










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de la Asociación Geológica Argentina
versión impresa ISSN 0004-4822versión On-line ISSN 1851-8249
Rev. Asoc. Geol. Argent. v.62 n.4 Buenos Aires oct./dic. 2007
Los depósitos diamictíticos cenozoicos de la cuenca James Ross, Antártida: una síntesis estratigráfica y nuevos hallazgos paleontológicos
Concheyro, A.1,2,3, Salani, F.M.2, Adamonis, S.3 y Lirio, J.M.1
1 Instituto Antártico Argentino. Instituto Antártico Argentino, Cerrito 1248, (C1010AAZ)
2 Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET)
3 Laboratorio de Bioestratigrafía de Alta Resolución. Departamento de Ciencias Geológicas, Universidad de Buenos Aires, Ciudad Universitaria, 1428, Buenos Aires.
RESUMEN
Se analizan los depósitos glaciarios cenozoicos de la cuenca James Ross, Antártida, se los caracteriza estratigráficamente, se precisan las localidades donde afloran y se dan a conocer nuevos hallazgos paleontológicos en diamictitas del sudeste de la isla James Ross, particularmente en cabo Hamilton. En esta región, las diamictitas son portadoras de una interesante megafauna y microfauna. La megafauna incluye el bivalvo Zygochlamys andersonni, braquiópodos, balánidos, serpúlidos y una rica asociación de briozoos compuesta por Cellarinella, Hornera, Membranicellaria, Melicerita, Microporella y Polirhabdotos y alguno de los cuales constituyen los primeros registros conocidos para el Mioceno Tardío de la Antártida. La edad de los briozoos es corroborada con foraminíferos y ostrácodos hallados en la misma localidad. La microfauna recuperada permite inferir condiciones de salinidad marina normal y temperaturas superiores a las actuales en las mismas latitudes e influencia de aguas frías.
Palabras clave: Diamictitas. Paleontología. Cenozoico. Cuenca James Ross. Antártida.
ABSTRACT
The Cenozoic diamictitic deposits of the James Ross Basin, Antarctica: A stratigraphic synthesis and new paleontological findings. The Cenozoic glaciar deposits from the James Ross Basin, Antarctica, are here studied. They are stratigraphically characterized, their localities and outcrops are mentioned, and new paleontological discoveries are presented from the Hamilton Cape, southeast James Ross Island. In this region, diamictites are fossilliferous in megafauna and microfauna. The megafauna contains Zygochlamys andersonni, brachiopods, balanids, serpulids and a rich bryozoan association, composed of Cellarinella, Hornera, Membranicellaria, Melicerita, Microporella y Polirhabdotos. Some of those genera constitute their first fossil record defined for the Late Miocene of Antarctica. The age is confirmed by foraminifera and ostracods found in the same locality. The microfauna allow establishing normal marine salinity conditions, higher temperatures than the present ones at the same latitude, and cold water influences.
Keywords: Diamictite. Paleontology. Cenozoic. James Ross Basin. Antarctica.
INTRODUCCIÓN
Los depósitos diamictíticos de Antártida revisten un especial interés dado que documentan el registro de las glaciaciones cenozoicas y permiten estudiar el comportamiento y evolución de su extensa calota glaciaria.
En el sector oriental de Antártida, en los montes Transantárticos y en Antártida occidental es posible reconocer depósitos glaciarios cenozoicos. En los primeros se destaca la Formación Sörsdal, con una edad de 4,1- 4,5 Ma (Lirio et al. 2003).
En la región de los montes Transantárticos se reconocen las Formaciones Prospect, Mesa Gravel, Scallop Hill y Taylor, con edades de 3 Ma, 2,6 Ma y 6.670 años AP, respectivamente (Lirio et al. 2003). En la región de los glaciares Beardmore, Darwin y los Dry Valley se concentra un gran número de depósitos glaciarios, algunos de ellos de difícil correlación, e incluyen al Grupo Sirius, con edades pliocenas que varían entre los 3,8 Ma (Ackert y Kurz 2004) y los 3,1 Ma a 2,5 Ma (Moriwaki et al. 1992).
La Antártida occidental puede ser dividida en dos sectores, el noroccidental de las islas Shetland del Sur y el sector sudoriental, representado por la península Antártica y la cuenca James Ross.
Los depósitos glaciarios de las islas Shetland del Sur están representados por las sedimentitas oligocenas de la Formación Polonez Cove (32-26 Ma) y las sedimentitas miocenas de la Formación Cape Melville (23-20 Ma) estudiadas por Birkenmajer et al. (2005).
Las diamictitas reconocidas al sur de la península Antártica, en la cuenca James Ross, han sido descriptas desde principio del siglo XX (Andersson 1906). Si bien, con posterioridad estos depósitos han sido tratados parcialmente por otros autores como Bibby (1966), Harwood (1986), del Valle et al. (1987a, b), Marenssi et al. (1987), Strelin et al. (1987, 1993, 1999), Gazdzicki y Webb (1996), Dingle et al. (1997), Lirio y del Valle (1999), Pirrie et al. (1997a), Jonkers (1998a, b), Jonkers y Kelley (1998), Bertels-Psotka et al. (2001), Jonkers et al. (2002), Lirio et al. (2003), Concheyro et al. (2005), la información de detalle es aún muy escasa. Los depósitos más conspicuos se encuentran en las islas James Ross, Cockburn, Marambio y Vega (Fig. 1).
Figura 1: Mapa de localización de la cuenca James Ross, Antártida. Principales localidades donde afloran diamictitas cenozoicas.

Como resultado de estas investigaciones y basándose en la megafauna, microfauna de foraminíferos y dataciones isotópicas 40K/39Ar, 40Ar/39Ar de rocas volcánicas y relaciones 87Sr/86Sr en conchillas de pectínidos, se postulan para la Antártida occidental al menos tres pulsos glaciarios situados en el Oligoceno Tardío, Mioceno Tardío y Plioceno Tardío (Birkenmajer y Gazdzicki 1986. Dingle et al. 1997, Jonkers 1998b, Jonkers et al. 2002).
El presente trabajo tiene por objeto efectuar una revisión de los depósitos glaciarios cenozoicos en el ámbito de la cuenca James Ross y dar a conocer nuevos hallazgos paleontológicos en diamictitas del sudeste de la Isla James Ross, particularmente en cabo Hamilton.
SÍNTESIS CRONOESTRATIGRÁFICA
MARCO GEOLÓGICO
La cuenca James Ross, situada en el sector nordeste de la península Antártica, entre los 65°S y 74°O, está caracterizada por un conjunto potente de sedimentitas depositadas en una cuenca de retroarco (Fig. 1). Ha sido rellenada con sedimentos clásticos deltaicos, de estuario, plataforma y talud, con edades jurásico tardías- paleógenas (Rinaldi et al. 1978, Rinaldi 1982, Olivero et al. 1986, Ineson et al. 1986, Lirio et al., 1989, Marenssi et al. 1991, 1998 y 2001, Medina et al. 1992, Pirrie et al. 1991, 1997b, Kiessling y Scasso 1996).
La cuenca posee la secuencia sedimentaria marina más completa y espesa descripta para el Cretácico de altas latitudes del hemisferio sur, que es cubierta en discordancia por vulcanitas y piroclastitas de composiciones basálticas del Grupo Volcánico Isla James Ross, de edad miocena-holocena (Adie 1953, Nelson 1975, Rex 1976, Pirrie y Sykes 1988, Sykes 1988, Smellie et al. 1998, Strelin 1990, Strelin et al. 1993, López 1994, Jonkers 1998a, b, De Angelis 1999, Carrizo et al. 1999, Marenssi et al. 2001).
Los depósitos glaciarios objeto de estudio se encuentran en general en la base de cada pulso eruptivo del Grupo Volcánico Isla James Ross.
UNIDADES LITOESTRATIGRÁFICAS
A continuación se incluyen los depósitos diamictíticos cenozoicos de la cuenca James Ross que figuran formalmente en la literatura y otros denominados informalmente, cuya edad y origen se precisan.
ISLA JAMES ROSS
a) Formación Hobbs Glacier (Pirrie et al.1997a)
La Formación Hobbs Glacier representa los depósitos glacigénicos más antiguos del Grupo Volcánico Isla James Ross reconocidos hasta el momento. Han sido mencionados informalmente por Andersson (1906), Bibby (1966) y Nelson (1975) y luego reexaminados por Marenssi, et al. (1987), Sykes (1988), Lirio et al. (1989), Strelin et al. (1999) y Bertels-Psotka et al. (2001).
Pirrie et al. (1997a) elevan este conjunto a la categoría formacional. Aflora en el sudeste de la isla James Ross, se apoya en discordancia erosiva sobre sedimentitas de las Formaciones Rabot (Lirio et al. 1989) o Snow Hill (Pirrie et al. 1997b) e infrayace las rocas volcánicas del Grupo Volcánico Isla James Ross.
Esta unidad, situada en la base del Grupo Volcánico Isla James Ross, comprende un sector inferior diamictítico y uno superior tobáceo con una potencia de cuatro metros en su localidad tipo, punta Rabot (Figs. 2 y 3). Estas sedimentitas han sido correlacionadas con las de otras áreas de las islas James Ross y Vega.
Figura 2: Perfiles estudiados de la Formación Hobbs Glaciar en el sudeste de la Isla James Ross en cabo Hamilton, su comparación con secciones de punta Rabot y punta Redonda.
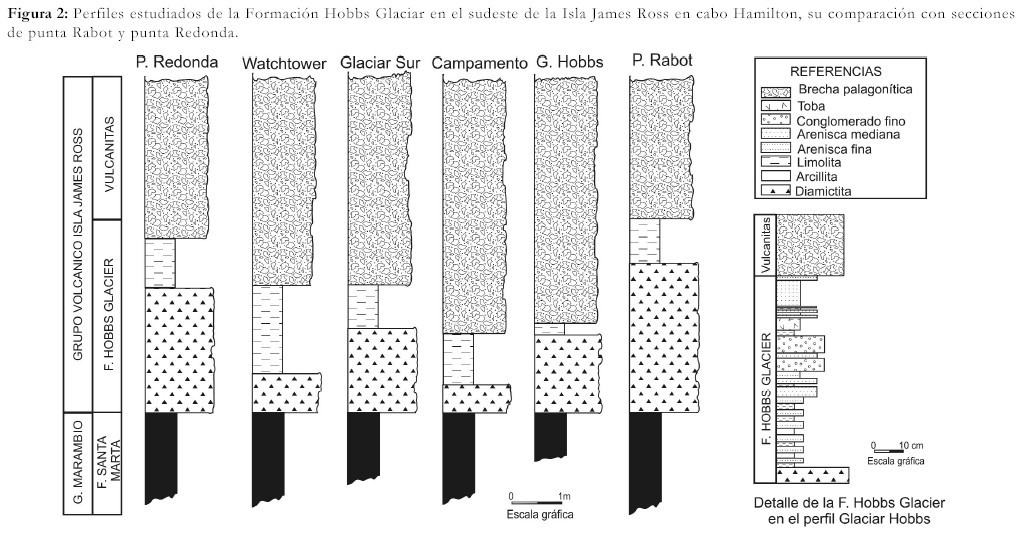
Figura 3: Principales afloramientos de la Formación Hobbs Glacier en diferentes localidades del SE de la isla James Ross e isla Vega. a) Vista general de la diamictita en punta Rabot, isla James Ross. b) Base de la diamictita en punta Rabot. c) Clastos dispersos de basaltos en la diamictita, punta Rabot. d). Estratificación de la sección superior tobácea de la Formación Hobbs Glacier en punta Rabot, isla James Ross. e) Diamictita basal con muy mala selección, perfil Glaciar Hobbs, cabo Hamilton. f) Estratificación de la sección superior tobácea de la Formación Hobbs Glacier en el perfil Glaciar Hobbs, cabo Hamilton. g) Vista general de la Formación Hobbs Glacier en punta Redonda, isla James Ross. h) Vista general de la diamictita basal en el sector de riscos Leal, cabo Lamb, isla Vega, los círculos indican la posición de uno de los niveles diamictíticos.

Se diferencian dos facies sedimentarias: una diamictita masiva sin estructura y otra laminada con intercalaciones de areniscas con estratificación entrecruzada y geometría lenticular. Hacia el tope de la formación predominan rocas piroclásticas, tobas arenosas, conglomerados tobáceos y limos atribuidos a cuatro facies sedimentarias (Pirrie et al. 1997a).
Estos depósitos son interpretados como depositación glacimarina cercana a una línea de costa, o por acción de un glaciar que desemboca al mar. Las estructuras sedimentarias y los clastos de composición plutónica y metamórfica sugieren que fueron transportados por un glaciar que fluía desde la península Antártica hacia el sudeste (Pirrie et al. 1997a).
La Formación Hobbs Glacier ha sido asignada al Mioceno Tardío sobre la base de los megafósiles (Pirrie et al. 1997a). Dingle y Lavelle (1998) utilizando estratigrafía de isótopos de Sr en conchillas de cirripedios recuperadas en punta Rabot, determinan una edad de 9,9 ± 0,97 Ma, Tortoniano (Mioceno Tardío).
En las localidades de cabo Hamilton y punta Redonda (Fig. 1), se reconocen depósitos glacimarinos ubicados en una posición estatigráfica similar a los de la localidad tipo de la Formación Hobbs Glacier; son descriptos y asignados por sus foraminíferos y diatomeas al Mioceno superior y Mioceno en sentido amplio, respectivamente (Bertels- Psotka et al. 2001, Marenssi et al. 1987).
b) Formación Belén (Lirio et al. 2003)
Del Valle et al. (1987a, b), Lirio y del Valle (1999) y Jonkers et al. (2002) hacen referencia a conglomerados con fósiles marinos que se apoyan en discordancia sobre sedimentitas cretácicas, sin otorgarle una denominación formal.
Posteriormente, Lirio et al. (2003) denominan Formación Belén a los conglomerados que forman un depósito diamíctitico, interestratificado con rocas volcánicas del Grupo Volcánico Isla James Ross. Aflora en la margen sur del fiordo Belén, en proximidades del cerro Terrapin y la península El Morro (The Naze), donde se halla el perfil y localidad tipo de la formación. Se encuentra en una posición estratigráfica ligeramente superior a la Formación Hobbs Glacier (Mioceno Tardío). Comprende numerosos bancos conglomerádicos granodecrecientes, con clastos subredondeados de hialobasaltos olivínicos y brechas palagoníticas del Grupo Volcánico Isla James Ross, escasos clastos de metamorfítas y areniscas cretácicas. La litología corresponde a conglomerados matriz-soportados, con matriz arena gruesa semiconsolidada y pobre selección. En la fracción conglomerádica el diámetro de los clastos mayores supera los 70 centímetros.
El depósito posee geometría lenticular, con un espesor de 5 m y un desarrollo lateral de aproximadamente 100 metros. El contacto basal es subhorizontal.
Sobre la costa este del fiordo, la formación está representada por un conjunto de estratos de 3 m de espesor máximo, en cuya parte superior se destacan bioclastos integrados por fragmentos de valvas de pectínidos (del Valle et al. 1987b). Posteriormente Jonkers (1998a, b) y Jonkers et al. (2002) determinaron que las valvas desarticuladas podrían pertenecer a la especie Zygochlamys anderssoni Henning.
Los foraminíferos de la Formación Belén son cassidulínidos en su mayoría, siendo Cassidulina crassa d'Orbigny el taxón dominante. Entre los foraminíferos planctónicos, la presencia de Globigerinita glutinata (Egger), permite inferir una edad no más antigua que miocena media terminal-miocena tardía.
Los análisis isotópicos de 87Sr/86Sr en conchillas de Zygochlamys anderssoni Henning indican una edad de 6,8 +1,3/-0,5 Ma, miocena tardía (Jonkers 1998b, Lirio et al. 2003). Un clasto de basalto incluído en la diamictita, proveniente de unidades inferiores del GVIJR y datado por el método 40Ar/39Ar (Jonkers et al. 2002 y Lirio et al. 2003), arroja una edad de 9,2 ± 0,5 Ma (Mioceno Tardío).
c) Formación Gage (Lirio et al. 2003)
Lirio et al. (2003) proponen la denominación de Formación Gage para los depósitos conglomerádicos que afloran entre cabo Gage y punta Ekelöf, al NE de la isla James Ross, anteriormente descriptos y datados por Jonkers et al. (2002).
Las mejores exposiciones se encuentran en los acantilados de la costa sur entre ambas localidades. La litología comprende una sección inferior formada por limolitas y areniscas y una sección superior compuesta por areniscas con estratificación entrecruzada, limolitas bioturbadas y conglomerados matriz- soportados. Los depósitos contienen abundante fauna de pectínidos, foraminíferos y ostrácodos (Jonkers et al. 2002), compuesta por Zygochlamys anderssoni Henning, Ammoelphidiella antarctica Conato y Sagre, Cytheropteron sp. nov. Jonkers, Patagonacythere devexa (Müller), Australicythere polylyca (Müller), que sugieren un paleoambiente de plataforma.
La formación se dispone en discordancia erosiva sobre las rocas cretácicas de la Formación López de Bertodano (Rinaldi et al. 1978) e infrayace las rocas del Grupo Volcánico Isla James Ross.
La presencia de Ammoelphidiella antarctica Conato y Sagre, un foraminífero exclusivo del Plioceno de Antártida, permite asignar una edad pliocena a la Formación Gage. Esta edad queda acotada por la datación por 40Ar/39Ar de 3,1 Ma (Plioceno Tardío) de un clasto basáltico incluído en estos depósitos (Jonkers et al. 2002) y la presencia de Zygochlamys anderssoni Henning, que desaparece de las aguas antárticas a los 2,4 Ma (Jonkers 1998b).
d) Formación Terrapin (Lirio et al. 2003)
En la costa sudoeste del fiordo Belén aflora un depósito diamictítico de 50 m de espesor, poco consolidado, compuesto por conglomerados de color verde oscuro, clasto- sostén y pobre selección. Presenta bloques angulosos, gravas de basaltos y brechas palagoníticas, en una matriz de arena gravosa gruesa a fina. Los bloques mayores (que alcanzan los 3 m de diámetro) se hallan en la base y otorgan al conjunto el aspecto de gradación normal; los 5 m superiores son portadores de fragmentos de bivalvos (pectínidos).
La Formación Terrapin apoya sobre una brecha palagonítica y está cubierta por tobas palagoníticas correspondientes a otro evento explosivo del Grupo Volcánico Isla James Ross (Lirio et al., 2003).
Los foraminíferos hallados pertenecen a las familias Cassidulinidae, Fursenkoinidae, que conjuntamente con los ostrácodos, sugieren un ambiente marino poco profundo y próximo al frente glaciario. Es frecuente el aporte de microfauna retrabajada del Cretácico superior, proveniente de sedimentitas del Grupo Marambio.
Los bivalvos están representados por Adamussium colbecki Smith y su datación con isótopos 87Sr/86Sr permite asignar a esta formación una edad de 1,95±0,52 Ma, Plioceno Tardío - Pleistoceno Temprano (Lirio et al. 2003).
La muy pobre selección del depósito, la estratificación masiva, la angulosidad y tamaño de los clastos reflejan una fuerte influencia glaciaria también sugerida por concreciones cretácicas con estrías glaciarias que aparecen en la base de la Formación.
e) Drift Refugio San Carlos
Con esta denominación Rabassa (1982) describe un depósito situado al oeste sudoeste del refugio San Carlos, en la bahía Brandy, constituido por un till de ablación que posee grandes bloques erráticos, de litología granítica y metamórfica, algunos con sus caras pulidas y estriadas, inmersos en una matriz arenosa y suelta.
Lirio et al. (2003) analizan este depósito, que presenta una escasa distribución areal, forma una pequeña meseta de baja altura y se apoya sobre las rocas sedimentarias cretácicas de la Formación Hidden Lake. Está compuesto por un conglomerado fino, suelto, con clastos redondeados, portador de gastrópodos y bivalvos, que es sucedido por una arenisca friable con abundantes valvas de Zygochlamys andersonni Henning. El depósito culmina con una diamictita de un metro de espesor, integrada por bloques volcánicos (90 %), metamórficos y plutónicos. El espesor de los estratos no supera los tres metros.
Para la sección basal del depósito, Lirio et al. (2003) interpretan un ambiente marino de playa y de tipo glaciario para el tope de la secuencia. Algunas conchillas de Zygochlamys andersonni Henning fueron datadas mediante el método isotópico 87Sr/86Sr y arrojaron una edad de 3,17-4,60 Ma (Kosler, com.verb. 2005).
f) Depósitos de cabo Lachman
En el noroeste de la isla James Ross, en cabo Lachman y alrededores, se han distinguido tres depósitos de origen glaciar y composición similar, con edades que varían del Plioceno al Pleistoceno. Estos afloran en: a) la costa occidental de cabo Lachman; b) norte del cabo y c) la meseta Lachman.
El depósito diamictítico de la costa occidental de cabo Lachman posee un espesor de 50-70 m y muestra una continuidad lateral de 4 km (Nelson 1975). Compuesto por un conglomerado matriz-sostén indurado en su base, más arenoso y friable hacia el techo, con fragmentos de pectínidos, que son abundantes en los sectores superiores y permiten establecer una edad pliocena o más joven. Suprayace a las rocas volcánicas del Grupo Volcánico Isla James Ross.
En el sector norte de cabo Lachman, cubriendo a los depósitos glaciarios anteriores, se dispone una diamictita polimíctica con grandes clastos de rocas metamórficas y plutónicas provenientes de la península Antártica. Los clastos alcanzan tamaños de varios metros de diámetro y algunos de ellos son encontrados a más de 100 m. de altura. Esta diamictita suprayace a los depósitos de cabo Lachman y localmente a las rocas volcánicas del extremo norte de cabo Lachman. Se infiere que podrían tener una edad pliocena.
El drift de la meseta Lachman ha sido descripto por Rabassa (1982). Está representado por grandes bloques erráticos de litología volcánica (local) y granítico metamórfica (exótica). Los bloques provenientes de la península Antártica poseen algunas de sus caras pulidas y estriadas; se disponen en forma aislada y apoyan directamente sobre el GVIJR. Su edad es incierta, aunque Rabassa (1982, 1987), sugiere una edad pleistocena temprana (?)-pleistocena media. Este depósito, su origen y edad es discutido por Malagnino et al. (1983).
g) Depósitos indiferenciados del Pleistoceno Superior - Holoceno
Los depósitos glaciarios cuaternarios de la isla James Ross han sido analizados por numerosos investigadores entre ellos: Malagnino (1979), Malagnino et al. (1981), Rabassa (1982, 1984, 1987), Ingólfsson et al. (1992), Björck et al. (1996), Hjort et al. (1997), Strelin et al. (1999) y Lirio et al. (2003). Los estudios de las sedimentitas se han concentrado en tres localidades, caleta Santa Marta, península El Morro y bahía Brandy.
En este trabajo se describen los depósitos glaciarios agrupándolos por rangos de edades, considerando las dataciones isotópicas disponibles. En particular, se mencionan los del noroeste de la isla James Ross, de donde se posee mejor información litológica y buen control estratigráfico.
h) Depósitos entre 36.000 y 34.000 años AP
En caleta Santa Marta, Rabassa (1982, 1987) reconoce depósitos glacimarinos, que contienen material fósil, datado por radiocarbono en 34.115 años AP.
Al sur de la caleta Santa Marta, Ingólfsson et al. (1992) estudian una espiga de aproximadamente un km de extensión, que interpretan como una antigua zona de morenas. El depósito comprende una arenisca con capas de gravilla, portadora de fragmentos de Laternula elliptica King y Broderip, con edades 14C de 36.240 años AP. Esta edad es confirmada por los estudios de Lirio et al. (2003).
En cabo Lachman, Ingólfsson et al. (1992) describen un complejo de till de alojamiento, portador de fragmentos mal preservados de Laternula elliptica King y Broderip con una edad 14C de 35.490 años AP.
i) Depósitos entre 25.000 y 24.000 años AP
Fukuda et al. (1992) mencionan terrazas costeras y fluvioglaciarias en los alrededores de caleta Santa Marta, que se encuentran situadas a cuatro alturas diferentes respecto del nivel del mar, entre los 35 y 3 m s.n.m. Dataciones efectuadas con 14C para estos niveles arrojan edades de 25.254 ± 64 años AP para las terrazas superiores y de 1.848 ± 98 años AP para las inferiores.
Para la misma región Lirio et al. (2003) describen dos depósitos similares a los estudiados por Fukuda et al. (1992). El primero corresponde a un till glacimarino situado a 22 m s.n.m. con Laternula elliptica King y Broderip y de edad 14C corregida de 23.300 años AP. El segundo, situado en la costa noroeste de la caleta Santa Marta, es un till basal, que grada a arenas fosilíferas con conchillas de Laternula elliptica King y Broderip, fragmentadas. El depósito presenta bloques de arenisca cretácica, de hasta 1 m de diámetro, con evidencias de estriaciones y orientación de los clastos, que sugieren el avance de un glaciar en dirección hacia el sudeste.
j) Depósitos entre 7.300-6.000 años AP
En la costa noroeste de la península El Morro, Ingólfsson et al. (1992) y Hjort et al. (1997) reconocen sedimentos glacimarinos que presentan pliegues de pequeña escala en su parte superior y fallas normales e inversas que indican desplazamiento desde la dirección sur. Las sedimentitas contienen abundantes conchillas de Laternula elliptica King y Broderip y Thracia meridionalis Smith, datados con 14C en 7.260 años AP. Hacia la parte superior se distingue un estrato arenolimoso con Laternula elliptica King y Broderip y Yoldia eighsii Courthouy, que indica una edad 14C de 6.720 años AP.
Las diamictitas estarían vinculadas a un glaciar de base húmeda con dirección de avance NE, partiendo desde bahía Croft hasta el Morro y la desglaciación habría comenzado con posterioridad a los 7.400 años AP (Hjort et al. 1997). Se destacan sedimentos originados por flujos gravitatorios o transportados por tracción, suspensión y por el hielo, producto de la desglaciación.
k) Depósitos entre 5.000-4.400 años AP
Rabassa (1982, 1984) define el Drift Bahía Bonita y lo data en 5.000 años AP aproximadamente. Hjort et al. (1997), incluyen seis nuevas dataciones con 14C, coincidentes con las de Rabassa (1984, 1987). Para ese momento se habría producido un gran avance glacial en el norte de la isla James Ross, que culmina cerca de los 5.000 años AP; y alcanza la boca de la bahía Brandy. Hjort et al. (1997) y Björck et al. (1996) proponen una rápida desglaciación evidenciada en los sedimentos lacustres acumulados en la costa de bahía Brandy con edades próximas a los 4.200 años AP.
Entre caleta Santa Marta y cabo Lachman aflora un depósito glacimarino portador de conchillas de moluscos cuya edad 14C corregida es de 6.350 años AP (Strelin et al. 1999). Han sido mencionados dos depósitos diamictíticos para caleta Santa Marta: uno glacimarino, portador de Laternula elliptica King y Broderip, con una edad 14C corregida de 5.080 años AP; y otro asociado a un delta representado por conglomerados con gradación normal y estructura entrecruzada asintótica, que pasan gradualmente a una arena fosilífera con Laternula elliptica King y Broderip (4.900 años AP).
l) Depósitos entre 2.000 años AP-Reciente
En el intervalo 2000 años AP hasta la actualidad existen escasos registros de depósitos glaciarios, con excepción de los sedimentos arenosos descriptos por Fukuda et al. (1992) en la caleta Santa Marta, con una edad corregida de 1.847 ± 98 años AP.
ISLA MARAMBIO
a) Formación Weddell Sea (Gazdzicki et al. 2004)
Distintos autores describen los depósitos glaciarios que coronan la meseta de la isla Marambio. Corresponde a Andersson (1906) la primera mención de grandes bloques erráticos, dispersos en la meseta de Marambio, con un origen glacial. Posteriormente Malagnino et al. (1981) y Elliot (1981) estudian y caracterizan estos depósitos, que apoyan sobre la Formación La Meseta.
Zinsmeister y De Vries (1983) utilizan informalmente la denominación Formación Weddell para referirse a estos depósitos y Gazdzicki et al. (2004) modifican el nombre de esta unidad por la de Formación Weddell Sea.
Rabassa (1984) analiza los depósitos y los subdivide en dos tipos litológicos: till alojado y till submarino.
La litología está representada por limolitas arenosas grises, pobremente seleccionadas con numerosos cadilitos; contiene gran número de palinomorfos retrabajados cretácicos y terciarios. Los componentes clásticos provienen mayormente de la cuenca James Ross, los de mayor tamaño son de rocas plutónicas y metamórficas, mientras que los menores están representados por basaltos y andesitas. En menor proporción los bloques corresponden a rocas sedimentarias cretácicas y terciarias fosilíferas, que también constituyen el componente principal de la matriz. Los espesores varían entre 4 y 20 metros.
La edad de estos depósitos ha sido asignada al Cuaternario por Malagnino et al. (1981) y por Elliot (1981) al Terciario Superior. Zinsmeister y De Vries (1983) sustentan la edad cuaternaria. Otros autores sugieren una edad post pliocena tardía para el inicio de la sedimentación, considerando asociaciones de foraminíferos similares a las de la Formación Cockburn Island (Gazdzicki et al. 2004). Extienden el rango temporal de la formación hasta el Pleistoceno teniendo en cuenta las glaciaciones posteriores a la trasgresión del Plioceno Tardío-Pleistoceno (2-3 Ma).
Malagnino et al. (1981) proponen un origen glacimarino para estos depósitos, en tanto que Gazdzicki et al. (2004) sugieren un origen continental.
Ivany et al. (2006) describen una serie de depósitos glaciales y un till de alojamiento para la isla Marambio, de posición infrayacente a las diamictitas de la Formación Weddell Sea. Consideran que éstas no pertenecen a las Formaciones La Meseta o Weddell Sea y acotan la edad del depósito con una importante asociación de dinoflagelados de edad eocena-oligocena a oligocena temprana hallada en el till analizado. Debido al intenso retrabajo de los depósitos próximos a la meseta de Marambio, es discutible si estas nuevas diamictitas no podrían ser parte de la Formación Weddell Sea, contaminada con microplancton marino de la Formación La Meseta.
De no ser así, este nuevo descubrimiento podría representar que el inicio de la glaciación cenozoica para la región de la Antártida occidental se remontaría al Eoceno cuspidal-Oligoceno.
ISLA COCKBURN
El "Conglomerado con Pecten" de la isla Cockburn constituye una de las unidades estratigráficas más citadas en la literatura antártica. Descripto inicialmente por Andersson (1906), aflora en la parte cuspidal de la isla Cockburn. El nombre informal que adquiere esta unidad lo debe al bivalvo pectínido Zygochlamys anderssoni Hennig, que aparece en abundancia en la localidad tipo. Hening (1910), Nelson (1975), Zinsmeister y Webb (1982) se ocupan de estas sedimentitas y es Jonkers (1998b) quien propone denominar a estos depósitos Formación Cockburn Island.
Litológicamente se compone por areniscas rojizas, conglomerados y bloques de hasta 1 metro. La matriz está constituida por clastos de basaltos redondeados. El conglomerado es portador de gastrópodos Patellidae, cirripedios balanidos y serpúlidos incrustantes.
La Formación Cockburn Island aflora en los acantilados de la isla entre los 190 m y los 280 m s.n.m., donde se ha determinado un espesor máximo de 10 m, aunque se describen afloramientos dispersos en la isla, que no superan los 2 m de espesor. Apoya en discordancia sobre las vulcanitas del Grupo Volcánico Isla James Ross. Sin embargo Strelin et al. (1999) plantean que las sedimentitas de la Formación Cockburn Island no coronan el Grupo Volcánico Isla James Ross, sino que se disponen en la base de los depósitos de la segunda época eruptiva del grupo o se intercalan con éstos y estarían vinculados a flujos gravitacionales submarinos.
Los foraminíferos, ostrácodos y macrofósiles de la isla Cockburn fueron comparados con los de otras localidades de la Antártida. Las asociaciones halladas indicarían un ambiente marino somero (menor a 100 m de profundidad) y condiciones interglaciarias (Gazdzicki y Webb 1996, Webb y Andreasen 1986, Szczechura 2001, Whatley et al. 2005).
La edad estimada por el método de isótopos de Sr para el Conglomerado con Pecten de la isla Cockburn es 2,8-2,4 Ma (Plioceno Tardío). Una muestra de basalto del Grupo Volcánico Isla James Ross, situada inmediatamente debajo del Conglomerado con Pecten, es datada por el método de 40K/39Ar en 3,65 ± 0,3 Ma, Plioceno Tardío (Jonkers, 1998b). Medina y del Valle (en Malagnino et al. 1983), obtienen una edad 14C de 36.000 años AP para pectínidos de la misma área, que debe ser considerada como "edad infinita" más allá del método 14C.
El contenido macro y micropaleontológico y la edad absoluta de la Formación Cockburn Island es similar al de la Formación Gage.
ISLA VEGA
En la región de cabo Lamb, en el sector correspondiente a los riscos Leal y Sandwich afloran tres depósitos diamictíticos de color gris verdoso y aspecto caótico (Marenssi et al. 2001). Las sedimentitas apoyan en discordancia sobre la secuencia marina cretácica o se encuentran interestratificadas con distintas unidades volcánicas. Los espesores medidos varían entre uno y 22 metros para los riscos Sandwich y Leal, respectivamente (Fig. 3). La geometría de los depósitos es lenticular. Los bancos son masivos, están compuestos por conglomerados friables, grises o verdosos, matriz soportados, con pobre selección. La fracción clástica, que no supera el 30%, está integrada por clastos de vulcanitas básicas y ácidas, metamorfitas, granitoides y areniscas; la matriz es una fangolita arenosa pobremente seleccionada.
Las diamictitas contienen restos de fósiles marinos, pectínidos, bivalvos y se las interpreta como un till de ablación subácueo depositado por fusión de una barrera de hielo (Marenssi et al. 2001, Lirio et al. en prensa). Las facies sedimentarias reconocidas en cabo Lamb son correlacionables con las facies 1 definidas por Pirrie et al. (1997a) para el sudeste de la isla James Ross. Su edad sería equivalente a los depósitos de cabo Hamilton y punta Rabot (Marenssi et al. 2001).
MICROPALEONTOLOGÍA
La mayoría de las contribuciones micropaleontológicas conocidas de Antártida hacen referencia a asociaciones vivientes de foraminíferos y ostrácodos. Entre los trabajos más relevantes sobre foraminíferos se destacan los de Andersson (1906), Buckman (1910), Hennig (1910), Holland (1910), Chapman (1916a, b), Birkenmajer y Gazdzicki (1986), y Gazdzicka y Gazdzicki (1994). Con respecto a los ostrácodos, la determinación de las especies actuales que habitan en la Antártida se debe a Brady (1880), Skogsberg (1928), Benson (1964), Neale (1967) y Hartmann (1987, 1989, 1990, 1991, 1992) quienes se han ocupado de analizar asociaciones antárticas y subantárticas actuales.
Los estudios sobre comunidades de ostrácodos fósiles se conocen a partir de los trabajos de Hennig (1910), Szczechura y Blaszyk (1996), como así también los de Gazdzicki y Webb (1996), y más recientemente otros de Szczechura (2001) y Whatley et al. (2005).
No obstante, el conocimiento de los microfósiles que habitaron el ambiente marino de la Antártida durante el Cenozoico, permanece casi inexplorado.
A continuación se dan a conocer en forma sintética las asociaciones de microfósiles halladas en las formaciones Belén, Gage, Terrapin, Cockburn Island, Weddell Sea como así también en otros depósitos diamictíticos que no poseen aún denominación formal.
a) Formación Belén
Los foraminíferos contenidos en la Formación Belén han sido estudiados por Jonkers et al. (2002) y posteriormente por Lirio et al. (2003). Se identifican 18 taxones con un marcado predominio de cassidulínidos, entre ellos: Cassidulina crassa d'Orbigny y Cassidulinoides parkerianus (Brady), siendo Cassidulina crassa d'Orbigny el taxón más importante. Definen Ammoelphidiella sp. nov. un nuevo taxón exclusivo del Mioceno, particularmente útil para datar sedimentitas miocenas en regiones australes y carentes de foraminíferos planctónicos diagnósticos (Lirio et al. 2003).
Jonkers et al. (2002), identificaron foraminíferos planctónicos, probablemente ejemplares juveniles de Globigerinita glutinata (Egger), taxón que fuera reconocido originalmente en aguas antárticas por Lipps y Krebs (1974). El hallazgo de Globigerinita glutinata (Egger) es de importancia pues es una especie que permite acotar la fauna al Mioceno Medio-Mioceno Tardío.
Entre los ostrácodos, Jonkers et al. (2002) reconocen exclusivamente Patagonacythere longiducta (Skogsberg), especie que habita los mares australes en la actualidad (cf. Hartmann, 1987). Este taxón ha sido descripto previamente en el "Conglomerado con Pecten" de la isla Cockburn por Szcechura y Blaszyk (1996), otorgándole una edad pliocena. Con el presente hallazgo el rango estratigráfico de la especie se extiende al Mioceno Tardío.
b) Formación Gage
En cabo Gage Lirio et al. (2003) señalan una mayor abundancia de las microfaunas respecto de las de Fiordo Belén. Entre los foraminíferos, se encuentran Ammoelphidiella antarctica Conato y Sagre, taxón que, hasta el presente, es exclusivo del Plioceno de Antártida (Jonkers et al. 2002). Los ostrácodos, resultan escasos aunque más diversos que en las formaciones precedentes y entre ellos se reconocen: Cytheropteron sp. nov. Jonkers, Patagonacythere devexa (Müller), Australicythere polylyca (Müller) y una especie que puede incluirse en la Familia Progonocytheridae la cual, es asignada al género Tumidoleberis sp. nov. Lirio et al.
Whatley et al. (2005) discuten la validez del género Tumidoleberis y consideran que éste no puede ser incluido en la Familia Progonocytheridae, sino que se trata de ejemplares pertenecientes al género Majungaella.
Para el Plioceno Tardío, el sustrato habría estado vegetado, indicando aguas de poca profundidad durante el período de depositación de la formación.
c) Formación Terrapin
En esta formación se han hallado foraminíferos, que a pesar de encontrarse muy fragmentados, poseen una buena a moderada preservación. Se reconocen seis taxones, asignables a las Familias Cassidulinidae y Fursenkoinidae (Jonkers et al. 2002), que incluyen: Elphidium excavatum (Terquem), Elphidium gunteri Cole, Cassidulina crassa (d'Orbigny), Globocassidulina subglobosa Brady, Fursenkoina earlandi (Parr) y Cibicides refulgens de Montfort. El número de cámaras de las especies de Elphidium es menor que las que caracterizan a los taxones encontrados en regiones extraantárticas.
La asociación de foraminíferos se acompaña de ejemplares de ostrácodos de Patagonacythere longiducta (Skogsberg) y es importante la contribución de microfauna del Cretácico Tardío cuspidal, probablemente retrabajada de la Formación López de Bertodano.
d) Formación Cockburn Island
Holland (1910) se ocupa por primera vez de los foraminíferos de esta formación, describe e ilustra 8 géneros y 11 especies. Harwood (1986) menciona una rica asociación de diatomeas junto con silicoflagelados, radiolarios y espículas de esponjas; reconoce 21 géneros y 57 especies y propone una edad de 2,8 a 2,0 Ma (Plioceno Tardío). Dicha edad concuerda con la presencia de Actinocyclus actinochilus (Ehrenberg) Simonsen y Thalassiosira complicata Gersonde, que indica una edad de 3 Ma (Gersonde y Burckle 1990, Harwood y Maruyama 1992). Contemporáneamente Webb y Andreasen (1986) estudian los foraminíferos y macrofósiles de la isla Cockburn e infieren un ambiente costero somero (menos de 100 m), en períodos interglaciarios.
En una revisión exhaustiva de la fauna de foraminíferos de esta isla, Gazdzicki y Webb (1996) encuentran los siguientes taxones: Patellina corrugata Williamson, Pyrgo fornasinii Chapman y Parr, Pyrgo elongata (d'Orbigny), Triloculina sp., Lenticulina gibba (d'Orbigny), Pseudonodosaria sp., Fissurina cf. F. semimarginata (Reuss), Fissurina cf. F. laevigata Reuss, Fissurina quadrata (Williamson), Oolina globosa (Montagu), Cassidulina crassa d'Orbigny, Cassidulinoides parkerianus (Brady), Globocassidulina subglobosa (Brady), Angulogerina angulosa (Williamson), Fursenkoina cf. earlandii (Parr), Epistominella vitrea Parker, Discorbinella sp., Cibicides refulgens de Montfort, Cibicides lobatulus (Walter y Jacob), Nonionella bradii (Chapman), Nonionella cf. turgida (Williamson), Melonis sp., Cribrononion sp., Ammoelphidiella antarctica Conato y Sagre.
La fauna de foraminíferos comparte muchas características con asociaciones pliocenas del sudoeste del mar de Ross-Montes Transantárticos y Larsemann Hills, próximo a bahía Prydzy, Antártida Oriental, y del sector de las islas Shetland del Sur. El paleoclima en la región de la isla Cockburn parece haber sido polar, moderado y se lo ha caracterizado con el término deglacial (Gazdzicki y Webb 1996).
Szczechura y Blaszyk (1996) reconocen 15 géneros y 8 familias de ostrácodos, cuatro de ellas endémicas. Entre las especies más importantes se destacan: Copytus caligula Skogsberg, ?Leptocythere sp., Pseudocythereis spinifera Skogsberg, ?Ramibilis, sp., Procythereis cf. P. robusta Skogsberg, Loxocythere sp. Australicythere polylyca (Müller), Patagonacythere cf. longiducta longiducta Skogsberg, Patagonacythere cf. tricostata Hartmann, ?Meridionalicythere sp., Antarctiloxconcha frigida (Neale), Semicytherura sp. Hemicytherura cf. reticulata Hartmann, Cytheropteron sp., ¿Loxoreticulatum sp. La fauna encontrada sugiere condiciones ambientales semejantes a las actuales y confirma una edad pliocena.
Whatley et al. (2005) revisan las asociaciones de ostrácodos de las Formaciones La Meseta y Cockburn Island, y concuerdan con Szczechura y Blaszyk (1996) acerca de las marcadas diferencias existentes entre ambas. Las asociaciones de la Formación Cockburn Island son modernas y cuatro de las especies halladas llegan a la actualidad o son semejantes a especies antárticas actuales.
e) Formación Weddell Sea
Malagnino et al. (1981) mencionan en estos depósitos la presencia de foraminíferos aglutinados, espículas de esponjas y fragmentos de conchillas de origen "autóctono" hallados en sedimentos finos. Determinan: Cyclammina sp., Haplophragmoides sp., Ammobaculites sp., Ammodiscus sp., Textularia sp. 1. y Textularia sp. 2., Trochammina? sp. y Cribrostomoides? sp.
Cabo Hamilton. Análisis y caracterización de los depositos diamictíticos y nuevos hallazgos paleontológicos
Se realizaron tres perfiles en las localidades: Glaciar Hobbs, (sector noreste de cabo Hamilton), Risco Campamento (sector central del cabo homónimo), Glaciar Sur (en la porción sudeste del cabo) que han sido comparados con observaciones complementarias efectuadas en Watchtower (sur de cabo Hamilton), punta Redonda y punta Rabot. Las diamictitas se encuentran en el sector basal de cada perfil levantado (Figs. 1, 2 y 3).
Litología
Los depósitos son de escaso espesor, entre 1 y 2 m y poseen geometría lenticular. Corresponden a conglomerados muy friables, de color verdoso y pardo, matriz-soportados, pobremente seleccionados, con clastos angulosos a subangulosos, cuya proporción no supera el 40%. Los mayores tamaños están representados por bloques de hasta 50 cm, pero predominan los clastos tamaño grava fina, de metamorfitas (esquistos y granitoides), vulcanitas basálticas vítreas y areniscas concrecionadas.
En el sector del perfil Glaciar Hobbs el espesor es de 1,55 m con base cubierta. Es un depósito caótico, irregular y mal seleccionado con clastos de tamaño y composición variada (Fig. 3). La matriz es pardo verdosa, arenosa fina a mediana con clastos mayores de tamaño grava fina y de hasta 3- 4 centímetros. Se encuentran aislados clastos mayores de orientación general entre Az 60° y 86° los que predominan en la seccion inferior. En la base se colectó una muestra portadora de microfósiles; en el nivel de 0,75 m se hallaron microfósiles y pectínidos, en tanto que a 1,15 m la megafauna es más diversa, la que incluye dos ejemplares pequeños de braquiópodos terebratúlidos, microgastrópodos y serpúlidos. En el tramo superior se intercala un nivel de 3 cm de toba amarillenta, consolidada, portadora de foraminíferos.
En el perfil Campamento el depósito presenta una ligera selección y estratificación. Los clastos menores varían entre los 1-3 cm, los mayores entre 5-10 cm y en ocasiones alcanzan los 50 cm y se hallan dispersos en la parte superior. Se reconocieron ejemplares aislados de Rotularia (Austrorotularia) fallax provenientes de las unidades cretácicas infrayacentes.
Las diamictitas del perfil Glaciar Sur muestran características similares a los depósitos anteriormente descriptos aunque el grado de preservación es menor. Su espesor alcanza los 2 metros.
La sección superior tobácea de la formación está integrada por un conjunto de piroclastitas (lapillitas, chonitas y tobas) interestratificadas con arcilitas limosas de colores verdosos con gran participación piroclástica, equivalentes a las ritmitas descriptas por Carrizo et al., (1999). El perfil Campamento ofrece las mejores exposiciones de la sección superior con un espesor de 1,20 metros. Se diferencian tres sectores: el basal con los primeros 50 cm compuestos por intercalaciones de arcilitas y limolitas, con laminación paralela muy marcada y estructuras de ondulitas escalonadas; le sucede una parte media lapillítica fina, granocreciente, de 30 cm de espesor, con geometría lenticular, que culmina en un estrato delgado de toba y una lapillita lítica, estratificada, en láminas de 1 cm; en contacto neto e irregular se dispone el sector superior con intercalaciones de chonitas y tobas que presentan ondulitas unidireccionales, estratificación convoluta y abundantes estructuras de carga.
La sección piroclástica en el perfil Glaciar Sur posee un espesor similar al descripto precedentemente mientras que en el perfil Glaciar Hobbs se reduce a 25 centímetros.
Paleontología
El intervalo basal de la Formación Hobbs Glacier en el sector norte de cabo Hamilton (Glaciar Hobbs) contiene dos horizontes fosilíferos, situados en los primeros centímetros del perfil y a un metro de la base. Es posible identificar numerosas aurículas bien preservadas y fragmentos de conchillas de valvas derechas de ejemplares juveniles de Zygochlamys anderssoni Henning, como así también fragmentos de conchillas con muy delgadas líneas de crecimiento de Chlamys sp., éstas últimas mencionadas para el área de punta Obelisco, isla James Ross por Jonkers (1998a).
Zygochlamys anderssoni Henning fue descripta inicialmente por Andersson (1906) en la isla Cockburn y datada mediante isótopos de Sr, con una edad pliocena temprana, entre los 5,3 y 3,5 Ma (Dingle et al. 1997). Su presencia en estratos de la Formación Hobbs Glacier indicaría una edad miocena tardía más antigua (Bertels-Psotka et al. 2001).
El hábitat de Zygochlamys andersonni Henning correspondería a un ambiente abierto de plataforma hasta los 100 m de profundidad, como ocurre con representantes actuales (Jonkers 1998a; b), aunque algunas formas con valvas más delgadas y ornamentación fina po-drían habitar ambientes más someros de plataforma interna (Medina, com. verb. 2000). El grado de fragmentación y desarticulación de las valvas de Zygochlamys anderssoni Henning, indica un ambiente proximal de alta energía, vinculado al proceso glaciario, con escaso transporte.
También se han recuperado dos fragmentos pequeños de un cirripedio balánido Fosterella sp. que caracteriza ambientes de aguas poco profundas (Pirrie et al. 1997a). Los dos braquiópodos terebratúlidos corresponden al género Bouchardia Davidson; se trata de ejemplares de tamaño pequeño (1 cm), sulcados, con sus dos valvas cerradas, lisas y forámen epitirido.
Los serpúlidos pueden ser asignados al género Serpula Linnaeus; se presentan como tubos orientados al azar, huecos o rellenos por sedimento y consolidados entre sí, de paredes muy delgadas, a veces se encuentran en contacto dos o más tubos; poseen colores blancos o grises y en corte transversal las paredes están compuestas por láminas concéntricas. Aparecen adheridos sobre fragmentos de conchillas de bivalvos, braquiópodos o sueltos. Algunos tubos son recubiertos por briozoos epibiontes, que aparecen adheridos a otros organismos o en colonias birramosas o unirramosas, con zoarios de 1 cm de longitud. En general se conservan muy recristalizados y son frecuentes los moldes secundarios calcíticos. Se han identificado diferentes géneros entre los cuales se reconocen Microporella, previamente referido por Pirrie et al. (1997a), como así también Cellarinella, Hornera, Melicerita, Membranicellaria y Polirhabdotos (Fig. 4). La presencia de briozoos al igual que el género Serpula indicaría condiciones marinas proximales y su incrustación en clastos de la diamictita podría ser el resultado de su colonización con posterioridad a la sedimentación en el ambiente marino.
Figura 4: Briozoos de la Formación Hobbs Glacier en cabo Hamilton, isla James Ross, Antártida. a) Cellarinella stellaepolaris?, CPBA N° 20054. Fragmento de zoario. b) Membranicellaria sp., CPBA N° 20056. Detalle de zooide. c-d) Melicerita sp., CPBA N° 20057 C. Detalle de zooide. D. fragmento de zoario. e) Hornera sp., CPBA N° 20058. Zoario. f) Microporella sp., CPBA N° 20055. Fragmento de zoario.

Sistemática de briozoos
Los briozoos hallados pertenecen a los órdenes Cyclostomata y Cheilostomata, ambos abundantes en el registro fósil y con representantes actuales en numerosas comunidades bentónicas marinas.
Los Cyclostomata (Ordovícico Tardío-Reciente) constituyen un grupo subordinado en cuanto a diversidad y abundancia con respecto a los Cheilostomata (Jurásico Superior-Reciente). Estos últimos a partir del Maastrichtiano desarrollan importantes innovaciones funcionales tales como la movilidad zoarial y estructuras altamente especializadas (avicularios, vibráculos, entre otras) que le permiten erigirse como grupo dominante de la fauna de briozoos desde el Cretácico Tardío hasta el presente. Estudios comparativos de los patrones de diversidad durante el Cenozoico evidencian que los Cheilostomata son más diversos que los Cyclostomata no sólo a nivel específico observándose una relación aproximada de 8:2, sino también en cuanto a la forma de las colonias. En el material hallado la proporción entre ambos es 5:1.
El orden Cheilostomata presenta zoarios incrustantes, erectos y libres, en general con predominio de los primeros y el orden Cyclostomata incluye formas incrustantes y erectas, siendo estas últimas las más abundantes.
Los Cheilostomata recuperados pertenecen a los subórdenes Ascophora (familias Metrarabdotosidae, Sclerodomidae y Microporellidae) y a Flustrina (familias Membranicellaridae y Cellaridae) y presentan zoarios erectos e incrustantes, con un predominio de los primeros. Los zoarios incrustantes (familia Microporellidae) se hallan tapizando pequeños fragmentos de poliquetos. El orden Cyclostomata está representado por ejemplares de la familia Horneridae (Suborden Cancellata) con zoarios erectos.
Para la determinación sistemática del material hallado se ha seguido a Hayward y Ryland (1979), Hayward (1995) y Bock (2000). La fuerte recristalización de calcita que exhibe el material dificulta la observación de caracteres diagnósticos que permitirían asignar los ejemplares a nivel específico. Entre ellos se destacan la morfología de las aberturas y las espinas orales en los autozooides, la presencia o ausencia y posición relativa de los avicularios (adventicios o vicarios), cantidad por zooide, la morfología de las mandíbulas, forma de los opesios, características de las ovicelas, etc. Los briozoos colectados se encuentran depositados en el Repositorio de la Cátedra de Paleontología de la UBA, CPBA 20054-20059.
Descripciones sistemáticas
Clase Gymnolaemata Allman 1856
Orden Cheilostomata Busk 1852
Suborden Ascophora Levinsen 1909
Superfamilia Lepralielloidea (Umbonuloidea) Vigneaux 1949
Familia Sclerodomidae Levinsen 1909
Género Cellarinella Waters 1904
Especie tipo. Cellarinella foveolata Waters 1904
Cellarinella stellaepolaris ? Moyano y Ristedt en Moyano et al. 2000 (Fig. 4a)
Material: fragmento de colonia. CPBA N° 20054.
Descripción: fragmento de zoario erecto, aplanado, ramificado dicotómicamente en un solo plano. Ramas cilíndricas de distinto grosor, compuestas por vueltas de autozooides. No se observa tipo de adherencia al sustrato. Superficie zoarial reticulada, producto del incremento de la calcificación y que enmascara los límites zoeciales. Aberturas observables, subcirculares a circulares y que podrían corresponder al espacio ocupado por el orificio zoecial y los avicularios asociados. No se distingue aviculario interno ni se observan ovicelas.
Observaciones: el género Cellarinella pertenece a la Familia Sclerodomidae y cuenta con 18 especies cuya distribución geográfica se halla circunscripta al área periantártica, a excepción de Cellarinella dubia Waters, que ha sido hallada sólo en localidades subantárticas de la provincia Magallánica (Moyano et al. 2000). Las características morfológicas utilizadas para la determinación sistemática a nivel específico son, el tipo de adherencia al sustrato, la morfología general del zoario, la forma de la abertura secundaria, la presencia de aviculario interno y el número, posición y forma de avicularios externos. Teniendo en cuenta que la mayoría de estas características no son observables en el material estudiado, la asignación a nivel específico es dudosa.
Sin embargo, merece destacarse que las especies de Cellarinella descriptas hasta la actualidad, sólo han sido registradas para el Reciente, por lo tanto, el hallazgo de este espécimen, ampliaría el biocrón del género hasta el Mioceno.
Distribución estratigráfica conocida: Reciente.
Distribución geográfica: Antártida, mar de Weddell, región de Magallanes argentina y chilena.
Superfamilia Schizoporelloidea Jullien 1883
Familia Microporellidae Hincks 1879
Género Microporella Hincks 1877
Especie tipo. Eschara ciliata Pallas 1766
Microporella sp.
(Fig. 4f)
Material: fragmento de colonia. CPBA N° 20055.
Descripción: fragmento de zoario incrustante de 8,0 mm de largo y 3,0 mm de ancho. Zooides hexagonales convexos separados por surcos, cuya longitud oscila entre los 0,9 mm y 0,7 mm y el ancho es de 0,6 mm aproximadamente. Pared frontal granular en la que no se observan poros. Se alcanzan a distinguir zooides, en cuyo extremo distal se reconoce lo que podría ser la base de ovicelas hiperestomiales. Orificio primario elíptico. No se observan espinas ni su zona de inserción. En algunos zoecios se observa lo que podría ser el ascoporo. Avicularios adventicios, uno por zooide, de posición próximo lateral, a la derecha o a la izquierda de los mismos. No se alcanzan a distinguir rostro ni mandíbula.
Observaciones: el género Microporella posee 39 especies de las cuales en Antártida se registran, Microporella stenoporta Hayward y Taylor, Plioceno-Reciente, Microporella antarctica Hu y Wang, Reciente (Bock 2006) y Microporella ciliata (Pallas). Zoarios asignados a esta última especie han sido citados para las Formaciones Polonez Cove y Cockburn Island (Gazdzicki y Pugaczewska 1984). Representantes de este género han sido también hallados en Punta Rabot, Formación Hobbs Glacier (Pirrie et al. 1997b). La preservación del material descripto en el presente trabajo no permite una asignación a nivel específico.
Distribución estratigráfica conocida: Terciario-Reciente.
Distribución geográfica: Antártida, Chile, Galápagos, Belice, golfo de Méjico, California, Alaska, océano Atlántico Occidental, océano Pacífico, Nueva Zelanda, Australia, Mauritania, Filipinas, Japón y mar Mediterráneo.
Superfamilia Lepralielloidea Vigneaux 1949
Familia Metrarabdotosidae Vigneaux 1949
Género Polirhabdotos Hayward and Torpe 1987
Especie tipo: Smittia inclusa Waters 1904
Polirhabdotos sp.
Material: fragmento de colonia. CPBA N° 20059.
Descripción: fragmento de colonia erecta cilíndrica ramificada distalmente, la longitud es de 5 mm y el ancho de 1 mm. Autozoecios elongados, anchos distalmente y angostos proximalmente, dispuestos en series longitudinales alternadas y formando vueltas de tres zooides alrededor del eje de la colonia. Pared frontal de los autozoecios calcificada y con poros marginales circulares. El grado de preservación no permite observar la morfología de las aberturas ni la presencia de avicularios, se distinguen conspicuos peristomas.
Observaciones: el género Polirhabdotos incluye sólo una especie, Polirhabdotos inclusum Waters, citada para el Holoceno de Antártida. El presente trabajo extiende el biocrón del género hasta el Mioceno.
Distribución estratigráfica conocida: Reciente.
Distribución geográfica: islas Shetland del Sur, archipiélago Palmer, mar de Bellingshausen, mar de Ross y Tierra de Oates.
Suborden Flustrina Smitt 1868
Superfamilia Cellarioidea Lamoroux 1868
Familia Membranicellariidae Levinsen 1909
Género Membranicellaria Levinsen 1909
Especie tipo. Melicerita dubia Busk 1884
Membranicellaria sp.
(Fig. 4b)
Material: fragmento de colonia. CPBA N° 20056.
Descripción: fragmento de colonia erecta de 3,5 mm de longitud y 1,25 mm de ancho. Autozoecios hexagonales cuya longitud es de 0,7 mm y cuyo ancho es de 0,4 mm. Opesios anteriores no terminales rodeadas por cryptocistos. No se observan ovicelas ni avicularios.
Observaciones: el género Membranicellaria incluye una única especie, Membranicellaria dubia Busk, citada para el Reciente del Atlántico Sudoccidental. El hallazgo de este género en Antártida constituiría la primera mención para el Mioceno.
Distribución estratigráfica conocida: Reciente.
Distribución geográfica: océano Atlántico sudoriental.
Familia Cellariidae Fleming 1828
Género Melicerita Milne-Edwards 1836
Especie tipo. Melicerita charlesworthii Morris 1843
Melicerita sp.
(Figs. 4c y d)
Material: fragmento de colonia. CPBA N° 20057.
Descripción: fragmento de zoario erecto, bilaminar, formado por una hoja aplanada simple con bordes afilados, cuya longitud es de 6,0 mm y el ancho es de 4,25 milímetros. No se observa ramificación dicotómica. Autozoecios de contorno frontal hexagonal, de 0,75 mm de largo y 0,5 mm de ancho, dispuestos en filas transversales alternadas compuestas por 8 autozoecios, que muestran una ligera curvatura hacia el extremo proximal del zoario en los bordes del mismo. No se observa la presencia de quenozooides. La forma de los autozooides se modifica hacia el márgen del zoario, quedando las aberturas en una posición excéntrica desplazada hacia la línea media frontal de la colonia y levemente girada. Criptocisto deprimido, granular, rodeado por paredes que en algunas partes del fragmento se ven crenuladas. Opesios en la mitad distal de los autozoecios, con aspecto reniforme, que en algunos ejemplares permite distinguir un incipiente labio proximal recto. En el material estudiado no se observan dentículos, ovicelas ni avicularios.
Observaciones: el género Melicerita está representado por 14 especies distribuidas en tres áreas, antártica, subantártica y australo- neozelandesa (Moyano 1997a; b). Las características diagnósticas utilizadas para la asignación a nivel específico son, presencia o ausencia de avicularios, posición de los mismos, dentículos aperturales, ausentes o presentes, en número de dos o cuatro, quenozooides ausentes o presentes y ancho del zoario. De las características mencionadas, es evidente la ausencia de avicularios y de quenozooides, rasgo que comparte, sólo con Melicerita subantarctica d'Hondt, mientras que el ancho del zoario menor a 5,0 mm es característico no sólo de la especie citada sino también de otras del mismo género. La preservación del material no permite observar los dentículos en el orificio primario. Este género ha sido citado para las Formaciones La Meseta (Hara 2001) y Polonez Cove (Gazdzicki y Pugaczewska 1984).
Distribución estratigráfica conocida: Terciario-Reciente.
Distribución geográfica: Antártida, océano Atlántico Sur, Chile, plataforma continental argentina, Australia, Nueva Zelanda, Nueva Caledonia, islas Kermedec y Europa.
Clase Stenolaemata Borg 1926
Orden Cyclostomata Busk 1852
Suborden Cancellata Gregory 1896
Familia Horneridae Gregory 189
Género Hornera Lamouroux 1821
(Fig. 4e)
Especie tipo: Hornera frondiculata La-mouroux 1821
Hornera sp.
Material: fragmento de colonia. CPBA N° 20058
Descripción: fragmento de zoario erecto de sección oval con esbozos de ramificación en su parte basal, media y superior, cuya longitud es de 5,25 mm y ancho es de 1,25 milímetros. Autozoecios dispuestos alternados y en filas longitudinales. Lado frontal del zoario con aberturas autozoeciales redondeadas y rodeadas en toda su extensión por peristomas. Canceli y sulci longitudinales poco conspicuos. Lado basal del zoario con sulci longitudinales y vacuolas elongadas. No se alcanzan a distinguir gimnozoecios.
Observaciones: de las especies descriptas para este género, Hornera antarctica Waters sensu Borg, 1944, ha sido registrada en las Formaciones La Meseta (Hara 2001), Polonez Cove y Cockburn Island (Gazdzicki y Pugaczewska 1984). El hallazgo de este género en la Formación Hobbs Glacier constituiría la primera mención para el Mioceno.
Distribución estratigráfica conocida: Terciario-Reciente.
Distribución geográfica: Antártida, Australia y Europa.
Micropaleontología
La microfauna recuperada es poco diversa y los valores de riqueza microfaunística son muy bajos. Se han hallado foraminíferos y ostrácodos, además de una gran cantidad de foraminíferos aglutinados alóctonos provenientes de estratos cretácicos de la Formación Santa Marta. La preservación de los caparazones es regular a mala.
Entre los foraminíferos encontrados figuran las especies Astacolus sp. aff. A. crepidulus (Fichtel y Moll), Lenticulina sp. aff. L. limbosa (Reuss), Cassidulinoides parkerianus (Brady), Globocassidulina suglobosa (Brady), Ammoelphidiella sp., y Nonionella bradi (Chapman). Se ha recuperado una sola valva de la especie de ostrácodo Australicythere polylyca (Müller) (Fig. 5).
Figura 5: Foraminíferos y ostrácodos de la Formación Hobbs Glacier en la localidad de Cabo Hamilton, isla James Ross: a) Astacolus sp. aff. A. crepidulus (Fitchel y Moll); FCEN-LM: 2920, la barra indica 0,15 mm. b) Lenticulina sp. aff. L. limbosa (Reuss) FCEN-LM: 2921, la barra indica 0,62 mm. c) Cassidulinoides parkerianus (Brady), FCEN-LM: 2922, la barra indica 0,15 mm. d) Globocassidulina subglobosa (Brady), FCEN-LM: 2923, la barra indica 0,075 mm. e) Amoelphidiella sp., FCEN-LM: 2924, la barra indica 0,22 mm. f) Nonionella bradi (Chapman), FCEN-LM: 2925, la barra indica 0,075 mm. g) Australicythere polylyca (Müller), FCEN-LM: 2926, la barra indica 0,85 mm.

La microfauna de foraminíferos permite correlacionar los estratos analizados con aquéllos de igual edad, Mioceno Superior, estudiados por Jonkers et al. (2002) y postular las siguientes condiciones depositacionales: salinidad normal marina, sobre la base de Lenticulina y Astacolus; temperaturas ligeramente superiores a las actuales en las mismas latitudes y con una efectiva influencia de aguas frías evidenciada por Cassidulinoides parkerianus (Brady). La profundidad de las aguas se correspondería con la de la plataforma interna, avalada por la marcada escasez de foraminíferos planctónicos y de nanofósiles calcáreos, y profundidad circalitoral en los estratos superiores.
Comentarios sobre la sistemática de foraminíferos y ostrácodos
Para el tratamiento sistemático de los foraminíferos y ostrácodos, se menciona el autor de cada especie y se efectúan algunas observaciones acerca del material recuperado. Los ejemplares analizados se encuentran depositados en el Laboratorio de Micropaleontología de la Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, bajo los números de respositorio FCEN-LM: 2920 a 2926.
Foraminíferos
Astacolus sp. aff. A. crepidularis (Fichtel y Moll) (Fig. 5a)
- aff. 1798 Nautilus crepidularis Fichtel y Moll: Testacea Microscopica, p.107, lám 19, figs. g-i.
Observaciones. El ejemplar es muy similar al ilustrado por Jonkers et al., (2002) bajo el nombre de Astacolus sp. La distribución geográfica hallada para el taxón por Boltovskoy et al. (1980) se extiende desde el Sur del Brasil hasta las islas Malvinas e indica que se trata de una especie euritérmica.
Lenticulina sp. aff. L. limbosa (Reuss) (Fig. 5b)
- aff. 1863 Robulina limbosa Reuss: S.-B. Akademie Wissenschaften Wien, v. 48, p. 55. lám. 6, fig. 69.
Observaciones. El ejemplar hallado muestra semejanzas con el ejemplar ilustrado por Boltovskoy et al. (1980) bajo el nombre de Robulus limbosus s.l. del golfo de San Jorge. Dicho autor señala que el taxón tiene una distribución latitudinal hasta aproximadamente los 48° de latitud Sur.
Cassidulinoides parkerianus (Brady)
(Fig. 5c)
-1884 Cassidulina parkeriana Brady: Challenger Exploration Reports Zoology v. 9, p. 432, lám. 54, figs. 11-16.
Observaciones. Según Loeblich y Tappan (1988) el rango estratigráfico del género es Pleistoceno-Holoceno, el que difiere de la edad miocena admitida en el presente trabajo.
La especie se distribuye desde la latitud del Río de la Plata hasta altas latitudes y es característica de aguas frías, en particular de la corriente de Malvinas (Boltovskoy, 1970).
Globocassidulina subglobosa (Brady)
(Fig. 5d)
-1881 Cassidulina subglobosa Brady: Quarterly Journal Micropaleontology Science v. 21, p. 60.
Observaciones. Según Boltovskoy et al. (1980) la distribución geográfica de la especie alcanza altas latitudes en sedimentos recientes. Citada por Jonkers et al. (2002) para la Formación Hobbs Glacier, donde constituye uno de los taxa dominantes, junto a Cassidulina crassa.
Ammoelphidiella sp.
(Fig. 5e)
-2001 Ammoelphidiella sp. nov. Jonkers et al., (2002)
Observaciones. Esta especie, de acuerdo con Jonkers et al. (2002) posee menor tamaño y un lado espiral más bajo que la especie tipo del género Ammoelphidiella antarctica Conato y Sagre. Jonkers et al. (2002) hallaron este taxón en estratos de la Formación Hobbs Glacier en las localidades de fiordo Belén y punta Rabot en la isla James Ross siendo, hasta el presente, exclusivo del Mioceno.
Nonionella bradi (Chapman)
(Fig. 5f)
- 1916 Nonionina scapha var. Bradi Chapman: British Antarctic Expedition 1907-9. Reports Scientific Investigations in Geology v. 2, pt. 3, p. 71, lám. 5, fig. 42.
Observaciones. La especie fue reconocida previamente por Jonkers et al. (2002) en la Formación Hobbs Glacier en punta Rabot (Mioceno) como así también en cabo Gage (Plioceno).
Ostrácodos
Australicythere polylyca (G. W. Müller, 1908)
(Fig. 5G)
-1908 Cythereis polylyca Müller n. sp. Deutschen Sudpolar expedition. 1901-1903. Wissenschaftlige Ergebnisse der Deutsche Sudpolar-expedition. Zoologie. 10, Band 2, p. 135, lám. 17, figs. 1, 5, 6, fig.- text. 1, 4.
Observaciones. Las características morfológicas responden a las descripciones del material viviente en particular las efectuadas por Benson (1964) y Hartmann (1990).
Según Hartmann (1990) la especie viviente habita sustratos duros, arenosos y con vegetación.
Australicythere polylyca fue citada reiteradamente en la Antártida en estratos del Mioceno, Plioceno y en sedimentos de fondo del Holoceno por diversos autores.
DISCUSIÓN Y CONCLUSIONES
La Antártida se ha encontrado englazada los últimos 34 millones de años, la calota de hielo ha variado considerablemente en su configuración y como consecuencia, ha contribuido a producir cambios en el nivel del mar y fluctuaciones climáticas durante el Cenozoico.
Los depósitos glaciarios neógenos de la cuenca James Ross son portadores de un importante registro paleontológico que atestigua estas variaciones.
La revisión de la información composicional y paleontológica disponible hasta el momento y los nuevos hallazgos paleontológicos permiten caracterizar los depósitos glaciarios cenozoicos en la región oriental de la península Antártica.
Las diamictitas de la Formación Hobbs Glacier en cabo Hamilton se caracterizan por dos grupos litológicos bien definidos: una diamictita basal, mal seleccionada y caótica, de espesor variable y una sección superior tobácea estratificada.
Las sedimentitas de cabo Hamilton presentan similitudes litológicas con las facies descriptas en el sudeste de la isla James Ross por Pirrie et al. (1997a). Los depósitos basales y masivos son comparables con la facies 1. En particular, en el perfil Campamento estas sedimentitas muestran una débil estratificación que permite correlacionarlas con la facies 2 propuesta por Pirrie et al. (1997a). La sección tobácea de la Formación Hobbs Glacier, en su porción inferior se corresponde con la facies 4 de tobas con laminación climbing, la parte media se equipara con la facies 5 de tobas arenosas y la sección terminal con la facies 3 de tobas interestratificadas areno-limosas con estructuras convolutas.
La megafauna está integrada por braquiópodos, briozoos, serpúlidos y fragmentos de conchillas juveniles de Zygochlamys anderssoni Henning y es indicadora de ambiente marino de plataforma. Sin embargo, la presencia de valvas delgadas y la importante fragmentación pueden dar indicios de condiciones menos profundas y de alta energía (Bertels-Psotksa et al. 2001).
Particularmente los briozoos Cellarinella, Hornera, Membranicellaria, Microporella y Polirhabdotos, descriptos para regiones antárticas en la actualidad, han sido encontrados en las sedimentitas de la Formación Hobbs Glacier y constituyen los primeros registros conocidos de los géneros para el Mioceno Tardío, extendiendo sus biocrones al Neógeno. En general los ejemplares se encuentran incrustados en clastos y sugieren que la colonización se produjo con posterioridad a la sedimentación glacimarina.
La microfauna de foraminíferos permite correlacionar la Formación Hobbs Glacier con otras del Mioceno Superior de igual edad, estudiadas por Jonkers et al. (2002).
Para definir el paleoambiente, el hallazgo de Lenticulina y Astacolus permite inferir condiciones de salinidad marina normal; temperaturas superiores a las actuales en las mismas latitudes e influencia de aguas frías, evidenciada por la presencia de Cassidulinoides parkerianus (Brady). En este sentido, Dingle y Lavelle (1998) interpretan un renovado deterioro climático en el océano Austral para el Mioceno Superior (aproximadamente 9,3 Ma) y el comienzo de ciclos glaciarios de extensión continental durante el Neógeno Superior, a partir de un incremento en la proporción de detritos transportados por el hielo y mayores valores de d18O.
La profundidad de las aguas en la región de cabo Hamilton se correspondería con la de una plataforma interna, corroborada por la presencia de briozoarios y serpúlidos.
En isla Vega, las diamictitas constituyen depósitos caóticos con litología similar a las del sudeste de la isla James Ross y equivalentes a la facies 1 de la Formación Hobbs Glacier descripta por Pirrie et al. (1997a) y Lirio et al. (2007).
El análisis estratigráfico de las Formaciones Belén y Gage y su contenido paleontológico indican que la sedimentación tuvo lugar durante intervalos interglaciarios en los que el cambio climático se manifestó preferentemente en las zonas costeras. Las condiciones ambientales resultaron propicias para el desarrollo de pectínidos en el continente Antártico, señalando el mejoramiento climático.
El estudio de los microfósiles de la Formación Fiordo Belén ha contribuido a ajustar el esquema bioestratigráfico de los depósitos y el hallazgo del ostrácodo Patagonacythere longiducta permite extender el biocrón de esta especie hasta el Mioceno Tardío.
Las características litológicas de Formación Terrapin y la presencia de clastos y concreciones con estrías glaciarias evidencian un avance glaciario hacia el final del Plioceno. La diversificada asociación de megafósiles y microfósiles recuperada de la Formación Cockburn Island ha permitido efectuar la correlación cronoestratigráfica con la Formación Gage y con unidades litoestratigráficas de las islas Shetland del Sur, con los montes Transantárticos y la Antártida oriental para el Plioceno. La mega y microfauna señalarían un ambiente marino somero y condiciones interglaciarias.
Con posterioridad al Plioceno, las sedimentitas glaciarias de la cuenca James Ross no reciben una denominación formal, por los afloramientos dispersos y su control estratigráfico se reduce a algunas dataciones realizadas sobre material fósil o clastos de basaltos incluidos en las diamictitas y comprenden el lapso 36.000 años AP a la actualidad. Los depósitos glaciarios reconocidos en la región de caleta Santa Marta y cabo Lachman, isla James Ross (36.000 y 25.000 años AP), se corresponden con un período interglaciario del continente antártico y condiciones climáticas relativamente cálidas, tal como fueran determinadas por Berkman et al. (1998) para el intervalo 35.000 y 20.000 años AP.
Las diamictitas de bahía Croft y El Morro coincidirían con la desglaciación propuesta por Ingólfsson et al. (1998, 2003) entre los 14.000-5.000 años AP.
En la isla James Ross se registra un avance glaciario en bahía Brandy entre los 5.000 y 4.500 años AP (Rabassa 1984, Hjört et al. 1997), como así también se sugiere un reavance contemporáneo del glaciar Croft a los 4.900 años AP.
Entre los 4.000 y 3.000 años AP, Ingólfsson et al. (1998) interpretan un óptimo climático a partir de sedimentos lacustres y flora, con condiciones climáticas cálidas y húmedas.
El conocimiento actual sobre el desarrollo glaciológico de la península Antártica permite concluir que después de los 3.000- 2.500 años AP, ocurrió un enfriamiento con una consecuente expansión de los glaciares y barreras de hielo (Björck et al. 1991, 1996). Para este intervalo existen escasos registros sedimentarios en el sector estudiado, con excepción de los que afloran en la caleta Santa Marta.
Finalmente, a partir de los 6.000 años AP se completa la transición de la etapa glacial a interglacial, la que puede caracterizarse por una configuración del hielo muy similar a la actual, con aparición de lagos de agua dulce en tierra firme y la acumulación de sedimentos lacustres.
AGRADECIMIENTOS
Los autores desean expresar su agradecimiento al Instituto Antártico Argentino y a la Fuerza Aérea Argentina por haber brindado los medios económicos y el soporte logístico para la realización de las campañas geológicas al área de estudio. Un especial recuerdo a la memoria de la Dra. A. Bertels- Psotka, quien determinó la microfauna del presente estudio. Al Dr. J. López Gappa por su valiosa colaboración en la revisión de las determinaciones sistemáticas de briozoos. Al Sr. N. Devincenzo, al Lic. F. Tricárico y al Servicio de Microscopia Electrónica del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" por las microfotografías de briozoos. Al Dr. A. Gadzicki por la importante bibliografía aportada, y a los Dres. S. Ballent y R. del Valle por sus valiosos comentarios. Este trabajo es una contribución al poyecto PICTA 16, IAA Programación 2004-2007.
TRABAJOS CITADOS EN EL TEXTO
Ackert, R.P. y Kurz, M.D. 2004. Age and uplift of Sirius Group sediments in the Dominion Range, Antarctica, from surface exposure dating and geomorphology. Global and Planetary Change 42(1-4): 207-225. [ Links ]
Adie, R. J. 1953. The rocks of Graham Land. Ph. D. Thesis, University of Cambridge, 259 p., Cambridge. [ Links ]
Andersson, J.G. 1906. On the Geology of Graham Land. Bulletin of the Geological Institution, University of Uppsala 7: 19-71. [ Links ]
Benson, R.H. 1964. Recent Cytheracean ostracodes from McMurdo sound and the Ross Sea, Antarctica. The University of Kansas Paleontological Contributions. Arthropoda. Article 6: 1-36. [ Links ]
Berkman, P.A., Andrews, J.T., Björck, S., Colhoun, E.A., Emslie S.D., Goodwin, I.D., Hall, B.L., Hart, C.P., Hirakawa, K., Igarashi, A., Ingolfsson, O., López-Martínez, J., Berry Lyons, W., Mabin, M.C.G., Quilty, P.G., Taviani, M. y Yoshida, Y. 1998. Circum-An-tarctic coastal environmental shifts during the Late Quaternary reflected by emerged marine deposits. Antarctic Science 10(3): 345 -362. [ Links ]
Bertels-Psotka, A., Concheyro, A. y Salani, F. M. 2001. Las Diamictitas de cabo Hamilton, isla James Ross, Antártida. Caracterización estratigráfica, sus microfósiles. 4° Congreso de Geología y Minería de la Sociedad Cubana de Geología, en CD, La Habana. [ Links ]
Bibby, J.S. 1966. The Stratigraphy of Part of North-East Graham Land and the James Ross Island Group. British Antarctic Survey Scientific Reports 53: 1-37. [ Links ]
Birkenmajer, K. y Gazdzicki, A. 1986. Oligocene age of the Pecten Conglomerate on King George Island, West Antarctica. Bulletin of the Polish Academy of Sciences, Earth Sciences 34: 219-226. [ Links ]
Birkenmajer, K., Gazdzick, A., K. Krajewski, K.P., Przybycin, A., Solecki, A., Tatur, A. y Yoon, H. 2005. First Cenozoic glaciers in West Antarctica. Polish Polar Research 26(1): 3-12. [ Links ]
Björck, S., Hjort, C., Ingólfsson, Ó. y Skog, S. 1991. Radiocarbon Dates from the Antarctic Peninsula Region - Problems and Potential. Quaternary Proceeding 1: 55-65. [ Links ]
Björck, S., Håkansson, H., Olsson, S. Ellis-Evans, C., Humlum, O. y Lirio, J.M. 1996. Late Holocene palaeoclimatic records from lake sediments on James Ross Island, Antarctica. Palaeogeography, Palaeoclimatology, Palaeoecology 113: 195-220. [ Links ]
Bock, P. 2000. Indexes To Bryozoan Taxa. Rmit University, School Of Civil and Chemical Engineering http://bryozoa.net. [ Links ]
Bock, P., 2006. Indexes To Bryozoan Taxa. Rmit University, School Of Civil and Chemical Engineering http://bryozoa.net. [ Links ]
Boltovskoy, E. 1970. Distribution of the littoral foraminifera in Argentina, Uruguay and Southern Brazil. Marine Biology 6(4): 335-344. [ Links ]
Boltovskoy, E. 1980. Masas de agua en el Atlántico Sudoccidental. En: Boltovskoy D. (Ed.): Atlas del Zooplancton del Atlántico Sudoccidental. Instituto Nacional de Investigación y Desarrollo Pesquero (INIDEP): 227-237. [ Links ]
Boltovskoy, E., Giussani, G., Watanabe, S. y Wright, R. 1980. Atlas of Benthic shelf foraminifera of the Southwest Atlantic: 1-147. Dr. W. Junk bv. Publishers. [ Links ]
Brady, G.S. 1880. Report on the Ostracoda dredged by the H. M. S. Challenger during the years 1873-1876. Reports Scientific Results Voyage of H. M. S. Challenger, Zoology 1(3): 1-184, London. [ Links ]
Busk, G. 1884. Report on the Polyzoa collected by H. M. S. Challenger during the years 1873- 1876. Part I. The Cheilostomata. Report on the Scientific Results of the Voyage of H.M.S. Challenger, Zoology 10(30): 1-216. [ Links ]
Buckman, S.S. 1910. Antarctic fossil Brachiopoda collected by the Swedish South Polar Expedition, 1901-03. Wissenschaftliche Ergebnise der Schwedischen Südpolar-Expedition, 1901-03, Geologie und Paläeotologie 3(7): 1-40. [ Links ]
Carrizo, H., Torielli, C., Strelin, J. y Muñoz, C. 1999. Ambiente eruptivo del Grupo Volcánico Isla James Ross en riscos Rink, isla James Ross, Antártida. Revista de la Asociación Geológica Argentina 53(4): 469-479. [ Links ]
Chapman, F. 1916a. Report on the Foraminifera and Ostracoda from elevated deposits on the shores of the Ross Sea: Report Scientific Investigation of the British Antarctic Expedition 1907-09, Geology, 2, Contribution. Palaeontology Petrology South Victoria Land, Ostracoda: 37-90. [ Links ]
Chapman, F. 1916b. Report on the Foraminifera and Ostracoda out of marine muds from soundings in the Ross Sea: Report Scientific Investigations British of the Antarctic Expedition 1907-09, Geology, 2, Contribution Palaeontology Petrology South Victoria Land, Ostracoda: 71-80. [ Links ]
Concheyro, A., Adamonis, S., Salani, F. M. y Lirio, J. M. 2005. La Formación Hobbs Glacier (Mioceno Superior) en el sudeste de la isla James Ross, Antártida: Localidades y contenido paleontológico. 5° Simposio Argentino y 1° Latinoamericano sobre Investigaciones Antárticas, en CD, Buenos Aires. [ Links ]
De Angelis, H. 1999. Ambiente Eruptivo del Grupo Volcánico James Ross Island en Riscos Lachman, Isla James Ross, Antártida. Trabajo Final de Licenciatura, Departamento de Ciencias Geológicas, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, (inédito), 127 p., Buenos Aires. [ Links ]
del Valle, R.A., Elliot, D.H. y Macdonald, D.I.M. 1992. Sedimentary basins on the east flank of the Antarctic Peninsula: proposed nomenclature. Antarctic Science 4: 477-478. [ Links ]
del Valle, R.A., Marshall, P.A., Lirio, J.M y Camerlingo, E.1987a. Estrías glaciarias bajo las piroclastitas del Terrapin Hill, isla James Ross. Antártida. Primera Reunión de Comunicaciones sobre Investigaciones Antárticas, Resúmenes s/p Buenos Aires. [ Links ]
del Valle, R.A., Marshall, P.A., Lirio, J.M. y Camerlingo, E. 1987b. Sobre la presencia de conglomerado con Pecten en el fiordo Belén. Primera Reunión de Comunicaciones sobre Investigaciones Antárticas, Resúmenes s/p, Buenos Aires. [ Links ]
Dingle, R. V. y Lavelle, M. 1998. Antarctic Peninsular Criosphere: Early Oligocene (c. 30 Ma) initiation and revised glacial chronology. Journal of the Geological Society of London 155: 433-437 [ Links ]
Dingle, R.V., McArthur, J.M. y Vroon, P. 1997. Oligocene and Pliocene interglacial events in the Antarctic Peninsula dated using strontium isotope stratigraphy. Journal of the Geological Society 154: 257-264, London. [ Links ]
Egger, J. G. 1893. Foraminiferen aus Meeresgrundproben, gelothet von 1874 bis 1876 von S. M. Sch.Gazelle. Abhandlungen der Bayerischen Akademie der Wissenschaften. München, Mathematik.-Physik 18(2):193- 458. [ Links ]
Elliot, D.H. 1981. Glacial geology of the Seymour Island. Antarctic Journal of the United States: 66-67. [ Links ]
Fleming, J. 1828. A history of British animals, exhibiting the descriptive characters and systematical arrangement of the genera and species of quadrupeds, birds, reptiles, fishes, mollusca, and radiata of the United Kingdom. Bell and Bradfute: 565 p., Edinburgh. [ Links ]
Fukuda, M., Strelin, J., Shimokawa, K., Takahashi, N., Sone, T. y Trombotto, D. 1992. Permafrost occurrence of Seymour Island and James Ross Island, Antarctic Peninsula Region. En Yoshida et al. (eds.) Recent Progress in Antarctic Earth Science: 745-750, TERRAPUB, Tokyo. [ Links ]
Gazdzicka, E. y Gazdzicki, A. 1994. Recycled Upper Cretaceous calcareous nannoplankton from the Pecten Conglomerate of Cockburn Island, Antarctica. Polish Polar Research 15: 3-13. [ Links ]
Gazdzicki, A. y Pugaczewska, H. 1984. Biota of the "Pecten conglomerate" (Polonez cove Formation, Pliocene) of King George Island( South Shetland Islands, Antarctica). En: Birkenmajer (ed.), Geological results of the Polish Antarctic Expeditions. Part IV. Studia geologica Polonica 79: 59-120, Warsaw. [ Links ]
Gazdzicki, A. y Webb, P.N. 1996. Foraminifera from Pecten Conglomerate (Pliocene) of Cockburn Island, Antarctic Península. En: Gazdzicki, A. (Ed.) Paleontological Results of the Polish Antarctic Expedition. Part II. Palaeontologia Polonica 55: 147-174. [ Links ]
Gazdzicki, A., Tatur, A., Hara, U. y del Valle, R. A. 2004. The Weddell Sea Formation: post- Late Pliocene terrestrial glacial deposits on Seymour Island, Antarctic Peninsula. Polish Polar Research 25(3-4): 189-204. [ Links ]
Gersonde, R. y Burckle, L.H. 1990. Neogene diatom biostratigraphy of ODP Leg 113, Weddell Sea (Antarctic Ocean). En Barker, P.F. Kennett, J.P. et al.(eds.) Proceedings of the Ocean Drilling Program, Scientific Results 113: 761-789. [ Links ]
Gregory, J.W. 1896. Catalogue of the fossil Bryozoa in the British Museum (Natural History), The Jurassic Bryozoa: 1-239, London. [ Links ]
Gregory, J.W. 1899. Catalogue of the fossil Bryozoa in the British Museum (Natural History), The Cretaceous Bryozoa 1, 1-14: 1-457, London. [ Links ]
Hara. U. 2001. Bryozoans from the Eocene of seymour Island, Antarctic Peninsula. En Gazdzicki, A. Ed. Paleontological results of the Polish Antarctic Expedition. Part III. Paleaontologia Polonica 60: 33-155, Warsow. [ Links ]
Hartmann, G. 1987. Antarktische Benthische Ostracoden II. Auswertung der fahrten der "Polarstern" Ant. III/2 und der reisen der "Walter Herwig"68/1 Und 2. 2. Teil: Elephant Island und Bransfield Strasse. Mitteilungen aus dem Hamburgischen Zoologische Museum und Institute 85: 115-156, Hamburg. [ Links ]
Hartmann, G. 1989. Antarktische bentische Ostracoden V. Auswertung der Südwinterreise von FS "Polarstern"(Ps9/-1) im Bereich Elephant Island und der Antarktischen Halbinsel. Mitteilungen aus dem Hamburgischen Zoologische Museum und Institute 86: 231-288, Hamburg. [ Links ]
Hartmann, G. 1990. Antarktische bentische Ostracoden 6. Auswertung der Reise der "Polarstern" Ant. VU-2 (1.Teil, Meiofauna und Zehnersevien) sowie Versuch einer vorläufigen Auswertung aller bislang vorliegenden Daten. Mitteilungen aus dem Hamburgischen Zoologische Museum und Institute 87: 191-245, Hamburg. [ Links ]
Hartmann, G. 1991. Antarktische bentische Ostracoden 7. Ostracoden aus dem Oberen Litoral von King George Island. Mitteilungen aus dem Hamburgischen Zoologische Museum und Institute. Band 88: 211-220. Hamburg. [ Links ]
Hartmann, G. 1992. Antarktische bentische Ostracoden. VIII. Auswertung der Reise der "Meteor"(Ant. 11/4) in die Gewässer um Elephhant Island und der Antarktischen halbinsel. Helgoländer Meeresunter 46: 406-424. [ Links ]
Harwood, D.M. 1986. Diatom biostratigraphy and paleoecology and a Cenozoic history of Antarctica, Philosophical Doctor Thesis, Ohio State University, 592 p. Columbus, Ohio. [ Links ]
Harwood, D. M. y Maruyama T. 1992. Middle Eocene to Pleistocene diatom biostratigraphy of Southern Ocean sdiments from the Kergelen Plateau, Leg 120. En: Wise, S.W., Schlich, R. et al. Proceeding of the Ocean Drilling Program, Scientific Results 120: 683- 733. [ Links ]
Hayward, P. J. 1995. Antarctic Cheilostomatous Bryozoa. Oxford University Press. 1-355. [ Links ]
Hayward, P. J. y Ryland, J. S. 1979. British Ascophoran bryozoans. Linnean Society Synopses of the British Fauna (new series) 14: 1-312. [ Links ]
Hayward, P.J. y Thorpe, J.P. 1987. The systematic position of Smittina inclusa Waters, an endemic Antarctic Bryozoan. Journal of Natural History 21: 1469-1476. [ Links ]
Hennig, A. 1910. Le conglomérat pleistocene a Pecten de l'ile Cockburn. Wissenschaftliche Ergebnisse der Schwedischen Südpolar-Expedition, 1901-1903. Geologie und Paläontologie 3(10): 1-73. [ Links ]
Hincks, T. 1877. On British Polyzoa, Part II. Classification. Annals and Magazine of Natural History 4(20): 520-532, London. [ Links ]
Hincks, T. 1879. On the classification of the British Polyzoa. Annals and Magazine of Natural History 5(3): 153-164, London. [ Links ]
Hjort, C., Ingolfsson, Ó., Möller, P. y Lirio, J. M. 1997. Holocene glacial history and sea-level changes on James Ross Island, Antarctic Peninsula. Journal of Quaternary Science 12: 259-273. [ Links ]
Holland, R. 1910. The fossil foraminifera. Wissenschaftliche Ergebnisse der Schwedischen Südpolar-Expedition, 1901-03, Geologie una Paläontologie 3(9): 1-12. [ Links ]
Ingólfsson, Ó, Hjort, C. y Humlum, O. 2003. Glacial and climate history of the Antarctic Peninsula since the Last Glacial Maximum. Arctic, Antarctic, and Alpine Research 35(2): 175-186. [ Links ]
Ingólfsson, Ó., Hjort, C., Björck, S. y Smith, R. I. L. 1992. Late Pleistocene and Holocene glacial history of James Ross Island, Antarctic Peninsula. Boreas 21: 209-222. [ Links ]
Ingólfsson, Ó., Hjort, C., Berkman, P.A., Björck, S., Colhoun, E., Goodwin, I.D., Hall, B. Hirakawa, K., Melles, M., Möller, P. y Prentice, M.L. 1998. Antarctic glacial history since the Last Glacial Maximum: an overview of the record on land. Antarctic Science 10(3): 326-344. [ Links ]
Ineson, J.R., Crame, A., y Thomson, M.R.A. 1986. Lithostratigraphy of the Cretaceous Strata of West James Ross Island, Antarctica. Cretaceous Research 7: 141-159. [ Links ]
Ivany, L., Van Simaeys, S., Domack, E. y Samson, S. 2006. Evidence for an earliest Oligocene ice sheet on the Antarctic Peninsula. Geology 34, 5: 377-380. [ Links ]
Jonkers, H.A. 1998a. Stratigraphy of Antarctic Late Cenozoic pectinind-bearing deposits. Antarctic Science 12: 161-170. [ Links ]
Jonkers, H.A. 1998b. The Cockburn Island Formation, Late Pliocene interglacial sedimentation in the James Ross Basin, northern Antarctic Peninsula. Newsletter Stratigraphy 36(2/3): 63-76. [ Links ]
Jonkers, H. A. y Kelley, S. P. 1998. A reassesment of the age of the Cockburn Island Formation, northern Antarctic Peninsula, and its palaeoclimatic implications. Journal of the Geological Society 155: 737-740, London. [ Links ]
Jonkers, H. A., Lirio, J. M., del Valle, R. A. y Kelley, S. P. 2002. Lithofacies, biofacies and age of late Neogene sedimentary units associated with the James Ross Island Volcanic Group, Antarctica. Geological Magazine 139: 577-594. Cambridge University Press. [ Links ]
Kiessling, W. y Scasso, R. 1996. Ecological Perspectives of Late Jurassic Radiolarian Fauna from the Antarctic Peninsula. En Riccardi, A.C. (ed.) Advances in Jurassic Research: 317 -326, Georesearch Forum, Zürich. [ Links ]
Lamouroux, J. 1821. Exposition méthodique des genres de l'ordre des Polypiers avec leur dercriptions et celles des principales espèces figurèes dans 84 planches; les 63 premières apartenant à l'Histoire naturelle des Zoophytes d'Ellis et Solander, Agasse, 115 p., Paris. [ Links ]
Levinsen, G. M. R. 1909. Morphological and systematic studies on the cheilostomatous Bryozoa, 431 p., Copenhagen. [ Links ]
Lipps, J. H. y Krebs, W. N. 1974. Planktonic Foraminifera associated with Antarctic Sea ice. Journal of Foraminiferal Research 4(2): 80- 85. [ Links ]
Lirio, J.M. y del Valle, R. A. 1999. Conglomerados con Pecten (Mioceno superior y Plioceno Superior) en la isla James Ross, Península Antártica. Cuartas Jornadas sobre Investigaciones AntárticasActas 298-304, Buenos Aires. [ Links ]
Lirio J.M., Nuñez, H.J., Bertels-Psotka, A. y del Valle, R.A. 2003. Diamictos fosilíferos (Mioceno- Pleistoceno): Formaciones Belén, Gage y Terrapin en la isla James Ross, Antártida. Revista de la Asociación Geológica Argentina 58(3): 298-310. [ Links ]
Lirio, J.M., Marenssi, S.A., Santillana, S.N., Marshall, P. y Rinaldi, C.A. 1989. Marambio Group at Southeastern part of James Ross Island, Antarctica. Contribución del Instituto Antártico Argentino 371: 1-46. [ Links ]
Lirio, J.M., Concheyro, A, Chaparro, M.A., Nyvlt, D. y Mlcoch, F. Diamictita Cabo Lamb, un nuevo depósito fosilífero marino cenozoico en la isla Vega, Península Antártica. VI° Simposio Argentino y III°. Latinoamericano sobre Investigaciones Antárticas. Buenos Aires 10 al 14 de septiembre del año 2007 (aceptado). [ Links ]
Loeblich, A. R. Jr. y Tappan, H. 1988. Foraminiferal genera and their classification. Van Nostrand Reinhold: 1- 970, New York. [ Links ]
López, A. 1994. Geología del Grupo Volcánico James Ross Island y geomorfología del sector comprendido entre las bahías Holluschickie y Villar Fabre. Trabajo Final de Licenciatura. Departamento de Ciencias Geológicas. Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires, (inédito), 106 p., Buenos Aires. [ Links ]
Malagnino, E., Olivero, E., Rinaldi, C. y Spikermann, J. 1979. Aspectos geológicos del borde occidental de la isla James Ross, Antártida. 7° Congreso Geológico Argentino, Actas 1: 489-504, Buenos Aires. [ Links ]
Malagnino, E., Olivero, E., Rinaldi, C. y Spikermann, J. 1981. Aspectos geomorfológicos de la isla Vicecomodoro Marambio, Antártida. 8° Congreso Geológico Argentino, Actas 2: 883-896. [ Links ]
Malagnino, E., Olivero, E., Rinaldi, C. A. y Spikermann, J. 1983. Depósitos glaciarios del Grupo de Isla James Ross, Antártida. Revista de la Asociación Geológica Argentina 38(1): 120-125. [ Links ]
Marenssi, S.A., Lirio, J.M., Santillana, S.N., Martinioni, D.R. y Palamarczuk, S. 1991. The Upper Cretaceous of southeastern James Ross Island, Antarctica. En Rinaldi, C.A. (ed.) Geología de la isla James Ross, Instituto Antártico Argentino: 89-99, Buenos Aires. [ Links ]
Marenssi, S.A., Salani, F.M. y Santillana, S.N. 2001. Geología de Cabo Lamb, Isla Vega, Península Antártica. Contribución del Instituto Antártico Argentino 530: 1-59. [ Links ]
Marenssi, S.A., Santillana, S.N. y Rinaldi, C.A. 1998. Paleoambientes sedimentarios de la Aloformación La Meseta (Eoceno), Isla Marambio (Seymour), Antártida. Contribución del Instituto Antártico Argentino 464: 1-51. Buenos Aires. [ Links ]
Marenssi, S.A., Lirio. J.M., Stinco L.P. y Macchiavello, J.C. 1987. Informe preliminar acerca de diamictitas en el Terciario de punta Redonda, Isla James Ross. En: Resúmenes, Primera Reunión de Comunicaciones sobre Investigaciones Antárticas, Actas 3 p., Buenos Aires. [ Links ]
Medina, F. A., Buatois, L., y López Angriman, A. 1992. Estratigrafía del Grupo Gustav en la isla James Ross, Antártida. En: Rinaldi, C.A. (Ed.): Geología de la isla James Ross: 167- 181. Instituto Antártico Argentino. Dirección Nacional del Antártico. Buenos Aires. [ Links ]
Milne-Edwards, H. 1836. Observations sur les Polypiers fossiles du genre Eschare. Annales des Sciences Naturelles, Zoologie 2 6: 321-45. [ Links ]
Moriwaki, K., Yoshida, Y. y Harwood, D.M. 1992. Cenozoic Glacial History of Antarctica, a correlative synthesis. En Yoshida, Y. et al. (eds.) Recent Progress in Antarctic Earth Science: 773-780, Tokyo. [ Links ]
Morris, J. 1843. A catalogue of British fossils, comprising all the genera and species hitherto described; with references to the geological distribution and to the localities in which they have been found. Van Voorst, 222 p. London. [ Links ]
Moyano, H.I. 1997a. Las especies Chilenas de Melicerita (Bryozoa:Cellaridae) con la descripción de una nueva especie. Gayana 61(1): 49-55. [ Links ]
Moyano G., H. I. 1997b. Revisión de la diversidad y de las conexiones zoogeográficas de los Briozoos Magallánicos. Gayana 61: 125-139. [ Links ]
Moyano, G., Hugo, I., y Ristedt, H. 2000. The Sclerodomidae (Bryozoa) of the eastern Weddell Sea collected during the Antarctic I, II, V Expeditions of the R/V Polarstern. Gayana 64: 83-107. [ Links ]
Neale, J.W. 1967. An Ostracod fauna from Halley Bay, Coats Land, British Antarctic Territory. British Antarctic Survey, Scientific Reports 58: 1-50. [ Links ]
Nelson, P.H.H. 1975. The James Ross Island Volcanic Group of Northeast Graham Land. British Antarctic Survey, Scientific Reports 54: 1-62. [ Links ]
Olivero, E.B., Scasso, R.A. y Rinaldi, C.A. 1986. Revisión del Grupo Marambio en la isla James Ross, Antártida / Revision of the Marambio Group, James Ross Island, Antarctica. Contribución del Instituto Antártico Argentino 331: 1-29. [ Links ]
Pallas, P.S. 1766. Elenchus Zoophytorum sistens generum adumbrationes generaliores et specierum cognitarum succinctas descriptiones, etc. (Petrum van Cleef, Hagae-Comitum) 28: 451 p. [ Links ]
Pirrie, D. y Sykes, M.A. 1987. Regional significance of proglacial delta-front reworked tuffs, James Ross Island area. British Antarctic Survey Bulletin 77: 1-12. [ Links ]
Pirrie, D., Crame, A. y Riding, J. 1991. Late Cretaceous Stratigraphy and Sedimentology of Cape Lamb, Vega Island, Antartica. Cretaceous Research 12: 227-258. [ Links ]
Pirrie, D., Crame, J.A. Lomas, S.A. y Riding, J.B. 1997a. Late Cretaceous stratigraphy of Admiralty Sound region, James Ross Basin, Antarctica. Cretaceous Research 18: 109-137. [ Links ]
Pirrie, D., Crame, A., Riding, J., Butcher, A. y Taylor, P. 1997b. Miocene glaciomarine sedimentation in the northern Antarctic Peninsula region: the stratigraphy and sedimentology of the Hobbs Glacier Formation, James Ross Island. Geological Magazine 136(6): 745-762. [ Links ]
Rabassa, J. 1982. Estratigrafía de los depósitos glacigénicos en el sector norte de la isla James Ross, Península Antártica, Argentina. 5° Congreso Latinoamericano de Geología, Actas 4: 237-251, Buenos Aires. [ Links ]
Rabassa, J. 1984. Nuevas consideraciones sobre la estratigrafía de los depósitos glacigénicos en el sector norte de la isla James Ross, península Antártica. Comentario. Revista de la Asociación Geológica Argentina 39(3-4): 310-318. [ Links ]
Rabassa J. 1987. Drumlins and drumloids forms in northern James Ross Island, Antarctic Peninsula. En Menzies, J. y Rose, J. (eds.) Drumlin Symposium: 267-288, Balkema, Rótterdam. [ Links ]
Rex, D.C. 1976. Geochronology in relation to the stratigraphy of the Antarctic Peninsula. British Antarctic Survey Bulletin 43: 49-58. [ Links ]
Rinaldi, C.A. 1982. The Upper Cretaceous in the James Ross Island Group. En Craddock, C. (ed.) Antarctic Geoscience: 331-337, The University of Wisconsin Press, Madison. [ Links ]
Rinaldi, C.A., Massabie, A., Morelli, J., Rosenman, H.L. y del Valle R.A. 1978. Geología de la isla Vicecomodoro Marambio. Contribución del Instituto Antártico Argentino 217: 1-44. [ Links ]
Skogsberg, T. 1928. Studies on marine ostracods: Part II. External morfology of the genus Cythereis with description of twenty-one new species. California Academy Science Occasional Papers 15: 1-55. [ Links ]
Smellie, J.L., Millar, I.L., Rex, D.C. y Butterworth, P.J. 1998. Subaqueous, basaltic lava dome and carapace breccia on King George Island, South Shetland Islands, Antarctica. Bulletin of Volcanology 59: 245-261. [ Links ]
Strelin, J.A.1990. Estudios preliminaries sobre el Grupo Volcánico Isla James Ross. Informe interno Instituto Antártico Argentino. Dirección Nacional del Antártico, (inédito), 34 p., Buenos Aires. [ Links ]
Strelin, J.A., Martino E. y Malagnino, E.C. 1987. El Cenozoico de la isla James Ross, En: Resúmenes, Primera Reunión de Comunicaciones sobre Investigaciones Antárticas, s/ p., Buenos Aires. [ Links ]
Strelin, J., Carrizo, H., López, A. y Torielli, C. 1993. Actividad volcánica holocenas en la Isla James Ross. Segundas Jornadas sobre Comunicaciones Antárticas: 335-340, Buenos Aires. [ Links ]
Strelin, J.A., Muñoz, C.E., Torielli, C.A., Carrizo H.C. y Medina, F. 1999. Las Diamictitas de la Isla James Ross, Antártida: Origen y probable relación con "Conglomerado con Pecten". Cuartas Jornadas sobre Investigaciones Antárticas: 328-335, Buenos Aires. [ Links ]
Sykes, M.A. 1988. New K-Ar age determinations on the James Ross Island Group, north-east Graham Land, Antarctica. British Antarctic Survey Bulletin 80: 51-56. [ Links ]
Szczechura, J. 2001. Ostracods from the Eocene of the Seymour Island, Antarctic Peninsula. En Gazdzicki, A. (ed.) Paleontological results of the Polish Antarctic Expedition. Part III. Palaeontologica Polonica 60: 157-180. [ Links ]
Szczechura, J. y Blaszyk, J. 1996. Ostracoda from the Pecten Conglomerate (Pliocene) of Cockburn Island, Antarctic Peninsula. En: Gázdzicki, A. (Ed.): Palaeontological Results of the Polish Antarctic Expeditions. Part II. Palaeontologia Polonica 55: 175-186. [ Links ]
Vigneaux, M. 1949. Révision des Bryozoaires néogènes du Bassin d'Aquitaine et essai de classification. Mémoires de la Société Geologique de France 28: 1-153. [ Links ]
Waters, A.W. 1904. Bryozoa. Resultats du Voyage du S. Y. Belgica en 1897-99. Zoologie, 114 p. [ Links ]
Webb, P.N. y Andreasen, J.E. 1986. Potasium/ Argon dating of volcanic material associated with Pliocene Pecten Conglomerate (Cockburn Island) and Scallop Hill Formation (McMurdo Sound). Antarctic Journal of the United States 21(5): 59. [ Links ]
Whatley, R., Ballent, S. y Szczechura, J. 2005. Antarctic Tertiary Progonocytheridae: The last refuge of Majungaella, the survivor of a long lineage, a geographical and physiological migration from low to high latitudes. Palaeogeography, Palaeoecology, Palaeoclimatology 225: 302-316. [ Links ]
Zinsmeister, W.J. y De Vries, T.J. 1983. Quaternary glacial marine deposits on Seymour Island. Antarctic Journal of the United States 18: 64-65. [ Links ]
Zinsmeister, W.J. y Webb, P.N. 1982. Cretaceous-Tertiary geology and Paleontology of Cockburn Island. Antarctic Journal of the United States 17 (5): 41-42. [ Links ]
Recibido: 29 de enero, 2007
Aceptado: 15 de agosto, 2007

