










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAmeghiniana
versão On-line ISSN 1851-8044
Ameghiniana vol.47 no.4 Buenos Aires oct./dez. 2010
ARTÍCULOS ORIGINALES
Conodontes sandbianos (Ordovícico Superior) en la Quebrada La Pola, Sierra de Villicum, precordillera de San Juan (Argentina)
Susana Heredia¹ y Juan Pablo Milana²
¹CONICET - Universidad Nacional de San Juan, IIM, Av. Libertador San Martín y Urquiza, 5400 San Juan, sheredia@unsj.edu.ar
²CONICET - Universidad Nacional de San Juan, INGEO, Av. De La Roza y Meglioli, 5400 San Juan, milana@unsj-cuim.edu.ar
Resumen. Se da a conocer nueva información bioestratigráfica y paleoambiental de la Formación La Pola, unidad siliciclástica con niveles mixtos de areniscas y lutitas que incluyen braquiópodos, que aparece en contacto con el techo de la Formación La Cantera en la Quebrada de La Pola (Sierra de Villicum). De los niveles bioclásticos superiores se ha rescatado una conodontofauna que se asigna a la Biozona de Amorphognathus tvaerensis, característica del Ordovícico Superior temprano (Sandbiano bajo). La determinación de Baltoniodus variabilis Bergström permite definir la subzona homónima. Las afinidades paleobiogeográficas de la conodontofauna corresponden claramente a la provincia del Midcontinent, aunque también se observan influencias noratlánticas, mientras que las características lito y biofaciales indican una plataforma siliciclástica media a media-somera.
Abstract. Sandbian (Late Ordovician) Conodonts at Quebrada La Pola, Sierra de Villicum, San Juan Precordillera (Argentina). In this contribution we better define the age and discuss the stratigraphic significance of a Late Ordovician brownish siliciclastic unit at the top of the La Cantera Formation at Quebrada La Pola (Sierra de Villicum), in the light of new biostratigraphic evidences here provided. The La Pola Formation includes siltstones and interbedded sandstones with brachiopod shells deposited in a proximal to intermediate shelf environment. A conodont association contained in this unit allows defining the Amorphognathus tvaerensis Zone, in the lower Late Ordovician. Within this zone, the unit can be assigned to the Baltoniodus variabilis Subzone, defining quite accurately the time of deposition. The conodont association suggests some paleobiogeographical affinities with the Midcontinent Province but some North Atlantic Province influence is also possible.
Palabras clave. Conodontes; Bioestratigrafía; Ordovícico Superior; San Juan; Precordillera; Argentina.
Key words. Conodonts; Biostratigraphy; Late Ordovician; San Juan; Precordillera; Argentina.
Al noreste de la Sierra de Villicum (provincia de San Juan), en el ámbito de la Precordillera Oriental (Ortiz y Zambrano, 1981), aparecen varias unidades litoestratigráficas clásticas del Paleozoico Inferior cuyo rango temporal abarca el Ordovícico-Devónico (figura 1). En Quebrada La Pola (figura 2), entre las formaciones La Cantera (zonas de Hustedograptus teretiusculus y Nemagraptus gracilis) (Peralta, 1993) y Don Braulio (Zona de Normalograptus persculptus - Fauna de Hirnantia) (Benedetto, 1986; Peralta y Baldis, 1990; Peralta y Carter, 1990, 1999), afloran depósitos siliciclásticos de color pardo claro con abundante macrofauna fósil. Estos depósitos fueron definidos por Astini (2001) como Formación La Pola, en base a consideraciones sedimentológicas. En particular ambos contactos con las unidades infra y suprayacentes fueron interpretados por Astini (2001) como fuertemente erosivos, lo que supone un hiato en su base y otro en su contacto con la unidad suprayacente. Astini (2001) interpretó esta unidad como un conjunto de depósitos de flujo de detritos, donde el contenido paleontológico exhibe claras evidencias de transporte, excepto en algunos niveles del tope que contienen braquiópodos, briozoos, bivalvos, gastrópodos y fragmentos de trilobites autóctonos (Benedetto, 2003). La asociación de estos braquiópodos indicaría una edad caradociana temprana (Benedetto, 2003). Previamente, Brussa (2000) identificó una asociación de graptolitos que permite reconocer el lapso comprendido entre las zonas de Nemagraptus gracilis y Climacograptus bicornis, y que según este autor podría extenderse hasta la Zona de Dicranograptus clingani. Debe mencionarse también el aporte de Sánchez et al. (2003), con el estudio de una fauna de bivalvos que aparece en clastos provenientes de esta unidad e incluidos en el conglomerado del Miembro Inferior de la Formación Don Braulio.
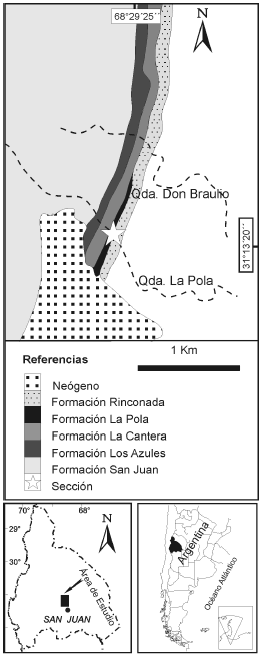
Figura 1. Mapa de ubicación (modificado de Benedetto, 2003) / location map (modified from Benedetto, 2003).
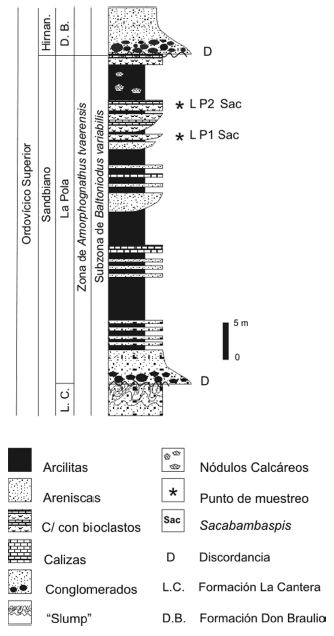
Figura 2. Sección estratigráfica con los niveles muestreados / stratigraphic section and sampled levels.
Las observaciones de campo ponen en evidencia que la Formación La Pola tiene una escasa distribución vertical y longitudinal. Si bien Astini (2001, 2003) la describió como con algo más de 1 km de extensión y con 48 metros máximos de espesor, el mapeo de esta unidad es ciertamente arduo de representar pues las dimensiones medidas sobre la localidad tipo resultan sustancialmente menores. La existencia de superficies erosivas es notablemente común, y su base suele ser una difícil elección entre diferentes superficies erosivas.
Para este estudio se realizaron observaciones en el sector de referencia localizado en S 31º13'20.9" y O 68º29'25.7". Se levantó una sección de la que se colectaron muestras para la obtención de conodontes, atendiendo al contenido macrofosilífero autóctono observado en los tramos medios y altos del perfil, según fue descrito por Benedetto (2003) (figura 2). El objetivo de esta contribución es dar a conocer los conodontes identificados en estos depósitos, los que brindan precisiones sobre la edad de la Formación La Pola, comparar la conodontofauna con otras de similar edad de la Precordillera, así como considerar los aspectos relacionados con la biofacies y el provincialismo de conodontes.
Estratigrafía
En la Quebrada de La Pola afloran las formaciones San Juan, Gualcamayo, La Cantera y Don Braulio; solo recientemente se ha adicionado la Formación La Pola (Astini, 2001). La Formación La Cantera (147 m de espesor) suprayace mediante discordancia a la Formación Gualcamayo e infrayace a la Formación La Pola. Peralta (1993) reconoció en la Formación La Cantera tres miembros: inferior, medio y superior, reflejando tres sucesiones grano- y estrato- decrecientes. El miembro inferior está definido por conglomerados y areniscas conglomerádicas, y pelitas verdes intercaladas; el miembro medio es transicional con respecto al miembro inferior y está conformado por pelitas y areniscas con gradación normal, estructuras sedimentarias (marcas de corriente y tool-marks) y escasas trazas fósiles pertenecientes a la icnofacies de Cruziana (sensu Peralta, 1993). Finalmente, el miembro superior está definido por una sucesión grano y estrato decreciente, y por una deformación sindepositacional intensa en su techo, la cual es de gran importancia para nuestra interpretación de la unidad ubicada al tope de la Formación La Cantera.
Por las características litológicas y por la distribución de sus afloramientos, los depósitos de la Formación La Pola representan un nuevo evento granoy estrato-decreciente. De la Formación La Cantera a la Formación La Pola se reconoce un cambio en el color que varía de verdoso a amarillento y gris amarillento. Este cambio de color de los depósitos como característica diagnóstica en el mapeo de las diferentes unidades, es el aspecto que se debe considerar como indicador para identificar estos niveles con un nombre formacional propio en el sentido de Astini (2001). Para estos depósitos se han medido unos 35 m de espesor máximo y una longitud máxima en afloramiento de 860 m (figura 2).
Las observaciones de campo efectuadas sugieren que el techo de la unidad infrayacente (miembro superior de la Formación La Cantera) es un conjunto de estratos que debido a un proceso de slumping (deslizamiento lento coherente) han sido moderadamente deformados a un nivel diferente al de los depósitos infra- y suprayacentes (figura 3.1). La deformación observada es coherente con el ambiente de depositación sugerido por la mayoría de los autores (un talud de pendiente moderada con frecuentes flujos gravitatorios), dada la conspicua presencia en este intervalo de turbiditas de grano grueso y de flujos de detrito entremezclados (figura 3.2). Por otra parte, la interpretación de este intervalo sedimentario como una unidad leve a moderadamente deformada por deslizamiento subácueo (slumping o submarine creep), explicaría la mezcla de deformaciones (principalmente pliegues) de pequeño y mediano radio de curvatura (decímetros y decámetros, figura 3.1), las que no podrían ser explicadas estructuralmente. A esto debe sumarse el hecho de que la vergencia de deformación parece no coincidir con la de las principales estructuras tectónicas del área.
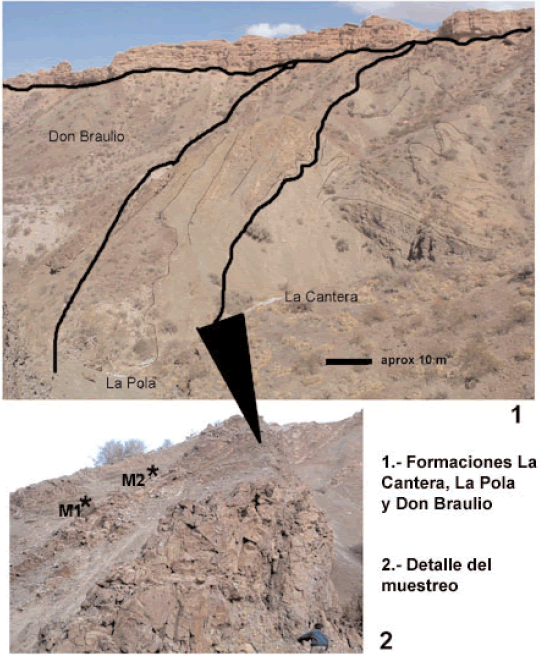
Figura 3. 1-2, afloramientos sandbianos de la Quebrada La Pola y su relación con las Formaciones La Cantera y Don Braulio, infra- y suprayacentes respectivamente. / Sandbian outcrop at Quebrada La Pola and its relation with overlying La Cantera Formation and the underlying Don Braulio Formation.
Es sobre estos tramos deformados que se produjo el depósito de la Formación La Pola, probablemente rellenando las áreas deprimidas dejadas por el proceso de deformación que afectó a los términos superiores de la Formación La Cantera. Esta interpretación permite explicar por qué las areniscas pardas de la Formación La Pola exhiben una muy limitada distribución areal y reducido espesor (figura 4). El lapso entre ambos eventos de depositación podría ser muy variable, pero en este caso, y de acuerdo con las precisiones bioestratigráficas que aportan los conodontes,éste lapso parece haber sido prácticamente nulo (figura 6). No se han encontrado evidencias paleontológicas que permitan argumentar la existencia de un hiato significativo entre los depósitos deformados del techo de la Formación La Cantera y los depósitos de la Formación La Pola.

Figura 4. Contacto irregular de la Formación la Pola con el techo Formación La Cantera / irregular contact of the La Pola Formation and the top of the La Cantera Formation.
Materiales y métodos
Se colectaron muestras en los niveles de areniscas carbonáticas (LP1 y LP2; figura 2) con el fin de obtener conodontes de la Formación La Pola, en la quebrada homónima. Tanto la muestra LP1 como la LP2 pueden ser caracterizadas como areniscas medianas a finas con cemento carbonático (especialmente LP1) por el contenido casi exclusivo de granos de cuarzo subredondeado. Las muestras fueron procesadas en laboratorio mediante los métodos convencionales con ácido acético (Jeppsson et al., 1985; Stone, 1987). De los residuos insolubles obtenidos se rescataron conodontes, placas de peces acorazados, briozoarios, mudas de trilobites, braquiópodos, artejos de crinoideos y gastrópodos.
El color que exhiben los conodontes recuperados, corresponde a un valor de 4 del CAI (Color Alteration Index de Epstein et al., 1977). La conservación de los conodontes es muy buena, pero la mayoría de los ejemplares más robustos aparecen rotos e incompletos, especialmente los que poseen en su morfología varios procesos. Sin embargo, las superficies se observan límpidas y sin evidencias de abrasión. Las observaciones realizadas sobre las superficies de las placas de peces acorazados que aparecen junto con los conodontes revelan que en la muestra LP1 se observa abrasión superficial intensa y desgaste de la morfología. Por el contrario, en la muestra LP2 las placas de pequeño tamaño, supuestamente ejemplares juveniles, aparecen sin evidencias de abrasión (figura 5.17), en tanto que en aquéllas de mayor tamaño se reconocen los efectos de una abrasión moderada.

Figura 5. 1-17, microfotografías (MEB) de conodontes del Sandbiano (Ordovícico Superior) de la Formación La Pola (LP1; LP2), Precordillera de San Juan / SEM microphotographs of Sandbian (Late Ordovician) conodonts from the La Pola Formation, (LP1; LP2), Precordillera of San Juan. 1, 6-7, Amorphognathus tvaerensis Bergström P y S, 1, vista lateral de elemento P / lateral view of element P; 6-7, vista lateral de elemento S / lateral view of element S ( LP2, CML-C 3012/1; 3;4). 2-4, Baltoniodus variabilis Bergström; 2, vista superior del proceso posterior de la forma Pb / upper view of posterior process of Pb element; 3-4, vistas laterales de los elementos S / lateral views of S elements (LP2, CML-C 3018/ 3; 4; 9); 5, 8, Eoplacognathus elongatus Bergström; 5, vista oral del elemento Pb / oral view of element Pb; 8, vista oral del fragmento de proceso lateral que podría pertenecer a una forma Pa / oral view of lateral process that could be a Pa element (LP2, CML-C 3015 / 1; 6); 9-10, Aphelognathus cf. politus (Hinde), vistas laterales de elementos Pa / lateral views of Pa elements (LP2, CML-C 3014/1; 4); 11-12, Erismodus typus Branson y Mehl, vista posterior de elementos S / posterior view of S elements (LP 2, CML-C 3016 / 2; 3); 13-14, Plectodina sp., vistas laterales de elementos S / lateral views of S elements (LP2, CML-C 3018/ 1; 5); 15, Oneotodus? ovatus (Stauffer). Vista postero-lateral / posterior-lateral view (LP1, CML-C 3011/1); 16, Protopanderodus varicostatus (Sweet y Bergström), vista lateral / lateral view (LP2, CML-C 3017/1); 17, Sacabambaspis janvieri Gagnier, vista superior de placa dérmica / upper view of dermic plate (LP 2, CMLC 3020/1). Escala gráfica / scale bar: 0,1 mm.
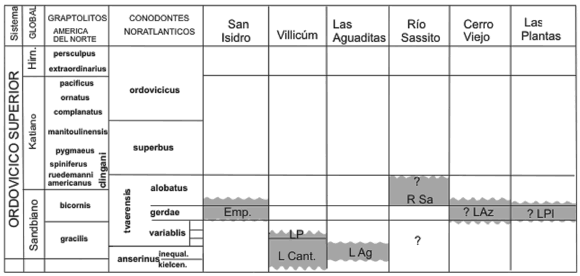
Figura 6. Cuadro de correlación de las unidades formales del Ordovícico Superior de la Precordillera / correlation chart of the Late Ordovician formations of the Precordillera. Acrónimos/acronyms: EMP: Empozada; LP: La Pola; L.Cant.: La Cantera; L.Ag.: Las Aguaditas; R Sa.: Río Sassito; LAz: Los Azules; LPl.: Las Plantas.
Todos los materiales de la Figura 5 corresponden a microfotografías tomadas por microscopia electrónica de barrido en el Laboratorio de Microscopía Electrónica del NOA (LAMENOA - Tucumán), los cuales fueron posteriormente depositados en la colección de microvertebrados Lillo-Conodontes de la Facultad de Ciencias Naturales e Instituto Miguel Lillo (CML-C), bajo los números de repositorio 3010-3062.
Consideraciones sobre la conodontofauna
Hasta el presente se han recuperado unos 50 elementos de conodontes entre ejemplares determinables y fragmentos, entre los cuales se han identificado 9 especies: Amorphognathus tvaerensis Bergström, Baltoniodus variabilis Bergström, Aphelognathus cf. politus (Hinde), Eoplacognathus elongatus (Bergström), Erismodus typus Branson y Mehl, Erismodus sp., Protopanderodus varicostatus (Sweet y Bergström), Oneotodus? ovatus (Stauffer) y Plectodina sp. Las muestras LP1 y LP2 proporcionaron, además de los citados conodontes, numerosas placas de arandáspidos similares a las reconocidas por Albanesi et al. (1995) como Sacabambaspis janvieri Gagnier, como así también braquiópodos, espinas de trilobites, briozoos, crinoideos y espículas de esponjas. Para sostener estas identificaciones se mencionan brevemente las características principales de estas especies, las que han sido descriptas y señaladas con frecuencia en la literatura especifica, por lo que no es necesario realizar una caracterización taxonómica de cada una de ellas.
Amorphognathus tvaerensis. Se han rescatado 5 elementos que corresponden a formas P y S (figura 5.1, 5.7- 8). Todos los ejemplares provienen de la muestra LP 2.
Baltoniodus variabilis. Se han separado 9 elementos entre formas Pb y S, (figura 5. 2, 5.3-4) en las muestras LP1 y LP2. En éstos ejemplares se ha reconocido que la morfología del elemento Pb corresponde a un estadio evolutivo intermedio, que permite identificar un intervalo bien definido dentro de la subzona homónima (Bergström y Carnes, 1976), pues este elemento Pb presenta un ensanchamiento de la plataforma y una disposición cercana a los 90°. Este cambio morfológico es característico de los tramos intermedios de la subzona, y corresponde al inicio de la transición de B. variabilis hacia la especie sucesora Baltoniodus gerdae Bergström.
Aphelognathus cf. politus. Se reconocieron varios elementos Pa (figura 5.9-10) caracterizados por la forma y distribución de los dentículos, así como por la amplitud y ubicación de la cavidad basal. Se han reconocido 6 elementos que provienen de la muestra LP 2. La gran similitud con A. politus que exhiben estas formas P, y los pocos elementos ramiformes (dos ejemplares) que aparecen asociados, sostienen esta identificación preliminar. Este registro constituye la primera mención de esta especie asociada a conodontes diagnósticos de zona y subzona, lo que permite una ubicación temporal de la misma en Precordillera.
Eoplacognathus elongatus. Se han rescatado 8 fragmentos que corresponden en su mayoría a formas Pb (figura 5.5), excepto un fragmento de proceso lateral que podría pertenecer a un elemento Pa (figura 5.6). Todo este material proviene de la muestra LP 2.
Erismodus typus. Está representado por 4 elementos S (figura 5.11-12) que fueron obtenidos de la muestra LP2. Un elemento S de gran tamaño (1,5 mm) de Erismodus sp. (sin figurar) ha sido identificado en la muestra LP1.
Plectodina sp. Se han separado 7 elementos de la muestra LP 2. Estas formas se asignan al género Plectodina por la disposición y tamaño de los dentículos, por la curvatura de la cúspide, y el diseño de la cavidad basal (figura 5.13-14).
Protopanderodus varicostatus. Se han obtenido dos ejemplares en la muestra LP 2 (figura 5.16). Éstos resultan diagnósticos por la disposición y morfología de las costillas laterales.
Oneotodus? ovatus. Se recuperó un solo ejemplar de la muestra LP1 (figura 5.15).
Bioestratigrafía de conodontes y edad de los depósitos de la Formación La Pola
Astini (2001), al momento de la descripción y definición de la Formación La Pola, y utilizando un criterio estratigráfico, sugirió para estos materiales una edad comprendida entre el Llandeiliano y el Ashgilliano (en el sentido de las series británicas). Brussa (2000), al analizar la graptofauna de los clastos (ver Astini, 2003) y al no encontrar especies diagnósticas de biozona, definió la asociación como perteneciente a un lapso comprendido entre las biozonas de Nemagraptus gracilis/Climacograptus bicornis con la posibilidad de extender la edad hasta la Zona de Dicranograptus clingani. Sánchez et al. (2003) sugirieron una edad caradociana temprana (Sandbiano temprano) para estos depósitos, en base a bivalvos.
Benedetto (2003) realizó una revisión de la edad de estos afloramientos y aportó nuevos datos basándose en el estudio de una colección de braquiópodos proveniente de los tramos cuspidales de la Formación La Pola. Este autor determinó una edad similar, o ligeramente más joven, a la de la Formación Las Plantas, la cual porta graptolitos de la Zona de Climacograptus bicornis (Ortega y Brussa, 1990).
En el marco de la presente contribución, los conodontes identificados permiten atribuir estos niveles a la Zona de Amorphognathus tvaerensis, Subzona de Baltoniodus variabilis (figura 2), lo que conlleva asignar una edad más antigua que la propuesta anteriormente para esta unidad (excepto por el aporte de Sánchez et al., 2003). De esta forma, el hiato sugerido por Astini (2001) entre la Formación La Cantera y la Formación La Pola carece de representatividad a nivel de biozonas de conodontes y graptolitos.
Debido a las características del depósito señaladas por Astini (2001), se podría argumentar que la conodontofauna ha sido heredada de niveles más antiguos y que éstos se han incorporado a la Formación La Pola mediante los clastos de diferentes granulometrías que componen la matrix y los clastos propiamente dichos, y que por lo tanto las edades mencionadas no corresponden al tiempo del depósito (elementos reelaborados), o bien que la conodontofauna ha sufrido transporte desde áreas más someras (faunas parautóctonas). Atendiendo a esta posibilidad, el muestreo se realizó en los mismos niveles donde se encuentran los braquiópodos descriptos como autóctonos por Benedetto (2003). En este marco puede sostenerse que el total de la asociación de conodontes rescatada es típica de los tramos medios de la Subzona de Baltoniodus variabilis. Además, no se observan evidencias de abrasión en los ejemplares, o una mezcla de elementos de conodontes a nivel taxonómico que indique diferentes procedencias dentro de las áreas de la plataforma (ver Consideraciones biofaciales y tafonómicas) o diferentes edades.
La presente conodontofauna define de manera precisa el tiempo de depósito de la Formación La Pola, la que se asigna al Sandbiano temprano, Zona de A. tvaerensis, Subzona de Baltoniodus variabilis. En términos de biozonas de graptolitos, correspondería a los tramos medios de la Zona de Nemagraptus gracilis.
Consideraciones biofaciales y tafonómicas
Los conodontes recuperados son formas robustas, algunos con procesos rotos, y no se han encontrado formas juveniles ni delicadas. Estas observaciones permiten interpretar la presente conodontofauna como conformada por elementos autóctonos de ambientes de alta energía (Armstrong y Owen, 2002). A esta interpretación se le suma la aparición de restos de Sacabambaspis janvieri Gagnier, ostracodermo ordovícico que suele estar vinculado a ambientes de plataforma somera de alta energía (Davies et al., 2007). La muestra LP1 contiene numerosas placas dérmicas de Sacabambaspis janvieri, presentando distinto grado de preservación superficial, algunas denotando una intensa abrasión, mientras que los conodontes de este nivel corresponden a formas robustas. Esto evidencia que la acumulación de estos niveles, representados por la muestra LP1, se realizó en un medio de alta energía.
El análisis de la muestra LP2 revela mayor porcentaje de cemento carbonático sobre la fracción clástica, con elementos bioclásticos constituidos por restos de braquiópodos, crinoideos, briozoos, gastrópodos y conodontes, interpretándose como un ambiente más calmo que el anterior. Desde el punto de vista de las Biofacies de Conodontes se determina aquí que el nivel del que proceden estos conodontes corresponde a depósitos de plataforma media, en base a la abundancia relativa del género Baltoniodus con respecto a los otros géneros presentes (Rassmusen y Stouge, 1995; Pålsson et al., 2002; Heredia et al., 2005; Heredia y Rosales, 2006). Se suma a esto la presencia de Aphelognathus, Erismodus y Plectodina como elementos clave para diagnosticar biotopos de plataforma (Nowlan y Haidl, 2001; Armstrong y Owen, 2002).
Sin embargo, Astini (2001) ha expresado que estos depósitos corresponden a un talud tectónicamente activo. Sobre esta aseveración es conveniente remarcar que, si bien es evidente una clinoforma con un cierto gradiente a fin de generar transporte gravitatorio de detritos, las condiciones bajo las cuales se depositaron estos sedimentos parecen indicar un ambiente de plataforma media a media-somera. Las evidencias que soportan esta afirmación son la presencia de conodontes típicos de estos hábitats y la manifiesta ausencia de otros elementos indicadores de aguas más frías y profundas, alejadas de ambientes de alta energía.
Consideraciones sobre provincialismo
Tal como fuera expresado por Benedetto (2003) en cuanto a las relaciones de provincialismo de los braquiópodos, las especies de conodontes identificadas revelan un provincialismo faunístico casi idéntico al mencionado por este autor. Se observan fuertes vínculos con la Provincia Midcontinent (Zhen y Percival, 2003) a los que se suman algunos elementos de procedencia noratlántica de clara signatura pandémica para el Ordovícico Superior, y que han sido mencionados en varias secciones de EE.UU. y Canadá, en donde se los ha identificado como faunas invasoras desde áreas noratlánticas (Nowlan, 1983). Si bien es cierto que existen elementos de extracción noratlántica tales como Amorphognathus, Baltoniodus y Eoplacognathus, éstos géneros (para el Ordovícico Medio alto y el Ordovícico Superior) tienen una gran dispersión en regiones atribuibles al Midcontinent (Nowlan, 1983; Bauer, 1994) y se consideran típicos de aguas oceánicas (Armstrong y Owen, 2002).
Aparentemente, los conodontes presentes en otras formaciones en las que se ha identificado la Biozona de A. tvaerensis en la Precordillera, tales como Las Aguaditas (Lehnert, 1995), Las Plantas (Albanesi y Ortega, 1998) y el tope de la Formación Los Azules (Albanesi y Ortega, 1998), poseen una impronta fuertemente pandémica dada por la posición de estos depósitos de plataforma distal/talud en la cuenca de Precordillera (Albanesi y Ortega, 1998).
Por otra parte, los registros de Sacabambaspis janvieri Gagnier se remiten a pocas localidades en América del Sur, se lo ha descripto para la Formación Anzaldo en Bolivia (Gagnier et al., 1986), donde se encuentra en depósitos mixtos del Ordovícico Superior (Egenhoff et al., 2007). En Argentina ha sido mencionado en calizas con una fuerte composición clástica y de edad supuestamente Ordovícico Medio-Superior baja? de la Cordillera Oriental de la Provincia de Jujuy (Albanesi y Astini, 2002), y en la Sierra de Mojotoro (Salta) del Ordovícico Superior (Moya et al., 2003). Dentro delámbito de la Precordillera, esta especie se ha mencionado en calizas alóctonas que aparecen como clastos en la Formación La Cantera (Albanesi et al., 1995) donde están presentes junto a conodontes de la Zona de Pygodus serra y de la Subzona de Eoplacognathus lindstroemi (figura 6), señalando una edad darriwiliana alta (Ordovícico Medio), constituyendo así el registro más antiguo para estos arandáspidos gondwánicos en América del Sur. En el presente trabajo, las placas de Sacabambaspis janvieri provenientes de las muestras LP1 y LP2 se han encontrado junto a una conodontofauna del Sandbiano temprano (Ordovícico Superior), la cual exhibe una fuerte impronta de la Provincia Midcontinent. Esto ofrece un panorama interesante pues S. janvieri ha sido definido como un elemento diagnóstico para determinar el provincialismo faunístico gondwánico (Young, 1997), situación que fue mencionada por Albanesi et al. (1995) dentro del contexto de la aloctonía de la Precordillera y el momento de su integración con Gondwana. Por otro lado, el gran número de placas rescatadas de los depósitos de la Formación La Pola indica que las poblaciones de arandáspidos eran numéricamente importantes en este sector de la cuenca.
El análisis paleobiogeográfico de los conodontes de la Formación La Pola sugiere una situación muy vinculada con la Provincia Midcontinent (Bergström, 1990; Sweet y Bergström, 1974; Zhen y Percival, 2003), fenómeno bastante común durante el Sandbiano temprano (Zona de A. tvaerensis) y que se hace más notorio en secciones cuyos biotopos se interpretan como depósitos de plataforma mediasomera (Sassito y La Pola) (Keller y Lehnert, 1998; Keller, 1999; esta contribución). Durante el Sandbiano tardío (Zona de A. superbus) esto se modifica cuando recibe una influencia ya definitiva de la región perigondwánica de la subprovincia Británica (Bergström, 1990), afinidad que continúa hasta el Katiano (Heredia et al., 1990; Heredia, 2008).
Durante el Darriwiliano tardío se inicia una de las transgresiones globales más importantes del Paleozoico (Barnes, 2003) y que durante el Sandbiano temprano llega a su punto máximo con la representación de un highstand de relieve global. Esta sería la respuesta al interrogante acerca de la dispersión de faunas de la Provincia Midcontinent sobre Gondwana durante el Ordovícico Superior (Heredia y Mestre, 2007).
Correlación
Astini (2003) y Brussa et al. (2003) correlacionaron la Formación La Pola con las formaciones Las Plantas/Las Vacas, Sassito, Empozada y Los Azules. Las edades de las formaciones clástico-carbonáticas sandbianas en la Precordillera (formaciones La Pola y Sassito) y las clásticas (formaciones Las Plantas/Las Vacas, Los Azules y Empozada) están acotadas por graptolitos y conodontes. Luego de un análisis detallado de estos datos paleontológicos se observa que no existe una correspondencia temporal estricta (referidas en zonas y subzonas de conodontes y graptolitos) entre todas las unidades mencionadas (figura 6).
Conclusiones
Se define la edad de los depósitos de la Formación La Pola, que afloran en la Quebrada La Pola (Sierra de Villicum) como sandbiana baja (Ordovícico Superior temprano) mediante el registro de la Zona de A. tvaerensis, subzona de B. variabilis. De esta manera, prácticamente se elimina el hiato sugerido entre las formaciones La Cantera y La Pola, siendo altamente probable que la Formación La Pola se haya depositado casi en continuidad sobre la unidad infrayacente. También se menciona el hallazgo de placas del arandáspido Sacabambaspis janvieri Gagnier, que contribuye a definir las características tanto biofaciales de esta unidad, sugiriendo una plataforma media a media-somera, como las paleobiogeográficas, que la vinculan a Gondwana. Por otro lado la conodontofauna sugiere un importante vínculo con la Provincia Midcontinent.
Agradecimientos
A W. Sweet, J. Bauer y S. Leslie por las excelentes sugerencias sobre los géneros Aphelognathus y Plectodina. A S. Finney por las discusiones sobre las Formaciones La Cantera y la Pola. A G. Sarmiento por revisar y mejorar el manuscrito. A un revisor anónimo por sus sugerencias. Al LAMENOA (UNT) por el Servicio de Microscopia Electrónica y a G.F. Aceñolaza por sostener económicamente este aspecto. A A. Mestre por su colaboración con las figuras y a M. González por su asistencia en las tareas de laboratorio.
Bibliografía
1. Albanesi, G. y Astini, R. 2002. Faunas de conodontes y Sacabambaspis janvieri (Vertebrata) en el Ordovícico Medio de la Cordillera Oriental argentina: implicancias estratigráficas y paleobiogeográficas. 8º Congreso Argentino de Paleontología y Bioestratigrafía (Corrientes), Resumenes, p. 17. [ Links ]
2. Albanesi, G. y Ortega, G. 1998. Conodont and Graptolite Faunas from the Las Plantas Formation and equivalent units (Caradoc) in the Argentine Precordillera. 7th International Conodont Symposium held in Europe (Bologna-Modena), Abstracts: 1-2. [ Links ]
3. Albanesi, G. y Ortega, G. 2002. Advances on conodont-graptolite biostratigraphy of the Ordovician System of Argentina. Serie de Correlación Geológica 16: 143-166. [ Links ]
4. Albanesi, G., Benedetto, J. y Gagnier, P.-Y. 1995. Sacabambaspis janvieri (Vertebrata) y conodontes del Llandeiliano temprano en la Formación La Cantera, Precordillera de San Juan, Argentina. Boletín de la Academia Nacional de Ciencias 60: 519-543. [ Links ]
5. Armstrong, H.A. y Owen, A.W. 2002. Euconodont diversity changes in a cooling and closing Iapetus Ocean. Special Publication of the Geological Society of London 194: 85-98. [ Links ]
6. Astini, R. 2001. Formación La Pola (Ordovícico Superior) relicto erosivo de la glaciación hirnantiana en la Precordillera argentina. Revista de la Asociación Geológica Argentina 56: 425 442. [ Links ]
7. Astini, R. 2003. The Ordovician Proto-Andean basins. En: Benedetto, J.L. (ed.). Ordovician fossils of Argentina. Secretaría de Ciencia y Tecnología, Universidad Nacional de Córdoba: 1- 74. [ Links ]
8. Barnes, C. 2003. Ordovician Oceans and Climate. En: B. Webby, F. Paris, M. Droser y I. Percival (eds.), The Great Ordovician biodiversification event, Columbia University Press (New York): 72- 76. [ Links ]
9. Bauer, J. 1994. Conodonts from the Bromide Formation (Middle Ordovician), south-central Oklahoma. Journal of Paleontology 68: 358-376. [ Links ]
10. Benedetto, J. L. 1986. The first typical Hirnantia Fauna from South America (San Juan Province, Argentine Precordillera. En: P.R. Racheboeuf y C.C. Emig (eds.), Les Brachiopodes fossiles et actuels: Biostratigraphie du Paleozoïque, v. 4: 439-447. Brest, Francia. [ Links ]
11. Benedetto, J. L. 2003. Braquiópodos del Caradociano temprano de la Formación La Pola, sierra de Villicúm, Precordillera de San Juan (Argentina). Ameghiniana 40: 33 52. [ Links ]
12. Bergström, S. 1990. Relations between conodont provincialism and changing palaeogeography during the Early Palaeozoic. En: W.S. McKerrow y C.R. Scotese (eds.), Palaeozoic Palaeogeography and Biogeography, Memoir of the Geological Society of London 12: 105-121. [ Links ]
13. Bergström, S. y Carnes, J.B. 1976. Conodont biostratigraphy and paleoecology of the Holston Formation (Middle Ordovician) and associated strata in Eastern Tennessee. Geological Association of Canada, Special Paper 15:27-57. [ Links ]
14. Brussa, E. 2000. Una nueva asociación de graptolitos caradocianos en la Formación La Cantera, sierra de Villicúm, Precordillera argentina. Ameghiniana 37 Suplemento: 7R [ Links ]
15. Brussa, E., Mitchell, C., Ortega, G. y Astini, R. 2003. Middle Ordovician Graptolite bioestratigraphy from the Los Azules Formation at Los Gatos Creek, Central Precordillera, Argentina. Serie Correlación Geológica, 18: 21-25. [ Links ]
16. Davies, N., Sansom, I., Albanesi, G. y Céspedes, R. 2007. Ichnology, palaeoecology and taphonomy of a Gondwanan early vertebrate habitat: Insights from the Ordovician Anzaldo Formation, Bolivia. Palaeogeography, Palaeoclimatology, Palaeoecology 249: 18-35 [ Links ]
17. Egenhoff, S., Webber, B., Lehnert O. y Maletz, J. 2007. Biostratigraphic precision of the Cruziana rugosa group: a study from the Ordovician succession of Southern and Central Bolivia. Geological Magazine 144: 289-303. [ Links ]
18. Epstein, A.G., Epstein, J.B. y Harris, L.D. 1977. Conodont color alteration - an index to organic metamorphism. U.S. Geological Survey Professional Paper 995: 1-27. [ Links ]
19. Finney, S. 2006. Global Series and Stages for the Ordovician System. Geologica Acta 3: 309-316. [ Links ]
20. Gagnier, P.Y., Blieck, A. y Rodrigo, G. 1986. First Ordovician vertebrates from South America. Geobios 19: 629-634. [ Links ]
21. Heredia, S. 2008. Primer registro de Conodontes ashgillianos en la Precordillera (Argentina). Ameghiniana 45: 59 - 68. [ Links ]
22. Heredia, S. y Mestre, A. 2007. Conodont Provincialism in the Upper Ordovician of the Cuyania terrane (Argentina). Cuadernos del Museo Geominero 8: 189-193. [ Links ]
23. Heredia, S. y Rosales, C. 2006. Biofacies de Conodontes de la Formación Ponón Trehué y la importancia bioestratigráfica como sección tipo para el límite del Ordovícico Medio- Ordovícico Superior de Cuyania (Argentina). Serie Correlación Geológica 21: 7-16. [ Links ]
24. Heredia, S., Gallardo, G. y Maldonado, A. 1990. Conodontes caradocianos en las calizas alóctonas del Miembro Superior de la Formación Empozada (Ordovícico Medio y Superior), San Isidro (Mendoza, Argentina). Ameghiniana 27: 197-2006. [ Links ]
25. Heredia, S., Rosales, C., Peralta S. y Beresi., M. 2005. Conodontes de la Formación Las Chacritas (Darriwiliano) y su significado tecto-sedimentario en el bloque Sierra de la Trampa-Los Blanquitos, Precordillera de San Juan. 16º Congreso Geológico Argentino (La Plata), Actas 3: 445-450. [ Links ]
26. Jeppsson, L., Fredholm, D. y Mattiasson, B. 1985. Acetic acid and phosphatic fossils: a warning. Journal of Paleontology 59: 952- 956. [ Links ]
27. Keller, M. 1999. Argentine Precordillera: Sedimentary and Plate Tectonic History of a Laurentian Crustal Fragment in South America. Geological Society of America, Special Paper 341: 1-131. [ Links ]
28. Keller, M. y Lehnert, O. 1998. The Rio Sassito sedimentary succession (Ordovician): a pinpoint in the geodynamic evolution of Argentine Precordillera. Geologische Rundschau 87: 326-344. [ Links ]
29. Keller, M., Eberlein, S. y Lehnert, O. 1993. Sedimentology of Middle Ordovician carbonates in the Argentine Precordillera: evidence of regional relative sea-level changes. Geologische Rundschau 82: 362-377. [ Links ]
30. Lehnert, O. 1995. Ordovizische Conodonten aus der Präkordillere Westergentinienes: Ihre Bedetung für Stratigraphie und Paläogeographie. Earlanger Geologische Abbandlungen 125: 1- 193. [ Links ]
31. Moya, M.C., Monteros, J.A., Malanca, S., Ortega, G. y Albanesi, G.L. 2003. The Mojotoro Range, Eastern Cordillera, Salta Province. Ordovician and Silurian of the Cordillera Oriental and Sierras Subandinas, NW Argentina. Instituto Superior de Correlación Geológica, Miscelánea 11: 17-22. [ Links ]
32. Nowlan, G. 1983. Biostratigraphic, paleogeographic and tectonic implications of Late Ordovician conodonts from the Grog Brook Group, northwestern New Brunswick. Canadian Journal of Earth Sciences 20: 651-671. [ Links ]
33. Nowlan, G. y Haidl, F. 2001. Biostratigraphy and paleoecology of Late Ordovician Conodonts from a composite section in the subsurface of Saskatchewan. Saskatchewan Geological Survey, Saskatchewan Energy Mines, Miscellanea Reprints 2001/4 1: 14- 31. [ Links ]
34. Ortega, G. y Brussa, E.D. 1990. La subzona de Climacograptus bicornis (Caradociano temprano) en la Formación Las Plantas en su localidad tipo, Precordillera de San Juan, Argentina. Ameghiniana 27: 281-288. [ Links ]
35. Ortiz, A. y Zambrano, J. 1981. La Provincia geológica Precordillera Oriental. 8º Congreso Geológico Argentino (San Luis) Actas 3: 59-74. [ Links ]
36. Pålsson, C., Månsson, K. y Bergström, S.M. 2002. Biostratigraphical and palaeoecological significance of graptolites, trilobites and conodonts in the Middle-Upper Ordovician Andersö Shale: an unusual "mixed facies" deposit in Jämtland, central Sweden. Transactions of the Royal Society of Edinburgh, Earth Sciences 93: 35-57. [ Links ]
37. Peralta, S.H. 1993. Estratigrafía y consideraciones paleoambientales de los depósitos marino-clásticos eopaleozoicos de la Precordillera Oriental de San Juan. 12º Congreso Geológico Argentino (Mendoza), Actas 1: 128-137. [ Links ]
38. Peralta, S.H. y Baldis, B.A. 1990. Glyptograptus persculptus en la Formación Don Braulio (Ashgilliano tardío-Llandoveriano temprano) en la Precordillera Oriental de San Juan, Argentina. Serie Correlación Geológica 7: 67-72. [ Links ]
39. Peralta, S.H. y Carter, C.H. 1990. La glaciación gondwánica del Ordovícico tardío: evidencias en fangolitas guijarrosas de la Precordillera de San Juan. 11º Congreso Geológico Argentino (San Juan), Actas 2: 181-185. [ Links ]
40. Peralta, S.H. y Carter, C.H. 1999. Don Braulio Formation (late Ashgillian-early Llandoverian, San Juan Precordillera, Argentina): stratigraphic remarks and paleoenvironmental significance. Acta Universitatis Carolinae, Geologica, 43: 225-228. [ Links ]
41. Rasmussen, J.A. y Stouge, S. 1995. Late Arenig-Early Llanvirn conodont Biofacies across the Iapetus Ocean. En: D.J. Cooper, M.L. Droser y S.C. Finney (eds.), Ordovician Odyssey: Short Papers for the Seventh International Symposium on the Ordovician System, 443-447. SEPM, Pacific Section, Book 77, Fullerton, California. [ Links ]
42. Sánchez, T., Marengo, L. y Astini, R. 2003. Late Ordovician Bivalvia Heteroconchia (Cycloconchidae and Glyptarcidae) from western Argentina. Ameghiniana 40: 433-440. [ Links ]
43. Stone, J. 1987. Review of investigative techniques used in the study of conodonts. En: R.L. Austin (ed.), Conodonts: Investigative techniques and applications, Ellis Horwood Limited, pp. 17- 34. [ Links ]
44. Sweet, W. 1979. Late Ordovician conodonts and biostratigraphy of the western Midcontinent Province. En: C. A. Sandberg y D. L. Clark (Eds.), Conodont Biostratigraphy of the Great Basin and Rocky Mountains. Brigham Young University Geology Studies 26: 45-85. [ Links ]
45. Sweet, W. y Bergström, S. 1974. Provincialism exhibited by Ordovician conodonts faunas. En: C.A. Ross (ed.), Paleogeographic provinces and provinciality, Society of EconomicPaleontologists and Mineralogists Special Publication 21: 189-202. [ Links ]
46. Young, G.C. 1997. Ordovician microvertebrates remains from the Amadeus Basin, central Australia. Journal of Vertebrate Paleontology 17: 1-25. [ Links ]
47. Zhen, Y. y Percival, I. 2003. Ordovician conodont biogeography - reconsidered. Lethaia 36: 357-369. [ Links ]
Recibido: 10 de junio de 2009.
Aceptado: 30 de mayo de 2010.

