










Serviços Personalizados
Journal

Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de la Asociación Geológica Argentina
versão impressa ISSN 0004-4822versão On-line ISSN 1851-8249
Rev. Asoc. Geol. Argent. v.62 n.4 Buenos Aires out./dez. 2007
Did the continent and sea have different temperatures in the northern Antarctic Peninsula during the Middle Eocene?
Cione, A.L., Reguero, M.A. and Acosta Hospitaleche, C.
CONICET. División Paleontología Vertebrados, Museo de La Plata, Paseo del Bosque s/n, 1900 La Plata
E-mails: acione@fcnym.unlp.edu.ar, regui@fcnym.unlp.edu.ar, acostacaro@fcnym.unlp.edu.ar
ABSTRACT
The Seymour Island beds include a remarkable representation of the continental flora and fauna and marine fauna of Eocene in southern high latitudes. We suggest that, at least during the deposition of the best sampled unit, the Cucullaea I Allomember of the La Meseta Alloformation in the Seymour Island area, a cold temperate terrestrial environment co-existed with relatively warmer temperatures in the adjacent shallow shelf sea. This is suggested by the fish and invertebrate fauna and could have been due to the presence of warmer waters of a current reaching the region from the north. The temperature drop proposed for the time of deposition of the uppermost part of the La Meseta Formation (Submeseta Allomember) appears to correspond to the global drop of the end of the Eocene and beginning of Oligocene and not to the establishment of the Antarctic Circumpolar Current.
Keywords: Climate. Eocene. Antarctica. La Meseta Formation. Seymour Island. Marine. Continental.
RESUMEN
Eran diferentes las temperaturas continentales y marinas durante el Eoceno Medio en el norte de la Península Antártica? Los estratos de la Isla Seymour incluyen una notable representación de flora y fauna continentales y fauna marina del Eoceno de altas latitudes australes. En este trabajo sugerimos que, un ambiente terrestre templado frío coexistió al menos durante la depositación de la unidad mejor muestreada, el Alomiembro Cucullaea I de la Aloformación La Meseta en el área de la Isla Seymour, con un ambiente marino costero y somero con temperaturas más cálidas. Esto es sugerido por la fauna de peces e invertebrados y pudo deberse a la presencia de una corriente de aguas cálidas que alcanzó el norte de la península. La caída de temperatura propuesta para el momento de depositación de la parte más superior de la Formación La Meseta (Alomiembro Submeseta) parece corresponder con la caída global a fines del Eoceno y comienzos del Oligoceno y no al establecimiento de la corriente circumpolar antártica.
Palabras clave: Clima, Eoceno. Península Antártica. Formación La Meseta. Isla Seymour. Marino. Continental.
INTRODUCTION
There is currently some debate about the climatic transition from the Cretaceous "greenhouse" to modern-day "icehouse" conditions (Pfuhl and McCave 2006). A fundamental record for this discussion is included in the Paleogene rocks of Seymour Island in Antarctica (Fig. 1). These beds are highly fossiliferous and yielded a remarkably diverse assemblage of plants, invertebrates and vertebrates that have provided the most detailed record of high-latitude southern Eocene organisms to date (e.g., Feldmann and Woodburne 1988). Nearly all phyla commonly preserved in the fossil record have been described from the unit (Schweitzer et al. 2005). Both continental and marine organisms indicate that the environment was completely different to that present in Antarctica today.
Figure 1: Map of Seymour (Marambio) Island showing locality IAA 1/90.
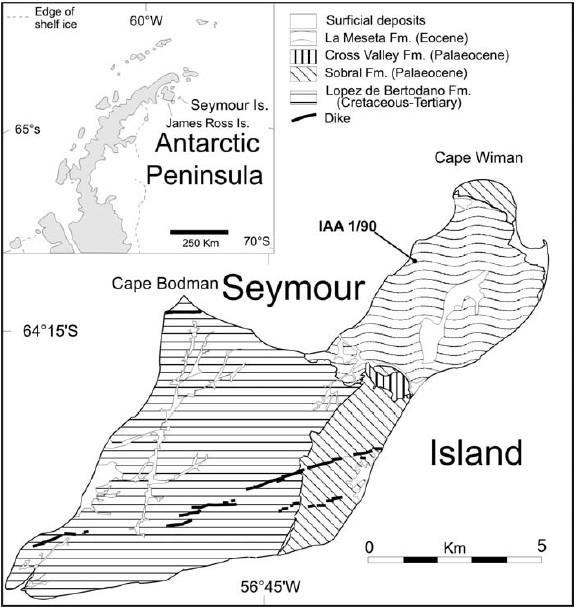
Zinsmeister (1979, 1982) proposed a biogeographic unit of shallow marine waters (the Weddellian Province) that encompassed the coasts of Australia, Tasmania, Antarctica and southern South America during the late Cretaceous through Eocene. The sea around Seymour Island was considered to be cold temperate during the Eocene by some authors (e.g., Case 1992, Long 1992a) although Long (1992c) noted the presence of some shallow water tropical migrants that occasionally entered the area. Case (1988) included in this biogeographic province the continental flora and fauna. These latter were characterized by the association of megafloral assemblages dominated by the Southern Beech, Nothofagus sp. and marsupials of the family Polydolopidae. Later, some new floral and faunal taxa were reported (see Reguero et al. 2002).
The Eocene rocks from Seymour Island contain a record of marine (and continental) organisms representing ecosystems with no Recent parallels, as they developed in relatively warm environments in a sunlight/ darkness-stressed setting (Parras et al. 2005).
Indeed, we found that proxies suggest large differences among the temperature preferences of marine and continental organisms present at least in some levels of the Eocene La Meseta Formation. We discuss them and propose a cause for explaining these differences in the oceanographic pattern present at that time.
METHODS
The fossil record in the La Meseta Formation is extremely diverse. In this paper we refer only to the Cucullaea I Allomember of that formation (Marenssi et al. 1998 a, b).
We selected some organisms as proxies due to their abundance and significance as indicators. Among fishes, the extreme abundance of neoselachians in the record is due to taphonomic reasons, as it is the case in other shallow marine deposits (see Cione 1988). For neoselachian we considered only the better sampled localities (IAA 1/90 and IAA 2/95, both close and correlative). Most of the material comes from IAA 1/90 which yielded by dry screening more than 8000 teeth in different campaigns.
Many sharks are highly mobile animals. They can (and do) follow the optimal environmental conditions by migrating along the coasts, crossing ocean stretches or displacing to deeper or shallower waters (see discussion in Long 1992a). Concerning migrating species, fossil representation corresponds to those that present their optimum in summer or in winter. This is a radical difference with fossil sessile or non-migratory organisms (most invertebrates, rays) which indicate tolerance to winter conditions in the area (Cione and Barla 1997, Cione et al. 2005). Migratory individuals from warm waters indicate summer temperatures (e.g., odontaspidid and isurid sharks; see Cione et al. 1994; Cione and Azpelicueta 2002). Migratory individuals of taxa from cold areas indicate winter temperatures (e.g., the porbeagle shark Lamna).
There is some debate about what is called warm temperate and cold temperate water in littoral populations. Stephenson (1953) as summarized by Knox (1960; also adopted by Briggs 1974) proposed that warm temperate waters have a minimal temperature (average for the warmest month) of between 12° and 20°C and a maximum summer range (average for the warmest month) near or somewhat exceeding 25°C. In cold temperate waters the temperature (average for the coldest month) should fall below 12°C in the winter, but usually not to zero; summer mean temperatures may approach 20°C. The Cold temperate region is divided into two sub-regions: Subantarctic cold temperate, with a mean temperature range from 3 to about 14-15°C and Cold temperate mixed waters with a mean temperature range of 7 to 18°C. Knox (1960) commented that thermal boundaries are not equal everywhere. Knox (1960) suggested that in the Antarctic waters the temperatures fall below zero in the winter and do not exceed 3-5°C in the summer (see also Charnock 1996).
In the southwestern Atlantic, the shelf Argentinian Province (Fig. 2; see below) reaches waters of about 8°C or less (in the coldest month). Knox (1960) considers this area as cold temperate. However, species present there are clearly different from those of the Magallanian Province and are similar to those inhabiting waters to the north of the Rio de la Plata that are identified as warm temperate by Knox (1960). Argentinian zoologists consider the Argentinian Province as warm temperate (e.g., López 1964, Menni 1981).
Figure 2: South American shelf marine biogeographic units (modified from López 1964) and the Subantarctic Province (modified from Cabrera and Willink 1973). Note that both units scarcely overlap in latitude.
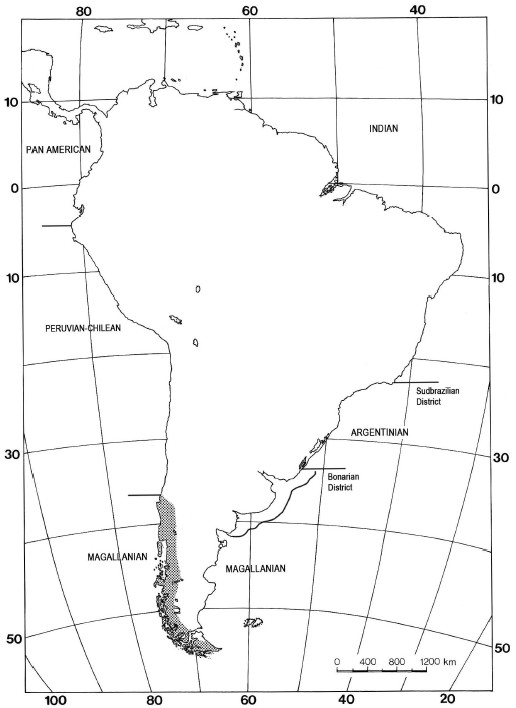
Abbreviations
ACC, Antarctic Circumpolar Current; DP, Drake Passage; LMF, La Meseta Formation; TG, Tasman Gateway.
SEYMOUR ISLAND GEOGRAPHIC AND PALEOGENE STRATIGRAPHIC SETTING
The early Eocene/early Oligocene La Meseta Alloformation is an unconformitybounded unit (La Meseta Alloformation of Marenssi et al. 1998a, 1998b). This unit has a maximum composite thickness of 720 meters filling up a 7 km wide valley cut down into the older sedimentary rocks of the island after the regional uplift and tilting of the Paleocene and Marambio Group beds (Fig. 3).
Figure 3: Comparison of temperature curve (according to Zachos et al. 2001) with La Meseta Formation unconformity-bounded internal units and ages (after Marenssi 2006).
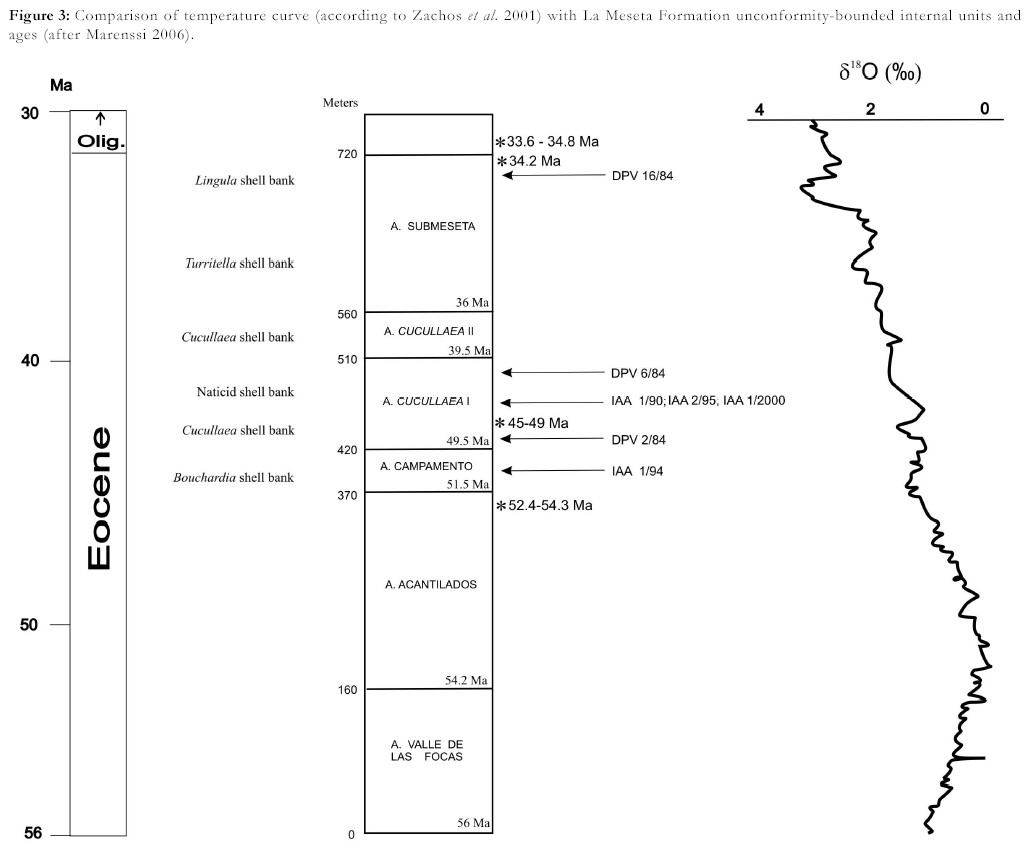
It comprises mostly poorly consolidated siliciclastic fine-grained sediments deposited in deltaic, estuarine and shallow marine environments as a part of a tectonically-controlled incised valley system (Marenssi 1995, Marenssi et al. 1998b). This unit is made up of several lens-shaped unconformity-bounded members representing different sedimentation stages (Marenssi et al. 2002).
Paleoenvironmental reconstructions indicate that the La Meseta Alloformation accumulated at the seaward end of an incised valley during an overall rise in sea level (Marenssi 1995, Marenssi et al. 1998a). Recent studies of the geometric relationships in the La Meseta Alloformation (Sadler 1988, Marenssi 1995, Marenssi et al. 1998b) show that the steep erosional boundaries are the margins of a large channel some 7 km in width that originated farther west. The head of the La Meseta incised valley was placed almost 60 km to the northwest, at the toe of the Antarctic Peninsula (Stilwell and Zinsmeister 1992).
87Sr/86Sr ratios of bivalves from the highest shell-bearing marine bed in the Submeseta Allomember yield ages of 33.57-34.78 Ma (Ivany et al. 2006), and are the youngest documented ages from the La Meseta Alloformation and confirm that the unit extends to the E-O boundary (33.9 Ma; Gradstein et al. 2004). The oldest dates, 52-54 Ma (based on 87Sr/86Sr isotopic ratios after Reguero et al. 2002), in the formation come from the 150 m level in the boundary of the Valle de las Focas and Acantilados allomembers of Marenssi and Santillana (1994). Consequently, the base of the La Meseta should be close to the beginning of the Eocene at 54.8 Ma (Gradstein et al. 2004).
The unit is unconformably overlain by Pliocene-Pleistocene till of the Weddell Sea Formation (Zinsmeister and DeVries 1983, Gazdzicki et al. 2004).
RECENT OCEANOGRAPHY AND CLIMATE
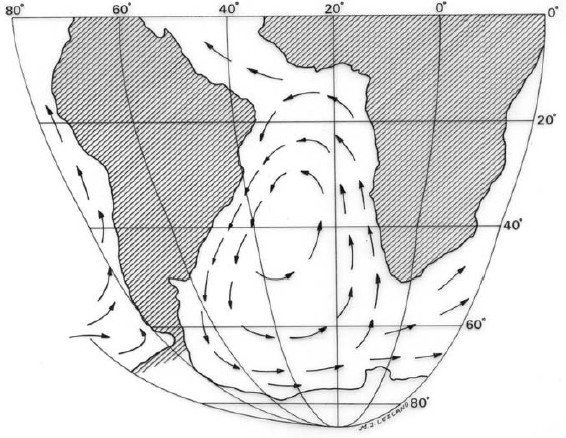
The dominant feature of the current system of the southern region is the Antarctic Circumpolar Current which influences the greatest part of the Antarctic oceanic region and the whole of the temperate oceanic region (Knox 1960, Fig. 4). It branches into currents flowing northerly off the coast of the major land masses. On the western part of South America it becomes in the Humboldt Current (also Peru Current). In the Drake Passage, the Antarctic Circumpolar Current is named Cape Horn Current which, after passing the southern tip of the continent, divides into two branches. The west branch passes on both sides of the Malvinas (Falkland) Islands and is called the Malvinas Current. It finds the warm southward Brazil Current at about 43°-46° S and turns east (López 1964). The Humboldt Current is much stronger than the Malvinas Current and its influence extends to near the Equator (Sverdrup et al. 1942). The combined effect of the northerly transport and upwelling is to produce that the average surface temperatures off the coast of Peru are something like 10°C lower than the theoretical values for the latitude (Knox 1960).
Warm currents are prevented (by the offshoots of the Antarctic Circumpolar Current) from circulating the sun's heat away from tropical and temperate latitudes into the polar region (Pielou, 1992).
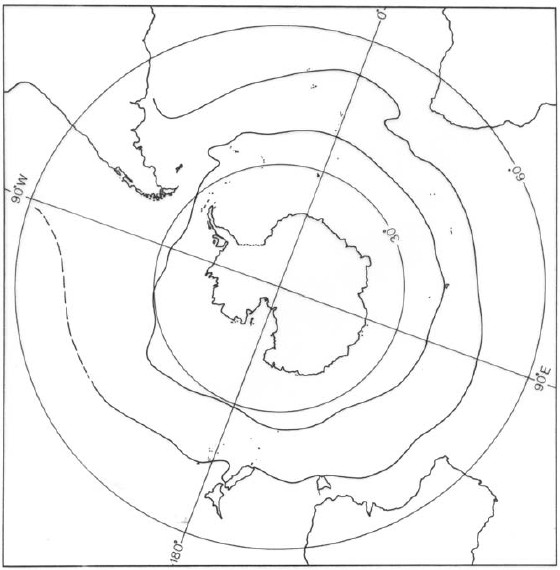
The southern ocean is divided into several regions on the basis of surface temperatures (Fig. 5). With increasing distance from the Antarctic continent temperature increases slowly until a region within which an increase of 2 or 3°C takes place in a very short distance (Knox 1960). This region is called the Antarctic Convergence. Further to the north, the surface temperature rises slowly until another region of rapid increase of about 4 or 5°C, the Subtropical Convergence. Both convergences limit the Antarctic region and the Subantarctic region (Knox 1960).
Figure 4: Recent marine circulation in southern Atlantic Ocean (modified from Sverdrup et al. 1942).
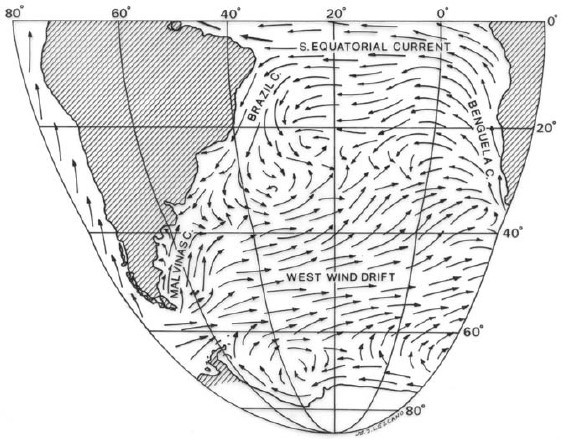
Figure 5: Antarctic and Subantarctic convergences (modified from Knox 1960).
RECENT BIOGEOGRAPHY OF ANTARCTICA AND SOUTH AMERICA
a) Marine
Different authors have divided the coastal marine waters of South America into biogeographic areas. Regan (1914) distinguished that the fishes determined a Temperate Zone with Argentinian and Chilean Districts and a Subantarctic Zone with the Magallanian and Antipode Districts (also an Antarctic Zone). Eckman (1953) distributed the whole marine fauna into Shelf Warm Waters Faunas, Temperate Waters Faunas, Warm-Temperate Waters Faunas and Antarctic Faunas.
López (1964) considered the following icthyogeographic units in South America: Magallanian Province (cold-temperate waters) with two districts, South Chilean and Patagonian; Argentinian Province (warm-temperate waters) with two districts, Bonarian and South Brazilian; West Indian Province (warm waters) with three districts, Gulf of Mexico and Caribbean, Antillean, and Brazilian (Fig. 2). Balech (1964) proposed a rather different scheme. Menni (1981, 1983) commented the biogeography of fishes and Martínez and del Río (2002) that of mollusks.
The Magallanian Province occupies the southernmost part of South America. According to López (1964), in the Atlantic coast it reaches 42°S and in the Pacific 40-41°S (Fig. 2). According to Balech (1964), it reaches 43°S and 40°S respectively. However, in the outer platform and talud the Magallanian Province reaches about 34°S. At latitude of 43° to 45°S separates from the coast.
The Argentinian Province ranges from about 42°S to 23°S according to López (1964).
However, boundaries between the Argentinian and Magallanian provinces could be presently changing due to the global temperature changes (see Góngora et al. 2003).
Waters around Antarctica, beyond the Antarctic Convergence, have a distinctive fauna.
b) Continental
Botanists and some general biogeographers consider an austral Recent biogeographic unit called Antarctic Region that encompasses part of South America and New Zealand (Cabrera and Willink 1973). Most zoogeographers do not accept that parts of South America were not included in the Neotropical Region (e.g., Ringuelet 1961). In South America, the Subantarctic Province of the Antarctic Region is recognized in southern Chile and westermost southern Argentina between 37°S and 55°S (Fig. 2). The climate is cold temperate and wet. Evergreen conifer/broad-leaf forests dominate, but there are also open areas. Rainfall in these areas is very high, mainly in summer and spring seasons, with an average precipitation from 1000-3000 mm/y and freezing temperatures can prevail during several months of the year. The unit is characterized floristically by the predominace of genera (or even families) of austral distribution. Genera most important are Nothofagus, Dacrydium, Laurelia, Lomatia and others (Cabrera and Willink 1973). The endemic taxa are a family (Misodendracea) and several genera (Fitzroya, Austrocedrus, Pilgerodendron, Macrachenium, Combera).
TAXA AND CLIMATE
The paleobotanical record of Paleogene Antarctic vegetation indicates the loss of warm-temperate vegetation and the increased dominance of other types more typical of high latitude environments (Askin 1997, Birkenmajer and Zastawniak 1989, Francis 1999).
Paleofloral data sets show that the middle late Eocene was warmer than present, but not as warm as the late Paleocene (Cross Valley Formation) through early Eocene (Acantilados Allomember, La Meseta Alloformation). Paleoflora from the middle part of the La Meseta Alloformation (RV 8200, IAA 1/90, and DPV 3/84 = C/88, from Cucullaea I Allomember) indicate a drop in temperature (Case 1988, Gandolfo et al. 1998) with respect to lower levels of the sequence.
Nothofagus is considered to be of critical importance as an paleoclimate indicator. Nothofagus is the predominant angiosperm taxon in each of the three paleofloras from the La Meseta Alloformation mentioned above. Gandolfo et al. (1998) reported N. serrulata and two indeterminate species of the same genus from the late early Eocene Acantilados Allomember (A/88). The former species is now restricted to southern South America, where it grows in a cool temperate climate. Nothofagus serrulata extends from southern Chile (Carmen Silva, Loreto, and Brush Lake Formations) and Argentina (Río Turbio, Río Guillermo, and Ñirihuau formations) to the southern shores of Tierra del Fuego Province (Cullen Formation), and thus provides a stronger geographical link to Antarctica than is suggested by other South American species (Fig. 2) (Reguero et al. 2002).
EVOLUTION OF CLIMATE DURING THE PALEOGENE
Paleoclimatic interpretations of the Paleogene of the James Ross Basin were proposed using different lines of evidence (sedimentology, fossils and stable isotope analysis). The idea of a temperate marine environment from the late Cretaceous through the late Eocene has been proposed some time ago based on the presence of particular taxa of marine mollusks (Weddellian Province of Zinsmeister 1982). It was recently confirmed by stable isotope analysis (Gazdzicki et al. 1992, Dichfield et al. 1994; Pirrie et al. 1998, Dutton et al. 2002). Francis (1991) interpreted a marked cooling of the climate during the Cretaceous-Paleocene transition on Seymour Island based on the study of growth ring patterns of tree trunks collected from the López de Bertodano and Sobral formations. By the time of deposition of the Cross Valley Formation (Upper Paleocene) the climate had returned to a warmer level (Francis 1991). Gandolfo et al. (1998) indicate the presence of subtropical forests based on the percentage of entire margin leaves (43%) from the same unit. Likewise, several lines of evidence point to indicate a seasonal cool-temperate rainy climate for the Middle Eocene of the Seymour Island with a cooling trend through the Eocene till the earliest Oligocene (Case 1988, Francis 1991, Torres et al. 1994, Gandolfo et al. 1996, 1998). Case (1988) suggested that the nothophyllous leaves of the lower part of the La Meseta Alloformation indicate a period of climatic amelioration with respect to the middle and upper part of the unit where microphyllous leaves were recovered. The palynoflora presents austral characteristics (Zamaloa et al. 1987) while taxonomic analysis indicates a mixture of Antarctic (dominant) and neotropical megafloral elements (Gandolfo et al. 1998). Morphological analysis of the leaves led Gandolfo et al. (1998) to interpret the presence of a mixed mesophitic forest (32% of entire margin leaves) developed under a seasonal cool-temperate climate with dry and rainy seasons. Mean annual paleotemperatures were calculated to be between 11° and 13°C and the mean of the coldest months between -3° and 2°C.
The marine temperature of the Seymour Island area, on the basis of oxygen isotope ratios of late Eocene invertebrate shells, was estimated as 8°C for the minimum winter temperature and 12°C for the minimum summer temperature (Kennett and Barker 1990).
Geochemistry and clay mineralogy of sedimentary rocks from Seymour Island were used to interpret the climatic evolution of the northern Antarctic Peninsula area since the Late Cretaceous (Dingle and Lavelle 1998). A cool period is indicated during the Early Paleocene before the climatic optimum of the Cenozoic (late Paleocene-early Eocene) followed by a climatic deterioration from very warm, non-seasonally wet conditions (early middle Eocene) to a latest Eocene cold, frost-prone and relatively dry stage (Dingle et al. 1998).
The sudden, widespread glaciation of Antarctica and the associated shift towards colder temperatures at the Eocene/Oligocene boundary (~34 million years ago) (Zachos et. al. 1996, 2001, Barrett 1996, Lear et al. 2000) is one of the most fundamental reorganizations of global climate known in the geologic record. The glaciation of Antarctica has hitherto been thought to result from the tectonic opening of Southern Ocean gateways, which enabled the formation of the Antarctic Circumpolar Current and the subsequent thermal isolation of the Antarctic continent (Kennett 1977).
The earliest Oligocene (ca. 33.5 Ma) is marked by a rapid and significant positive shift in the oxygen isotope value of marine carbonates, and the appearance of ice-rafted debris in the Southern Ocean that corresponds to the first major expansion of Antarctic ice in the Cenozoic (Zachos et al. 1992).
The opening of Southern Ocean gateways and the formation of the Antarctic Circumpolar Current undoubtedly cooled high southern latitudes. However, the simulated effect of a 20% change in ocean heat transport associated with the opening of Drake Passage is shown to have a smaller effect than that expected in the transition from a 'greenhouse' to an 'icehouse' climate. In our model, the opening of Drake Passage can only be a potential trigger for glacial inception when atmospheric CO2 is within a relatively narrow range, reinforcing the importance of pCO2 as a fundamental boundary condition for Cenozoic climate change (DeConto and Pollard 2003).
ESTABLISHMENT OF THE DRAKE PASSAGE
A highly debated issue is the establishment of the modern circulation around Antarctica. Actually, the Antarctic Circumpolar Current is a strong, wind-driven current, which in parts reaches the ocean-floor and significantly limits meridional heat-exchange in the Southern Hemisphere (Barker and Thomas 2004, Pfuhl and McCave 2006). The Antarctic Circumpolar Current inception depended on changes in atmospheric circulation patterns but was determined by the separation of Antarctica after the establishment of the Tasman Gateway south of Australia and the Drake Passage south of America to deep throughflow (Lawver and Gahagan 2003, Pfuhl and McCave 2006). Major plate motions based on dated seafloor spreading anomalies and distinct fracture zone lineations constrain the age of the opening of a seaway between the South Tasman Rise and Antarctica as very close to the Eocene/Oligocene boundary, with an unrestricted opening deeper than 2000 m dating from ~ 32 Ma.
Timing of the opening of Drake Passage is more circumstantial because the exact motions of certain micro-continental fragments are not known. Uncertainty of motion of the South Georgia and South Orkney microcontinents and other possible continental fragments make an exact time for opening of Drake Passage difficult to ascertain. Even so, the early Oligocene position of the Antarctic Peninsula with respect to South America requires a throughgoing, deepwater seaway to have been open at Drake Passage prior to 28 Ma, even given the unconstrained motion of various highstanding crustal fragments in the Scotia Sea. With reasonable assumptions concerning motion of the crustal fragments in the western and central Scotia Sea, it is likely that Drake Passage or passage through Powell Basin was open to deep water circulation by ~ 31 ± 2 Ma. Recent investigations in the Leg 189 to the Tasman Gateway, suggest that a homogenous Southern Ocean water mass did not exist prior to the mid-Oligocene interval (Pfuhl and McCave 2006, see also Beu et al. 1999).
MARINE AND CONTINENTAL TEMPERATURES DURING THE DEPOSITION OF CUCULLAEA I ALLOMEMBER
Global temperatures during the Eocene were in general much higher than today (see above).
Warm temperate conditions during the deposition of the Cucullaea I Allomember are suggested by the presence of typically warm-water taxa such as Cucullaea, Polinices, Autratoma, Ringicula, ficids and ranellids (Stilwell and Zinsmeister 1992). Additional support for relatively warm temperatures is provided by the presence of a number of extinct austral taxa such as the bivalve Lahillia, and the gastropods Struthiolarella, Taioma and Struthioptera, which are known from early Tertiary warm-temperate facies of Australia and New Zealand. Shells of the bivalve Cucullaea are very abundant and form banks in the Cucullaea I and II allomembers. Shells of brachiopods Glottidia and Bouchardia form banks in several levels of the La Meseta Alloformation including Cucullaea I Allomember (Emig and Bittner 2005). Two fossil species of Bouchardia appears to occur in most localities cited in La Meseta Alloformation (Wiedman et al. 1992). A close relative of these two species, B. rosea, is currently found living in shallow waters off the South American coast at the latitude of Rio de Janeiro and perhaps to the south (Manceñido and Griffin 1988). These waters correspond to the Sudbrazilian District of the Argentinian Province (Fig. 2).
Cirripeds also indicate warm waters. Austrobalanus is represented by a single extant species restricted to the warm, temperate southeast coast of Australia. Extant Scoliodobalanus achieves its greatest diversity in the tropical and subtropical Indo-West Pacific region, but species are also known from warmer regions of the Atlantic Ocean (Zullo et al. 1988).
We only considered neoselachian proportions from the best sampled locality (IAA 1/90; and in part IAA 2/95) which correspond to a bank of naticids (mostly Polynices) in the Cucullaea I Allomember. Actually, most of the fish taxa mentioned here are found in many different levels of the La Meseta Alloformation excepting the upper levels; however, they are currently being restudied. The most abundant (according to tooth number) taxa are Squatina (37.89%), Pristiophorus (22.45%), Odontaspididae (17.24%), Myliobatis (6.70%), Squalus (6.81 %), Rajidae (4.72) and Holocephali (2.68 %). The other taxa are present in a much smaller proportion (Centrophorus, Stegostoma fasciatum, Cetorhinus, Dalatias licha, Carcharocles auriculatus, Isurus praecursor, Lamna, Scoliodon). The shark assemblage suggests shallow waters (discussed in Long 1992c. There are some species with affinities to deeper waters but these taxa have also been found into shallow water deposits from other parts of the world (e.g., Miocene of Patagonia; Cione 1988). Most neoselachian families and genera that dominate are typical of the Argentinian province of central Argentina, Uruguay and Brazil: odontaspidids (represented in La Meseta Alloformation mainly by Striatolamia macrota, Odontaspis rutoti and Odontaspis winkleri), squatinids (Squatina), and myliobatids (Myliobatis) (see Menni 1981, Cousseau and Perrotta 1992). These families have almost no representation in the Magallanian province. Other common taxa such genus Squalus (represented by Squalus woodburnei and S. weltoni) and pristiophorids are wide-ranging in warm and cold temperate seas (Compagno et al. 2005). Pristiophorid sharks became extinct in southern Atlantic after the Miocene (Cione and Azpelicueta 2002). Remarkably, there are several taxa that presently inhabit warm temperate to tropical areas (e.g., the sharks Stegostoma, Isurus, the carcharhinids Carcharhinus and Scoliodon) and Pristis, the teleosts Oplegnathus, and Trichiuridae; Long 1991, 1992b, 1992c. Cione and Re-guero 1994, Cione et al. 1994, Kriwet 2005). The porbeagle shark Lamna occurs both cold and warm temperate waters (Compagno et al. 2005) but is rare in the unit. Consequently, ichthyological evidence suggests that sea temperatures would have been similar to those of the recent marine Argentinian Province especially those of the Sudbrazilian District (e.g., Cione 1978, Long 1992 b, Cione and Reguero 1994. Cione et al. 1994, Fig. 2).
On the basis of oxygen isotope ratios on Late Eocene shells in Seymour Island areas, mean temperatures appear to have been above 8°C in winter and above 15°C in summer (Kennett and Barker 1990).
On the continent, plant (Gandolfo et al. 1998), geological and palynological (Askin 1997, Dingle et al. 1998) evidence suggest that the climate in Seymour Island would have been similar to that of the recent Subantarctic Province (the Valdivian forest) of southern Chile and Argentina (see also Case 1988, Reguero et al. 2002, Poole and Cantrill 2006, Fig. 2). Moreover, microbiotheriid mammals still live in this province. The Subantarctic Province has a cold temperate and wet climate, with annual mean temperatures between 11°C and 13°C and the coldest month mean between -3°C and 2°C (Gandolfo et al. 1998). Certainly, temperatures could be colder in higher altitudes and in the southernmost area.
The northern part of the Subantarctic Province scarcely overlaps the latitude of the southern part of the Argentine Province (Fig. 2).
Palynological and geological continental evidence agree with a dramatic shift to colder conditions both in the marine and continental environments at the top of the La Meseta Alloformation (Dingle et al. 1998, Poole and Cantrill 2006). Invertebrate diversity is low in the basal part but steadily increases to a peak in the abundant shell lenses of Cucullaea I Allomember (Stilwell and Zinsmeister 1992). However, a sharp decrease in diversity occurs near the boundary Cucullaea II and Submeseta allomembers. In agreement, in the Submeseta Allomember, the largest diversity of penguins occurs (Tambussi et al. 2006), there is a dramatic diminution of diversity of those selachians that dominated below, there are no teleost taxa characteristic of warm water (e.g., Labridae, Oplegnathidae, Xiphiorhynchidae), sharks dramatically decreases in diversity and quantity (a few Pristiophorus and odontaspidid teeth) and begin to predominate some sharks (Lamna) and teleosts with species characteristic of colder waters (e.g., gadiforms of the informal genus "Mesetaichthys") are recorded.
Marine temperatures depend on latitude, but also on the current pattern. Presently, the two branches of the Antarctic Circumpolar Current that run along the eastern and western coasts of South America are different in strength (Fig. 4). This difference cause that temperatures are very different on both sides of the continent and that biogeographic pattern is different.
During the Eocene, notwithstanding that some circulation could exist in Drake area (Wrenn and Hart 1992), there was not a well developed Antarctic Circumpolar Current and consequently no author proposed the existence of the equivalent to Humboldt and Malvinas currents. Smith and Briden (1977 fide Manceñido and Griffin 1988; see also Parrish and Curtis 1982) suggest a model of circulation for the Paleogene that involves a warm current running southward along the eastern coast of South America at least to the present Drake Passage latitude (Fig. 6). Both the terrestrial fauna of southern South America and Seymour Island suggest that a terrestrial connection existed at least until de early Eocene. Mammal taxa in the Vacan Subage (early Eocene) permit to propose a terrestrial communication for the sparnotheriodontids in Antarctica near the Vacan Subage (early Eocene) or Riochican Age (late Paleocene; Bond et al. 2006).
Figure 6: A land connection between South America and Antarctica was suggested for the Cretaceous (Case et al. 1988). Does it persist until the Early Eocene as mammal evidence suggest? A model of marine circulation during Eocene times previous to the establishment of the Antarctic Circumpolar Current is depicted.
In the Weddell area, a counter current that flowed northward along the eastern coast of the Antarctic Peninsula exists today (Sverdrup et al. 1942). There is no evidence of its presence in the Eocene and the southward current could have extended into the southernmost regions. Actually, the Weddell area could have some similarity to the present northeastern Atlantic area, where the North Atlantic Current warms higher latitude areas (Sverdrup et al. 1942). In summary, we suggest that, at least during the deposition of the Cucullaea I Allomember of the La Meseta Alloformation in the Seymour Island area, a cold temperate terrestrial environment coexisted with relatively warmer temperatures in the adjacent shallow shelf sea. This could have been due to the presence of waters of a warm current reaching the region. The temperature drop proposed for the time of deposition of the uppermost part of the La Meseta Formation (Submeseta Allomember) appears to correspond to the global drop of the end of the Eocene and beginning of Oligocene and not to the establishment of the Antarctic Circumpolar Current.
THE WEDDELLIAN PROVINCE
The Weddellian Province was proposed as a shallow water and temperate marine shelf region that encompassed New Zealand, Tasmania, Australia, Antarctica and southern South America during the late Cretaceous through Eocene (Zinsmeister 1979, 1982, Woodburne and Zinsmeister 1984, Zinsmeister and Feldmann 1984). At this time, these continents were proximate and shared many marine taxa.
This biogeographic hypothesis was expanded to include the terrestrial fauna and flora (Case 1988). The province was based on the predominance of species of the Southern Beech Nothofagus and caenolestid marsupials. However, marine and continental biogeographic (both large and small) units do not necessarily have the boundaries at the same latitude. In South America, continental units (see Cabrera and Willink 1973) hardly have something to do with shelf units (see López 1964). Actually, in southern South America there is an approximate coincidence in the northern boundary of the marine Magallanian Province with the continental Subantarctic forest in the western side of South America. However, the northern boundary of the marine Magallanian Province does not agree with a continental biogeographic boundary on the eastern side of South America (López 1964, Ringuelet 1961, Cabrera and Willink 1973, see Fig. 2). In this case, as in many others, the marine organism distribution is greatly affected by marine currents, the western and stronger Humboldt Current and the eastern Malvinas Current. On the other hand, the continental topography and climate greatly determine the continental organism distribution (Ringuelet 1961, Cabrera and Willink 1973).
We found that the marine fauna appears to correspond to warmer environments that the continental in the Eocene of Seymour Island (see above). Moreover, plant taxa present in the La Meseta Formation do not appear to differ much from those of the recent Subantarctic Province of southern Chile and Argentina (i.e., a flora dominated by species of genus Nothofagus; Cabrera and Willink 1973). A typical inhabitant is also a small marsupial, the microbiotherid Dromyciops gliroides.
Consequently, it appears more appropriate to restrict the term Weddellian Province for the marine realm and perhaps to propose a different one for the continent.
ACKNOWLEDGMENTS
We especially acknowledge the Instituto Antártico Argentino and Fuerza Aérea Argentina, which provided logistical support for our participation in the Antarctic fieldwork and the Agencia Nacional de Promoción Científica y Tecnológica, Consejo Nacional de Investigaciones Científicas y Técnicas, and Universidad Nacional de La Plata for partial finantial support for this project.
WORKS CITED IN THE TEXT
Askin, R. A. 1997. Eocene-¿earliest Oligocene terrestrial palynology of Seymour Island. In: Ricci, C. A. (ed.). The Antarctic Region: Geological Evolution and Processes. Terra Antarctica Publication 993-996, Siena. [ Links ]
Balech, E. 1964. Caracteres biogeográficos de la Argentina y Uruguay. Boletín del Instituto de Biología Marina de Mar del Plata 7: 107-112. [ Links ]
Barker, P. F. and Thomas, E. 2004. Origin, signature and palaeoclimatic influence of the Antarctic Circumpolar Current. Earth-Science Reviews 66: 143-162. [ Links ]
Barrett, P. J. 1996. Antarctic paleoenvironment through Cenozoic times-a review. Terra Antarctica 3: 103-119. [ Links ]
Beu, A.G., Griffin, M. and Maxwell, P.A. 1999. Opening of Drake Passaje gateway and late Miocene to Pleistocene cooling reflected in Southern Ocean molluscan dispersal: evidence from New Zealand and Argentina. Tectonophysics 281: 83-97. [ Links ]
Birkenmajer, K. and Zastawniak, E. 1989. Late Cretaceous-Early Neogene vegetation history of the Antarctic Peninsula sector, Gondwana breakup and Tertiary glaciations. Bulletin of the Polish Academy of Sciences (Earth Sciences) 37: 63-88. [ Links ]
Bond, M., Reguero, M. A., Vizcaíno, S. F., and Marenssi, S. A. 2006. A new "South American ungulate" (Mammalia: Litopterna) from the Eocene of the Antarctic Peninsula. In Francis, J. E., Pirrie, D., and Crame, J. A. (eds.). Cretaceous - Tertiary High Latitude Palaeonvironments, James Ross Basin, Antarctica. The Geological Society, Special Publications 258:163-176, London. [ Links ]
Briggs, J. C. 1974. Marine zoogeography. Mc-Graw Hill, 475 p. [ Links ]
Cabrera, A. L. and Willink, A. 1973. Biogeografía de América Latina. Programa de Desarrollo Científico y Tecnológico. OEA, Serie Biología 13: 1-120. [ Links ]
Case, J. 1988. Paleogene floras from Seymour Island, Antarctic Peninsula. In: Feldmann, R. M. and Woodburne, M. O. (eds.) Geology and Paleontology of Seymour Island, Antarctic Peninsula. Memoir of the Geological Society of America 169: 523-530. [ Links ]
Case, J. A. 1992. Evidence from fossil vertebrates for a rich Eocene Antarctic marine environment. Antarctic Research Series 56: 119- 130. [ Links ]
Charnock, H. 1996. The atmosphere and the ocean. In: Summerhayes, C.P. and Thorpe, S.A. (eds.) Oceanography, 27-40, Manson Publishing, London. [ Links ]
Cione, A. L. 1988. Los peces de las formaciones marinas del Cenozoico de Patagonia. Ph.D Dissertation, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata (unpublished), 536 p. [ Links ]
Cione, A. L. and Azpelicueta, M. M. 2002. An oplegnathid fish from the Miocene of Patagonia. Geobios 35: 367-373. [ Links ]
Cione, A. L., Azpelicueta, M. M., and Bellwood, D. 1994. A knifejaw (Osteichthyes: Oplegnathidae) from the Eocene of Antarctica. Palaeontology 37: 931-940. [ Links ]
Cione, A. L., Azpelicueta, M. M., Casciotta, J.R. and Dozo M.T. 2005. Tropical freshwater teleosts from Miocene beds of eastern Patagonia, southern Argentina. Geobios 38: 29-42. [ Links ]
Cione, A. L. and Barla, M.J. 1997. A new locality for the synbranchid eel Synbranchus marmoratus (Teleostei: Percomorpha) in southern Buenos Aires province, Argentina. Neotrópica 43: 113-115. [ Links ]
Cione, A. L. and Reguero, M. 1994. New records of the sharks Isurus and Hexanchus from the Eocene of Seymour Island, Antarctica. Proceedings of the Geologist's Association 105: 1-14. [ Links ]
Cione, A. L. and Reguero, M. 1998. A middle Eocene basking shark (Lamniformes, Cetorhinidae) from Antarctica. Antarctic Science 10: 83-88. [ Links ]
Compagno, L., Dando, M. and Fowler, S. 2005. A field guide to the sharks of the world. Collins, 368 p., London. [ Links ]
Cousseau, M.B. and Perrotta, R.G. 1998. Peces marinos de Argentina. Instituto Nacional de Investigación y Desarrollo Pesquero, 167 p., Mar del Plata. [ Links ]
DeConto, R. M. and Pollard, D. 2003. Rapid Cenozoic glaciation of Antarctica triggered by declining atmospheric CO2. Nature 421:245-249. [ Links ]
Ditchfield, P. W., Marshall, J. D., and Pirrie, D. 1994. High latitude palaeotemperature variation: new data from the Tithonian to Eocene of James Ross Island, Antarctica. Palaeogeography, Palaeoclimatology, Palaeoecology 107: 79-101. [ Links ]
Dingle, R. and Lavelle, M. 1998. Antarctic Peninsula cryosphere: early Oligocene (c. 30 Ma) initiation and a revised glacial chronology. Journal of the Geological Society 155: 433- 437, London. [ Links ]
Dingle, R. V., Marenssi, S., and Lavelle, M. 1998. High latitude Eocene climate deterioration: evidence from the northern Antarctic Peninsula. Journal of South American Earth Sciences 11:571-579. [ Links ]
Dutton, A. L., Lohmann, K., and Zinsmeister, W. J. 2002. Stable isotope and minor element proxies for Eocene climate of Seymour Island, Antarctica. Paleoceanography 17(2): 1- 13. [ Links ]
Eckman, S. 1953. Zoogeography of the sea. Sidgwick and Jackson, 417 p., London. [ Links ]
Emig, C. and Bittner, M.A. 2005. Glottidia (Brachiopoda: Lingulidae) from the Eocene La Meseta Formation, Seymour Island, Antarctica. Palaeontology 48: 423-431. [ Links ]
Feldmann, R. and Woodburne, M.O. (eds.) 1988. Geology and Paleontology of Seymour Island, Antarctic Peninsula. The Geological Society of America, Memoir 169: 1-566. [ Links ]
Francis, J.E. 1991. Palaeoclimatic significance of Cretaceous/early Tertiary fossil forests of the Antarctic Peninsula. In Thomson, M.R. A., Crame, J.A. and Thomson, J.W. (eds.) Geological Evolution of Antarctica: 623-627, Cambridge University Press, New York. [ Links ]
Francis, J.E. 1999. Evidence from fossil plants for Antarctic paleoclimates over the past 100 million years. Terra Antarctica Reports 3: 43-52. [ Links ]
Gandolfo, M.A, Marenssi, S.A., and Santillana, S.N. 1998. Flora y paleoclima de la Formación La Meseta (Eoceno medio), isla Marambio (Seymour), Antártida. In Casadío, S. (ed.) Paleógeno de América del Sur y de la Península Antártica. Asociación Paleontológica Argentina, Publicación Especial 5: 155-162. [ Links ]
Gazdzicki, A. J., Gruszczynski, M., Hoffman, A., Malkowski, K., Marenssi, S. A., Halas, S., and Tatur, A. 1992. Stable carbon and oxygen isotope record in the Paleogene La Meseta Formation, Seymour Island, Antarctica. Antarctic Science 4: 461-468. [ Links ]
Gazdzicki, A., Tatur, A., Hara, U., and Valle, R. A. D. 2004. The Weddell Sea Formation: Post-late Pliocene terrestrial glacial deposits on Seymour Island, Antarctic Peninsula. Polish Polar Research 25: 189-204. [ Links ]
Góngora, M.E., Bovcon, N., Cochia, P. and Gosztonyi, A.E. 2003. Aportes al conocimiento de la ictiofauna del golfo San Jorge. 5° Jornadas Nacionales de Ciencias del Mar (Mar del Plata, 8-12 de diciembre de 2003) Resumen: 117. [ Links ]
Gradstein, F., Ogg, J., and Smith, A. 2004. A geologic time scale 2004. Cambridge University Press, 589 p., Cambridge. [ Links ]
Ivany, L.C., Van Simaeys, S., Domack, E.W., and Samson, S.D. 2006. Evidence for an earliest Oligocene ice sheet on the Antarctic Peninsula. Geology 34(5): 377-380. [ Links ]
Kennett, J.P. 1977. Cenozoic evolution of Antarctic glaciation, the circum-Antarctic oceans and their impact on global paleoceanography. Journal of Geophysic Research 82: 3843-3859. [ Links ]
Kennett, J. P. and Barker, P. F. 1990. Latest Cretaceous to Cenozoic climate and oceanographic developments in the Weddell Sea, Antarctica: an ocean-drilling perspective. In: Barker, P. F. and Kennett, J. P. (eds.) Scientific Results. College Station Texas, Ocean Drilling Program, Proceedings of the Ocean Drilling Program 865-880, Houston. [ Links ]
Kriwet, J. 2005. Additions to the Eocene selachian fauna of Antarctica with comments on Antarctic selachian diversity. Journal of Vertebrate Paleontology 25(1):1-7. [ Links ]
Knox, G. A. 1960. Marine Biology. Littoral ecology and biogeography of the southern oceans. Proceedings of the Royal Society 152: 577-624. [ Links ]
Lawver, L. A. and Gahagan, L. 2003. Evolution of Cenozoic seaways in the circum-Antarctic region. Palaeogeography, Palaeoclimatology, Palaeoecology 198: 11-37. [ Links ]
Lear, C. H., Elderfield, H., and Wilson, P. A. 2000. Cenozoic deep-sea temperatures and global ice volumes from Mg/Ca in benthic foraminiferal calcite. Science 287:269-272. [ Links ]
Long, D.J. 1991. Fossil cutlassfish (Perciformes: Trichiuridae) teeth from the La Meseta Formation (Eocene), Seymour Island, Antarctic Peninsula. Paleobios 13: 3-4. [ Links ]
Long, D.J. 1992a. The shark fauna from La Meseta Formation (Eocene), Seymour Island, Antarctic Peninsula. Journal of Vertebrate Paleontology 12:1-32. [ Links ]
Long, D.J. 1992b. An Eocene wrasse (Perciformes; Labridae) from Seymour Island. Antarctic Science 4:235-237. [ Links ]
Long, D.J. 1992c. Paleoecology of Eocene Antarctic sharks, In: Kennett, J.P. and Warnke, D.A. (eds): The Antarctic paleoenvironment: a perspective on global change, Antarctic Research Series 56: 131-139. [ Links ]
López, R. 1964. Problemas de la distribución geográfica de los peces marinos suramericanos. Boletín del Instituto de Biología Marina 7: 57-63, Mar del Plata. [ Links ]
Marenssi, S.A., 1995. Sedimentología y paleoambientes de sedimentación de la Formación La Meseta, isla Marambio, Antártida. Ph.D. Dissertation, Universidad de Buenos Aires, (unpublished) 1: 1-330., 2: 1-172. [ Links ]
Marenssi, S.A. 2006. Eustatically controlled sedimentation recorded by Eocene strata of the James Ross Basin, Antarctica. In Francis, J. E., Pirrie, D., and Crame, J. A. (eds.) Cretaceous - Tertiary High Latitude Palaeonvironments, James Ross Basin, Antarctica. Geological Society, Special Publications 258:125- 133, London. [ Links ]
Marenssi, S.A., Net, L.I., and Santillana, S.N. 2002. Provenance, depositional and paleogeographic controls on sandstone composition in an incised valley system: the Eocene La Meseta Formation, Seymour Island, Antarctica. Sedimentary Geology 150: 301-321. [ Links ]
Marenssi, S.A. and Santillana, S.N. 1994. Unconformity-bounded units within the La Meseta Formation, Seymour Island, Antarctica: a preliminary approach. In 21° Polar Symposium (Warszawa) Polish Academy of Sciences, Abstracts 33-37, Warsaw. [ Links ]
Marenssi, S.A., Santillana, S.N., and Rinaldi, C.A. 1998a. Paleoambientes sedimentarios de la Aloformación La Meseta (Eoceno), Isla Marambio (Seymour), Antártida. Instituto Antártico Argentino, Contribución 464: 1-51. [ Links ]
Marenssi, S.A., Santillana, S.N., and Rinaldi, C.A. 1998b. Stratigraphy of the La Meseta Formation (Eocene), Marambio (Seymour) Island, Antarctica. In Casadio, S. (ed.) Paleógeno de América del Sur y de la Península Antártica, Asociación Paleontológica Argentina, Publicación Especial 5: 137-146, Buenos Aires. [ Links ]
Martínez, S. and del Río, C. 2002. Las provincias malacológicas miocenas y recientes del Atlántico sudoccidental. Anales de Biología 24: 121-130. [ Links ]
Manceñido, M. O. and Griffin, M. 1988. Distribution and palaeonvironmental significance of the genus Bouchardia (Brachiopoda, Terebratellidina): Its bearing on the Cenozoic evolution of the South Atlantic. Revista Brasileira de Geociências 18:201-211. [ Links ]
Menni, R. C. 1981. Sobre la distribución de los peces marinos de la Argentina. 6° Jornadas Zoológicas Argentinas, Symposia: 57-74, La Plata. [ Links ]
Menni, R. 1983. Los peces en el medio marino. Estudio Sigma S.R.L., 169 p., La Plata. [ Links ]
Parras, A., Casadío, S., Griffin, M., Marenssi, S., and Santillana, S. 2005. Borers and encrusters as giveaways to the presence of hermit crabs in Antarctic Eocene gastropods. Ameghiniana 42: 38R. [ Links ]
Parrish, J. and Curtis, R.J. 1982. Atmospheric circulation, upwelling and organic-rich rocks in the Mesozoic and Cenozoic eras. Palaeogeography, Palaeoclimatology, Palaeoecology 40: 31-66. [ Links ]
Pfuhl, H.A. and McCave, I.N. 2006. Investigating the link between Antarctic Circumpolar Current inception and events at the Oligocene-Miocene transition. Geophysical Research Abstracts 8: 6047. [ Links ]
Pielou, E. C. 1992. After the Ice Age. The University of Chicago Press, 366 p., Chicago. [ Links ]
Pirrie, D., Marshall, J.D. and Crame, J.A. 1998. Marine high Mg calcite cements in Teredolites bored fossil wood: evidence for cool palaeoclimates in the Eocene La Meseta Formation, Seymour Island, Antarctica. Palaios 13: 276-286. [ Links ]
Poole, I. and Cantrill, D. J. 2006. Cretaceous and Cenozoic vegetation of Antarctica integrating the fossil wood record. In Francis, J. E., Pirrie, D. and Crame, J. A. (eds.) Cretaceous - Tertiary High Latitude Palaeonvironments, James Ross Basin, Antarctica, The Geological Society, Special Publications 258: 63-81, London. [ Links ]
Regan, C.T. 1914. British Antarctic ("Terra Nova") Expedition 1910. Natural History Report 1: 1-54. [ Links ]
Reguero, M.A., Marenssi, S.A. and Santillana, S.N. 2002. Antarctic Peninsula and Patagonia Paleogene terrestrial environments: biotic and biogeographic relationships. Palaeogeography, Palaeoclimatology, Palaeoecology 2776: 1-22. [ Links ]
Ringuelet, R.A. 1961. Rasgos fundamentales de la zoogeografía de la Argentina. Physis 22: 151-170. [ Links ]
Sadler, P. 1988. Geometry and stratification of uppermost Cretaceous and Paleogene units on Seymour Island, northern Antarctic Peninsula. In Feldmann, R. M. and Woodburne, M. O. (eds.) Geology and Paleontology of Seymour Island, Antarctic Peninsula, Geological Society of America, Memoir 169: 303-320, Boulder. [ Links ]
Schweitzer, C., Feldmann, R., Marenssi, S., and Waugh, D.A. 2005. Remarkably preserved annelid worms from the La Meseta Formation (Eocene), Seymour Island, Antarctica. Palaeontology 48: 1-13. [ Links ]
Stilwell, J.D. and Zinsmeister, W.J. 1992. Molluscan systematics and biostratigraphy: Lower Tertiary La Meseta Formation, Seymour Island, Antarctic Peninsula. Antarctic Research Series 55: 1-152. [ Links ]
Sverdrup, H.U., M.W. Johnson, and R.H. Fleming. 1942. The oceans. Prentice-Hall, Inc., Englewood Cliffs, 1087 p., New Jersey. [ Links ]
Tambussi, C.P., Acosta Hospitaleche, C.I., Reguero M.A., and Marenssi, S.A. 2006. Late Eocene penguins from West Antarctica: systematics and biostratigraphy. In Francis, J. E., Pirrie, D. and Crame, J.A. (eds.) Cretaceous - Tertiary High Latitude Palaeonvironments, James Ross Basin, Antarctica. The Geological Society, Special Publications 258: 145-161, London. [ Links ]
Torres, T., Marenssi, S.A., and Santillana, S.N. 1994. Maderas fósiles de la isla Seymour, Formación La Meseta, Antártica. Serie Científica del INACH 4: 17-38, Santiago de Chile. [ Links ]
Woodburne, M.O. and Zinsmeister, W.J. 1984. The first land mammal from Antarctica and its biogeographic implications. Journal of Paleontology 58: 913-948. [ Links ]
Zachos, J.C., Breza, J.R., and Wise, S.W. 1992. Early Oligocene ice sheet expansion on Antarctica: Stable isotope and sedimentological evidence from Kerguelen Plateau, southern Indian Ocean. Geology 20: 569-573. [ Links ]
Zachos, J.C. and Kump, L.R. 2005. Carbon cycle feedbacks and the initiation of Antarctic glaciation in the earliest Oligocene Global and Planetary Change 47: 51-66. http://dx. doi.org/10.1016/j.gloplacha.2005.01.001. [ Links ]
Zachos, J., Pagani, M., Sloan, L., and Thomas, E. 2001. Trends, rhythms, and aberrations in global climate 65Ma to present. Science 292: 686-693. [ Links ]
Zachos, J.C., Quinn, T.M., and Salamy, K.A. 1996. High-resolution (104 years) deep-sea foraminiferal stable isotope records of the Eocene-Oligocene climate transition. Paleoceanography 11: 251-266. [ Links ]
Zamaloa, M.C., Romero, E.J., and Stinco, L. 1987. Polen y esporas de la Formación La Meseta (Eoceno superior-Oligoceno) de la isla Marambio (Seymour), Antártida. 7° Congreso Argentino de Paleobotánica y Palinología, Actas: 199-203. [ Links ]
Zinsmeister, W.J. 1979. Biogeographic significance of the late Mesozoic and early Tertiary molluscan faunas of Seymour Island (Antarctic Peninsula) to the final breakup of Gondwanaland. In: Gray, J. and Boucot, A. (eds.) Historical biogeography, plate tectonics, and the changing environment, Oregon State University 349-355. [ Links ]
Zinsmeister, W.J. 1982. Late Cretaceous-Early Tertiary molluscan biogeography of southern Circum-Pacific. Journal of Paleontology 56: 84-102. [ Links ]
Zinsmeister, W.J. and DeVries, T.E. 1983. Observations on the stratigraphy of the Lower Tertiary Seymour Island Group, Seymour Island, Antarctic Peninsula. Antarctic Journal of the United States 17: 71-72. [ Links ]
Zinsmeister, W.J. and Feldmann. R.M. 1984. Cenozoic high latitude heterochroneity of southern Hemisphere marine faunas. Science 224: 281-283. [ Links ]
Zullo, V.A., Feldmann, R.M., and Wiedman, L.A. 1988. Balanomorph Cirripedia from the Eocene La Meseta Formation, Seymour Island, Antarctica. In Feldmann, R.M. and Woodburne, M.O. (eds.) Geology and Paleontology of Seymour Island, Antarctic Peninsula, Geological Society of America, Memoir 169: 459-464. [ Links ]
Recibido: 30 de noviembre, 2006
Aceptado: 11 de septiembre, 2007

