










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de la Asociación Geológica Argentina
versión impresa ISSN 0004-4822versión On-line ISSN 1851-8249
Rev. Asoc. Geol. Argent. v.63 n.4 Buenos Aires oct./dic. 2008
Archosaur evolution during the Jurassic: a southern perspective
Oliver W. M. Rauhut and Adriana Lopez-Arbarello
Bayerische Staatssammlung für Paläontologie und Geologie, München, Germany.
E-mails: o.rauhut@lrz.uni-muenchen.de, a.lopez-arbarello@lrz.uni-muenchen.de
ABSTRACT
The fossil record of archosaurs - crocodylomorphs, pterosaurs and dinosaurs - from the Jurassic of the Southern Hemisphere is critically reviewed, and its evolutionary implications are evaluated. Although several important faunas and also isolated finds are known from Gondwana, the record in total is still very patchy, and any evolutionary scenario based on this record should be seen as tentative. Compared to the Northern Hemisphere, southern archosaurs are much more poorly known, which is especially true for terrestrial crocodiles and pterosaurs. Marine crocodiles are rather well represented in south-western South America, whereas the report of terrestrial archosaurs is currently best for Africa. However, in South America, important and especially promising archosaur faunas are known from the Callovian Cañadón Asfalto and the (?)Tithonian Cañadón Calcáreo formations of Chubut province, Argentina. Early and Middle Jurassic Gondwanan archosaurs demonstrate that the faunas of that period still had a generally Pangean distribution, whereas first indications of differential archosaur evolution in the Northern and Southern Hemispheres are evident in Late Jurassic Gondwanan faunas.
Keywords: Crocodylomorphs; Pterosaurs; Dinosaurs; Gondwana; Fossil record.
RESUMEN: La evolución de los arcosaurios durante el Jurásico: una perspectiva desde el Sur. El presente trabajo incluye una revisión crítica del registro fósil de los arcosaurios - cocodrilos, pterosaurios y dinosaurios - del Jurásico del Hemisferio Sur. Se evalúan además, sus implicancias evolutivas. Aunque se conocen varias faunas importantes y hallazgos aislados, el registro, en su totalidad, es todavía incompleto y cualquier escenario evolutivo basado en este registro debería verse como tentativo. En comparación con la situación en el Hemisferio Norte, es mucho menos lo que sabemos sobre los arcosaurios del Hemisferio Sur, y esto es especialmente cierto para los cocodrilos terrestres y pterosaurios. Los cocodrilos marinos están bastante bien representados en el sudoeste de Sudamérica, en tanto que el registro de arcosaruios terrestres es actualmente mejor en África. No obstante, en Sudamérica, se conocen faunas importantes, y especialmente prometedoras, del Caloviano de la Formación Cañadón Asfalto y del Titoniano (?) de la Formación Cañadón Calcáreo, en la provincia de Chubut, Argentina. Las faunas de arcosaurios del Jurásico Temprano y Medio de Gondwana muestran una distribución pangeica, en tanto que los primeros indicios de diferenciación en la evolución de los arcosaurios de los Hemisferios Norte y Sur se evidencian en las faunas gondwánicas del Jurásico Tardío.
Palabras clave: Cocodrilos; Pterosaurios; Dinosaurios; Gondwana; Registro fósil.
INTRODUCTION
Archosaurs - crocodylomorphs, pterosaurs, dinosaurs, and several basal, mainly Triassic groups - are certainly one of the most prolific clades of Mesozoic terrestrial vertebrates, and their living representatives, modern crocodiles and birds, are important components of recent vertebrate faunas. All three derived groups (crocodylomorphs, pterosaur, dinosaurs) originated in the later stages of the Triassic, but experienced their first and most important radiations in the Jurassic. In crocodylomorphs, the Jurassic saw the origin and diversification of the Mesoeucrocodylia, the largest clade, which also includes the modern forms (Fig. 1, e.g. Benton and Clark 1988, Tykoski et al. 2002, Pol and Gasparini 2007). In pterosaurs, the early "rhamphorhynchoid" faunas reached their peak diversity during this period, and the pterydactyloids, to which almost all Cretaceous pterosaurs belong, originated and first diversified in the Upper Jurassic (Fig. 2, Kellner 2003, Unwin 2003, 2006). Dinosaurs started with only a few clades into the Jurassic, almost none of which survived this period. However, by the end of this period, basically all important lineages were established (Fig. 3, Sereno 1999a, Weishampel et al. 2004). The Jurassic thus witnessed important faunal turnovers in all major clades of archosaurs and is therefore of greatest importance for our understanding of the evolutionary history of these groups.

Figure 1: Phylogeny of crocodiles showing the origin and first diversification of the Mesoeucrocodylia during the Jurassic. Based on Pol and Gasparini (2007). Dotted lines indicate uncertain limits of the stratigraphic range of the bearing sediments of a certain taxon.
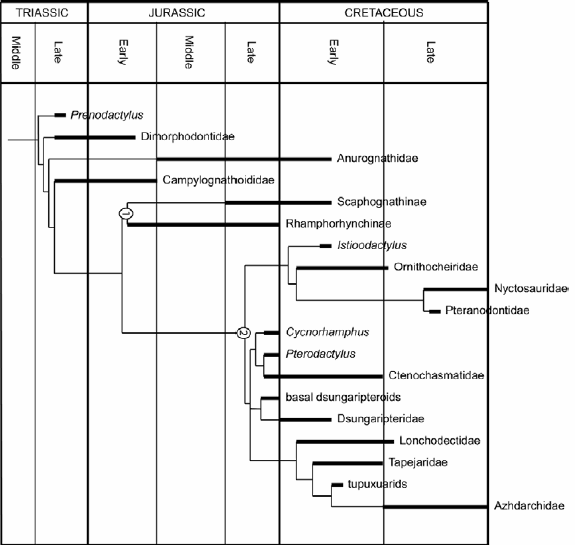
Figure 2: Phylogeny of pterosaurs showing the origin and first diversification of the two main lineages, Ramphorhynchidae (node 1) and the Pterodactyloidea (node 2) during the Jurassic (based on Unwin 2006).
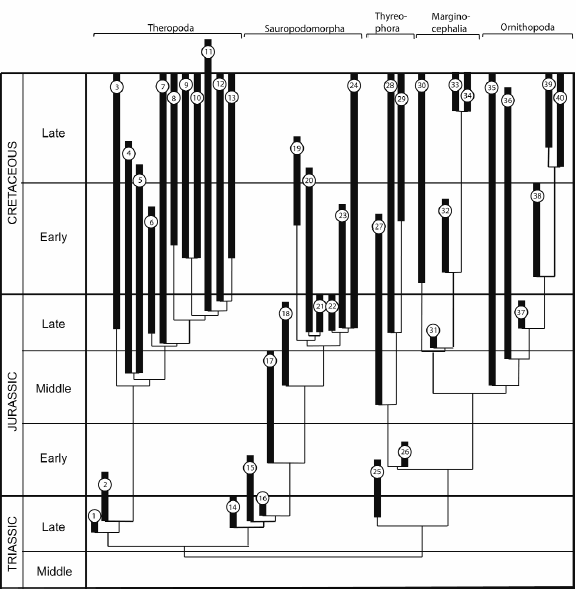
Figure 3: Phylogenetic relationships and stratigraphic distribution of the main dinosaur clades, showing the diversification of almost all main dinosaur lineages during the Jurassic. Minor clades or single taxa have been considered if of stratigraphic importance. Composite cladogram, Theropoda (based on Rauhut 2003), Sauropodomorpha on Wilson (2002) and Smith and Pol (2007), and Ornithischia on Weishampel, Dodson and Osmólska (2004) and Butler et al. (2008). 1) Herrerasauridae, 2) Coelophysoidea, 3) Ceratosauria, 4) Allosauroidea, 5) Spinosauroidea, 6) Coeluridae, 7) Tyrannosauroidea, 8) Ornithomimosauria, 9) Oviraptorosauria, 10) Therizinosauroidea, 11) Avialae, 12) Dromaeosauridae, 13) Troodontidae, 14) Plateosauridae, 15) Massospondylidae, 16) Antetonitrus + Lessemsaurus, 17) Cetiosauridae (sensu Upchurch et al. 2004), 18) Omeisauridae, 19) Rebbachisauridae, 20) Dicraeosauridae, 21) Diplodocidae, 22) Camarasaurus spp., 23) Brachiosauridae, 24) Titanosauria, 25) Heterodontosauridae, 26) Emausaurus, 27) Stegosauria, 28) Ankylosauridae, 29) Nodosauridae, 30) Pachycephalosauria, 31) Yinlong, 32) Psittacosauridae, 33) Centrosaurinae, 34) Chasmosaurinae, 35) "Hypsilophodontidae" (non-monophyletic group of basal ornithopods, the interrelationships of which are still unclear), 36) Dryosauridae, 37) Camptosaurus spp., 38) Iguanodontidae, 39) Hadrosaurinae, 40) Lambeosaurinae.
It is still rather unclear what caused these important changes in Jurassic archosaur faunas. In the Jurassic the initial break-up of the supercontinent of Pangea took place, during which the western Tethian and northern Atlantic oceans opened and finally separated Gondwana from the northern continents in the Late Jurassic (Scotese 1991, Smith et al. 1994, Scotese et al. 1999, Ford and Golonka 2003). Vicariance as a consequence of this separation has been proposed as an important aspect of dinosaurian radiations (Russell 1993, Sereno 1999a, b), and evidence for continent-wide vicariance was found in empirical analyses of dinosaurian biogeography (Upchurch et al. 2002). Furthermore, these changes in geography brought about other modifications, such as the end of the megamonsoonal climate that had dominated large parts of Pangea in the Triassic and early stages of the Jurassic (Parrish 1993, Scotese et al. 1999). Archosaur radiations and faunal turnovers certainly have to be seen in the context of these environmental changes (see Forster 1999), but even the timing and the modes of these evolutionary events are still largely unknown, owing to the rather poor Jurassic fossil record in many groups. Furthermore, our current ideas about Jurassic faunal changes are almost entirely based on the fossil record of the Northern Hemisphere, making it basically impossible to check possible effects of the separation of Gondwana on the evolutionary history of the different groups.
In the current paper, we critically evaluate the Jurassic archosaur fossil record of the Southern Hemisphere and its importance for our current understanding of Jurassic archosaur evolution. In the following, the colloquial term "crocodile" is used to refer to crocodylomorphs in general. True members of the Crocodylia are so far unknown from the Jurassic and only appear in the Cretaceous.
Institutional abbreviations: MACN: Museo Argentino de Ciencias Naturales "Bernardino Rivadavia", Buenos Aires, MB: Museum für Naturkunde, Berlin, MPEF: Museo Paleontológico Egidio Feruglio, Trelew, Argentina.
JURASSIC ARCHOSAURS FROM THE SOUTHERN HEMISPHERE
Jurassic terrestrial archosaurs from the Southern Hemisphere are known only from a handful of localities (Fig. 4), and most finds are isolated and/or fragmentary. In the following, the Gondwanan archosaur fossil record will be briefly outlined and compared with that from the Northern Hemisphere. Only the body fossil record will be dealt with in detail here. Although tracks can give important insights into faunal compositions and other aspects of archosaur biology, they are usually less diagnostic than body fossils on a lower taxonomic level. Thus, the, for some stages, more numerous track record is only taken into consideration in cases where it inidcates the presence of otherwise not represented groups.

Figure 4: Palaeogeographic maps (modified from Mollewide Plate Tectonic Maps available at http://jan.ucc.nau.edu/~rcb7/globaltext2.html) indicating the main Jurassic archosaur localities in Laurasia and Gondwana. a) Early Jurassic: Elliot and Clarens formations, upper Stormberg Group, Upper Karoo Supergroup, South Africa (1), Toundoute continental series, High Atlas, Morocco (2), Kota Formation, Prahita-Godavari valley, India (3), Hanson Formation, Transantarctic Mountains, Antarctica (4), Kayenta Formation and Navajo Sandstone and equivalent beds, USA (5), Newark Supergroup, North America (6), Posidonia Shale in Germany and comparable strata in England and France (7), Lufeng Formation, China (8). b) Middle Jurassic: Durham Downs, Queensland, Australia (1), Tilougguit Formation and equivalent beds of Morocco (2), Isalo III Formation, Madagascar (3), Cañadón Asfalto Formation, Chubut, Argentina (4), Lautaro Formation, Chile (5), various marine localities in England and France (6), Balabansai Formation, Kirghizia (7), Lower Shaximiao Formation, Sichuan, China (8). C. Late Jurassic: Vaca Muerta Formation and equivalent beds, Neuquén Basin, Argentina (1), Cañadón Calcáreo Formation, Chubut, Argentina (2): Tendaguru Beds, Tanzania (3), Kadsi Formation, Zimbabwe (4), Morrison Formation, USA (5), various localities in Portugal (6), Solnhofen Limestone and equivalent beds in Germany (7), Shishougou Formation, Xinjiang, China (8), Upper Shaximiao Formation of China (9).
Early Jurassic
The Early Jurassic Gondwanan archosaur record is strongly biased towards Africa, with one important fauna known from India, and isolated occurrences from South America and Antarctica (Fig. 4a).
The most important Lower Jurassic Gondwanan vertebrate faunas are certainly those of the Upper Karoo Supergroup, namely the upper Stormberg Group, comprising the Upper Elliot and Clarens formations [Fig. 4a(1), see Knoll 2005], and contemporaneous units in Zimbabwe, such as the Forest Sandstone. Several taxa of crocodylomorphs have been described especially from the Upper Elliot and Clarens formations, but many of them seem to be junior subjective synonyms of other taxa (see review in Knoll 2005). Taxa currently regarded as valid (Knoll 2005) include the 'sphenosuchians' Sphenosuchus (Haughton 1915, Walker 1990) and Litargosuchus (Clark and Sues 2002), and the protosuchids Protosuchus (Gow 2000) and Orthosuchus (Nash 1968, Pol et al. 2004). Pterosaur remains are currently unknown from the Elliot and Clarens Formations, though this group might be represented by tracks (Knoll 2005).
Dinosaurs are certainly the best-represented and best-known archosaur group from the upper Stormberg Group and equivalent strata in southern Africa (South Africa, Lesotho and Zimbabwe). Since the first description of the "prosauropod" Massospondylus from the Upper Elliot Formation by Owen (1854), a plethora of dinosaurs, mainly basal sauropodomorphs, have been described from these units. However, most of these taxa are considered to be junior synonyms of Massospondylus (Cooper 1981), which remains the best known Early Jurassic "prosauropod" from the Southern Hemisphere, and is well-documented from the upper Stormberg (e.g. Gow 1990, Gow et al. 1990, Sues et al. 2004, Yates and Vasconcelos 2005, Barrett and Yates 2006), including remains of eggs and embryos (Reisz et al. 2005), but also known from other Early Jurassic units in southern Africa, mainly Zimbabwe (Cooper 1981). However, although Massospondylus certainly represents the most common sauropodomorph in the upper Stormberg, at least one other "prosauropod" and two basal sauropods are also present in this unit (Barrett 2004, Yates et al. 2004, 2007), and the basal sauropod Vulcanodon has been described from strata of Early Jurassic age in Zimbabwe (Raath 1972, Cooper 1984). Theropod dinosaurs from the Early Jurassic of southern Africa include the small coelophysoid Coelo-physis rhodesiensis, which represents one of the best-known Early Jurassic theropods (Raath 1969, 1977, 1985, 1990, Galton 1971, Chinsamy 1990, Bristowe and Ra-ath 2004). Most material of this theropod comes from the Forest Sandstone of Zimbabwe (Raath 1969, 1977), but the taxon also seems to be represented in the Upper Elliot Formation (Raath 1980). Recently, Yates (2005) described a new taxon of theropod from the Upper Elliot Formation, Dracovenator. Recent fieldwork indicates, however, that this specimen might be derived from the basal part of the overlying Clarens Formation (OR, pers. obs. 2008).
The Upper Elliot Formation has yielded the most diverse Early Jurassic ornithischian fauna known. Taxa represented include the heterodontosaurids Heterodontosaurus, Lycorhinus and Abrictosaurus (Haughton 1924, Crompton and Charig 1962, Hopson 1975, see also Knoll 2005), the probably basal thyreophoran Lesothosaurus (Thulborn 1970, 1972, Galton 1978, Sereno 1991, Butler et al. 2008), and the basal cerapodan Stormbergia (Knoll 2002a, b, Butler 2005).
Apart from the Karoo basin, Early Jurassic archosaurs from Africa have only been described from Morocco [Fig. 4a(2)]. Material from the Toundoute continental series of the High Atlas include the basal sauropod Tazoudasaurus (Allain et al. 2004) and the probable abelisauroid theropod Berberosaurus (Allain et al. 2007). Furthermore, Taquet (1985) reported and figured a partial theropod hindlimb from an unnamed Early Jurassic unit in Morocco.
Outside Africa, the most important Early Jurassic Gondwanan archosaur localities are found in the Prahita-Godavari valley of eastern India, mainly from the Kota Formation [Fig. 4a(3)]. However, it might be noted that, although the Kota Formation is generally considered to be Early Jurassic in age (Bandyopadhyay et al. 2002, Weishampel et al. 2004, Bandyopadhyay and Sengupta 2006), there is some uncertainty about the dating of this unit, and it might be as young as Early Cretaceous (Prasad et al. 2004, Prasad and Manhas 2007).
Again, it was Owen (1854) who first reported archosaur remains from this area, from levels now assigned to the Kota Formation. The remains described by Owen (1854) represent a marine, thalattosuchian crocodile (Kutty et al. 1987, Bandyopadhyay et al. 2002), and are still the only crocodile remains described from the Gondwana sequence of this basin. However, it might be noted that a partial skull from the Lower Jurassic Kota Formation of India, referred to an ankylosaurian dinosaur by Nath et al. (2002: pl. 1a-e), most probably represents a crocodilian, as indicated by the pitted bone surface and the general shape of the fragments. Judged by the general morphology, this fragment most probably represents a terrestrial crocodile, but further interpretation has to await a detailed reanalysis of this specimen.
Jain (1974) described the campylognathoid pterosaur Campylognathoides indicus from the upper levels of the Kota Formation, which still represents the oldest pterosaur known from the Southern Hemisphere. Dinosaurs from the Kota Formation were described by Jain et al. (1975, 1979) and Yadagiri (1988, 2001), who named and described the sauropods Barapasaurus and Kotasaurus, respectively. The material referred to the latter taxon includes at least one further taxon of sauropod (OR, pers. obs. 2005). Nath et al. (2002) described fragmentary dinosaur remains from the Kota Formation and referred them to an ankylosaurian thyreophoran, which would represent the oldest record of this group. However, this identification is questionable, since more than one taxon is represented in the material (see comments on a probable crocodylomorph included in this material above), but some of the scutes described and figured by Nath (2002: pl. 1fj) might indeed represent a thyreophoran. Apart from these remains, theropods are represented in the Kota Formation by undiagnostic material (Bandyopadhyay et al. 2002, Prasad and Manhas 2007).
Just recently, Kutty et al. (2007) described two taxa of sauropodomorphs, the "prosauropod" Pradhania and the probable basal sauropod Lamplughasaura, and mentioned the presence of a third sauropodomorph taxon in the Upper Dharmaran Formation (Sinemurian), which underlies the Kota Formation.
Bradmeister et al. (2006) described a vertebrate fauna, including probable theropod and sauropod teeth, from the Isalo II Formation of Madagascar. Although they stated in the abstract that the vertebrate fauna indicates an Early Jurassic age for this fauna, the faunal association, including probable colobodontid fishes and aetosaurian and phytosaurian archosaurs, is more indicative of a Late Triassic age.
Only fragmentary archosaur material has been described from the Early Jurassic of South America so far. Huene (1927) described Lower Jurassic crocodile remains from Mendoza province in Argentina, South America. The material consists of two dorsal vertebrae, which he made the type of a new species of the thalattosuchian Steneosaurus, Steneosaurus gerthi, but this species is currently considered to be a nomen dubium (Pol and Gasparini 2007). An articulated partial metatarsus from the Sinemurian of Alto de Varas in the Cordillera de Domeyko in Chile was described by Chong-Diaz and Gasparini (1972) and also referred to a thalattosuchian (see also Gasparini 1985, Gasparini et al. 2000). Early Jurassic dinosaur remains have been described from the probably Lower Jurassic La Quinta Formation of Venezuela. Russell et al. (1992) and Sánchez-Villagra and Clark (1994) reported ornithischian material from this formation, which they referred to the southern African genus Lesothosaurus, and Moody (1997) described some unidentifiable theropod teeth. However, a recent revision of this material shows that the ornithischian material is not referable to Lesothosaurus, and that a further taxon of probable basal saurischian was present (Barrett et al. 2008). Martínez (1999) referred some material from the probably Early Jurassic Cañón de Colorado Formation of the province of San Juan, Argentina, to Massospondylus, though reexamination of this material indicates that it represents a new genus (Pol, pers. com. 2008). Another basal sauropodomorph has recently been reported from the Las Leoneras Formation of the province of Chubut, Argentina (Pol and Garrido 2007), but these remains have not been described in detail yet.
A noteworthy Early Jurassic vertebrate fauna, including dinosaurs, from Hanson Formation in the Transantarctic Mountains of Antarctica [Fig. 4a(4)] was made known by Hammer and Hickerson (1994). This southernmost archosaur fauna discovered so far includes a humerus of a pterosaur, the theropod Cryolophosaurus, the basal sauropodomorph Glacialisaurus, and an undescribed sauropod (Hammer and Hickerson 1994, 1999, Smith and Pol 2007, Smith et al. 2007a, b).
Middle Jurassic
From Australia, only a distal theropod tibia and some fragmentary sauropod remains of Middle Jurassic age have been reported so far, and were named Ozraptor and Rhoetosaurus, respectively (Longman 1926, 1927, Long & Molnar 1998). Ozraptor comes from the shallow marine Colalura Sandstone (Bajocian) of western Australia, whereas Rhoetosaurus was found in terrestrial deposits at Durham Downs in Queensland [Fig. 4b(1)].
Middle Jurassic strata in Africa have yielded a few dinosaur remains from the Bathonian-Callovian Tilougguit Formation and equivalent beds of Morocco [Fig. 4b(2)], including the sauropods "Cetiosaurus" mogrebiensis and Atlasaurus imelakei (Lapparent 1955, Monbaron et al. 1999), as well as the sauropod Chebsaurus algeriensis from indeterminate Middle Jurassic rocks of Algeria (Mahammed et al. 2005). Isolated vertebrae from the Bathonian of Morocco that were originally described as a theropod, Megalosaurus mersensis, by Lapparent (1955), probably represent a thalattosuchian crocodile (Chabli 1985). However, theropods are represented in the Bathonian of Morocco by isolated and unidentified teeth (Monbaron et al. 1999). Remains of the thalattosuchian Steneosaurus are found in the Bathonian of Tunisia (Mueller-Töwe 2006).
The Bathonian Isalo III Formation of Madagascar [Fig. 4b(3)] has yielded a rich, though mainly fragmentary archosaur fauna. Taxa reported include a possible atoposaurid crocodilian, a "rhamphorhynchoid" pterosaur, indeterminate ornithischians, theropods, all represented by teeth only (Flynn et al. 2006), and the sauropods Lapparentosaurus madagascariensis and Archaeodontosaurus descouensi (Bonaparte 1986a, Buffetaut 2005, Maganuco et al. 2005). Flynn et al. (2006) also listed the sauropod species Bothriospondylus madagascariensis in the Isalo III Formation, but the material of this taxon was referred to the new genus Lapparentosaurus by Bonaparte (1986a, see also Buffetaut 2005), and furthermore referred several teeth to an unspecified neosauropod, though they noted that the apparent differences to the teeth of Archaeodontosaurus might be due to preservation, so this assignment is questionable. Thus, only two sauropod taxa are recognized from this formation, Lapparentosaurus and Archaeodontosaurus. Maganuco et al. (2005) referred theropod teeth to several different groups of theropods, including advanced coelurosaurs, but Flynn et al. (2006) considered all this material as Theropoda indet. In addition to the fauna from the Isalo III Formation, material of the thalattosuchian Steneosaurus has been reported from the Callovian of Madagascar (Mueller-Töwe 2006).
Middle Jurassic archosaurs from South America are mainly known as isolated finds, with the exception of the fauna from the Cañadón Asfalto Formation of Chubut, Argentina [Figs. 4b(4), 5]. This Callovian unit has yielded so far unidentified and unstudied crocodile remains, as well as a diverse, but also still unstudied pterosaur fauna, comprising at least three taxa, possibly including the oldest pterodactyloid known from Gondwana (Rauhut et al. 2001, Unwin et al. 2004, Rauhut, unpublished data). Most of the remains come from a single pterosaur bonebed, which has yielded disarticulated, but very well-preserved and three-dimensional material (Fig. 6), which is very rare in Jurassic pterosaurs, potentially making this the most important Middle Jurassic pterosaur locality globally. Most noteworthy, however, is the dinosaur fauna, which is the second most diverse Middle Jurassic dinosaur fauna known, following the Bathonian Laurasian fauna from the Lower Shaximiao Formation of China (Peng et al. 2005). Taxa described include the sauropod Patagosaurus and Volkheimeria, and the theropods Piatnitzkysaurus and Condorraptor (Bonaparte 1979, 1986b, Rauhut 2005a), and at least one further taxon of sauropod is present (Rauhut 2003a). Ornithischian dinosaurs have not been described from this formation so far, but are represented by an isolated ungual from the locality of Cerro Cóndor (Fig. 7) and some teeth from the microvertebrate locality Queso Rallado.

Figure 5: Schematic stratigraphic column of the archosaurbearing units of the Middle Jurassic Cañadón Asfalto Formation and the Late Jurassic Cañadón Calcáreo Formation of Chubut province, Argentina.

Figure 6: Pterosaur material from the Middle Jurassic Cañadón Asfalto Formation, Chubut province, Argentina. a) Left mandible in lateral view. b) Left mandible in medial view. c) Right humerus in anterior view. d) Proximal end of wing phalanx 1. Scale bars: 1 cm.

Figure 7: Isolated pedal phalanx of a probably basal cerapodan ornithischian from the Middle Jurassic Cañadón Asfalto Formation of Chubut, Argentina, MACN CH 223. a) Dorsal view. b) Lateral view. c) Ventral view. Scale bar: 1 cm.
Apart from the fauna of the Cañadón Asfalto Formation, the most important archosaur remains from South America are marine crocodiles from western Argentina and Chile, which include the oldest record of the genus Metriorhynchus from the Bajocian Lautaro Formation of Chile [Fig. 4b(5), Gasparini et al. 2000]. Two species of Metriorhynchus were described from the Callovian of Chile, Metriorhynchus casamiquelai from an unnamed unit in the Quebrada Sajasa in northern Chile (Gasparini and Chong Díaz 1977, Gasparini 1985), and Metriorhynchus westermanni from the Mina Chica Formation of northern Chile (Gasparini 1980, 1985) [Fig. 4b(5)]. The latter was regarded as a synonym of the former by Gasparini et al. (2000), but new material shows that it is a valid taxon (Gasparini et al. in press). Further thalattosuchian oc-currences in Chile were discussed by Gasparini et al. (2000). Recently, Gasparini et al. (2005) reported a fragmentary skull of Metriorhynchus from the Bathonian of the Los Molles Formation of Neuquén province, Argentina.
Apart from the fauna of the Cañadón Asfalto Formation, Middle Jurassic dinosaurs from South America are only known from two localities in Argentina. Cabrera (1947) described fragmentary sauropod remains from Pampa de Agnia, Chubut, as Amygdalodon patagonicus. The material comes from the Cerro Carnerero Formation, which is most probably of Toarcian-Aalenian age, and thus represents the oldest Middle Jurassic archosaur-bearing unit in South America.Additional material from the same locality was referred to this taxon and even thought to represent the same individual as the type by Casamiquela (1963), but Rauhut (2003b) showed that more than one individual is represented by these remains. Nevertheless, Amygdalodon still represents the oldest sauropod record from South America. The recent find of a partial sauropod sacrum from the Upper Aalenian Remoredo Formation of Neuquén, Argentina (Salgado and Gasparini 2004), demonstrates that these animals were already widely distributed in South America at the beginning of the Middle Jurassic.
Only isolated finds of fragmentary dinosaur material have been reported from the Middle Jurassic of India so far (see overview in Moser et al. 2006). Most of the material reported, which comes from different units in north-western India and ranges in age from the Aalenian-Bajocian to the Callovian, seems to represent sauropod dinosaurs, but only a few fragmentary remains from Khadir Island have been described in detail so far (Moseret al. 2006). Mainly on the basis of the morphology of a proximal fibula, Moseret al. (2006) concluded that some of this material represents a camarasauromorph sauropod, which would represent the oldest record of a neosauropod from the Southern Hemisphere. However, better material is needed to substantiate this claim, especially since the shape of the proximal articular surface of the element in question is rather reminiscent of sauropod metacarpals II-IV (see e.g. Ostrom and McIntosh 1966: pl. 56-58, 60-61), as is its size in comparison to other metacarpal fragments found at the same locality (Moser et al. 2006: figs. 4.1, 4.2).
Late Jurassic
Although the Late Jurassic is often seen as a well-known stage in the history of terrestrial vertebrates, mainly due to numerous localities in the Northern Hemisphere, such as the famous Morrison Formation of the western USA, very little is still known about Late Jurassic terrestrial vertebrates from Gondwana, and only few localities yielding archosaur material are known.
Late Jurassic archosaurs from South America were mainly known from marine crocodiles until recently. With the exception of one vertebra from Chile, reported by Gasparini (1985), all marine crocodile remains from the Late Jurassic of South America come from the Neuquén Basin in Argentina, from the Tithonian Vaca Muerta Formation and equivalent beds [Fig. 4C(1), Gasparini 1985, 1992, 1996, Gasparini and Fernandez 1997, Gasparini et al. 2006]. Taxa reported from these units include the metriorhynchids Geosaurus araucanensis, Metriorhynchus potens, and Dakosaurus andiniensis.
Pterosaurs are still very poorly known from the Late Jurassic of South America. Casamiquela (1975) reported a partial skeleton of a supposed theropod dinosaur, Herbstosaurus pigmaeus, found in an isolated boulder, which was later reinterpreted as a pterosaur, demonstrably from Upper Jurassic (Tithonian) strata of the Vaca Muerta Formation in Neuquén province, Argentina (Bonaparte 1981, Codorniú and Gasparini 2007). Further, mainly fragmentary fossils have been described since, including Tithonian pterosaur remains from the same beds in Argentina (Gasparini et al. 1987, Codorniú et al. 2006) and from coastal sediments of Chile (Martill et al. 2000), although the latter might be Early Cretaceous in age.
Dinosaurs were almost unknown from the Late Jurassic of South America until recently, with the exception of a single sauropod vertebra from unspecified Late Jurassic sediments of northern Colombia (Langston and Durham 1955) and undescribed dinosaur remains from the Late Jurassic of Chile, reported by Chong Díaz and Gasparini (1976). More recently, an isolated sauropod knee joint was reported from the Kimmeridgian Tordillo Formation of Neuquén, Argentina (García et al. 2003), and theropod teeth have been found in the Tacurembó Formation in Uruguay (Perea et al. 2003), which is probably also Late Jurassic in age (Soto, pers. comm. 2007). Salgado et al. (2008) described some theropod remains from the Tithonian Toqui Formation of southern Chile. This material, which includes at least one taxon of tetanurans, thus represents the southernmost Late Jurassic dinosaur body fossils known.
The most important, but still rather poorly studied Late Jurassic dinosaur fauna reported from South America so far is that of the Cañadón Calcáreo Formation of Chubut province, Argentina [Figs. 4c(2), 5]. The age of this unit is still somewhat uncertain: whereas a radiometric date puts its basal part in the Tithonian (see Rauhut 2006a), new palynological evidence seems to indicate a lowermost Cretaceous age for at least parts of the formation (Silva Nieto et al. 2007). More geological and stratigraphic work is needed to tie down the stratigraphic position of the dinosaurs from this formation. The first dinosaur described from the Cañadón Calcáreo Formation was Tehuelchesaurus, though originally thought to be from the Middle Jurassic Cañadón Asfalto Formation (Rich et al. 1999). This taxon was first considered to be closely related to the Middle Jurassic Chinese sauropod Omeisaurus (Rich et al. 1999, Upchurch et al. 2004), but new data indicates that it is a member of the Macronaria and close to or within the Titanosauriformes (Rauhut 2002, Rauhut et al. 2005, Carballido et al. 2007). More recently, the dicraeosaurid Brachytrachelopan mesai (Fig. 8) and an unidentified brachiosaurid were reported from the same formation (Rauhut et al. 2005, Rauhut 2006a).

Figure 8: Brachytrachelopan mesai from the (?)Tithonian Cañadón Calcáreo Formation, Chubut province, Argentina. a) Vertebral column as preserved (MPEF PV 1716). b) Outline reconstruction showing preserved elements.
The most important Late Jurassic terrestrial vertebrate fauna from Gondwana known so far comes undoubtedly from the famous Tendaguru Beds of Tanzania [Fig. 4C(3), Maier 2003]. Archosaurs from three different levels within the Tendaguru Beds include crocodile teeth, isolated pterosaur remains, and numerous taxa of dinosaurs (Janensch 1914, Russell et al. 1980, Aberhan et al. 2002, Maier 2003). The ages of the three different beds are probably Oxfordian for the Lower Saurian Bed, Kimmeridgian-Early Tithonian for the Middle Saurian Bed, and Late Tithonian to earliest Cretaceous for the Upper Saurian Beds (Aberhan et al. 2002).
Crocodiles are so far only represented by isolated teeth from the Upper Saurian Beds, which seem to be similar to those of the Early Cretaceous neosuchian Bernissartia (Maier 2003: 315). Pterosaurs are represented by numerous, but mainly isolated and often fragmentary elements. Most pterosaur remains come from the Upper Saurian Beds, though they are also present in the Middle Saurian Beds (Reck 1931). Reck (1931) described four species on the basis of this material, though mainly based on uncomparable remains: Rhamphorhynchus tendagurensis, Pterodactylus maximus, Pterodactylus brancai, and Pterodactylus arningi, all from the Upper Saurian Beds. However, the validity of all of these species is highly questionable (Unwin and Heinrich 1999). Galton (1980a) referred another specimen, probably from the Middle Saurian Bed, to P. brancai and transferred this species to the Early Creatceous Asian genus Dsungaripterus. Interestingly, Unwin and Heinrich (1999) described a pterosaurian jaw fragment from the Middle Saurian Beds as a new species, Tendaguripterus recki, which they also referred to the Dsungaripteroidea. However, since the type and referred specimens of "Dsungaripterus" brancai are tibiotarsi, more material is necessary to establish whether only one, or two taxa of dsungaripteroids were present in the Tendaguru fauna.
The dinosaur fauna from Tendaguru is dominated by sauropods, which are represented by at least seven species, the brachiosaurid Brachiosaurus brancai, the probably basal titanosaurs Janenschia robusta and Tendaguria tanzaniensis (which might be conspecific), the dicraeosaurids Dicraeosaurus hansemanni and D. sattleri, and the diplodocids Tornieria africana and Australodocus bohetii (Janensch 1914, 1929, Bonaparte et al. 2000, Upchurch et al. 2004, Remes 2006, 2007). Theropod dinosaurs are represented by the basal neoceratosaurElaphrosaurus bambergi (Fig. 9) and at least six more taxa (Janensch 1920, 1925, Rauhut 2005b, c, 2006b). The theropod fauna seems to be dominated by basal tetanurans and neoceratosaurs, whereas coelurosaur remains could not be identified so far (Rauhut 2005b, c, 2006b). Ornithischians are much less diverse and are represented only by the stegosaur Kentrosaurus aethiopicus and the small ornithopod Dysalotosaurus lettowvorbecki (Hennig 1925, Janensch 1955). Dinosaur remains are not evenly distributed in all levels at Tendaguru. The Lower Saurian Beds have only yielded a few theropod teeth attributed to "Megalosaurus" ingens (Janensch 1925) and fragmentary sauropod remains, possibly representingBrachiosaurus (Aberhan et al. 2002). The Middle and Upper Saurian Beds are much richer in vertebrate remains, but differ slightly in their dinosaur fauna. The Middle Saurian Beds contain remains of the sauropods Brachiosaurus brancai, Dicraeosaurus hansemanni, Tornieria africana, and Australodocus bohetii (Aberhanet al. 2002, Remes 2006, 2007), the theropodsElaphrosaurus bambergi and at least four more taxa, including two tetanurans and two ceratosaurs (Janensch 1925, Rauhut, unpublished data). Both ornithischians known from Tendaguru, Kentrosaurus aethiopicus and Dysalotosaurus lettowvorbecki, are also present in the Middle Saurian Beds (Aberhan et al. 2002). The Upper Saurian Beds have yielded the sauropods Brachiosaurus brancai, Janenschia robusta, Tendaguria tanzaniensis, Dicraeosaurus sattleri, and Tornieria africana, at least two ceratosaurian and two tetanuran theropods (Rauhut 2005c, unpublished data), and the stegosaur Kentrosaurus aethiopicus (Aberhan et al. 2002).
Figure 9:
Outline reconstruction showing preserved elements of the ceratosaurian theropod Elaphrosaurus from the Middle Saurian Beds (Kimmeridgian) of Tendaguru, Tanzanía, based on the holotype MB dd unnumbered. Scale bar: 50 cm.
Another dinosaur fauna reported from the Upper Jurassic of Africa comes from the Kadsi Formation of Zimbabwe [Fig. 4c(4), Raath and McIntosh 1987], but the material collected so far is fragmentary and identifications of taxa have to be treated with caution. Raath and McIntosh (1987) identified at least four, possibly five sauropod taxa from these beds, and referred them to the genera Brachiosaurus, Janenschia (=Tornieria in the cited work, for the convoluted taxonomic history of this genus see Wild 1991), Dicraeosaurus, Barosaurus, and, possibly, Camarasaurus. However, only the remains referred to Brachiosaurus might be diagnostic on generic level, the rest of the material would better be understood as basal titanosaur indet., two diplodocoids indet. (possibly a dicraeosaurid and a diplodocid), and an unidentified sauropod. Two theropod femora, tentatively identified as ?allosaurid by Raath and McIntosh (1987: 109, 113 fig. 4) might rather represent a large neoceratosaur, as indicated by the low, aliform lesser trochanter with a well-developed trochanteric shelf.
A further Late Jurassic unit from Africa has yielded archosaur remains, the Mugher Mudstone Formation of Ethiopia, which is of probably Tithonian age (Goodwin et al. 1999). Archosaur fossils from this formation include a possibly goniopholid crocodile and theropod, sauropod, and probable ornithischian teeth (Werner 1995, Goodwin et al. 1999). One theropod tooth fragment was identified as cf. Acrocanthosaurus by Goodwin et al. (1999: 735-736), but the only character given in support of this assignment (chisel-shaped denticles) actually represents the plesiomorphic condition in theropods, so this specimen should be regarded as Theropoda indet. From slightly older levels close to the city of Harrar in Ethiopia (Oxfordian-Kimmeridgian), Huene (1938) described a partial mandibular symphysis as a new pliosaur, Simolestes nowackianus, but this specimen was later assigned to the thalattosuchian crocodyliform Machimosaurus by Bardet and Hua (1996), thus representing the oldest record of this group in Africa.
JURASSIC ARCHOSAURS FROM THE NORTHERN HEMISPHERE
Jurassic archosaurs from the Northern Hemisphere are known from a number of localities.
In the Early Jurassic, the probably most important Laurasian localities are the Kayenta Formation and Navajo Sandstone and equivalent beds of the western USA [Fig. 4A(5)], the Newark Supergroup of eastern North America [Fig. 4a(6)], the Posidonia Shale in Germany and comparable strata in England and France [Fig. 4a(7)], and the Lufeng Formation of south-eastern China [Fig. 4a(8)].
In the Middle Jurassic, important faunas are known especially from various, mainly marine units in England and France [Fig. 4b(6)] and the Lower Shaximiao Formation of Sichuan, China [Fig. 4b(8)]. Furthermore, the Upper Shaximiao Formation of China [Fig. 4c(9)], often considered to be Late Jurassic in age (e.g. Weishampel et al. 2004, Peng et al. 2005), might also be of late Middle Jurassic (Callovian) age (Averianov et al. 2005, 2007). Averianov et al. (2005) cited the supposed close relationships of the Chinese sauropod Omeisaurus with the supposedly Middle Jurassic South American sauropod Tehuelchesaurus as evidence for a Middle Jurassic age, but, as noted above, Tehuelchesaurus neither comes from Middle Jurassic sediments, nor is it closely related to Omeisaurus. However, Averianov et al. (2007) further mentioned invertebrate evidence for a Middle Jurassic age the Upper Shaximiao Fm. On the other hand, the fauna of the Upper Shixamiao Formation shares the dinosaur genera Sinraptor and Mamenchisaurus with the Shishougou Formation of Xinjiang, China, which is considered to be Oxfordian in age (Xu et al. 2006a, Xu, pers. comm. to OR 2006), so the age of this unit is best considered to be uncertain.
The most important Late Jurassic faunas are certainly those of the Morrison Formation of the western USA [Fig. 4c(5)], various Kimmeridgian units in Portugal [Fig. 4c(6)], the Solnhofen Limestone in southern Germany [Fig. 4c(7)], and various units in China, including the Shishougou Formation of Xinjiang [Fig. 4c(8)].
Early Jurassic
In the Northern Hemisphere, crocodiles are well known from marine forms, as in the Southern Hemisphere. The oldest northern records of thalattosuchians are contemporaneous to those in the Southern Hemisphere and were found in the Sinemurian of France (see Gasparini et al. 2000, Mueller-Töwe 2006). Thalattosuchians are especially well-represented in the Toarcian of the Posidonia Shale and contemporaneous sediments in Europe, from where the genera Pelagosaurus, Platysuchus and Steneosaurus are known, the latter with several species (Mueller-Töwe 2006). In this stage, fragmentary thalattosuchian remains are also known from North America and China (Mueller-Töwe 2006).
Terrestrial Early Jurassic crocodiles are known from the western US, from where several taxa of "sphenosuchians" (the monophyly of which is still controversial, see Clark and Sues 2002 and Sues et al. 2003) and protosuchids and the oldest known goniopholid, Calsoyasuchus, were reported (Colbert and Mook 1951, Crompton and Smith 1980, Sues et al. 1994, Clark and Sues 2002, Tykoski et al. 2002). A similar faunal composition, with protosuchids and "sphenosuchians" is also found in the McCoy Brook Formation of the Newark Supergroup of eastern North America (Shubin et al. 1994, Sues et al. 1996) and the Lufeng Formation of China (Simmons 1965, Luo and Wu 1994, Wu and Sues 1996, Harris et al. 2000).
Pterosaurs are known from a number of Early Jurassic Laurasian localities, though most remains are fragmentary, such as the type of Rhamphinion from the Kayenta Formation of the western USA (Padian 1984, Sues et al. 1994). However, such specimens might provide important anatomical information as well (e.g. Clark et al. 1998). Well-preserved pterosaur remains have been found in the Posidonia Shale of Germany (see Padian and Wild 1992) and the Lias of Lyme Regis in England (see Padian 1983). These remains represent the families Dimorphodontidae and Campylognathidae.
Dinosaurs are also known from a variety of Early Jurassic localities in the Northern Hemisphere. Taxa recorded include basal thyreophoran ornithischians from western North America (Colbert 1981, Padian 1989), Europe (Owen 1863, Haubold 1990) and Asia (Simmons 1965, Dong 2001, Norman et al. 2007). Theropods are represented by coelophysoids in North and Central America (Tykoski and Rowe 2004, Munter and Clark 2005, Tykoski 2005), Europe (Carrano and Sampson 2004) and Asia (Irmis 2004) and "dilophosaurs" in North America (Welles 1954, 1984) and Asia (Hu 1993, see also Smith et al. 2007). A therizinosauroid theropod reported from the Lufeng Formation (Xu et al. 2001) is of doubtful affinities, and might represent a prosauropod (Rauhut 2003c). Basal sauropodomorphs ("prosauropods") are well-documented from the Lufeng Formation of China (Young 1941, 1951, Zhang and Yang 1995), but they are also present in both western and eastern North America (Attridge et al. 1985, Galton 1976, Yates 2004). Furthermore, Early Jurassic sauropods are known from China (e.g. He et al. 1998, Barrett 1999, Upchurch et al. 2007) and Europe (Wild 1978).
Middle Jurassic
As for the Southern Hemisphere, the Middle Jurassic archosaur record from the Northern Hemisphere is rather poor, and, most notably, only one skeletal remain of an archosaur from that period, the poorly preserved type specimen of the crocodyliform Entradasuchus, has been described from North America (Hunt and Lockley 1995), though dinosaurs are known from track evidence (e.g. Kvale et al. 2001). Middle Jurassic crocodiles from Europe are represented by isolated teeth attributed to atoposaurids and goniopholids in England (Evans and Milner 1994) and France (Kriwet et al. 1997) and marine thalattosuchians from various countries (Mueller-Töwe 2006). The record of these marine crocodyliforms is poor in the Aalenian and Bajocian, although the first metriorhynchids are known from the latter stage (Mueller-Töwe 2006). In the Bathonian and Callovian, both teleosaurid and metriorhynchid thallatosuchians are wellrepresented especially in England and France, although some remains are also known from central Asia (Mueller-Töwe 2006). Middle Jurassic terrestrial crocodiles from Asia include a goniopholid and the enigmatic Hsisosuchus from the Lower Shixaniao Formation (Peng et al. 2005), as well as a "sphenosuchian" from the Wuceiwan Formation (Clark et al. 2004) and a goniopholid from the Totunhe Formation in China (Maisch et al. 2003), and a report of a goniopholid from Kirghizia (Averianov 2000).
The most important Middle Jurassic dinosaur fauna from the Northern Hemisphere is that of the (?) Bathonian Lower Shaximiao Formation of China (Peng et al. 2005), which includes stegosaurian (Maidment and Wei 2006) and basal cerapodan ornithischians (Barrett et al. 2005), theropods, and sauropods. Theropods include basal tetanurans, such as Xuanhanosaurus and "Szechuanosaurus" zigongensis (Dong 1984, Gao 1993), and the possible basal coelurosaur Gasosaurus (Dong and Tang 1985, Holtz et al. 2004). The sauropod fauna is dominated by mamenchisaurids (Peng et al. 2005), but also includes the basal sauropod Shunosaurus and the possibly oldest known neosauropods (Upchurch et al. 2004). Another close relative of neosauropods or a basal member of this clade is also known from the Callovian Balabansai Formation of Kirghizia [Fig. 4b(7), Alifanov and Averianov 2003]. Ornithischians are represented in the same formation by an indeterminate stegosaur (Averianov et al. 2007) and isolated teeth that were described as the oldest known pachycephalosaur (Averianov et al. 2005). However, the teeth are rather poorly preserved and the identification is questionable.
Although some of the first dinosaurs to be described were Middle Jurassic taxa from Europe (e.g. Buckland 1824), Middle
Jurassic dinosaurs from this continent are still poorly understood, mainly due to the fragmentary nature of most remains. However, the fauna includes non-neosauropodan sauropods (Upchurch and Martin 2002, 2003), the basal spinosauroids Eustroptospondylus (Walker 1964, Holtz et al. 2004), Dubreuillosaurus (Allain 2002, 2005) and Poekilopleuron (Allain and Chure 2002), and one of the oldest coelurosaurs, Proceratosaurus (Woodward 1910, Holtz 2000, Rauhut 2003c). Ornithischian dinosaurs are represented by stegosaurs (Galton and Powell 1983, Galton 1990), ankylosaurs (Galton 1983) and basal ornithopods (Galton 1980b, Evans and Milner 1994, Kriwet et al. 1997, Ruiz-Omeñaca et al. 2005).
Late Jurassic
One of the most important Late Jurassic archosaur faunas is certainly that of the Morrison Formation of western North America (see Foster 2003). Crocodiles from this unit include some of the last "sphenosuchians" known (Walker 1970, Göhlich et al. 2005), a cursorial basal mesoeucrocoylian and other cursorial crocodylomorphs of unspecified systematic position, the probable atoposaurid Hoplosuchus, several species of Goniopholis, and the goniopholid Eutretauranosuchus (Tykoski et al. 2002, Foster 2003, Pol and Gasparini 2007: fig. 5.5). Pterosaurs are poorly represented in the terrestrial deposits of the Morrison Formation, but include at least one "rhamphorhynchoid" and several taxa of pterodactyloids (Foster 2003). The dinosaur fauna of the Morrison Formation is the most diverse fauna known from the Jurassic, and one of the most diverse Mesozoic dinosaur faunas known in general (see Weishampel et al. 2004). The dinosaur fauna includes several species of diplodocid sauropods, the basal macronarian Camarasaurus as the most abundant taxon among the sauropods, the basal titanosauriform Brachiosaurus altithorax, Haplocanthosaurus, which either represents a basal diplodocoid, or a basal macronarian, and the probably basal diplodocoid Amphicoelias (Wilson 2002, Foster 2003, Upchurch et al. 2004). The theropod fauna includes rare ceratosaurs (Marsh 1884, Galton 1982, Madsen and Welles 2000, Chure 2001) and basal spinosauroids (Britt 1991, Holtz et al. 2004), abundant allosaurids (Madsen 1976, Foster 2003), and a variety of coelurosaurs (Foster 2003, Holtz et al. 2004, Carpenter et al. 2005). Ornithischians are represented by rare heterodontosaurids (Galton 2005), several stegosaurian and ankylosaurian thyreophorans (Kirkland and Carpenter 1994, Carpenter et al. 1998, 2001, Galton and Upchurch 2004) and diverse ornithopods (Foster 2003, Norman et al. 2004, Norman 2004, Galton 2005).
Another northern fauna of particular interest in biogeographic terms is that from central America and the Caribbean (see e.g. Gasparini and Iturralde-Vinent 2006). Taxa reported from that area include metriorhynchid crocodiles (Gasparini and Iturralde-Vinent 2001, Buchy et al. 2006, 2007) and rhamphorhynchid pterosaurs (Colbert 1969, Gasparini et al. 2004).
In Europe, several units of Kimmeridgian age in Portugal have yielded a fauna that is closely comparable to that of the Morrison Formation. Furthermore, the Kimmeridgian-Tithonian lithographic limestones of southern Germany have yielded a rich fauna of mainly non-dinosaurian archosaurs, marine crocodiles are known from a number of Late Jurassic formations, and several isolated archosaur remains have been reported from various localities.
European Late Jurassic crocodiles include goniopholids (e.g. Schwarz 2002), atoposaurids (e.g. Wellnhofer 1971), teleosaurid and metriorhynchid thalattosuchians (see Mueller-Töwe 2006), and some mesoeucrocodylian taxa of uncertain affinities, such as the enigmatic Lisboasaurus and Lusitanosuchus (Buscalioni et al. 1996, Krebs and Schwarz 2000, Schwarz and Fechner 2004). Pterosaurs are especially represented by the rich faunas of the Solnhofen limestone (see especially Wellnhofer 1970, 1975), but are also known from fragmentary material from other localities (e.g. Wiechmann and Gloy 2000, Fastnacht 2005). Taxa reported in-clude the enigmatic "frogmouthed" anuroganthids (Wellnhofer 1975, Bennett 2007), scaphognathine and rhamphor-hynchine rhamphorhynchids (Wellnhofer 1975), ctenochasmatoids (Wellnhofer 1970, Unwin 2003, Jouve 2004), and dsungariptoroids (Wellnhofer 1970, Un-win 2003, Fastnacht 2005).
Late Jurassic dinosaurs from Europe include ceratosaurian, basal spinosauroid, allosauroid, and various coelurosaurian theropods, brachiosaurid and diplodocid sauropods, and stegosaurian and ornithopodan ornithischians from Portugal (Antunes and Mateus 2003, Mateus 2006, Mateus et al. 2006), basal coelurosaurs and the basal bird Archaeopteryx from the lithographic limestones of southern Germany (Ostrom 1978, Wellnhofer 1992, Mayr et al. 2005, Göhlich and Chiappe 2006), and some other finds of similar taxa from various localities (see Weishampel et al. 2004).
In China, Late Jurassic archosaurs are mainly known from the Upper Shaximiao Formation (though see comments on the age of this formation above) and the Shishougou Formation. Crocodiles reported include basal crocodyliforms (Peng et al. 2005) and goniopholids (see Wu et al. 1996), as well as another species of Hsisosuchus (Peng et al. 2005). Late Jurassic pterosaurs from Asia are rare, but include at least the ctenochasmatid Huanhepterus (Dong 1982).
Dinosaurs from the Late Jurassic of Asia include sinraptorid allosauroids, mamenchisaurid sauropods, and stegosaurian and ornithopodan ornithischians (Weishampel et al. 2004, Peng et al. 2005, Maidment and Wei 2006). Furthermore, the oldest ceratopsian ornithischians are known from the Tithonian Tuchengzi and the Oxfordian Shishougou formations of China (Zhao et al. 1999, Xu et al. 2006b), and the latter unit has also yielded the basal tyrannosauroid Guanlong (Xu et al. 2006a) and the first ceratosaurs from Asia (Xu and Clark 2006).
JURASSIC ARCHOSAUR EVOLUTION: A SOUTHERN PERSPECTIVE
Quality of the Jurassic archosaur fossil record
From the above, it is clear that the Jurassic archosaur fossil record is far from complete, both stratigraphically and geographically (Figs. 4, 10), and our understanding of Jurassic archosaur evolution is actually based on surprisingly few localities (Fig. 4). Whereas several important faunas are known in the Early Jurassic (Upper Elliot and Clarens Formations in Africa, Navajo Sandstone, Kayenta Formation and equivalent beds in North America, Posidonia Shale and equivalent beds in Europe, Lufeng Formation in Asia) and the Late Jurassic (Tendaguru Beds in Africa, Morrison Formation in North America, various units in Portugal and Solnhofen limestones in Europe, Shishougou Formation in Asia), Middle Jurassic archosaurs are almost entirely known from isolated finds, often also fragmentary. This is especially true for the early Middle Jurassic (Aalenian-Bajocian), whereas at least one important fauna is known from the Bathonian-Callovian, that of the Lower Shaximiao Formation of China (Peng et al. 2005). This underscores the potential importance of the Late Middle Jurassic Cañadón Asfalto Formation of Chubut, Argentina. Although mainly dinosaurs and mammals have been described from this formation so far (Bonaparte 1979, 1986, Rauhut et al. 2002, Rauhut 2003a, 2005a, Rougier et al. 2007a, b), abundant remains of other vertebrates have also been found, but not described in detail yet, including well-preserved pterosaurs (Unwin et al. 2004) and remains of crocodiles (Rauhut et al. 2001), potentially making this unit a southern counterpart of the Lower Shaximiao Formation.

Figure 10: Stratigraphic completeness of the archosaur fossil record of Gondwana. a) Crocodiles. b) Pterosaurs. c) Dinosaurs. Dotted lines indicate uncertain limits of the stratigraphic range of a certain geological unit. Some minor localities with fragmentary and uninformative material have been omitted, in the chart for dinosaurs, only localities that have yielded at least two identifiable taxa are shown. India includes Madagascar, since these two areas were geographically united in the Jurassic.
However, on stage level, biases in stratigraphic distribution are also found in other epochs of the Jurassic. Thus, Oxfordian archosaurs are still very poorly known, with the Shishougou Formation of China basically representing the only productive formation of that age. Likewise, several stages of the Early Jurassic might be underrepresented, though interpretation is hampered here by the uncertain dating of almost all known terrestrial vertebrate localities of this epoch. Concerning the geographic distribution of Jurassic archosaurs, their fossil record is strongly biased towards faunas from the Northern Hemisphere, which becomes evident when looking at the dinosaur record from the Jurassic (Fig. 11). For the entire Jurassic, 121 valid species of dinosaurs are recognized from Laurasian continents, whereas only 27 have been described from Gondwana (as of October 2007, only formally named taxa have been considered, OR & ALA, unpublished data). For the different epochs of the Jurassic, the numbers are more even for the Early Jurassic (19 species in Laurasia, 10 species in Gondwana), but very strongly biased towards northern faunas in the Middle (34 to 8) and especially the Late Jurassic (68 to 9). Thus, most scenarios of Jurassic archosaur evolution are largely or even entirely based on the fossil record of the Northern Hemisphere. This makes interpretation of evolutionary events often very difficult, and especially the hypothesis of increased endemism in terrestrial archosaurs in response to the initial breakup of Pangea is hard to test in the absence of suitable localities in the Southern Hemisphere. For example, until recently it was difficult to evaluate the observed differences between the Laurasian Late Jurassic dinosaur fauna and that from the Tendaguru Beds in Tanzania, since, in the absence of other Late Jurassic dinosaur faunas from Gondwana, it was impossible to decide whether these differences reflect differential evolution of the faunas, or maybe just different ecological settings of the localities. Indeed, given this paucity in comparative faunas, more emphasis has been laid on the supposed similarities of the Tendaguru fauna and those of the Morrison Formation and Late Jurassic Portuguese localities, and a variety of biogeographical hypotheses have been proposed to explain these similarities (e.g. Lull 1915, Galton 1977, 1982, Mateus 2006). Fortunately, the newly recognized fauna from the Cañadón Calcáreo Formation of Argentina (Rich et al. 1999, Rauhut et al. 2005, Rauhut 2006a, c) might help to put the Tendaguru fauna in a Gondwanan context, once more material is excavated and studied (see also below).
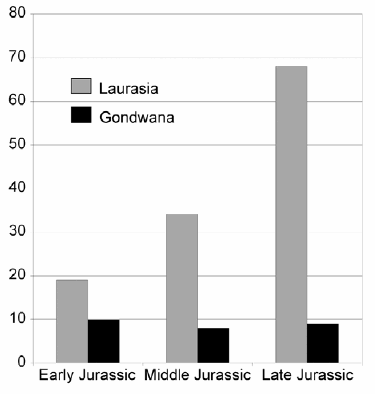
Figure 11: Valid dinosaur species from the Jurassic of the Northern and Southern Hemisphere (as of October 2007, OR & ALA unpublished data).
Taking both the stratigraphic and geographic bias in Jurassic archosaur distribution into consideration, especially the Gondwanan archosaur record of this time can only be called patchy (Figs. 4, 10). In terms of marine crocodiles, South America certainly has the most complete record of all Gondwanan continents, with thalattosuchians known from most stages from the Sinemurian to the Tithonian. Concerning terrestrial Jurassic archosaur faunas, Africa has the most complete record, with remains known from almost the entire Early Jurassic, the latter stages of the Middle Jurassic, and the upper stages of the Late Jurassic, though a lot of especially the Middle Jurassic finds are isolated occurrences. In South America, only the later Middle Jurassic is well represented by the fauna from the Cañadón Asfalto Formation, though with great potential for new discoveries in both the Early and the Late Jurassic in Argentina. All other Gondwanan continents have only isolated occurrences, in the Early Jurassic in India and Antarctica, and in the Middle Jurassic in Australia. However, it must be noted that even for the Jurassic stages with faunas known in the Gondwanan continents, these are usually single localities or formations, and comparable material from other parts of the continent is usually absent.
Faunal composition, evolutionary considerations and biogeography
a) Marine crocodiles: The evolutionary and biogeographic significance of the thalattosuchian crocodiles, especially of South America, has been discussed at length by Zulma Gasparini and co-workers (e.g. Gasparini 1985, 1992, 1996, Gasparini and Fernandez 1997, Gasparini et al. 2000, 2005, 2006), so only the most important aspects will be outlined here.
The occurrence of thalattosuchian crocodiles in the Sinemurian of Chile (Chong Diaz and Gasparini 1972) and western Argentina (Huene 1927), and, probably, the Early Jurassic of India (Bandyopadhyay et al. 2002) demonstrates that this clade did not only have a circumtethyan distribution in the Early Jurassic, but also already entered the Pacific region. This dispersal most probably happened through the Hispanic corridor (Gasparini et al. 2000), which was sporadically open since at least the Pliensbachian (Aberhan 2001), though the contemporaneous occurrence of the first thalattosuchians in the Sinemurian of Europe and Chile might indicate that marine faunal exchanges may have been possible even before.
With the repeated opening and closing of the Hispanic corridor during the Middle Jurassic (Westermann 1993, Moyne et al. 2004), there might have been an exchange of marine crocodiles between the Pacific and the Tethys every time the corridor was in effect. This exchange was probably not unidirectional; whereas the oldest representative of Metriorhynchus is found in Chile (Gasparini et al. 2000), and the genus only later appears in Europe (Mueller-Töwe 2006), Geosaurus is first known in Europe, and only subsequently in South America (Pol and Gasparini 2007). The evolution of thalattosuchians in the Middle Jurassic might thus be characterized by periods of dispersal separated by periods of cladogeneses in separation in the Tethys and the Pacific. Thus, the Hispanic corridor might have been a "motor" of thalattosuchian evolution in the Middle Jurassic, though this hypothesis needs further testing by new finds and rigorous cladistic and biogeographic analyses of all species of Middle and Late Jurassic thalattosuchians.
The exchange between the Tethyan region and the Pacific continued during the Late Jurassic, when the Hispanic corridor was permanently open (Ford and Golonka 2003), as evidenced by shared genera of metriorhynchids, such as Geosaurus and Dakosaurus. It is interesting, though, that, whereas all three genera of metriorhynchids (Metriorhynchus, Geosaurus and Dakosaurus) are present in both areas, no teleosaurids have been reported from the Late Jurassic of western South America so far, although they are present both in the northern and southern part of the Tethys during this period (e.g. Krebs 1967, Bardet and Hua 1996). This might be due to the more fully marine environments represented by the thalattosuchian-bearing sediments in Chile and Argentina (Gasparini et al. 1999), and further finds of marine reptiles from nearshore environments will have to show whether this group was truly absent from the Pacific coast of South America.
b) Terrestrial crocodiles: Interpretation of the evolution of terrestrial crocodiles in Gondwana during the Jurassic is hampered by the abysmal fossil record of these animals in the Middle and Late Jurassic of the Southern Hemisphere. In the Early Jurassic, the first successful radiations of the crocodylomorph lineage, the "sphenosuchians" and basal crocodyliforms ("protosuchians"), had a Pangean distribution, as evidenced by the occurrence of these groups in southern Africa, North America and China during this time. Phylogenetic relationships among crocodylomorphs, especially the presence of the advanced mesoeucrocodylian lineages Thalattosuchia and Goniopholidae in the Early Jurassic, indicate that the major radiation of mesoeucrocodylians already happened during this period (Pol and Gasparini 2007), but most lineages are poorly represented globally before the Late Jurassic. The presence of possible atoposaurids in the Middle Jurassic in Madagascar and a possible goniopholid in the Late Jurassic of Tanzania, if the identity of these fragmentary remains can be confirmed, might show that these more advanced mesoeucrocodylians were also globally distributed before the final separation of the continents of the Northern and Southern Hemisphere. Furthermore, the atoposaurid record from Madagascar (Flynn et al. 2006) is roughly contemporaneous with the oldest records of this group in the Northern Hemisphere (e.g. Kriwet et al. 1997), indicating that derived members of this lineage might originated already considerably earlier.
However, the lack of two important mesoeucrocodylian lineages, which are especially widely distributed in the Southern Hemisphere during the Cretaceous, the notosuchians and the peirosaurids (Fig. 1, see Pol and Gasparini 2007), during the Jurassic is striking. Two possible scenarios might explain the lack of these clades in the fossil record during the Jurassic. First, both groups might have been genuinely rare during the Jurassic and were maybe restricted to a small region, and only experienced their great radiation in the Cretaceous. Alternatively, both groups might have been largely restricted to the Southern Hemisphere already in the Jurassic, but not necessarily been rare, and have not been reported so far due to the extremely poor record of terrestrial vertebrates in the Jurassic of Gondwana. Which of these scenarios, which might not be completely exclusive, is correct can only be demonstrated by more material of terrestrial crocodiles from the Southern Hemisphere.
c) Pterosaurs: As in the case of the terrestrial crocodiles, the extremely poor Jurassic Gondwanan record of pterosaurs severely restricts any interpretation of their evolutionary history in the Southern Hemisphere. From the scarce remains, it is evident that pterosaurs were already present in Gondwana in the Early Jurassic. However, pterosaur remains are unknown from the best known Early Jurassic vertebrate fauna from the Southern Hemisphere, that of the Upper Stormberg Group of South Africa, despite the fact that the Upper Elliot Formation frequently preserves small vertebrate remains in often exquisite detail. This might indicate that these animals might have been very rare in southern Gondwana at that time. That pterosaurs had colonized even southern Gondwana by the Early Jurassic is shown by the isolated humerus found in the Hanson Formation of Antarctica (Smith and Pol 2007).
If confirmed, the occurrence of a pterodactyloid in the Callovian Cañadón Asfalto Formation of Argentina would be the earliest record of this clade globally, which, in the Northern Hemisphere appears abruptly in the fossil record with many different clades in the Kimmeridgian- Tithonian (Unwin 2006). Once properly studied, the pterosaur fauna from the Cañadón Asfalto Formation might prove to be of great importance for our understanding of pterosaur evolution, since it represents one of the very few strictly terrestrial faunas that have yielded abundant and well-preserved pterosaur material (see Buffetaut 1995).
From the Late Jurassic, only pterodactyloid pterosaurs have been reported from South America so far (Martill et al. 2000, Codorniú and Gasparini 2007), but both "rhamphorhynchoids" and pterodactyloids are present in eastern Africa (Reck 1931, Galton 1980, Unwin and Heinrich 1999). Again, the wide distribution of pterydactyloids indicates that their evolution and radiation was well under way by the time the first members of this clade appear in the fossil record. Furthermore, the occurrence of dsungaripteroids in Europe (see Unwin 2003, Fastnacht 2005), Africa (Unwin and Heinrich 1999) and, possibly, South America (Martill et al. 2000) indicates a still global distribution of these animals in the Late Jurassic, as might be expected from clade of actively flying animals, and is also still found in the Cretaceous for several pterosaur clades, despite the separation of continental masses (Unwin 2006).
Again, more fossil finds from Gondwanan localities are needed to better understand pterosaur evolution during the Jurassic.
d) Dinosaurs: Compared to the other groups discussed above, dinosaurs certainly have the best Jurassic fossil record in the Southern Hemisphere, though their record is certainly far from complete, and even very poor if compared with that of the Northern Hemisphere (see above). Thus, all interpretations of Jurassic dinosaur evolution in Gondwana should be regarded as tentative, and might easily change in the light of new discoveries.
Early Jurassic dinosaurs faunas, especially that of the Elliot and Clarens formations and equivalent beds in Zimbabwe in southern Africa highlight the close faunal relationships in different parts of Pangea at that time. Basal thyreophoran ornithischians are present in basically all important Early Jurassic dinosaur localities, in southern Africa (Butler et al. 2008), the Kota Formation of India (Nath et al. 2002, see above), the Kayenta Formation of western North America (Colbert 1981, Padian 1989), the Liassic of Europe (Owen 1863, Haubold 1990), and the Lufeng Formation of China (Norman et al. 2007). This global distribution gives testimony to the rapid evolution of thy-reophorans, which seem to represent one of the first successful radiations of ornithischians, soon after the origin of this group in the Late Triassic.
Another group of ornithischians from southern Africa is of special interest as well, the heterodontosaurids. Heterodontosaurids represent the most diverse and abundant group of ornithischians in the Upper Elliot and Clarens Formations (Butler 2005, Knoll 2005), but they seem to be rare in the Northern Hemisphere, from where only one undescribed heterodontosaurid from the Kayenta Formation has been reported (Sereno pers. com. in Butler 2005). The fact that heterodontosaurids probably represent the most basal clade and thus the earliest radiation of ornithischian dinosaurs (Butler et al. 2008), together with the occurrence of the basalmost ornithischians known in the Late Triassic of South America (Casamiquela 1967, Ferigolo and Langer 2007), including the oldest known heterodontosaurid (Báez and Marsicano 2001), and southern Africa (Butler et al. 2007), indicates that the origin and early diversification of ornithischians is probably found in southern Gondwana. The phylogenetic position of Lesothosaurus as the basalmost thyreophoran known and of Stormbergia as the basalmost cerapodan (Butler et al. 2008) is in general accordance with this interpretation (Fig. 12).

Figure 12: Phylogeny of early ornitischians (based on Butler et al. 2007, 2008), showing the geographic distribution of early ornitischians.
Thus, the poor Gondwanan fossil record in the Middle Jurassic might account for our still rather poor understanding of the early Mesozoic radiation of ornithischians, since the major radiation of this group, in which basically all important lineages were established, took place during this time (Sereno 1999a, Butler et al. 2008). It is interesting, however, to note that ornithischians seem to have been rather rare in the Middle Jurassic of Gondwana, where they have only been reported in the best known fauna, that of the Cañadón Asfalto Formation (see above), and here they seem to be amongst the rarest vertebrate groups represented in general (OR, pers. obs.). Nevertheless, the wide distribution and diversity of ornithischians on higher taxonomic level in the Late Jurassic indicates that much of this radiation took place during Pangean times in the Early or Middle Jurassic, and so it should not be too surprising to find similar groups of ornithischians in the faunas of the Northern and Southern Hemisphere even after the final separation of the continents of Laurasia and Gondwana at the Middle-Late Jurassic boundary.
Early Jurassic saurischians from Gondwana are also indicative of a global Pangean dinosaur fauna at that time (see also Smith et al. 2007b). In theropods, coelophysoids are found on all major landmasses from which theropods have been reported (Raath 1969, 1977, Rowe 1989, Carrano and Sampson 2004, Irmis 2004, Tykoski 2005), with the exception of Antarctica. Furthermore, the recently recognized dilophosaur clade (Smith et al. 2007a, b) also shows a global distribution, with representatives known from North America, China, South Africa, and Antarctica. Likewise, close relatives of the southern African sauropodomorph Massospondylus are known from the Early Jurassic of North (Attridge et al. 1985) and South America (Martínez 1999), and Smith and Pol (2007) found close relationships of this genus with the Chinese Lufengosaurus and the Antarctic Glacialisaurus, so that the Massospondylidae (sensu Smith and Pol 2007) represent another successful Pangean radiation of dinosaurs in the early Mesozoic. Thus, especially if the radiation of dinosaurs really followed a Late Triassic extinction event, as argued by Olsen et al. (2002), this Pangean distribution of many groups already in the Early Jurassic indicates high evolutionary dynamics and great dispersal potential of these animals. It might be noted, however, that high dinosaur diversity already in the Triassic in other parts of the world, including South America, and inferred ghost lineages argue against such a sudden radiation after a major extinction event at the T-J boundary.
Interesting is the report of a probable abelisauroid from the Early Jurassic of Gondwana (Allain et al. 2007). This lineage is almost entirely restricted to the Southern Hemisphere in the Late Jurassic and Cretaceous (see Tykoski and Rowe 2004, Rauhut 2005b), so its occurrence already in the Early Jurassic would represent a surprising case of endemism of this group, especially considering the northern Gondwanan (and thus more or less central Pangean) position of this find and the global distribution of most other groups.
Of special interest are furthermore the sauropods, which originated in the Triassic (see Yates and Kitching 2003), but seem to have experienced their first important radiation across the Triassic-Jurassic boundary. Gillette (2003: 686) suggested that Early Jurassic sauropods had a distribution in eastern Gondwana and southern and eastern Laurasia, and were restricted to middle latitudes, and that sauropods reached North America and western Gondwana only in the Middle or even Late Jurassic. Most of this argument was based on negative evidence, i.e. the lack of sauropod fossils from these regions during the Early Jurassic. Although some non-eusauropodan taxa were included in Gillette's analysis (e.g. Vulcanodon), most of this scenario invoves eusauropods, and, given the current confusion about the definition of Sauropoda, the following discussion will mainly consider Eusauropoda. New discoveries and paleogeographic considerations contradict the idea of restricted Early Jurassic faunas and an only Middle to Late Jurassic global distribution of sauropods. First, the discovery of probable sauropod remains in Antarctica (Smith and Pol 2007) might show that sauropods had reached low southern latitudes by that time, although a detailed study of this material is needed to establish its sauropod affinities and answer the question whether this material represents a basal taxon, or a eusauropod. Furthermore, the occurrence of Amygdalodon at around the Early-Middle Jurassic boundary in Argentina (Rauhut 2003b) and other fragmentary eusauropod remains from the Aalenian of this country (Salgado and Gasparini 2004) indicate that eusauropods were also present in western Gondwana early in the Jurassic. Considering palaeogeographic reconstructions of the Early Jurassic (Scotese 1991, Smith et al. 1994), it seems furthermore unlikely that a dispersal of eusauropods from eastern Gondwana to eastern Laurasia (or vice versa) could have happened withouth these animals entering North America as well. Indeed, the presence of Anchisaurus, which, according to Yates (2004), represents an early, non-eusauropodan sauropod (see also Fedak and Galton 2007), in the Early Jurassic of North America indicates that immediate eusauropodan outgroups already had a global distribution during this time. This is also in agreement with the find of the probable primitve eusauropod Ohmdenosaurus in central Europe (Wild 1978), a region that was geographically close to the eastern coast of North America. The absence of eusauropods from western North America might thus have pelaeoecological reasons, or might simply be due to incomplete sampling of these faunas. Thus, it seems most probable that eusauropods also reached a Pangean distribution already in the Early Jurassic.
In the Middle Jurassic, the change in theropod faunas from coelophysoid/basal theropod dominated Early Jurassic faunas to basal tetanuran faunas seems to have been paralleled in the Southern Hemisphere (Rauhut 2005a), though the timing and mode of this change remains uncertain due to the very poor Middle Jurassic theropod record of Gondwana, which, in terms of identifiable taxa, is restricted to the Cañadón Asfalto Formation. Basal, non-neosauropodan eusauropods were widely distributed in Gondwana during the Middle Jurassic, and seem to represent several lineages of an early radiation of eusauropods, some of which might go back to the early Jurassic. This notion is supported by the association of archaic forms in the later Middle Jurassic, such as Archaeodontosaurus (Buffetaut 2005) or Volkheimeria (Bonaparte 1986b, 1999), with much more progressive forms of sauropods, some of them already close to or within Neosauropoda, such as Atlasaurus (Monbaron et al. 1999, Upchurch et al. 2004). Lapparentosaurus is a taxon of problematic position, since it shows a combination of plesiomorphic and apomorphic characters (Upchurch et al. 2004). Upchurch (1998) found this taxon to represent a brachiosaurid, which would be the oldest record of a derived neosauropod, and argued that the primitive character of the vertebrae might be due to the juvenile state of the type material. However, the latter notion seems to be unlikely, since similar sized, juvenile specimens referred to Patagosaurus (MACN Ch 932, 934), for example, already show more complicated vertebral structures (OR pers. obs.). Thus, the phylogenetic position of this interesting taxon has to remain unresolved, pending a revision of the known, or the discovery of new material. Being intercalated within Laurasian taxa, or having close relatives in Laurasia, these Middle Jurassic sauropods furthermore indicate a generally Pangean sauropod fauna in the late Middle Jurassic.
Of special interest are, of course, the dinosaur faunas of the Late Jurassic of the Southern Hemisphere, since it is in these faunas that we would expect to find first consequences of the separation of Gondwana from the continents of the Northern Hemisphere. However, analysis of these effects has been hampered by the fact that, until very recently, only one dinosaur fauna had been reported from this period in Gondwana, that of the Tendaguru Beds in Tanzania, so that material for comparisons was missing (see above). This had another effect in the description of the fauna from Tendaguru: due to the lack of comparable material from the Southern Hemisphere, the taxa from this locality have generally been compared and often even been referred to taxa known from the Northern Hemisphere, which led to the emphasis on faunal similarities between the fauna of Tendaguru and especially that of the Morrison Formation. However, although this similarity certainly exists on higher taxonomic level, this might not be too significant, since most lineages represented probably reach back to the Middle Jurassic and thus to the time before the separation of Gondwana (see Remes and Rauhut 2005, Remes 2006, 2007). Recent work has furthermore shown that taxonomic assignment to Laurasian forms on lower taxonomic level is often questionable (Remes 2006), and the same is true for those forms still kept in the same genera as comparable taxa from the Morrison Formation. In the case of Brachiosaurus brancai, for example, a rigorous phylogenetic analysis of the taxa referred to the family Brachiosauridae would be necessary to test whether this species really is more closely related to Brachiosaurus altithorax from the Morrison Formation, or whether the latter might rather group with other Laurasian brachiosaurids, such as Sauroposeidon. The same is true for Dysalotosaurus lettowvorbecki (currently kept in the North American genus Dryosaurus, see Galton 1977, Norman 2004). In this case, the problem is even more acute, since Dysalotosaurus and Dryosaurus represent the only well-known dryosaurids, wheras all other taxa within the Dryosauridae are based on isolated elements or very fragmentary remains. Thus, no analysis of the interrelationships of dryosaurids has ever been attempted. In the case of Elaphrosaurus, the North American specimens referred to this taxon (Galton 1982, Chure 2001) might represent ceratosaurs, but cannot be referred to this genus (Rauhut and Carrano in prep). These examples again highlight the need for rigorous phylogenetic analyses as the basis for biogeographic interpretations, as already emphasized by Forster (1999). In the absence of such phylogenetic analyses for the taxa concerned, a faunistic approach might shed some light on the biogeographic affinities of the Tendaguru fauna. Although similar in terms of taxonomic composition at higher taxonomic levels, the relative abundance of taxa and overall composition of the fauna from this locality is strikingly different from that of the Morrison Formation. In the latter, the most common sauropod isCamarasaurus, with its several species, and diplodocids, whereas titanosauriforms (only known from brachiosaurids in this formation) are very rare (Dodson et al. 1980, Foster 2003). In Tendaguru, in contrast, brachiosaurids and other basal titanosauriforms are among the most common sauropods (Russell et al. 1980), which would be in good accordance with the titanosauriform dominated Cretaceous faunas in Gondwana. Although this difference might be assigned to palaeoecological reasons, the same seems to be true for the Kadsi Formation of Zimbabwe (Raath and McIntosh 1987) and, maybe, the Cañadón Calcáreo Formation of Argentina (with Tehuelchesaurus representing a basal titanosauriform or an immediate titanosauriform outgroup, Rauhutet al. 2005, Carballido et al. 2007), though more sampling is needed to confirm this. In the latter case, the paleoecological setting seems to be different from that of Tendaguru, and this locality was placed considerably further south than the latter in Late Jurassic times.
In addition to this relative abundance of specimens, differences in the faunal composition are also reflected in the taxonomic composition of the sauropod faunas (Fig. 13). For this comparison, only the sauropod fauna of the Morrison Formation (Foster 2003, Upchurch et al. 2004) and the Late Jurassic of Portugal (Antunes and Mateus 2003) have been considered for the Northern Hemisphere, since eastern Asia was separated from western Laurasia and Gondwana by epicontinental seas from at least the Bathonian (Smith et al. 1994), and, thus, the Chinese Late Jurassic sauropod fauna shows a rather high endemism and can thus not be regarded as a "typical Laurasian" fauna (Upchurch et al. 2002). For the Southern Hemisphere, the Tendaguru fauna and the two named taxa from the Cañadón Calcáreo Formation have been considered together as representatives of the Late Jurassic Gondwanan fauna. Taxa have been counted on species level.
Looking thus at sauropod faunal composition, diplodocids constitute more than half of the known sauropod taxa from the Northern Hemisphere (11 of 21 species), whereas basal titanosauriforms (2 species) account for only ten percent of the species. Camarasurids are the second most important faunal element, with 14% (3 species). In contrast, basal titanosauriforms (or immediate titanosauriform outgroups) account for almost half of the species in the Southern Hemisphere (4 of 9 species), followed by dicraeosaurids (3 of 9 species), whereas only two species of diplodocids have been described from Gondwana. The most remarkable aspect of this faunal comparison, however, is the lack of one of the most common groups of sauropods from the Morrison Formation, the camarasaurids, in Gondwana, and, in turn, the lack of an important Gondwanan lineage, the dicraeosaurids, in Laurasia. Although the former might still be accounted for by the poor Gondwanan fossil record and the very different sample sizes, the latter is especially noteworthy, since dicraeosaurids have been reported from all three Gondwanan localities that have yielded identifiable Late Jurassic sauropods (Raath and McIntosh 1987, Aberhan et al. 2002, Rauhut et al. 2005), even though they are geographically, latitudinally (and thus probably climatically) and, as far as can be said at the current state of knowledge, ecologically disparate. Since the phylogenetic relationships of both camarasaurids and dicraeosaurids indicate that these groups must have originated in Pangean times as well, their absence in the Late Jurassic in one of the Hemispheres must either be due to the failure to disperse to the other Hemisphere in Middle Jurassic times, or to differential extinction in Laurasia and Gondwana (see also Harris and Dodson 2004). Although this question can only be answered by new discoveries, the lack of these important faunal components in the respective other Hemisphere might thus represent a first sign of differential evolution of the dinosaur faunas of the two Hemispheres due to limitations in dispersal potential, following the separation of Gondwana.
CONCLUSIONS
Jurassic archosaurs from the Southern Hemisphere are still rather poorly known. Although this is especially true for terrestrial crocodiles and pterosaurs, the dinosaur record from Gondwana is also still far from complete. The most important archosaur faunas from the Jurassic of the Southern Hemisphere are that of the Hettangian-Sinemurian Elliot and Sinemurian-Pliensbachian Clarens formations of southern Africa, the (?) Hettangian-Toarcian Kota Formation of India, the Early Jurassic Hanson Formation of Antarctica, the Callovian Cañadón Asfalto Formation of Argentina, and the Kimmeridgian-Tithonian Saurian Beds of Tendaguru, Tanzania. Marine crocodiles are especially well-represented in the western part of southern South America, from where different taxa are known from units ranging from the Sinemurian to the Tithonian. Multiple events of faunal exchange through the sporadically open Hispanic corridor during the Early and Middle Jurassic might have been an important factor in thalattosuchian evolution.
Terrestrial archosaur faunas show a Pangean distribution in the Early Jurassic, and, most probably in the Middle Jurassic (for which almost no terrestrial crocodiles have been reported from the Southern Hemisphere). The rather high diversity of basal ornithischian dinosaurs in the Early Jurassic of southern Africa might indicate that the early radiation of this clade took place in southern Gondwana. In the Late Jurassic, especially sauropod faunas might already show the first indications of differential faunal evolution between the Southern and Northern Hemisphere.
However, given the patchy archosaur record from the Jurassic of Gondwana, all current interpretations of archosaur evolution and faunal differentiation during this epoch must be seen as tentative. More material is needed, especially from the Southern Hemisphere, to better understand archosaur evolution, in particular the effects that the initial continental breakup during the Jurassic might have had on the evolutionary history of this group.
ACKNOWLEDGEMENTS
We thank Ana Maria Zavattieri, Laura Giambiaggi, and Victor Ramos for the invitation to participate in the Jurassic Symposium in Mendoza and to contribute to this special volume. Too many colleagues to mention everybody have contributed to this work, with access to specimens, discussions, and critical comments on ideas expressed in meetings or personal discussions over the years, but we are thankful to each and every one of them for this input. Special thanks are due to the Museo Paleontológico Egidio Feruglio, Trelew, for the support during fieldwork in the Middle and Late Jurassic of Chubut Province, Argentina, and to Wolf Volkheimer for many insights into the Jurassic history of South America. The manuscript greatly benefited from critical comments by Diego Pol and Max Langer.
Parts of the research that led to this review paper were financially supported by a DAAD fellowship, grants from Fundación Antorchas, the Jurassic Foundation, BBC Horizon, and German Research Foundation (DFG) grants RA 1012/1-1, 2-1, 8-1, and 9-1 to OR, and DFG grant LO 1401/1-2 to ALA.
WORKS CITED IN THE TEXT
1. Aberhan, M. 2001. Bivalve palaeobiogeograohy and the Hispanic Corridor: time of opening and effectiveness of a proto-Atlantic seaway. Palaeogeography, Palaeoclimatology, Palaeoecology 165: 375-394. [ Links ]
2. Aberhan, M., Bussert, R., Heinrich, W.-D., Schrank, E., Schultka, S., Sames, B., Kriwet, J. and Kapilima, S. 2002. Palaeoecology and depositional environments of the Tendaguru Beds (Late Jurassic to Early Cretaceous, Tanzania). Mitteilungen aus dem Museum für Naturkunde Berlin, Geowissenschaftliche Reihe 5: 19-44. [ Links ]
3. Alifanov, V.R. and Averianov, A.O. 2003. Ferganasaurus verzilini gen. et sp. nov., a new neosauropod (Dinosauria, Saurischia, Sauropoda) from the Middle Jurassic of Fergana Valley, Kirghizia. Journal of Vertebrate Paleontology 23(2): 358-372. [ Links ]
4. Allain, R. 2002. Discovery of megalosaur (Dinosauria, Theropoda) in the Middle Bathonian of Normandy (France) and its implications for the phylogeny of basal Tetanurae. Journal of Vertebrate Paleontology 22(3): 548-563. [ Links ]
5. Allain, R. 2005. The postcranial anatomy of the megalosaur Dubreuillosaurus valesdunensis (Dinosauria, Theropoda) from the Middle Jurassic of Normandy, France. Journal of Vertebrate Paleontology 25(4): 850-858. [ Links ]
6. Allain, R. and Chure, D.J. 2002. Poekilopleuron bucklandii, the theropod dinosaur from the Middle Jurassic (Bathonian) of Normandy. Palaeontology 45(6): 1107-1121. [ Links ]
7. Allain, R., Aquesbi, N., Dejax, J., Meyer, C., Monbaron, M., Montenat, C., Richir, P., Rochdy, M., Russell, D. and Taquet, P. 2004. A basal sauropod dinosaur from the Early Jurassic of Morocco. Comptes Rendus Palevol 3: 199-208. [ Links ]
8. Allain, R., Tykoski, R.S., Aquesbi, N., Jalil, N.-E., Monbaron, M., Russell, D.A. and Taquet, P. 2007. An abelisauroid (Dinosauria: Theropoda) from the Early Jurassic of the High Atlas Mountains, Morocco, and the radiation of ceratosaurs. Journal of Vertebrate Paleontology 27(3): 610-624. [ Links ]
9. Antunes, M.T. and Mateus, O. 2003. Dinosaurs of Portugal. Comptes Rendus Palevol 2: 77-95. [ Links ]
10. Attridge, J., Crompton, A.W. and Farish A. Jenkins, J. 1985. The southern African Liassic prosauropod Massospondylus discovered in North America. Journal of Vertebrate Paleontology 5(2): 128-132. [ Links ]
11. Averianov, A.O. 2000. Sunosuchus sp. (Crocodylomorpha, Goniopholididae) from the Middle Jurassic of Kirghisia. Journal of Vertebrate Paleontology 20(4): 776-779. [ Links ]
12. Averianov, A.O., Bakirov, A.A. and Martin, T. 2007. First definitive stegosaur from the Middle Jurassic of Kyrgyzstan. Paläontologische Zeitschrift 81(4): 440-446. [ Links ]
13. Averianov, A.O., Martin, T. and Bakirov, A.A. 2005. Pterosaur and dinosaur remains from the Middle Jurassic Balabansai Svita in the Northern Fergana Depression, Kyrgyzstan (Central Asia). Palaeontology 48(1): 135-155. [ Links ]
14. Bandyopadhyay, S. and Sengupta, D.P. 2006. Vertebrate faunal turnover during the Triassic-Jurassic transition: an Indian scenario. New Mexico Museum of Natural History and Science Bulletin 37: 77-85. [ Links ]
15. Bandyopadhyay, S., RoyChowdhury, T.K. and Sengupta, D.P. 2002. Taphonomy of some Gondwana vertebrate assemblages of India. Sedimentary Geology 147: 219-245. [ Links ]
16. Bardet, N. and Hua, S. 1996. Simolestes nowackianus Huene, 1938 from the Upper Jurassic of Ethiopia is a teleosaurid crocodile, not a pliosaur. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 1996 (2): 65-71. [ Links ]
17. Barrett, P.M. 1999. A sauropod dinosaur from the Lower Lufeng Formation (Lower Jurassic) of Yunnan Province, People's Republic of China. Journal of Vertebrate Paleontology 19(4): 785-787. [ Links ]
18. Barrett, P.M. 2004. Sauropodomorph dinosaur diversity in the upper Elliot Formation (Massospondylus range zone: Lower Jurassic) of South Africa. South African Journal of Science 100: 501-503. [ Links ]
19. Barrett, P.M. and Yates, A.M. 2006. New information on the palate and lower jaw of Massospondylus (Dinosauria: Sauropodomorpha). Palaeontologia Africana 41: 123-130. [ Links ]
20. Barrett, P.M., Butler, R.J., Novas, F.E., Moore-Fay, S.C., Moody, J.M., Clark, J.M. and Sánchez-Villagra, M.R. 2008. Dinosaur remains from the La Quinta Formation (Lower or Middle Jurassic) of the Venezuelan Andes. Paläontologische Zeitschrift 82(2): 163-177. [ Links ]
21. Barrett, P.M., Upchurch, P. and Knoll, F. 2005. Small-bodied ornithischian dinosaurs from the Middle Jurassic of Sichuan, China. Journal of Vertebrate Paleontology 25(4): 823-834. [ Links ]
22. Bennett, S.C. 2007. A second specimen of the pterosaur Anurognathus ammoni. Paläontologische Zeitschrift 81(4): 376-398. [ Links ]
23. Benton, M.J. and Clark, J.M. 1988. Archosaur phylogeny and the relationships of the Crocodylia. In Benton, M.J. (ed.) The phylogeny and classification of the tetrapods, Volume 1: Amphibians, reptiles, birds. Clarendon Press, 295-338, Oxford. [ Links ]
24. Bonaparte, J.F. 1979. Dinosaurs: a Jurassic assemblage from Patagonia. Science 205: 1377-1379. [ Links ]
25. Bonaparte, J.F. 1981. Inventario de los vertebrados Jurasicos de America del Sur. En Volkheimer, W. and Musacchio, E.A. (eds.) Cuencas sedimentarias del Jurásico y Cretácico de América del Sur, Comité Sudamericano del Jurásico y Cretácico 2: 661-684, Buenos Aires. [ Links ]
26. Bonaparte, J.F. 1986a. The early radiation and phylogenetic relationships of the Jurassic sauropod dinosaurs, based on vertebral anatomy. In Padian, K. (ed.), The Beginning of the Age of Dinosaurs. Cambridge University Press 245-258, Cambridge. [ Links ]
27. Bonaparte, J.F. 1986b. Les Dinosaures (Carnosaures, Allosauridés, Sauropodes, Cétiosauridés) du Jurassique moyen de Cerro Cóndor (Chubut, Argentine). Annales de Paléontologie 72: 247-289, 326-386. [ Links ]
28. Bonaparte, J.F. 1999. Evolución de las vértebras presacras en Sauropodomorpha. Ameghiniana 36(2): 115-187. [ Links ]
29. Bonaparte, J.F., Heinrich, W.-D. and Wild, R. 2000. Review of Janenschia Wild, with the description of a new sauropod from the Tendaguru beds of Tanzania and a discussion on the systematic value of procoelous caudal vertebrae in the Sauropoda. Palaeontographica A 256(1-3): 25-76. [ Links ]
30. Bradmeister, K.C., Flynn, J.J., Parrish, J.M. and Wyss, A.R. 2006. Paleogeographic and biostratigraphic implications of new early Mesozoic vertebrates from Poamay, central Morondava Basin, Madagascar. New Mexico Museum of Natural History and Science Bulletin 37: 457-475. [ Links ]
31. Bristowe, A. and Raath, M.A. 2004. A juvenile coelophysoid skull from the Early Jurassic of Zimbabwe, and the synonymy of Coelophysis and Syntarsus. Palaeontologia africana 40: 31-41. [ Links ]
32. Britt, B.B. 1991. Theropods of Dry Mesa Quarry (Morrison Formation, Late Jurassic), Colorado, with emphasis on the osteology of Torvosaurus tanneri. BYU Geology Studies 37: 1-72. [ Links ]
33. Buchy, M.-C., Stinnesbeck, W., Frey, E. and González, A.H. 2007. Première mention du genre Dakosaurus (Crocodyliformes, Thalattosuchia) dans le Jurassique supérieur du Mexique. Bulletin de la Societé géologique de la France 178(5): 391-397. [ Links ]
34. Buchy, M.-C., Vignaud, P., Frey, E., Stinnesbeck, W., López Oliva, J.G. and Götte, M. 2006. A new thalattosuchian crocodyliform from the Tithonian of northeastern Mexico. Comptes Rendus Palevol 5: 785-794. [ Links ]
35. Buckland, W. 1824. Notice on the Megalosaurus or great fossil lizard of Stonesfield. Transactions of the Geological Society, Series 2, 1: 391-396. [ Links ]
36. Buffetaut, E. 1995. The importance of "Lagerstätten" for our understanding of the evolutionary history of certain groups of organisms: the case of pterosaurs. In 2° International Symposium on Lithographic Limestones (Lleida-Cunca), Universidad Autónoma de Madrid, Extended Abstracts: 49-52, Madrid. [ Links ]
37. Buffetaut, E. 2005. A new sauropod dinosaur with prosauropod-like teeth from the Middle Jurassic of Madagascar. Bulletin de la Societé géologique de la France 176(5): 467-473. [ Links ]
38. Buscalioni, A.D., Ortega, F., Pérez-Moreno, B.P. and Evans, S.E. 1996. The Upper Jurassic maniraptoran theropod Lisboasaurus estesi (Guimarota, Portugal) reinterpreted as a crocodylomorph. Journal of Vertebrate Paleontology 16(2): 358-362. [ Links ]
39. Butler, R.J. 2005. The 'fabrosaurid' ornithischian dinosaurs of the Upper Elliot Formation (Lower Jurassic) of South Africa and Lesotho. Zoological Journal of the Linnean Society 145: 175-218. [ Links ]
40. Butler, R.J., Smith, R.M.H. and Norman, D.B. 2007. A primitive ornithischian from the Late Triassic of South Africa, and the early evolution and diversification of Ornithischia. Proceedings of the Royal Society London B 274: 2041-2046. [ Links ]
41. Butler, R.J., Upchurch, P. and Norman, D.B. 2008. The phylogeny of the ornithischian dinosaurs. Journal of Systematic Palaeontology 6(1): 1-40. [ Links ]
42 Báez, A.M. and Marsicano, C.A. 2001. A heterodontosaurid ornithischian dinosaur from the Upper Triassic of Patagonia. Ameghiniana 38 (3): 271-279. [ Links ]
43. Cabrera, A. 1947. Un saurópodo nuevo del Jurásico de Patagonia. Notas del Museo de La Plata, Paleontología 12(95): 1-17. [ Links ]
44. Carballido, J.L., Pol, D. and Rauhut, O.W.M. 2007. Relaciones filogenéticas de Tehuelchesaurus benitezii Rich, Gimenez, Cúneo, Puerta y Vacca (Dinosauria, Eusauropoda). Ameghiniana 44(4): 90R. [ Links ]
45. Carpenter, K., Miles, C. and Cloward, K. 1998. Skull of a Jurassic ankylosaur (Dinosauria). Nature 393: 782-783. [ Links ]
46. Carpenter, K., Miles, C.A. and Cloward, K. 2001. New primitive stegosaur from the Morrison Formation, Wyoming. In Carpenter, K. (ed.) The Armored Dinosaurs, Indiana University Press 55-75, Bloomington and Indianapolis. [ Links ]
47. Carpenter, K., Miles, C. and Cloward, K. 2005. New small theropod from the Upper Jurassic Morrisson Formation of Wyoming. In Carpenter, K. (ed.) The Carnivorous Dinosaurs, Indiana University Press, 23-48, Bloomington and Indianapolis. [ Links ]
48. Carrano, M.T. and Sampson, S.D. 2004. A review of coelophysoids (Dinosauria: Theropoda) from the Early Jurassic of Europe, with comments on the late history of the Coelophysoidea. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 2004 (9): 537-558. [ Links ]
49. Casamiquela, R.M. 1963. Consideraciones acerca de Amygdalodon Cabrera (Sauropoda, Cetiosauridae) del Jurásico Medio de la Patagonia. Ameghiniana 3(3): 79-95. [ Links ]
50. Casamiquela, R.M. 1967. Un nuevo dinosaurio ornitisquio Triasico (Pisanosaurus mertii, Ornithopoda) de la Formacion Ischigualasto, Argentina. Ameghiniana 5(2): 47-64. [ Links ]
51. Casamiquela, R.M. 1975. Herbstosaurus pigmaeus (Coeluria, Compsognathidae) n. gen. n. sp. del Jurásico medio del Neuquen (Patagonia septentrional). Uno de los más pequeños dinosaurios conocidos. Actas del Congreso Internacional de Paleontologia y Biostratigrafia 1: 87-101. [ Links ]
52. Chabli, S. 1985. Données nouvelles sur un "Dinosaurien" Jurassique Moyen du Maroc: Megalosaurus mersensis Lapparent 1955, et sur les Megalosaurides en général. In Les Dinosaures de la Chine à la France. Musée d'Histoire Naturelle de Toulouse: 66-72, Toulouse. [ Links ]
53. Chinsamy, A. 1990. Physiological implications of the bone histology of Syntarsus rhodesiensis (Saurischia: Theropoda). Palaeontologia africana 27: 77-82. [ Links ]
54. Chong Diaz, G. and Gasparini, Z.B.d. 1972. Presencia de Crocodilia marinos en el Jurásico de Chile. Revista de la Asociación Geológica Argentina 27(4): 406-409. [ Links ]
55. Chong Diaz, G. and Gasparini, Z.B.d. 1976. Los vertebrados Mesozoicos de Chile y su aporte geo-paleontológico. 6° Congreso Geológico Argentino, Actas 1: 45-67. [ Links ]
56. Chure, D.J. 2001. The second record of the African theropod Elaphrosaurus (Dinosauria, Ceratosauria) from the Western Hemisphere. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 2001(9): 565-576. [ Links ]
57. Clark, J.M., Hopson, J.A., Hernández R., R., Fastovsky, D.E. and Montellano, M. 1998. Foot posture in a primitive pterosaur. Nature 391: 886-889. [ Links ]
58. Clark, J.M. and Sues, H.-D. 2002. Two new basal crocodylomorph archosaurs from the Lower Jurassic and the monophyly of the Sphenosuchia. Zoological Journal of the Linnean Society 136: 77-95. [ Links ]
59. Clark, J.M., Xu, X., Forster, C.A. and Wang, Y. 2004. A Middle Jurassic "sphenosuchian" from China and the origin of the crocodylian skull. Nature 430: 1021-1024. [ Links ]
60. Colbert, E.H. 1969. A Jurassic pterosaur from Cuba. American Museum Novitates 2370: 1-26. [ Links ]
61. Colbert, E.H. 1981. A primitive ornithischian dinosaur from the Kayenta Formation of Arizona. Bulletin of the Museum of Northern Arizona 53: 1-61. [ Links ]
62. Colbert, E.H. and Mook, C.C. 1951. The ancestral crocodilian Protosuchus. Bulletin of the American Museum of Natural History 97: 143-182. [ Links ]
63. Cooper, M.R. 1981. The prosauropod dinosaur Massospondylus carinatus Owen from Zimbabwe: its biology, mode of life and phylogenetic significance. Occasional Papers, National Museum of Rhodesia B 6(10): 689-840. [ Links ]
64. Cooper, M.R. 1984. A reassessment of Vulcanodon karibaensis Raath (Dinosauria: Saurischia) and the origin of the Sauropoda. Palaeontologia africana 25: 203-231. [ Links ]
65. Codorniú, L. and Gasparini, Z. 2007. Pterosauria. In Gasparini, Z., Salgado, L. and Coria, R.A. (eds.), Patagonian Mesozoic reptiles. Indiana University Press, 143-166, Bloomington and Indianapolis. [ Links ]
66. Codorniú, L., Gasparini, Z. and Paulina-Carabajal, A. 2006. A Late Jurassic pterosaur (Reptilia, Pterodactyloidea) from northwestern Patagonia, Argentina. Journal of South American Earth Sciences 20: 383-389. [ Links ]
67. Crompton, A.W. and Charig, A.J. 1962. A new ornithischian from the Upper Triassic of South Africa. Nature 196: 1074-1077. [ Links ]
68. Crompton, A.W. and Smith, K.K. 1980. A new genus and species of crocodilian from the Kayenta Formation (Late Triassic?) of northern Arizona. In Jacobs, L.L. (ed.) Aspects of Vertebrate History, Museum of Northern Arizona 193-217, Flagstaff. [ Links ]
69. Dodson, P., Behrensmeyer, A.K., Bakker, R.T. and McIntosh, J.S. 1980. Taphonomy and paleoecology of the dinosaur beds of the Jurassic Morrison Formation. Paleobiology 6(2): 208-232. [ Links ]
70. Dong, Z. 1982. On a new Pterosauria (Huanhepterus quingyangensis gen. et sp. nov.) from Ordos, China. Vertebrata PalAsiatica 20 (2):115-121 [ Links ]
71. Dong, Z. 1984. A new theropod dinosaur from the Middle Jurassic of Sichuan Basin. Vertebrata PalAsiatica 22(3): 213-218 (in Chinese). [ Links ]
72. Dong, Z.-M. 2001. Primitive armored dinosaur from the Lufeng Basin, China. In Tanke, D.H. and Carpenter, K. (ed.), Mesozoic vertebrate life. Indiana University Press, 237-242, Bloomington and Indianapolis. [ Links ]
73. Dong, Z.-M. and Tang, Z. 1985. A new mid- Jurassic theropod (Gasosaurus constructus gen. et sp. nov.) from Dashanpu, Zigong, Sichuan Province, China. Vertebrata PalAsiatica 23 (1): 78-83 (in Chinese). [ Links ]
74. Evans, S.E. and Milner, A.R. 1994. Middle Jurassic microvertebrate assemblages from the British Isles. In Fraser, N.C. and Sues, H.-D. (eds.) In the shadow of the dinosaurs. Early Mesozoic tetrapods, Cambridge University Press, 303-321, Cambridge. [ Links ]
75. Fastnacht, M. 2005. The first dsungaripterid pterosaur from the Kimmeridgian of Germany and the biomechanics of pterosaur long bones. Acta Palaeontologica Polonica 50(2): 273-288. [ Links ]
76. Fedak, T.J. and Galton, P.M. 2007. New information on the braincase and skull of Anchisaurus polyzelus (Lower Jurassic, Connecticut, USA, Saurischia: Sauropodomorpha): implications for sauropodomorph systematics. Special Papers in Palaeontology 77: 245-260. [ Links ]
77. Ferigolo, J. and Langer, M.C. 2007. A Late Triassic dinosauriform from south Brazil and the origin of the ornithischian predentary bone. Historical Biology 19(1): 23-33. [ Links ]
78. Flynn, J.J., Fox, S.R., Parrish, J.M., Ranivoharimanana, L. and Wyss, A.R. 2006. Assessing diversity and paleoecology of a Middle Jurassic microvertebrate assemblage from Madagascar. New Mexico Museum of Natural History and Science Bulletin 37: 476-489. [ Links ]
79. Ford, D. and Golonka, J. 2003. Phanerozoic paleogeography, paleoenvironment and lithofacies maps of the circum-Atlantic margins. Marine and Petroleum Geology 20: 249-285. [ Links ]
80. Forster, C.A. 1999. Gondwanan dinosaur evolution and biogeographic analysis. Journal of African Earth Sciences 169: 169-185. [ Links ]
81. Foster, J.R. 2003. Paleoecological analysis of the vertebrate fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, USA. Bulletin of the New Mexico Museum of Natural History and Science 23: 1-95. [ Links ]
82. Galton, P.M. 1971. Manus movements of the coelurosaurian dinosaur Syntarsus and opposability of the theropod hallux. Arnoldia 5 (15): 1-8. [ Links ]
83. Galton, P.M. 1976. Prosauropod dinosaurs (Reptilia: Saurischia) of North America. Postilla 169, 98 p. [ Links ]
84. Galton, P.M. 1977. The ornithopod dinosaur Dryosaurus and a Laurasia - Gondwanaland connection in the Upper Jurassic. Nature 268: 230-232. [ Links ]
85. Galton, P.M. 1978. Fabrosauridae, the basal family of ornithischian dinosaurs (Reptilia: Ornithopoda). Paläontologische Zeitschrift 52 (1/2): 138-159. [ Links ]
86. Galton, P.M. 1980a. Avian-like tibiotarsi of pterodactyloids (Reptilia: Pterosauria) from the Upper Jurassic of East Africa. Paläontologische Zeitschrift 54 (3/4): 331-342. [ Links ]
87. Galton, P.M. 1980b. European Jurassic ornithopod dinosaurs of the families Hypsilophodontidae and Camptosauridae. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 160(1): 73-95. [ Links ]
88. Galton, P.M. 1982. Elaphrosaurus, an ornithomimid dinosaur from the Upper Jurassic of North America and Africa. Paläontologische Zeitschrift 56(3/4): 265-275. [ Links ]
89. Galton, P.M. 1983. Armored dinosaurs (Ornithischia: Ankylosauria) from the Middle and Upper Jurassic of Europe. Palaeontographica A 182(1-3): 1-25. [ Links ]
90. Galton, P.M. 1990. Stegosauria. In Weishampel, D.B., Dodson, P. and Osmólska, H. (eds.),The Dinosauria. University of California Press, 435-455, Berkeley. [ Links ]
91. Galton, P.M. 2005. Teeth of ornithischian dinosaurs (mostly Ornithopoda) from the Morrison Formation (Upper Jurassic) of the western United States. In Carpenter, K. (ed.), Horns and Beaks. Ceratopsian and ornithopod dinosaurs. Indiana University Press, 17- 48, Bloomington and Indianapolis. [ Links ]
92. Galton, P.M. and Powell, H.P. 1983. Stegosaurian dinosaurs from the Bathonian (Middle Jurassic) of England, the earliest record of the family Stegosauridae. Geobios 16(2): 219-229. [ Links ]
93. Galton, P.M. and Upchurch, P. 2004. Stegosauria. In Weishampel, D.B., Dodson, P. and Osmólska, H. (eds.) The Dinosauria. Second Edition. University of California Press, 343- 362, Berkeley. [ Links ]
94. Gao, Y. 1993. A new species of Szechuanosaurus from the Middle Jurassic of Dashanpu, Zigong, Sichuan. Vertebrata PalAsiatica 31(4): 308-314 (in Chinese). [ Links ]
95. García, R., Salgado, L. and Coria, R.A. 2003. Primeros restos de dinosaurios saurópodos en el Jurásico de la Cuenca Neuquina, Patagonia, Argentina. Ameghiniana 40 (1): 123-126. [ Links ]
96. Gasparini, Z. 1980. Un nuevo cocodrilo marino (Crocodylia, Metriorhynchidae) del Caloviano del norte de Chile. Ameghiniana 17(2): 97-103. [ Links ]
97. Gasparini, Z. 1985. Los reptiles marinos Jurásicos de America del Sur. Ameghiniana 22(1- 2): 23-34. [ Links ]
98. Gasparini, Z. 1992. Marine reptiles of the circum-Pacific region. In Westerman, G.E.G. (ed.), The Jurassic of the Circum-Pacific. Cambridge University Press, 361-365, Cambridge. [ Links ]
99. Gasparini, Z. 1996. Biogeographic evolution of the South American crocodilians. Münchener Geowissenschaftliche Abhandlungen 30: 159- 184. [ Links ]
100. Gasparini, Z. and Chong Diaz, G. 1977. Metriorhynchus casamiquelai n. sp. (Crocodylia, Thalattosuchia), a marine crocodile from the Jurassic (Callovian) of Chile, South America. Neues Jahrbuch für Geologie und Paläontologie, Abhandulngen 153(3): 341-360. [ Links ]
101. Gasparini, Z. and Fernández, M.S. 1997. Tithonian marine reptiles of the eastern Pacific. In Callaway, J.M. and Nicholls, E.L. (eds.) Ancient marine reptiles. Academic Press, 435- 450, London. [ Links ]
102. Gasparini, Z., Cichowolski, M. and Lazo, D.G. 2005. First record of Metriorhynchus (Reptilia: Crocodyliformes) in the Bathonian (Middle Jurassic) of the eastern Pacific. Journal of Paleontology 79(4): 801-805. [ Links ]
103. Gasparini, Z. and Iturralde-Vinent, M. 2001. Metriorhynchid crocodiles (Crocodyliformes) from the Oxfordian of Western Cuba. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 2001(9): 534-542. [ Links ]
104. Gasparini, Z. and Iturralde-Vinent, M.A. 2006. The Cuban Oxfordian herpetofauna in the Caribbean Seaway. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 240 (3): 343-371. [ Links ]
105. Gasparini, Z., Fernández, M. and De la Fuente, M. 2004. A new pterosaur from the Jurassic of Cuba. Palaeontology 47(4): 919-927. [ Links ]
106. Gasparini, Z., Leanza, H.A. and Garate Zubillaga, J.I. 1987. Un pterosaurio de las calizas litograficas Tithonianas del area de los Catutos, Neuquen, Argentina. Ameghiniana 24(1- 2): 141-143. [ Links ]
107. Gasparini, Z., Paulina-Carabajal, A. and Chong, G. 2008. Un nuevo especimen de cocodrilo marino del Jurásico medio del norte de Chile: revalidación de Metriorhynchus westermanni (Crocodyliformes: Metriorhynchidae). Revista Geológica de Chile (in press). [ Links ]
108. Gasparini, Z., Pol, D. and Spalletti, L.A. 2006. An unusual marine crocodyliform from the Jurassic-Cretaceous boundary of Patagonia. Science 311: 70-73. [ Links ]
109. Gasparini, Z., Spalletti, L., Fernández, M. and de la Fuente, M. 1999. Tithonian marine reptiles from the Neuquén basin: diversity and palaeoenvironments. Revue de Paléobiologie 18(1): 335-345. [ Links ]
110. Gasparini, Z., Vignaud, P. and Chong, G. 2000. The Jurassic Thalattosuchia (Crocodyliformes) of Chile: a paleobiogeographic approach. Bulletin de la Societé géologique de la France 171(6): 657-664. [ Links ]
111. Gillette, D.D. 2003. The geographic and phylogenetic position of sauropod dinosaurs from the Kota Formation (Early Jurassic) of India. Journal of Asian Earth Sciences 21: 683-689. [ Links ]
112. Goodwin, M.B., Clemens, W.A., Hutchinson, J.H., Wood, C.B., Zavada, M.S., Kemp, A., Duffin, C.J. and Schaff, C.R. 1999. Mesozoic continental vertebrates with associated palynostratigraphic dates from the northwestern Ethiopian Plateau. Journal of Vertebrate Paleontology 19(4): 728-741. [ Links ]
113. Gow, C.E. 1990. Morphology and growth of the Massospondylus braincase (Dinosauria, Prosauropoda). Palaeontologia africana 27: 59-75. [ Links ]
114. Gow C.E. 2000. The skull of Protosuchus haughtoni, an early Jurassic Crocodyliform from southern Africa. Journal of Vertebrate Paleontology 20: 49-56. [ Links ]
115. Gow, C.E., Kitching, J.W. and Raath, M.A. 1990. Skulls of the prosauropod dinosaur Massospondylus carinatus Owen in the collections of the Bernard Price Institute for Palaeontological Research. Palaeontologia africana 27: 45- 58. [ Links ]
116. Göhlich, U.B. and Chiappe, L.M. 2006. A new carnivorous dinosaur from the Late Jurassic Solnhofen archipelago. Nature 440: 329-332. [ Links ]
117. Göhlich, U.B., Chiappe, L.M., Clark, J.M. and Sues, H.-D. 2005. The systematic position of the Late Jurassic alleged dinosaur Macelognathus (Crocodylomorpha: Sphenosuchia). Canadian Journal of Earth Sciences 42: 307-321. [ Links ]
118. Hammer, W.R. and Hickerson, W.J. 1994. A crested theropod dinosaur from Antarctica. Science 264: 828-830. [ Links ]
119 Hammer, W.R. and Hickerson, W.J. 1999. Gondwana dinosaurs from the Jurassic of Antarctica. National Science Museum Monographs 15: 211-217. [ Links ]
120. Harris, J.D. and Dodson, P. 2004. A new diplodocoid sauropod dinosaur from the Upper Jurassic Morrison Formation of Montana, USA. Acta Palaeontologica Polonica 49(2): 197-210. [ Links ]
121. Harris, J.D., Lucas, S.G., Estep, J.W. and Li, J. 2000. A new and unusual sphenosuchian (Archosauria: Crocodylomorpha) from the Lower Jurassic Lufeng Formation, People's Republic of China. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 215(1): 47-68. [ Links ]
122. Haubold, H. 1990. Ein neuer Dinosauria (Ornithischia, Thyreophora) aus dem Unteren Jura des nördlichen Mitteleuropa. Revue de Paléobiologie 9(1): 149-177. [ Links ]
123. Haughton S.H. 1915. Investigations in South African fossil reptiles and Amphibia: a new thecodont from the Stormberg Beds. Annals of the South African Museum 12: 98-105. [ Links ]
124. Haughton, S.H. 1924. The fauna and stratigraphy of the Stormberg Series. Annals of the South African Museum 12: 323-497. [ Links ]
125. He, X., Wang, C., Liu, S., Zhou, F., Liu, T., Cai, K. and Dai, B. 1998. A new species of sauropod from the Early Jurassic of Gongxian Co., Sichuan. Acta Geologica Sichuan 18: 1-7. [ Links ]
126. Hennig, E. 1925. Kentrurosaurus aethiopicus. Die Stegosaurier-Funde vom Tendaguru, Deutsch -Ostafrika. Palaeontographica, Supplement 7, 1(1): 103-253. [ Links ]
127. Holtz, T.R.Jr. 2000. A new phylogeny of the carnivorous dinosaurs. Gaia 15: 5-61. [ Links ]
128. Holtz, T.R.Jr., Molnar, R.E. and Currie, P.J. 2004. Basal Tetanurae. In Weishampel, D.B., Dodson, P. and Osmólska, H. (eds.), The Dinosauria. Second Edition. University of California Press, 71-110, Berkeley. [ Links ]
129. Hopson, J.A. 1975. On the generic separation of the ornithischian dinosaurs Lychorhinus and Heterodontosaurus from the Stormberg Series (Upper Triassic) of South Africa. South African Journal of Science 71: 302-305. [ Links ]
130. Hu, S. 1993. A new Theropoda (Dilophosaurus sinensis sp. nov.) from Yunnan, China. Vertebrata PalAsiatica 31(1): 65-69 (in Chinese). [ Links ]
131. Huene, F.von 1927. Beitrag zur Kenntnis mariner mesozoischer Wirbeltiere in Argentinien. Zentralblatt für Mineralogie, Geologie und Paläontologie Abt. B 1: 22-29. [ Links ]
132. Huene, F.von 1938. Ein Pliosauride aus Abessinien. Zentralblatt für Mineralogie, Geologie und Paläontologie, B, 1938 (10): 370-376. [ Links ]
133. Hunt, A.P. and Lockley, M.G. 1995. A nonmarine tetrapod from the Middle Jurassic of the United States: a primitive crocodyliform from the Entrada Sandstone of eastern Utah. Journal of Vertebrate Paleontology 15(3): 554-560. [ Links ]
134. Irmis, R.B. 2004. First report of Megapnosaurus (Theropoda: Coelophysoidea) from China. PaleoBios 24(3): 11-18. [ Links ]
135. Jain, S.L. 1974. Jurassic pterosaur from India. Journal of the Geological Society of India 15(3): 330-335. [ Links ]
136. Jain, S.L., Kutty, T.S., Roy-Chowdhury, T. and Chatterjee, S. 1975. The sauropod dinosaur from the Lower Jurassic Kota Formation of India. Proceedings of the Royal Society of London A 188: 221-228. [ Links ]
137. Jain, S.L., Kutty, T.S., Roy-Chowdhury, T. and Chatterjee, S. 1979. Some characteristics of Barapasaurus tagorei, a sauropod dinosaur from the Lower Jurassic of Deccan, India. In 4° International Gondwana Symposiu (1977, Calcutta), 204-216, Calcutta. [ Links ]
138. Janensch, W. 1914. Übersicht über die Wirbeltierfauna der Tendaguruschichten, nebst einer kurzen Charakterisierung der neu aufgeführten Arten von Sauropoden. Archiv für Biontologie 3(1): 81-110. [ Links ]
139. Janensch, W. 1920. Ueber Elaphrosaurus bambergi und die Megalosaurier aus den Tendaguru- Schichten Deutsch-Ostafrikas. Sitzungsberichte der Gesellschaft naturforschender Freunde zu Berlin, 1920: 225-235. [ Links ]
140. Janensch, W. 1925. Die Coelurosaurier und Theropoden der Tendaguru-Schichten Deutsch-Ostafrikas. Palaeontographica, Supplement 7, 1(1): 1-99. [ Links ]
141. Janensch, W. 1929. Material und Formengehalt der Sauropoden in der Ausbeute der Tendaguru- Expedition. Palaeontographica, Supplement 7, 1 (2): 3-34. [ Links ]
142. Janensch, W. 1955. Der Ornithopode Dysalotosaurus der Tendaguruschichten. Palaeontographica, Supplement 7. 1(3): 105-176. [ Links ]
143. Jouve, S. 2004. Description of a skull of Ctenochasma (Pterosauria) from the latest Jurassic of eastern France, with a taxonomic revision of European Tithonian Pterodactyloidea. Journal of Vertebrate Paleontology 24(3): 542- 554. [ Links ]
144. Kellner, A.W.A. 2003. Pterosaur phylogeny and comments on the evolutionary history of the group. In Buffetaut, E. and Mazin, J.-M. (eds.) Evolution and Palaeobiology of Pterosaurs. Geological Society, Special Publications, 105- 137, London. [ Links ]
145. Kirkland, J.I. and Carpenter, K. 1994. North America's first pre-Cretaceous ankylosaur (Dinosauria) from the Upper Jurassic Morrison Formation of western Colorado. Brigham Young University, Geology Studies 40: 25-42. [ Links ]
146. Knoll, F. 2002a. A nearly complete skull of Lesothosaurus (Dinosauria: Ornithischia) from the Upper Elliot Formation (Lower Jurassic: Hettangian) of Lesotho. Journal of Vertebrate Paleontology 22(2): 238-243. [ Links ]
147. Knoll, F. 2002b. New skull of Lesothosaurus (Dinosauria: Ornithischia) from the Upper Elliot Formation (Lower Jurassic) of southern Africa. Geobios 35: 595-603. [ Links ]
148. Knoll, F. 2005. The tetrapod fauna of the Upper Elliot and Clarens formations in the main Karoo Basin (South Africa and Lesotho). Bulletin de la Societé géologique de la France 176(1): 81-91. [ Links ]
149. Krebs, B. 1967. Der Jura-Krokodilier Machimosaurus H. v. Meyer. Paläontologische Zeitschrift 41(1/2): 46-59. [ Links ]
150. Krebs, B. and Schwarz, D. 2000. The crocodiles from the Guimarota mine. In Martin, T. and Krebs, B. (eds.) Guimarota. A Jurassic Ecosystem. Verlag Dr. Friedrich Pfeil, 69-74, Munich. [ Links ]
151. Kriwet, J., Rauhut, O.W.M. and Gloy, U. 1997. Microvertebrate remains (Pisces, Archosauria) from the Middle Jurassic (Bathonian) of southern France. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 206(1): 1- 28. [ Links ]
152. Kutty, T.S., Chatterjee, S., Galton, P.M. and Upchurch, P. 2007. Basal sauropodomorphs (Dinosauria: Saurischia) from the Lower Jurassic of India: their anatomy and relationships. Journal of Paleontology 81(6): 1218- 1240. [ Links ]
153. Kutty, T.S., Jain, S.L. and RoyChowdhury, T.K. 1987. Gondwana sequence of the northern Pranhita-Godavari valley: its stratigraphy ans vertebrate faunas. The Palaeobotanist 36: 214- 229. [ Links ]
154. Kvale, E.P., Johnson, G.D., Mickelson, D.L., Keller, K., Furer, L.C. and Archer, A.W. 2001. Middle Jurassic (Bajocian and Bathonian) dinosaur megatracksites, Bighorn Basin, Wyoming, U.S.A. Palaios 16: 233-254. [ Links ]
155. Langston, W. and Durham, J.W. 1955. A sauropod dinosaur from Columbia. Journal of Paleontology 29(6): 1047-1051. [ Links ]
156. Lapparent, A.F.d. 1955. Étude paléontologique des vertébrés du Jurassique d'el Mers. Notes et Mémoires du Service Géologique du Maroc 124: 1-36. [ Links ]
157. Long, J.A. and Molnar, R.E. 1998. A new Jurassic theropod dinosaur from western Australia. Records of the Western Australian Museum 19: 121-129. [ Links ]
158. Longman, H.A. 1926. A giant dinosaur from Durham Downs, Queensland. Memoirs of the Queensland Museum 8(3): 183-194. [ Links ]
159. Longman, H.A. 1927. The giant dinosaur: Rhoetosaurus brownei. Memoirs of the Queensland Museum 9(1): 1-18. [ Links ]
160. Lull, R.S. 1915. Sauropoda and Stegosauria of the Morrison of North America compared with those of Europe and Eastern Africa. Bulletin of the Geological Society of America 26: 323-334. [ Links ]
161. Luo, Z. and Wu, X.-C. 1994. The small tetrapods of the Lower Lufeng Formation, Yunnan, China. In Fraser, N.C. and Sues, H.-D. (eds.) In the shadow of the dinosaurs. Early Mesozoic tetrapods. Cambridge University Press, 251-270, Cambridge. [ Links ]
162. Madsen, J.H. 1976. Allosaurus fragilis: a revised osteology. Utah Geological and Mineralogical Survey Bulletin 109: 3-163. [ Links ]
163. Madsen, J.H.J. and Welles, S.P. 2000. Ceratosaurus (Dinosauria, Theropoda). A revised osteology. Miscellaneous Publication, Utah Geological Survey 00-2: 1-80. [ Links ]
164. Maganuco, S., Cau, A. and Pasini, G. 2005. First description of theropod remains from the Middle Jurassic (Bathonian) of Madagascar. Atti della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale in Milano 146(2): 165-202. [ Links ]
165. Mahammed, F., Läng, É., Mami, L., Mekahli, L., Benhamou, M., Boumediène, B., Kacemi, A., Chérief, S.-A., Chaouati, H. and Taquet, P. 2005. The 'giant of Ksour', a Middle Jurassic sauropod dinosaur from Algeria. Comptes Rendus Palevol 4: 707-714. [ Links ]
166. Maidment, S.C.R. and Wei, G. 2006. A review of the Late Jurassic stegosaurs (Dinosauria, Stegosauria) from the People's Republic of China. Geological Magazine 143(5): 621-634. [ Links ]
167. Maier, G. 2003. African dinosaurs unearthed. Indiana University Press, 380 p, Bloomington. [ Links ]
168. Maisch, M.W., Matzke, A.T. and Stöhr, H. 2003. Sunosuchus (Archosauria, Crocodyliformes) from the Toutunhe Formation (Middle Jurassic) of the southern Junggar Basin (Xinjiang, NW-China). Geobios 36: 391-400. [ Links ]
169. Marsh, O.C. 1884. Principal characters of American Jurassic dinosaurs. Part VIII. The order Theropoda. American Journal of Science (series 3) 27: 329-340. [ Links ]
170. Martill, D.M., Frey, E.D., Chong Diaz, G. and Bell, C.M. 2000. Reinterpretation of a Chilean pterosaur and the occurrence of Dsungaripteridae in South America. Geological Magazine 137(1): 19-25. [ Links ]
171. Martínez, R.N. 1999. The first Southamerican record of Massospondylus (Dinosauria: Sauropodomorpha). Journal of Vertebrate Paleontology 19 (supplement to 3): 61A. [ Links ]
172. Mateus, O. 2006. Late Jurassic dinosaurs from the Morrison formation (USA), the Lourinha and Alcobaça Formations (Portugal), and the Tendaguru Beds (Tanzania): A Comparison. New Mexico Museum of Natural History and Science Bulletin 36: 1-9. [ Links ]
173. Mateus, O., Walen, A. and Antunes, M.T. 2006. The large theropod fauna of the Lourinha Formatuion (Portugal) and its similarity to that of the Morrison Formation, with a description of new species of Allosaurus. New Mexico Museum of Natural History and Science Bulletin 36: 123-129. [ Links ]
174. Mayr, G., Pohl, B. and Peters, D.S. 2005. A wellpreserved Archaeopteryx specimen with theropod features. Science 310: 1483-1486. [ Links ]
175. Monbaron, M., Russell, D.A. and Taquet, P. 1999. Atlasaurus imelakei n. g., n. sp., a brachiosauridlike sauropod from the Middle Jurassic of Morocco. Comptes Rendus de l'Academie des Sciences de Paris 329: 519-526. [ Links ]
176. Moody, J.M. 1997. Theropod teeth from the Jurassic of Venezuela. Boletín de la Sociedad Venezolana de Geólogos 22(2): 37-42. [ Links ]
177. Moser, M., Mathur, U.B., Fürsich, F.T., Pandey, D.K. and Mathur, N. 2006. Oldest camarasauromorph sauropod (Dinosauria) discovered in the Middle Jurassic (Bajocian) of the Khadir Island, Kachchh, western India. Paläontologische Zeitschrift 80(1): 34-51. [ Links ]
178. Moyne, S., Neige, P., Marchand, D. and Thierry, J. 2004. Repartition mondial des faunes d'ammonites au Jurassique moyen (Aalenien superieur a Bathonien moyen): relations entre biodiversite et paleogeographie. Bulletin de la Societé géologique de la France 175(5): 513-523. [ Links ]
179. Mueller-Töwe, I.J. 2006. Anatomy, phylogeny, and palaeoecology of the basal thalattosuchians (Mesoeucrocodylia) from the Liassic of central Europe. Johannes Gutenberg-Universität, Fachbereich Chemie, Pharmazie und Geowissenschaften, PhD thesis (unpublished), 1-422, Mainz. [ Links ]
180. Munter, R.C. and Clark, J.M. 2006. Theropod dinosaurs from the Early Jurassic of Huizachal Canyon, Mexico. In Carrano, M.T., Gaudin, T.J., Blob, R.W. and Wible, J.R. (eds.), Amniote paleobiology: perspectives on the evolution of mammals, birds, and reptiles. University of Chicago Press, 53-75, Chicago. [ Links ]
181. Nash D.S. 1968. A crocodile from the Upper Triassic of Lesotho. Journal of Zoology 156: 163-179. [ Links ]
182. Nath, T.T., Yadagiri, P. and Moitra, A.K. 2002. First record of armoured dinosaur from the Lower Jurassic Kota Formation, Pranhita- Godavari Valley, Andrah Pradesh. Journal of the Geological Society of India 59: 575-577. [ Links ]
183. Norman, D.B. 2004. Basal Iguanodontia. In Weishampel, D.B., Dodson, P. and Osmólska, H. (eds.) The Dinosauria. Second Edition. University of California Press, 413-437, Berkeley. [ Links ]
184. Norman, D.B., Butler, R.J. and Maidment, S.C.R. 2007. Reconsidering the status and affinities of the ornithischian dinosaur Tatisaurus oehleri Simmons, 1965. Zoological Journal of the Linnean Society 150: 865-874. [ Links ]
185. Norman, D.B., Sues, H.-D., Witmer, L.M. and Coria, R.A. 2004. Basal Ornithopoda. In Weishampel, D.B., Dodson, P. and Osmólska, H. (eds.) The Dinosauria. Second Edition. University of California Press, 393-412, Berkeley. [ Links ]
186. Olsen, P.E., Kent, D.V., Sues, H.-D., Koeberl, C., Huber, H., Montanari, A., Rainforth, E.C., Fowell, S.J., Szajna, M.J. and Hartline, B.W. 2002. Ascent of dinosaurs linked to an iridium anomaly at the Triassic-Jurassic boundary. Science 296: 1305-1307. [ Links ]
187. Ostrom, J.H. 1978. The osteology of Compsognathus longipes Wagner. Zitteliana, 4: 73- 118. [ Links ]
188. Ostrom, J.H. and McIntosh, J.S. 1966. Marsh's dinosaurs. The collections from Como Bluff. Yale University Press, 388 p, New Haven. [ Links ]
189. Owen R. 1854. Descriptive catalogue of the fossil organic remains of reptilia and pisces contained in the Museum of the Royal College of Surgeons of England. British Museum (Natural History), 184 p, London. [ Links ]
190. Owen, R. 1863. A monograph of the fossil reptilia of the Liassic Formations. II. Scelidosaurus harrisonii. Continued. Palaeontographical Society Monographs 14: 1-26. [ Links ]
191. Padian, K. 1983. Osteology and functional morphology of Dimorphodon macronyx (Buckland) (Pterosauria: Rhamphorhynchoidea) based on new material in the Yale Peabody Museum. Postilla 189: 1-44. [ Links ]
192. Padian, K. 1984. Pterosaur remains from the kayenta Formation (Early Jurassic) of Arizona. Palaeontology 27: 407-413. [ Links ]
193. Padian, K. 1989. Presence of the dinosaur Scelidosaurus indicates Jurassic age for the Kayenta Formation (Glen Canyon Group, northern Arizona). Geology 17: 438-441. [ Links ]
194. Padian, K. and Wild, R. 1992. Studies of Liassic Pterosauria, I. The holotype and referred specimens of the Liassic pterosaur Dorygnathus banthensis (Theodori) in the Petrefaktensammlung Banz, northern Bavaria. Palaeontographica A 225(1-3): 59-77. [ Links ]
195. Parrish, J.T. 1993. Climate of the supercontinent Pangea. Journal of Geology 101: 215-233. [ Links ]
196. Peng, G., Ye, Y., Yuhui, G., Shu, C. and Jiang, S. 2005. Jurassic dinosaur faunas in Zigong. Zigong Dinosaur Museum, 236 p., Zigong (in Chinese). [ Links ]
197. Perea, D., Ubilla, M. and Rojas, A. 2003. First report of theropods from the Tacuarembó Formation (Late Jurassic-Early Cretaceous), Uruguay. Alcheringa 27: 79-83. [ Links ]
198. Pol, D. and Garrido, A.C. 2007. Un nuevo dinosaurio sauropodomorfo basal de la Formación Las Leoneras (Jurásico inferior), Chubut, Argentina. Ameghiniana 44 (4): 64R. [ Links ]
199. Pol, D. and Gasparini, Z. 2007. Crocodyliformes. In Gasparini, Z., Salgado, L. and Coria, R.A. (eds.), Patagonian Mesozoic reptiles. Indiana University Press, 116-142, Bloomington and Indianapolis. [ Links ]
200. Pol, D., Ji, S., Clark, J.M. and Chiappe, L.M. 2004. Basal crocodyliforms from the Lower Cretaceous Tugulu Group (Xinjiang, China), and the phylogenetic position of Edentosuchus. Cretaceous Research 25: 603-622. [ Links ]
201. Prasad, G.V.R. and Manhas, B.K. 2007. A new docodont mammal from the Jurassic Kota Formation of India. Palaeontologia electronica 10(2): 1-11. [ Links ]
202. Prasad, G.V.R., Manhas, B.K. and Arratia, G. 2004. Elasmobranch and actinopterygian remains from the Jurassic and Cretaceous of India. In Arratia, G. and Tintori, A. (eds.), Mesozoic fishes 3 - systematics, paleoenvirnments and biodiversity. Verlag Dr. Friedrich Pfeil 625-638, Munich. [ Links ]
203. Raath, M.A. 1969. A new coelurosaurian dinosaur from the Forest Sandstone of Rhodesia. Arnoldia 4(28): 1-25. [ Links ]
204. Raath, M.A. 1972. Fossil vertebrate studies in Rhodesia: a new dinosaur (Reptilia: Saurischia) from near the Trias-Jurassic boundary. Arnoldia 30(5): 1-37. [ Links ]
205. Raath, M.A. 1977. The anatomy of the Triassic theropod Syntarsus rhodesiensis (Saurischia: Podokesauridae) and a consideration of its Biology. Rhodes University, PhD thesis (unpublished), Grahamstown. [ Links ]
206. Raath, M.A. 1980. The theropod dinosaur Syntarsus (Saurischia: Podokesauridae) discovered in South Africa. South African Journal of Science 76: 375-376. [ Links ]
207. Raath, M.A. 1985. The theropod Syntarsus and its bearing on the origin of birds. In Hecht, M. K., Ostrom, J.H., Viohl, G. and Wellnhofer, P. (eds.) The beginning of birds. Freunde des Jura Museums, 219-227, Eichstätt. [ Links ]
208. Raath, M.A. 1990. Morphological variation in small theropods and its meaning in systematics: evidence from Syntarsus rhodesiensis. In Carpenter, K. and Currie, P.J. (eds.), Dinosaur systematics. Approaches and perspectives. Cambridge University Press, 91-105, Cambridge. [ Links ]
209. Raath, M.A. and McIntosh, J.S. 1987. Sauropod dinosaurs from the Central Zambesi Valley, Zimbabwe, and the age of the Kadzi Formation. South African Journal of Geology 90(2): 107-119. [ Links ]
210. Rauhut, O.W.M. 2002. Dinosaur evolution in the Jurassic: A South American perspective. Journal of Vertebrate Paleontology 22 (suppl. to 3): 89A. [ Links ]
211. Rauhut, O.W.M. 2003a. A dentary of Patagosaurus (Sauropoda) from the Middle Jurassic of Patagonia. Ameghiniana 40(3): 425-432. [ Links ]
212. Rauhut, O.W.M. 2003b. Revision of Amygdalodon patagonicus Cabrera, 1947 (Dinosauria: Sauropoda). Mitteilungen aus dem Museum für Naturkunde in Berlin, Geowissenschaftliche Reihe 6: 173-181. [ Links ]
213. Rauhut, O.W.M. 2003c. The interrelationships and evolution of basal theropod dinosaurs. Special Papers in Palaeontology 69: 1-213. [ Links ]
214. Rauhut, O.W.M. 2005a. Osteology and relationships of a new theropod dinosaur from the Middle Jurassic of Patagonia. Palaeontology 48(1): 87-110. [ Links ]
215. Rauhut, O.W.M. 2005b. Postcranial remains of "coelurosaurs" (Dinosauria, Theropoda) from the Late Jurassic of Tanzania. Geological Magazine 142(1): 97-107. [ Links ]
216. Rauhut, O.W.M. 2005c. Theropod dinosaurs from the Late Jurassic of Tendaguru, Tanzania. Kaupia 14: 94. [ Links ]
217. Rauhut, O.W.M. 2006a. A brachiosaurid sauropod from the Late Jurassic Cañadón Calcàreo Formation of Chubut, Argentina. Fossil Record 9(2): 226-237. [ Links ]
218. Rauhut, O.W.M. 2006b. Theropod dinosaurs from the Late Jurassic of Tanzania and the origin of Cretaceous Gondwanan theropod faunas. Journal of Vertebrate Paleontology 26 (Suppl. to 3): 113A. [ Links ]
219. Rauhut, O.W.M. 2006c. Dinosaurier aus dem oberen Jura Südamerikas. Freunde der Bayerischen Staatssammlung für Paläontologie und Historische Geologie e.V., Jahresbericht und Mitteilungen 34: 56-70. [ Links ]
220 Rauhut, O.W.M., López-Arbarello, A., Puerta, P. and Martin, T. 2001. Jurassic vertebrates from Patagonia. Journal of Vertebrate Paleontology 21 (suppl. to 3): 91A. [ Links ]
221. Rauhut, O.W.M., Martin, T., Ortiz-Jaureguizar, E. and Puerta, P. 2002. A Jurassic mammal from South America. Nature 416: 165-168. [ Links ]
222. Rauhut, O.W.M., Remes, K., Fechner, R., Cladera, G. and Puerta, P. 2005. Discovery of a shortnecked sauropod dinosaur from the Late Jurassic period of Patagonia. Nature 435: 670- 672. [ Links ]
223. Reck, H. 1931. Die deutschostafrikanischen Flugsaurier. Centralblatt für Mineralogie, Geologie und Paläontologie B 1931 (7): 321-336. [ Links ]
224. Reisz, R.R., Scott, D., Sues, H.-D., Evans, D.C. and Raath, M.A. 2005. Embryos of an Early Jurassic prosauropod dinosaur and their evolutionary significance. Science 309: 761-764. [ Links ]
225. Remes, K. 2006. Revision of the Tendaguru sauropod Tornieria africana (Fraas) and its relevance for sauropod paleobiogeography. Journal of Vertebrate Paleontology 26(3): 651-669. [ Links ]
226. Remes, K. 2007. A second Gondwanan diplodocid dinosaur from the Upper Jurassic Tendaguru Beds of Tanzania, East Africa. Palaeontology 50(3): 653-667. [ Links ]
227. Remes, K. and Rauhut, O.W.M. 2005. New insights into the origin and evolution of diplodocoid sauropods. Journal of Vertebrate Paleontology 25 (Suppl to 3): 104A. [ Links ]
228. Rich, T.H., Vickers-Rich, P., Gimenez, O., Cúneo, R., Puerta, P. and Vacca, R. 1999. A new sauropod dinosaur from Chubut Province, Argentina. National Science Museum Monographs 15: 61-84. [ Links ]
229. Rougier, G.W., Garrido, A., Gaetano, L., Puerta, P., Corbitt, C. and Novacek, M.J. 2007. First Jurassic triconodont from South America. American Museum Novitates 3580: 1-20. [ Links ]
230. Rougier, G.W., Martinelli, A.G., Forasiepi, A.M. and Novacek, M.J. 2007. New Jurassic mammals from Patagonia, Argentina: a reappraisal of australosphenidan morphology and interrelationships. American Museum Novitates 3566: 1-54. [ Links ]
231. Rowe, T. 1989. A new species of the theropod dinosaur Syntarsus from the Early Jurassic Kayenta Formation of Arizona. Journal of Vertebrate Paleontology 9(2): 125-136. [ Links ]
232. Ruiz-Omeñaca, J.I., Pereda Suberbiola, X. and Galton, P.M. 2005. Callovosaurus leedsi, the earliest dryosaurid dinosaur (Ornithischia: Euornithopoda) from the Middle Jurassic of England. In Carpenter, K. (ed.) Horns and Beaks. Ceratopsian and ornithopod dinosaurs. Indiana University Press, 3-16, Bloomington and Indianapolis. [ Links ]
233. Russell, D.A. 1993. The role of central Asia in dinosaurian biogeography. Canadian Journal of Earth Sciences 30: 2002-2012. [ Links ]
234. Russell, D.A., Béland, P. and McIntosh, J.S. 1980. Paleoecology of the dinosaurs of Tendaguru (Tanzania). Mémoires de la Societé géologique de la France, Nouvelle Série 139: 169-175. [ Links ]
235. Russell, D.E., Odreman Rivas, O., Battail, B. and Russell, D.A. 1992. Découverte de vertébrés fossils dans la Formation de La Quinta, Jurassique du Vénézuela Occidental. Comptes rendus de l'Académie des Sciences, Paris, Série 2, 314: 1247-1252. [ Links ]
236. Salgado, L., de la Cruz, R., Suárez, M., Fernández, M.S., Gasparini, Z., Palma-Heldt, S. and Fanning, M. 2008. First Late Jurassic dinosaur bones from Chile. Journal of Vertebrate Paleontology 28(2): 529-534. [ Links ]
237. Salgado, L. and Gasparini, Z. 2004. El registro más antiguo de Dinosauria en la Cuenca Neuquina (Aaleniano, Jurásico Medio). Ameghiniana 41(3): 505-508. [ Links ]
238. Sánchez-Villagra, M.R. and Clark, J.M. 1994. An ornithischian from the Jurassic of the Venezuelan Andes. Journal of Vertebrate Paleontology 14(3, Suppl.): 44A. [ Links ]
239. Schwarz, D. 2002. A new species of Goniopholis from the Upper Jurassic of Portugal. Palaeontology 45(1): 185-208. [ Links ]
240. Schwarz, D. and Fechner, R. 2004. Lusitanosuchus, a new generic name for Lisboasaurus mitracostatus (Crocodylomorpha: Mesoeucrocodylia), with a description of new remains from the Upper Jurassic (Kimmeridgian) and Lower Cretaceous (Berriasian) of Portugal. Canadian Journal of Earth Sciences 41: 1259-1271. [ Links ]
241. Scotese, C.R. 1991. Jurassic and Cretaceous plate tectonic reconstructions. Palaeogeographc, Palaeoclimatology, Palaeoecology 87: 493- 501. [ Links ]
242. Scotese, C.R., Boucout, A.J. and McKerrow, W.S. 1999. Gondwanan palaeogeography and palaeoclimatology. Journal of African Earth Sciences 28(1): 99-114. [ Links ]
243. Sereno, P.C. 1991. Lesothosaurus, "fabrosaurids", and the early evolution of Ornithischia. Journal of Vertebrate Paleontology 11(2): 168-197. [ Links ]
244. Sereno, P.C. 1999a. The evolution of dinosaurs. Science 284: 2137-2147. [ Links ]
245. Sereno, P.C. 1999b. Dinosaurian biogeography: vicariance, dispersal and regional extinction. National Science Museum Monographs 15: 249-257. [ Links ]
246. Shubin, N.H., Olsen, P.E. and Sues, H.-D. 1994. Early Jurassic small tetrapods from the McCoy Brook Formation of Nova Scotia, Canada. In Fraser, N.C. and Sues, H.-D. (eds.) In the shadow of the dinosaurs. Early Mesozoic tetrapods. Cambridge University Press, 242-250, Cambridge. [ Links ]
247. Silva Nieto, D.G., Cabaleri, N.G., Armella, C., Volkheimer, W. and Gallego, O. 2007. Hipótesis sobre la evolución tecto-sedimentaria de los depocentros de la cuenca de Cañadón Asfalto (Jurásico-Cretácico), provincia del Chubut. Ameghiniana 44(4): 67R. [ Links ]
248. Simmons, D.J. 1965. The non-therapsid reptiles of the Lufeng Basin, Yunnan, China. Fieldiania: Geology 15(1): 1-93. [ Links ]
249. Smith, A.G., Smith, D.G. and Funnell, B.M. 1994. Atlas of Mesozoic and Cenozoic coastlines. Cambridge University Press, 99 p., Cambridge. [ Links ]
250. Smith, N.D., Makovicky, P.J., Hammer, W.R. and Currie, P.J. 2007. Osteology of Cryolophosaurus ellioti (Dinosauria: Theropoda) from the Early Jurassic of Antarctica and implications for early theropod evolution. Zoological Journal of the Linnean Society 151: 377-421. [ Links ]
251. Smith, N.D., Makovicky, P.J., Pol. D., Hammer, W.R. and Currie, P.J. 2007. The dinosaurs of the Early Jurassic Hanson Formation of the Central Transantarctic Mountains: phylogenetic review and synthesis. In Cooper, A.K. and Raymond, C.R. (eds.) Antarctica: A keystone in a changing world - Online Proceedings of the 10th ISAES. USGS Open-File Report 2007-1047, Short Research paper 003, 5 p. [ Links ]
252. Smith, N.D. and Pol, D. 2007. Anatomy of a basal sauropodomorph dinosaur from the Early Jurassic Hanson Formation of Antarctica. Acta Palaeontologica Polonica 52(4): 657-674. [ Links ]
253. Sues, H.-D., Clark, J.M. and Jenkins, F.A. 1994. A review of the Early Jurassic tetrapods from the Glen Canyon Group of the American southwest. In Fraser, N.C. and Sues, H.-D. (eds.) In the shadow of the dinosaurs. Early Mesozoic tetrapods. Cambridge University Press, 284-294, Cambridge. [ Links ]
254. Sues, H.-D., Olsen, P.E., Carter, J.G. and Scott, D.M. 2003. A new crocodylomorph archosaur from the Upper Triassic of North Carolina. Journal of Vertebrate Paleontology 23(2): 329-343. [ Links ]
255. Sues, H.-D., Reisz, R.R., Hinic, S. and Raath, M.A. 2004. On the skull of Massospondylus carinatus Owen, 1854 (Dinosauria: Sauropodomorpha) from the Elliot and Clarens Formations (Lower Jurassic) of South Africa. Annals of the Carnegie Museum 73(4): 239- 257. [ Links ]
256. Sues, H.-D., Shubin, N.H., Olsen, P.E. and Amaral, W.W. 1996. On the cranial structure of a new protosuchid (Archosauria: Crocodyliformes) from the McCoy Brook Formation (Lower Jurassic) of Nova Scotia, Canada. Journal of Vertebrate Paleontology 16(1): 34-41. [ Links ]
257. Taquet, P. 1985. Two new Jurassic specimens of coelurosaurs (Dinosauria). In Hecht, M.K., Ostrom, J.H., Viohl, G. and Wellnhofer, P. (eds.) The beginning of birds. Freunde des Jura Museums, 229-232, Eichstätt. [ Links ]
258. Thulborn, R.A. 1970. The skull of Fabrosaurus australis, a Triassic onithischian dinosaur. Palaeontology 13(3): 414-432. [ Links ]
259. Thulborn, R.A. 1972. The post-cranial skeleton of the Triassic ornithischian dinosaur Fabrosaurus australis. Palaeontology 15(1): 29-60. [ Links ]
260. Tykoski, R.S. 2005. Anatomy, ontogeny, and phylogeny of coelophysoid theropods. PhD thesis, University of Texas, (unpublished), 572 p., Austin. [ Links ]
261. Tykoski, R.S. and Rowe, T. 2004. Ceratosauria. In Weishampel, D.B., Dodson, P. and Osmólska, H. (eds.), The Dinosauria. Second Edition. University of California Press, 47-70, Berkeley. [ Links ]
262. Tykoski, R.S., Rowe, T.B., Ketcham, R.A. and Colbert, M.W. 2002. Calsoyasuchus valliceps, a new crocodyliform from the Early Jurassic Kayenta Formation of Arizona. Journal of Vertebrate Paleontology 2(3): 593-611. [ Links ]
263. Unwin, D.M. 2003. On the phylogeny and evolutionary history of pterosaurs. In Buffetaut, E. and Mazin, J.-M. (eds.), Evolution and Palaeobiology of Pterosaurs. Geological Society, Special Publications, 139-190, London. [ Links ]
264. Unwin, D.M. 2006. The pterosaurs from deep time. PI Press, 347 p, New York. [ Links ]
265. Unwin, D.M. and Heinrich, W.-D. 1999. On a pterosaur jaw from the Upper Jurassic of Tendaguru (Tanzania). Mitteilungen aus dem Museum für Naturkunde zu Berlin, Geowissenschaftliche Reihe 2: 121-134. [ Links ]
266. Unwin, D.M., Rauhut, O.W.M. and Haluza, A. 2004. The first "rhamphorhynchoid" from South America and the early history of pterosaurs. In Reitner, J., Reich, M. and Schmidt, G. (eds.) Geobiologie. 74. Jahrestagung der Paläontologischen Gesellschaft, Göttingen, 02. bis 08, Kurzfassungen der Vorträge und Poster. Universitätsdrucke Göttingen, 235- 237, Göttingen. [ Links ]
267. Upchurch, P. 1998. The phylogenetic relationships of sauropod dinosaurs. Zoological Journal of the Linnean Society 124: 43-103. [ Links ]
268. Upchurch, P., Barrett, P.M. and Dodson, P. 2004. Sauropoda. In Weishampel, D.B., Dodson, P. and Osmólska, H. (eds.) The Dinosauria. Second Edition. University of California Press, 259-322, Berkeley. [ Links ]
269. Upchurch, P., Barrett, P.M., Zhao, X. and Xu, X. 2007. A re-evaluation of Chinshakiangosaurus chunghoensis Ye vide Dong 1992 (Dinosauria, Sauropodomorpha): implications for cranial evolution in basal sauropod dinosaurs. Geological Magazine 144(2): 247-262. [ Links ]
270. Upchurch, P., Hunn, C.A. and Norman, D.B. 2002. An analysis of dinosaurian biogeography: evidence for the existence of vicariance and dispersal patterns caused by geological events. Proceedings of theRoyal Society of London B 269: 613-621. [ Links ]
271. Upchurch, P. and Martin, J. 2002. The Rutland Cetiosaurus: the anatomy and relationships of a Middle Jurassic British sauropod dinosaur. Palaeontology 45(6): 1049-1074. [ Links ]
272. Upchurch, P. and Martin, J. 2003. The anatomy and taxonomy of Cetiosaurus (Saurischia, Sauropoda) from the Middle Jurassic of England. Journal of Vertebrate Paleontology 23(1): 208-231. [ Links ]
273. Walker, A.D. 1964. Triassic reptiles from the Elgin area: Ornithosuchus and the origin of carnosaurs. Philosophical Transactions of the Royal Society of London, Series B 248 (744): 53- 134. [ Links ]
274. Walker, A.D. 1970. A revision of the Jurassic reptile Hallopus victor (Marsh), with remarks on the classification of crocodiles. Philosophical Transactions of the Royal Society of London B, 257: 323-372. [ Links ]
275. Walker, A.D. 1990. A revision of Sphenosuchus acutus Haughton, a crocodylomorph reptile from the Elliot Formation (late Triassic or early Jurassic) of South Africa. Philosophical Transactions of the Royal Society of London B, 330: 1-120. [ Links ]
276. Weishampel, D.B., Barrett, P.M., Coria, R.A., Le Loeuff, J., Xu, X., Zhao, X., Sahni, A., Gomani, E.M.P. and Noto, C.R. 2004. Dinosaur distribution. In Weishampel, D.B., Dodson, P. and Osmólska, H. (eds.) The Dinosauria. Second Edition. University of California Press, 517-606, Berkeley. [ Links ]
277. Weishampel, D.B., Dodson, P. and Osmólska, H. (eds.) 2004. The Dinosauria. Second edition. University of California Press, 861 p., Berkeley. [ Links ]
278. Welles, S.P. 1954. New Jurassic dinosaur from the Kayenta Formation of Arizona. Bulletin of the Geological Society of America 65: 591- 598. [ Links ]
279. Welles, S.P. 1984. Dilophosaurus wetherilli (Dinosauria, Theropoda). Osteology and comparisons. Palaeontographica A 185(4-6): 85-180. [ Links ]
280. Wellnhofer, P. 1970. Die Pterodactyloidea (Pterosauria) der Oberjura-Plattenkalke Süddeutschlands. Abhandlungen der Bayerischen Akademie der Wissenschaften, mathematischnaturwissenschaftliche Klasse, Neue Folge, 141: 1-133. [ Links ]
281. Wellnhofer, P. 1971. Die Atoposauridae (Crocodylia, Mesosuchia) der Oberjura-Plattenkalke Bayerns. Palaeontographica A 138(5-6): 133- 165. [ Links ]
282. Wellnhofer, P. 1975. Die Rhamphorhynchoidea (Pterosauria) der Oberjura-Plattenkalke Süddeutschlands. Palaeontographica A, 148, 149: 1-33, 132-186, 1-30. [ Links ]
283. Wellnhofer, P. 1992. A new specimen of Archaeopteryx from the Solnhofen limestone. In Campbell, K.E.J. (ed.) Papers in Avian Paleontology honoring Pierce Brodkorb. Natural History Museum of Los Angeles County, Science Series, 3-23, Los Angeles. [ Links ]
284. Werner, C. 1995. Neue Funde von mesozoischen Wirbeltieren in Äthiopien. Berliner geowissenschaftliche Abhandlungen (E) 16(1): 377- 383. [ Links ]
285. Westermann, G.E.G. 1993. Global bio-events in mid-Jurassic ammonites controlled by seaways. In House, M.R. (ed.), The Ammonoidea: Environment, Ecology and Evolutionary Change. Clarendon Press, 187-226, Oxford. [ Links ]
286. Wiechmann, M.F. and Gloy, U. 2000. Pterosaurs and urvogels from the Guimarota mine. In Martin, T. and Krebs, B. (eds.), Guimarota. A Jurassic Ecosystem. Verlag Dr. Friedrich Pfeil, 83-86, Munich. [ Links ]
287. Wild, R. 1978. Ein Sauropoden-Rest (Reptilia, Saurischia) aus dem Posidonienschiefer (Lias, Toarcium) von Holzmaden. Stuttgarter Beiträge zur Naturkunde, Serie B 41: 1-15. [ Links ]
288. Wild, R. 1991. Janenschia n. g. robusta (E. Fraas 1908) pro Tornieria robusta (E. Fraas 1908) (Reptilia, Saurischia, Sauropodomorpha). Stuttgarter Beiträge zur Naturkunde, Serie B, 173: 1-3. [ Links ]
289. Wilson, J.A. 2002. Sauropod dinosaur phylogeny: critique and cladistic analysis. Zoological Journal of the Linnean Society 136: 217-276. [ Links ]
290. Woodward, A.S. 1910. On a skull of Megalosaurus from the Great Oolite of Minchinhampton (Gloucestershire). Quarterly Journal of the Geological Society of London 66: 111-115. [ Links ]
291. Wu, X.-C. and Sues, H.-D. 1996. Reassessment of Platyognathus hsui Young, 1944 (Archosauria: Crocodyliformes) from the Lower Lufeng Formation (Lower Jurassic) of Yunnan, China. Journal of Vertebrate Paleontology 16(1): 42-48. [ Links ]
292. Wu, X.-C., Brinkman, D.B. and Russell, A.P. 1996. Sunosuchus junggarensis sp. nov. (Archosauria: Crocodyliformes) from the Upper Jurassic of Xinjiang, People's Republic of China. Canadian Journal of Earth Sciences 33(4): 606-630. [ Links ]
293. Xu, X. and Clark, J.M. 2006. New ceratosaurs from the Jurassic Shishugou Formation of western China. Journal of Vertebrate Paleontology 26 (suppl. to 3): 142A. [ Links ]
294. Xu, X., Clark, J.M., Forster, C.A., Norell, M.A., Erickson, G.M., Eberth, D.A., Jia, C. and Zhao, Q. 2006. A basal tyrannosauroid dinosaur from the Late Jurassic of China. Nature 439: 715-718. [ Links ]
295. Xu, X., Forster, C.A., Clark, J.M. and Mo, J. 2006. A basal ceratopsian with transitional features from the Late Jurassic of northwestern China. Proceedings of the Royal Society London B, 273: 2135-2140. [ Links ]
296. Xu, X., Zhao, X.-J. and Clark, J.M. 2001. A new therizinosaur from the Lower Jurassic Lufeng Formation of Yunnan, China. Journal of Vertebrate Paleontology 21(3): 477-483. [ Links ]
297. Yadagiri, P. 1988. A new sauropod Kotasaurus yamanpalliensis from Lower Jurassic Kota Formation of India. Journal of the Geological Survey of India 11: 102-127. [ Links ]
298. Yadagiri, P. 2001. The osteology of Kotasaurus yamanpalliensis, a sauropod dinosaur from the Early Jurassic Kota Formation of India. Journal of Vertebrate Paleontology 21(2): 242- 252. [ Links ]
299. Yates, A.M. 2004. Anchisaurus polyzelus (Hitchcock): the smallest known sauropod dinosaur and the evolution of gigantism among sauropodomorph dinosaurs. Postilla 230: 1-58. [ Links ]
300. Yates, A.M. 2005. A new theropod dinosaur from the Early Jurassic of South Africa and its implications for the early evolution of theropods. Palaeontologia africana 41: 105-122. [ Links ]
301. Yates, A.M. and Kitching, J.W. 2003. The earliest known sauropod dinosaur and the first steps towards sauropod locomotion. Proceedings of the Royal Society of London B, 270: 1753- 1758. [ Links ]
302. Yates, A.M. and Vasconcelos, C.C. 2005. Furculalike clavicles in the prosauropod dinosaur Massospondylus. Journal of Vertebrate Paleontology 25(2): 466-468. [ Links ]
303. Yates, A.M., Bonnan. M. and Neveling, J. 2007. A new diverse dinosaur assemblage from the Early Jurassic of South Africa. Journal of Vertebrate Paleontology 27 (Suppl. to 3): 169A. [ Links ]
304. Yates, A.M., Hancox, P.J. and Rubidge, B.S. 2004. First record of a sauropod dinosaur from the upper Elliot Formation (Early Jurassic) of South Africa. South African Journal of Science 100: 504-506. [ Links ]
305. Young, C.-C. 1941. Gyposaurus sinensis Young (sp. nov.), a new prosauropod from the Upper Triassic beds at Lufeng, Yunnan. Bulletin of the Geological Society of China 21 (2-4): 207- 233. [ Links ]
306. Young, C.C. 1951. The Lufeng saurischian fauna in China. Palaeontologia Sinica (Series C), 13: 1-96. [ Links ]
307. Zhang, Y. and Yang, Z. 1995. A complete osteology of Prosauropoda in Lufeng Basin, Yunnan, China. Jingshanosaurus. Yunnan Science and Technology Publishing House, 100 p., Kunming. [ Links ]
308. Zhao, X.-J., Cheng, Z.-W. and Xu, X. 1999. The earliest ceratopsian from the Tuchengzi Formation of Liaoning, China. Journal of Vertebrate Paleontology 19(4): 681-691. [ Links ]
Recibido: 14 de marzo, 2008.
Aceptado: 30 de junio, 2008.

