










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de la Asociación Geológica Argentina
versión impresa ISSN 0004-4822versión On-line ISSN 1851-8249
Rev. Asoc. Geol. Argent. v.62 n.2 Buenos Aires abr./jun. 2007
Carbonatos subtropicales de la Formación Calabozo (Caloviano) y su diagénesis, Cuenca Neuquina, Mendoza
Palma, R.M.1,2, Mehdli, M.3, Bressan, G.S.1 y Kietzmann, D.A.1
1Departamento de Ciencias Geológicas, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Intendente Güiraldes 2160, Ciudad Universitaria Pabellón II, C1428 EGA, Buenos Aires. E-mails: palma@gl.fcen.uba.ar; gbressan@gl.fcen.uba.ar, diegokietzmann@fibertel.com.ar
2 Consejo Nacional de Investigaciones Científicas y Técnicas.
3 Université Ibn Tofail, Faculte des Sciences, Department de Geologie, BP 133, Kénitra, Marruecos. E-mail: mehdimina@hotmail.com.
RESUMEN
El estudio de los carbonatos calovianos de la Formación Calabozo (30 m) procedente del centro-oeste de Argentina revela la abundancia de partículas carbonáticas bentónicas producidas por organismos fotodependientes y de granos no-esqueletales tales como ooides y peloides. La biota está representada por bivalvos, dasicladáceas (Salpingoporella annulata Carozzi, Cylindroporella annulata Carozzi, y Heteroporella sp.), corales escleractínidos, equinodermos, cianobacterias (Cayeuxia (Rivularia) piae Rech-Frollo, y Cayeuxia (Rivularia) kurdistanensis Elliot), oncoides, gastrópodos, foraminíferos, calciesferas y en menor cantidad, briozoos y ostrácodos. Las partículas aloquímicas sugieren para las calizas marinas someras de la Formación Calabozo una asociación Photozoan de aguas subtropicales. Las características petrográficas permitieron reconocer siete microfacies diferenciadas de acuerdo al tipo de partículas, fósiles y textura depositacional: 1) bindstone criptalgal; 2) wackestone bioclástico-peloidal; 3) wackestone bioclástico-intraclástico; 4) wackestone bioclástico-peloidal-terrígeno, 5) boundstone coralino, 6) packstone bioclástico-peloidal-intraclástico, y 7) packstone/grainstone oolítico-bioclástico-peloidal. El reconocimiento de grupos de microfacies constituyen dos asociaciones paleoambientales: rampa interna y rampa media. Los procesos de micritización y precipitación de micrita peloidal y cemento acicular son sinsedimentarios. Estos cementos son previos a la fase de cementación de calcita granular y sintaxial. Estudios de cátodoluminiscencia permitieron identificar tres tipos de cementos: (1) acicular, (2) calcita granular, y (3) cemento sintaxial en partículas de equinodermos. La cátodoluminiscencia permitió determinar cementos no luminiscente, cemento con luminiscencia opaca (roja/naranja) y cemento con luminiscencia amarilla brillante. El cemento acicular es no luminiscente (marino), en tanto que la luminiscencia opaca a brillante típica de las cavidades inter-intraparticulares y del cemento sintaxial indica un ambiente diagenético meteórico-freático.
Palabras clave: Microfacies; Rampa carbonática; Asociación photozoan; Diagénesis; Jurásico.
ABSTRACT
Subtropical carbonates from the Callovian Calabozo Formation, and their diagenesis, Neuquen Basin, Mendoza province. Detailed study of Callovian carbonates of the Calabozo Formation (30 m) in western Argentina, reveals the abundance of benthic carbonates particles produced by organisms that are light dependent, and non-skeletal grains such as ooids and peloids. The biota consists of bivalves, dasyclads (Salpingoporella annulata Carozzi, Cylindroporella annulata Carozzi, and Heteroporella sp.), scleractinian corals, echinoderms, cyanobacteria (Cayeuxia (Rivularia) piae Rech-Frollo, and Cayeuxia (Rivularia) kurdistanensis Elliot), oncoids, gastropods, forams, and calcispheres as well as ostracods, and bryozoans in relatively minor quantities. Allochemical particles suggest for the shallow marine limestones of the Calabozo Formation a Photozoan Association under subtropical seawater conditions. The petrographic features allow to recognize seven microfacies, differentiated by particle types, fossils and depositional texture: 1) criptalgal bindstone; 2) bioclastic-peloidal wackestone; 3) bioclastic-intraclastic wackestone; 4) bioclastic-peloidal-terrigenous wackestone, 5) coral framestone, 6) peloidal-bioclastic-intraclastic packstone, and 7) oolitic-bioclastic-peloidal packstone-grainstone. The recognition of microfacies groups constitute two palaeoenvironmental associations: inner ramp and middle ramp settings. Diagenetic studies are based on examination of stained thin sections, through conventional petrography and under cathodoluminescence. Early diagenetic processes are micritization and precipitation of peloidal micrite and acicular calcite. These cements predate granular and syntaxial calcite cements. Cathodoluminescent petrography allows to identify three morphologic types of clear cements: (1) fine acicular crystals, (2) granular cements, and (3) syntaxial cement rims on echinoderm particles. Description of the CL signature has evolved to discrimination as either, non-luminescent, dully luminescent red/orange, or brightly luminescent yellow. The earlier acicular calcite cement is non-luminescent, on the other hand, the predominantly dully luminescent red/orange, or brightly luminescent yellow of interparticular and intraparticular granular calcite and syntaxial cements indicate a shallow burial meteoric-phreatic diagenetic environment.
Keywords: Microfacies; Carbonate ramp; Photozoan Association; Diagenesis; Jurassic.
INTRODUCCIÓN
El relleno de la cuenca Neuquina ha sido subdividido en diferentes mesosecuencias sobre la base de discontinuidades estratigráficas de carácter regional, mayormente influenciadas por eventos eustáticos (Legarreta et al. 1993). En este esquema, la Formación Calabozo se incluye dentro de la Mesosecuencia Cuyo, equivalente al Ciclo Cuyano (Gulisano et al. 1984a), limitado en base y techo por discontinuidades regionales. Dichas discontinuidades son la Intra-liásica (Gulisano et al. 1984b) en la base y la Intra-caloviana en el techo (Dellapé et al. 1979).
En la provincia de Mendoza, la Mesosecuencia Cuyo (Fig. 1) está compuesta, en orden ascendente, por los depósitos continentales de la Formación El Freno, seguidos por los depósitos marinos de plataforma de la Formación Puesto Araya, y las lutitas negras (offshore) de la Formación Tres Esquinas. A continuación de estas últimas se encuentran los depósitos marinos de la Formación Lajas y los carbonáticos de la Formación Calabozo. La sucesión culmina con las evaporitas de la Formación Tábanos, que registran una caida global del nivel del mar (Riccardi et al. 2000).
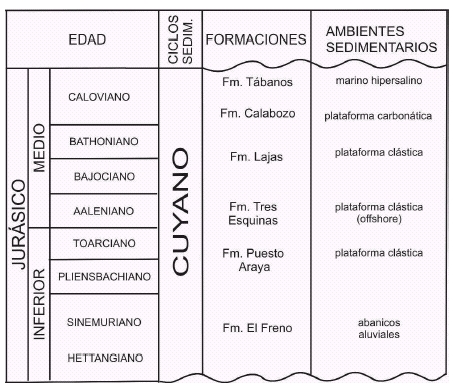
Figura 1: Cuadro estratigráfico del ciclo cuyano en la cuenca Neuquina.
La Formación Calabozo está compuesta por una amplia variedad de rocas calcáreas que incluyen wackestones-packstones bioclásticos-peloidales, packstones-grainstones oolíticos, estromatolitos y framestone coralino. El contenido fosilífero está representado por bivalvos, algas verdes calcáreas, corales escleractínidos, equinodermos, cianobacterias, oncoides, gastrópodos, foraminíferos, calciesferas, briozoos y ostrácodos. Los componentes no esqueletales están representados por ooides, peloides e intraclastos. Estos componentes aloquímicos se encuentran asociados a una escasa proporción de partículas siliciclásticas (Palma et al. 2000). Su espesor alcanza a 30 metros.
En el ámbito de la cuenca Neuquina se han realizado numerosos trabajos de carácter regional (Legarreta y Gulisano 1989, Legarreta et al. 1993, entre otros) sin embargo, es escasa la información existente sobre la composición y variabilidad de las microfacies de los depósitos carbonáticos de la Formación Calabozo, a excepción de Palma et al. (2000, 2005) y Armella et al. (2005). En este último artículo los autores mencionan la presencia de Halimeda (algas verdes del Orden Caulerpales-Chlorophyta) en depósitos de rampa interna. Cabe destacar que una interpretación de esta naturaleza es errónea, ya que Halimeda tiene su biocrón desde el Cretácico a la actualidad, en tanto que a la Formación Calabozo le corresponde una edad caloviana media inferior.
El objetivo de este artículo es mostrar la composición de los depósitos carbonáticos en base a un estudio detallado de microfacies carbonáticas, su diagénesis y sus implicancias ambientales. Para ello se ha tenido en cuenta además, la asociación de partículas aloquímicas y su significado desde un punto de visto paleoclimático, de acuerdo a las características de las asociaciones Photozoan versus Heterozoan (James 1997).
MARCO GEOLÓGICO
Los afloramientos de la Formación Calabozo estudiados se encuentran localizados a unos 50 km al oeste de la ciudad de Malargüe, en el sur de la provincia de Mendoza. Los mismos incluyen a los depósitos aflorantes en el arroyo Calabozo, localidad tipo de esta unidad, y observaciones adicionales en aquellos ubicados a unos 5 km al sur del primero, en las inmediaciones del arroyo Plomo (mina Chuquichenque) (Fig. 2).
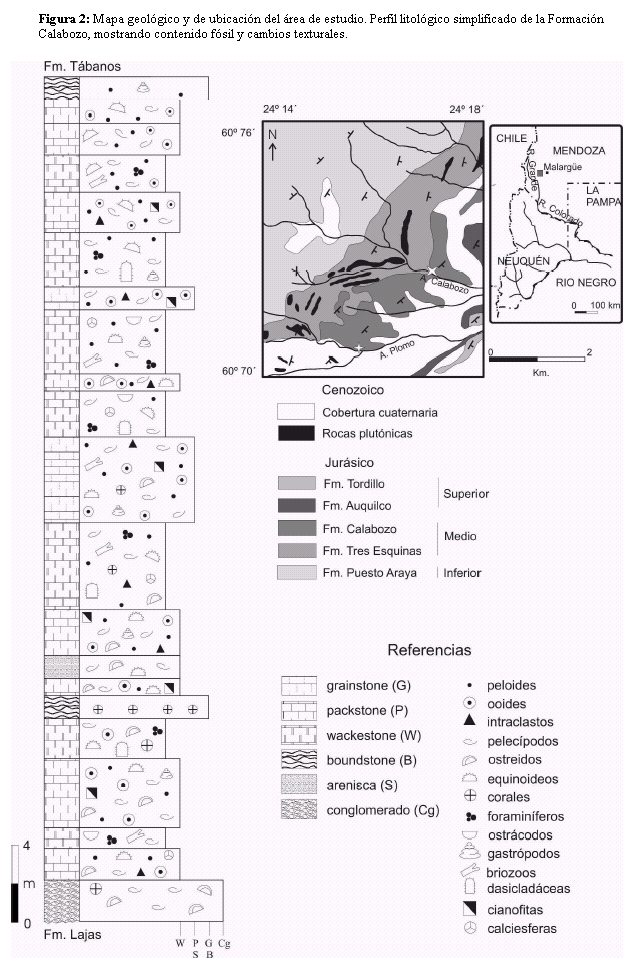
Figura 2: Mapa geológico y de ubicación del área de estudio. Perfil litológico simplificado de la Formación Calabozo, mostrando contenido fósil y cambios texturales.
La Formación Calabozo apoya sobre las facies marinas de cara de playa de la Formación Lajas (Bathoniano-Caloviano), a través de un contacto erosivo e irregular; y es cubierta en forma concordante por las evaporitas de la Formación Tábanos. El control bioestratigráfico de la Formación Calabozo está basado en la fauna de amonites, representada por Rehmannia sp., Choffatia sp., y Glossouvira sp. que caracteriza una edad caloviana media inferior (Riccardi et al. 2000).
METODOLOGÍA
En el estudio de los carbonatos de la Formación Calabozo se realizó un muestreo a escala centimétrica, con el objeto de determinar los cambios durante su evolución ambiental. En dicho muestreo se tuvo en cuenta el espesor de las litofacies y su contenido paleontológico, a la vez que se realizó un estudio cualitativo que permitió diferenciar componentes esqueletales y no esqueletales. Se estudiaron un total de sesenta láminas delgadas, provenientes de muestras extraídas del sector inferior, medio y superior de cada capa. Las características composicionales y atributos diagenéticos de la sucesión de microfacies han sido observadas mediante microscopía normal y de cátodoluminiscencia con la finalidad de determinar las diferentes fases cementantes y sus respectivas condiciones diagenéticas de formación. La microscopía de cátodoluminiscencia (fría-Technosyn) se realizó en la Facultad de Ciencias Geológicas de la Uni-versidad Complutense de Madrid, España, y sus condiciones operativas fueron mantenidas a 11-16 kV; intensidad 200-500 µA y vacío: 0,2-0,1 Torr. Los análisis de fueron realizados en el servicio de microscopía electrónica de CITEFA.
MICROFACIES E INTERPRETACIÓN AMBIENTAL
La Formación Calabozo (Fig. 2) constituye una rampa carbonática cuyos depósitos se disponen en sucesiones de somerización (Palma et al. 2000, 2005). Las litofacies están representadas por wackestones-packstones bioclásticos y peloidales, interestratificados con grainstones-packstones oolíticos-bioclásticos y ocasionales framestones coralinos. El conjunto se dispone en capas de espesores centimétricos a decimétricos con tonalidades gris oscuras, generalmente macizas, con gradación normal o localmente con estratificación cruzada tabular planar y bioturbación. Los fósiles están representados por una abundante fauna marina, que incluye bivalvos, algas verdes calcáreas (dasicladáceas), corales, equinodermos, cianobacterias, oncoides, gastrópodos, foraminíferos, además de calciesferas, briozoos y ostrácodos (Palma et al. 2000, 2005).
Sobre la base del análisis de litofacies (Palma et al. 2005) y un detallado estudio petrográfico, composicional y diagenético se han reconocido siete microfacies que caracterizan rampa interna y rampa media.
RAMPA INTERNA
Estos depósitos son los más representados e incluyen una amplia variedad de calizas, de colores gris claro y espesores que oscilan entre 50 y 90 cm. Se trata de wackestones y packstones, bioclásticos, peloidales y oolíticos, y con escasos corales. Se han reconocido seis microfacies (1, 2, 3, 4, 5, y 6); tres de las cuales (microfacies 2, 3, 4) pertenecen a la litofacies de wackestones bioclásticos peloidales, que es la más abundante en el área de estudio.
Estas microfacies fueron identificadas como estromatolitos (M-1), wackestone bioclático peloidal (M-2), wackestone bioclástico intraclástico (M-3), wackestone bioclásticopeloidal terrígeno (M-4) framestone coralino (M-5), packstone bioclástico peloidal intraclástico (M-6).
Microfacies 1: Corresponde a capas estromatolíticas con láminas milimétricas, planas a crenuladas y que alternan con láminas de automicrita y microesparita, además de peloides, gastrópodos (Nerinea sp.) y escasos ostrácodos. Su espesor varía entre 20 y 35 cm. La naturaleza planar-ondulante de la fábrica estromatolítica se debe a la actividad de las cianobacterias, y sus frecuentes discontinuidades son debidas a la presencia de fenestras paralelas a la estratificación (Fig. 3a) y al desarrollo de micro-tepees, grietas de desecación y bioturbación, probablemente causada por la acción de moluscos.
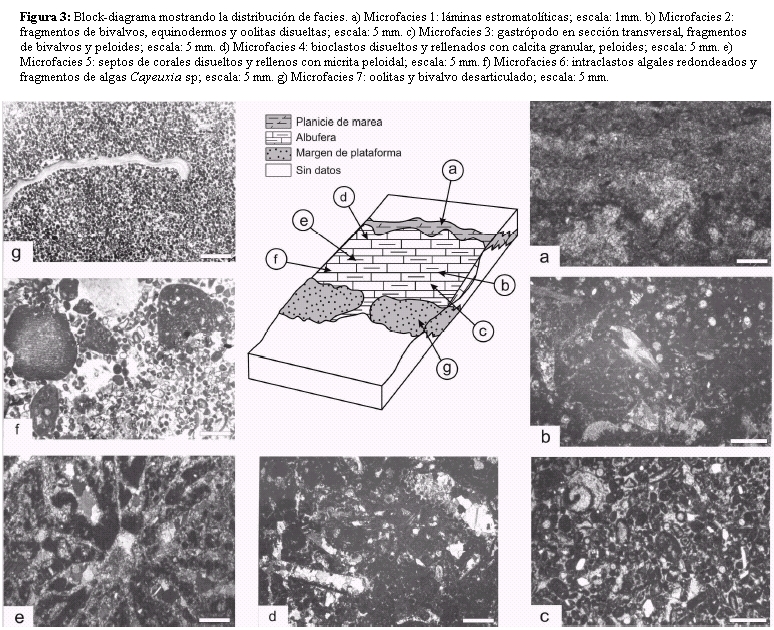
Figura 3: Block-diagrama mostrando la distribución de facies. a) Microfacies 1: láminas estromatolíticas; escala: 1mm. b) Microfacies 2: fragmentos de bivalvos, equinodermos y oolitas disueltas; escala: 5 mm. c) Microfacies 3: gastrópodo en sección transversal, fragmentos de bivalvos y peloides; escala: 5 mm. d) Microfacies 4: bioclastos disueltos y rellenados con calcita granular, peloides; escala: 5 mm. e) Microfacies 5: septos de corales disueltos y rellenos con micrita peloidal; escala: 5 mm. f) Microfacies 6: intraclastos algales redondeados y fragmentos de algas Cayeuxia sp; escala: 5 mm. g) Microfacies 7: oolitas y bivalvo desarticulado; escala: 5 mm.
Las fenestras planares pudieron haber sido generadas por la descomposición de la materia orgánica o por contracción de las láminas algales debido a la alternancia de períodos húmedos y secos (Shinn 1968), y se encuentran rellenadas con cristales de calcita granular y también microesparita. Por su parte, los micro-tepees y grietas de desecación sugieren períodos de exposición subaérea. Corresponde a un bindstone criptalgal y sugiere condiciones restringidas en el área intertidal superior de la rampa interna (Shinn 1968) como lo sugiere además, la abundancia y poca diversidad de gastrópodos (Brookfield y Brett 1988).
Microfacies 2: Esta microfacies está compuesta por partículas esqueletales muy finas no identificables y peloides, originando una textura heterogénea (Fig. 3b). Su matriz es de tipo alomicrita. La fauna está representada principalmente por bivalvos desarticulados, disueltos y rellenados por cemento de calcita granular (Fig. 4a), fragmentos de equinodermos, miliólidos y calciesferas. La mayor parte de las partículas esqueletales se encuentran afectadas por una intensa micritización producto de la actividad endolítica de algas, e inclusive se observan perforaciones, sobre todo en moluscos y equinodermos. La micritización es el resultado de la obliteración de la microestructura carbonática original, a través de un proceso de alteración a una textura criptocristalina (Reid y Macintyre 2000).

Figura 4: Microfacies: a) Microfacies 2: fragementos de moluscos disueltos y espinas de equinodermos; escala: 1 mm. b) Microfacies 3: fragmentos de bivalvos, oolitas disueltas y micritizadas; escala: 1 mm. c) Microfacies 3: foraminífero con enroscamiento tipo quinqueloculino; escala: 0,5 mm. d) Microfacies 4: sección longitudinal de gastrópodo disuelto y relleno con peloides, micrita peloidal y partículas terrígenas; escala: 1 mm. e) Microfacies 5: fragmento de bivalvo afectado por fuerte bioerosión; escala: 1 mm. f) Microfacies 6: sección transversal de briozoos incrustado con costras microbianas; escala: 1 mm. g) Microfacies 7: oolitas, fragmentos de bivalvos, intraclastos y briozoos; escala: 1 mm.
Las calciesferas, de forma más o menos esféricas y sin apertura, son interpretadas como órganos reproductores de algas dasicladáceas (Marszalek 1975) e indican un ambiente marino somero con una salinidad más o menos normal (Berger y Kaever 1992).
La naturaleza peloidal de estos wackestones, similar en todos los wackestones estudiados, así como la presencia de valvas desarticuladas, aunque sin abrasión, y el alto contenido de matriz, reflejan un ambiente subtidal somero de baja energía. Con respecto a los peloides, su origen puede estar vinculado principalmente a la fragmentación de partículas esqueletales. Se identifica a esta microfacies como wackestone bioclástico-peloidal.
Microfacies 3: Está compuesta por fragmentos de conchillas de bivalvos de composición aragonítica original, y calcita con bajo contenido en magnesio (LMC), gastrópodos (Fig. 3c) y secciones transversales de placas y espinas de equinodermos, entre los principales componentes. Los moluscos aragoníticos presentan en sus bordes internos y externos desarrollo de cemento acicular, en tanto que la cavidad interparticular está rellena por calcita granular. En algunos casos se observan crecimientos lamelares de la calcita que evidencian su neomorfismo, de manera similar a la descripta por Maliva y Dickson (1992).
Además aparecen calciesferas, escasos ooides (tipo 2, de Strasser 1986), algunos disueltos (Fig. 4b). Presentan envolturas concéntricas, fuertemente micritizadas y con frecuencia total o parcialmente recristalizadas. Otros componentes adicionales son miliólidos (Fig. 4c), y algas verdes calcáreas, representadas principalmente por Salpingoporella annulata Carozzi (Fig. 5a).

Figura 5: Algas de la Formación Calabozo. a) Salpingoporella annulata Carozzi, sección transversal; escala: 0,5 mm. b) Heteroporella sp., sección transversal; escala: 0,5 mm. c) Cylindroporella annulata Carozzi, sección transversal; escala 0,5 mm. d) Cayeuxia (Rivularia) kurdistanensis Elliot, sección longitudinal; escala: 1 mm. e) Cayeuxia (Rivularia) piae Rech-Frollo, sección transversal; escala: 2 mm.f) Cayeuxia (Rivularia) piae Rech-Frollo, sección longitudinal; escala: 1 mm.
Entre los componentes no esqueletales se reconocen peloides e intraclastos. Los peloides aparecen redondeados y con buena selección y su origen podría ser detrítico, es decir por la destrucción de valvas de moluscos micritizadas (peloides bioerosionales) o bien a partir de la abrasión de algas (peloides algales) (Flügel 2004). Los intraclastos son subangulares y tienen en general una composición similar a esta microfacies, aunque aparecen otros de packstones peloidales o bioclásticos. Todos los componentes bioclásticos aparecen como cortoides (Flügel 2004) mostrando una fuerte micritización. La matriz es tipo alomicrita asociada con peloides.
Los especímenes algales están generalmente fragmentados, probablemente por la acción de corrientes, en tanto que los intraclastos son producto de la acción de corrientes erosivas de tormentas, sobre un sustrato semiconsolidado.
Los microfósiles y algas calcáreas sugieren condiciones intermitentes de energía, en un área subtidal somera de la rampa interna, por detrás de un margen oolítico. Se interpreta que los escasos ooides fueron transportados durante la acción de tormentas. Esta microfacies corresponde a wackestone bioclástico-intraclástico.
Microfacies 4: Constituye una variedad de las microfacies 2 y 3, pero se caracteriza por la aparición de componentes terrígenos que alcanzan hasta un 15% (Fig. 3d). Es notable la presencia de intraclastos y bioclastos representados por bivalvos de composición aragonítica original, recristalizados, y de calcita con bajo contenido en magnesio (grifeidos), además de gastrópodos (Fig. 4d), placas y espinas de equinodermos, calciesferas, peloides y escasos ooides.
Los peloides aparecen con formas esféricas y bien seleccionados, o bien, irregulares, probablemente originados a partir de la fragmentación de partículas aloquímicas fuertemente micritizadas (Tucker y Wright 1990, Flügel 2004). La matriz aparece recristalizada y muestra por sectores una gradual transición a los componentes peloidales.
La fauna presente y la ausencia de rasgos de exposición subaérea sugieren para esta microfacies un ambiente subtidal. Condiciones de baja energía, por debajo del nivel de olas de buen tiempo, predominaron durante la depositación de los wackestones peloidales. Sin embargo, la presencia de material terrígeno (cuarzo, feldespatos y fragmentos volcánicos) de tamaño fino, subangulares a angulares, probablemente está relacionada a la acción de tormentas que erosionaban depósitos siliciclásticos cercanos. Esta microfacies corresponde a wackestone bioclástico-peloidal-terrígeno.
Microfacies 5: Está compuesta por pequeñas colonias de corales de hasta 8 cm, en posición de vida. Estas colonias presentan un crecimiento cerioide, cálices pequeños y poligonales, asignándose probablemente a Actinastrea sp. (cf. Morsch 1990). El espacio entre las colonias lo rellena un fango micritico, donde se observan fragmentos de corales, bivalvos, equinodermos, peloides y cianobacterias calcáreas.
La mayoría de los corales aparecen con su estructura original obliterada por la destrucción sinsedimentaria producida por organismos perforadores, y por fenómenos de disolución de la microestructura original cuyas cavidades se encuentran rellenadas con micrita peloidal y peloides (Fig. 3e). Generalmente la micrita peloidal rellena las cavidades de disolución, pudiendo aparecer en posición geopetal y antigeopetal. Es frecuente la presencia de envolturas micríticas alrededor de los septos. Los fragmentos de bivalvos y equinodermos localizados en las áreas de intercolonia aparecen con poca abrasión, pero intensamente afectados por bioerosión (Fig. 4e) probablemente causada por bivalvos, serpúlidos o esponjas (Edinger y Risk 1996). Estas cavidades se encuentran también rellenadas por micrita peloidal. El origen de la misma estaría relacionado con la actividad micriobiana en el fondo marino (Chafetz 1986, Sun y Wright 1989).
El grado de fragmentación de la mayor parte de los fósiles y la matriz sugieren un ambiente de depositación somero con turbulencia moderada, en las inmediaciones de pequeños patch reefs. Esta microfacies corresponde a un framestone coralino.
Microfacies 6: Está compuesta por packstones bien estratificados, gris oscuros, en capas que oscilan entre 80 y 100 cm. Se distinguen bivalvos recristalizados, grifeidos, oncoides, y fragmentos de corales escleractínidos. Entre los componentes no esqueletales aparecen intraclastos algales (Fig. 3f) oolitas micritizadas y peloides.
Los bivalvos incluyen, tanto a aquellos originalmente aragoníticos, usualmente disueltos y rellenados con calcita granular, como a los que preservan su estructura filamentosa original (grifeidos). Se reconocen además placas y espinas de equinodermos, algas verdes (dasicladáceas), cianobacterias, gastrópodos y briozoos incrustados por costras microbianas (Fig. 4f).
Las algas verdes son dasicladáceas, identificadas como Cylindroporella annulata Carozzi (Fig. 5c) y por diferentes formas de cianobacterias calcáreas, que incluyen a Cayeuxia (Rivularia) kurdistanensis Elliot (Fig. 5d) y Cayeuxia (Rivularia) piae Rech-Frollo (Figs. 5e, f). Estas últimas dos especies son algas con ramificaciones dicótomas, que fueron mencionadas originalmente como solenoporáceas (Legarreta et al. 1993) pero en realidad corresponden a cianobacterias, de acuerdo al modo de bifurcación y el ángulo entre los filamentos (cf. Dragastan 1985). Desde un punto de vista tafonómico, las algas en esta microfacies como en las otras mencionadas, presentan incrustación (común en las cianobacterias), bioerosión (en dasicladáceas), fragmentación y abrasión (en ambos grupos); y recristalización obliterando sus características (en dasicladáceas).
Los intraclastos son redondeados a subangulares y están compuestos por fragmentos reelaborados de wackestones peloidales-bioclásticos o packstone-grainstones bioclásticosoolíticos, y por intraclastos algales. Se reconocen dos tipos de peloides, aquellos bien redondeados y con buena selección, y otros con formas irregulares y mala selección, que probablemente deben su génesis a la fragmentación de partículas aloquímicas fuertemente micritizadas (Flügel 2004). Corresponden a peloides bioerosionales.
Esta microfacies sugiere un ambiente subtidal influenciado por condiciones de energía moderadas a altas (Hermann 1996). En efecto, se advierten procesos de erosión y retrabajo de depósitos previos localizados en áreas subtidales someras, como lo evidencian los intraclastos. La abundancia de oolitas micríticas y la frecuente micritización en otras partículas sugieren una baja tasa de sedimentación. El notable aumento en el contenido de algas permite suponer una mayor colonización del fondo de la rampa interna. Formas similares de algas dasicladáceas fueron mencionadas para el Jurásico y Cretácico de Europa y Libano (Basson y Edgell 1971, Elliot 1975) e indican un ambiente marino protegido, somero y cálido. La abundancia de Cayeuxia sp., e incrustando con frecuencia oncoides, refleja asimismo un ambiente somero y agitado (Schmid 1996). Esta microfacies corresponde a packstone bioclástico-peloidal-intraclástico.
RAMPA MEDIA
Estos depósitos consisten principalmente de packstones-grainstones oolíticos-bioclásticos, dispuestos en capas ligeramente lenticulares y localmente con estratificación cruzada tabular planar, en espesores que oscilan entre 50 y 90 cm. Constituyen la microfacies 7.
Microfacies 7: Esta microfacies (Fig. 3g) está constituída principalmente por ooides, bioclastos, intraclastos, oncoides y peloides. Los ooides, son de formas esféricas a elípticas, reconociéndose aquellos micríticos, concéntricos y radial-concéntricos, con núcleos de fragmentos esqueletales o peloides y tamaños entre 1 y 3 mm. Las envolturas de los ooides son principalmente concéntricas y están afectadas por una intensa micritización, en tanto que otros ooides aparecen a menudo total o parcialmente disueltos por lo que la porosidad móldica resultante quedó parcialmente rellena por cemento de calcita granular.
Los componentes esqueletales están representados por abundantes fragmentos de bivalvos recristalizados, grifeidos (Fig. 4g), equinodermos (placas y espinas) y oncoides, además de gastrópodos, escasos foraminíferos tipo miliólidos y talos de Cayeuxia (Rivularia) kurdistanensis Elliot (Fig. 5d) y fragmentos escasos de dasicladáceas, probablemente pertenecientes a Heteroporella sp. (Fig. 5b), muchos de ellos con notable abrasión.
Los oncoides varían desde esféricos a elongados, con diámetros entre 2 y 10 mm. Sus núcleos están constituidos por fragmentos redondeados de bivalvos, intraclastos y fragmentos de Cayeuxia sp. Su abundancia asociada a los oncoides responde probablemente a un incremento en la salinidad del agua.
Los intraclastos están redondeados y derivan de las microfacies de wackestone-packstones, y de la erosión de sus propios depósitos. La presencia de ooides e intraclastos coincide con un incremento en el tamaño de partículas. La composición de esta microfacies sugiere una depositación en un ambiente somero y de alta energía. El grado de esfericidad de los ooides, sumado al buen desarrollo de las láminas de sus envolturas, indican que podrían haberse formado en un ambiente continuamente agitado, de forma similar a como ocurre actualmente en ambientes modernos (Loreau y Purser 1973). Asimismo, los oncolitos sugieren una periódica turbulencia en el ambiente depositacional (Tucker y Wright 1990).
Se interpreta a éstos depósitos como barreras oolíticas que ocupaban la rampa media, y cuyo notable desarrollo como la naturaleza de los ciclos resultantes (Palma et al. 2005) responden a un aumento en el espacio de acomodación como consecuencia de un ascenso relativo del nivel del mar que habría favorecido el desarrollo de estas barras. Estas condiciones habrían controlado el cambio en el tipo de facies, incrementando el porcentaje de oolitas respecto a los bioclastos.
La presencia de ooides con estructuras fibro-radial y formando parte de intraclastos, sugiere que derivan de otra microfacies. En efecto, la génesis de intraclastos de origen subtidal, refleja el retrabajo del fondo por acción de tormentas, evidenciadas además por grainstones oolíticos centimétricos con base plana e intercalados con wackestone-packstones, y delgadas intercalaciones de concentraciones de valvas (cf. Aigner 1985). Cabe destacar que las envolturas concéntricas en oolitas de ambientes sedimentarios actuales tienen una composición aragonítica, por lo que siendo este mineral muy inestable, es común que las facies con este tipo de oolitas presenten una porosidad móldica y recristalización más desarrollada.
La abundancia de Cayeuxia sp., como así también su presencia incrustando oncoides reflejan un ambiente somero y agitado (Gebelein 1976, Schmid 1996). Por su parte, los fragmentos de dasicladáceas presentes en esta microfacies se consideran como retrabajados, ya que las mismas caracterizan un ambiente tranquilo y protegido (Wray 1977). Esta microfacies corresponde a packstone grainstone oolítico-bioclástico-peloidal.
ASOCIACIÓN DE PARTÍCULAS Y SUS IMPLICANCIAS AMBIENTALES
Desde hace décadas se sabe que actualmente los carbonatos se acumulan también fuera de las latitudes que engloban el cinturón arrecifal (Less 1975). De esta manera los carbonatos no tropicales o de aguas frías documentados en el registro geológico por Nelson (1988) y Rao (1990) entre otros, se desarrollaron bajo un conjunto de condiciones diferentes a las de los carbonatos tropicales.
Sobre la base de las asociaciones de partículas esqueletales y no esqueletales que caracterizan a los depósitos carbonáticos, los carbonatos tropicales han sido denominados Photozoan (>22ºC); y los no tropicales, Heterozoan (<18ºC; James 1997). Estas asociaciones se encuentran controladas por la temperatura del agua, salinidad, nutrientes, transparencia del agua, concentración de oxígeno y mineralogía original de las partículas, además de la naturaleza del sustrato que requiere la comunidad biótica, que en definitiva influye en la producción carbonática (Pomar et al. 2004, Hallock y Schlager 1986).
Actualmente, la distinción entre depósitos carbonáticos tropicales y no tropicales se manifiesta como consecuencia de un fuerte gradiente latitudinal entre las altas y bajas latitudes. En efecto, la asociación Photozoan se distribuye en un dominio intertropical, en tanto que la asociación Heretozoan se extiende desde los límites de la fábrica tropical (30ºL) hasta latitudes polares (James 1997, Schlager 2000). Sin embargo, esta distribución puede cambiar cuando los carbonatos de aguas frías reemplazan a aquellos tropicales, como consecuencia de la dilución del agua de mar debido a la influencia de aguas dulces (Less 1975).
El análisis de las microfacies de la Formación Calabozo permitió reconocer una asociación de tipo Photozoan (Fig. 6) caracterizada por la presencia, entre las partículas esqueletales, de corales escleractínidos, dasicladáceas, foraminíferos, cianobacterias, bivalvos, equinodermos, gastrópodos y briozoos en menor proporción. En tanto que las partículas no esqueletales están representadas por abundantes oolitas y peloides. Estas últimas partículas se encuentran ausentes en la asociación Heterozoan, al igual que las algas dasicladáceas y los corales escleractínidos. La mineralogía de las partículas aloquímicas en esta asociación está caracterizada por calcita con bajo contenido en magnesio (LMC) al igual que sus contrapartes fósiles.

Figura 6: Principales partículas esqueletales y no esqueletales identificadas en los depósitos de la Formación Calabozo, y sus implicancias ambientales (modificado de Less y Buler 1972, Less 1975, James 1997).
La abundancia de algas dasicladáceas en los depósitos estudiados, consideradas por Okla (1991) y Ott (1967) como típicas de ambiente marino normal, requiere de una comparación con la distribución de las mismas en ambientes actuales (Valet 1979, Berger y Kaever 1992). Las condiciones óptimas para el desarrolllo de estas algas, en la actualidad, están controladas por la temperatura y la naturaleza del sustrato (fijo o móvil). Si bien éste no jugaría un rol fundamental en su distribución (Valet 1979), la temperatura tiene una influencia mayor, ya que las algas dasicladáceas se desarrollan en aguas tropicales a subtropicales, y en consecuencia su tolerancia está limitada alrededor de los 20ºC (Milliman 1974). Rangos de temperatura más bajos inhibirían los procesos de calcificación estrictamente vinculados con la intensidad de la luz.
Por su parte, las variaciones en la composición mineralógica de los componentes esqueletales durante el Fanerozoico dependieron de las oscilaciones de la razón Ca/Mg del agua de mar (Stanley y Hardie 1998), motivo por el cual la temperatura del agua influyó en la mineralogía carbonática de la fauna marina somera. En efecto, aragonita y calcita con alto contenido en magnesio (HMC) dominan en temperaturas mayores de 25ºC, en tanto que HMC y calcita con bajo contenido en magnesio (LMC) ocurren a temperaturas menores a 25ºC (Rao 1993) y solamente ésta última a temperaturas menores a 5ºC (Rao 1981).
De igual forma, la mineralogía de las partículas no esqueletales también está controlada por la geoquímica del agua de mar y la precipitación de aragonita o calcita (Sandberg 1983), por lo que se puede inferir que durante el Fanerozoico hubo mares con diferentes composiciones. El desarrollo de mares calcíticos fue común durante los períodos de greenhouse (Mesozoico), en tanto que los aragoníticos caracterizan períodos de icehouse (Paleozoico Tardío y Cenozoico Tardío) de acuerdo a James (1997). Sobre la base de esta hipótesis, se considera que los carbonatos neríticos de la Formación Calabozo se habrían depositado en un mar calcítico (James 1997), donde fueron comunes los procesos de disolución de partículas originalmente aragoníticas (cf. Palmer et al. 1988).
Teniendo en cuenta las características de las asociaciones de partículas esqueletales y no esqueletales se acepta para la Formación Calabozo, que la producción de carbonatos fue controlada por una asociación de tipo Photozoan, desarrollada bajo condiciones climáticas subtropicales. Los datos paleomagnéticos basados en el promedio de los seis polos del Jurásico Medio a Superior existentes para América del Sur indican una paleolatitud de 31º1´ (Iglesia Llanos et al. 2003). El error paleomagnético permite moverse entre las latitudes de 37º 4´y 25º 6´ (Rapalini, com. pers.).
En función de estas evidencias se puede inferir que, las paleotemperaturas para el intervalo que representa la Formación Calabozo fueron subtropicales. La asociación de partículas esqueletales, principalmente algas verdes y corales, como aquellas no esquetales (ooides y peloides) sugieren un ambiente de aguas someras y cálidas, con un incremento en la productividad de las aguas superficiales, a juzgar por el desarrollo de escasos arrecifes en parches (Esteban 1996). Esta nutrificación (Hallock et al. 1993, Hallock y Schlager 1986) está reflejada en la disminución de los corales y en el incremento de algas, cianobacterias y bioerosión.
DIAGÉNESIS
Los diferentes procesos diagenéticos observados involucran la presencia de micritizacion, micrita peloidal y variedades de calcita que aparece como cemento isopáquico, granular y sintaxial.
La micritización (Fig. 4d) es un proceso muy común en todas las microfacies estudiadas y aparece como una delgada y continua envoltura micritica en la periferia de las partículas esqueletales o bien generando una intensa a completa micritización de ooides (oolitas micriticas; Fig. 4b). La micritización es un proceso que tuvo lugar durante e inmediatamente después de la depositación, y es atribuible a la actividad de algas perforantes u hongos (Bathurst 1975). Simultáneamente se produjo la cementación de una micrita peloidal que aparece rellenado las cavidades de disolución de corales y /o moluscos, en muchos casos desarrollando una estructura geopetal. Esta micrita peloidal estaría relacionada a la actividad microbiana en el lecho marino (Chafetz 1986, Insalaco 1996, Flügel 2004).
La siguiente fase de cementación (Figs. 4c, d, 7a, b, c) está representada por cristales de calcita acicular que crecen en forma perpendicular sobre la superficie de bioclastos y ooides, apareciendo asimismo como un delgado anillo en el interior de las cavidades de disolución de bioclastos de composición aragonítica original y oolitas (oolitas disueltas). Este tipo de cemento es típico de ambiente marino freático (Goldstein 1991). Esta fase de cementación no fue reconocida en todas las microfacies estudiadas, ya que la misma fue disuelta y reemplazada por cristales de calcita con bajo contenido en magnesio (LMC).

Figura 7: Microfotografías obtenidas a microscopía normal (PN) y de cátodoluminiscencia (CL). Escala: 1 mm. a) (PN) cemento acicular (1) y cemento de calcita granular (2); b) (CL) desarrollo de fase no luminiscente (1) y fase con luminiscencia opaca clara (2 a), luminiscencia brillante (2 b) y luminiscencia opaca oscura (2 c). Cementación en fracturas (3). Observar ooides con luminiscencia opaca clara (LOC); c) (PN) fases de cementación con cristales aciculares (1) y de calcita granular (2); d) (CL) cemento acicular no luminiscente (1) y desarrollo de calcita granular con luminiscencia opaca clara (2 a), luminiscencia brillante (2 b) y luminiscencia opaca oscura (2 c). Observar (LOC) en ooides; e) (PN) cemento sintaxial (2) en placa de equinodermo; f) (CL) zonación de la cementación sintaxial que varía desde luminiscencia opaca clara (2 a), luminiscencia brillante (2 b) y luminiscencia opaca oscura (2 c).
La fase de recristalización de este cemento acicular y su transición a aquel de tipo granular pudo haber ocurrido en una fase temprana de diagénesis freática de aguas dulces. Este cemento (Figs. 4a, d; 7a, b, c) está representado por cristales anhedrales a subhedrales, generalmente límpidos, aunque ocasionalmente con algunas inclusiones, a juzgar por su aspecto turbio. Sus límites intercristalinos son planos a rectos y crecen hacia el centro de las cavidades intraparticulares; como así también en los poros interparticulares o bioclastos recristalizados. Este tipo de cemento se forma en la zona freática activa de aguas dulces (Longman 1980). Asociada a la influencia de fluidos porales de esta zona diagenética es común la disolución de organismos originalmente aragoníticos como así también de ooides (oolitas disueltas, Fig. 4b).
Otra fase cementante característica es el cemento sintaxial (Fig. 7e) que se desarrolla sobre fragmentos de equinodermos. Existen diferentes interpretaciones sobre el origen de este tipo de cemento, sin embargo se acepta comúnmente que el mismo puede desarrollarse bajo influencia de diagénesis freática de aguas dulces (Longman 1980). Los efectos diagenéticos producto de soterramiento incluyen fracturación de las partículas (Fig. 7b) (compactación física) y desarrollo de microeslilolitas y contactos interpenetrantes entre las partículas (compactación química; Fig. 7d). La fase de compactación física es posterior a la fase inicial de cemento, y previa a la diagénesis freática de aguas dulces; en tanto que la compactación química corresponde a una fase tardía de soterramiento.
CÁTODOLUMINISCENCIA
Los estudios de cátodoluminiscencia (CL) en las rocas de la Formación Calabozo muestran una compleja historia no revelada a través de estudios de petrografía estandar. Bien es sabido que la luminiscencia en calcita esta influenciada por la presencia de elementos que actúan como activadores o inhibidores (Barbin 2000, Machel 2000). En efecto, el elemento que actúa en la calcita como principal activador es el Mn2+, en tanto que el principal inhibidor es el Fe2+. Sin embargo, tal como reconoce Machel (1985) algunos elementos trazas pueden actuar de igual forma. A los efectos de estandarizar las descripciones de las diferentes fases de cementación, éstas han sido identificadas como: 1) no luminiscente (NL), 2) luminiscencia opaca clara (LOC); 3) luminiscencia amarillenta brillante (LB); y 4) luminiscencia opaca oscura (LOO). Estas diferentes fases de luminiscencia en cementos calcíticos son atribuidas a la composición geoquímica de los fluidos porales, es especial a la razón Fe/Mn, más que la concentración absoluta de estos cationes (Frank et al. 1982, Miller 1986).
Los ooides identificados en la facies de barrera tienen una luminiscencia opaca clara (LOC) (Figs. 7b, f) que podría ser original y relacionada con un microambiente reductor (Weiss y Wilkinson 1988). Sin embargo, esta característica llama la atención ya que los ooides modernos carecen de luminiscencia. A juzgar por la frecuente presencia de oolitas disueltas, se asume que su composición pudo haber sido aragonita o calcita con alto contenido en magnesio (HMC), por lo cual se descarta una luminiscencia primaria, en consecuencia, ésta sería producto de la alteración diagenética en aguas marinas con bajo contenido en oxígeno por debajo de la interfase agua-sedimento (Major et al. 1988). Los análisis realizados en los ooides revelan un contenido de manganeso de 0,08-0,15 wt%, y un contenido en hierro menor de 1 wt%. Las envolturas micríticas tan notables en todas las facies presentan una luminiscencia similar (LOC) y se atribuye un origen similar.
Por su parte, la fase de cementación cristales de calcita con hábito acicular es no luminiscente (Figs. 7b, d) y tiene un porcentaje de manganeso similar a los ooides, pero un incremento en el contenido de Fe2+ que alcanza a 1,8 wt%. Su naturaleza no luminiscente (NL) indica que este cemento podría haberse formado en un ambiente anóxico marino freático (Coleman 1985).
El cemento granular (Figs. 7b, d) presenta una luminiscencia zonada representada por las fases (opaca clara-LOC-2a), 3 (amarilla brillante-LB-2b) y 4 (opaca oscura-LOO-2c). Estos cementos muestran un contenido de Mn2+ que varía entre 0,08-0,18 wt% y el Fe2+ en pocentaje menor al 1,0 wt%. El patrón de luminiscencia zonada se puede interpretar en función de la evolución del potencial de óxido-reducción de los fluidos porales durante etapas progresivas de soterramiento. Esta evolución comenzó con condiciones anóxicas, seguida por una fase subóxica (luminiscencia brillante-LB) y finalmente anóxica en la cual precipita la fase con luminiscencia opaca oscura (LOO), que es la que caracteriza también a la micrita peloidal.
Por su parte, la cementación sintaxial (Fig. 7f) registra una fase de evolución similar pasando de condiciones anóxicas a subóxicas. La fase de luminiscencia brillante sugiere una relación baja Fe/Mn dentro de la zona freática de aguas dulces (Frank et al. 1982, Coleman 1985).
Las fracturas producto de la compactación mecánica (Fig. 7b) están rellenadas con cristales de calcita granular que revelan una zonación similar (fases 2,3,4) y son posteriores a la cementación no luminiscente (NL).
La combinación de estudios de petrografía normal, CL y EDAX de las diferentes fases diagenéticas estudiadas ha permitido reconstruir la secuencia de eventos. Los iniciales involucran una diagénesis marina (micritización, precipitación de micrita peloidal y cemento acicular) concomitante con la sedimentación de la Formación Calabozo en una rampa carbonática.
La interacción de aguas meteóricas con los carbonatos de aguas marinas favoreció la disolución y neomorfismo de las partículas y/o cementos aragoníticos o de calcita con alto contenido en magnesio (HMC). La disolución favoreció el enriquecimiento de las soluciones favoreciendo la precipitación de calcita granular en la porosidad primaria remanente y la posorosidad secundaria de disolución, además de cemento sintaxial. Por último y durante el soterramiento, se desarrollan procesos de compactación mecánica y química.
CONCLUSIONES
El análisis microfacial de los carbonatos de la Formación Calabozo ha permitido reconocer una notable variedad de microfacies depositadas en una rampa carbonática de tipo homoclinal, donde se han reconocido depósitos de rampa interna y media. Las calizas son principalmente wackestones/packstones bioclásticos y grainstones oolíticos. Las partículas esqueletales están representadas por bivalvos, algas dasicladáceas (Salpingoporella annulata, Carozzi, Cylin-droporella annulata Carozzi, y Heteroporella sp.), corales escleractínidos, equinodermos, cianobacterias (Cayeuxia (Rivularia) piae Rech-Frollo y Cayeuxia (Rivularia) kurdistanensis Elliot), oncoides, gastrópodos, foraminíferos (miliólidos), calciesferas y en menor cantidad, briozoos y ostrácodos. Entre las partículas no esqueletales se reconocen abundantes ooides y peloides. El conjunto de partículas aloquímicas sugiere una asociación de tipo Photozoan desarrollada bajo condiciones subtropicales.
Los estudios de petrografía normal, CL y EDAX han permitido caracterizar eventos de diagénesis marina (micritización, precipitación de micrita peloidal y cemento acicular) sinsedimentaria. La interacción de aguas meteóricas favoreció la disolución y neomorfismo de las partículas y/o cementos aragoníticos o de calcita con alto contenido en magnesio (HMC) y la precipitación de calcita granular y cemento sintaxial. Procesos de compactación mecánica y química caracterizan la fase de soterramiento.
TRABAJOS CITADOS EN EL TEXTO
Aigner, T. 1985. Storm depositional systems. Lecture Notes in Earth Sciences. Springer Verlag, 174 p., Berlin. [ Links ]
Armella, C., Cabaleri, N., Cagnoni, M., Ramos, A., y Valencio S. 2005. Paleoambientes de la Formación Calabozo (Caloviano Inferior), en el río Potimalal, Mendoza, Argentina. 16º Congreso Geológico Argentino, Actas 3: 117-124, La Plata. [ Links ]
Barbin, V. 2000. Cathodoluminescence of Carbonate Shells: Biochemical vs Diagenetic process. En Pagel M. Barbin V. Blanc P. y Ohnenstetter D. (eds) Cathodoluminescence in Geosciences. Springer-Verlag, 303-330. Heidelberg. [ Links ]
Bathurst, R.G.C. 1975. Carbonate sediments and their diagenesis. Elsevier, 658 p., Amsterdam. [ Links ]
Basson, P.W. y Edgell, H.S. 1971. Calcareous algae from the Jurassic and Cretaceous of Lebanon. Micropaleontology 17 (4): 411-433. [ Links ]
Berger, S. y Kaever, M.J. 1992. Dasycladales. An illustrated monograf of a fascinating algal order. 247 p. Stuttgart. [ Links ]
Brookfield, M.E. y Brett. C.E. 1988. Paleoenvironments of the Mid-Ordovician (Upper Caradocian) Trenton limestones of southern Ontario, Canada: storm sedimentation on a shoal-basin shelf model. Sedimentary Geology 57: 75-105. [ Links ]
Chafetz, H.S. 1986. Marine peloids: a product of bacterially induced precipitation of calcite. Journal of Sedimentary Petrology 56 (3): 812-817. [ Links ]
Coleman, M.L. 1985. Geochemistry of diagenetic non-silicicate minerals: kinetic considerations. Phil. Transational Royal Society of London, A 315: 39-56. [ Links ]
Dragastan, O. 1985. Review of Tethyan Mesozoic Algae of Romania. En Toomey, D.F. y Nitecki, M.A. (eds.) Paleoalgology, Springer Verlag, 101-161 p. Berlín. [ Links ]
Dellapé, D.A., Mombrú, C., Riccardi, C.A., Uliana, M.A. y Westermann, G.E.G. 1979. Edad y correlación de la Formación Tábanos en Chacay Melehue y otras localidades de Neuquén y Mendoza. Con consideraciones sobre la distribución y el significado de las sedimentitas Lotenianas. Museo La Plata, Obra Centenario 5: 81-105. [ Links ]
Edinger, E.N. y Risk, M.J. 1996. Sponge borehole size as a relative measure of bioerosion and paleoproductivity. Lethaia 29: 275-285. [ Links ]
Elliot, G. F. 1975. Transported algae as indicators of different marine habitats in the english Middle Jurassic: Paleontology, 18: 351-366. [ Links ]
Esteban, M. 1996. An overview of Miocene reefs from Mediterranean areas: general trends and facies models. En Fransen, E.K. Esteban, M. Ward, W.C. y Rouchy, J.M. (eds.) Models for Carbonate Stratigraphy from Miocene Reef Complexes of Mediterranean Regions. Society of Economic Paleontologists and Mineralogists (SEPM) Concepts Sedimentology, Paleontology 5: 3-53. [ Links ]
Frank, J.R. Carpenter, A.B. y Oglesby, T.W. 1982. Cathodoluminescence and composition of calcite cement in the Taum Sauk limestone (Upper Cambrian), Southeast Missouri. Journal of Sedimentary Petrology 52: 631-638. [ Links ]
Flügel, E. 2004. Microfacies analysis of limestones. Springer Verlag, 976p., Berlin. [ Links ]
Gebelein, C.D. 1976. Open marine subtidal and intertidal stromatolites (Florida, The Bahamas and Bermuda). En Walker, M.R. (ed.) Stromatolites, Springer-Verlag, 381-388 p., Amsterdam. [ Links ]
Goldstein, R.H. 1991. Practical aspects of cement stratigraphy with illustrations from Pennsylvanian limestone and sandstones, New Mexico and Kansas. Society of Economic Paleontologists and Mineralogists (SEPM) Short Course 25:123-132, Kansas. [ Links ]
Gulisano, C.A., Gutiérrez Pleimling, A. y Digregorio, R.E. 1984a. Esquema estratigráfico de la secuencia jurásica del oeste de la provincia de Neuquén. 9º Congreso Geológico Argentino, Actas 1: 236-259, San Carlos de Bariloche. [ Links ]
Gulisano, C.A. Gutiérrez Pleimling, A. y Digregorio, R.E. 1984b. Análisis estratigráfico del intervalo Tithoniano-Valanginiano (Formaciones Vaca Muerta, Quintuco y Mulichinco) en el suroeste de la provincia del Neuquén. 9º Congreso Geológico Argentino, Actas 1: 221 -235, San Carlos de Bariloche. [ Links ]
Hermann, R. 1996. Entwicklung einer oberjurassichen karbonatplattform: Biofazies, Riffe und sedimentologie im Oxfordium der Zentralen Dobrogea (Ost Rumänien). Berliner Geowissenschaften, Abhandlung E 19: 1-101. [ Links ]
Hallock, P., Müller Karger, F.E. y Halas, J.C. 1993. Coral reef decline-anthropogenic nutrients and degradation of western Atlantic and Caribbean coral reefs. Research Exploration, 9: 358-378. [ Links ]
Hallock, P. y Schlager, W. 1986. Nutrient excess and the demise of coral reefs and carbonate platforms. Palaios 1: 389-398. [ Links ]
Iglesia Llanos, M., Leanza, R., Riccardi, A.C., Geuna, S., Laurenzi, M.A. y Ruffini, R. 2003. Paleomagnetic study of the El Quemado Complex and Marifil Formation, Patagonian Jurassic igneous province, Argentina. Geophysical Journal International 154: 599-617. [ Links ]
Insalaco, E. 1996. Upper Jurassic microsolenid biostromes of norther and central Europe: facies and depositional environment. Palaeogeography, Palaeoclimatology, Palaeoecology 121: 169-194. [ Links ]
James, N.P. 1997. The cool-water carbonate depositional realm. En James, N.P. y Charke, J.A.D. (eds.) Cool-water carbonates. Society Economists Paleontologists Mineralogists, Special Publication 6:1-20, Tulsa. [ Links ]
Legarreta, L. y Gulisano, C.A. 1989. Análisis estratigráfico secuencial de la Cuenca Neuquina (Triásico superior - Terciario inferior), Argentina. En Chebli, G.A. y Spalleti, L. A. (eds.) Cuencas Sedimentarias Argentinas, Serie Correlación Geológica 66: 221-243. [ Links ]
Legarreta, L. Gulisano, C.A. y Uliana, M.A. 1993. Las secuencias sedimentarias jurásicas-cretácicas. En Ramos, V.A. (ed.) Actas 12º Congreso Geológico Argentino y 2º Congreso de Exploración de Hidrocarburos. Geología y Recursos Naturales de Mendoza, Relatorio 9: 87-114. [ Links ]
Less, A. 1975. Possible influences of salinity and temperature on modern shelf carbonate sedimentation. Marine Geology 19:159-198. [ Links ]
Les, A. y Buller, A.T. 1972. Modern temperate water and warm water shelf carbonate sediments contrasted. Marine Geology 13:1767-1773. [ Links ]
Loreau, J.P. y Purser, B.H. 1973. Distribution and ultrastructure of Holocene ooids in the Persian Gulf. En Purser, B.H. (ed.) The Persian Gulf, 279-328, Berlin. [ Links ]
Longman, M.W. 1980. Carbonate diagenesis textures from near surface diagenetic environments. American Association Petroleum Geologists, Bulletin 64: 461-487. [ Links ]
Machel, H.G. 1985. Cathodoluminescence in calcite and dolomite and its chemical interpretation. Geoscience Canada 12: 139-147. [ Links ]
Machel, H.G. 2000. Application of Cathodoluminescence to Carbonate Diagenesis En Pagel M, Barbin V, Blanc P, Ohnenstetter D (eds.) Cathodoluminescence in Geosciences. Springer-Verlag, 271-302, Heidelberg. [ Links ]
Major, R.P., Halley, R.B. y Lukas, K.J. 1988. Cathodoluminescent bimineralic ooids from the Pleistocene of the Florida continental shelf. Sedimentology 35: 843-855. [ Links ]
Maliva, R.G. y Dickson, J.A.D. 1992. The mechanism of skeletal aragonite neomorphism: evidence from neomorpfosed molluscs from the upper Purbeck Formation (Late Jurassic-Early Cretaceous) Southern England. Sedimentary Geology 76: 221-232. [ Links ]
Maliva, R.G. y Siever, R. 1988. Mechanism and controls of silicification of fossils in limestones. Journal of Geology 96: 387-398. [ Links ]
Marszalek, D.S. 1975. Calcisphere ultrastructure and skeletal aragonite from the alga Acetabularia antillana. Journal of Sedimentary Petrology 45: 266-271. [ Links ]
Miller, J. 1986. Facies relationships and diagenesis in Waulsortion mud mounds from the lower Carboniferous of Ireland and England. En Schroeder, J.H. y Purser, B.H. (eds.) Reef Diagenesis, Springer-Verlag, 311-335, Berlin. [ Links ]
Milliman, J.D. 1974. Marine carbonates. Springer-Verlag, 375 p., New York. [ Links ]
Morsch, S.M. 1990. Corales (Scleractinia) de la extremidad sur de la Sierra de la Vaca Muerta, Formación La Manga (Oxfordiano), Provincia del Neuquén, Argentina. Ameghiana 27(1-2): 19-28. [ Links ]
Nelson, B.S. 1988. An introductory perspective on non-tropical shelf carbonates. Sedimentary Geology 60: 2-12. [ Links ]
Okla, S.M. 1991. Dasycladacean algae from the Jurassic and Cretaceous of central Saudi Arabi. Micropaleontology 37(2): 183-190. [ Links ]
Ott, E. 1967. Segmentierte Kalschwämme (Sphinctozoa) aus der Alpinen Mittelrais und ihre Bedeutung als Riffbilder im Wettersteinkalk. Bayerische Akademische Wissenschafter, Abhandlungen 131:96 p. [ Links ]
Palma, R.M., Lo Forte, G.L. y Lanes, S.G. 2000. Ciclos depositacionales en la Formación Calabozo (Caloviano temprano), Cuenca Neuquina, Mendoza, Argentina. 2º Congreso Latinoamericano de Sedimentología y 8º Reunión Argentina de Sedimentología, Actas 138-139, Mar del Plata. [ Links ]
Palma, R.M., Lo Forte, G.L., Mehdli, M., Piethé, R.D. y Lanés, S. 2005. Depositional cycles of the Callovian Calabozo Formation, Neuquén Basin, Argentina: a preliminary approach. En Riccardi, A.C. (ed.) New insights into the Jurassic record of South America (Andes and Patagonia), Geologica Acta 3:119-132, Barcelona. [ Links ]
Palmer, T.J. Hudson, J.D. y Wilson, M.A. 1988. Palaeoecological evidence for early aragonite dissolution in ancient calcite seas. Nature 335: 809-810. [ Links ]
Pomar, L., Brandano, M. y Westphal, H. 2004. Environmental factors influencing skeletal grain sediment associations: a critical review of Miocene examples from the western Mediterranean. Sedimentology 51: 627-651. [ Links ]
Rao, C.P. 1981. Criteria for recognition of coldwater carbonate sedimentation: Berriedale Limestone (Lower Permian), Tasmania, Australia. Journal of Sedimentary Petrology 51: 591-506. [ Links ]
Rao, C.P. 1990. Geochemical characteristics of cool-temperate carbonates, Tasmania, Australia. Carbonates and Evaporites 5: 209-221. [ Links ]
Rao, C.P. 1993. Carbonate minerals, oxygen and carbon isotopes in modern temperate bryozoa, eastern Tasmania, Australia. Sedimentary Geology 88: 123-135. [ Links ]
Reid, R.P. y Macintyre, I.G. 2000. Microboring versus recrystallization: Further insight into the micritization process. Journal of Sedimentary Petrology 70: 24-28. [ Links ]
Riccardi, A.C., Leanza, H.A., Damborenea, S.E., Manceñido, M.O., Ballent, S.C. y Zeiis, A. 2000. Marine Mesozoic biostratigraphy of the Neuquen Basin. En Miller, H. y Hervé, F. (cords.) Zeitschrift für Angewandte Geologie. Sonderhelft SH1. 31st. International Geological Congress, Proceedings: 103-108, Rio de Janeiro. [ Links ]
Sandberg, P.A. 1983. An oscillating trend in Phanerozoic non-skeletal carbonate mineralogy. Nature 305: 19-22. [ Links ]
Schlager, W. 2000. Sedimentation rates and growth potential of tropical, cold-water and mud-mound carbonate systems. En Insalaco, E. Skelton, P.W. y Palmer, T.J. (eds.) Carbonate Platform Systems: Components and interactions. Geological Society of London, Special Publication 178: 217.227. [ Links ]
Schmid, D.V. 1996. Marine Mikrobiolitithe und Mikroinkrustierter aus dem Oberjura. Profil 9: 101-251, Stuttgart. [ Links ]
Shinn, F.A. 1968. Practical significance of birdeye structure in carbonate rocks. Journal of Sedimentary Petrology 3: 215-223. [ Links ]
Stanley, S.M. y Hardie, L.A. 1998. Secular oscillations in the carbonate mineralogy of reefbuilding and sediment-producing organisms driven by tectonically forced shifts in seawater chemistry. Palaeogeography, Palaeoclimatology, Palaeoecology 144: 3-19, Amsterdam. [ Links ]
Strasser, A. 1986. Ooids in Purbeck Limestones (lowermost Cretaceous) of the Swiss and French Jura. Sedimentology 33: 711-728. [ Links ]
Sun, S.G. y Wright, V.P. 1989. Peloidal fabrics in the Upper Jurassic reefal limestones, Weal Basin, Southern England. Sedimentary Geology 65:165-181. [ Links ]
Tucker, M. y Wright, P. 1990. Carbonate Sedimentology. Blackwell Scientific Publications, 482 p., London. [ Links ]
Valet, G. 1979. Approche paléoécologique du monde des Dasycladales á partir d' écologie des formes actuelles. Bulletin Centre Research Elf-Aquitaine Exploration-Production, 3:859-866, París. [ Links ]
Weiss, C.P. y Wilkinson, B. 1988. Holocene cementation along the central Texas coast. Journal of Sedimentary Petrology 58: 468-478. [ Links ]
Wray, J.L. 1977. Calcareous Algae. Development in Palaeontology and Stratigraphy, Elsevier, 4: 185 p., Amsterdam. [ Links ]
Recibido: 2 de febrero, 2006
Aceptado: 1 de enero, 2007

