










Serviços Personalizados
Journal

Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAmeghiniana
versão On-line ISSN 1851-8044
Ameghiniana vol.47 no.4 Buenos Aires oct./dez. 2010
ARTÍCULOS ORIGINALES
The oldest lower Upper Cretaceous plesiosaurs (Reptilia, Sauropterygia) from southern Patagonia, Argentina
José Patricio O'Gorman1 and Augusto Nicolás Varela2
1División Paleontología Vertebrados, Museo de La Plata, Universidad Nacional de La Plata - CONICET. Paseo del bosque s/n 1900 La Plata, Argentina. joseogorman@fcnym.unlp.edu.ar
2Centro de Investigaciones Geológicas, Universidad Nacional de La Plata - CONICET, calle 1 Nº 644, 1900 La Plata, Argentina. augustovarela@cig.museo.unlp.edu.ar
Abstract. Plesiosaurs are recorded for the first time from the lower section of Mata Amarilla Formation, Santa Cruz Province, Patagonia, Argentina. The stratigraphic succession consists of mudstones and siltstones interbedded with medium to finegrained sandstone, deposited in a littoral environment during the Cenomanian-Santonian; therefore the material is the oldest record of plesiosaurs from the lower Late Cretaceous rocks of Argentina. The remains include teeth, some vertebrae, and one propodium assigned to Elasmosauridae indet. and Plesiosauria indet. The status of Polyptychodon patagonicus Ameghino, 1893, as well as its stratigraphic position are discussed, leading to the conclusion that the material described by Ameghino is probably from the Mata Amarilla Formation and can only be referred to Plesiosauria indet. Analysis of sedimentologic features suggests that the material described here was deposited in an estuar-ine environment, strongly influenced by tides. The characters of the inferred environment are consistent with the type of preservation of the materials.
Resumen. Los plesiosaurios (Reptilia, Sauropterygia) más antiguos del Cretácico Superior bajo de la Patagonia austral, Argentina. Se dan a conocer los primeros registros claramente referibles a plesiosau-rios de la sección inferior de la Formación Mata Amarilla en la provincia de Santa Cruz, Argentina. Los niveles portadores están compuestos por limolitas y arcillitas grises y negruzcas, que alternan con bancos de areniscas finas y medianas. Dichos niveles se depositaron en un ambiente litoral durante el intervalo Cenomaniano-Santoniano, siendo por tanto el registro más antiguo de plesiosaurios del Cretácico Superior de Argentina. Los restos hallados corresponden a dientes, vértebras y un propodio asignados a Elasmosauridae indet. y a Plesiosauria indet. Se discute el estatus de Polyptychodon patagonicus Ameghino, 1893 y su posición estratigráfica, concluyendo que el material descripto por Ameghino es sólo referible a Plesiosauria indet. y que proviene de la sección inferior de la Formación Mata Amarilla. El análisis sedimentológico indica que los materiales que se describen aquí fueron depositados en un ambiente estuarino con fuerte influencia mareal. Las características del ambiente de depositación de esta sección son consistentes con el estado de preservación de los materiales.
Key words. Plesiosaurs; Mata Amarilla Formation; Upper Cretaceous; Santa Cruz; Argentina.
Palabras clave. Plesiosaurios; Formación Mata Amarilla; Cretácico Superior; Santa Cruz; Argentina.
Introduction
Plesiosaurs are a monophyletic group of reptile that exhibit extreme adaptation to marine life (Brown 1981; O'Keefe, 2001). Cretaceous elements of this clade have been previously recorded in Campanian-Maastrichtian rocks in Patagonia (Chubut and Río Negro and Mendoza (Gasparini and de la Fuente, 2000; Gas parini et al., 2007; Lazo and Cichowolski, 2003; Salgado et al., 2007).
The record of Cretaceous plesiosaurs in Santa Cruz is restricted to a set of teeth which were used by Ameghino (1893) to describe a new species, i.e., Polyptychodon patagonicus Ameghino, 1893. His material was collected near Lago Argentino in beds included in Ameghino's "formación Santacruceña" (Ameghino, 1893, p. 82) and subsequently included in the "Sehuenense" stage (Ameghino, 1906).
The "Sehuenense" stage of Ameghino (1906) has been synonymized with the Mata Amarilla Formation (Cione et al., 2007; Varela et al., 2008; Varela, 2009). This unit was deposited during the Cenomanian-Santonian (Poiré et al., 2007; Varela and Poiré, 2008; Varela et al., 2008), an age based on stratigraphic relations. Subsequent authors disagreed with Ameghino's taxonomic identification of the teeth as Polyptychodon patagonicus (Cabrera, 1941; Welles, 1962). In a summary of South American plesiosaurs, Gasparini and Goñi (1985) mentioned Polyptychodon patagonicus, but expressing serious doubts about its inclusion among plesiosaurs.
The present contribution is the first report of material from the Mata Amarilla Formation that can be clearly referable to plesiosaurs. It is also the first lower Late Cretaceous record in Argentina. The purpose of this paper is to describe this new plesiosaur material and characterize and interpret the sedimentary paleoenvironments in which the remains were preserved. The equivalence of the lower section of the Mata Amarilla Formation and the "Sehuenense", and the taxonomic status of Polyptychodon patagonicus Ameghino, 1893, are also discussed.
Geological setting
The Austral Basin, also known as Magallanes Basin, is located in the southwestern region of the South American plate and comprises an area of ap-proximately 230,000 km2, covering the southern end of the Argentinean and Chilean territories. The basin is elongated in a N-S direction. The eastern edge is parallel to the Chico River and runs into the Atlantic Ocean as the "Rio Chico Dorsal" or "Dungeness Arch". The western tectonic edge is the Patagonian-Fueguian Andes, and the southern boundary is the Scotia plate (figure 1).
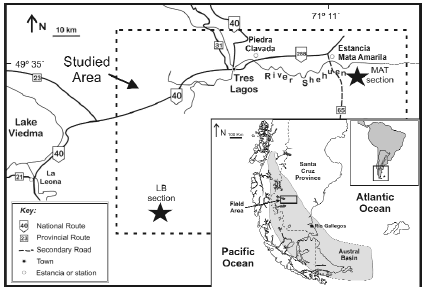
Figure 1. Geographic location of the Austral Basin and study area / ubicación geográfica de la Cuenca Austral y el área de estudio.
The geological history of the Austral Basin is related to three main tectonic stages. The first is the rift stage, the second is the thermal subsidence stage or sag, and the third one corresponds to the foreland stage (Arbe, 1989, 2002; Biddle et al., 1986). The Mata Amarilla Formation was deposited during the foreland stage, more precisely during the second episode of deformation, which could be related to the closure of the marginal Rocas Verdes Basin (Biddle et al., 1986).
The "Mata Amarilla Strata" (Feruglio, in Fossa Mancini et al., 1938) or Mata Amarilla Formation (Bianchi, 1967; Leanza, 1972; Russo and Flores, 1972) corresponds to Ameghino's "Sehuenense" (1906) and is one of the most representative units of the Upper Cretaceous of the Austral Basin. The Mata Amarilla Formation is up to 350 m thick, consisting of siltstones, and grey and blackish mudstones, interbedded with thin whitish sandstones deposited in coastal and continental environments (Russo and Flores, 1972; Russo et al., 1980; Arbe, 1989, 2002; Poiré et al., 2004; Varela and Poiré, 2008). The type section is located South of the Shehuen or Chalía River, in the vicinity of the Estancia Mata Amarilla (Estancia La Soriana), 23 km east from the town of Tres Lagos. The Mata Amarilla Formation overlies the Piedra Clavada Formation; the contact between them is transitional. It underlies the La Anita Formation (Varela and Poiré, 2008) (figure 2). The unit can be divided into three sections: a lower littoral section, a middle continental one, and an upper section that ranges from littoral to continental depending on the location within the study area (Varela, 2009). This unit was deposited during the early Late Cretaceous and spans the Cenomanian-Santonian (Poiré et al., 2007; Varela and Poiré, 2008; Varela, 2009) (figure 2).

Figure 2. Chronostratigraphic setting of the Austral Basin modified from Poiré et al., 2006 and Varela et al., 2008b / esquema cronoestratigráfico de la Cuenca Austral modificado de Poiré et al., 2006 y Varela et al.,2008b.
The area where the plesiosaurs were discovered is located in the southwestern corner of Santa Cruz Province (Patagonia, Argentina), east of Lake Viedma and near the town of Tres Lagos and the "Parador La Leona" (figure 1). This study was carried out in the lower section of the Mata Amarilla Formation, at its type locality (MAT section) and at Estancia La Blanca (LB section) (figure 1). The section at Estancia Mata Amarilla is located near the margin of the basin, while the one at Estancia La Blanca is located in the central area (Varela and Poiré, 2008).
In addition to the described plesiosaurs, the lower section of the Mata Amarilla Formation also bears the bivalves Exogyra guaranitica (Ihering, 1899), Cor-bula sehuena Ihering, 1907 and the gastropod Potamides patagoniensis Ihering, 1897 (Ihering, 1907, Wilckens, 1907, Bonarelli and Nágera, 1921; Feruglio, 1936, 1938). The presence of one specimen of the Santonian ammonite Placenticeras sp. has been mentioned for this formation, specifically at the locality of Cerro Índice (Blasco et al., 1980). However, the exact stratigraphic position was not determined as this genus was never reported again in the studied area.
Lungfish, amphibians, aquatic turtles, and crocodilians have been also recorded in the Mata Amarilla Formation (Cione et al., 2007; Goin et al., 2002). The middle and upper sections of the Mata Amarilla Formation are dominated by distinct continental tetrapods such as the ornithischian Talenkauen santacrucensis Novas, Cambiaso, and Ambrosio, 2004, the saurischian theropod Orkoraptor burkei Novas, Ezcurra, and Leucona, 2008, and the sauropod Puertasaurus reuili Novas, Salgado, Calvo, and Agnolin, 2005 (Lacovara et al., 2004; Novas et al., 2004a, 2004b, 2005, 2008).
Angiosperms and a few algae have been recorded in this formation too (Arrondo, 1983). Recently, Iglesias et al. (2007) described an abundant and diverse taphoflora dominated by angiosperms deposited in two levels, corresponding to the lower and middle sections of the Mata Amarilla Formation.
Material and methods
The material described in this paper consists of isolated teeth and bones with evidence of transport.
Welles's indices were used in the description of the vertebral centra (Welles, 1952). This index first considers the vertebral length in millimeters (L), the ratio between height (H) and length (H/L) and finally, the ratio between breadth (B) and length (B/L). Measurements of the height and breadth are taken on the posterior articular face of the centrum. Both (B/L) and (B/L) are given as percentages. The three values are always given in the same order (L, H/L, B/L). For propodial elements, we used the breadth / length index (B:L index), which is equal to the ratio between the anteroposterior distal length, and the proximodistal length expressed as percentage (Welles, 1952). The teeth were analyzed at the SEM facility in La Plata Museum (Jeol JSM-6360LV low vacuum SEM).
Two vertical lithostratigraphic sections at a scale of 1:100 were logged (MAT and LB section, see figure 3), describing the sedimentary facies (lithology and sedimentary structure). Sedimentary units were defined by means of facies association analysis and lithosome arquitecture, in order to interpret the sedimentary palaeoenvironments. Special emphasis was given to the lower section of the Mata Amarilla Formation, where the plesiosaur remains described in this paper were collected.
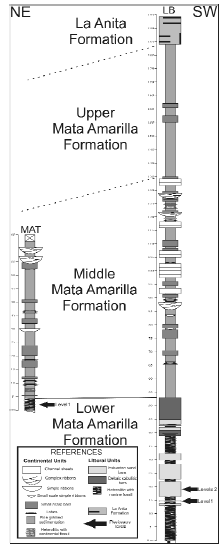
Figure 3. Schematic sedimentolgical columns of sedimentary units (facies association and lithosome architecture) of the Mata Amarilla Formation. Location of the levels with plesiosaurs are indicated with black arrows / perfiles esquemáticos de las unidades sedimentarias (asociaciones de facies y arquitectura de los litosomas) de la Formación Mata Amarilla. La ubicación de los niveles con plesiosaurios se indica con flechas negras.
Institutional abbreviations. ANSP, Academy of Natural Sciences of Philadelphia, USA; CM Zfr, Canterbury Museum, Christchurch, New Zealand; MACN, Museo Argentino de Ciencias Naturales "Bernardino Rivadavia", Ciudad Autónoma de Buenos Aires, Argentina; MLP, Museo de La Plata, La Plata, Buenos Aires Pro-vince, Argentina; MML, Museo Municipal de Lamarque, Río Negro Province, Argentina; MPEF, Museo Paleontológico Egidio Feruglio, Chubut Province, Argentina; MPM, Museo Regional Provincial Padre Manuel Jesús Molina, Río Gallegos, Santa Cruz Province, Argentina.
Systematic paleontology
Subclass Sauropterygia Owen, 1860
Order Plesiosauria Blainville, 1835
Material. MPM-PV 1871-2 (figure 4.1-2), one cervical vertebra; MPM-PV 1871-1-1, 54 teeth; MPM-PV 1871-1-2, one tooth; MPM-PV 1871-1-3 (figure 5), one tooth; MPM-PV 1870-2 (figure 4.3), one cervical vertebra; MPM-PV 1870-3 (figure 4.7-9), one pectoral vertebra; MPM-PV 1869-1 (figure 4.4-6), one anterior dorsal vertebra; MPM-PV 1869-2, one fragment of dorsal? vertebra; MPM-PV 1870-1(figure 4.10-12), one propodium.
Locality and horizon. All specimens are from the Mata Amarilla Formation (Cenomanian-Santonian), southwestern Santa Cruz Province, Patagonia, Argentina. Localities of collections are as follows: type locality of the Mata Amarilla Formation (MAT section): MPM-PV 1869-1 (figure 4.4-6) and MPM-PV 1869-2; Estancia La Blanca, East of Lake Viedma, near the town of Tres Lagos and the "Parador La Leona"(LB section) Level 1: MPM-PV 1870-2 (figure 4.3), MPM-PV 1870-3 (figure 4.7-9), and MPM-PV 1870-1 (figure 4.10-12); Level 2: MPM-PV 1871-1-1; MPM-PV 1871-1-2; MPM-PV 1871-1-3, and MPM-PV 1871-2 (figure 4.1-2).
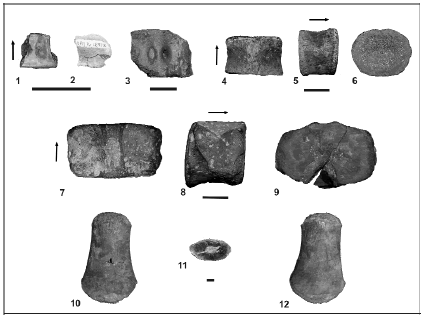
Figure 4. 1-12, Plesiosaur vertebrae and propodium from the Mata Amarilla Formation, Santa Cruz Province, Argentina / vértebras y propodio de plesiosaurios provenientes de la Formación Mata Amarilla, Santa Cruz, Argentina. 1-2, (MPM-PV 1871-2). 1, cervical vertebra in dorsal view, 2, posterior view / 1, vértebra cervical en vista dorsal y, 2, vista posterior; 3 (MPM-PV 1870-2) cervical vertebra in ventral view / vértebra cervical en vista ventral. 4-6 (MPM-PV 1869-1). Anterior dorsal vertebra in dorsal (4) right lateral (5) and ventral (6) views / vértebra dorsal anterior en vista dorsal (4) lateral derecha (5) y posterior (6) .7-9 (MPM-PV 1870-3). Pectoral vertebra in dorsal (7), lateral (8), and anterior (9) views / vértebra pectoral en vista dorsal (7), lateral (8) y anterior (9). 10-12 (MPM-PV1870-1). Propodium in dorsal (10), ventral (12) views, and in cross section (11) / propodio en vista dorsal (10), ventral (12) y en sección transversal (11).
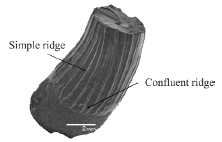
Figure 5. MPM-PV 1871-1-3. Plesiosauria indet. tooth in axial view / diente de Plesiosauria indet. en vista axial. Scale bar 2 mm / escala 2 mm
Family Elasmosauridae Cope, 1869 Elasmosauridae indet.
MPM-PV 1871-2 is a cervical vertebra with no neural arch or ribs (figure 4.1-2). The proportions of the vertebral body are 16, 106:150; hence, it is higher than long and wider than high. One foramen at the base of the neural channel connects to a large cavity inside the body. The body is 9 mm long, 8 mm high, and more than 14 mm wide. The facets of articulation for the neural arch are depressed and extend anteroposteriorly (figure 4.1). The articular face is slightly depressed with a slight dumbbell-shape as the ventral notch is not very pronounced (figure 4.2). A small foramen is observed ventrally. The body is damaged, and it seems likely that originally there may have been two foramina.
This vertebral body shows juvenile features, such as small size and lack of fusion between the neural arch and ribs to the body. The dumbbell-shaped face is considered as a synapomorphy of Elasmosauridae (Tarlo, 1960; Sato and Storrs, 2000). Although proportions of MPM-PV 1871-2 are not the typical of elasmosaurids, the difference could be due to the position along the neck or more probably to ontogenetic changes as already recorded in plesiosaurs (O'Keefe, 2006).
Plesiosauria indet.
MPM-PV 1871-1-1, 54 teeth; MPM-PV 1871-1-2, one tooth; MPM-PV 1871-1-3 (figure 5), one tooth. These isolated teeth are mostly incomplete because their roots are broken, and some lack the apex of the crown. In some cases, only crown fragments were collected. Ornamentation is clearly visible in some, but quite unclear in others. Teeth height range is 10-25 mm, whereas the diameter at the boundary between the crown and the root measures 3-10 mm.
Teeth are curved and thus present a concave (lingual) and a convex (labial) face. Their surface is covered by striae (about 20 per tooth). The striation is more pronounced on the lingual side, softer and shorter on the labial side (figure 5), or almost absent in some teeth. The striation arises at the base of the crown or more distally. In some cases, two striae converge near the boundary between the crown and the root.
Descriptions and illustrations of Cretaceous plesiosaur teeth from Patagonia are scarce. In the polycotylid Sulcusuchus erraini Gasparini and Spalletti, 1990, (MPEF: 650) from the La Colonia Formation, late Campanian-early Maastrichtian, Chubut Province (Gasparini and de la Fuente, 2000), teeth are acuminate, curved and with striae. One anterior tooth shows abrasion and lacks striation on the labial surface. These characteristics are compatible with the morphology of the material from Estancia La Blanca, from the lower section of the Mata Amarilla Formation. Isolated plesiosaur teeth were also recorded in the Loncoche Formation from CalmuCo, Mendoza Province, and assigned to the late Campanian-early Maastrichtian (Previtera and González Riga, 2008). These teeth and those collected at Estancia La Blanca share the shape and the presence of striae. In a tooth from CalmuCo, the striation is absent on the labial side as in the teeth of Estancia La Blanca. It appears that tooth morphology found in specimens from Estancia La Blanca is common among plesiosaurs and has been previously recorded in Argentina.
Tooth features have been used in Plesiosauria systematics (Bardet et al., 1999; O'Keefe, 2001; Smith, 2003). The most commonly used features are the shape, the presence of flat faces, and the pattern of ornamentation. Teeth can be gracile with long roots, robust with short roots, or small and needle-like (Brown, 1981). Flat faces are present in some pliosaurs from the Upper Jurassic, in which teeth cross-section is subtriangular (Tarlo, 1960). Observed striation patterns are basically three: no striation, only lingual striation, or striae around the entire pe-riphery (Tarlo, 1960). Teeth from Estancia La Blanca are gracile, with circular cross-sections and lingual striation (labial striation reduced or absent). No species with the latter characters (gracile, cylindrical, labial striation) were present in the data matrix used by O' Keefe (2001) for phylogenetic analysis of the clade Plesiosauria. Phylogenetic analyses by Smith (2003) show that only Terminonatator ponteixensis (Sato, 2003) has the same three states observed in the material from Santa Cruz. However, it is necessary to emphasize that there are many missing data in the matrix used, especially those related to striae arrangement. General shape is similar to that observed in Plesiosauroidea, but it has been observed in Polycotylidae too. Thus, because Polycotylidae has a changing position in plesiosaur phylogeny (O'Keefe, 2001; Druckenmiller, 2006) and because of the absence of characters defining or justifying familial or generic assignation, the dental material from Estancia La Blanca (MPM-PV 1871-1-1; MPM-PV 1871-1-2; MPM-PV 1871-1-3) must be referred to Plesiosauria indet.
MPM-PV 1870-2, one incomplete cervical vertebral body (figure 4.3). The proportions of the body are 40, 113:150; therefore, it is higher than long and wider than high. Ventral surface is slightly concave and has two vascular foramina separated by a rounded ridge. There are traces of single-headed fused ribs in the ventrolateral area of the body.
Vertebral body is longer than those observed in polycotylids and pliosaurids, and lacks the prominent ring around the articular face observed in polycotylids (Tarlo, 1960; Sato and Storrs, 2000). On the other hand, proportions are not those of typical elasmosaurid cervical bodies, which tend to be longer than high. The latter character has been used as a synapomorphy of the family Elasmosauridae (Persson, 1963; Bardet et al., 1999). However, when plotting the values of neck length, height, and width in two Elasmosauridae, it is obvious that the last cervical vertebrae are shorter than those located in the anterior region and midlength of neck. Graphs (figure 6) show measurements of the cervical centra in Elasmosaurus platyurus Cope, 1869, a typical elasmosaurid with strong elongation of the cervical vertebrae (figure 6.1), and Mauisaurus haasti Hector, 1874, a more generalized elasmosaur in relation to the vertebral elongation (figure 6.2). In both cases, the posterior cervical centra are higher than long. Moreover, the articular surface, which is very damaged, seems to be nearly flat -an elasmosaur feature (Bardet et al., 1999). It is possible that vertebra MPM-PV 1870-2 may belong to an elasmosaur, but poor preservation prevents definite confirmation. Thus, we prefer to refer this vertebra to Plesiosauria indet.
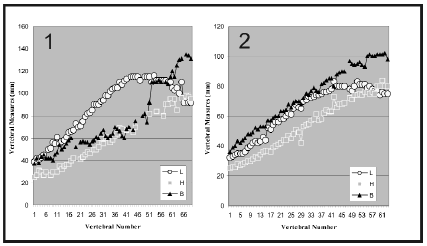
Figure 6. 1-2. Graphic length (L, ), height (H, ), and breadth (B, ) of the cervical vertebrae of two Upper Cretaceous elasmosaurs / gráfico de los valores del largo (L, ), alto (H, ) y ancho (B, ) de las vértebras cervicales de dos elasmosaurios del Cretácico Superior. 1, Elasmosaurus platyurus Cope, 1869. A.N.S.P. nº 10081 (data taken from Welles, 1952, p. 55); and 2, Mauisaurus haasti Hector, 1874. CM Zfr 115 (Data taken from O'Keefe, 2006, p. 228) / 1, Elasmosaurus platyurus Cope, 1869. A.N.S.P. nº 10081 (Datos tomados de Welles, 1952, p. 55) y 2, Mauisaurus haasti Hector, 1874 . CM Zfr 115 (Datos tomados de O'Keefe, 2006, p. 228).
MPM-PV 1870-3 (figure 4.7-9), one pectoral vertebral body without ribs or fused neural arches. Proportions of the vertebral body are 76, 118:161. The base of the neural canal becomes wider toward the rear end. On both sides, areas of articulation of the neural arch are depressed and confluent with the parapophysis so there is no clear boundary between the parapophysis and the areas of articulation of the neural arch (figure 4.8). This parapophysis occupies much of the sides of the vertebral body, being slightly depressed at the central zone and convex at the periphery. The two zones -the parapophysis and the zone of articulation of the neural arch- are crossed by foramina. The anterior margin of the parapophysis coincides with the anterior edge of the lateral surface of the vertebral body. The posterior margin of the parapophysis does not contact the posterior border of the lateral surface of the vertebral body (figure 4.8). The anterior articular face is subelliptic and has three dorsal concavities: one corresponding to the neural canal, and two corresponding to lateral areas produced by the articular zones of the neural arch (figure 4.9). The posterior articular face of the vertebral body is subelliptic with a slight dorsal concavity corresponding to the neural canal. Both articular faces are slightly depressed. There are at least two ventral foramina.
MPM-PV 1869-1 (figure 4.4-6), one dorsal vertebral body with a nonfused neural arch. Proportions of the centrum are 32, 122:153, rendering it wider than high and higher than long. Two foramina present at the base of the neural canal. The articulation surface of the neural arch is a unique depression that extends up to the dorsal part of the lateral surface of the vertebral body. There is a small "lump" on the lateral surface which is slightly elongated in an anteroventral direction (figure 4.5). Articular faces of the centrum are elliptical and slightly depressed, with a small prominence in the central zone. Ventral surface of the body rounded, with one small central and two lateral foramina.
MPM-PV 1869-2, fragment of dorsal(?) vertebra, corresponding to the articular face. It measures 38 mm high and 42 mm wide. Therefore, the articular face is subcircular and presents a marked central depression.
The dorsal vertebral bodies (MPM-PV 1869-1 and MPM-PV 1869-2) are amphiplatyan (MPM.-PV 1869-1) to slightly amphicoelic (MPM-PV 1869-2), showing the typical morphology of Plesiosauria. A neural arch not fused to the vertebral body in MPM-PV 1869-1 can be interpreted as indicator of a juvenile or sub-adult ontogenetic stage (Brown, 1981).
MPM-PV 1870-1(figure 4.10-12), one propodium of a juvenile specimen. As the humerus and femur are often similar in plesiosaurs, the assignment of the propodium is difficult. This difficulty becomes more problematic in juveniles, where these two bones are more closely similar than in adults (Brown, 1981). The propodium is robust with proximal and distal expansions. Measurements: 158 mm long, 91 mm distal expansion; B:L index: 57. For orientation of the propodium it was assumed that the most important muscular scar is the ventral scar, which is displaced toward the posterior margin (figure 4.12). On the dorsal surface of the propodium there is another muscular scar which is smaller and located in a more central position (figure 4.10), as well as a third elon-gated muscular scar at the distal half of the posterior margin.
The capitulum is elliptical in proximal view, with the major axis anteroposteriorly oriented, and surrounded by a slight ridge. The entire surface of the capitulum is pierced by foramina which indicate the presence of transphyseal canals, and therefore a vascularized cartilaginous cap. Although there is no well developed trochanter or tuberosity, the dorsal surface has a slightly marked projection, which could be an incipient trochanter/tuberosity. The distal end is rounded and devoid of articular facets. The material was found broken across the middle of the shaft, exposing a dense and compact cortex, a well defined medulla, and canals that connect the medulla with the outer surface (figure 4.11).
A juvenile ontogenetic stage is inferred on the basis of the following features: the robustness, the lack of differentiation between capitulum and tuberosity or trochanter, and the absence of articular facets at the distal end. Furthermore, the internal structure of the bones, particularly of the propodium, has been suggested as a possible indicator of ontogenetic stage. Compact and well-defined cortex surrounding the medulla has been recorded in juvenile individuals. The cortex has a cancellous structure in adults, so the boundary between medulla and cortex is diffuse (Wiffen et al., 1995; Wahl, 2006). The material described herein displays the typical structure observed in juveniles. This is congruent with the other morphological features mentioned above.
Foramina in the capitulum indicate the presence of transphyseal canals which suggest a vascularized cap. These canals have been found in fast growing species that reach large sizes (Rhodin, 1985). They have been observed in the turtle Dermochelys coriacea and interpreted as related to its large size. Vascularization of the epiphyses has not been observed in smaller turtles (Rhodin, 1985).
Propodium morphology (MPM-PV 1870-1) closely resembles that of MML Pv 5, an indeterminate Plesiosauria found in the Jagüel Formation (late Maastrichtian) in Salinas de Trapalcó, Río Negro Province (Gasparini et al., 2003a, 2007). MML Pv 5 and MPM-PV 1870-1 from Estancia La Blanca share the robustness of the humerus and femur, the lack of differentiation between capitulum and tuberosity or trochanter, and the absence of articular facets at the distal end. All these characters have been associated with juvenile stages (Brown, 1981; Wahl, 2006); therefore, their taxonomic value is doubtful. However, a propodium with similar features found in Río Negro (MML-Pv 193) indicates that larger individuals (and therefore, probably older) retain part of the mentioned features (Gasparini et al., 2007). Propodia of similar characteristics have been collected in the Maastrichtian of Central Chile ever since the 19th century (Gay, 1848, pl. 2) and assigned to Plesiosaurus chilensis Gay (1848), afterwards named Pliosaurus chilensis (Gay, 1848) by Deecke, 1986. However, the latter has been considered a nomen vanum by subsequent authors and probably requires revision (Welles, 1962).
Comments on Polyptychodon patagonicus Ameghino, 1893
Polyptychodon patagonicus was described by Ameghino in 1893. The type material of Polyptychodon patagonicus consists of a group of teeth -described but never illustrated- coming from what Ameghino called and understood as "formación Santacruceña" in 1893. Ever since the 19th century there have been different opinions about the age and correct assignment of this material. The discovery of plesiosaur vertebrae and teeth in the lower section of the Mata Amarilla Formation leads to the analysis of the history of Florentino Ameghino's ideas on the age of the layers bearing the remains, and how this influenced the assignation of the materials to other reptile taxa.
Ameghino made the following description of the original materials of Polyptychodon patagonicus: [The teeth of Polyptychodon patagonicus are open at theirbase, conicalcylindrical in shape, with sharp-pointed apex, and strongly curved. The outer surface is covered by pronounced longitudinal ridges of enamel, separated by deep furrows; the ridges start at the base, all of them at the same level, ending at different distances from the apex, nearly in the same way as in Polyptychodon interruptus, Ow. I do not know any complete tooth. The largest tooth, which almost lacks the entire base occupied by the pulp cavity, is nearly 3 cm high; and at the broken part at the base, it measures one centimeter in diameter approximately] (Ameghino, 1893, p. 82; translated from Spanish).
However, subsequent authors considered Ameghino's identification (1893) with doubts, probably because of two reasons. First, plesiosaur teeth have been often confused with crocodile or fish teeth, because of their similar morphological characteristics. Second, the association of Polyptychodon patagonicus teeth with Cenozoic mammalian teeth in the same for-mation was assumed. For this reason Cabrera (1941) stated that if the teeth of Polyptychodon patagonicus were contemporaneous with mammalian remains, then they should be assigned to crocodilians. Welles (1962) followed the same line of thought and mentioned Cabrera (1941), claiming that the teeth could belong to crocodilians. Finally, in a summary of South American plesiosaurs, Gasparini and Goñi (1985) mentioned Polyptychodon patagonicus as a plesiosaur, but with serious doubts about its assignation.
Unfortunately, the original material described by Ameghino in 1893 is not available for study. Although there is a record of two teeth of Polyptychodon patagonicus from the "Sub-Patagoniano" of Lago Argentino in the MACN, this material (catalogued as MACN-A 5809) could not be located in the collection.
Concerning the stratigraphic and geographic provenance of his material, Ameghino (1893, p. 76) wrote: [The formation that I have named Santacruceña, covers most of the Austral Patagonian region that is crossed by the Santa Cruz, Sehuen, and Gallegos Rivers] (Ameghino, 1893, p. 76; translated from Spanish). It is clearly evident that the "formación Santacruceña" named by Ameghino, is equivalent only in part to what is nowadays known as the Santa Cruz Formation, as the Santa Cruz Formation is not exposed along the Shehuen or Chalia River (Sehuen River of Ameghino). Contrarily, the Mata Amarilla Formation is the unit exposed along the mentioned river (Feruglio, in Fossa Mancini et al., 1938; Arbe, 1989, 2002; Varela and Poiré, 2008). For this reason it is evident why Ameghino included the Mata Amarilla Formation in his "formación Santacruceña" (Cione et al., 2007). Ameghino's opinion is quite understandable taking into account the lithological similarity of the Mata Amarilla and the Santa Cruz formations.
In relation to the age, Ameghino (1893, p. 76) assigned his "formación Santacruceña" to the "Eoceno inferior (Paleoceno)". Due to the problematic associa-tion of primates and other Cenozoic mammals with Cretaceous taxa in the "formación Santacruceña", Ameghino wrote in the final discussion: [This fact (referring to the identifications carried out in the paper and the associated Cretaceous age) is meaningful and enough to decidedly tilt the scales in favor of those who believe that the "formación Santacruceña" is even older than the "lower Eocene", as I considered it from the beginning. It is possible that it becomes necessary to refer the lower part of it to the larámico or upper Cretaceous] (Ameghino, 1893, p. 84; translated from Spanish).
So what seemed to be an association between Polyptychodon patagonicus and mammalian teeth was just a lithostratigraphic misunderstanding of Ameghino, who rectified himself in 1906. It is important to emphasize that the original purpose of the 1893 paper was not to solve the stratigraphical problem, but to add new descriptions of the material sent by his brother, Carlos Ameghino. However, since 1893 Ameghino supposed that his "formación Santacruceña" included units of different age.
In his work published in 1906, Ameghino separated the outcrops at Río Shehuen initially belonging to the "formación Santacruceña", and named them "Sehuenense stage". Ever since then Polyptychodon patagonicus was listed together with taxa from the "Sehuenense stage" instead of being included with those from the "formación Santacruceña", thus solving the stratigraphic problem. Hence, it can be assumed that there is no real association between Polyptychodon patagonicus and Cenozoic taxa.
It is necessary to point out that different authors refer to the "Sehuenense" in different ways. Ameghino (1906) used "Sehuenense", while Cabrera (1941) used "Sehuense", and Arbe (1989, 2002) "Shehuenense". We use the first spelling in this contribution because we refer to Ameghino's original concept (1906).
Under the light of the discussion above, and with the new record showing the existence of abundant dental material from the Mata Amarilla Formation assignable to Plesiosauria, there appears to be strong evidence that Ameghino was the first to publish material referred to Plesiosauria from lower Late Cretaceous rocks in Santa Cruz.
Since the original material is not available, the relation between the teeth referred to Polyptychodon patagonicus and the new records described here can only be discussed based on Ameghino's description and the diagnosis of the genus Polyptychodon Owen, 1841
Tooth morphology described by Ameghino (1893) is common in plesiosaurs: conical, slightly curved teeth, with marked striae. Polyptychodon teeth illustrated by Owen in 1841 seem to bear striae on the entire outer surface
The fact that Ameghino never mentioned any difference between his material and the teeth illustrated by Owen could indicate that his material had the same features shown by Polyptychodon. Therefore, the presence of striae on the entire outer surface is a difference between the original material of Polyptychodon patagonicus and the materials described in this paper, which have reduced or even absent lingual striation.
The reasons stated by other authors (Cabrera, 1941; Welles, 1962) to reject the identification of the teeth made by Ameghino are thus not valid. The assignment of Ameghinos's material to Polyptychodon, a pliosaur from the Upper Cretaceous (Cenomanian-Campanian?) in England, Germany, Czech Republic, and USA (Welles and Slaughter, 1963, Bardet and Godefroint, 1995) is not valid because no characters support this assignation. Hence, Polyptychodon patagonicus is a nomen vanum and the type material, as well as the teeth described in this paper, must be referred to Plesiosauria indet.
Paleoenvironment interpretation
The lower section of the Mata Amarilla is well exposed in LB section, where the plesiosaur remains were found in white sandstone facies with herring-bone cross-stratification, showing changes in the direction of palaeocurrents; according to Boyd et al. (2006) this is interpreted as produced by tidal action. So, following the criteria of Boyd et al. (2006), these facies were interpreted as subtidal bars characterizing the shallowest part of estuaries. Finally, this lower section ends with a prograding bayhead delta.
In the MAT section, the lower section of the Mata Amarilla Formation exposes only five meters (figure 3). The skeletal remains of plesiosaurs in the MAT section were found in heterolithic, mixed stratified (flaser and wavy) facies, showing alternating decantation and traction processes. Such structures are mostly dominated by shales, characteristic of estuarine central areas, according to Boyd et al. (2006). The paleoenvironment of the lower section of the Mata Amarilla Formation where the plesiosaurs were found is therefore interpreted as a tidal dominated estuary.
All the plesiosaur bones and teeth show signs of transport. The material is disarticulated with no evidence that they belonged to the same single specimen. It is interesting to note that the remains are predominantly elements resistant to fragmentation, such as the vertebral centra, the propodium, and teeth. However, the teeth are incomplete, most with-out roots and some lacking the apex of the crown. Bone damage consists of fractures and elimination of superficial bone but no perforations or fouling were observed. Therefore, the preservation status of the skeletal remains is consistent with the energetic conditions of tidal action, which is consistent with the sedimentological interpretation.
In vertebra MPM-PV 1871-2 (Level 2, LB section), the internal cavity is filled with sediment forming a natural cast of the missing part of the vertebra. Clearly, the fracture occurred after lithification, suggesting that part of the damage observed in the material could have been caused by reworking or recent weathering.
Finally, although very scarce, some remains of the lower Mata Amarilla Formation are young specimens, possibly lying near the coast, probably an estuary. It has been often mentioned that young specimens, at least those of plesiosauroids, could live in rivers and estuaries (Wiffen et al., 1995).
Biogeographic significance of the new record
Late Cretaceous plesiosur records in southern Gondwana were limited to the Campanian-Maastrichthian of Argentina (Cabrera, 1941; Gasparini and Goñi, 1985, Gasparini and Salgado, 2000; Gasparini et al., 2003a,b; 2007), Chile (Gay, 1849; Casamiquela, 1969; Suárez and Fritis, 2002), Antarctic Peninsula (Gasparini et al., 1984; Chatterjee and Small, 1989; Fostowicz-Frelik and Gazdzicki, 2001; Martin et al., 2007), New Zealand (Wiffen and Moisley, 1986; Cruickshank and Fordyce, 2002; Hiller et al., 2005), and the Turonian-Cenomanian of Australia (Kear, 2003). According to those records it is evident that while the early Late Cretaceous is recorded in Oceania, there are no records for the same period in southern South America and Antarctica. The new plesiosaurian material from the lower section of the Mata Amarilla Formation contributes toward completing this gap in the record (figure 9).

Figure 7. Late Creatceous paleomap showing some of the records of plesiosaurs in Southern Gondwana (modified from Gasparini et al., 2003a). Lower Early Creataceous record; upper upper Late Cretaceous record / paleomapa del Cretácico Superior donde se muestran algunos de los registros del Cretácico Superior en el Sur de Gondwana. registros correspondientes al Cretácico Superior bajo; registro correspondiente al Cretácico Superior alto.
Conclusion
The oldest lower Late Cretaceous plesiosaur record in Argentina is presented, becoming the first record of these reptiles for Santa Cruz Province. Analysis of the literature provides strong evidence that Polyptychodon patagonicus Ameghino, 1893, is based on remains of Plesiosauria from the Mata Amarilla Formation. However, the status of nomen vanum is maintained for P. patagonicus because the isolated teeth are not considered sufficient to diagnose a genus or species. The lower section of the Mata Amarilla Formation in the study area was deposited in an estuarine environment with strong tidal influence. The conservation and lack of articulation of the remains is congruent with the expected level of transport in that environment.
Acknowledgments
The authors thanks T. Martin (Institut für Palaeontologie, Universität Bonn), F. Goin and J. Gelfo (Museo de La Plata) for providing logistical support in field work, in which also participated J. Cuitiño, G. Pedersen, A. Barrueco, M. Giacobelli, L. Chornogubsky, S. Hoffmann, and J. Schultz. They also thank Dr. Adan Tauber (Museo Regional Provincial Padre Manuel Jesús Molina, Río Gallegos) for authorization to study the material from Santa Cruz, and L. Acosta Burllaile for the preparation of material. C. Deschamps and J. Echevarria (Museo de La Plata) for reading the English version. Likewise they tanks their dissertation advisors, Z. Brandoni de Gasparini, L. Salgado, and D. Poiré for critically reading of the manuscript and for financial support (projects PICT 25276 and CONICET PIP 6237/05). Finally they thank the referees, N. Bardet (Muséum national d' Histoire naturelle, Paris) and J. Martin (Museum of Geology, South Dakota School of Mines and Technology, USA), whose comments have substantially improved both the scientific content and the language of this paper.
References
1. Ameghino, F. 1893. Sobre la presencia de vertebrados de aspecto mesozoico en la formación Santacruceña de la Patagonia austral. Revista del Jardín Zoológico de Buenos Aires 1:76-84 [ Links ]
2. Ameghino, F. 1906. Les formations sédimentaires du Crétacé Superieur et du Tertiaire de Patagonia, avec an paralèlle entre leurs faunes mammalogiques et celles de l' ancien continent. Anales del Museo Nacional de Buenos Aires, 15 (serie 3, 8): 1-508. [ Links ]
3. Arbe, H.A. 1989. Estratigrafía, discontinuidades y evolución sedimentaria del Cretácico en la Cuenca Austral, Prov. de Santa Cruz. Instituto Superior de Correlación, Universidad Nacional de Tucumán, Serie de Correlación Geológica 6: 419-442. [ Links ]
4. Arbe, H.A. 2002. Análisis estratigráfico del Cretácico de la Cuenca Austral. In: M.J. Haller (ed.), Geología y Recursos Naturales de Santa Cruz. 15º Congreso Geológico Argentino, Relatorio: 103-128. [ Links ]
5. Arrondo, O.G. 1983. [Informe estudio paleontológico. Yacimientos Petrolíferos Fiscales, Buenos Aires. Unpublished report. [ Links ]].
6. Bardet, N. and Godefroint, P. 1995. Plesiosaurus houzeaui Dollo, 1909 from the Upper Campanian of Ciply (Belgium) and a review of the Upper Cretaceous plesiosaur from Europe. Sciences de la Terre 65: 179-186. [ Links ]
7. Bardet, N., Godefroit, P. and Sciau, J. 1999. A new elasmosaurid plesiosaur from the Lower Jurasic of southern France. Palaeontology 42: 927-952. [ Links ]
8. Bianchi, J.L. 1967. [Informe preliminar acerca de los perfiles estratigráficos realizados en el sector occidental de la Cuenca Austral, durante las campañas 1964-65 y 1965-66. Yacimientos Petrolíferos Fiscales, Buenos Aires. Unpublished report. [ Links ]].
9. Biddle, K., Uliana, M., Mitchum, R .Jr., Fitzgerald, M., and Wright, R. 1986. The stratigraphic and structural evolution of central and eastern Magallanes Basin, Southern America. In: P. Allen, and P. Homewood (eds.), Foreland basins. International Association of Sedimentologists, Special Publication 8: 41-61. [ Links ]
10. Blainville, H.D. de. 1835. Description de quelques espèces de reptiles de la Californie, precédée de l'analyse d'un système général d'herpetologie et d'amphibiologie. Nouvelles Archives du Museum d'Histoire Naturelle 4: 233-296. [ Links ]
11. Blasco, G.B., Nullo, F. and Proserpio, C. 1980. Santoniano-Campaniano: Estratigrafía y contenido amonitífero, Cuenca Austral. Revista Asociación Geológica Argentina 35: 467-493. [ Links ]
12. Bonarelli, G. and Nagera, J.J. 1921. Observaciones geológicas en las inmediaciones del lago San Martín (territorio de Santa Cruz). Dirección General de Minas, Boletín 27b: 1-39. [ Links ]
13. Boyd, R., Dalrymple, R.W. and Zaitlin, B.A. 2006. Estuarine and Incised-Valley Facies Model. In: H.W. Posamentier and R.G. Walker (eds.), Facies Model Revisited. Society for Sedimentary Geology, Special Publication 84: 171-235. [ Links ]
14. Brown D. S.1981. The English Upper Jurassic Plesiosauroidea (Reptilia) and a review of the phylogeny and classification of the Plesiosauroidea. Bulletin of British Museum of Natural History, (Geology) 35: 253-347. [ Links ]
15. Cabrera, A. 1941. Un Plesiosaurio nuevo de Cretáceo del Chubut. Revista del Museo de la Plata (Nueva Serie) 2: 113-130. [ Links ]
16. Casamiquela, R.M. 1969. La presencia de Aristonectes Cabrera (Plesiosauria) del Maastrichtense del Chubut, Argentina. Edad y carácter de la transgresión Rocanense. 4º Jornadas Geología Argentinas (Mendoza), Actas 1:199-213. [ Links ]
17. Chatterjee, S. and Small, B.J. 1989. New plesiosaur from the Upper Cretaceous of Antarctica. In: J.A. Crame (ed.), Origins and Evolution of the Antarctic Biota, Special Publications of the Geological Society 47: 197-215. [ Links ]
18. Cione, A.L., Gourie, S., Goin, F., and Poiré, D. 2007. Atlantoceratodus, a new genus of lungfish from Upper Cretaceous of South America and Africa. Revista del Museo de La Plata, Paleontología 10: 1-12. [ Links ]
19. Cruickshank, A.R.I. and Fordyce, R.E .2002. A new marine reptile (Sauropterygia) from New Zealand: Further evidence for a Late Cretaceous Austral radiations of cryptoclidid plesiosaur. Palaeontology 45: 557-575. [ Links ]
20. Cope, E.D. 1869. Synopsis of the extinct Batrachia, Reptilia and Aves of North America. Transaction of the American Philosophical Society (new series) 14: 1-252. [ Links ]
21. Deecke, H.V. 1896. Über Saurier-Reste aus dem Quiriquina-Schichten; Neues Jahrbuch für Mineralogie, Geologie und Paläontologie 10: 32-63 [ Links ]
22. Druckenmiller, P.S. 2006. [Early Cretaceous plesiosaurs (Sauropterygia: Plesiosauria) from the northern Alberta palaeoenvironmen-tal and systematic implications. Unpublished Thesis. Department of Biological Sciences, Alberta, University of Calgary. 581 pp. [ Links ]].
23. Feruglio, E. 1936. Paleontographia Patagonica. Istituto Geologico della Università di Padova, Memorie 11: 1-384. [ Links ]
24. Feruglio, E. 1938. El Cretáceo superior del lago San Martín y de las regiones adyacentes. Physis 12: 293-342. [ Links ]
25. Fossa Mancini, E., Feruglio, E. and Yussen de Campana, J.C. 1938. Una Reunión de geólogos de Y.P.F. y el problema de la Terminología Estratigráfica. Boletín de Informaciones Petroleras 171: 31-95. [ Links ]
26. Fostowicz-Frelik, L. and Gazdziki A. 2001. Anatomy and histol-ogy of plesiosaur bones from the Late Cretaceous of Seymour Island, Antarctic Peninsula. Palaeontologia Polonica 60: 7-32. [ Links ]
27. Gasparini, Z., Bardet, N., Martin, J.E. and Fernandez, M. 2003b. The elasmosaurid plesiosaur Aristonectes Cabrera from the Latest Cretaceous of South America and Antarctica. Journal of Vertebrate Paleontology 23: 104-115. [ Links ]
28. Gasparini, Z. and de la Fuente, M. 2000. Tortugas y plesiosaurios de la Formación La Colonia (Cretácico Superior) de Patagonia, Argentina. Revista Española de Paleontología 15: 23-35. [ Links ]
29. Gasparini, Z., del Valle, R. and Goñi, R. 1984. Un Elasmosaurido (Reptilia, Plesiosauria) del Cretácico Superior de la Antártida. Contribuciones del Instituto Antártico Argentino 305: 1-24. [ Links ]
30. Gasparini, Z. and Goñi, R. 1985. Los plesiosaurios el Cretácico de America del Sur y del continente antártico. Coletânea de Trabalhos Paleontológicos 27: 55-63. [ Links ]
31. Gasparini, Z., and Salgado, L. 2000. Elasmosauridos (Plesiosauria) del Cretácico Tardío del norte de Patagonia. Revista Españo de Paleontología 15: 13-21. [ Links ]
32. Gasparini, Z., Salgado, L., and Casadío, S., 2003a. Maastrichtia plesiosaurs from northern Patagonia. Cretaceous Research 2 147-170. [ Links ]
33. Gasparini Z., Salgado, L. and Parras, A. 2007. Late Cretaceous plesiosaurs from northern Patagonia, Argentina. Geologic Journal 42: 185-202. [ Links ]
34. Gasparini, Z, and Spalletti, L.A. 1990. Un nuevo cocodrilo en lo depósitos mareales maastrichtianos de la Patagonia noroccidental. Ameghiniana 27: 141-150. [ Links ]
35. Gay, C. 1848. Historia Física y Política de Chile. Zoología 3-4. Maulde et Renou, París, pp. 371. [ Links ]
36. Goin, F.J., Poiré, D.G., de la Fuente, M.S., Cione, A.L., Nova F.E., Bellosi, E.S., Ambrosio, A., Ferrer, O., Canessa, N.D Carloni, A., Ferigolo, J., Ribeiro, A.M., Sales Viana, M.S Pascual, R., Reguero, M., Vucetich M.G., Marenssi, S., D Lima Filho, M. and Agostinho, S. 2002. Paleontología Geología de los sedimentos del Cretácico Superior aflorant al sur del Río Shehuen (Mata Amarilla, Prov. de Santa Cru Argentina). 15º Congreso Geológico Argentino (El Calafate) Actas: 603-608. [ Links ]
37. Hector, J. 1874. On the Fossil Reptilia of New Zealand. Transactions and Proceedings of the New Zealand Institute 6: 333-358. [ Links ]
38. Hiller, N., Mannering, A.A, Jones, C.M, and Cruickshank, A.R. 2005. The nature of Mauisaurus haasti Hector, 1874 (Reptilia:Plesiosauria). Journal of Vertebrate Paleontology 25: 588-601 [ Links ]
39. Iglesias, A., Zamuner, A.B., Poiré, D.G, and Larriestra, F. 200 Diversity, taphonomy and palaeoecology of an angiosper flora from the Cretaceous (Cenomanian-Coniacian) in Southern Patagonia, Argentina. Palaeontology 50: 445-466. [ Links ]
40. Ihering, H., von. 1897. Os molluscos dos terrenos terciarios de Patagonia. Revista do Museu Paulista 2 : 217-227. [ Links ]
41. Ihering, H., von. 1899. Die Conchylien der patagonischen Forma tion. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie 1899, 1-46. [ Links ]
42. Ihering, H., von. 1907. Les mollusques fossiles du Tertiare et du Crétacé superieur de l' a Argentine. Anales Museo Nacional Buenos Aires 7: 1-611. [ Links ]
43. Kear, B.P. 2003. Cretaceous marine reptiles of Australia: a revie of taxonomy and distribution. Cretaceous Research 24: 277-30 [ Links ]
44. Lacovara, K., Harris, J., Lammana, M., Novas, F., Martinez, R. and Ambrosio, A. 2004. An enormous sauropod from the Maastritchtian Pari Aike Formation of southernmost Patagonia. Journal of Vetebrate Paleontology 24: 81A. [ Links ]
45. Lazo, D.G. and Cichowolski, M. 2003. First plesiosaur remain from the Lower Cretaceous of the Neuquén Basin, Argentina Journal of Paleontology 77: 784-789. [ Links ]
46. Leanza, A.F. 1972. Andes Patagónicos Australes. In: A.F. Leanza (ed.), Geología Regional Argentina. Academia Nacional de Ciencias (Córdoba), pp. 689-706. [ Links ]
47. Martin, J.E., Sawyer, J.F, Reguero, M. and Case, J.A. 2000 Occurence of a young elasmosaurid plesiosaur skeleton from the Late Cretaceous (Maastrichtian) of Antartica. Online Proceedings of the 10th ISAES. USGS Open Report 2007-104, Short Research Paper 066, pp.4 [ Links ]
48. Novas, F.E., Cambiaso, A.V. and Ambrosio, A. 2004a. A new basal iguanodontian (Dinosauria, Ornithischia) from the Upper Cretaceous of Patagonia. Ameghiniana 41: 75-82. [ Links ]
49. Novas, F.E., Ezcurra, M.A. and Leucona, A., 2008. Orkorapt burkei nov. gen. et sp., a large theropod from the Maastrichtia Pari Aike Formation, Southern Patagonia, Argentina Cretaceous Research 29: 468-480. [ Links ]
50. Novas, F.E., Lecuona, A., Calvo, J. and Porfiri, J. 2004b. Un terópodo del Cretácico Superior de la provincia de Santa Cruz Ameghiniana Suplemento Resúmenes 41: 58R. [ Links ]
51. Novas, F.E., Salgado, L., Calvo, J.O. and Agnolín, F.L. 2005. Giant titanosaur (Dinosauria, Sauropoda) from the Late Cretaceou of Patagonia. Revista del Museo Argentino de Ciencias Naturales (nueva serie) 7: 37-41. [ Links ]
52. Owen, R. 1841. Odontography 1. London. Hippolyte Bailliere Press, London 655 pp. [ Links ]
53. Owen, R. 1860. On the orders of fossil and recent Reptilia, and their distribution in time. Reports of the British Association for the Advancement of Science 29: 153-166. [ Links ]
54. O'Keefe, F.R. 2001. A cladistic analysis and taxonomic revision of the Plesiosauria (Reptilia: Sauropterygia). Acta Zoologica Fennica 213: 1-63. [ Links ]
55. O'Keefe, F.R. 2006. Morphologic and ontogenetic patterns in elas-mosaur neck length, with comments on the taxonomic utility of neck length variables. Paludicola 5: 206-229. [ Links ]
56. Persson, P.O. 1963. A Revision of the classification of the Plesiosauria with synopsis of the stratigraphical and geo-graphical distribution of the group. Lunds Universitets Arsskrift 59: 1-60. [ Links ]
57. Poiré, D.G., Franzese, J.R., Spalletti, L.A. and Matheos, S.D. 2007. [Estratigrafía de las rocas reservorios de la Cuenca Austral en el sector cordillerano, provincia de Santa Cruz, Argentina. Guía de Campo, Centro de Investigaciones Geológicas, La Plata, 112 pp. Unpublished. [ Links ]].
58. Poiré, D.G., Zamuner, A.B., Goin, F., Iglesias, A., Canessa, N., La-rriestra, C.N., Varela, A.N., Calvo Marcillese, L. and Larriestra, F. 2004. Ambientes sedimentarios relacionados a las tafofloras de las formaciones Piedra Clavada y Mata Amarilla (Cretácico), Tres Lagos, Cuenca Austral, Argentina. 10º Reunión Argentina de Sedimentología (San Luis) Actas: 140-141. [ Links ]
59. Previtera, E. and Gonzalez Riga, B.J. (2008). Vertebrados cretácicos de la Formación Loncoche en CalmuCo, Mendoza, Argentina. Ameghiniana 45: 349-359. [ Links ]
60. Rhodin, A.G.J. 1985. Comparative chondro-osseous development and growth of marine turtles. Copeia 3: 752-771. [ Links ]
61. Russo, A. and Flores, M.A. 1972. Patagonia austral extraandina. In: A.F. Leanza (ed.), Geología Regional Argentina. Academia Nacional de Ciencias (Córdoba), pp. 707-725. [ Links ]
62. Russo, A., Flores, M.A. and Di Benedetto, H. 1980. Patagonia austral extraandina. In: J.C.M. Turner (ed.), Segundo Simposio de Geología Regional, Academia Nacional de Ciencias (Córdoba), pp. 1431-1462. [ Links ]
63. Salgado, L., Parras, A. and Gasparini, Z. 2007. Un plesiosaurio de cuello corto (Plesiosauroidea, Polycotylidae) del Cretácico Superior del norte de Patagonia. Ameghiniana 44: 349-358. [ Links ]
64. Sato, T. 2003. Terminonator ponteixensis, a new elasmosaur (Reptilia; Sauropterygia) from the Upper Cretaceous of Saskatchewan. Journal of Vertebrate Paleontology 23: 89-103. [ Links ]
65. Sato, T. and Storrs, G.W. 2000. An Early Polycotylid plesiosaur (Reptilia: Sauropterygia) from the Cretaceous of Hokkaido, Japan. Journal of Paleontology 74: 907-914. [ Links ]
66. Smith, A.S. 2003. [Cladistic Analysis of the Plesiosauria (Reptilia: Sauropterygia).Unpublished Thesis. Department of Earth Sciences, University of Bristol. pp.81. [ Links ]].
67. Suarez, M.E. and Fritis, O. 2002. Nuevo registro de Aristonectes ( Plesiosauroidea, incertae sedis) del Cretácico Tardío de la Formacion Quiriquina, Cocholgüe, Chile. Boletín de la Sociedad de Biología de Concepción 73: 87-93. [ Links ]
68. Tarlo, L.B.H. 1960. A Review of the Upper Jurassic Pliosaurs. Bulletin of the British Museum (Natural History) 4: 145-189. [ Links ]
69. Varela, A.N. 2009. Accommodation / sediment supply fluvial deposition controlled by base level changes and relative sea level fluctuations in the Mata Amarilla Formation (Early Upper Cretaceous), Southern Patagonia, Argentina. Abstracts 9th International Conference on Fluvial Sedimentology. Acta Geologica Lilloana 21:66. [ Links ]
70. Varela, A.N. and Poiré, D.G. 2008. Paleogeografía de la Formación Mata Amarilla, Cuenca Austral, Patagonia, Argentina. 12ª Reunión Argentina de Sedimentología, Actas: 183. [ Links ]
71. Varela, A.N., Richiano, S. and Poiré, D.G. 2008. Análisis paleoambiental de la Formación Mata Amarilla a partir de su malaco-fauna, Cuenca Austral, Patagonia, Argentina. In: Schiuma M. (ed.), Trabajos Técnicos, 7ª Congreso de Exploración y Desarrollo de Hidrocarburos (Mar del Plata), pp. 601-605. [ Links ]
72. Wahl, W.R. 2006. Juvenile plesiosaur (Reptilia: Sauropterygia) assemblage from the Sundance Formation (Jurassic), Natrona County, Wyoming. Paludicola 5: 255-261. [ Links ]
73. Welles, S.P. 1952. A Review of the North American Cretaceous Elasmosaurs. University of California Publications in Geological Sciences 29: 144 [ Links ]
74. Welles, S.P. 1962. A new species of Elasmosaur from the Aptian of Colombia and a review of the Cretaceous plesiosaurs. University of California Publications in Geological Sciences 44: 96 [ Links ]
75. Welles, S,P. and Slaughter, B.H. 1963. The first record of the ple-siosaurian genus Polyptychodon (Pliosauridae) from the new world. Journal of Paleontology 37: 131-133. [ Links ]
76. Wiffen, J., Buffrenil, V., Ricqlès, A.D. and Mazin, J. 1995. Ontogenetic evolution of bone structure in Late Cretaceous Plesiosauria from New Zealand. Geobios 7: 625-636. [ Links ]
77. Wiffen, J. and Moisley, W.L. 1986. Late Cretaceous reptiles (families Elasmosauridae and Pliosauridae) from the Manga-houange Stream, North Island, New Zealand. New Zealand Journal of Geology and Geophysics 29: 205-252. [ Links ]
78. Wilckens, O. 1907. Die Lamellibranchiaten, gastropoden etc, der oberen Kreide Südpatagoniens. Berichte der Naturforschungen Gesellschaft Freiburg 15: 1-70. [ Links ]
Recibido: 2 de febrero de 2009.
Aceptado: 28 de abril de 2010.

