










Serviços Personalizados
Journal

Artigo
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAmeghiniana
versão On-line ISSN 1851-8044
Ameghiniana vol.47 no.3 Buenos Aires set. 2010
ARTÍCULOS ORIGINALES
Taphonomy of Bouchardia rosea (Rhynchonelliformea, Brachiopoda) shells from Ubatuba Bay, Brazil: implications for the use of taphonomic signatures in (paleo)environmental analysis
Sabrina Coelho Rodrigues1 and Marcello Guimarães Simões2
1Faculdade de Ciências Integradas do Pontal, Ciências Biológicas, Universidade Federal de Uberlândia, Avenida José João Dib, 2545, Progresso, 38302-000, Ituiutaba, MG, Brasil scrodrigues@pontal.ufu.br.
2Instituto de Biociências, Universidade Estadual Paulista, Distrito de Rubião Junior, CP. 510, 18.610-000, Botucatu, SP, Brasil. btsimoes@ibb.unesp.br
Abstract. This study analyzes taphonomic alteration of brachiopod shells, i.e., Bouchardia rosea (Mawe) from Ubatuba Bay, northern coast of São Paulo State, Brazil. The average age of the studied shell deposits is 674 years BP. The underlying hypothesis is that taphonomic signatures are not uniformly distributed along a bathymetric gradient. Assemblages were sampled using a Van Veen grab sampler (31 x 40cm) along transect of fourteen stations from zero (beach) to 35 m water depth. Four sedimentary environments were recognized: (a) beach, (b) protected bay with clay bottom, (c) protected bay with sandy bottom, and (d) inner shelf with sandy bottom. All brachiopod shells (i.e., total fraction including fragments) from each of the five stations with appreciable accumulations (up to 1607 shells) were investigated following a protocol of ten taphonomic signatures. Six of these ten taphonomic signatures (articulation, fragmentation, edge alteration, color alteration, bioerosion, and encrustation) were homogeneously distributed among studied environments, indicating that these general types of taphonomic signatures per se are not sensitive to environmental conditions in this case. On the other hand, shell surface texture varied along the bathymetric gradient, thus showing a high potential to be used in taphofacies analyses. Several examples of taphonomic generalizations available in the literature are neither applicable nor comparable to our results, which further confirm that overgeneralizations should be avoided in taphonomic studies.
Resumen. Tafonomía de los depósitos de Bouchardia rosea (Rhynchonelliformea, Brachiopoda) de la Bahía de Ubatuba, Basil: implicancias para el uso de índices tafonómicos en análisis (paleo)ambientales. En este estudio se analizó la alteración tafonómica del braquiópodo Bouchardia rosea (Mawe), en la bahía de Ubatuba, litoral norte de São Paulo, Brasil. Los depósitos de conchillas de estos braquiópodos registran una edad media de 674 años AP. La hipótesis subyacente al análisis fue que los índices tafonómicos no están uniformemente distribuidos a lo largo del gradiente batimétrico evaluado. Las conchillas se muestrearon mediante una draga puntal tipo Van Veen (31 x 40 cm) a lo largo de un gradiente batimétrico de 0 (playa) a 35 m de profundidad, incluyendo 14 estaciones de muestreo. Se reconocieron cuatro ambientes sedimentarios: (a) playa, (b) bahía protegida de fondo arcilloso, (c) bahía protegida de fondo arenoso, y (d) plataforma interna de fondo arenoso. Los braquiópodos de cada una de las estaciones (incluida la fracción total de fragmentos) fueron investigados siguiendo un protocolo que contiene 10 índices tafonómicos. Seis de estos índices (fragmentación, alteración de borde, alteración del color, bioerosión e incrustación) se distribuyen homogéneamente entre los depósitos de los ambientes estudiados, indicando que estos índices tafonómicos no son sensibles a las condiciones ambientales en este caso. Por otro lado, la textura de la conchilla mostró un alto potencial para la interpretación ambiental, pareciendo útiles como una herramienta de análisis de tafofacies. Varios ejemplos de generalizaciones tafonómicas disponibles en la literatura no son comparables ni aplicables a nuestros resultados, confirmando además que las generalizaciones de las normas y modelos aplicables a todos los casos sin restricción deben evitarse en los estudios tafonómicos.
Keywords. Bouchardia rosea; Brachiopods; Taphonomic signatures; Shell texture; Ubatuba Bay; Holocene.
Palabras clave. Bouchardia rosea; Braquiópodos; Firmas tafonómicas; Textura de la conchilla; Bahía Ubatuba; Holoceno.
Introduction
Benchmark studies in taphonomy (Davies et al., 1989; Staff and Powell, 1990; Kowalewski et al., 1994; Kidwell et al., 2001) during the last two decades have demonstrated that taphofacies and time-averaging are fundamental parameters for paleoenvironmental analyses (Walker and Bambach, 1971; Brett and Baird, 1986; Speyer and Brett, 1986, 1988; Kidwell and Bosence, 1991; Kidwell et al., 2001; Flessa and Kowalewski, 1994; Kowalewski et al., 1998; Behrensmeyer et al., 2005). Taphonomic signatures (sensu Fürsich and Oschmann, 1993) such as bioclast fragmentation (= skeletal remains) may vary in a predictable fashion along environmental gradients (e.g., shallow to deep water). Thus, taphonomic data (e.g., taphofacies) are powerful tools in stratigraphic studies (see Holz and Simões, 2004, for a review), providing independent information, especially when geochemical and sedimentological data are lacking or dubious. However, recent taphonomic studies in modern marine settings have shown that some taphonomic signatures can be generated by several distinct mechanisms (e.g., Best and Kidwell, 2000a, b; Kidwell et al., 2001; Callender et al., 2002; Staff et al., 2002; Nielsen, 2004), and so the end products may be hard to distinguish (Fürsich and Oschmann, 1993). Indeed, as shown in detail by various authors (see Tomašových and Zuschin, 2009, and references therein), variation in skeletal preservation of brachiopods are not linked only to environmental variables (extrinsic factor), but also to shell-specific (intrinsic), methodological (analytical), and temporal (time-averaging) factors.
In addition to the issues mentioned above, other factors may account for loss of accuracy of taphofacies analysis and extrapolation of present-day data to the fossil record. For instance, taphonomic knowledge is still limited to certain environments (i.e., continental shelves), taxa (mostly mollusks) and climatic zones (subtropical). These issues have been recognized by a number of authors (Kidwell, 1998; Kowalewski et al., 2003) that eloquently emphasized the need for additional studies on death assemblages formed from biologic remains other than mollusks. Indeed, studies dealing with the taphonomy of Holocene rhynchonellid and terebratulid brachiopods became available only very recently (Tomašových and Zuschin, 2009). These authors, however, centered the data in brachiopod death assemblages found in shallow water carbonate environments of tropical/subtropical settings in the Red Sea and the Gulf of Aden. Contrarily, in this paper we investigated the taphonomy of shells (surface death assemblages) of the terebratellidine brachiopod Bouchardia rosea (Mawe) in low-latitude, siliciclatic-dominated, shallow settings of the inner portion of the Brazilian shelf (western South Atlantic Ocean). Hence, this study focused on a taxonomic group (living terebratellidine brachiopods) from a geographic region that has received little attention from taphonomists.
Background
Bouchardia rosea (Mawe), the most abundant living rhynchonelliform brachiopod in the southern sector of the Brazilian shelf, was reported to occur in coarse bioclastic-rich substrates in shallow marine water (= photic zone) (Tommasi 1970; Kowalewski et al., 2002; Simões et al., 2004). Shells of B. rosea are calcitic, endopunctate (organic-rich) with a thin prismatic primary layer, a thick fibrous-fabric secondary layer, and no tertiary layer. The prismatic layer is twice as thick as the fibrous one in the ventral valve, and the fibrous layer is ten times thicker than the prismatic one in the case of the dorsal valve. Shells of B. rosea have tightly interlocked hinges, with the animal lying on the sediment/water interface resting on the dorsal valve (Simões et al., 2007a). Juvenile specimens are found attached to granules and other sedimentary particles (Simões et al., 2004), but adults are free-lying (Simões et al., 2007a, p. 143, figure 1). Hence, B. rosea shells are prone to be transported and sorted by waves, currents and storm flows (see Simões et al., 2007b) and/or bored, encrusted, or corroded by taphonomic agents present in the taphonomic active zone (TAZ, Davies et al., 1989).
Species belonging to this genus were also found in Paleogene and Neogene assemblages from South America and Antarctica (see Manceñido and Griffin, 1988; Rodrigues et al., 2008); therefore, taphonomic study of B. rosea shells can help paleontologists better understand the sedimentological and paleoecological data locked in such fossil concentrations (see also Simões et al., 2005).
Study Area
The study area is located in Ubatuba Bay, on the northern coast of São Paulo State (23º - 24ºS and 44º -46ºW, figure 1). This region is within a transition zone between humid subtropical and tropical climates. It is influenced by tropical and equatorial water masses, but suffers incursions of polar water masses too. Ubatuba Bay has eight small beaches bordered by promontories of igneous and metamorphic rocks. The bay mouth is oriented eastwards. The area covered by the bay is approximately 8 km2 and its width is 4.5 km, tapering landwards.
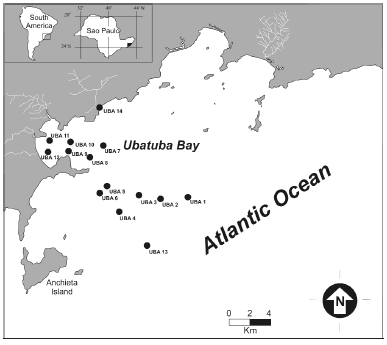
Figure 1. Location map of stations sampled in Ubatuba region, northern coast of São Paulo State, Brazil / mapa de ubicación de las estaciones de muestreo en la región de Ubatuba, costa norte del estado de São Paulo, Brasil.
Siliciclastic sediments (fine to very fine sands) predominate (table 1) along the northern coast of São Paulo State. Coarse, well-sorted sediments are present in the outer portions of the bay. These coarser sediments are probably related to the reworking of relict deposits during storm events (Rodrigues et al., 2002). However, in some areas, especially near islands and rocky shores, bioclast-rich sediments mostly made-up of shells of B. rosea are common.
Table 1. Data from the stations sampled in Ubatuba Bay, northern coast of São Paulo State, Brazil / información de las estaciones de muestreo en la Bahía de Ubatuba, costa norte del estado de São Paulo, Brasil.
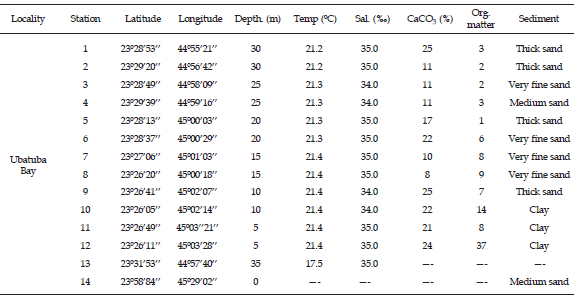
In the study area, four distinct sedimentary environments were recognized on the basis of substrate type, depth, and hydrodynamic conditions: (a) beach, particularly Itamambuca Beach (12 km north of the city of Ubatuba) (figure 1), characterized by two distinct sedimentary regimes (dissipative to the south and reflexive to the north). Beach sediments are composed of coarse (33.3%), medium (55.6%) and fine (11.1%) sands, which are well (22.2%) to moderately well (77.8%) sorted (Souza, 1997); (b) inner shelf, muddy, organic-rich bottoms (5 to 10 m deep); (c) inner shelf, sandy bottoms (10 to 20 m deep); and (d) outer shelf, sandy bottoms, over 20 m deep (figure 1).
Material and methods
Sampling procedures
A total of 14 collecting stations were sampled at five meter intervals (5—35 m water depth). The geographic position and depth of each station was determined using GPS and an ecobathymeter (figure 1, table 1). Sea floor was sampled (at least eight liters of sediment) using both a 300x400 mm dredge and a 310x400 mm Van Veen grab sampler (=0.12m2).
Brachiopod shells from Itamambuca beach were sampled in the foreshore zone too, using randomly distributed quadrants of nearly one square meter. A shovel was used to collect the surface layers of beach sediments (up to 30 mm). Bioclasts present in the quadrants were carefully picked, independently of their taphonomic state.
After sampling, sediments from each station were sieved through eight, six and two milllimeter meshes, and dried. Five sample localities (UBA 1, UBA 4, UBA 5, UBA 9, UBA 14) had sufficient material of B. rosea to conduct taphonomic analyses. For taphonomic analyses, subsamples of 150 shells per sieve fraction were obtained via sample splitting (see Kidwell et al., 2001) and all sieve fractions were analyzed.
Out of the 14 sampling stations along the bathymetric gradient, only five stations (UBA 1, UBA 4, UBA5, UBA 9 and UBA 14) yielded a sufficient number of brachiopod shells for taphonomic analysis.
Taphonomic protocol
The taphonomic protocol employed in this study is based on Davies et al. (1989), Best and Kidwell (2000a), Kidwell et al. (2001), Callender et al. (2002), Staff et al. (2002), Rothfus (2004), and Rodrigues et al. (2009). In order to avoid the so-called observer error (see Rothfus, 2004), taphonomic observations were carried out by one person, using a stereomicroscope at 10x magnification. Taphonomic signatures were determined for each bioclast (including fragments), and SEM analyses were also used for some specimens. Chi-Square analysis was applied to all taphonomic signatures (p=0.05).
Our protocol examined ten taphonomic signatures (sensu Fürsich and Oschmann, 1993) as presented in table 2, including: (1) articulation, (2) valve type, (3) fragmentation, (4) abrasion, (5) dissolution, (6) edge chipping, (7) color alteration, (8) bioerosion, (9) encrustation, and (10) superficial shell texture.
Table 2. Taphonomic signatures and their states (condition), investigated for brachiopod shells / índices tafonómicos de las valvas de braquiópodos y sus estados (condiciones) analizados.

States of each taphonomic signature (table 2) were determined. Articulation distinguished disarticulated valves from articulated specimens (table 2, figures 2.1-3). For disarticulated shells, valve type (dorsal or ventral, table 2, figures 2.2-3) was identified when possible. Fragmentation was established by tallying breaks independent of their origin (physical, chemical and/or biological). Fragmented valves denote shells with less than 90% of its original shell material preserved.
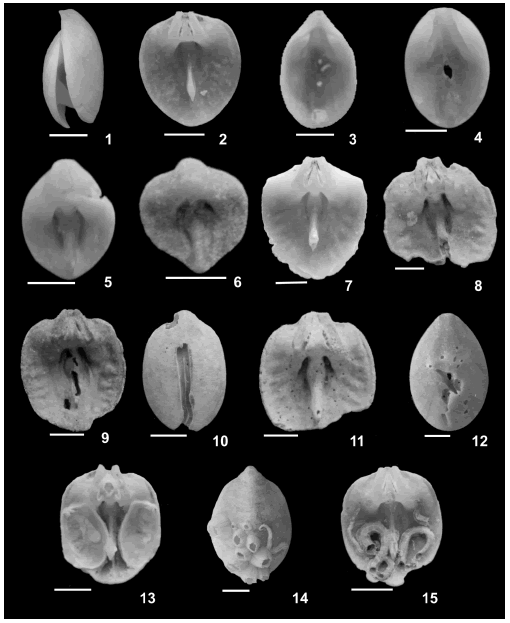
Figure 2. Bouchardia rosea (Mawe) shells showing different taphonomic signatures / valvas mostrando los diferentes índices tafonómicos. 1, articulated shell / valva articulada; 2, dorsal disarticulated valve / valva dorsal desarticulada; 3, ventral disarticulated valve / valva ventral desarticulada; 4-5, abrasion / abrasión; 6, dissolution / disolución; 7, valve with irregular edge / valva con borde irregular; 8-9, shell color alteration / valva con alteración de color; 8, bleaching / blanqueadas; 9, gray / gris; 10-12, bioerosion by polychaetes / bioerosión por poliquetos; 11-12, sponge attacks / ataque de esponjas; 13-15, encrustation by bivalve mollusks, barnacles and serpuliids / incrustaciones por moluscos bivalvos, cirripedios y poliquetos. Scale bar / escala gráfica = 1cm.
Degree of abrasion (table 2, figures 2.4-5) was divided into three states: natural or without abrasion (state 0); eroded shells with polished morphological features (state 1), and intensely eroded with shell features (teeth, umbones, ornamentation) completely abraded (state 2). Some shells yielding state 2 abrasion may have erosional holes on the most convex part of the valves. Degree of dissolution (table 2, figure 2.6) also was tallied using three states: natural or with no dissolution (state 0); partially dissolved shells with isolated dissolution foci present only on the outermost shell layers (state 1); highly dissolved with prominent shell structures (ornament) lost and dissolution holes present (state 2). Although abrasion and dissolution have distinct origins, they produce similar taphonomic alterations. In other words, both abrasion (physical agents) and dissolution (chemical/biological process) can create holes and rounding of edges and other surface relief (see figures 2.4-6). Distinguishing between abrasion and dissolution signatures can be difficult. However, shell color alteration and the superficial shell textures were useful in recognizing abraded vs. dissolved shells (see below) in many doubtful cases.
Shell edge alteration (table 2, figure 2.7) was tallied as unaltered, chipped or rounded (see Davies et al., 1989; Kowalewski et al., 1994; Best and Kidwell, 2000a; Callender et al., 2002 e Staff et al., 2002). Color alteration patterns of brachiopod shells (table 2, figures 2.8-9) are the same as those described in the literature for mollusk shells (e.g., Cutler, 1995; Callender et al., 2002; Staff et al., 2002), including white, gray, brown and black colors.
In the case of bioerosion (figures 2.10-12) and encrustation (figures 2.13-15), the organisms causing alteration (bioerosion or encrusting agents) and the proportion of shell area affected by those processes (<10% shell, between 10% and 50%, and >50%, table 2) were both tallied. These signatures were investigated by considering the interior and exterior of each shell.
Shell surface textures (see Cutler, 1987, 1995; Nielsen, 2004) were grouped into five categories based on taphonomic process as follows: (a) natural texture, (b) biogenic textures, (c) physical textures, (d) chemical textures, and (e) palimpsest textures (table 2, figure 3). An unaltered valve surface is tallied as a natural texture (figures 3.1-2). Among biogenic textures, four distinct types were recognized: (a) gallery texture (figures 3.3-4), when shells have Caulostrepsis-like tubes; (b) cratered texture (figures 3.5-6), in shells with numerous, large, vertical holes (>2mm); (c) perforated texture (figures 3.7-8), for specimens with small (>2mm), vertical holes; (d) etched texture (figures 3.9-10), in shells with several sinuous, ramified etched areas, usually showing a radial distribution—generally produced by filaments of algae and fungi (fixation sites on the shell, see Cutler, 1995 for similar examples in bivalve mollusk shells). Perforated and cratered textures were produced by attacks or infestation by sponges (Cliona, see Cutler, 1995 for similar examples in bivalve mollusks), whereas polychaetes were responsible for gallery textures (Polydora, see Rodrigues et al., 2008).

Figure 3. Superficial texture of Bouchardia rosea shells / textura superficial de las valvas. 1, natural texture / textura natural; 2, detail of natural texture / detalle de la textura natural; 3, gallery texture / textura en galería; 4, detail of gallery texture / detalle de la textura en galería; 5, cratered texture / textura poceada; 6, detail of cratered texture / detalle de la textura poceada; 7, perforated texture / textura perforada; 8, detail of perforated texture / detalle de la textura perforada; 9, etching texture / textura en "etching"; 10, detail of etching texture / detalle de la textura en "etching"; 11, granular texture / textura granular; 12, detail of granular texture / detalle de la textura granular; 13, exfoliated texture / textura exfoliada; 14, detail of exfoliated texture / detalle de la textura exfoliada; 15, palimpsest texture showing the overprinting of granular and exfoliated textures in the same shell / textura palimpsesto mostrando la superposición de las texturas granular y exfoliada en la misma valva; 16, detail of palimpsest texture / detalle de la textura palimpsesto. Scale bar / escala gráfica = 1 mm.
Chemical texture renders a granular texture (figures 3.11-12) characterized by a chalky appearance. It results from loss of the primary shell layer and separation of radial-fibrous mineral crystals in the secondary shell layer. Physical texture is represented by exfoliation (figures 3.13-14) and includes shells with abraded surfaces that expose the secondary shell layer in the most convex portions of valves (umbones and beaks). Finally, palimpsest textures denote over-printing of all above mentioned textures on a single shell (figures 3.15-16).
Results
Beach environment (UBA 14)
All shells (n = 406) are disarticulated, and 94.8% are ventral valves. Fragmentation was observed in 15.6% of bioclasts and abrasion is high with 76.4% of abraded shells. Dissolution was also observed in 48.3% of valves. Most shells (76.1%) show edge modification, and color alteration is present in 45.1% of them. Shells are commonly bioeroded (62.8%) and encrusted (83%). The predominant superficial texture is exfoliated (occurring on 47.2% of shells), followed by granular (24.5%), biogenic (17.6%) and natural (10.7%) textures.
Inner shelf, sandy bottoms (UBA 9 and UBA 5)
Almost all shells (n = 637) were disarticulated (99% at UBA 9; 100% at UBA 5). Dorsal valves predominate (61%) at UBA 9 station, whereas ventral valves prevailed (74.8%) at UBA 5 station. Fragmentation is moderate (34%), as is abrasion (21.3% at UBA 9; 30.9% at UBA 5). Dissolution is intense, and observed in 56% (UBA 9) to 89% (UBA 5) of the examined specimens. Most bioclasts bear shelledge modification (86.3% at UBA 9; 96.8% at UBA 5) and color alteration (71.2% at UBA 9; 91% at UBA 5). Bioerosion is high (53.7%) at UBA 9, but low (7.7%) at UBA 5. Encrustation is present on most shells at both stations (50.3% at UBA 9; 89% at UBA 5). Granular (31% at UBA 9; 60% at UBA 5) and biogenic (18% at UBA 9; 0.6% at UBA 5) shell textures predominate.
Outer shelf, sandy bottoms (UBA 4 and UBA 1)
The vast majority of the shells (n = 564) are disarticulated (98%, UBA 4; 100%, UBA 1), and dorsal valves predominate (54% at UBA 4; 79.6% in UBA 1). Shell fragmentation is moderately frequent (23.7%, UBA 4 to 43.4%, UBA 1), whereas abrasion is low to moderate (17.5%, UBA 4 to 39.8%, UBA 1). Dissolution is intense, affecting over 80% of shells. Almost all bioclasts showed edge modification (94.7%, UBA 4 to 98%, UBA 1) and color alteration (90.0%, UBA 4 to 93.8%, UBA 1). Bioerosion varies from 22% (UBA1) to 44.2% (UBA 4), and encrustation is low to moderate (16%, UBA 4 to 33.6%, UBA 1). Granular texture predominates (40%, UBA 4 to 70.6%, UBA 1), followed by natural (24.7%, UBA 4 to 36.7%, UBA 1), biogenic (2.4%, UBA 4 to 16.6%, UBA 1) and exfoliated textures (2.3%, UBA 4 to 6.7%, UBA 1).
Discussion
Origin and (paleo)environmental implications of the studied taphonomic signatures
Signatures such as articulation, fragmentation, bioerosion, encrustation, abrasion, dissolution, color alteration, and edge modification, are typically employed in taphonomic studies to recognize biotic and abiotic conditions of the environment where bioclasts are deposited (e.g., Brett and Baird, 1986; Davies et al., 1989; Fürsich and Oschmann, 1986, 1993; Best and Kidwell, 2000a; Callender et al., 2002; Staff et al., 2002; Parsons-Hubbard, 2005; Best, 2008; Tomašových and Zuschin, 2009). However, as shown in figure 4, most of these taphonomic signatures have a nearly homogenous distribution along the bathymetric trend investigated in this case.

Figure 4. Frequency of taphonomic signatures at each sampled station / frecuencia de los índices tafonómicos en cada estación de muestreo. 1, disarticulation / desarticulación; 2, fragmentation / fragmentación; 3, shell color alteration / alteración de color; 4, shell edge alteration / alteración del borde de la valva; 5, bioerosion / bioerosión; 6, encrustation / encrustación; 7, abrasion / abrasión; 8, dissolution / disolución.
Valve disarticulation is, for instance, commonly used in taphonomic studies. Bouchardia rosea has a tightly interlocked hinge (see Brunton, 1996; Brunton et al., 1996) preventing wide opening and rotation of ventral and dorsal valves (Brunton, 1996, p. 234), even during life. Thus, under natural post-mortem conditions, articulation is favored in shells of this species. At first glance, an articulated condition seems to agree with that of Brett and Baird (1986): "...certain brachiopods (e.g., atrypids and rynchonellids) possessed interlocking hinge dentitions which enhanced articulation and increased the period of time during which they remained intact after death. Their disarticulation implies rough water conditions...". However, our results indicate that post-mortem disarticulation is extremely common in the study area even though Ubatuba Bay is protected from the action of the main S-SW wave sets of the region, due to the eastern orientation of the bay mouth. As a result, reworking of bottom sediments occurs only sporadically (Mahiques et al., 1998). Among our sampling sites, the beach environment (UBA 14) is the most energetic, with lower energy conditions to the south (Souza, 1997), where brachiopod assemblages are being reworked (Simões et al., 2007b). In other words, based on the intrinsic and extrinsic factors affecting the shells of Bouchardia rosea, high disarticulation indexes were expected only for the beach environment. Contrary to such expectation, valve disarticulation indexes are not significantly distinct among our sampled sites (Χ2 = 7.022; k = 3; p = 0.071). Indeed, valve disarticulation is not limited to beach facies. Stochastic variation in some variables such as shell transportation, physical and biological sediment reworking, and rates of sedimentation and shell residence time at the TAZ may account for the pattern recorded. Indeed, radiocarbon dates from more than 100 brachiopod shells (Bouchardia rosea) as well as cooccurring bivalve mollusk shells (Semele casali Doello-Jurado, Glycymeris sp.) from death assemblages at the same sites indicate that temporal mixing of hundreds to thousands years is a common phenomenon (Carroll et al., 2003; Krause et al., 2004; Simões et al., 2004; Barbour Wood et al., 2006; Krause et al., 2010). The average age of the studied shells is in the order of 674 YBP (Krause et al., 2010). Probably, prolonged residence time in the TAZ, associated with intermittent action of waves and currents (see Simões et al., 2007b) and intrastratal biological activity and predation (Simões et al., 2007a; Rodrigues et al., 2008) favors the disarticulation of Bouchardia rosea shells.
Fragmentation rates are also high at almost all stations (figure 4.2). In other words, fragmentation is not restricted to a particular subenvironment, and is not significantly different between UBA 4, UBA 5 and UBA 9 stations (Χ2 = 3.261; k = 2; p = 0.196) (see also Best and Kidwell, 2000a for a similar observation, with accumulations of bivalve mollusks from the Caribbean Coast of Panama). Fragmentation can be generated by several physical, chemical and/or biological processes (Zuschin et al., 2003). For example, fragmentation of shells found in the beach environment (UBA 14 station) is caused by onshore transport and reworking of shells during high energy events (storms, see Simões et al., 2007b). In the quiet waters of the inner shelf (UBA 9 station), fragmentation is mainly caused by the attack of durophagous predators, especially brachyuran crabs. In fact, Pires-Vanin et al. (1995) noted that intense predation events may occur as a result of activity of those crabs in Ubatuba Bay. These observations and the presence of wedge marks on the margins of shells, which are signs of brachyuran crab attacks (see Simões et al., 2001), indicate a biological origin for most fragmentation observed in shells of Bouchardia rosea recovered from this part of the bay. In contrast, fragmentation of shells is caused by intense dissolution in the outer shelf (UBA 1 station). These data agree with Fürsich and Oschmann (1993) and Zuschin et al. (2003) on the multiple origins of fragmentation and on the need of identifying the origin of this taphonomic feature. Hence, in our case study fragmentation per se has little or no interpretative value.
Encrustation and bioerosion rates are significantly different among studied sites (Χ2 = 661.31; k = 4; p < 0.001; Χ2 = 248.728; k = 4; p < 0.001, respectively), and commonly found in shells from sites where there seems to be a high primary productivity (UBA 9). Such is the case near some islands, probably because of either "the island mass effect" (Doty and Oguri, 1956) or else the high concentration of nutrients in those areas (Simões et al., 2009). Thus, our data agree with observations recently made by Tomašových and Zuschin (2009) on brachiopod shells of death assemblages in Red Sea and Gulf of Aden carbonatic settings. According to these authors, levels of shell encrustation and bioerosion in terebratulid brachiopods decrease with depth because of a drop in abundance of light-dependent borers, and heterotrophic borers and encrusters.
Superficial shell textures as a tool for (paleo)environmental analysis
Among the taphonomic signatures examined for this study, superficial shell textures seem to provide powerful proxies for (paleo)environmental interpretation. Shell textures are useful because some of them have a restricted environmental distribution and others can be overprinted on individual shells, offering valuable information about the taphonomic pathways or history for each bioclast. For example, bioclasts bearing exfoliated shell texture are commonly found at the beach site (UBA 14) and are rare in the other environments (figure 5.2). The exfoliated texture is a result of intense mechanical abrasion of the shells, and is noted in material exposed to intense reworking (UBA 14).

Figure 5. Frequency of shell superficial texture at each sampled station / frecuencia de las texturas superficiales de las valvas en cada estación de muestreo. 1, natural / natural; 2, exfoliated / exfoliada; 3, granular / granular; 4, biogenic / biogénica.
Although observed in shells from all environments, the granular texture is frequently recorded (more than 50%) in inner shelf sandy bottoms (figure 5.3). Granular textures are a result of chemical dissolution. Shells with granular texture have a chalky appearance with loss of original color and luster. Observations in modern marine environments and laboratory experiments with shells of Bouchardia rosea suggest that granular texture is a result of dissolution by interstitial water (Rodrigues et al., 2006; Simões et al., 2007b). As noted by Burone (2002) in the study area, inner-shelf sediments below 10 cm depth are organic-rich and anoxic, favoring shell dissolution.
Shells yielding biogenic shell textures are found in high proportions (more than 50%, figure 5.4) on the inner-shelf sandy bottoms of Ubatuba Bay (UBA 9 and UBA 4 stations, respectively). These textures are produced by several biological agents, including the attack or infestation by sponges, polychaetes and algae. Based on observations in shallow and deep marine environments, and also on laboratory results, some authors have suggested that infestations may occur in very short periods (up to 15 days, see Cutler, 1995). However, experimental observations by Asgaard and Bromley (1991) in the Mediterranean indicate that a bioclast must remain in the TAZ with no significant reworking for several years, in order to be infested. For shells of Bouchardia rosea, gallery textures result from infestation by polychaetes when the brachiopod is still alive. Hence, this signature is not a faithful indicator of prolonged exposure of the shells to environmental conditions in the TAZ (see Rodrigues et al., 2008).
Determining the taphonomic pathway of a single shell
Palimpsest shell texture is a noteworthy taphonomic signature exhibited by shells of B. rosea. Palimpsest textures are a result of the overprinting of new shell textures over the old ones (see Cutler, 1995, for a similar phenomenon in Recent bivalve shells from the Gulf of California). Although many taphonomic processes may occur concomitantly in the shells of a given environment, some may occur preferentially only under certain environmental conditions (mechanical abrasion, dissolution). High durability and long residence time in the TAZ favor the imprinting of some of these environment-specific signatures on certain shells. Indeed, shells of B. rosea in the studied bioclastic concentrations are time-averaged in orders of magnitude of 102 to 104 years (Carroll et al., 2003; Barbour Wood et al., 2006; Krause et al., 2010), thus contributing to the development of palimpsest shell textures.
Palimpsest shell texture can provide robust evidence of the taphonomic pathway followed by individual shells; a shell specimen (DZP 17538, see figure 3.8) will be used here as an example of this. The specimen was sampled in the inner-shelf sandy bottom, at 20 m depth (UBA 5), and shows a palimpsest shell texture characterized by overprinting of exfoliated (physical) and granular (chemical) textures. As previously discussed, exfoliation is common on shells collected on the beach, and is a result of intense mechanical abrasion (see Simões et al., 2007b). Thus, exfoliation indicates that this specimen was in a beach environment for some time during its post-mortem history. However, the presence of granular texture exposing the prismatic outer shell layer and the fibrous inner layer indicates shell dissolution, which is commonly observed in those shells lying in sandy organic-rich substrates, a condition observed in inner-shelf areas such as station UBA 5. This indicates that during its taphonomic history this shell was transported onshore and laid on the beach where the exfoliated shell texture was acquired. Subsequently, this shell was transported offshore into organic-rich substrates, where the granular texture was acquired (see Simões et al., 2007b, for the onshore/offshore shell transportation in the study area).
Final remarks
As shown above, taphonomic signatures such as articulation, fragmentation, dissolution, bioerosion, and encrustation found in the studied brachiopod shells may have multiple origins. The usage of these signatures without previous investigations on their origin may blur taphofacies analysis. In our case study, superficial shell textures seem to carry the best potential for precise environmental interpretations. However, as well noted by Tomašových and Zuschin (2009), brachiopod shell preservation is not a product of steady state alteration processes over the course of their taphonomic history. Stochastic variation in some intrinsic (productivity, see Simões et al., 2009) and extrinsic (background and episodic sedimentation) factors in the course of temporal mixing—plus the fact that the studied brachiopod shells are durable and are present in a very particular siliciclastic environment—may better explain the taphonomic behavior of the studied shells. In other words, taphonomic patterns found in the brachiopod-rich accumulations of Ubatuba Bay are a product of particular (extrinsic and intrinsic) conditions and can not be extrapolated to all brachiopod fossil assemblages of the Phanerozic geological record. However, since bouchardiid shells are ubiquitous in various Paleogene and Neogene fossil concentrations of South America and Antarctica their use in paleoenvironmental analyses should be viewed under the light of the taphonomic data discussed in this contribution.
Acknowledgements
The authors are grateful to A. Fransozo (Institute of Biosciences, São Paulo State University - UNESP, Botucatu Campus, São Paulo State, Brazil) for the help during field studies and to Sr. Isaac Sayeg (Laboratory of Scanning Electron Microscopy, Institute of Geosciences, University of São Paulo -USP, São Paulo State, Brazil) for the help with SEM images. Thanks to Dr. Katia Facuri (Federal University of Uberlandia, Ituiutaba Campus, Brazil) for statistic analysis. This contribution was supported by the State of São Paulo Research Foundation - FAPESP (Processes 00/12659-7, 02/13552-7, 06/00169-1 and 06/59416-8) and National Council for Scientific and Technological Development - CNPq (Processes 300448/03-01, 306601/2006-0 and 154937/2006-1). The final version of this contribution was substantially improved, thanks to careful and constructive comments of D.G. Lazo and an anonymous reviewer.
References
1. Asgaard, U. and Bromley, R.G. 1991. Colonization by micromorph brachiopods in the shallow subtidal of eastern Mediterranean sea. In: D.I. MacKinnon, D.E. Lee and J.D. Campbell (eds.), Brachiopod through time, Proceedings of 2º International Brachiopod Congress (Dunedin): 261-264. [ Links ]
2. Barbour Wood, S.L., Krause, R.Jr., Kowalewski, M., Wehmiller, J.F. and Simões, M.G. 2006. Aspartic acid (Asp) racemization dating of Holocene calcitic brachiopods and aragonitic bivalves from the southern Brazilian shelf, South Atlantic. Quaternary Research 66: 323-331. [ Links ]
3. Behrensmeyer, A.K., Fürsich, F.T., Gastaldo, R.A., Kidwell, S.M., Kosnik, M.A., Kowalewski, M., Plotnick, R.E., Rogers, R.R. and Alroy, J. 2005. Are the most durable shelly taxa also the most common in the marine fossil record? Paleobiology 31: 607-623. [ Links ]
4. Best, M.M.R. 2008. Contrast in preservation of bivalve death assemblages in siliciclastic and carbonate tropical shelf settings. Palaios 23: 796-809. [ Links ]
5. Best, M.M.R. and Kidwell, S.M. 2000a. Bivalve taphonomy in tropical mixed siliciclastic-carbonate settings. I. Environmental variation in shell condition. Paleobiology 26: 80-102. [ Links ]
6. Best, M.M.R. and Kidwell, S.M. 2000b. Bivalve taphonomy in tropical mixed siliciclastic-carbonate settings. II. Effect of bivalve life habits and shell types. Paleobiology 26: 103-115. [ Links ]
7. Brett, C.E. and Baird, G.C. 1986. Comparative taphonomy: A key for paleoenvironmental reconstruction. Palaios 1: 207-227. [ Links ]
8. Brunton, C.H.C. 1996. The functional Morphology of the Recent brachiopod Bouchardia rosea. Acta Zoologica 77: 233-240. [ Links ]
9. Brunton, C.H.C., Alvarez, F. and Mackinnon, D.I. 1996. Morphological terms used to describe the cardinalia of articulate brachiopods: homologies and recommendations. Historical Biology 11: 9-42. [ Links ]
10. Burone, L. 2002. [Foraminíferos bentônicos e parâmetros físico-químicos da Enseada de Ubatuba, São Paulo: estudo ecológico em uma região com poluição orgânica. Tese de Doutorado, Instituto Oceanográfico, Universidade São Paulo, São Paulo, 279 pp. Inédito. [ Links ]].
11. Callender, W.R., Staff, G.M., Parsons-Hubbard, K.M., Powell, E.N., Rowe, G., Walker, S.E., Brett, C.E., Raymond, A., Carlson, D.D., White, S. and Heise, E.A. 2002. Taphonomic trends along a forereef slope: Lee Stocking Island, Bahamas. I. Location and water depth. Palaios 17: 50-65. [ Links ]
12. Carroll, M., Kowalewski, M., Simões, M.G. and Goodfriend, G.A. 2003. Quantitative estimates of time-averaging in brachiopod shell accumulations from a modern tropical shelf. Paleobiology 29: 382-403. [ Links ]
13. Cutler, A.H. 1987. Surface textures of shells as taphonomic indicators. Palaeontological Society Special Publication 2: 164-176. [ Links ]
14. Cutler, A.H. 1995. Taphonomic implications of shell surface textures in Bahia la Choya, northern Gulf of California. Palaeogeography, Palaeoclimatology, Palaeoecology 114: 219-240. [ Links ]
15. Davies, D.J., Powell, E.N. and Stanton, R.J.Jr. 1989. Taphonomic signature as a function of environmental process: shells and shell beds in a hurricane-influenced inlet on the Texas coast. Palaeogeography, Palaeoclimatology, Palaeoecology 72: 317-352. [ Links ]
16. Doty M.S. and Oguri, M. 1956. The island mass effect. Journal du Conseil International pour l'Exploration de la Mer 22: 33-37. [ Links ]
17. Flessa, K.W. and Kowalewski, M. 1994. Shell survival and time-averaging in nearshore and shelf environments: estimates from the radiocarbon literature. Lethaia 27: 153-165. [ Links ]
18. Fürsich, F.T. and Oschmann, W. 1986. Storm shell beds of Nanogyra virgula in the Upper Jurassic of France. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 172: 141-161. [ Links ]
19. Fürsich, F.T. and Oschmann, W. 1993. Shell beds as tools in basin analysis: the Jurassic Kachchh, western India. Journal of Geological Society of London 150: 169-185. [ Links ]
20. Holz, M. and Simões, M.G. 2002. Elementos fundamentais de Tafonomia. Editora da UFRGS, 240 pp. Porto Alegre [ Links ]
21. Kidwell, S.M. 1998. Time-averaging in the marine fossil record: overview of strategies and uncertainties. Geobios 30: 977-995. [ Links ]
22. Kidwell, S.M. and Bosence, D.W.J. 1991. Taphonomy and time-averaging of marine shelly faunas. In: P.A. Allison and D.E.G. Briggs (eds.), Taphonomy: releasing the data locked in the fossil record, Plenum Press, New York, pp. 115-209. [ Links ]
23. Kidwell, S.M., Rothfus, T.A. and Best, M.M.R. 2001. Sensitivity of taphonomic signatures to sample size, sieve size, damage scoring system, and target taxa. Palaios 16: 25-52. [ Links ]
24. Kowalewski, M., Flessa, K.W. and Aggen, J.A. 1994. Taphofacies analysis of Recent shelly cheniers (beach ridges), northeastern Baja California, Mexico. Facies 31: 209-242. [ Links ]
25. Kowaleski, M., Goodfriend, G.A. and Flessa, K.W. 1998. High-resolution estimates of temporal mixing within shell beds: the evils and virtues of time-averaging. Paleobiology 24: 287-304. [ Links ]
26. Kowalewski, M., Simões, M.G., Carroll, M. and Rodland, D.L. 2002. Abundant brachiopods on a tropical, upwelling-influenced shelf (Southeast Brazilian Bight, South Atlantic). Palaios 17: 274-283. [ Links ]
27. Kowalewski, M., Carroll, M., Casazza, L., Gupta, N.S., Hannisdal, B., Hendy, A., Krause, R.A., Labarbera, M., Lazo, D.G., Messina, C., Puchalski, S., Rothfus, T.A., Sälgeback, J., Stempien, J., Terry, R.C. and Tomaových, A. 2003. Quantitative fidelity of brachiopod-mollusk assemblages from modern subtidal environments of San Juan Islands, USA. Journal of Taphonomy 1: 43-65. [ Links ]
28. Krause, R.A., Barbour Wood, S.L., Wehmiller, J.F., Kowalewski, M. and Simões, M.G. 2004. A comparative analysis of time averaging for bivalves and brachiopods from a modern Tropical Shelf. Annual Meeting Geological Society of America (Denver), Abstracts with Programs 36:383. [ Links ]
29. Krause, R.A., Barbour Wood, S.L., Kowalewski, M., Kaufman, D.A., Romanek, C.S., Simões, M.G. and Wehmiller, J.F. 2010. Comparative time-averaging: age mixing among sympatric bivalves and brachiopods from modern tropical shelf. Paleobiology 36: 428-452. [ Links ]
30. Mahiques, M.M., Tessler, M.G. and Furtado, V.V. 1998. Characterization of energy gradient in enclosed bays of Ubatuba region, southeastern Brazil. Estuarine, Coastal and Shelf Science 47: 431-446. [ Links ]
31. Manceñido, M.O. and Griffin, M. 1988. Distribution and palaeoenvironmental significance of the genus Bouchardia (Brachiopoda, Terebratellidina): its bearing on the Cenozoic evolution of the South Atlantic. Revista Brasileira de Geociências 18: 201-211. [ Links ]
32. Nielsen, J.K. 2004. Taphonomy in the light of intrinsic shell properties and life habits: marine bivalves from the Eemian of northern Russia. Paläontologische Zeitschrift 78: 53-72. [ Links ]
33. Parsons-Hubbard, K. 2005. Molluscan taphofacies in recent carbonate reef/lagoon systems and their application to sub-fossil samples from reef cores. Palaios 20: 175-191. [ Links ]
34. Pires-Vanin, AM.S., Jorge, J.P.S. and Sartor, S. 1995. Variação diária e sazonal da fauna bêntica de plataforma continental no litoral Norte do Estado de São Paulo. Publicação Especial do Instituto Oceanográfico 11: 107-114. [ Links ]
35. Rodrigues, M., Mahiques, M.M. and Tessler, M. 2002. Sedimentação atual nas enseadas de Ubatumirim e Picinguaba, região norte de Ubatuba, Estado de São Paulo, Brasil. Revista Brasileira de Oceanografia 50: 27-45. [ Links ]
36. Rodrigues, S.C., Neves, J.P.; Simões, M.G. and Kowalewski, M. 2006. Dissolution of Bouchardia rosea shells: implications for taphonomic bias in rhynchonelliform brachiopod assemblages. 9° Congreso Argentino de Paleontología y Bioestratigrafía (Córdoba), Resúmenes: 266. [ Links ]
37. Rodrigues, S.C., Simões, M.G.; Kowalewski, M.; Petti, M.A.V., Nonato, E.F., Martinez, S. and Del Rio, C.J. 2008. Biotic interaction between spionid polychaetes and bouchardiid brachiopods: paleoecological, taphonomic and evolutionary implications. Acta Palaeontologica Polonica 53: 657-668. [ Links ]
38. Rodrigues, S.C., Simões, M.G. and Pires-Domingues, R.A. 2009. Viés analítico em estudos tafonômicos com macroin-vertebrados: implicações (paleo)ambientais e (paleo)ecológicas. Geologia USP, Série Científica 9: 101-114. [ Links ]
39. Rothfus, T.A. 2004. Clams and Brachiopods: Chips that pass out of sight. Palaios 19: 507-513. [ Links ]
40. Simões, M.G., Marques, F.P.L., Fransozo, A. and Mello, L.H.C. 2001. Brachyuran (Xanthidae) crab predation on Bouchardia rosea (Brachiopoda, Terebratulida) and its paleobiological and taphonomic importance. Paleo-2001, Reunião Anual da SBP (Rio Claro), Boletim de Resumos: 14. [ Links ]
41. Simões, M.G., Kowalewski, M., Mello, L.H.C., Rodland, D. and Carroll, M. 2004. Recent brachiopods from the Southern Brazilian shelf: palaeontological and biogeographical implications. Palaeontology 47: 515-533. [ Links ]
42. Simões, M.G.; Rodrigues, S.C.; Leme, J.M. and Bissaro Jr., M.C. 2005. The settling pattern of brachiopod shells: stratigraphic and taphonomic implications to shell bed formation and paleoecology. Revista Brasileira de Geociências 35: 383-391. [ Links ]
43. Simões, M.G., Rodrigues, S.C. and Kowalewski, M. 2007a. Comparative analysis of drilling frequencies in Recent Brachiopod-Mollusk associations from the Southern Brazilian Shelf. Palaios 22: 143-154. [ Links ]
44. Simões, M.G., Rodrigues, S.C., Leme, J.M. and Pires-Domingues, R.A. 2007b. Brachiopod shells on the beach: taphonomic overprinting in a fair-weather shell accumulation and their paleoecological implications. Journal of Taphonomy 5: 205-225. [ Links ]
45. Simões, M.G., Rodrigues, S.C. and Kowalewski, M. 2009. Bouchardia rosea, a vanishing brachiopod species of the Brazilian platform: taphonomy, historical ecology and conservation paleobiology. Historical Biology 21: 123-137 [ Links ]
46. Souza, C.R.G. 1997. [As células de deriva litorânea e a erosão no Estado de São Paulo. Tese de Doutoramento, Instituto de Geociências, Universidade São Paulo, São Paulo, 358 pp. Inédito. [ Links ]].
47. Speyer, S.E. and Brett, C.E. 1986. Trilobite taphonomy and Middle Devonian taphofacies. Palaios 3: 312-327. [ Links ]
48. Speyer, S.E. and Brett, C.E. 1988. Taphofacies models for epeiric sea environments: middle paleozoic examples. Palaeogeography, Palaeoclimatology, Palaeoecology 63: 225-262. [ Links ]
49. Staff, G.M. and Powell, E.N. 1990. Onshore-offshore trends in community structural attributes: death assemblages from the shallow continental shelf of Texas. Continental Shelf Research 19: 717-756. [ Links ]
50. Staff, G.M., Callender, W.R., Powell, E.N., Parsons-Hubbard, K.M., Brett; C.E., Walker, S.E., Carlson, D.D., White, S., Raymond, A. and Heise, E.A. 2002. Taphonomic trends along a forereef slope: Lee Stocking Island, Bahamas. II. Time. Palaios 17: 66-83. [ Links ]
51. Tommasi, L.R. 1970. Sobre o braquiópode Bouchardia rosea (Mawe, 1823). Boletim do Instituto Oceanográfico 19: 33-42. [ Links ]
52. Tomaových, A. and Zuschin, M. 2009. Variation in brachiopod preservation along a carbonate shelf-basin transect (Red Sea and Gulf of Aden): environmental sensitivity of taphofacies. Palaios 24: 697-716. [ Links ]
53. Walker, K.R. and Bambach, R.K. 1971. The significance of fossil assemblages from fine-grained sediments: Time-averaged communities. Annual Meeting Geological Society of America Abstracts with Programs 3: 783-784. [ Links ]
54. Zuchin, M., Stachowitsch, M. and Stanton, R.J.Jr. 2003. Patterns and processes of shell fragmentation in modern and ancient marine environments. Earth-Science Reviews 63: 33-82. [ Links ]
Recibido: 27 de Julio de 2009.
Aceptado: 2 de Febrero de 2010.

