










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana vol.47 no.4 Buenos Aires oct./dic. 2010
ARTÍCULOS ORIGINALES
Distribución de Enneadocysta (Dinoflagellata) en el Paleógeno del Atlántico Sudoccidental: implicancias paleoceanográficas
G. Raquel Guerstein1 y Gloria Daners2
1Instituto Geológico del Sur - Consejo Nacional de Investigaciones Científicas y Técnicas - Departamento de Geología, Universidad Nacional del Sur. San Juan 670, 8000 Bahía Blanca. Argentina raquel.guerstein@uns.edu.ar
2Departamento de Evolución de Cuencas, Facultad de Ciencias, Iguá 4225, 11400 Montevideo, Uruguay. glo@fcien.edu.uy
Abstract. Paleogene distribution of Enneadocysta (Dinoflagellata) in the Southwestern Atlantic Ocean: paleoceanographic implications. The study of Middle to Late Eocene dinoflagellate cyst assemblages from subtropical Southwest Atlantic basins shows the presence of species of Enneadocysta, including E. dictyostila (Menéndez) Stover and Williams emend. Fensome et al., E. brevistila Fensome et al. and E. multicornuta (Eaton) Stover and Williams. The species Enneadocysta dictyostila has been widely recorded in middle Paleogene sections from Southern Hemisphere high latitudes. High representativity of Enneadocysta spp., along with other components of the Southern Ocean endemic dinoflagellate flora, have been observed in samples from Colorado Basin (~38°S), Argentina and Punta del Este Basin (~36°S), Uruguay. Previous research in the Tasman area has related the presence of these endemic taxa at mid-lat-itudes to the formation of a strong clockwise subpolar gyre ("cold trap") favoured by the continental blockage of the Tasman Gateway. In this paper we propose that the observed dinoflagellate cyst distribution in the Southwest Atlantic basins can be explained by a similar dynamical mechanism operating close to the present Weddell Gyre. The blockage of the Tasman Gateway and a partially open Drake Passage both contributed to the formation of a strong western-intensified clockwise gyre that may have transported the endemic flora components, mainly represented by Enneadocysta spp., from the Antarctic continent northward along the Southwest Atlantic Shelf during the Middle - Late Eocene.
Resumen. En las asociaciones de quistes de dinoflagelados del Eoceno medio a tardío de cuencas subtropicales del Atlántico Sudoccidental se registra la presencia de especies de Enneadocysta, entre ellas: E. dictyostila (Menéndez) Stover y Williams emend Fensome et al., E. brevistila Fensome et al. y E. multicornuta (Eaton) Stover y Williams. La especie Enneadocysta dictyostila fue registrada en secciones del Paleógeno medio de altas latitudes del Hemisferio Sur. Se hallaron numerosos ejemplares de Enneadocysta spp. junto con otros componentes de la flora de dinoflagelados endémica del Océano Austral en las cuencas del Colorado (~38°S), Argentina y Punta del Este (~36°S), Uruguay. Previamente, en un trabajo realizado en el área de Tasmania, otros autores relacionaron la presencia de componentes de dicha flora en latitudes medias con el desarrollo de giros oceánicos supbolares ("cold traps") favorecidos por la obstrucción continental del conducto de Tasmania. El presente trabajo propone que la distribución de quistes de dinoflagelados en las cuencas del Atlántico Sudoccidental puede ser explicada por una dinámica similar que habría funcionando como un proto-Giro de Wedell. Tanto el conducto de Tasmania cerrado como un Pasaje de Drake parcialmente abierto habrían contribuido a la formación de giros en sentido horario con intensos flujos en los bordes occidentales. Estos flujos serían los responsables de transportar parte de la flora endémica, principalmente representada por Enneadocysta spp., desde las costas del continente Antártico hacia el norte a lo largo de la plataforma sudamericana durante el Eoceno medio-tardío.
Key words. Dinoflagellate cysts; Circum-Antarctic endemic flora; Eocene; Southwest Atlantic Ocean; Oceanic paleocirculation.
Palabras clave. Quistes de dinoflagelados; Flora endémica circum-antártica; Eoceno; Océano Atlántico Sudoccidental; Paleocirculación oceánica.
Introducción
Los quistes de dinoflagelados de pared orgánica muestran un alto potencial tanto en la resolución de problemas estratigráficos como en las reconstrucciones paleoambientales. Esto ha promovido un gran desarrollo de la palinología marina en las últimas décadas, en particular el estudio de las asociaciones del Paleógeno y la utilización de los quistes de dinoflagelados como indicador indirecto (proxy-data) en el análisis detallado de la transición desde el modo climático invernadero al modo glacial (Sluijs et al., 2005; Pross y Brinkhuis, 2005).
En los últimos años se ha incrementado notablemente el estudio de dinoflagelados en varias localidades circum-antárticas (Brinkhuis et al., 2003a, 2003b; Sluijs et al., 2003; Williams et al., 2004). Brinkhuis y colaboradores documentaron las asociaciones de dinoflagelados en cinco testigos profundos del Mar de Tasmania, con el objetivo de definir con mayor precisión la cronología y características de la apertura del Pasaje de Tasmania e instalación de la Corriente Circumpolar Antártica (CCA). Este evento tectónico y su posible relación con el inicio de la glaciación antártica han sido investigados por Huber et al. (2004) y Stickley et al. (2004a, 2004b) para determinar cuáles fueron los principales mecanismos responsables de la mayor transición climática del Paleógeno.
Gran parte de la información sobre dinoflagelados del Eoceno en el Hemisferio Sur proviene del sector oriental de la Antártida (Cranwell et al., 1960; Cranwell, 1964; Hannah, 1997, 2006; Hannah et al., 1998; Levy y Hardwood, 2000; MacPhail y Truswell, 2004), el Mar de Ross (Kemp, 1975), Australia (Wilson, 1967; Stover, 1975; Martin, 1991, 1993), la Plataforma de Tasmania, (Truswell, 1997; Brinkhuis et al., 2003a, b; Sluijs et al., 2003), Nueva Zelanda (Haskel y Wilson, 1975; Wilson, 1984, 1985, 1988; Clowes, 1985), y la Península Antártica (Cookson y Eisenack, 1967; Wrenn y Beckmann, 1982; Wrenn y Hart, 1988; Mohr, 1990; Cocozza y Clarke, 1992; Mao y Mohr, 1995). Comparativamente, los estudios sobre dinoflagelados del Paleógeno medio en el Atlántico Sudoccidental son escasos (Menéndez, 1965; Pöthe de Baldis, 1966; Archangelsky, 1969; Gamerro y Archangelsky, 1981; Goodman y Ford, 1983; Guerstein y Junciel, 2001; Guerstein et al., 2002, 2003, 2008, 2010; Guler y Guerstein, 2002; Daners y Guerstein, 2004).
El género Enneadocysta es un elemento característico de las asociaciones de quistes de dinoflagelados del Eoceno medio y tardío del Océano Austral (Guerstein et al., 2008, 2010). Este género fue recientemente enmendado por Fensome et al. (2006) a partir del estudio de asociaciones de la plataforma continental del este de Canadá y de Tierra del Fuego, Argentina. La especie más abundante en el Atlántico Sudoccidental es E. dictyostila (Menéndez) Fensome et al. [como Areosphaeridium diktyoplokum (Klumpp) Eaton o Enneadocysta partridgei Stover y Williams]. Junto con esta especie, se reporta una escasa representación de E. brevistila (Fensome et al.), E. multicornuta (Eaton) Stover y Williams, y Enneadocysta sp. A de Brinkhuis et al., 2003b. Este conjunto de especies está consistentemente presente en las asociaciones del Eoceno medio a tardío de altas latitudes del Hemisferio Sur y es un importante indicador bioestratigráfico y paleoambiental (Guerstein et al., 2008).
En el presente trabajo se muestra la distribución del género Enneadocysta en el Atlántico Sudoccidental y se discute su presencia en latitudes medias teniendo en cuenta información proveniente de las cuencas del Colorado (Argentina) y Punta del Este (Uruguay). Asimismo, se analiza su abundancia relativa y distribución en relación con los modelos de paleocirculación oceánica para el Océano Austral propuestos para configuraciones previas a la instalación de la Corriente Circumpolar Antártica (Kennett et al., 1975; Kennet, 1977; Huber y Sloan, 2001; Huber et al., 2004).
Marco geológico y paleogeográfico
La apertura del Atlántico Sur se relaciona con la última fase de la ruptura de Gondwana. Es el resultado de un complejo proceso de ruptura continental y magmatismo sobre los márgenes conjugados de la parte sur de África y América del Sur que se dio en forma diacrónica de sur a norte (Uchupi y Emery, 1991). La ruptura inicial ocurrió aproximadamente a los 130 Ma (M 10) entre los 44º S y 45º S, propagándose hacia el norte (Ghidella et al., 2006). El margen continental argentino ha sido definido como un típico margen pasivo de rift volcánico con una amplia plataforma y talud desarrollado sobre extensos flujos basálticos y rocas volcaniclásticas (Hinz et al., 1999). Análisis de reflectores sísmicos profundos en distintas transectas transversales a la plataforma y el talud continental confirman las características de margen pasivo volcánico para el margen argentino-uruguayo (Franke et al., 2006, 2007).
La Cuenca del Colorado es la más austral de una serie de cuencas cuyos ejes están orientados en forma transversal al margen continental. Otras cuencas incluidas en esta serie son las del Salado y Punta del Este, desarrolladas sobre la plataforma argentina y uruguaya, respectivamente. La Cuenca Punta del Este se extiende hacia el norte hasta el Alto de Polonio que la separa de la de Pelotas y limita al sur a través del Alto Martín García con la del Salado (figura 1).

Figura 1. Área de estudio en el Atlántico Sudoccidental. Mapa de distribución de los sitios analizados. Modificado de Urien y Zambrano (1996). / study area in the Southwestern Atlantic. Distribution map of compiled sites. After Urien and Zambrano (1996).
Desde el punto de vista tectono-estratigráfico se reconocen tres fases en la evolución de las cuencas: rift, hundimiento térmico y margen pasivo o deriva (Stoakes et al., 1991; Frydlund et al., 1996; Ucha et al., 2003). Durante la fase de rift se depositan areniscas y conglomerados rojos correspondientes a facies aluviales, restringidas mayormente a los hemigrabens de orientación regional noroeste-sudeste, en el caso del Atlántico Sudoccidental. Franke et al. (2007) señalaron que la finalización de esta fase corresponde a un notable horizonte sísmico que representa una superficie erosiva observable a lo largo de toda la plataforma externa de Argentina y Uruguay. Por encima de esta discontinuidad, las edades de las unidades basales de los depósitos de post-rift varían entre el Aptiano y el Albiano (Stoakes et al., 1991; Juan et al., 1996; Ucha et al., 2003). La fase de hundimiento térmico está caracterizada por sedimentitas depositadas entre el Albiano y el Campaniano (Daners et al., 2004, Guerstein et al., 2005), que señalan una primera transgresión atlántica. Franke et al. (2007) distinguieron un horizonte sísmico como marcador del límite entre las fases de hundimiento térmico y margen pasivo, al que denominaron "equivalente Pedro Luro" (PLe). Por encima de dicho marcador las pelitas de la Formación Pedro Luro en la Cuenca del Colorado contienen el límite Cretácico/Paleógeno y constituyen la unidad basal de la fase de deriva (Frydlund et al., 1996). Finalmente, la fase de deriva está representada por una espesa secuencia cenozoica que corresponde a la sedimentación a lo largo del recientemente formado Océano Atlántico Sudoccidental.
A partir del final del Cretácico (Maastrichtiano) y durante el Cenozoico, la Patagonia extraandina y la plataforma continental sudamericana fueron afectadas por cinco transgresiones marinas atlánticas de carácter regional (Malumián, 1999). Las unidades marinas resultantes alternan con depósitos continentales que incluyen hiatos importantes. El primer ciclo transgresivo-regresivo tuvo lugar en el Maastrichtiano - Daniano y está ampliamente documentado en todas las cuencas del Atlántico Sudoccidental (Malumián, 1999). Luego de un período erosivo se reconoce un segundo ciclo asignado al Eoceno medio sobre la base de foraminíferos y dinoflagelados (Malumián, 1999; Guerstein et al., 2008, 2010). Este evento está mejor documentado al sur de los 40°S en la Cuenca Austral, aunque se lo ha reconocido también en las cuencas del sud-sudeste de Brasil, Punta del Este, Salado y Colorado (Guerstein et al., 2003). Con excepción de la Cuenca Austral, donde el Eoceno medio a tardío aflora en el este y noreste de Tierra del Fuego (formaciones Leticia y Cerro Colorado) y en el sudoeste de Santa Cruz (Formación Río Turbio), los depósitos correspondientes a este ciclo provienen exclusivamente de perforaciones y testigos profundos. El tercer ciclo se inicia con unidades marinas, asociadas a la transición Eoceno-Oligoceno, ampliamente distribuidas en Patagonia, mientras que la fase regresiva está representada por depósitos continentales asignados al Oligoceno temprano. El cuarto ciclo se caracteriza por dos eventos transgresivos muy significativos en términos del área de cobertura. Uno de ellos es el informalmente denominado "Patagoniano", que tuvo lugar durante el Oligoceno tardío-Mioceno temprano, y el otro, referenciado como "Entrerriense", fue datado como Mioceno medio-tardío. El quinto y último ciclo se relaciona con la fase orogénica y levantamiento de los Andes Patagónicos y corresponde al final del Mioceno-Plioceno temprano (Malumián, 1999).
El género Enneadocysta se ha reconocido hasta el momento en las unidades del segundo ciclo y en la parte inferior del tercero. La presencia de ejemplares en los niveles superiores del tercer ciclo se interpreta como material redepositado (Archangelsky, 1969; Guerstein et al., 2008). En la figura 2 se indica la correlación de las formaciones correspondientes al segundo y tercer ciclo en el área de estudio y se grafica la curva eustática propuesta por Miller et al. (2005) para el intervalo comprendido entre los 42 y los 29 Ma.

Figura 2. Correlación de las unidades litoestratigráficas en las cuencas del Atlántico Sudoccidental: Austral, Colorado y Punta del Este. Compilado y modificado de Malumián, 1999; Guler y Guerstein, 2002; Daners et al., 2004; Malumián y Olivero, 2006; Olivero y Malumián, 2008 / correlation of lithostratigraphic units from the Southwest Atlanctic basins: Austral, Colorado and Punta del Este. Compiled and modified from Malumián, 1999; Guler and Guerstein, 2002; Daners et al., 2004; Malumián and Olivero, 2006; Olivero and Malumián, 2008.
Materiales y métodos
Los materiales analizados provienen de las perforaciones Pejerrey-1, Cruz del Sur-1 y C x-1 (plataforma continental argentina, Cuenca del Colorado) y Gaviotín-1 (plataforma continental uruguaya, Cuenca Punta del Este) (figura 1). Las muestras fueron procesadas para el análisis de dinoflagelados en los laboratorios palinológicos del Servicio Geológico Canadiense, Dartmouth, Nova Scotia (GSC, Atlantic), Museo Argentino de Ciencias Naturales "Bernardino Rivadavia", Buenos Aires, Argentina (MACN), INGE-OSUR, Bahía Blanca y Universidad de la República, Montevideo, Uruguay. En la tabla 1 se indican las muestras en las que se registró el género Enneadocysta en términos de la profundidad de perforación, el laboratorio de extracción físicoquímica donde se procesaron las muestras y la colección donde se encuentran archivados los preparados palinológicos.
Tabla 1. Distribución y frecuencias de las especies de Enneadocysta registradas en las distintas perforaciones analizadas en la Cuenca Punta del Este y Cuenca del Colorado/ Enneadocysta species distribution and frequencies recorded in Punta del Este Basin and Colorado Basin boreholes.

En todos los casos, el tratamiento de extracción físicoquímica consistió en la eliminación de la fracción inorgánica con ácidos clorhídrico y fluorhídrico. La fracción orgánica se concentró mediante filtrados con tamices de 10, 25 y 180 µm. En el caso de las muestras provenientes de las perforaciones Pejerrey -1 y C x-1 los residuos fueron teñidos con Bismarck C; los residuos de las muestras del pozo Gaviotín-1, Pejerrey-1 y Cruz del Sur-1 fueron montados en gelatina glicerina mientras que los de las muestras de la perforación C x-1, procesadas en el GSC, fueron montados en celulosa Hidroxietil y pegados con elvacita.
Otros materiales observados incluidos en la discusión de este trabajo corresponden a (1) muestras originales del Pozo La Sara x-1, estudiadas por Menéndez (1965); (2) muestras provenientes de la Formación Leticia (sección Río de la Turba y Cabo Campo del Medio), Tierra del Fuego, Argentina; (3) residuos optimizados de la Formación Río Turbio (muestras originalmente estudiadas por Archangelsky, 1969) y (4) muestras del testigo DSDP 511, Plateau de Malvinas, cedidas por D. Goodman. El holotipo y los preparados de Menéndez (1965) están depositados en la Colección Palinológica del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia", Buenos Aires, Argentina.
Las fotomicrografías fueron tomadas utilizando un microscopio Nikon Eclipse 600 nº 772751 en el IN-GEOSUR, Departamento de Geología de la Universidad Nacional del Sur. Se indican las coordenadas de la escala Vernier de dicho microscopio y las referencias England Finder para cada ejemplar ilustrado.
Se utilizó un microscopio electrónico de barrido ElectroScan E3 del Servicio Geológico Canadiense Atlántico (Dartmouth) bajo presión parcial de vapor de agua. Para el estudio bajo microscopio electrónico los especímenes fueron previamente recubiertos con oro. En el Apéndice I se listan las especies mencionadas en el texto de acuerdo al Índice de Dinoflagelados de Fensome y Williams (2004) y Fensome et al. (2008). La escala de tiempo utilizada es la propuesta por Gradstein et al. (2004).
Enneadocysta dictyostila y especies afines: su historia
Enneadocysta dictyostila, fue descripta por Menéndez (1965) como una especie de Hystrichosphaeridium en material del Eoceno proveniente de la perforación La Sara x-1 en el norte de la Isla Grande de Tierra del Fuego (figura 3.1-3). Subsecuentemente, Pöthe de Baldis (1966) describió e ilustró el mismo morfotipo, recuperado de sedimentos paleógenos de otra perforación de Tierra del Fuego y, sin hacer referencia a la especie de Menéndez, lo asignó a la especie alemana Hystrichosphaeridium diktyoplokum Klumpp (figura 4.1). Cranwell (1964) y Wilson (1967) identificaron el morfotipo argentino como Cordosphaeridium diktyoplokum en depósitos paleógenos de Antártida. Cookson y Cranwell (1967) asignaron los especímenes de "Hystrichosphaeridium dictyostilum" presentes en el Paleógeno del sur de Chile a Cordosphaeridium diktyoplokum, considerando formalmente a esta especie el sinónimo taxonómico senior de Hystrichos-phaeridium dictyostilum. Tanto Archangelsky (1969) como Fasola (1969) aceptaron esta sinonimia al estudiar depósitos del sur de Chile y Argentina, así como Eaton (1971, p. 359) al transferir la especie descripta por Klumpp a su nuevo género Areosphaeridium.

Figura 3.1-2. Holotipo de Enneadocysta dictyostila, vista antapical de un espécimen de la perforación La Sara LSX4, Tierra del Fuego, Argentina, muestra 1301, 1 BA PB: 20-89,5- (microscopio Watson 90.128), England Finder K38/2; 1, representa un foco intermedio tomado con un microscopio Nikon Eclipse 600 usando un sistema óptico de contraste de interferencia; 2-3, dibujos del mismo ejemplar; 2, superficie antapical; 3, superficie apical (cara interna) /Enneadocysta dictyostila, holotype, antapical view of specimen from Borehole La Sara LSX4, Tierra del Fuego, Argentina, sample 1301, slide 1 BA PB: 20-89,5- (microscope Watson 90.128), England Finder K38/2; 1, represents an intermediate focal level using an interference contrast optical system; 2-3, Sketches of the same specimen; 2, representing the antapical surface; 3, representing the apical surface (internal face).
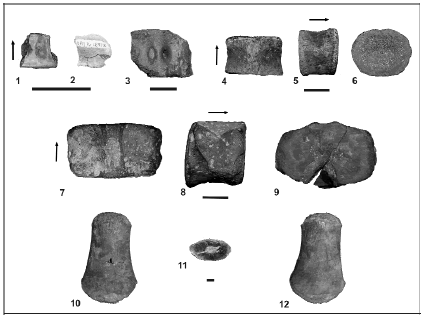
Figura 4.1-6. Comparación entre los géneros Areosphaeridium y Enneadocysta; 1, 4, Bosquejos mostrando la vista apical de ambos géneros; 1, A. diktyoplokum (de Williams et al., 2004 fig. P1, 16); 4, Topotipo de E. dictyostila del miembro Lb de la Formación Leticia; muestra de afloramiento PT57, Gr. La Despedida, Cabo Campo del Medio, sudeste de Tierra del Fuego, Argentina, Preparado P36632-01: 34.5 x 104.0, England Finder Q42/1; 2-3, 5-6, Dibujos esquemáticos mostrando la distribución de los procesos (círculos llenos) en relación a la tabulación interpretada en especies de Areosphaeridium y Enneadocysta; 2-3, A. diktyoplokum; 2, vista apical; 3, vista antapical; 5-6, E. dictyostila y E. brevistila; 5, vista apical; 6, vista antapical. Los asteriscos indican que la designación de la placa se basa en homología con el patrón gonyaulacoideo estándar, no con la interpretación Kofoideana (ver Fensome et al., 1993)/ comparison between the genera Areosphaeridium and Enneadocysta; / 1, 4, Sketches showing ventral views of both genera; 1, A. diktyoplokum (from Williams et al., 2004 fig. P1, 16); 4, E. dictyostila topotypic specimen from the Lb member of the Leticia Formation; outcrop sample no. 57, La Despedida Group at Cabo Campo del Medio, southeastern Tierra del Fuego, Argentina, specimen on slide P36632-01: 34.5 x 104.0, England Finder Q42/1; 2-3, 5-6, Schematic drawings showing the distribution of processes (filled circles) relative to interpreted tabulation in species of Areosphaeridium and Enneadocysta; 2-3, A. diktyoplokum; 2, apical view; 3, antapical view; 5-6, E. dictyostila and E. brevistila; 5, apical view; 6, antapical view. Asterisks indicate that plate designation is based on homology with the standard gonyaulacalean pattern rather than on a literal Kofoidian interpretation (see Fensome et al., 1993).
Por su parte Sarjeant (1981) enmendó la diagnosis de Hystrichosphaeridium (como Areosphaeridium) dictyostilum y propuso retener esta especie separadamente. Lentin y Williams (1985) aceptaron esta propuesta, pero debido al bajo grado de preservación de los ejemplares y a la consecuente imposibilidad de determinar la paratabulación sin observar el material tipo, propusieron que el nombre fuera retenido sólo para el holotipo. Goodman y Ford (1983) registraron especímenes de "Areosphaeridium sp. cf. A. diktyoplokum" del Eoceno terminal-Oligoceno inferior del testigo DSDP 71-sitio 511, sobre el Plateau de Malvinas. Estos autores diferenciaron este taxon de Areosphaeridium diktyoplokum teniendo en cuenta los márgenes irregulares de las plataformas distales de los procesos, de manera equivalente a lo que posteriormente Fensome et al. (2006) definieron como ragged clypeate processes (text-fig. 3 M-N). Goodman y Ford (1983, p. 865) destacaron que ninguna de las formas registradas hasta entonces en el Hemisferio Sur "debería ser atribuida a A. diktyoplokus s.s".
Stover y Williams (1995), tal como lo habían señalado anteriormente otros autores, notaron diferencias entre los especímenes del Hemisferio Norte y los del Hemisferio Sur asignados a Areosphaeridium diktyoplokum. Sin embargo, Stover y Williams reconocieron formalmente esas diferencias creando la especie Enneadocysta partridgei para ubicar a la mayoría de los especímenes del Hemisferio Sur que habían sido previamente asignados a Areosphaeridium diktyoplokum. En la misma contribución Stover y Williams transfirieron Hystrichosphaeridium dictytostilum a Enneadocysta, considerando esta asignación provisional ante la imposibilidad de definir el tipo de arqueopilo y la distribución de los procesos.
Fensome et al. (2006) reestudiaron el material tipo de Menéndez, junto con gran cantidad de ejemplares de Enneadocysta dictyostila provenientes de muestras del norte y sudeste de Tierra del Fuego. La distribución de los procesos, particularmente en el área antapical, así como la evidente compresión dorso-ventral y el desplazamiento del sulcal notch hacia la izquierda permitió diferenciarlo claramente de Areosphaeridium diktyoplokum. Estas dos últimas son características de la familia Areoligeraceae a la cual fue transferido el género Enneadocysta (figura 4). Asimismo, Fensome et al. (2006) consideraron a E. partridgei un sinónimo junior de E. dictyostila.
Entre las especies asociadas a Enneadocysta dictyostila pero registradas en frecuencias menores en las asociaciones del Hemisferio Sur, se encuentran E. arcuata, E. multicornuta, E. brevistila y Enneadocysta sp. A de Brinkhuis et al., 2003b (figuras 5 y 6). En trabajos recientes, a los efectos de cuantificar las variaciones en las asociaciones de quistes de dinoflagelados, estas especies son agrupadas como Enneadocysta spp. (Sluijs et al., 2005; Warnaar, 2006; Guerstein et al., 2008). En todos los casos el complejo de especies está ampliamente dominado por E. dictyostila pero puede observarse una importante variabilidad morfológica con un gran número de ejemplares ubicados en posiciones intermedias entre los distintos extremos del complejo.

Figura 5. 1-6. Enneadocysta dictyostila; 1-2, vista ventral/ ventral view, UNSLP PEJ-1650: 45x102 (EF L31/3); 1, foco alto sobre superficie ventral / high focus on ventral surface; 2, foco bajo sobre superficie dorsal / low focus on ventral surface; 3, vista lateral derecha, foco alto / right lateral view, high focus, UNSLP PEJ-1650: 50.4x101 (EF K25/3); 4, vista interna de un opérculo / internal view of an operculum, UNSLP PEJ-1650: 37.7x101.7 (EF L38/2); 5, vista ventral, foco intermedio, el opérculo permanece adherido / ventral view, intermediate focus, the operculum remains attached. Especimen de la Formación Río Turbio, Sudoeste de Santa Cruz, Argentina / specimen from the Río Turbio Formation, southwestern Santa Cruz Province, Argentina, UNSLP 230 (+25)-A: 50.8X104.7 (EF O25/1); 6, Ejemplar con procesos relativamente cortos, foco alto/ specimen with shorter processes, high focus, UNSLP PEJ-1600: 35.5x101 (EF K41/3). 7-9, Enneadocysta brevistila; 7-8, vista ventral / ventral view, Gr. La Despedida en Cabo Campo del Medio, sudeste de Tierra del Fuego, Argentina, UNSLP P36632- 01: 47.1x100.5 (EF K34/4); 7, foco alto sobre superficie ventral / high focus on ventral surface; 8, foco bajo sobre superficie dorsal/ low focus on dorsal surface; 9, vista ventral, foco alto sobre superficie ventral, ejemplar completo con opérculo / ventral view, high focus on ventral surface, complete specimen with operculum. UNSLP Cx-1 1000-05-1: 39.3x94.6 (EF D37/0); 10, Enneadocysta multicornuta, foco intermedio / intermediate focus, DPFC Gaviotín -1 1576-D: 34x110.5 (EF U42); 11-12, Enneadocysta sp. A (Brinkhuis et al., 2003b), vista lateral derecha, ejemplar con opérculo/ right lateral view, operculum attached, DPFC Gaviotín -1 1576-E: 43.4x102 (EF L33/3); 11, foco alto sobre superficie laterla derecha / high focus on right lateral surface; 12, foco bajo sobre superficie lateral izquierda / high focus on left lateral surface. Todas las imágenes fueron tomadas usando un sistema óptico de contraste de interferencia. EF, England Finder. Escala gráfica: 20 μm / all digital images were taken using an interference contrast optical system. EF, England Finder references. Scale bar: 20 μm.

Figura 6. 1-4. Enneadocysta dictyostila; 1, vista lateral derecha/ right lateral view; 2, vista lateral derecha/ right lateral view; 3, Vista antapical/ antapical view; 4, Opérculo / operculum. 5-6, Enneadocysta brevistila; 5, vista lateral derecha / right lateral view; 6, vista dorsal oblicua / oblique dorsal view. Fotomicrografías tomadas con MEB de ejemplares del miembro Lb de la Formación Leticia, Grupo La Despedida en Cabo Campo del Medio, sudeste de Tierra del Fuego, Argentina; muestra de afloramiento PT57, 220 por encima de la base del miembro Lb / SEM photomicrographs of specimens from the Lb member of the Leticia Formation, La Despedida Group at Cabo Campo del Medio, southeastern Tierra del Fuego, Argentina; outcrop sample PT57, 220m above the base of the Lb member. Escala gráfica: 20 μm / scale bar: 20 μm.
Distribución en las cuencas del Atlántico Sudoccidental
En la tabla 1 se muestran los registros de las especies de Enneadocysta halladas en latitudes medias del Atlántico Sudoccidental. Se indica también la distribución de frecuencias semicuantitativas de cada especie y las distintas bioestratigrafías propuestas (Quattrocchio y Guerstein, 1994; Archangelsky et al., 1997; Guerstein y Junciel, 2001; Guler y Guerstein, 2002; Daners y Guerstein, 2004).
Debido a las características del muestreo (cuttings), el análisis bioestratigráfico se basa fundamentalmente en datos de últimas ocurrencias o techos estratigráficos, dado que las bases o primeras ocurrencias suelen estar distorsionadas por la contaminación que sufre el material al ascender por las paredes del pozo. En cuanto a la información semicuantitativa, Enneadocysta está presente de manera abundante (entre 21 y 30 ejemplares por preparado) en las asociaciones del Eoceno medio en los pozos Pejerrey-1 y Gaviotín-1, y es un elemento común (hasta 20 ejemplares por preparado) en las asociaciones del Eoceno medio de los pozos Cruz del Sur-1 y Estrella-1. En los pozos Cx-1 y Dx-1 no está representado el Eoceno medio y el género Enneadocysta se registra sólo en niveles asignados a la transición Eoceno-Oligoceno donde se hallaron menos de 5 ejemplares por preparado.
El único registro de Enneadocysta en la Cuenca del Colorado anterior al Eoceno es el mencionado por Quattrocchio y Sarjeant (1996), quienes consideraro "in situ" la presencia de E. dictyostila (como A. diktyo plokum) en una asociación asignada al Daniano en e pozo Puerto Belgrano 20 (figura 1). Dichos autore analizaron dos testigos laterales, por lo que se inter preta que la presencia de dos únicos ejemplares ha llados en sólo una de las muestras extraídas de la paredes del pozo puede deberse a la contaminació con material de niveles superiores durante el ascens de la muestra.
En la figura 7 se muestra la distribución de Enneadocysta en el Atlántico Sudoccidental, incluyendo las proporciones máximas sobre el total de quistes de dinoflagelados que se registraron en distintas secciones de la Cuenca Austral. En esta última, las asociaciones analizadas provienen de afloramientos y testigos de sedimento por lo que las frecuencias relativas corresponden a recuentos estadísticamente significativos. En la Formación Leticia, Eoceno medio de Tierra del Fuego (figura 7.1), Enneadocysta dictyostila es claramente dominante dentro del complejo, tanto en las barrancas del Río de la Turba como en la sección Cabo Campo del Medio. En Río de la Turba las mayores frecuencias se observan en la parte media de la sección con un máximo de 304 ejemplares por preparado, que representa un 76% del total de quistes de dinoflagelados (Guerstein et al., 2008; tabla 1, pág. 87; fig 7., pág. 88). En el miembro medio de la Formación Leticia, Cabo Campo del Medio, se registró un máximo de 276 ejemplares por preparado que constituye el 54% del total de dinoflagelados. En los miembros inferior y superior las frecuencias varían entre 5 y 10%, respectivamente (RG, obs. pers.)

Figura 7. Reconstrucciones paleogeográficas para el Eoceno medio-tardío (7.1) y transición Eoceno-Oligoceno (7.2) en la Cuencas del Atlántico. Líneas punteadas indican paleolíneas de costa y márgenes continentales (modificado de Malumián, 1999) / palaeogeographical reconstructions for the middle-late Eocene (7.1) and the Eocene-Oligocene transition (7.2) in the Southwest Atlantic basins. Dashed lines indicate palaeo- shorelines and continental margins (modified from Malumián, 1999).
Archangelsky (1969) estudió las asociaciones de quistes de dinoflagelados de la Formación Río Turbio provenientes de testigos de Yacimientos Carboníferos Fiscales extraídos en el sudoeste de la provincia de Santa Cruz. Recientemente se optimizaron los residuos palinológicos de dichas muestras y su análisis permitió determinar que las altas abundancias del morfotipo asignado por Archangelsky a Cordosphaeridium diktyoplokum corresponde a Enneadocysta dictyostila, junto a unos pocos ejemplares de E. brevistila. En los niveles inferiores de la Formación Río Turbio este morfotipo supera el 75% del total de paleomicroplancton, proporciones que disminuyen gradualmente hacia la parte media en la que se registra la última ocurrencia certera. El único ejemplar hallado en los niveles superiores ha sido interpretado como material redepositado (Archangelsky, 1969, pág. 209, cuadro II).
En las muestras de la transición Eoceno-Oligoceno en la Cuenca Austral, Enneadocysta se encuentra presente en frecuencias muy bajas (se contabilizaron hasta 17 ejemplares por preparado), valores que no superan el 10% del total de dinoflagelados (Guerstein et al., 2008, tablas 2-4; figs. 8-10) (figura 7.2). Asimismo, Enneadocysta dictyostila fue hallada en el Sitio 511 del testigo DSDP 71 sobre el margen oriental del Plateau de Malvinas. Los 185 metros superiores del testigo fueron asignados al Eoceno tardío-Oligoceno temprano, de acuerdo con el contenido de foraminíferos planctónicos, nanofósiles calcáreos y radiolarios (Basov et al., 1983). Goodman y Ford (1983) señalaron la presencia de Enneadocysta dictyostila (como Areosphaeridium sp. cf. A. dyktyoplokum) desde los niveles atribuidos al Eoceno tardío (muestra 18-2, 40-42 por debajo del límite Eoceno/Oligoceno) hasta la muestra 3-2, 96-98, asignada al Oligoceno temprano.
Se analizó un set de preparados palinológicos del testigo 511 cedidos por D. Goodman con el objeto de obtener datos cuantitativos. Se reconocieron 20 ejemplares de Enneadocysta dictyostila y 2 de E. brevistila en la muestra 18-2, 40-42 y la presencia de un par de ejemplares mal preservados de Enneadocysta spp. en uno de los niveles superiores del testigo en la muestra 5-2, 55-57 (Guerstein et al., 2010; Cayulef, com. pers. ) Estos datos muestran las mismas tendencias que las de los resultados analizados precedentemente, confirmando que las especies de Enneadocysta, cuando presentes en el Eoceno tardío, se caracterizan por frecuencias muy bajas. En la plataforma de Brasil, Regali et al. (1974a, 1974b) y Regali (1980) registraron la presencia de "Cordosphaeridium diktyoplokus"en el Eoceno medio de las cuencas de Bahía, Espirito Santo y Sergipe, del Sud-Sudeste de Brasil y en la Cuenca de Ceará, en el área ecuatorial de Brasil. Según Arai y Lana (2007, com. pers.), todos los ejemplares de la plataforma brasilera asignados originalmente a "Cor-dosphaeridium diktyoplokus" corresponden a Enneadocysta dictyostila. Dichos autores consideraron que los registros mencionados por Regali y colaboradores son consistentes en el Eoceno medio. Sin embargo, no hay datos de abundancia proveniente de los estudios realizados en la década del 70. Recientemente, Ferreira (2004) realizó un estudio detallado del Paleoceno-Eoceno en la Cuenca de Sergipe-Alagoas y registró la presencia de Enneadocysta dictyostila (como E. partridgei) y Enneadocysta arcuata en el Eoceno medio y tardío, respectivamente. Gran parte de los materiales analizados provienen de muestras de corte con alternancia de testigos; estos últimos permiten ajustar tanto los datos de distribución estratigráfica como la información cuantitativa. Las frecuencias de Enneadocysta son siempre inferiores al 10% del total de dinoflagelados con excepción de uno de los pozos (SG-11), donde alcanza valores cercanos al 20% sólo en una de las asociaciones. Si bien Ferreira (2004) no presenta datos crudos, los valores relativos son consistentes con la distribución observada en las cuencas del Colorado y Punta del Este.
De acuerdo con la información analizada, es evidente que Enneadocysta es un elemento característico de las asociaciones del Eoceno medio en todas las cuencas del Atlántico Sudoccidental. Sin embargo, es importante destacar que mientras en la Cuenca Austral las frecuencias superan el 50% del total de quistes de dinoflagelados en los sitios de latitudes medias y bajas las abundancias y proporciones registradas son significativamente menores.
Discusión
Significado bioestratigráfico
En la plataforma continental del sudeste de Brasil, Ferreira (2004) estudió las asociaciones palinológicas de nueve perforaciones profundas realizadas en la Cuenca de Sergipe-Alagoas. La información palinológica fue calibrada con una bioestratigrafía independiente basada en nanofósiles calcáreos y foraminíferos. De acuerdo con esta información, Enneadocysta arcuata se registra sólo en la base de la zona N470 de Antunes et al. (2004), que corresponde a la parte media del Priaboniano (figura 8). Enneadocysta dictyostila (como E. partridgei) está presente desde la base del Lutetiano, dentro de la zona N430 hasta la zona N450, con la última ocurrencia en la mitad del Bartoniano. Por otra parte, Enneadocysta spp. se registra desde la base de la zona N437 hasta el techo de la zona N450, con la última ocurrencia en el límite Bartoniano/Priaboniano (figura 8).

Figura 8. Distribución estratigráfica de las distintas especies de Enneadocysta registradas en el Hemisferio Sur / stratigraphic distribution of Enneadocysta species recorded in the Southern Hemisphere.
Enneadocysta dictyostila es la especie más representativa del complejo, y tiene una amplia distribución en el Hemisferio Sur. Además de las localidades del Atlántico Sudoccidental mencionadas, ha sido hallada in situ en depósitos del Eoceno de Antártida, Mar de Tasmania, Australia y Nueva Zelanda, principalmente en paleolatitudes altas. Como ya fue comentado, hasta 1995 fue asignada y/o comparada con Areosphaeridium diktyoplokum, como Hystrichosphaeridium, Cordosphaeridium o Areosphaeridium (Stover y Williams, 1995; apéndice 2: F-1 y F-2, pág. 140). Posteriormente, fue registrada como Enneadocysta partridgei en el Eoceno del margen continental de Tasmania (Truswell, 1997); en la transición Eoceno-Oligoceno de Antártida, en el testigo CIROS-1, con la última ocurrencia confirmada aproximadamente a los 32 Ma, en el magnetocrón C12n (Hannah, 1997; Wilson et al., 1998); en el Eoceno de McMurdo Sound, Antártida (Levy y Harwood, 2000); en el Eoceno medio a tardío de los testigos ODP 189 sitios 1170, 1171 y 1172, plataforma de Tasmania (Brinkhuis et al., 2003a, b; y Sluijs et al., 2003), y en el Eoceno medio? a tardío en el Sitio 1166, en el este de Antártida (MacPhail y Truswell, 2004).
Los rangos estratigráficos más precisos obtenidos hasta el momento para las especies de Enneadocysta en el Océano Austral son los determinados por Brinkhuis et al. (2003b) y Stickley et al. (2004a) en el testigo ODP 189-sitio 1172 (figura 8). Sobre este testigo se estableció un control de edad independiente basado en magnetoestratigrafía y bioestratigrafía. Entre los grupos fósiles estudiados se incluyen foraminíferos, nanofósiles calcáreos, diatomeas, radiolarios y dinoflagelados. Estos datos permitieron correlacionar los eventos de quistes de dinoflagelados con la escala de tiempo de polaridad geomagnética (GPTS). Brinkhuis et al. (2003b) registraron la primera ocurrencia de Enneadocysta dictyostila (como E. partridgei) dentro del magnetocrón C21r (48,5 Ma) y la última dentro del magnetocrón C13r (33,5 Ma). En el Sitio 1168, también calibrado con magnetoestigrafía, el registro más joven corresponde a los 32,8 Ma, dentro del magnetocrón C12r (Brinkhuis et al., 2003a). El rango estratigráfico se extiende entonces desde el Eoceno medio temprano hasta el comienzo del Rupeliano.
Enneadocysta sp. A está presente en el Sitio 1172 desde el magnetocrón C21n (46,6 Ma) hasta el magnetocrón C13r (33,5 Ma). Por otra parte, Enneadocysta brevistila fue hallada en el mismo sitio en el testigo 40, secciones 2, 3 y 4 (Fensome et al., 2006). Según Stickley et al. (2004a) este intervalo corresponde al magnetocrón C16r2r, equivalente a los 36,4 Ma (Priaboniano temprano). Asimismo, el registro más alto de E. pectiniformis coincide con el magnetocrón C16r2r, mientras que la última ocurrencia en forma abundante de Enneadocysta spp. se reconoce dentro del magnetocrón C16n2n, equivalente a los 35,7 Ma, aproximadamente 2 Ma antes del último registro confirmado del género (Brinkhuis et al., 2003b).
Las asociaciones de quistes de dinoflagelados de la Cuenca Austral en Tierra del Fuego y Santa Cruz, así como en la plataforma continental en las cuencas Punta del Este y Colorado muestran los máximos valores de Enneadocysta en el Eoceno medio (figuras 7.1; 8). En la Cuenca Austral las últimas ocurrencias en forma abundante se registran hasta el Bartoniano, mientras que en Colorado y Punta del Este la posición estratigráfica de los niveles con valores relativamente más altos de Enneadocysta se asigna al Eoceno medio, pero no puede definirse con mayor precisión. Por su parte, Ferreira (2004) señaló que hay un máximo de E. dictyostila en la Cuenca de Sergipe, registrado en la zona de nanofósiles calcáreos N470, aproximadamente a los 43,5 Ma (figura 8).
Significado paleoambiental
Según Cocozza y Clarke (1992, figura 3), Enneadocysta dictyostila (como A. diktyoplokum) representa hasta el 50% de las asociaciones de quistes de dinoflagelados en la parte alta del Eoceno en la Formación La Meseta, en el norte de la Isla Seymour. Asimismo, Mao y Mohr (1995, figura 4) describieron frecuencias superiores al 50% del total de quistes de dinoflagelados en una asociación del Eoceno medio proveniente de un testigo del Banco Brunce en el Mar de Scotia, sobre la plataforma antártica.
Wrenn y Beckmann (1982) definieron a la "Flora Transantártica" a partir de bloques erráticos de Mc-Murdo Sound como una flora endémica integrada por elementos mayoritariamente restringidos a altas latitudes del Hemisferio Sur. Subsecuentemente, Wrenn y Hart (1988) describieron su composición y distribución, reconociendo a esta flora en distintos sitios del Océano Austral. Es importante destacar que en trabajos publicados con anterioridad a 1995, la abundancia de Enneadocysta dictyostila (como Areosphaeridium diktyoplokum) fue considerada una característica de las palinofloras del Terciario temprano, en particular del Eoceno del Océano Austral. Sin embargo, Enneadocystaictyostila no fue originalmente considerada un miemro de esta flora endémica debido a su asignación a reosphaeridium diktyoplokum, una especie con amplia istribución en el Hemisferio Norte. Esta confusión taonómica enmascaró el potencial de la especie argentina como indicador paleoambiental hasta que Stover Williams (1995) diferenciaron taxonómicamente a E. ictyostila (como E. partridgei) de A. diktyoplokum. ambién establecieron una diferenciación espacial entre ambas especies basada en el reconocimiento de que odos los registros de esta última corresponden al emisferio Norte mientras que E. dictyostila (como E. artridgei) se restringe al Hemisferio Sur.
Subsecuentemente, Truswell (1997) revisó la distribución de las floras de quistes de dinoflagelados e altas latitudes del Eoceno-Oligoceno y destacó ue Enneadocysta dictyostila (como E. partridgei) era un omponente común de la denominada Flora ransantártica de Wrenn y Beckmann (1982) y de la isma manera fue considerada por Hannah (1997). os resultados de los estudios realizados sobre los estigos ODP 189 al sur de Tasmania permitieron onfirmar que esta especie es un componente importante de la flora endémica (Sluijs et al., 2003).
Röhl et al. (2004) registraron dominancias alternantes de Enneadocysta spp. y Deflandrea spp. duran te parte del Eoceno medio en el testigo ODP 189 sitio 1172, al este de Tasmania. Estos autores correlaciona ron las máximas frecuencias de Enneadocysta spp. con las mayores concentraciones de nanoplancton calcá reo, mientras que los máximos de Deflandrea spp. son coincidentes con los intervalos caracterizados po floraciones de diatomeas y altas concentraciones d nutrientes. De acuerdo con esta información, Röhl e al. (2004) propusieron que los máximos de Ennea docysta podrían reflejar aguas más cálidas, oligotróficas, en áreas de plataforma, mientras que los d Deflandrea indicarían condiciones eutróficas en am bientes someros. Las fluctuaciones observadas en e sitio 1172, calibrado con magnetoestratigrafía, fueron interpretadas como la respuesta a variaciones orbitales.
Sobre la base de observaciones personales (R.G. se determinó un máximo de Enneadocysta spp. en e miembro medio de la Formación Leticia en Tierra de Fuego, coincidente con el único intervalo en el que s hallaron nanofósiles calcáreos (Malumián et al., 1994 Olivero, com. pers., 2004). De acuerdo con estos da tos y considerando que Fensome et al. (2006) transfi rieron el género Enneadocysta a la familia Areoli geraceae, puede inferirse que Enneadocysta dictyostil y especies afines habrían tenido hábitos autotrófico con óptimo desarrollo en aguas superficiales bien iluminadas.
Sobre la base de lo expuesto hasta el momento podría considerarse que Enneadocysta habría preferi do aguas relativamente cálidas y que por lo tanto podría haber sido un grupo de dinoflagelados caracte rístico de latitudes medias a bajas. Sin embargo, la altas proporciones de Enneadocysta dictyostila y especies afines sólo se observan en la Cuenca Austral y en sitios circum-antárticos, lo que pone de manifiesto que este grupo de dinoflagelados fue un constituyente mayoritario de la flora endémica durante e Eoceno medio. Estos resultados sugieren una clara preferencia de Enneadocysta dictyostila y especies afines por aguas frías dentro de una variabilidad climática de gran escala, donde la alternancia con Deflandrea spp. podría reflejar condiciones locales y una variabilidad de escala menor tal como la dada por los ciclos de Milankovitch.
Por otra parte, las muestras de testigos estudiada para palinología y micropaleontología del pozo SE-2 de la Cuenca de Sergipe, Brasil fueron analizadas pa ra establecer relaciones isotópicas de oxígeno y carbono (Ferreira, 2004). El nivel con la mayor represen tación de Enneadocysta coincide con un aumento en la relación δ18O con valores cercanos a -2,3, indicando un enfriamiento de las aguas superficiales. Esta auto ra registró las mayores frecuencias de Enneadocysta y el aumento en los valores isotópicos de oxígeno asociados a un importante evento paleoceanográfico que tuvo lugar en el Lutetiano (Zona N447, ca. 43,5 Ma.). Dicho evento se evidencia por la abundancia de Tasmanites, un aumento en el aporte de palinomorfos de origen continental (esporas y polen), presencia de palinoforaminíferos y foraminíferos bentónicos (Uvigerina). Según Ferreira (2004), este conjunto de datos sugiere temperaturas de aguas marinas más frías que las registradas, tanto en los niveles inferiores como en los inmediatamente superiores.
Implicancias paleoceanográficas
De acuerdo con los patrones de distribución establecidos empíricamente, Enneadocysta dictyostila y especies afines conforman un elemento característico del Océano Austral, propio de regiones al sur de los 45°S (Warnaar, 2006). Considerando la distribución de Enneadocysta en el Atlántico Sudoccidental (figuras 7 y 8), así como la representación significativamente mayor en la Cuenca Austral y en áreas circumantárticas, es importante analizar el significado de la presencia de este grupo de dinoflagelados en las asociaciones de las cuencas del Colorado, Punta del Este y en la plataforma de Brasil.
Dado que la velocidad promedio con la que se trasladan los dinoflagelados modernos en el estado móvil es de aproximadamente de 3 x 10-4 m/s (Smayda, 2002), es evidente que su distribución depende, entre otros factores, de los patrones de circulación oceánica. Es necesario revisar, entonces, los modelos de paleocirculación propuestos para tiempos paleógenos previos a la instalación de la Corriente Circumpolar Antártica (CCA).
Los procesos de apertura y profundización de los pasajes de Drake y Tasmania en el Océano Austral jugaron un rol preponderante en la instalación de la CCA y, de acuerdo con Kennett (1977), en el consecuente aislamiento térmico de Antártida, considerado hasta entonces la causa principal del enfriamiento ocurrido en el límite Eoceno-Oligoceno. Según Kennett et al. (1975) y Kennett (1977), en un escenario previo a la apertura y profundización de los pasajes en el Océano Austral, las corrientes provenientes de latitudes bajas habrían llegado hasta latitudes altas transportando calor y manteniendo las costas antárticas relativamente cálidas. Exon et al. (2004) analizaron la circulación en el área al sur de Tasmania con anterioridad a los 33 Ma y propusieron la existencia de una corriente cálida equivalente a una proto-Corriente Australiana Oriental que, según estos autores, alcanzaría paleloatitudes cercanas a los 70°S (Exon et al., 2004, pág. 35 fig. 7 A y B).
Sin embargo, el sistema climático terrestre se comporta de manera tal que la circulación oceánica superficial responde a los patrones de circulación at mosférica (Siedler et al., 2001; Marshall y Plumb 2008). De acuerdo con los modelos generales de circulación, la corriente que se desplaza hacia el sur de Giro Subtropical (equivalente a la Corriente de Brasil) habría alcanzado como máximo la latitud donde el rotor del viento se anula (~ 45°S). Esto no se cumple en el modelo de Kennett y de hecho, la distribución de quistes de dinoflagelados registra dos para altas latitudes del Eoceno medio en e Océano Austral es incompatible con el escenario propuesto por dicho modelo. Sin una efectiva barre ra física separando las aguas del Océano Austral de las aguas de latitudes medias surgen como interro gantes ¿qué mecanismo habría favorecido tan marcado endemismo durante el Paleógeno hasta el fina del Eoceno? y ¿cómo se compatibilizan los patrone de circulación propuestos por Kennett y colaborado res con la presencia confirmada de especies de Enneadocysta y otros elementos endémicos al Océano Austral en latitudes medias y bajas del Atlántico Sudoccidental?
Contrariamente a la hipótesis propuesta por Kennett, Huber et al. (2004) combinaron los resultados de experimentos de modelos acoplados de circulación general (CGCM) con información de los testigos ODP 189. El esquema de circulación oceánica superficial propuesto para el Eoceno del Océano Austral es considerablemente diferente a las hipótesis previas y explica los distintos grados de endemismo mostrados por los patrones de distribución de quistes de dinoflagelados. En lugar de corrientes oceánicas cálidas dirigiéndose hacia el sur, los resultados de los experimentos de Huber et al. (2004) mostraron amplios giros en sentido horario alrededor de la Antártida (figura 9). El endemismo puede haber sido el resultado de un mecanismo selectivo intrínseco provocado por el "cold trap". Por otra parte, estos giros se intensifican en sus bordes occidentales produciendo corrientes relativamente frías, dirigidas hacia el norte a lo largo del margen oriental de Australia y de América del Sur. La primera, denominada Corriente de Tasmania (CT) por Huber et al. (2004) y Stickley et al. (2004b, fig. 5; pág. PA4027), explicaría los altos porcentajes de quistes de dinoflagelados endémicos en los sitios al sur y este de Tasmania, frente a muy bajos porcentajes registrados en el sitio 1168, sobre la plataforma occidental de Tasmania bañada por una Proto-Corriente de Leeuwin (PCL), relativamente más cálida.

Figura 9. Mapa paleogeográfico para la región Circumpolar Antártica durante el Eoceno medio (40 Ma - Bartoniano). Mapa generado por Plate Tectonic / Reconstructions On-line Paleogeographic Mapper [http://www.serg.unicam.it/Reconstructions. htm]. Áreas en gris indican corteza continental. Líneas en negro representan circulación oceánica esquemática (modificado de Sloan y Rea, 1996; Huber y Sloan, 2001). Abreviaturas: p- GW, proto-Giro de Weddell; CT, Corriente de Tasmania; p-CL, proto-Corriente de Leeuwin / Circumpolar Antarctic paleogeographic map during the Middle Eocene (40 Ma - Bartonian). Map derived from Plate Tectonic Reconstructions On-line Paleogeographic Mapper (Schettino and Scotese, 2005) [http://www.serg.unicam.it/Reconstructions. htm]. Gray areas indicate continental crust. Black lines indicate schematic oceanic circulation (modified after Sloan and Rea, 1996; Huber and Sloan, 2001). Abbreviations: p-GW, proto-Weddell Gyre; CT, Tasman Current; p-CL, proto-Leeuwin Current.
Los experimentos de Huber et al. (2004) fueron analizados fundamentalmente en el Pacífico Sur e Índico. Sin embargo, es posible considerar que el mecanismo propuesto ("cold trap") habría funcionado igualmente para el Atlántico Sur (Sloan y Rea, 1996; Huber y Sloan, 2001). En este caso habría existido un giro en sentido horario, equivalente a un Proto-Giro de Weddell (PGW). La intensificación del borde occidental de este giro habría causado la circulación de aguas relativamente frías sobre la plataforma sudamericana hacia latitudes medias del Atlántico Sudoccidental (figura 9) transportando especies de Enneadocysta y otros taxa considerados endémicos de altas latitudes hacia las cuencas del Colorado y Punta del Este.
Hacia el final del Eoceno se produjeron cambios en la circulación oceánica como resultado de la apertura del Conducto de Tasmania y el Pasaje de Drake. La reducción de Enneadocysta y de otros dinoflagelados endémicos durante la transición Eoceno-Oligoceno puede ser explicada por la disrupción local de los sistemas de giros descriptos por Huber et al. (2004). Las corrientes hacia el norte en los bordes occidentales habrían sido afectadas por la invasión de aguas cálidas a través de los pasajes restringiendo el "cold trap" como mecanismo inductor del endemismo. Stickley et al. (2004b) mostraron que el flujo de la Proto-Corriente de Leeuwin habría sido responsable de un aumento de temperatura en las aguas superficiales en el sector oriental de Tasmania a partir de la profundización del Conducto de Tasmania. Conco mitantemente, la apertura del Pasaje de Drake habría permitido el flujo de aguas más cálidas proveniente del Pacífico Sudoriental hacia el Atlántico interrum piendo el Proto-Giro de Weddell. El progresivo en friamiento e instalación de un sistema con hielo oceánico permanente habría causado la extinción de la típica asociación endémica de dinoflagelados en la mitad del Oligoceno. Esta asociación fue reemplaza da por dinoflagelados heterotróficos, incluyendo unos pocos nuevos taxa endémicos (Warnaar, 2006 Guerstein et al., 2010).
Conclusiones
El género Enneadocysta es un elemento caracterís tico de las asociaciones de quistes de dinoflagelado del Eoceno medio a tardío en el Atlántico Sudoc cidental. Está representado por las especies: E. dict yostila, E. arcuata, E. multicornuta, E. brevistila y Enneadocysta sp. A (de Brinkhuis et al., 2003b).
Las mayores frecuencias de Enneadocysta se regis tran en el Eoceno medio; es abundante en las asocia ciones de Tierra del Fuego y Santa Cruz, es común en las cuencas del Colorado y Punta del Este, y ha sido reportada en las cuencas del sudeste de Brasil en pro porciones menores. La abundancia de este grupo dis minuye considerablemente en las asociaciones de la transición Eoceno-Oligoceno.
El rango estratigráfico propuesto para las especie de Enneadocysta a partir de secciones calibradas en la plataforma de Tasmania por Brinkhuis et al., (2003b permite ajustar las edades de las asociaciones estu diadas en las cuencas del Atlántico Sudoccidental. En los sitios de perforaciones profundas en las cuenca de Colorado y Punta del Este se utiliza principal mente el dato de última presencia establecido en e Eoceno tardío (33,5 Ma). Asimismo, el horizonte de última presencia abundante registrado en la mitad del Priaboniano (35,7 Ma), constituye un importante evento bioestratigráfico correlacionable a través de los distintos sitios analizados en latitudes altas. En las cuencas ubicadas en latitudes medias y bajas se registra Enneadocysta de manera frecuente sólo en el Eoceno medio. Ferreira (2004) determinó que el máximo registrado dentro de la zona de nanoplancton calcáreo N447 corresponde aproximadamente a lo 43,5 Ma.
La distribución de Enneadocysta en el Atlántico Sudoccidental y en otros sitios del Océano Austra confirma a E. dictyostila y especies afines como miembros de la denominada "Flora Transantártica". Sobre la base de esta información se infiere que estos taxa habrían preferido aguas superficiales frías en ambientes costeros y de plataforma somera. Los datos isotópicos y palinológicos provenientes de la Cuenca de Sergipe en el sudeste de Brasil permiten corroborar esta interpretación.
La distribución de Enneadocysta en el Atlántico Sudoccidental resulta consistente con el modelo de paleocirculación de Huber et al. (2004) en el cual proponen la existencia de un sistema de circulación en el océano Austral, denominado "cold trap", gobernado por giros horarios en los océanos Pacífico Sur e Índico-Atlántico Sur. Este sistema de circulación permite explicar no sólo el endemismo de la denominada "Flora Transantártica" sino también la presencia de algunos componentes de dicha flora en latitudes medias y bajas del Atlántico Sudoccidental. Durante la transición Eoceno-Oligoceno la instalación de la Corriente Circumpolar Antártica habría debilitado los giros subpolares con la consecuente disminución en la representación de la flora endémica, la que se extinguió a mediados del Oligoceno.
Lista de nombres de géneros y especies citadas en el texto con referencias completas. Mayor información sobre taxonomía y nomenclatura en relación a estos taxa en Fensome y Williams (2004) y Fensome et al. (2008) / list of generic and specific names used in text with complete authorships. Further information on the taxonomy and nomenclature of these taxa can be found in Fensome and Williams (2004) and Fensome et al. (2008).
Areosphaeridium Eaton 1971
Areosphaeridium dictyostilum (Menéndez 1965) Sarjeant 1981
Areosphaeridium diktyoplokum (Klumpp 1953) Eaton 1971
Areosphaeridium multicornutum Eaton 1971
Cordosphaeridium diktyoplokum (Klumpp 1953) Eisenack 1963
Enneadocysta (Stover y Williams 1995) emend Fensome et al. 2006
Enneadocysta arcuata (Eaton 1971) Stover y Williams 1995
Enneadocysta brevistila Fensome et al. 2006
Enneadocysta dictyostila (Menéndez 1965) Stover y Williams
1995 emend Fensome et al. 2006
Enneadocysta multicornuta (Eaton 1971) Stover y Williams 1995
Enneadocysta partridgei Stover y Williams 1995
Enneadocysta pectiniformis (Gerlach 1961) Stover y Williams 1995
Enneadocysta sp. A de Brinkhuis et al. 2003b
Hystrichosphaeridium Deflandre 1937
Hystrichosphaeridium diktyoplokum Klumpp 1953
Hystrichosphaeridium dictyostilum Menéndez 1965
Agradecimientos
Los autores agradecen a O. Cárdenas y B. J. Crilley por el procesamiento de las muestras palinológicas y A. Henry por la asistencia técnica brindada en la utilización del MEB. Los comentarios de M. Prámparo y J. Helenes Escamilla en calidad de revisores contribuyeron significativamente a mejorar este trabajo. Este estudio fue realizado mediante proyectos financiados por la Agencia Nacional de Promoción Científica y Tecnológica (PICT 07-26057), Consejo Nacional de Investigaciones Científicas y Técnicas (PIP 6416) y la Secretaría de Ciencia y Tecnolología de la Universidad Nacional del Sur.
Bibliografía
1. Antunes, R.L., Shimabukuro, S., Oliveira, L.C.V., Zucatti da Rosa, A.L., Oliveira Costa, S., Cunha, A.S. y Oliveira Lima, F.H. 2004. Em busca da bioestratigrafia de alta resolução. Boletim Geociências da Petrobras 12: 421-427. [ Links ]
2. Archangelsky, S. 1969. Estudio del paleomicroplancton de la Formación Río Turbio (Eoceno), Provincia de Santa Cruz. Ameghiniana 3: 181-218. [ Links ]
3. Archangelsky, S., Guerstein, G.R. y Quattrocchio, M.E. 1997. [Pejerrey x-1 offshore well, Paynological Report. Shell Capsa, Buenos Aires, 19 pp. Inédito. [ Links ]].
4. Basov, I.A., Ciesielsky, P.F., Krasheninnikov, V.A., Weaver, F.M. y Wise, S.W. Jr. 1983. Biostratigraphic and Paleontologic Synthesis: Deep Sea Drilling Project Leg 71, Falkland Plateau and Argentine Basin. En: W.J. Ludwig, V.A. Krasheninnikov, et al., (eds.), Initial Reports of the Deep Sea Drilling Project 71: 445-460. [ Links ]
5. Berggren, W.A., Kent, D.V., Swisher, III, C.C., y Aubry, M.P. 1995. A revised Cenozoic geochronology and chronostratigraphy. En: W.A Berggren, D.V. Kent, M.-P. Aubry, and J. Hardenbol, (eds.), Geochronology, Time scales and Global Stratigraphic Correlation. SEPM Special Publication 54, p. 129-212. [ Links ]
6. Brinkhuis, H., Munsterman, D. M., Sengers, S., Sluijs, A., Wanaar, J. y Williams, G.L. 2003a. Late Eocene to Quaternary dinoflagellate cysts from ODP Site 1168, off western Tasmania, 189. 1-36. doi:10.2973/odp.proc.sr.189.105.2003 [ Links ]
7. Brinkhuis, H., Sengers, S., Sluijs, A., Warnaar, J. y Williams, G.L. 2003b. Latest Cretaceous to earliest Oligocene, and Quaternary dinoflagellate cysts from ODP Site 1172, East Tasman Plateau. En: N.F. Exon, J.P. Kennetty M. Malone (eds.), Proceedings of the Ocean Drilling Program, Scientific Results 189, pp.1 - 48. doi:10.2973/odp.proc.sr.189.105.2003 [ Links ]
8. Clowes, C.D. 1985. Stoveracysta, a new gonyaulacacean dinoflagellate genus from the upper Eocene and lower Oligocene of New Zealand. Palynology 9: 27-35. [ Links ]
9. Cocozza, C.D. y Clarke, C.M. 1992. Eocene microplankton from La Meseta Formation, northern Seymour Island. En: A.M. Duane, D. Pierrie, y J.B. Riding, (Eds.), Palynology of the James Ross Island area, Antarctic Peninsula. Antartic Science 4: 355-362. [ Links ]
10. Cookson, I.C. y Cranwell, L.M. 1967. Lower Tertiary microplankton, spores and pollen grains from southernmost Chile. Micropaleontology 13: 204-216. [ Links ]
11. Cookson, I.C. y Eisenack, A. 1967. Some early Tertiary microplankton and pollen grains from a deposit near Strahan, western Tasmania. Proceedings of the Royal Society of Victoria 80: 131-145. [ Links ]
12. Cranwell L.M., Harrington H.J. y Speden I.G. 1960. Lower Tertiary microfossils from McMurdo Sound, Antarctica. Nature 186: 700-702. [ Links ]
13. Cranwell, L.M. 1964. Hystrichospheres as an aid to Antarctic da-ting with special reference to the recovery of Cordosphaeridium in erratics at McMurdo Sound. Grana Palynologica 5: 397-405. [ Links ]
14. Daners, G. y Guerstein G.R. 2004. Quistes de dinoflagelados de las sedimentitas marinas del Cretácico tardío-Eoceno en la plataforma continental uruguaya. En: G. Veroslavsky, M. Ubilla y S. Martínez (eds.), Cuencas sedimentarias de Uruguay; Geología, Paleontología y Recursos Naturales, Cenozoico. Edición: División Relaciones y Actividades Culturales de Facultad de Ciencias, Montevideo. pp. 37-69. [ Links ]
15. Daners, G., Guerstein, G.R., Guler, M.V. y Veroslavsky, G. 2004. Las transgresiones del Maastrichtiense-Daniense y Eoceno medio en la cuenca Punta del Este y su correlación regional basada en dinoflagelados. 4º Congreso Uruguayo de Geología - II Reunión de Geología Ambiental y Ordenamiento Territorial del Mercosur (Montevideo), SUG, 9 páginas, Actas (Cd-Rom). [ Links ]
16. Eaton, G.L. 1971. A morphogenetic series of dinoflagellate cysts from the Bracklesham Beds of the Isle of Wight, Hampshire, England. En: A. Farinacci, (ed.), Proceedings of the Second Planktonic Conference, Rome, 1970, Roma, Edizioni Tecnoscienza, p. 355-379. [ Links ]
17. Exon, N.F., Kennett, J.P. y Malone, M.J. 2004. Leg 189 synthesis: Cretaceous-Holocene history of the Tasmanian Gateway. En: N.F. Exon, J.P. Kennett y M.J. Malone (eds.), Proceedings of the Ocean Drilling Program, Scientific Results, 189, 1: 1-37 doi:10.2973/odp.proc.sr.189.101.2004 [ Links ]
18. Fasola, A. 1969. Estudio palinológico de la Formación Loreto (Terciario Medio), provincia de Magallanes, Chile. Ameghiniana 6: 3-49. [ Links ]
19. Fensome, R.A. y Willliams, G.L. 2004. The Lentin and Williams index of Fossil Dinoflagellates 2004 edition. American Association of Stratigraphic Palynologists Contributions Series 42, 909 pp. [ Links ]
20. Fensome, R.A., Guerstein, G.R. y Williams, G.L. 2006. The Paleogene dinoflagellate cyst genera Enneadocysta and Licracysta gen. nov.: new insights based on material from offs-hore eastern Canada and southern Argentina. Micro-paleontology 52: 385-410. [ Links ]
21. Fensome, R.A., MacRae, R.A. y Williams, G.L. 2008. DINOFLAJ2, Version 1. American Association of Stratigraphic Palynologists, Data Series no.1. http://dinoflaj.smu.ca/wiki/Main_Page [ Links ]
22. Fensome, R.A., Taylor, F.J.R., Norris, G., Sarjeant, W.A.S., Wharton, D.I. y Williams, G.L. 1993. A classification of fossil and living dinoflagellates. Micropaleontology Press Special Publication 7: 1-351. [ Links ]
23. Ferreira, E.P. 2004. [Palinoestratigrafia e caracterização de paleoambientes da seção paleocênica-eocênica da Bacia de Sergipe. Instituto de Geociências - UFRJ. Tese Doutoral . 253pp., 63 figs. Inédito. [ Links ]].
24. Franke, D., Neben, S., Schreckenberger, B., Schulze, A., Stiller, M. y Kravczyk, C. 2006. Crustal structure across the Colorado Basin, offshore Argentina. Geophysical Journal International 165: 850-864. doi:10.1111/j.1365-246X.2006.02907.x. [ Links ]
25. Franke, D., Neben, S., Ladage, S., Schreckenberger, B. y Hinz, K. 2007. Margin segmentation and volcano-tectonic architecture along the volcanic margin off Argentina/Uruguay, South Atlantic. Marine Geology 244: 46-67. doi:10.1016/j.mar-geo.2007.06.009. [ Links ]
26. Fryklund, R., Marshall, A. y Steves, J. 1996. La Cuenca del Colorado. En: V.A. Ramos y M.A. Turic (eds.), Geología y Recursos Naturales de la Plataforma Continental Argentina. 13° Congreso Geológico Argentino y 3° Congreso de Exploración de Hidrocarburos (Buenos Aires), Relatorio 8: 135-158. [ Links ]
27. Gamerro, J.C. y Archangelsky, S. 1981. Palinozonas Neocretácicas y Terciarias de la plataforma continental Argentina en la Cuenca del Colorado. Revista Española de Micropaleontología 13: 119-140. [ Links ]
28. Ghidella, M.E., Schreckenberger, B., Paterlini, C.M. y Abraham, D.A. 2006. Anomalías magnéticas en el margen argentino. Reunión de Tectónica (San Luis), Actas en CD-ROM. [ Links ]
29. Goodman, D.K. y Ford, L.N. Jr. 1983. Preliminary dinoflagellate biostratigraphy for the middle Eocene to lower Oligocene from the southwest Atlantic Ocean. En: W.J. Ludwig, V.A. Krasheninnikov, et al. (eds.). Initial Reports of the Deep Sea Drilling Project 17: 859-877. [ Links ]
30. Gradstein, F.M., Ogg, J.G. y Smith, A.G. 2004. A Geologic Time scale 2004. Cambridge University Press, Cambridge, 589 pp. [ Links ]
31. Guerstein, G.R y Junciel, G.L. 2001. Quistes de dinoflagelados del Cenozoico de la Cuenca del Colorado, Argentina. Ameghiniana 38: 299-316. [ Links ]
32. Guerstein, G.R., Chiesa, J.O., Guler, M.V. y Camacho, H.H. 2002. Bioestratigrafía basada en quistes de dinoflagelados de la Formación Cabo Peña (Eoceno terminal - Oligoceno temprano), Tierra del Fuego, Argentina. Revista Española de Micropaleontologia 34: 105-115. [ Links ]
33. Guerstein, G.R., Guler, M.V., Daners, G. y Archangelsky, S. 2003. Quistes de dinoflagelados del Eoceno medio del Atlántico Sudoccidental y su correlación con otros sitios del Hemisferio Sur. Ameghiniana 40: 87R. [ Links ]
34. Guerstein, G.R., Junciel, G.L., Guler, M.V. y Daners, G. 2005. Diconodinium lurense sp. nov., a late Maastrichtian to Danian dinoflagellate cyst from Southwest Atlantic basins. Ameghiniana 42: 329-338. [ Links ]
35. Guerstein, G.R., Guler, M.V., Williams, G.L., Fensome, R.A. y Chiesa, J.O. 2008. Mid Palaeogene dinoflagellate cysts from Tierra del Fuego, Argentina: biostratigraphy and palaeo environments. Journal of Micropalaeontology 27: 75-94. [ Links ]
36. Guerstein, G.R., Guler, M.V., Brinkhuis, H. y Warnaar, J. 2010. Mid Cenozoic Palaeoclimatic and Palaeoceanographic trends in the Southwest Atlantic Basins, a Dinoflagellate view. The Paleontology of Gran Barranca. En: R.H. Madden, A.A. Carlini, M.G. Vucetich y R.F. Kay (eds.), The Paleontology of Gran Barranca: Evolution and Environmental Change through the Middle Cenozoic of Patagonia, p. 398-409, Cambridge, Cambridge University Press. [ Links ]
37. Guler, M.V. y Guerstein, G.R. 2002. Bioestratigrafía del Oligoceno-Plioceno temprano de la Cuenca del Colorado (Argentina), basada en quistes de dinoflagelados. Revista Española de Micropaleontología 34: 359-371. [ Links ]
38. Hannah, M.J. 1997. Climate controlled dinoflagellate distribution in Late Eocene-Earliest Oligocene strata from CIROS-1 drill-hole, McMurdo Sound, Antarctica. Terra Antartica 4: 73-78. [ Links ]
39. Hannah, M.H. 2006. The palynology of ODP Site 1165, Prydz Bay, East Antarctica: A record of Miocene glacial advance and retreat. Palaeogeography, Palaeoclimatology, Palaeoecology 231: 120-133. [ Links ]
40. Hannah, M.J., Wrenn, J.H. y Wilson, G.L. 1998. Early Miocene and Quaternary marine palynomorphs from Cape Roberts Project CRP-1, McMurdo Sound, Antarctica. Terra Antarctica 5: 527-538. [ Links ]
41. Haskell, T.R. y Wilson, G.J. 1975. Palynology of sites 280-284, Deep Sea Drilling Project Leg 29, off southeastern Australia and western New Zealand. En: J.P. Kennett et al. (eds.). Initial Reports of the Deep Sea Drilling Project 29: 723-741. [ Links ]
41. Hinz, K., Neben, S., Schreckenberger, B., Roeser, H.A., Block, M., Gonzalvez de Souza, K. y Meyer, H. 1999. The Argentine continental margin north of 48°S: sedimentary successions, volcanic activity during breakup. Marine and Petroleum Geology 16: 1-25. [ Links ]
42. Huber, M. y Sloan, L.C. 2001. Heat transport, deep waters, and thermal gradients: Coupled simulation of an Eocene ''Greenhouse'' climate. Geophysical Research Letters 28: 3481-3484. [ Links ]
43. Huber, M., Brinkhuis, H., Stickley, C.E., Döös, K., Sluijs, A., Warnaar, J., Schellenberg, S.A. y Williams, G.L. 2004. Eocene circulation of the Southern Ocean: Was Antarctica kept warm by subtropical waters? Paleoceanography 19: PA4026, doi: 10.1029/2004PA001014. [ Links ]
44. Juan, R., de Jager, J., Russell, J. y Gehard, I. 1996. Flanco norte de la Cuenca del Colorado. En: V.A. Ramos y M.A. Turic (eds.), Geología y Recursos Naturales de la Plataforma Continental Argentina. 13° Congreso Geológico Argentino y 3° Congreso de Exploración de Hidrocarburos (Buenos Aires), Relatorio 7: 117-133. [ Links ]
45. Kemp, E.M. 1975. Palynology of Leg 28 drill sites, Deep Sea Drilling Project. En: D.E. Hayes y L.A. Frakes (eds.), Initial Reports of the Deep Sea Drilling Project, 28: 599-623. [ Links ]
46. Kennett, J.P. 1977. Cenozoic evolution of Antarctic glaciation, the Circum-Antarctic Ocean, and their impact on global paleoceanography. Journal of Geophysical Research 82: 3843-3860. [ Links ]
47. Kennett, J.P., Houtz, R.E., Andrews, P.B. y Edwards, A.R. 1975. Development of the circum-Antarctic current. Science 186: 144-147. [ Links ]
48. Lentin, J.K. y Williams, G.L. 1985. Fossil dinoflagellates: index to genera and species 1985 edition. Canadian Technical Report of Hydrography and Ocean Sciences 60, 449 pp. [ Links ]
49. Levy, R.H. y Hardwood, D.M. 2000. Tertiary marine palynomorphs from the McMurdo Sound erratics, Antarctica. En: J.D. Stilwell y R.M. Feldmann (eds.) Paleobiology and paleoenvironments of Eocene rocks, McMurdo Sound, East Antarctica. Antarctic Research Series 76: 183-242. [ Links ]
50. MacPhail, M.K. y Truswell, E.M. 2004. Palynology of Site 1166, Prydz Bay, East Antarctica. En: A.K. Cooper, P.E. O'Brien, y C. Richter (eds.), Proceedings of the Ocean Drilling Program, Scientific Results 188: 1-43. [ Links ]
51. Malumián, N. 1999. La sedimentación y el volcanismo terciarios en la Patagonia Extraandina. En: R. Caminos (ed.), Geología Argentina, Instituto de Geología y Recursos Minerales, Anales 29: 557-612. [ Links ]
52. Malumián, N. y Olivero, E.B. 2006. El Grupo Cabo Domingo, Tierra del Fuego: Bioestratigrafía, Paleoambientes y acontecimienos del Eoceno-Mioceno marino. Revista de la Asociación Geológica Argentina 61: 139-160. [ Links ]
53. Malumián, N., Olivero, E.B. y Concheyro, A. 1994. Eocene microfossils from the Leticia Formation, Tierra del Fuego Island, Argentina. Ameghiniana 31: 398R. [ Links ]
54. Mao, S. y Mohr, B.A.R. 1995. Middle Eocene dinocysts from Bruce Bank (Scotia Sea, Antarctica) and their paleoenvironmental and paleogeographic implications. Review of Palaeobotany and Palynology 86: 235-263. [ Links ]
55. Marshall, J. y Plumb. R.A. 2008. Atmosphere, Ocean and Climate Dynamics: An Introductory Text. Academic Press, Londres, 344 pp. [ Links ]
56. Martin, H.A. 1991. Dinoflagellate and spore pollen biostratigraphy of SADME MC63 bore, western Murray Basin. Alcheringa 15: 107-144. [ Links ]
57. Martin, H.A. 1993. Middle Tertiary dinoflagellate and spore/pollen biostratigraphy and palaeoecology of the Mallee Cliffs Bore, central Murray Basin. Alcheringa 17: 91-124. [ Links ]
58. Menéndez, C.A. 1965. Microplancton fósil de sedimentos Terciarios y Cretácicos del norte de Tierra del Fuego (Argentina). Ameghiniana 4: 7-18. [ Links ]
59. Miller, K.G., Kominz, M.A., Browning, J.V., Wright, J.D., Mountain, G.S., Katz, M.E., Sugarman, P.J., Cramer, B.S. Christie-Blick, N. y Pekar, S.F. 2005. The Phanerozoic record of global sea-level change. Science 310: 1293-1298. [ Links ]
60. Mohr, B.A.R. 1990. Eocene and Oligocene Sporomorphs and dinoflagellate cysts from leg 113 drill sites, Weddell Sea, Antarctica. En: P.F. Barker y J.P. Kennet (eds.), Proceedings of the Ocean Drilling Program, Scientific Results 113: 595-611 U.S. Government Printing Office, College Station, Texas. [ Links ]
61. Olivero, E.B. y Malumián, N. 2008. Mesozoic-Cenozoic stratigraphy of the Fueguian Andes, Argentina. Geologica Acta 6: 5-18. [ Links ]
62. Pöthe de Baldis, E.D. 1966. Microplancton del Terciario de Tierra del Fuego. Ameghiniana 4: 219-228. [ Links ]
63. Pross, J. y Brinkhuis, H. 2005. Organic-walled dinoflagellate cysts as paleoenvironmental indicators in the Paleogene; a synopsis of concepts. Paläontologische Zeitschrift 79: 53-59. [ Links ]
64. Quattrocchio, M. y Guerstein, G.R. 1994. [Palynological analysis of Cruz del Sur -1 Borehole. Colorado Basin, Argentina. Informe técnico presentado a Union Texas Petroleum Co. 25 pp. Inédito. [ Links ]].
65. Quattrocchio, M.E. y Sarjeant, W.A.S. 1996. Early Paleocene (Danian) dinoflagellates from the Colorado Basin, Argentina. Revista Española de Micropaleontología 28: 111-138. [ Links ]
66. Regali, M.S.P., Uesugui, N. y Santos, A.D.S. 1974a. Palinologia dos sedimentos Meso-Cenozoicos do Brazil (I). Boletim Tecnico da Petrobras 17: 177-190 [ Links ]
67. Regali, M.S.P., Uesugui, N. y Santos, A.D.S. 1974b. Palinologia dos sedimentos Meso-Cenozoicos do Brazil (II). Boletim Tecnico da Petrobras 17: 263-301 [ Links ]
68. Regali, M.S.P. 1980. Palinoestratigrafia da Bacia do Ceará. Congresso Brasileiro de Geologia, Camboriú, Anais 5: 3118-3129. [ Links ]
69. Röhl, U., Brinkhuis, H., Stickley, C.E., Fuller, M., Schellenberg, S.A., Wefer, G. y Williams, G.L. 2004. Sea level and astronomically induced environmental changes in middle and late Eocene sediments from the East Tasman Plateau. En: N.F. Exon, M. Malone, y J.P. Kennett (Eds.), Climate evolution of the Southern Ocean and Australia's northward flight from Antarctica. American Geophysical Union, Geophysical Monograph Series 151: 127-151. [ Links ]
70. Sarjeant, W.A.S. 1981. A restudy of some dinoflagellate cyst holotypes in the University of Kiel Collections II. The Eocene holotypes of Barbara Klumpp (1953); with a revision of the genus Cordosphaeridium Eisenack, 1963. Meyniana 33: 97-132. [ Links ]
71. Schettino, A. y Scotese, C.R. 2005. Apparent polar wander paths for the major continents (200 Ma-Present Day): A paleomagnetic reference frame for global plate tectonic reconstructions. Geophysical Journal International 163: 727-759. [ Links ]
72. Siedler, G., Gould, W.J. y Church, J. 2001. Ocean Circulation and Climate: Observing and Modelling the Global Ocean. International Geophysics Series 77, Academic Press, Londres, 715 pp. [ Links ]
73. Sloan, L.C. y Rea, D.K. 1996. Atmospheric carbon dioxide and Early Eocene climate: A general circulation modeling sensitivity study. Palaeogeography, Palaeoclimatology, Palaeoecology 119: 275-292. [ Links ]
74. Sluijs, A., Brinkhuis, H., Stickley, C.E., Warnaar, J., Williams, G.L. y Fuller, M. 2003. Dinoflagellate cysts from the Eocene / Oligocene transition in the Southern Ocean; results from ODP Leg 189. En: N.F. Exon, J.P. Kennett y M.J. alone, (eds.), Proceedings of the Ocean Drilling Program, Scientific Results 189: 1-42 doi:10.2973/odp.proc.sr.189.104.2003. [ Links ]
75. Sluijs, A., Pross, J. y Brinkhuis, H. 2005. From greenhouse to ice-house; organicwalled dinoflagellate cysts as paleoenviron-mental indicators in the Paleogene. Earth Science Reviews 68 281-315. [ Links ]
76. Smayda, 2002. Turbulence, watermass stratification and harmfu algal blooms: an alternative view and frontal zones as 'pelagic seed banks'. Harmful Algae 1: 92-112. [ Links ]
77. Stickley, C.E., Brinkhuis, H., McGonigal, K.L., Chaproniere G.C.H., Fuller, M., Kelly, D.C., Nürnberg, D., Pfuhl, H.A. Schellenberg, S.A., Schoenfeld, J., Suzuki, N., Touchard, Y. Wei, W., Williams, G.L., Lara, J., y Stant, S.A. 2004a. Late Cretaceous-Quaternary biomagnetostratigraphy of ODP Sites 1168, 1170, 1171, and 1172, Tasmanian Gateway En: N.F. Exon J.P. Kennett, y M.J. Malone (eds.), Proceedings of the Ocean Drilling Program, Scientific Results, 189: 1-57 doi 10.2973/odp.proc.sr.189.111.2004. [ Links ]
78. Stickley, C.E., Brinkhuis, H., Schellenberg, S.A., Sluijs, A., Röhl U., Fuller, M., Grauert, M., Huber, M., Warnaar, J. y Williams G.L. 2004b. Timing and nature of the deepening of the Tasmanian Gateway. Paleoceanography 19, PA4027, doi:10.1029/2004PA001022. [ Links ]
79. Stoakes, F.A. Campbell, C.V. Cass, R. y Ucha, N. 1991. Seismic stratigraphic analysis of the Punta del Este Basin, offshore Uruguay, South America. American Association of Petroleum Geologists Bulletin 75: 219-240. [ Links ]
80. Stover, L.E. 1975. Observations on some Australian Eocene dinoflagellates. Geoscience and Man 11: 35-45. [ Links ]
81. Stover, L.E. y Williams, G.L. 1995. A revision of the Paleogene dinoflagellate genera Areosphaeridium Eaton 1971 and Eatonicysta Stover and Evitt 1978. Micropaleontology 41: 97-141 [ Links ]
82. Truswell, E.M. 1997. Palynomorph assemblages from marine Eocene sediments on the West Tasmanian continental margin and the South Tasman Rise. Australian Journal of Earth Sciences 4: 633-654. [ Links ]
83. Ucha, N., de Santa Ana, H. y Veroslavsky, G. 2003. La cuenca Punta del Este: geología y potencial hidrocarburífero. En: G Veroslavsky, M. Ubilla, y S. Martínez (eds.), Cuencas Sedimentarias de Uruguay: Geología, Paleontología y Recursos Naturales, Mesozoico. División Relaciones y Actividades Culturales de Facultad de Ciencias, Montevideo, pp. 171-190. [ Links ]
84. Uchupi, E. y Emery, K. 1991. Pangaean divergent margins: historical perspective. Marine Geology 102: 1-28. [ Links ]
85. Urien, C.M. y Zambrano, J.J. 1996. Estructura del margen continental. En: V. Ramos y M. Turic (eds.), Geología y recursos naturales de la plataforma continental Argentina. Asociación Geológica Argentina, Buenos Aires, Argentina, pp. 29-65. [ Links ]
86. Warnaar, J. 2006. [Climatological implications of Australian -Antarctic separation. Universidad de Utrecht, Utrecht. PhD Tesis .143 pp. Inédito. [ Links ]].
87. Williams, G.L., Brinkhuis, H., Pearce, M.A., Fensome, R.A. y Weegink, J.W. 2004. Southern Ocean and global dinoflagellate cyst events compared: Index events for the late Cretaceous-Neogene. En: N. Exon y J.P. Kennett (eds.), Proceedings of the Ocean Drilling Program, Scientific Results 189: 1-98. [ Links ]
88. Wilson, G.J. 1967. Some new species of lower Tertiary dinoflagellates from McMurdo Sound, Antarctica. New Zealand Journa of Botany 5: 57-83. [ Links ]
89. Wilson, G.J. 1984. New Zealand late Jurassic to Eocene dinoflagellate biostratigraphy -a summary. Newsletters in Stratigraphy 13: 104-117. [ Links ]
90. Wilson, G.J. 1985. Dinoflagellate biostratigraphy of the Eocene Hampden Section North Otago, New Zealand. New Zealand Geological Survey Record 8: 93-101. [ Links ]
91. Wilson, G.J. 1988. Paleocene and Eocene dinoflagellate cysts from Waipawa, Hawkes Bay, New Zealand. New Zealand Geological Survey Paleontological Bulletin 57: 1-96. [ Links ]
92. Wilson, G.S., Roberts, A.P., Verosub, K.L., Florindo, F. y Sagnotti, L. 1998. Magnetobiostratigraphic chronology of the Eocene-Oligocene transition in the CIROS-1 core, Victoria Land margin, Antarctica: Implications for Antarctic glacial history. Geological Society of America Bulletin 110: 35-47. [ Links ]
93. Wrenn, J.H. y Beckman, S.W. 1982. Maceral, total organic carbon, and palynological analyses of Ross Ice Shelf Project site J9 cores. Science 216: 187-189. [ Links ]
94. Wrenn, J.H. y Hart, G.F. 1988. Paleogene dinoflagellate cyst biostratigraphy of Seymour Island, Antarctica. En: R.M. Feldmann y M.O. Woodburne (eds.), Geology and paleontology of Seymour Island, Antarctic Peninsula. Geological Society of America Memoir 169: 321-447. [ Links ]
Recibido: 2 de agosto de 2009.
Aceptado: 5 de mayo de 2010.

