft ripe fruits taking few or no bites, thereby swallowing the seeds whole (Kinzey and Norconk, 1990). The former would be expected to have large canines whilst the latter may not. Folivores, by contrast, are expected to have larger roots on more posteriorly-positioned cheek teeth than on anterior teeth because most leaf-processing occurs posteriorly and should involve repeated chewing cycles.
Within our sample of extant platyrrhines, two comparisons of root area are particularly interesting: that between two atelids, Alouatta guariba and Ateles geoffroyi, and that between two callitrichine cebids Callithrix jacchus and Saguinus fuscicollis. In each of these pairs, there are differences in diet and/or food procurement. Alouatta guariba consumes a diet rich in leaves, immature leaves are preferred but mature ones are consumed in times of need, and only a small percentage of fruit is consumed (Chiarello, 1994). By contrast, Ateles geoffroyi consumes primarily ripe fruit and does not split open the seeds contained within the fruits (Cant, 1990; Chapman and Chapman, 1991; Strier, 1992). We would expect that postcanine tooth roots have larger areas in Alouatta than in Ateles due to the need for masticating tough leaves in Alouatta. We predict that canine root area (and incisor root area, for which we have no data) might be greater in Ateles due to the need for considerable anterior tooth use in processing fruits, but with the caveat that such differences may be masked by sexual selection and also depend on the manner in which the fruit is consumed (see footnote).
Callithrix jacchus uses its lower incisors and incisi-form lower canines to scrape and gouge tree bark in search of plant exudates, whereas Saguinus fuscicollis does not use its anterior teeth in this way (Sussman and Kinzey, 1984; Ferrari, 1993). We predict that anterior tooth roots (i.e., the canine root) will have a larger surface area in Callithrix to accommodate forceful gouging. Vinyard and colleagues found that gouging in these callitrichids is not associated with large forces (Vinyard et al., 2001) or with especially strong jaw adductor muscles (Taylor and Vinyard, 2004). Therefore, if our prediction is not met, it may be because gouging does not require large forces and this may be reflected in tooth root area.
A final prediction stems from ecological studies that suggest that Pithecia masticates extremely resistant seeds (Kinzey and Norconk, 1990; Kinzey, 1992). We predict that tooth root area in Pithecia will be great relative to the other extant species in our sample.
Material. Acronyms: CORD, Universidad Nacional de Córdoba, Córdoba, Argentina; NMNH, National Museum of Natural History, Smithsonian Institution, Washington D.C., U.S.A.; MACN, Museo Argentino de Ciencias Naturales "Bernardino Rivadavia", Buenos Aires, Argentina; MPM, Museo Regional Provincial "Padre M.J.Molina", Río Gallegos, Argentina.
All measurements were taken from CT scans of skulls of extant platyrrhines and crania of extinct platyrrhines. A single specimen was scanned for each species; all are museum specimens.
We collected data on nine extant platyrrhine species: Alouatta guariba, Ateles geoffroyi, Callithrix jacchus, Cebus nigrivittatus, Cebuella pygmaea, Callicebus torquatus, Mico argentatus, Pithecia sp., and Saguinus fuscicollis (appendix 1).
We collected data on three extinct platyrrhine species: Homunculus patagonicus, Dolichocebus gaimanensis, and Tremacebus harringtoni (appendix 1). Root areas were measured on two specimens of Homunculus (MPM-PV 3501 and 3502), but data for MPM-PV 3501 were omitted from the analysis because many of the tooth roots in this specimen are damaged. To estimate masticatory leverage in Homunculus, it was necessary to use a scaled composite of three specimens: one cranium (MPM-PV 3502), and two fragmentary mandibles (MACN A 5969 and MPM-PV 3504) to perform this analysis on Homunculus patagonicus (figure 2). We discuss below our reasons for allocating MPM-PV 3502 to H. patagonicus (appendix 2).

Figure 2. Composite skull of Homunculus patagonicus / cráneo compuesto de Homunculus patagonicus. The cranium is a CT image of MPM-PV 3502 / el cráneo es una imagen de tomografía computada del MPM-PV 3502. The anterior part of the lower jaw is MPM-PV 3504 and the posterior part is a cast of MACN A 5969 / la parte anterior de la mandíbula pertenece a MPM-PV 3504 y la parte posterior es un calco del MACN A 5969. Photographs of these specimens were scaled so that the maxillary teeth would occlude with the mandibular ones / Las fotografías de estos especímenes fueron llevadas a la misma escala de modo que los dientes maxilares ocluyan con los mandibulares. MACN A 5969 has been flipped horizontally (the specimen is a fragment of a left mandible) / MACN A 5969 ha sido reflejada horizontalmente (el espécimen es un fragmento de mandíbula izquierda). It was necessary to reconstruct the posterior part of the mandibular angle to estimate the insertion areas for the masseter and medial pterygoid, and the coronoid process to estimate the insertion area for the temporalis / fue necesario reconstruir la parte posterior del ángulo mandibular para estimar el área de inserción del masetero y del pterigoides medio y el proceso coronoides para estimar el área de inserción del temporal. Scale bar = 1 cm / escala = 1 cm.
MPM-PV 3502 is from Killik Aike Norte, Argentina, and is the only relatively complete and undistorted cranial specimen for the species. We took representative measurements on a second, somewhat distorted skull (MPM-PV 3501) from Puesto Estancia La Costa, to confirm that the patterns we saw in root size for MPM-PV 3502 (see below - Results) were also present in other specimens of Homunculus. Despite the presence of another primate species of similar size from Killik Aike Norte, Killikaike blakei (Tejedor et al., 2006), we allocate MPM-PV 3502 to H. patagonicus because it strongly resembles specimens of the latter from other localities (see Discussion).
The mandibular specimens used in our reconstruction come from Estancia La Costa (MPM-PV 3504) and Monte Observación (now Cerro Observatorio, Marshall, 1976) (MACN A-5969). MPM-PV 3504 is very similar in all comparable respects to the neotype designated for Homunculus patagonicus, also from Corriguen Aike (Tejedor and Rosenberger, 2008). The analyses of Tremacebus and Dolichocebus are based on the type specimens of those taxa.
Methods
Skulls were scanned in the high-resolution X-ray CT facility at the University of Texas (Austin). Slice thickness varied between 0.04 mm and 0.1 mm for specimens of different size. Stacks of 8-bit jpg files were assembled in VGStudio Max 1.2 (Volume Graphics, Heidelberg, Germany). Lateral views of three-dimensional renderings of skull surfaces were used for orientation. Tooth root area measurements were taken from volume renderings of the tooth roots and leverage measurements were taken as two-dimensional lateral projections using the skull surface rendering, employing the "non-perspective" setting. Measurements were calibrated using caliper measurements of the original skulls and using the associated CT scan data.
Roots
We measured root area of all maxillary teeth from the canine to the most posterior molar from CT scans. We used the segmentation function in VGStudio Max 1.2 to outline the roots in three dimensions. Outlines of roots were selected on each slice (slice thickness = 0.05-0.01 mm) and the entire root segment was compiled at the end to yield root area. The boundary between root and crown was estimated using the base of the enamel cap; the enamel of the tooth crown is distinguishable in CT scans based on its density.
Following Spencer (1999, 2003), tooth root areas were scaled to a geometric mean of five cranial measurements (see table 1). These measurements are all taken from the face; therefore, any possible differences in brain size between fossil and extant platyrrhines would not affect this geometric mean. Spencer also scaled root area to crown area; however, crown area itself varies with diet: folivores and insectivores, in general, have larger tooth crowns than do frugivores (Kay, 1975; Anthony and Kay, 1993). Therefore, we did not use crown area as a scaling variable.
Table 1. Measurements used as a proxy for body size (mm) / medidas utilizadas como indicadores para tamaño corporal (mm). BB, bicondylar breadth / ancho bicondilar; PB, palate breadth / ancho palatal; TFL, temporal foramen length / largo del foramen temporal; TRL, tooth row length / largo de la serie dentaria; IH, infratemporal height / altura infratemporal; GM, geometric mean of the five measurements above / media geométrica de las cinco medidas mencionadas arriba. See Spencer (1999) for explanations of measurements / ver Spencer (1999) para una explicación sobre las medidas. For all specimens, TRL was measured from the P2 to the posterior-most molar, whether the M2 or the M3 / para todos los especímenes, TRL fue tomada desde P2 al molar más posterior. See the appendix for abbreviations of species names and specimen numbers / ver apéndice para las abreviaturas de las especies y números de especímenes.

All statistical analyses were performed in JMP 7.0 (JMP, Version 7. SAS Institute Inc., Cary, NC, 1989-2007).
Chewing force: muscle dimensions and leverage
The physiological cross-sectional area (PCSA) of a muscle gives an estimate the amount of force it is capable of producing (Weijs and Hillen, 1985). PCSA is an estimate of the amount of muscle that contracts in parallel to generate a load, and can be calculated from preserved muscles (Perry and Wall, 2008). PC-SA indicates the potential force of a muscle; the actual force depends on many other variables, including activity patterns (Hylander et al., 2000). Despite the availability of data on chewing muscle PCSAs for a few species of extant platyrrhines (Schumacher, 1961; Anapol et al., 2008), we do not know enough about the anatomical correlates of PCSA in platyrrhines (e.g., attachment scars on the skull) to reconstruct chewing muscle PCSA in fossil platyrrhines.
In the absence of directly measured chewing muscle cross-sectional areas (as is true for all fossil species), the best approximation of muscle force is estimated muscle leverage. Chewing muscle leverage can be estimated from the centroids of attachment areas of the muscles when they are visible on the bony specimens. By comparing the leverages of the chewing muscles across species, one can gauge the relative effectiveness of the jaw adductor muscles. To estimate jaw adductor leverage for the extant platyrrhines and for Homunculus we took the following steps:
- Based on the muscle scars, we traced the estimated areas of origin and insertion of the temporalis, masseter, and medial pterygoid muscles onto two-dimensional lateral images of skulls that had been generated from CT volumes. Published descriptions of dissections of platyrrhine chewing muscles (Stark, 1933; Schumacher, 1961; Schön, 1968; Madeira and de Oliveira, 1979) and a dissection of the chewing muscles in Cebuella pygmaea (by JMGP) guided this procedure.
- Using the software ImageJ (United States National Institutes of Health, public domain), we calculated centroids for all areas of attachment. We traced a line of action for each muscle by joining the centroid of the origin area to the centroid of the insertion area. The lever arm for each muscle is a perpendicular line drawn from the line of action to the center of the mandibular condyle. By convention, moments were taken about the mandibular condyle; no joint reaction force was calculated (Stern, 1974; Tattersall, 1974; Hylander, 1975; Demes and Creel, 1988; Vizcaíno et al., 1998).
- Onto the same images, we drew load arms parallel to the occlusal plane (that is, normal to an assumed vertical bite force, see figure 3) (Vizcaíno and Bargo, 1998; Vizcaíno et al., 1998; De Iuliis et al., 2000; Bargo, 2003; Vizcaíno et al., 2006). The distance from the condyle to bite points on the upper teeth, measured parallel to the occlusal plane, was used to estimate the length of the load arm for those bite points. The tip of the paracone was selected as the bite point for each postcanine tooth.
- The leverage for a given muscle at a given bite point was determined by dividing the lever arm length of the muscle by the load arm length at the bite point.

Figure 3. Reconstructed CT image of the skull of a female Mico argentatus (NMNH 238463) in lateral view / imagen reconstruida de tomografía computada del cráneo de una hembra de Mico argentatus (NMNH 238463) en vista lateral. Each small black dot is the centroid for an area of attachment of the masseter, and the white line connecting the small black dots is the hypothetical line of action of the muscle / cada punto negro pequeño es el centroide del área de inserción del masetero y la línea blanca que conecta los puntos negros es la línea hipotética de acción del músculo. The large black dot represents a center of rotation (at the condyle), and the black line joining the condyle to the line of action is the lever arm of the masseter / el punto negro grande representa el centro de rotación (en el cóndilo) y la línea negra que une el cóndilo con la línea de acción representa el brazo de momento del masetero. Two white lines are drawn perpendicular to the occlusal plane: one through the paracone of the M2, another through the apex of the canine / se dibujan dos líneas blancas perpendiculares al plano occlusal: una a través del paracono del M2, la otra a través del ápice del canino. A black line drawn from the condyle to each of these lines is the load arm for each bite point / la línea negra trazada desde el cóndilo a cada una de estas líneas es el brazo de carga para cada punto de mordida. The length of the lever arm divided by the length of the load arm at a particular point is the leverage of the masseter for that bite point in a vertical bite / el largo del brazo de palanca dividido por el largo del brazo de carga a un punto particular es el brazo de palanca del masetero para ese punto de mordida en una mordida vertical. Scale bar = 1 cm / escala = 1 cm.
Results
Roots
Root surfaces as reconstructed in Homunculus MPM-PV 3502 are illustrated in figure 4. Using the least squares model, log tooth root area of the canine and cheek teeth was regressed against log geometric mean of the five cranial measurements for all specimens in the sample (table 2, figures 5 and 6).

Figure 4. Comparison of tooth roots in Homunculus (above) and Pithecia (below) / comparación de las raíces dentarias de Homunculus (arriba) y Pithecia (abajo). Scale bar = 1 mm / escala = 1 mm.
Table 2. Descriptive statistics for LS regressions against the cranial geometric mean / estadísticos descriptivos para las regresiones LS contra las medias geométricas del cráneo. RA, root area (mm2) / RA, area de la raíz (mm2); GM, cranial geometric mean (mm) / GM, media geométrica del cráneo (mm). All data were logged (base ten) prior to regression analyses / todos los datos fueron transformados a logaritmo en base 10 antes del análisis de regresión. Expected slope for regressions is 2 / pendiente esperada para la regresión es 2. For the M3, n = 7; in all other cases, n = 12 / para el M3, n = 7; en todos los otros casos, n= 12. The sum is composed of root areas C-M2 / la suma está compuesta por las áreas de las raíces C-M2.
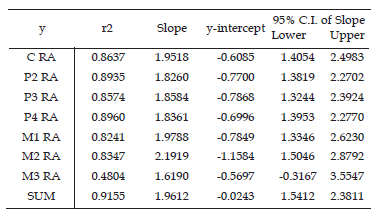
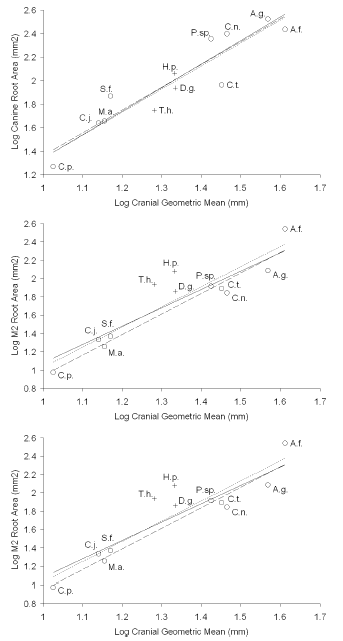
Figure 5. Tooth root size / tamaño de la raíz dentaria. Least squares (LS) regressions of root area versus the geometric mean (GM) of five cranial measurements, in log space / regresiones de cuadrados mínimos del área de la raíz dentaria versus la media geométrica de cinco medidas del cráneo, transformadas a logaritmo. Raw data for root area are in mm2 and for the geometric mean are in mm / unidades del área de la raíz en mm2 y de la media geométrica en mm. The theoretical slope of isometry is 2.0 as indicated by the solid line, while the regression line is shown by the dotted line / la pendiente esperada para isometría es 2.0 como lo indica la línea continua, mientras que la línea de regresión se indica en línea punteada. The dashed line is fitted to only extant taxa / la línea entrecortada es la recta de regresión utilizando sólo los taxones actuales. Abbreviations for species names as in table 1 / abreviaturas de las especies como en la tabla 1. Descriptive statistics are listed in table 2 / estadísticos descriptivos listados en la tabla 2. Extinct species are marked with crosses / las especies extintas están indicadas con cruces. 1, canine root area / área de la raíz del canino, 2, M2 root area / área de la raíz del M2, 3, sum of root areas C to M2 / suma de las áreas de las raíces del C al M2.
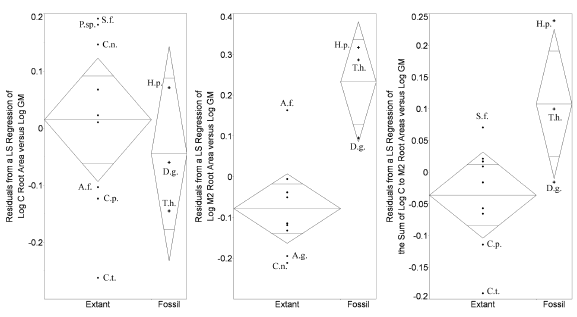
Figure 6. Comparison of tooth root area residuals for extant and extinct taxa / comparación de los residuos de las áreas de las raíces dentarias para los taxones vivientes y extintos. Plots showing residuals from least squares regressions of log root area versus log geometric mean / diagramas de los residuos de las regresiones de cuadrados mínimos de los log del área de la raíz versus el log de la media geométrica. Abbreviations for species as in table 1 / abreviaturas de las especies como en tabla 1. Extinct species are marked with crosses / las especies extintas están indicadas con cruces. The middle horizontal line depicts the group mean and the vertical tips of the diamonds represent the 95% confidence limits / la línea horizontal representa la media del grupo y los extremos verticales de los diamantes los límites de confianza de 95 %. 1, canine / canino, 2, M2 / M2, 3, sum of C-M2 / suma de C-M2. Among these comparisons, the difference between fossil and extant forms is significant only for M2 (p=0.02, Wilcoxon 1-way test), but approaches significance for the sum of C-M2 (p=0.079) / entre estas comparaciones, la diferencia entre las formas fósiles y vivientes es significativa sólo para el M2 (p=0.02, Wilcoxon 1-way test), pero la suma de C-M2 (p=0.079) está próxima a un valor significativo. Homunculus lies beyond the confidence interval for extant primates for all tooth positions, except the canine / Homunculus cae fuera del intervalo de confianza de los primates vivientes para todas las piezas dentarias, excepto el canino.
Root area scales isometrically to the geometric mean for all teeth; that is, the 95% confidence interval for the slope encompasses the slope that is expected by isometry (i.e., a slope of 2.0).
In all cases among platyrrhines, canine root area is greater than that of any premolar (table 3). However, canine root area is often less than that of the M1 and M2. There is no consistent trend in root area across the premolars. In all but one case, M1 root area > M2 root area > M3 root area. The exception is Alouatta in which M2 > M1> M3.
Table 3. Root areas (mm2) for all teeth examined / areas de las raíces (mm2) de todos los dientes examinados. Abbreviations for species names as in table 1 / abreviaturas de las especies como en la tabla 1. The M3's are broken away in Dolichocebus / los M3 están rotos en Dolichocebus.

Callithrix jacchus and Saguinus fuscicollis are similar-sized callitrichines; in the former, tree exudates form a much larger part of the diet than in the latter (Coimbra-Filho and Mittermeier, 1976). Callithrix jacchus generally has larger tooth root areas at each tooth position (relative to the geometric mean) thandoes Saguinus fuscicollis. This difference is most pronounced in the most posterior teeth, whereas canine root area is larger in Saguinus.
Ateles geoffroyi and Alouatta guariba are similarly-sized atelids; the former is mainly frugivorous, where-as the latter is mainly folivorous (Schoeninger et al., 1997). Relative to the geometric mean, canine root area is greater in Ateles than in Alouatta; however, for all other tooth positions, root area is greater in Alouatta than in Ateles. We used female specimens for both species.
Total C-M2 root area relative to the geometric mean in Homunculus patagonicus is greater than in any other platyrrhine in this sample (figures 5.3, 6.3). The difference is especially pronounced for the molars (figures 5.2, 6.2). The only exception is the canine root, which is small compared to some extant platyrrhines (e.g., Saguinus).
Tremacebus resembles Homunculus in having relatively larger M2 roots than any living platyrrhine in our sample (figures 5.2, 6.2). Dolichocebus also follows this pattern: its M2 roots are larger than those of any extant platyrrhine in our sample, with the exception of Alouatta guariba. Total C-M2 root size of Tremacebus and Homunculus likewise exceed those of extant platyrrhines (figures 5.1, 6.1) but that of Dolichocebus overlaps the extant platyrrhine range because its canine and premolars are more modest in size (figures 5.1, 6.1).
Tooth root surface area differs between the two Homunculus specimens measured (table 3). The canine, especially, has a much greater surface area in MPM-PV 3501 than in MPM-PV 3502. Canine root cross-sectional area and length are slightly greater in the former specimen, but these small differences amount to a large difference in surface area.
Leverage
Most of the extant platyrrhines in our sample have similar patterns of muscle leverage. Lever arm lengths are given for the temporalis, masseter, and medial pterygoid muscles in table 4, as are load arm lengths at all measured bite points.
Table 4. Lever arms for the three muscle groups and load arms for all tooth positions (mm) / brazos de palanca de los tres grupos musculares y brazos de carga de todas las posiciones de los dientes (mm). Temp., temporalis muscle / músculo temporal; Mass., masseter muscle / músculo masetero; M.P., medial pterigoid muscle/ músculo pterigoides medio. The load arm is measured to the paracone / el brazo de carga se midió al paracono. All data are in mm / todas las medidas en mm. Abbreviations for species as in table 1 / abreviaturas de las especies como en tabla 1.

Figure 7 shows the variation in leverage at the canine and the M2. Callithrix has the greatest masseter and medial pterygoid leverage and also has considerable temporalis leverage. Saguinus, Callicebus, and Alouatta have strong masseter and medial pterygoid leverage, but weak temporalis leverage. Additionally, Alouatta has the greatest masseter leverage in our sample, at all tooth positions. Cebuella reverses this pattern with relatively high temporalis leverage and relatively low masseter and medial pterygoid leverage. Among extant platyrrhines, Cebus has the greatest temporalis leverage at all bite points; but has very poor medial pterygoid leverage and only moderate masseter leverage.
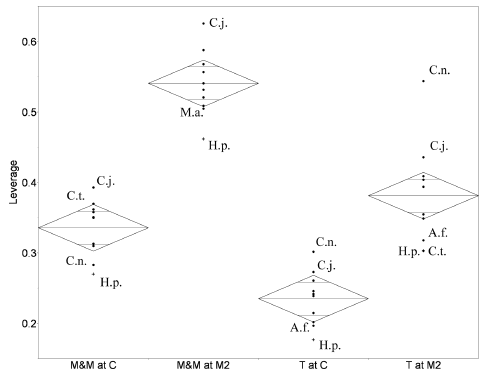
Figure 7. Comparison of muscle leverage / comparación del esfuerzo de torsión de los músculos. Plots showing muscle leverage for temporalis and for masseter plus medial pterygoid at two different bite points / diagramas mostrando el esfuerzo de torsión de los músculos temporal y masetero, más el pterigoides medio en dos puntos de mordida diferentes. Leverage is lever arm length divided by load arm length / el esfuerzo de torsión es la longitud del brazo de palanca dividida por la longitud del brazo de la carga. Abbreviations for species as in table 1 / abreviaturas de las especies como en tabla 1. Extinct species are marked with crosses / las especies extintas están indicadas con cruces. Diamonds as in figure 6. / diamantes como en la figura 6. Data are in table 4 / datos en tabla 4. M&M, masseter and medial pterygoid; T, temporalis / M&M, músculos masetero y pterigoides medio; T, músculo temporal.
The most striking result is that Homunculus has the poorest muscle leverage in the sample for all muscles at all tooth positions.
Discussion
Roots
Our method for measuring tooth root area captures true root area, even for very complicated tooth root shapes. Spencer (2003) admitted that his method using two dimensional projections of roots failed to account for parts of the root that were concealed behind the roots in the foreground.
Using a geometric mean of cranial measurements as a scaling variable is superior to using crown area when the aim is to detect dietary effects on tooth root area. This is because crown area is known to vary with diet in primates (Kay, 1975; Anthony and Kay, 1993). Crown area may be useful for understanding how bite pressure is related to tooth root area (Demes and Creel, 1988). However, bite pressure is likely only loosely related to crown area; the area of interest is actually the contact area between teeth and food. The latter may be more or less than crown area, depending on such additional variables as tooth relief and the ductility of food.
As predicted, there appears to be a signal for folivory versus frugivory in tooth root size. Folivorous Alouatta has larger postcanine tooth roots relative to its cranial geometric mean than does frugivorous Ateles. The former also has larger postcanine tooth crowns than the latter. Spencer (2003) suggested that large molar roots might be associated with leaf processing. The large size of the tooth roots - especially the molars - of Alouatta relative to Ateles supports this suggestion. It is not clear whether heavy loads or repeated loads are more likely to select for greater root area, or whether leaf-processing requires heavy loads or just repeated loads. Either way, both loading situations likely favor large roots.
Our prediction regarding canine root area in Ateles versus Alouatta also holds: canine root area is greater in Ateles than in Alouatta. This is consistent with the explanation that fruit-processing favors large canine roots.
Canine root area is smaller in Callithrix jacchus than in Saguinus fuscicollis, contrary to our initial expectations based on the assumption that gouging behavior should impose heavy loads on the anterior dentition. Indeed, our findings accord with those of Taylor and Vinyard (2004) who report that physiological cross-sectional area of the masseter muscle is actually less in Callithrix jacchus than in Saguinus oedipus, contrary to the prediction that gouging involves high bite forces. Anterior tooth root shape might provide a better signal for gouging in callitrichids as the canine and incisor roots of Callithrix are more mesiodistally compressed than those of Saguinus.
This may be an adaptation to resisting bending in the anteroposterior plane.
The Callithrix specimen in our sample is a female. Although little sexual dimorphism is expected for callitrichids, we measured the tooth roots of a male Callithrix jacchus cranium (USNM 503885) to rule out the possibility that sexual dimorphism explains the difference in tooth root area between C. jacchus and S. fuscicollis. Canine root area of the male Calithrix is greater than that of the female Callithrix; however, it is still less than that of the male Saguinus. Therefore, the difference in root size between Saguinus and Callithrix cannot be explained by sexual dimorphism alone.
Another comparison of interest is that of Pithecia to the other extant platyrrhines. Pithecia is a hard-object feeder (Kinzey, 1992; Peres, 1993). This species is expected to have large anterior tooth roots compared to the other platyrrhines in our sample. We found that canine root area is great in Pithecia compared to its cranial geometric mean (see figure 5). However, among the extant platyrrhines, Saguinus has a relatively larger canine root than Pithecia. Furthermore, premolar root area is not particularly great in Pithecia compared to the other extant platyrrhines in our sample. Although the foods consumed by Pithecia are very resistant to fracture, the closely related genus Chiropotes breaks open fruits that are considerably more resistant than those consumed by Pithecia (Kinzey and Norconk, 1990; Kinzey, 1992). Spencer (2003) found that the canine roots of Chiropotes are considerably larger than those of Pithecia (relative to a cranial geometric mean).
The great tooth root surface area observed in the Miocene platyrrhines, especially Homunculus, suggests that these roots were able to resist considerable bite loads. Root surface areas for the posterior dentition are especially great in Homunculus, suggesting this taxon may have used powerful and/or repeated bites at the molars to break down its food. However, this dietary signal is not supported by the estimates of muscle leverage or by the sizes of the muscle scars (see below).
Differences between MPM-PV 3501 and MPM-PV 3502 in root surface area are small and variable for the posterior teeth; this variation is likely due in part to root damage and distortion in the former specimen. The difference in canine root surface area, however, is considerable. Because these two specimens are anatomically very similar in most other respects (see appendix 2), this difference in canine root size might be attributed to canine sexual dimorphism. Root dimensions in a third specimen (MPM-PV 3503 from PLC) are similar to those of MPM-PV 3502 (from Killik Aike); these specimens might represent females while MPM-PV 3501 from PLC would repre-sent a male. More detailed comparisons are required to test this hypothesis.
Leverage
The pattern of chewing muscle leverage in platyrrhines is likely related to cranial morphology and diet. The lever arms for the masseter and medial pterygoid are generally long in primates that have the mandibular condyle positioned well above the occlusal plane; whereas, the temporalis lever arm is short (Greaves, 2000). Callicebus and Alouatta both have deep jaws with high condyles; their masseter and medial pterygoid muscles have considerably more leverage than the other platyrrhines in the sam-ple, but their temporalis leverage is poor. Alouatta's considerable masseter leverage might be an adaptation for generating great bite force at narrow gapes (e.g., during mastication of flat food items like leaves).
The temporalis muscle is likely important in producing force when the mouth is gaped widely (Perry, 2008), for example, when a large food item is bitten or during gouging. This may partly explain why Callithrix has considerable temporalis leverage; nevertheless, Callithrix also has considerable leverage for the medial pterygoid and masseter.
Species rankings for leverage are very different for the temporalis versus the masseter/medial pterygoid at all tooth positions. However, for a given muscle, species rankings vary little from one tooth position to another. This suggests that the teeth are similarly spaced along the tooth row in this sample of extant platyrrhines, but that the locations of the jaw adductor attachments vary. One notable exception is that at the canine Ateles has greater medial pterygoid leverage than Alouatta, but at the M3 Ateles has much less medial pterygoid leverage than Alouatta. The same is true to a lesser degree for masseter leverage in these two taxa. This result is consistent with the expectation that chewing muscle leverage is maximized at points along the tooth row where food is most heavily loaded. Thus these differences in leverage likely reflect Alouatta's emphasis on leaves and Ateles's emphasis on fruits.
Jaw adductor leverage in Homunculus is poor relative to all other platyrrhines examined. This is partly a consequence of the length of its rostrum. Homunculus is prognathic relative to the extant platyrrhines (figure 8). Therefore, its load arm at any given bite point is longer than that of the other monkeys. Nevertheless, in addition to having long load arms, Homunculus also has short lever arms.

Figure 8. Skull of Callithrix jacchus (NMNH 503895f) superimposed on the composite skull of Homunculus patagonicus / Cráneo de Callithrix jacchus (NMNH 503895f) sobrepuesto en el cráneo compuesto de Homunculus patagonicus. This image shows the degree of prognathism of Homunculus in relation to Callithrix and the more posterior extent of the temporalis origin / esta imagen muestra el grado de prognatismo de Homunculus en relación a Callithrix y la ubicación más posterior del músculo temporal. The original images are scaled to equal skull height at the anterior edge of the postorbital bar (black vertical line at left) and were superimposed at the mandibular condyles / las imágenes originales fueron llevadas a la misma escala para igualar la altura del cráneo en el borde anterior de la barra postorbital (línea negra vertical de la izquierda) y fueron superpuestas a nivel del cóndilo mandibular.
The origin of the temporalis is more posteriorly located in Homunculus than in extant platyrrhines, but the temporalis insertion is no more anteriorly located. To a lesser degree, the insertions of masseter and medial pterygoid are also more posteriorly located, but the masseter and medial pterygoid origins are no more anteriorly located. This means that the line of action of each muscle passes closer to the condyle than in the extant species, and therefore, the lever arms in Homunculus are shorter.
Based solely on information about the origins of these muscles (but not on their insertions), it is likely that Dolichocebus and Tremacebus also had poor chew-ing muscle leverage because both crania are relatively prognathic, and the temporalis origin is situated posteriorly on the cranium.
Homunculus patagonicus has large maxillary tooth roots relative to the extant platyrrhines studied. Also, the cheek teeth are heavily worn in most specimens of this species. These signs point to a diet of very tough foods. However, there is no reason to believe that Homunculus had especially large chewing muscles. There is no sagittal crest and the temporal lines are not especially prominent. The muscle scars are not especially deep, and the zygomatic arches are only moderately flared, leaving only a modest amount of space for the chewing muscles in the temporal fossa. Jaw adductor leverage in Homunculus is not as great as in the extant platyrrhines studied here. This finding complements the above inference about overall muscle size. Assuming modest chewing muscle cross-sectional area and activity, and in spite of its large tooth roots, this Miocene primate was probably not especially well-adapted to producing heavy bite loads during chewing.
Inferences about masticatory function and diet
The following features characterize the masticatory system of Homunculus: moderate molar shearing (Fleagle et al., 1997), heavy postcanine tooth wear(Tauber, 1991), large postcanine tooth roots, and modestly developed musculature with poor leverage. One possible explanation for the cooccurrence of these traits is that Homunculus consumed both fruits and leaves, but that neither required very forceful mastication. Perhaps during mastication, bite forces were never high.
A diet of fruit and leaves may not be sufficient to explain the high degree of tooth wear seen in most specimens of Homunculus. Leaves and other plant matter ingested by Homunculus might have been heavily protected by silica inclusions and/or might have been covered with exogenous abrasives (Sanson et al., 2007).
Other interpretations are possible: Homunculus may have eaten foods that required more masticatory cycles than typically seen in living platyrrhines. This would lead to the inference that its foods were more fibrous than those of most platyrrhines. Another interpretation relates to seasonality. Homunculus is found as far south as 55 degrees South latitude, almost 3000 kilometers south of the southern edge of the distribution of living platyrrhines, and just 1,200 kilometers north of the Antarctic Circle. The paleolatitude of this locality was several degrees even further south in the early Miocene. An abrasive diet may have been a feature of the long season of low plant productivity at this latitude.
Tauber (1991, 1999) described a new cranium of Homunculus and made dietary inferences based on cranial features. He suggested that Homunculus had a very powerful masticatory system, more powerful than that of pitheciines. This was based partly on rugosity for the attachment of the buccinator muscle and partly on dental morphology. Even if the size of the buccinator muscle can be judged from scars on bone, it is uncertain what, if anything, it has to do with food properties. The buccinator is located within the cheek and aids the tongue to maintain a food bolus on the teeth and likely plays an important role in suckling (Tomo et al., 2002); its bony attachments are slight. Because the buccinator muscle does not generate force to fracture foods, the physical properties of foods have likely had no adaptive influence of the muscle.
Tauber also suggested that the origin of the temporalis from the cranial surface indicates that this was a powerful muscle. We have examined all known crania of Homunculus patagonicus and have found nothing to suggest that the temporalis muscles were especially large compared to those of extant platyrrhines. The temporal lines are slight, the lateral surface of the skull for the origin of the deep temporalis is only moderately rugose, and the temporal fossa (through which the temporalis passes toward its insertion) is small (figure 9). Homunculus probably did not have a powerful masticatory system compared to extant pitheciines. The former has poor masticatory leverage and its cranial anatomy suggests only modestly-sized chewing muscles. Although the diet of Homunculus probably included very abrasive, resistant foods, at least seasonally, it was probably not capable of generating masticatory forces like those used by extant pitheciines.
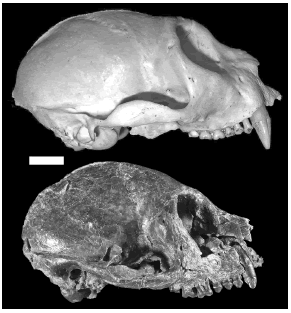
Figure 9. Comparison of cranium of Homunculus MPM-PV 3501 (below) to cranium of Pithecia NP5 (above) / comparación de los cráneos de Homunculus MPM-PV 3501 (abajo) y de Pithecia NP5 (arriba). The bony surface associated with the masticatory muscles is not obviously more developed in Homunculus / la superficie del hueso asociada con los músculos masticatorios no está más desarrollada en Homunculus. The extent of the scar for the origin of temporalis is similar in both specimens / el área de la cicatriz del origen del músculo temporal es similar en ambos especímenes. Compare also with specimen MPM-PV 3502 in figure 2 / comparar también con el espécimen MPM-PV 3502 en figura 2. Scale bar = 1 cm / escala = 1 cm.

Allocation of MPM-PV 3502 to Homunculus patagonicus
A critical aspect of our reconstructions is to assure that the specimens used in the reconstructions all belong to the same taxon, Homunculus patagonicus. This particularly is an issue with the skull MPM-PV 3502, which has not yet been described. All relevant species and specimens have been examined by one of us (RFK) and a detailed description and assessment of their allocation to Homunculus is in progress (Kay et al., 2005). Until 2006 the consensus was that all specimens of primates from coastal exposures of the Santa Cruz Formation belonged to a single species, Homunculus patagonicus. Two events have modified this picture. Tejedor et al. (2006) named a new genus Killikaike blakei from Killik Aike Norte on the estuary of the Río Gallegos. The type, MPM-PV 5000, is the rostral part of a cranium with most of its upper cheek teeth. Because the type specimen of Homunculus patagonicus is now lost, Tejedor and Rosenberger (2008) proposed a neotype, MACN-A 5757, a mandible from Puesto Estancia La Costa. Unfortunately, the teeth of the neotype are too heavily worn to discern most of the details of the dental anatomy and the type specimen of Killikaike preserves no mandibular dentition. Therefore, only specimens in the hypodigm of Homunculus, as proposed by Tejedor and Rosenberger (2008), can be compared to the type of Killikaike. One member of the hypodigm of Homunculus patagonicus is a partial skull CORD-PZ 1130, described by Tauber (1991) from the same locality as the neotype. Another is a distorted and broken orbit and maxilla, MACN-A 5968, also from Corriguen Aike, preserving virtually the same anatomy as CORD-PZ 1130. Two other adult skulls come from this same locality: a virtually complete but distorted skull, MPM-PV 3501 (figured by Tafforeau et al., 2006), and another less complete skull (MPM-PV 3503) preserving the same parts as CORD-PZ 1130. We have chosen to compare our material mostly with the published specimen CORD-PZ 1130. Examination of the anatomy of all these specimens together suggests that we are dealing with one species or at most two species of the same genus and that MPM-PV 3502 is most similar to CORD-PZ 1130 and less so to MPM-PV 5000.
In the original description, Tejedor et al. (2006) do not offer a differential diagnosis of Killikaike blakei from Homunculus patagonicus, but in a later paper, Tejedor and Rosenberger (2008) list the following differences.
1) Homunculus is said to have larger canines than Killikaike. This does not appear to be a diagnostic difference between the two species: the canine occlusal dimensions of CORD-PZ 1130 (Homunculus patagonicus from Puesto Estancia La Costa) (mesiodistal: 2.85 mm x buccolingual 3.10 mm) are nearly identical to those of the type of K. blakei, MPM-PV 5000 (2.8 mm x 3.4 mm). MPM-PV 3502 has slightly larger canines than K. blakei. Canine dimensions in MPM-PV 3501 and MPM-PV 3502 are similar, though somewhat greater in the former. Canine length in MPM-PV 3501 is 13.34 mm as compared to 12.63 mm in MPM-PV 3502. Canine cross-sectional area at the cementoenamel junction is 8.33 mm2 in MPM-PV 3501 and 7.99 mm2 in MPM-PV 3502. This small difference represents a great difference in root surface area. Nevertheless, canine dimensions of all specimens from both named taxa fall within the range of variation of non-sexually dimorphic platyrrhines like Aotus (see Kay et al., 1987).
2) They state that the interorbital region is relatively wider in Homunculus than it is in Killikaike. The interorbital breadth of four specimens from PLC has a mean of 5.27 mm and a range of 5.06 to 5.52 (that of CORD-PZ 1130 is 5.2 mm). Interorbital breadth is 4.4 mm in MPM-PV 5000. The interorbital dimension in MPM-PV 3502 is 4.2 mm, closer to K. blakei. However, there is considerable variation in interorbital dimensions even in non-sexually dimorphic platyrrhine species. Hartwig (1993) gives a coefficient of variation of 7.0 for seventeen specimens of the extant platyrrhine Callicebus hofmannsi. In his sample of four specimens of Callicebus personatus, the measured interorbital breadths range from 4.24 mm to 5.46 mm, overlapping virtually the entire range of published breadths of H. patagonicus and K. blakei. Pending recovery of more specimens there does not appear to be sufficient variation in this dimension to warrant recognition of two taxa.
3) The profile of the frontal bone is said to be ‘low' in Homunculus whereas it is more vaulted in Killikaike. MPM-PV 3502 resembles Homunculus in this respect and, like CORD-PZ 1130 and the other frontal specimens from PLC, has a slightly depressed area posterior to the glabellar point, whereas the corresponding area in Killikaike is slightly convex.
4) The postorbital constriction is said to be narrower in Homunculus than in Killikaike. This is difficult to determine accu rately in CORD-PZ 1130. Nevertheless, MPM-PV 3502 has a far narrower postorbital constriction than does MPM-PV 5000, and in that respect resembles Homunculus.
A detailed comparison of other cranial and dental proportions of MPM-PV 5000 and CORD-PZ 1130 reveal some other differences.
1) The anterior root of the zygomatic arch of CORD-PZ 1130 is deeper dorsoventrally than in MPM-PV 5000. MPM-PV 3502 agrees with CORD-PZ 1130 in that respect.
2) The M2 is more transverse in MPM-PV 3502 than in Killikaike (MPM-PV 5000 and MPM-PV 1607, a referred specimen). The ratio of M2 buccolingual breadth to mesiodistal length is 1.57 in MPM-PV 3502 versus 1.33 and 1.38 in MPM-PV 5000 and MPM-PV 1607.
Thus MPM-PV 3502 repeats in the M2 the shape difference shown between Killikaike and Homunculus in the P3.
Overall, Killikaike and Homunculus are strikingly similar in most details of dental and cranial morphology. Most of the morphological differences mentioned above are subtle and have yet to be placed in the context of within-species variation in samples of living platyrrhines. Several other cranial and dental characters could be compared, but that is beyond the scope of this study. Furthermore, given that the holotype of Killikaike blakei is a subadult, many of the diagnostic characters of this species, such as the shape of the frontal and the length of the rostrum are of limited comparative value as they are likely to change through ontogeny. Nevertheless, Killikaike and Homunculus (as represented by CORD-PZ 1130) do differ in a number of ways. And where they differ, MPM-PV 3502 more closely resembles Homunculus. Therefore, we are confident that MPM-PV 3502 should be allocated to Homunculus patagonicus.
Acknowledgements
We thank the Patagonia field crews, seasons 2003 to present, for their efforts in collecting Miocene mammals, including primates, from the Santa Cruz Formation of coastal Argentina. We thank the Battini family for their hospitality during the fieldwork. We thank the staff of the Museo Regional Provincial "Padre Manuel Jesús Molina" (Río Gallegos, Argentina), the staff of the National Museum of Natural History (particularly L. Gordon), and J. Fleagle (Stony Brook University), for allowing us to borrow specimens of platyrrhines for CT scanning and study. Special thanks to M. Colbert and the staff of the University of Texas (Austin) MicroCT Facility for making the scans used in this study. Ch. Wall and R. Madden provided insightful suggestions throughout the execution of this study. We thank two reviewers and the editorial staff for helping us to improve the quality of this paper. This project was funded by the National Science Foundation, the National Geographic Society, the Agencia Nacional de Promoción Científica y Tecnológica (PICT 26219), the Universidad Nacional de La Plata (N474), and Midwestern University.
References
1. Ameghino, F. 1891. Los monos fósiles del Eoceno de la República Argentina. Revista Argentina de Historia Natural 1: 383-397. [ Links ]
2. Anapol, F., Shahnoor, N. and Ross, C.F. 2008. Scaling of reduced physiologic crosssectional area in primate muscles of mastication. In: C.J. Vinyard, M.J. Ravosa and C.E. Wall (eds.), Primate Craniofacial Function and Biology, Springer US, New York, pp. 201-216. [ Links ]
3. Anthony, M.R.L. and Kay, R.F. 1993. Tooth form and diet in ateline and alouattine primates: reflections on the comparative method. American Journal of Science 293A: 356-382. [ Links ]
4. Bargo, M.S. 2003. Biomechanics and palaeobiology of the Xenarthra: the state of the art. Senckenbergiana Biologica 83: 41-50. [ Links ]
5. Beertsen, W., McCulloch, C.A.G. and Sodek, J. 1997. The periodontal ligament: a unique, multifunctional connective tissue. Periodontology 13: 20-40. [ Links ]
6. Bluntschli, H. 1931. Homunculus patagonicus und die ihm zugereihten Fossilfunde aus den Santa-Cruz-Schichten Patagoniens. Morphologisches Jahrbuch 67: 811-892. [ Links ]
7. Butler, P.M. 1973. Molar wear facets of early Tertiary North American primates. In: W. Montagna and M.R. Zingeser (eds.), Symposia of the Fourth International Congress of Primatology 3, Karger, Basel, pp. 1-27. [ Links ]
8. Cant, J.G.H. 1990. Feeding ecology of spider monkeys (Ateles geof-froyi) at Tikal, Guatemala. Human Evolution 5: 269-281. [ Links ]
9. Chapman, C.A. and Chapman, L.J. 1991. The foraging itinerary of spider monkeys: when to eat leaves? Folia Primatologica 56: 162-166. [ Links ]
10. Chiarello, A.G. 1994. Diet of the brown howler monkeyAlouatta fusca in a semideciduous forest fragment of southeastern Brazil. Primates 35: 25-34. [ Links ]
11. Coimbra-Filho, A.F. and Mittermeier, R.A. 1976. Exudate-eating and tree gouging in marmosets. Nature 262: 630. [ Links ]
12. Corruccini, R.S. and Beecher, R.M. 1982. Occlusal variation related to soft diet in a non-human primate. Science 218: 74-76. [ Links ]
13. Covert, H.H. 1986. Biology of Early Cenozoic primates. In: editores, Comparative Primate Biology, Volume 1: Systematics, Evolution, and Anatomy, Alan R. Liss, New York, pp. 335-359. [ Links ]
14. De Iuliis, G., Bargo, M.S. and Vizcaíno, S.F. 2000. Variation in skull morphology and mastication in the fossil giant armadillos Pampatherium spp. and allied genera (Mammalia: Xenarthra: Pampatheriidae), with comments on their systematics and distribution. Journal of Vertebrate Paleontology 20: 743-754. [ Links ]
15. Demes, B. and Creel, N. 1988. Bite force, diet, and cranial morphology of fossil hominids. Journal of Human Evolution 17: 657-670. [ Links ]
16. Ferrari, S.F. 1993. Ecological differentiation in the Callitrichidae. In: A.B. Rylands (ed.), Marmosets and Tamarins: Systematics, Behaviour, and Ecology, Oxford University Press, Oxford, pp. 314-328. [ Links ]
17. Fleagle, J.G. and Kay, R.F. 1989. The dental morphology of Dolichocebus gaimanensis, a fossil monkey from Argentina. American Journal of Physical Anthropology 78: 221. [ Links ]
18. Fleagle, J.G. and Kay, R.F. 1997. Platyrrhines, catarrhines, and the fossil record. In: W. G. Kinzey (ed.), New World Primates: Ecology, Evolution, and Behavior, Walter de Gruyter, Inc., New York, pp. 3-24. [ Links ]
19. Fleagle, J.G. and Tejedor, M.F. 2002. Early platyrrhines of southern South America. In: W.C. Hartwig (ed.), The Primate Fossil Record, Cambridge University Press, Cambridge, pp. 161-174. [ Links ]
20. Fleagle, J.G., Kay, R.F. and Anthony, M.R.L. 1997. Fossil New World monkeys. In: R.F. Kay, R.H. Madden, R.L. Cifelli and J.L. Flynn (eds.), Vertebrate Paleontology in the Neotropics: The Miocene Fauna of La Venta, Colombia, Smithsonian Institution Press, Washington, D.C., pp. 473-495. [ Links ]
21. Flynn, J.L., Wyss, A.R. and Swisher III, C.C. 1995. An early Miocene anthropoid skull from the Chilean Andes. Nature 373: 603-607. [ Links ]
22. Ford, S.M. 1990. Locomotor adaptations of fossil platyrrhines. Journal of Human Evolution 19: 141-173. [ Links ]
23. Ford, S.M. and Davis, L.C. 1992. Systematics and body size: Implications for feeding adaptations in New World Monkeys. American Journal of Physical Anthropology 88: 415-468. [ Links ]
24. Gordon, K.D. 1982. A study of microwear on chimpanzee molars: implications for dental microwear analysis. American Journal of Physical Anthropology 59: 195-215. [ Links ]
25. Greaves, W.S. 2000. Location of the vector of jaw muscle force in mammals. Journal of Morphology 243: 293-299. [ Links ]
26. Hartwig, W.C. 1993. Comparative Morphology, Ontogeny, and Phylogenetic Analysis of the Platyrrhine Cranium. University of California, Berkeley, Berkeley, CA, 628 pp. [ Links ]
27. Hershkovitz, P. 1970. Notes on Tertiary platyrrhine monkeys and description of a new genus from the Late Miocene of Colombia. Folia Primatologica 12: 1-37. [ Links ]
28. Hershkovitz, P. 1974. A new genus of Late Oligocene monkey (Cebidae, Platyrrhini) with notes on postorbital closure and platyrrhine evolution. Folia Primatologica 21: 1-35. [ Links ]
29. Hershkovitz, P. 1981. Comparative anatomy of platyrrhine mandibular cheek teeth dpm4, pm4, m1 with particular reference to those of Homunculus (Cebidae), and comments on platyrrhine origins. Folia Primatologica 35: 179-217. [ Links ]
30. Hershkovitz, P. 1984. More on the Homunculus dpm4 and m1 and comparisons with Alouatta and Stirtonia (Primates, Platyrrhini, Cebidae). American Journal of Primatology 7: 261-283. [ Links ]
31. Hoffstetter, R. 1969. Un primate de l'Oligocene inférieur sudamericain: Branisella boliviana gen. et sp. nov. Comptes Rendus de l'Academie des Sciences - Series D 269: 434-437. [ Links ]
32. Hylander, W.L. 1975. The human mandible: lever or link? American Journal of Physical Anthropology 43: 227-242. [ Links ]
33. Hylander, W.L., Ravosa, M.J., Ross, C.F., Wall, C.E. and Johnson, K.R. 2000. Symphyseal fusion and jaw-adductor muscle force: an EMG study. American Journal of Physical Anthropology 112: 469-492. [ Links ]
34. Janis, C.M. 1984. Prediction of primate diets from molar wear patterns. In: D.J. Chivers, B.A. Wood and A. Bilsborough (eds.), Food Acquisition and Processing in Primates, Plenum Press, New York, pp. 331-340. [ Links ]
35. Kay, R.F. 1975. The functional adaptations of primate molar teeth. American Journal of Physical Anthropology 43: 195-216. [ Links ]
36. Kay, R.F. 1977. The evolution of molar occlusion in the Cercopithecidae and early catarrhines. American Journal of Physical Anthropology 46: 327-352. [ Links ]
37. Kay, R.F. and Cozzuol, M.A. 2006. A new platyrrhine from the Solimões Formation (late Miocene, Acre State, Brazil) with comments about other Miocene monkeys from that region. Journal of Human Evolution 50: 673-686. [ Links ]
38. Kay, R.F. and Frailey, C.D. 1993. Large fossil platyrrhines from the Rio Acre local fauna, late Miocene, western Amazonia. Journal of Human Evolution 25: 319-327. [ Links ]
39. Kay, R.F., Plavcan, J.M., Glander, K.G. and Wright, P.C. 1987. Sexual selection and canine dimorphism in New World monkeys. American Journal of Physical Anthropology 77: 385-397. [ Links ]
40. Kay, R.F., Williams, B.A. and Anaya, F. 2002. The adaptations of Branisella boliviana, the earliest South American monkey. In: J.M. Plavcan, R.F. Kay, W.L. Jungers and C.P. van Schaik (eds.), Reconstructing Behavior in the Primate Fossil Record, Kluwer Academic / Plenum Publishers, New York, pp. 339-370. [ Links ]
41. Kay, R.F., Schmitt, D.O., Vinyard, C.J., Perry, J.M.G., Shigehara, N., Takai, M. and Egi, N. 2004. The paleobiology of Amphipithecidae, South Asian late Eocene primates. Journal of Human Evolution 46: 3-25. [ Links ]
42. Kay, R.F., Vizcaíno, S.F., Tauber, A.A., Bargo, M.S., Williams, B.A., Luna, C. and Colbert, M.W. 2005. Three newly discovered skulls of Homunculus patagonicus support its position as a stem platyrrhine and establish its diurnal arboreal folivorous habits. American Journal of Physical Anthropology 40S: 127. [ Links ]
43. Kay, R.F., Fleagle, J.G., Mitchell, T.R.T., Colbert, M.W., Bown, T.M. and Powers, D.W. 2008a. The anatomy of Dolichocebus gaimanensis, a stem platyrrhine monkey from Argentina. Journal of Human Evolution 54: 323-382. [ Links ]
44. Kay, R.F., Vizcaíno, S.F., Bargo, M.S., Perry, J.M.G., Prevosti, F. and Fernicola, J.C. 2008b. Two new fossil vertebrate localities in the Santa Cruz Formation (late early Miocene, Argentina), ~51 degrees South latitude. Journal of South American Earth Sciences 25: 187-195. [ Links ]
45. Kinzey, W.G. 1992. Dietary and dental adaptations in the Pitheciinae. American Journal of Physical Anthropology 88: 499-514. [ Links ]
46. Kinzey, W.G. and Norconk, M.A. 1990. Hardness as a basis of fruit choice in two sympatric primates. American Journal of Physical Anthropology 81: 5-15. [ Links ]
47. Kraglievich, J.L. 1951. Contribuciones al conocimento de los primates fósiles de la Patagonia. I. Diagnosis previa de un nuevo primate fósil del Oligoceno superior (Colhuehuapiano) de Gaiman, Chubut. Insituto Nacional de Investigaciones de las Ciencias Naturales 2: 55-82. [ Links ]
48. Kupczik, K. and Dean, M.C. 2007. Comparative observations on the tooth root morphology of Gigantopithecus blacki. Journal of Human Evolution 54: 196-204. [ Links ]
49. Lucas, P.W. 2004. Dental Functional Morphology: How Teeth Work. Cambridge University Press, Cambridge, 355 pp. [ Links ]
50. MacFadden, B.J. 1990. Chronology of Cenozoic primate localities in South America. Journal of Human Evolution 19: 7-21. [ Links ]
51. Madeira, M.C. and de Oliveira, J.A. 1979. Anatomical aspects of the masticatory muscles of the tufted capuchin (Cebus apella). Okajimas Folia Anatomica, Japan 56: 35-44. [ Links ]
52. Marshall, L.G. 1976. Fossil localities for Santacrucian (Early Miocene) mammals, Santa Cruz Province, southern Patagonia, Argentina. Journal of Paleontology 50: 1129-1142. [ Links ]
53. Meldrum, D.J. 1993. Postcranial adaptations and positional be-havior in fossil platyrrhines. In: D.L. Gebo (ed.), Postcranial Adaptation in Non-human Primates, Northern Illinois University Press, DeKalb, IL, pp. 235-251. [ Links ]
54. Peres, C.A. 1993. Notes on the ecology of Buffy Saki Monkeys (Pithecia albicans, Gray 1860): a canopy seed-predator. American Journal of Primatology 31: 129-140. [ Links ]
55. Perry, J.M.G. 2008. The Anatomy of Mastication in Extant Strepsirrhines and Eocene Adapines. Duke University, Durham. 490pp. [ Links ]
56. Perry, J.M.G. and Wall, C.E. 2008. Scaling of the chewing muscles in prosimians. In: C.J. Vinyard, M.J. Ravosa and C.E. Wall (eds.), Primate Craniofacial Function and Biology, Springer US, New York, pp. 217-240. [ Links ]
57. Rosenberger, A.L. 1978. New data on Branisella and Homunculus. American Journal of Physical Anthropology 48: 431. [ Links ]
58. Rosenberger, A.L. 1992. Evolution of feeding niches in New World Monkeys. American Journal of Physical Anthropology 88: 525-562. [ Links ]
59. Rosenberger, A.L., Setoguchi, T. and Shigehara, N. 1990. The fossil record of callitrichine primates. Journal of Human Evolution 19: 209-236. [ Links ]
60. Rosenberger, A.L., Hartwig, W.C. and Wolff, R.G. 1991. Szalatavus attricuspis, an early platyrrhine primate. Folia Primatologica 56: 225-233. [ Links ]
61. Rosenberger, A.L., Tejedor, M.F., Cooke, S.B. and Pekar, S. 2009. Platyrrhine ecophylogenetics in space and time. In: P.A. Garber, A. Estrada, J.C. Bicca-Marques, E.W. Heymann and K.B. Strier (eds.), South American Primates, Springer Science, New York, pp. 69-113. [ Links ]
62. Rusconi, C. 1933. Nuevos restos de monos fósiles del Terciario antiguo de la Patagonia. Anales de la Sociedad Científica Argentina 116: 286-289. [ Links ]
63. Rusconi, C. 1935. Las especies de primates del oligoceno de Patagonia (gen. Homunculus). Revista Argentina de Paleontología y Anthropología Ameghinia 1: 39-68, 71-100,103-125. [ Links ]
64. Sanson, G.D., Kerr, S.A. and Gross, K.A. 2007. Do silica phytoliths really wear mammalian teeth? Journal of Archaeological Science 34: 526-531. [ Links ]
65. Schneider, H. 2000. The current status of the New World monkey phylogeny. Anais da Academia Brasileira de Ciências 72: 165-172. [ Links ]
66. Schoeninger, M.J., Iwaniec, U.T. and Glander, K.G. 1997. Stable isotope ratios indicate diet and habitat use in New World Monkeys. American Journal of Physical Anthropology 103: 69-83. [ Links ]
67. Schön, M.A. 1968. The muscular system of the red howling monkey. United States National Museum Bulletin 273: 1-185. [ Links ]
68. Schumacher, G.H. 1961. Funktionelle Morphologie der Kaumuskulatur. Fisher, Jena, 189 pp. [ Links ]
69. Spencer, M.A. 1999. Constraints on masticatory system evolution in anthropoid primates. American Journal of Physical Anthropology 108: 483-506. [ Links ]
70. Spencer, M.A. 2003. Toothroot form and function in platyrrhine seedeaters. American Journal of Physical Anthropology 122: 325-335. [ Links ]
71. Stark, D. 1933. Die kaumuskulatur der platyrrhinen. Gegenbaurs Morphologisches Jahrbuch 72: 212-285. [ Links ]
72. Stern, J.T. 1974. Biomechanical significance of the instantaneous center of rotation: The human temporomandibular joint. Journal of Biomechanics 7: 109. [ Links ]
73. Stirton, R.A. 1951. Ceboid monkeys from the Miocene of Colombia. University of California Press, Berkeley, 355 pp. [ Links ]
74. Strait, S.G. 1993. Molar morphology and food texture among small-bodied insectivorous mammals. Journal of Mammalogy 74: 471-484. [ Links ]
75. Strier, K.B. 1992. Atelinae adaptations: Behavioral strategies and ecological constraints. American Journal of Physical Anthropology 88: 515-524. [ Links ]
76. Sussman, R.W. and Kinzey, W.G. 1984. The ecological role of the Callitrichidae: a review. American Journal of Physical Anthropology 64: 419-449. [ Links ]
77. Tafforeau, P., Boistel, R., Boller, E., Bravin, A., Brunet, M., Chaimanee, Y., Cloetens, P., Feist, M., Hoszowska, J., Jaeger, J.-J., Kay, R.F., Lazzari, V., Marivaux, L., Nel, A., Nemoz, C., Thibault, X., Vignaud, P. and Zabler, S. 2006. Applications of X-ray synchrotron microtomography for non-destructive 3D studies of paleontological specimens. Applied Physics A: Materials Science & Processing 83: 195-202. [ Links ]
78. Takai, M. and Anaya, F. 1996. New specimens of the oldest fossil platyrrhine, Branisella boliviana, from Salla, Bolivia. American Journal of Physical Anthropology 99: 301-317. [ Links ]
79. Tattersall, I. 1974. Facial structure and mandibular mechanics in Archaeolemur. In: R.D. Martin, G.A. Doyle and A.C. Walker (eds.), Prosimian Biology, Duckworth, London, pp. 563-577. [ Links ]
80. Tauber, A.A. 1991. Homunculus patagonicus Ameghino, 1891 (Primates, Ceboidea), Mioceno Temprano, de la costa Atlántica austral, Provincia de Santa Cruz, República Argentina. Academia Nacional de Ciencias, Córdoba, Miscelánea 82: 1-32. [ Links ]
81. Tauber, A.A. 1999. Los vertebrados de la Formación Santa Cruz (Mioceno Inferior-medio) en el extremo sureste de la Patagonia y su significado paleoecológico. Revista Española de Paleontología 14: 173-182. [ Links ]
82. Taylor, A.B. and Vinyard, C.J. 2004. Comparative analysis of masseter fiber architecture in tree-gouging (Callithrix jacchus) and nongouging (Saguinus oedipus) callitrichids. Journal of Morphology 261: 276-285. [ Links ]
83. Teaford, M.F. and Walker, A.C. 1984. Quantitative differences in dental microwear between primate species with different di-ets and a comment on the presumed diet of Sivapithecus. American Journal of Physical Anthropology 64: 191-200. [ Links ]
84. Tejedor, M.F. 2005a. New specimens of Soriacebus adrianae, with comments on pitheciin primates from the Miocene of Patagonia. Ameghiniana 42: 249-251. [ Links ]
85. Tejedor, M.F. 2005b. New fossil platyrrhine from Argentina. Folia Primatologica 76: 146-150. [ Links ]
86. Tejedor, M.F. 2008. The origin and evolution of Neotropical Primates. Arquivos do Museu Nacional, Rio de Janeiro 66: 251-269. [ Links ]
87. Tejedor, M.F. and Rosenberger, A.L. 2008. A neotype for Homunculus patagonicus Ameghino, 1891, and a new interpretation of the taxon. PaleoAnthropology 2008: 68-82. [ Links ]
88. Tejedor, M.F., Tauber, A.A., Rosenberger, A.L., Swisher III, C.C. and Palacios, M.E. 2006. New primate genus from the Miocene of Argentina. Proceedings of the National Academy of Sciences of the USA 103: 5437-5441. [ Links ]
89. Tomo, S., Tomo, I., Nakajima, K., Townsend, G.C. and Hirata, K. 2002. Comparative anatomy of the buccinator muscle in cat Felis domestica. The Anatomical Record 267: 78-86. [ Links ]
90. Vinyard, C.J., Wall, C.E., Williams, S.H., Schmitt, D.O. and Hylander, W.L. 2001. A preliminary report on the jaw mechanics during tree gouging in common marmosets (Callithrix jacchus). In: A. Brooks (ed.), Dental Morphology 2001: Proceedings of the 12th International Symposium on Dental Morphology, Sheffield Academic Press, Sheffield, pp. 283-297. [ Links ]
91. Vizcaíno, S.F. and Bargo, M.S. 1998. The masticatory apparatus of the armadillo Eutatus (Mammalia, Cingulata) and some allied genera: paleobiology and evolution. Paleobioliology 24: 371-383. [ Links ]
92. Vizcaíno, S.F., De Iuliis, G. and Bargo, M.S. 1998. Skull shape, masticatory apparatus, and diet of Vassallia and Holmesina (Mammalia: Xenarthra: Pampatheriidae). When anatomy constrains destiny. Journal of Mammalian Evolution 5: 291-322. [ Links ]
93. Vizcaíno, S.F., Bargo, M.S. and Cassini, G.H. 2006. Dental occlusal surface area in relation to body mass, food habits and other biological features in fossil xenarthrans. Ameghiniana 43: 11-26. [ Links ]
94. Weijs, W.A. and Hillen, B. 1985. Physiological cross-section of the human jaw muscles. Acta Anatomica 121: 31-35. [ Links ]
95. Wolff, R.G. 1984. New specimens of the primate Branisella boliviana from the early Oligocene of Salla, Bolivia. Journal of Vertebrate Paleontology 4: 570-574. [ Links ]
96. Wood, B.A., Abbott, S.A. and Uytterschaut, H. 1988. Analysis of the dental morphology of Plio-Pleistocene hominids IV. Mandibular postcanine root morphology. Journal of Anatomy 156: 107-139. [ Links ]
97. Zhang, G., Tsou, Y.-c. and Rosenberger, A.L. 2000. Reconstruction of the Homunculus skull using a combined scanning and stereolithography process. Rapid Prototyping Journal 6: 267-275. [ Links ]
Recibido: 17 de abril de 2009.
Aceptado: 7 de enero de 2010.

