










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de la Asociación Geológica Argentina
versión impresa ISSN 0004-4822versión On-line ISSN 1851-8249
Rev. Asoc. Geol. Argent. v.63 n.4 Buenos Aires oct./dic. 2008
El Jurásico de la Argentina y sus amonites
Alberto C. Riccardi
CONICET, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata, provincia de Buenos Aires.
Email: riccardi@museo.fcnym.unlp.edu.ar.
RESUMEN
El estudio de los amonites se halla íntimamente ligado al avance del conocimiento del Jurásico de la Argentina. Ello ha permitido, entre 1870 y la actualidad, documentar la existencia en facies marinas de todos los pisos estratigráficos del Jurásico - excepto el Kimmeridgiano-. Como consecuencia ha sido posible definir diferencias paleobiogeográficas con otras regiones y establecer un marco cronoestratigráfico de referencia regional para el lapso Hettangiano - Tithoniano integrado por una sucesión de 45 unidades bioestratigráficas. Ello ha posibilitado a su vez definir la existencia y alcance de numerosas discontinuidades estratigráficas, establecer relaciones espacio-temporales entre unidades litoestratigráficas y limitadas por discontinuidades, determinar ciclos transgresivo-regresivos y cambios ambientales asociados, y ubicar en un contexto global, tanto zonaciones basadas en otros organismos fósiles como ciclos sedimentarios originados en la mecánica orbital del planeta, registros de inversiones en el campo magnético terrestre, desplazamientos de la curva de deriva polar aparente de América del Sur y modificaciones temporales en relaciones isotópicas. La vinculación global del esquema bioestratigráfico establecido con las faunas de amonites a nivel regional y el anclaje directo que el mismo da a edades absolutas incorporadas a la escala geocronológica internacional, resulta de importancia en la datación de unidades y eventos no-marinos, e.g. continentales, ígneos y metamórficos atribuibles al Jurásico.
Palabras clave: Amonites; Jurásico; Bioestratigrafía; Cronoestratigrafía.
ABSTRACT: The Jurassic of Argentina and its ammonites. Ammonites have been fundamental for improving the knowledge of the Argentinean Jurassic. Since 1870 they have been fundamental to prove the existence, in marine facies, of all Jurassic stages, except the Kimmeridgian. Paleobiogeographic distinction provided the basis to establish a regional biostratigraphic framework of 45 biostratigraphic units for the Hettangian - Tithonian, which has helped to define, existence and scope of a number of stratigraphic discontinuities, space and time relationships among lithostratigraphic and dicontinuity-bounded units, and transgressive-regressive cycles and related environmental changes, and to locate within the global time scale, biozones based on other fossil groups as well as astronomically controlled sedimentary cycles, paleomagnetic reversals, changes in the apparent polar wander path of South America, and isotopic excursions. Anchorage of radimetric ages to regional ammonite biostratigraphy and linkage to the global time scale, provide a basis for dating Jurassic non-marine events, e.g. continental, igneous, metamophic.
Keywords: Ammonites; Jurassic; Biostratigraphy; Chronostratigraphy.
INTRODUCCIÓN
En la Argentina, el Jurásico se halla ampliamente representado (véase Fig. 1) por una variedad de facies marinas y continentales (Riccardi 1983, 1993, Riccardi et al. 1992, Vicente 2005, 2006).

Figura 1: Distribución de rocas del Jurásico en la Argentina (modificado de Stipanicic 1996).
Al sur de los 39° S, el Jurásico está usualmente formado por rocas volcánicas y, en menor proporción, por sedimentitas continentales y marinas. Las facies marinas corresponden al Pliensbachiano-Toarciano Inferior y al Jurásico más alto (Tithoniano), expuestas en el centro y sur de la Patagonia, respectivamente.
Entre los 39° S y los 31° S, el Jurásico, mayormente marino, se halla representado a lo largo de una faja orientada en sentido norte-sur, aproximadamente coincidente con el límite entre la Argentina y Chile, que más al norte se restringe al territorio chileno. Al oeste de esta faja, el Jurásico se halla también expuesto en la Cordillera de la Costa de Chile, conformando una espesa secuencia volcánica en la cual existen algunas intercalaciones de rocas sedimentarias marinas. Facies sedimentarias y volcánicas continentales del Jurásico Superior también se hallan en el nordeste de la Argentina, en la cuenca Chaco-Paranense, que se extienden al Uruguay, Paraguay y sur del Brasil, donde forman parte de las Formaciones Botucatú y Serra Geral (Riccardi 1988).
El Jurásico marino de la Argentina generalmente se dispone en discordancia sobre, sedimentitas marinas del Triásico Superior (centro-oeste de la Argentina) y volcanitas del Jurásico Superior (Patagonia austral) o Paleozoico Superior (Patagonia central). Solamente en la primera región mencionada, en el río Atuel, provincia de Mendoza (Fig. 2), se ha reconocido la existencia de una transición entre rocas marinas del Triásico y Jurásico (véase Riccardi et al. 2004). En otras áreas de la misma región, el Jurásico marino comienza con niveles más modernos, usualmente del Pliensbachiano, y se dispone discordantemente sobre rocas continentales y volcánicas del Triásico, o más antiguas.

Figura 2: Distribución de las principales unidades litoestratigráficas del Jurásico y Cretácico Inferior en la cuenca Neuquina.
De acuerdo con lo expuesto, la sucesión mas completa y representativa del Jurásico está expuesta en el centro-oeste de la Argentina, i.e. en la cuenca Aconcagüo- Neuquina o, abreviando, la cuenca Neuquina (Figs 1-2), donde todos los pisos, desde el Hettangiano al Oxfordiano, están representados en facies marinas clásticas, con carbonatos y evaporitas subordinados. Por encima se disponen rocas volcánicas y sedimentarias continentales del Kimmeridgiano y niveles marinos del Tithoniano, que pasan transicionalmente al Cretácico Inferior. Las diferentes unidades litoestratigráficas reconocidas (Fig. 3) han sido referidas a los Grupos pre- Cuyo y Cuyo (Retiano-Calloviano), Lotena (Calloviano Medio-Oxfordiano), y Mendoza (Kimmeridgiano-Barremiano) (Figs 2-3).

Figura 3: Relaciones cronoestratigráficas entre las principales unidades litoestratigráficas (grupos y formaciones) del Jurásico del centro-oeste de la Argentina.
Los amonites presentes en esta sucesión han sido objeto de numerosos estudios, especialmente en los últimos 35 años y, como resultado, se ha obtenido un esquema bioestratigráfico detallado, que puede ser correlacionado con bastante precisión con el estándar europeo (véase Riccardi, en Riccardi et al. 1990a-c, 2000, Hillebrandt et al. 1992). El mismo es aplicable a unidades marinas expuestas en Patagonia central y austral, aunque allí el registro de la fauna de amonites es relativamente pobre.
Esta zonación bioestratigráfica ha sido usada (Riccardi 1984, 1991b, Westermann y Riccardi 1985, Riccardi y Gulisano 1992, Leanza 1996, Riccardi, en Riccardi et al. 2004) para determinar la existencia de todos los pisos del Jurásico y delimitar los límites inferior y superior de este sistema, para datar y correlacionar unidades litoestratigráficas, secuencias o unidades depositacionales genéticas, unidades limitadas por discontinuidades y, finalmente, para reconstruir la historia del relleno marino de la cuenca Neuquina (véase Legarreta y Gulisano 1989, Legarreta y Uliana 1996, 1999, Vicente 2005, 2006). También ha servido de referencia a zonaciones hechas con otros grupos fósiles (cf. Riccardi et al. 1990a, 1990b, 1990c). Adicionalmente, la distribución geográfica y estratigráfica de los amonites ha provisto la base para otros estudios paleontológicos y geológicos. Así se han documentado casos de gradualismo filético, equilibrio puntuado y de procesos heterocrónicos en el Jurásico Medio (Riccardi 1985, 1996, 2000), ha sido posible construir (Iglesia Llanos y Riccardi 2000) una sección magnetoestratigráfica compuesta para el Jurásico Inferior, proponer una estratigrafía isotópica del Pliensbachiano (Valencio et al. 2005), establecer la evolución tectonosedimentaria del registro inicial de la transgresión marina jurásica en el oeste de la Argentina (Riccardi et al. 1988a, Lanés 2005) y datar evidencias de un acontecimiento oceánico anóxico global (cf. Riccardi 2005).
Una serie de estudios sobre la sistemática y bioestratigrafía de los amonites andinos, mayormente publicada en los últimos 15 años, ha servido de base a la presente síntesis, en la que se presenta el esquema bioestratigráfíco actualizado de los amonites del Jurásico del centro-oeste de la Argentina y se lo vincula con la evolución paleogeográfica de la región.
BIOESTRATIGRAFÍA DE AMONITES
Se reconocen 45 unidades bioestratigráficas dentro del Jurásico del centro-oeste de la Argentina, 22 en el Jurásico Inferior, 14 en el Jurásico Medio y 9 en el Jurásico Superior (Oxfordiano y Tithoniano). Un análisis de las mismas, sobre la base de la representación de géneros - categoría con un nivel adecuado de estabilidad taxonómica y resolución estratigráfica- muestra que el rango y la longevidad genérica media (total = 137), analizada en términos de las zonas andinas, es de 1,4.
Si se usan los valores numéricos de edades incluidos en las escalas estratigráficas publicadas por Remane (2000) y Ogg (2004), la duración promedio de una zona del Jurásico Andino da longevidades genéricas de 0,8 a 1,3 Ma. Cuando se discrimina el Jurásico Inferior, Medio y Superior (este último restringido al Oxfordiano y Tithoniano), las longevidades genéricas medias son, respectivamente, de 0,6-1,3, 1,0-1,5 y 1,2 Ma. Los valores para los diferentes pisos varían entre valores mínimos de 0,5 (Hettangiano), 0,7-1,5 (Calloviano), y 0,8-1,0 (Tithoniano) a 3,5- 4,5 (Sinemuriano) y 2,0-2,7 (Oxfordiano), como es de esperar, considerando que en los dos últimos casos los amonites están, en comparación, pobremente preservados y/o estudiados.
Al margen de lo expuesto, la exactitud de la correlación de la zonación local con la estándar de Europa varía también, a través de los diferentes pisos, debido a que está estrechamente relacionada a cambios biogeográficos controlados por modificaciones en el nivel del mar y en las rutas migratorias entre el "Tethys" y el Pacífico oriental (véase Westermann y Riccardi 1985, Riccardi 1991a).

Figura 4: Zonación de amonites del Jurásico Inferior del centro-oeste de la Argentina (duración de los pisos tomada de Remane 2000).
Figura 5:
Amonites del Jurásico Inferior: Hettangiano: 1, Psiloceras cf. crassicostatum (Guex) (MLP 22247), arroyo Malo, 2, Storthoceras cf. frigga (Wahner) (MLP 22251), arroyo Malo, 3, Schlotehimia? cf. cachinensis Hill. (MLP 22250), arroyo Malo, 4-5, Badouxia canadensis (Freb.) (MLP 22255), arroyo Malo, Sinemuriano: 6-7, cf. Paracaloceras varaense Hill. (MLP 22259), arroyo Malo, 8-9, Coroniceras (C.) cf. alcinoe (Rey.) (MLP 22264), arroyo Malo, 10-11, Orthechioceras cf. incaguasiense Hill. (MLP 24486), Las Chilcas, Pliensbachiano: 12-13, Miltoceras chilcaense Hill. (TUB 790212/2/2), Las Chilcas, 14-15, Meridiceras cf. externum Hill. (TUB 790213/3/4), cerro La Brea, 16-17, Eoamaltheus parameridianus Hill. (TUB 790209/6/1), Puesto Araya, 18-19, Austromorphites behrendseni (Jaw.) (GPIUB, MLP 11004), 20, Fanninoceras cf. carlottense McLearn (MLP 11076), Rajapalo, 21, Canavaria (Tauromeniceras) sp. (NHMB J 21661a, MLP 18161), Toarciano: 22, Hildaites cf. murleyi (Moxon) (MLP 15458), Rajapalo, 23, Peronoceras pacificum Hill. (CPBA 17529) La Ramada, 24-25, Collina chilensis Hill. (CPBA 17459), La Ramada, 26-27, Phymatoceras cf. pseudoerbaense (Gabilly) (CPBA 16514), La Ramada, 28, Phlyseogrammoceras? tenuicostatum (Jaw.) (TUB 790222/2), arroyo Negro, 29-30, Sphaerocoeloceras brocchiiforme Jaworski (GPIUB, MLP 17037), arroyo Negro, 31, Pleydellia cf. lotharingica (Branco) (TUB 680109/10), río de los Patos. Repositorios: MLP, Museo de La Plata, CPBA, Universidad de Buenos Aires, GPIUB, Instituo Geológico y Paleontológico de la Universidad de Bonn, NHMB, Museo de Historia Natural de Basilea, TUB, Universidad Técnica de Berlin. Figs. 12-17, 21, 28, 31 modificadas de Hillebrandt, 1987, 2006, 23-27, modificadas de Álvarez, 1996. Todas las figuras X 0,5, excepto Figs. 1-3, 10-13, 18-21, 29-30, X 1.
Asociaciones fósiles del Hettangiano y Sinemuriano solamente se hallan en elárea del río Atuel, sur de la provincia de Mendoza (formaciones Arroyo Malo y Puesto Araya/El Cholo). Asociaciones del Pliensbachiano y Toarciano están representadas a lo largo del centro-oeste de la Argentina desde 30º a 40º S, mayormente en las provincias de Mendoza y Neuquén (formaciones Puesto Araya y Los Molles y equivalentes). Fuera de la cuenca Neuquina se han registrado amonites del Plienbachiano Tardío y Toarciano Temprano en Patagonia central.
Hettangiano
La existencia de amonites del Retiano y Hettangiano en la Argentina ha sido documentada (Riccardi, en Riccardi et al. 1988a, 1991b, 2004, Riccardi e Iglesia Llanos 1999) en las formaciones Arroyo Malo y Puesto Araya, en la región del río Atuel. Tanto los amonites como la bioestratigrafía de la misma región han sido también consideradas por Hillebrandt (2000), en un estudio monográfico dedicado a los amonites de América del Sur.
1) Zona de Asociación de Psiloceras rectocostatum. Definida originalmente para el norte de Chile (Hillebrandt 1988, 2000), ha sido reconocida en el área del río Atuel, donde se han hallado ejemplares comparables a la especie nominal (véase Riccardi, en Riccardi e Iglesia Llanos 1999 y Riccardi et al. 2000, 2004). Hettangiano temprano, parte superior de la Subzona de Plicatulus e inferior de la de Johnstoni de la Zona de Planorbis. En la Argentina amonites equivalentes a la Subzona de Planorbis no han sido documentados, aunque en la región del río Atuel podrían hallarse en un tramo aparentemente no fosilífero de c. 80 m ubicado entre niveles con Choristoceras cf. marshi Hauer (Retiano) y P. cf. rectocostatum Hill. Resulta significativo en tal sentido el hallazgo de material suelto, proveniente de los niveles en los que se encontró C. cf. marshi, que resulta comparable con Psiloceras erugatum Buck. y P. pressum Hill., especies indicativas de la parte inferior y media de la Zona de Planorbis (véase Riccardi, en Riccardi et al. 2004).
2) Zona de Asociación de Kammerkarites bayoensis. Definida para el norte de Chile (Hillebrandt 2000), en la Argentina parece hallarse representada en la región del río Atuel (Riccardi, en Riccardi et al. 1988a, 1991b, como "Zona de Asociación de Psiloceras"), con K. cf. bayoensis Hill., K. cf. chinchillaensis Hill. (MLP 22245), P. cf. crassicostatum (Guex) (ML P 22247) (Fig. 5.1), P. sp. (MLP 22248), y "Alsatites cf. liasicus (d`Orb.)". Hettangiano Medio, Zona de Liasicus temprano.
3) Zona de Asociación de Discamphiceras reissi. Definida para el norte de Chile (Hillebrandt 2000). En la Argentina algunas de las especies incluidas parecen estar presentes en la región del río Atuel (Riccardi, en Riccardi et al. 1988a, 1991b, parte inferior de la "Zona de Asociación de Waehneroceras-Schlotheimia"), e.g. Kammerkarites cf. diploptychoides Guex (ML P 22249) y Storthoceras cf. frigga (Wahner) (MLP 22251) (Fig. 5.2). Hettangiano Medio, Zona de Liasicus tardío.
4) Zona de Asociación de Sunrisites peruvianus. Zona definida para el norte de Chile (Hillebrandt 2000). Hasta la fecha solo una especie de las citadas para esta zona parece estar representada en la región del Río Atuel, Argentina (Riccardi, en Riccardi et al. 1988a, 1991b, como parte superior de la "Zona de Asociación de Waehneroceras-Schlotheimia"), i.e. Schlotheimia? cf. cachinensis Hillebrandt (MLP 22250, 22252) (Fig. 5.3), que se halla asociada a Paracaloceras cf. coregonense (Sow.). Hettangiano Medio a Tardío, zonas de Liasicus tardío y Angulata temprano.
5) Zona de Asociación de Badouxia canadensis. Originalmente introducida para el Hettangiano Superior de los Lagos Taseko, Canadá, está representada en la región del río Atuel (Riccardi, en Riccardi et al. 1988a, 1991b). En el norte de Chile (Hillebrandt 2000) ha sido dividida en dos subzonas, desde abajo, las subzonas de Schlotheimia cuevitensis y Paracaloceras varaense, que difieren por la presencia-ausencia de S. cuevitensis. En la Argentina, la asociación de B. canadensis incluye, además de la especie nominal (Figs. 5.4-5), Paracaloceras cf. rursicostatum Frebold, Angulaticeras cf. marmoreum (Oppel), y Paracaloceras sp. (Hettangiano ?a Sinemuriano), Zona(s) de Angulata tardío (?a Bucklandi temprano).
6) "Zona de Asociación de Vermiceras". Introducida por Riccardi (en Riccardi et al. 1988a, 1991b) sobre la base de "Vermiceras cf. gracile (Spath) y Vermiceras spp.", y considerada por Hillebrandt (2000) como equivalente a la parte superior de la Subzona de P. varaense, Zona de Canadensis (véase más arriba). Según Hillebrandt (2000) el material comparado con V. gracile (véase Figs. 5.6-7) podría ser referido a Paracaloceras varaense Hill. (2000, p. 79), pero esto implicaría que esta especie se extiende mucho mas arriba del último registro de B. canadensis, Paracaloceras cf. rursicostatum y Angulaticeras cf. marmoreum. Consecuentemente, no se puede descartar reconocer dos biozonas sobre la base del rango de este (y otros?) Arietitidae. La resolución de este tema debe esperar la finalización del estudio sistemático, actualmente en ejecución, de la fauna de Arietitidae presente en la sección del arroyo Malo. (Hettangiano? a) Sinemuriano, Zona(s) de (?Angulata tardío a) Bucklandi temprano.
Sinemuriano
En la Argentina, los amonites del Sinemuriano están restringidos a la región del Río Atuel, y fueron ilustrados por Riccardi (en Riccardi et al. 1988a, 1991b) y Hillebrandt (1990). Material del Sinemuriano Superior de la sección del arroyo Las Chilcas fue descripto e ilustrado por Hillebrandt (2002) en un estudio monográfico dedicado a los amonites del Sinemuriano Tardío de América del Sur. Sobre esta base Hillebrandt (2002) reconoció para América del Sur once horizontes, cuatro equivalentes a la Zona Estándar de Obtusum, dos a la Zona de Oxynotum, y cinco a la Zona de Raricostatum. Hasta la fecha únicamente tres de los últimos cinco han sido reconocidos en la Argentina (véase más abajo).
7) Zona de Asociación de Coroniceras-Arnioceras. Nombre propuesto en reemplazo de "Zona de Asociación de Agassiceras" de Riccardi (en Riccardi et al. 1988a, 1991b, 2000). Zona bien representada en la sección del arroyo Malo, con Coroniceras (C.) cf. alcinoe (Rey.) (Figs. 5.8-9), Coroniceras spp. y Pseudaetomoceras? cf. arcuatum Hill. (MLP 22262). Material pobremente representado, referible en parte a Arnioceras spp. se halla presente inmediatamente al este, en la sección del arroyo El Pedrero. Sinemuriano Temprano, zonas de Bucklandi y Semicostatum (?a Obtusum). Pseudaetomoceras arcuatum ha sido usado por Hillebrandt (2000) para reconocer un horizonte faunístico en el límite Hettangiano -Sinemuriano, por encima de la Zona de B. canadensis y subyaciendo a la sucesión caracterizada por Arietitidae de la Zona de Bucklandi. Material comparable a esta especie aparece en la sección del arroyo Malo en niveles ubicados muy por encima de la última Badouxia, a continuación de horizontes que contienen material comparable, en parte, con P. varaense (véase más arriba en "Zona de Asociación de Vermiceras").
8) Fáunula de Orthechioceras-Paltechioceras. Nombre propuesto como reemplazo de la "Fáunula de Epophioceras" de Riccardi (véase Riccardi 1984 y Riccardi et al. 1988a, 1991b, 2000), arroyo Las Chilcas, región del río Atuel. Es equivalente a tres horizontes definidos por Hillebrandt (2002), i.e., desde abajo: Horizonte con Orthechioceras incaguasiense y Plesechioceras domeykoense, Horizonte con Paltechioceras cf. romanicum, y Horizonte con Paltechioceras oosteri y P. cf. tardecrescens. Esta fáunula esta caracterizada por Orthechioceras incaguasiense Hillebrandt (MLP 22263) (Figs. 5.10-11), P. domeykoense Hill., P. cf. romanicum (Uhlig), P. oosteri (Dumortier) y P. cf. tardecrescens (Hauer). Sinemuriano Tardío, Zona de Raricostatum.
Pliensbachiano
Amonites del Pliensbachiano se encuentran ampliamente distribuidos en el centro- oeste de la Argentina (véase Riccardi et al. 1991b). Material nuevo del Pliensbachiano de la Argentina, especialmente de la zona del río Atuel, fue descripto e ilustrado por Hillebrandt (2006) en un estudio monográfico dedicado a los amonites del Pliensbachiano de América del Sur. Las zonas definidas son, desde abajo:
9) Zona de Asociación de Miltoceras chilcaense. Introducida por Hillebrandt (2006) para una asociación presente en el área del río Atuel y típicamente representada en la sección del arroyo Las Chilcas. Equivalente a la "Fáunula de Miltoceras" de Riccardi (véase Riccardi 1984 y Riccardi et al. 1991b, 2000) con M. chilcaense Hill. (Figs. 5.12-13), M. reductispinosum Hill., M.? sp. A, Tropidoceras? hamulatum Hill., T. cf./aff. demonense (Gemm.). Pliensbachiano Temprano, Zona de Jamesoni temprano. A la base de esta zona en la margen norte del río Atuel corresponde el estratotipo regional del Pliensbachiano (cf. Westermann y Riccardi 1989).
10) Zona de Asociación de Meridiceras externum. Introducida por Hillebrandt (2006) para una asociación típicamente representada en la sección de Puesto Araya, río Atuel, Argentina. Equivalente a la parte inferior de la "Zona de Asociación de Dubariceras" de Riccardi (en Riccardi et al. 1990a) o "Zona de Asociación de Tropidoceras" de Hillebrandt (1987) y Riccardi (en Riccardi et al. 2000). Contiene M. cf. externum Hill. (Figs 5.14-15), M. celericrescens Hill., M. amolanasense Hill., M. densicostatum Hill., Tropidoceras cf. demonense (Gemm.), T. flandrini cf. flandrini (Dum.), T. f. cf. obtusum (Futt.), T. cf. mediterraneum (Gemm.), T. cf./ex gr. zitteli Fuc., T. atuelense Hill., Andicoeloceras sp. A, Dubariceras argentinicum Hill., Liparoceras (L.) cf. cheltiense (Murch.), Phylloceras hebertinum (Rey.), Partschiceras striatocostatum (Meneg.), Juraphyllites libertus australis Hill., J. sp. A. Pliensbachiano Temprano, zonas de Jamesoni tardío e Ibex temprano.
11) Zona de Asociación de Eoamaltheus meridianus. Introducida por Hillebrandt (1987, 2006) para el norte de Chile y para la región del río Atuel, Argentina. Equivalente a la parte superior de la "Zona de Asociación de Dubariceras" de Riccardi (en Riccardi et al. 1990a, 2000), con E. meridianus Hill., E. parameridianus Hill. (Figs. 5.16-17), E. paraovatus Hill., E. multicostatus Hill., E. ovatoides Hill., E. intermissus Hill., E. blancoensis Hill., E. arayaensis Hill., Calliphylloceras cf. bicicolae (Meneg.), Juraphyllites cf. planispira (Reynes). Pliensbachiano Temprano, zonas de Ibex tardío y Davoei temprano.
12) Zona de Asociación de Austromorphites behrendseni. Introducida por Hillebrandt (1987, 2006) y ampliamente representada en el centro-oeste de la Argentina. Es equivalente a la parte inferior de la "Zona de Asociación de Fanninoceras" de Riccardi (1984 y Riccardi et al. 1990a, 2000), con A. behrendseni (Jaworski, sinónimo de A. spinatus Leanza y Blasco 1990) (Figs. 5.18-19), A. volkheimeri (Hill.), A. multiformis (Hill.), A. prorsiflexus (Hill.), A. carinatus (Hill.), A. piutrinensis (Hill.), Zetoceras cf. indicarense (Haas), Fanninoceras sp. A. Liparoceras (Becheiceras) cf. bechei (J. Sow.). Parte tardía del Pliensbachiano Temprano, Zona de Davoei.
13) Zona de Asociación de Fanninoceras fannini. Introducida por Hillebrandt (1987, 2006) para el norte de Chile. En el centro-oeste de la Argentina es equivalente a la parte media de la "Zona de Asociación de Fanninoceras" de Riccardi (1984, Riccardi et al. 1990a, 2000), con F. fannini McLearn, F. leptodiscus (Behr.), F. cf. latum McLearn, F. carlottense McLearn (Fig. 5.20). Parte final del Pliensbachiano Temprano y temprana del Pliensbachiano Tardío, Zona de Davoei tardío y Margaritatus temprano.
14) Zona de Asociación de Fanninoceras disciforme. Introducida por Hillebrandt (1987, 2006) para el norte de Chile. Es equivalente a la parte superior de la "Zona de Asociación de Fanninoceras" de Riccardi (1984, Riccardi et al. 1990a, 2000), en el centro-oeste de la Argentina incluye F. disciforme Hill., Canavaria (C.) cf. finitima (Fucini), C. (C.) cf. naxensis (Gemm.), C. (C.) cf. pulcherrima (Fuc.), C. (Tauromeniceras) sp. (Fig. 5.21), Neolioceratoides sp., Arieticeras sp., Protogrammoceras sp. También se halla representada en Chubut. Pliensbachiano Tardío, zonas de Margaritatus tardío y Spinatum.
Toarciano
Aunque han sido poco estudiados, los amonites toarcianos se hallan ampliamente distribuidos en el centro-oeste de la Argentina (véase Riccardi et al. 1990a). Las zonas definidas incluyen, desde abajo:
15) Zona Estándar de Tenuicostatum. Esta Zona Estándar de Europa fue reconocida en los Andes por Hillebrandt y Schmidt-Effing (1981). En la Argentina parece estar escasamente representada, aunque existen evidencias de su presencia en los arroyos Lapa y Ñiraico (Neuquén) y en Chubut, con Dactylioceras (Eodactylites) sp., D. (Orthodactylites) sp. Toarciano Temprano.
16) Zona de Asociación de Dactylioceras hoelderi. Introducida por Hillebrandt y Schmidt-Effing (1981) para Chile, y representada también en la Argentina, desde el sur de San Juan hasta Catan Lil (Neuquén) y en Pampa de Agnia (Chubut) (véase Riccardi 1984, Hillebrandt 1987, Riccardi et al. 1990a, 2000, Hillebrandt et al. 1992), con D. (Orthodactylites) hoelderi Hill. y Schm.-Eff., Nodicoeloceras sp., Harpoceratoides cf. alternatus (Simps.), Hildaites cf. murleyi (Moxon) (Fig. 5.22). Toarciano Temprano, Zona de Serpentinum.
17-19) Zonas de Asociación de Peronoceras largaense, P. pacificum y Collina chilensis. Introducidas por Hillebrandt y Schmidt-Effing (1981) para Chile, se hallan representadas en el centro-oeste de la Argentina (véase Hillebrandt 1987, Riccardi, en Riccardi et al. 1990a, 2000, Álvarez 1996) con Peronoceras cf. subarmatum (Young y Bird), P. cf. vortex (Simps.), P. pacificum Hill. y Schm.-Eff. (Fig. 5.23), Harpoceras cf. subexaratum Bon., H. subplanatum (Oppel), Frechiella cf. helvetica Renz, Collina chilensis Hill. y Schm.-Eff. (Figs. 5. 24-25), Maconiceras sp., Polyplectus sp. Toarciano Temprano tardío-Tardío temprano, zonas de Bifrons y Variabilis temprano.
20) Zona de Asociación de Phymatoceras. Equivalente a las zonas de P. toroense, y P. copiapense de Hillebrandt (1987). En la Argentina ha sido documentada desde el sur de San Juan al arroyo La Jardinera (Neuquén) (Riccardi 1984, Riccardi, en Riccardi et al. 1990a, 2000) con P. copiapense (Moricke), P. ex gr. lilli (Hauer), P. ex gr. erbaense (Hauer), P. cf. pseudoerbaense (Gab.) (Figs. 5.26-27). Toarciano Tardío, zonas de Variabilis y Thouarsense.
21) Zona de Asociación de Phlyseogrammoceras tenuicostatum. Bien representada en la Argentina, donde se conoce desde el sur de San Juan al arroyo Lapa (Neu-quén) (Riccardi 1984, Riccardi, en Riccardi et al. 1990, 2000, Álvarez 1996), con P. tenuicostatum (Jaw.) (Fig. 5.28), "Witchellia" obscurecostata Jaw., Hammatoceras insigne (Schubler), Sphaerocoeloceras cf. brochiiforme Jaw. Toarciano Tardío, Zona de Dispansum.
22) Zona de Asociación de Dumortieria. Equivalente a las zonas de Pleydellia lotharingica y P. fluitans de Hillebrandt (1987). En la Argentina se conoce desde Paso del Espinacito (San Juan) al sur del Neuquén (Riccardi 1984, 2000, Riccardi, en Riccardi et al. 1990a, 2000) con D. pusilla Jaw., Sphaerocoeloceras brochiiforme Jaw. (Figs. 5.29-30), Pleydellia spp. (Fig. 5.31), Hammatoceras sp. Toarciano Tardío, zonas de Pseudoradiosa tardío y Aalensis.
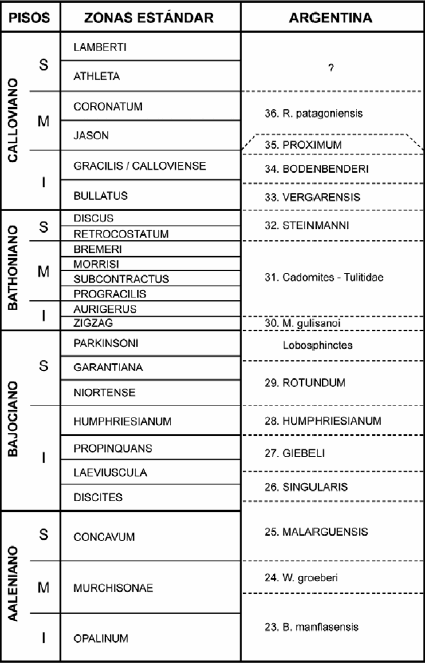
Figura 6: Zonación de amonites del Jurásico Medio del centro-oeste de la Argentina (duración de los pisos tomada de Remane 2000).
Figura 7:
Amonites del Jurásico Medio. Aaleniano: 1, Bredyia cf. manflasensis West. (MLP 24502), arroyo Overas, 2. Westermanniceras groeberi (West. y Ricc.) (MLP 25536), cerro Granito, 3, Puchenquia malarguensis (Burck.) (MLP12228), Picún Leufú, Bajociano: 4, Pseudotoites singularis (Gottsche) (SEGEMAR 349b, MLP 14936), Carro Quebrado, 5-6. Emileia (Chondromileia) giebeli (Gottsche) (MLP 12547), Charahuilla, 7, Stephanoceras (S.) pyritosum (Quenstedt) (MLP 10666), Paso del Espinacito, 8, Megaspaheroceras magnum Ricc. y West. (MLP 23931), Chacay Melehue, Bathoniano: 9-10, Morphoceras gulisanoi Ricc. y West. (MLP 20047), Agua del Ñaco, 11, Cadomites ex gr. rectelobatus (Hauer) (MLP 21166), Chacay Melehue, 12, Lilloettia steinmanni (Spath) (MLP 12703), Chacay Melehue, 13-14, Choffatia gottschei (Steinmann) (MLP 21170), Chacay Melehue, Calloviano: 15-16, Eurycephalites vergarensis (Burck.) (MLP 12658), Chacay Melehue, 17, Neuqueniceras (Frickites) bodenbenderi (MLP 23997), Chacay Melehue, 18-19, Eurycephalites rotundus (Tornquist) (MLP 12694), Chacay Melehue, 20-21, Hecticoceras (H.) proximum Elmi (MLP 12808), Chacay Melehue, 22-23, Rehmannia (R.) brancoi (Steinmann) (MLP 13046), Chacay Melehue, 24-25, Rehmannia (Loczyceras) patagoniensis (Weaver) (MLP 23986), Picún Leufú. Repositorios: MLP, Museo de La Plata, SEGEMAR, Servicio Geológico Minero Argentino. Todas las figuras X 0,5, excepto 2, 3, 22, 23, X 1, 8, X0,3 y 17, X0,4.
Asociaciones de amonites del Jurásico Medio están presentes en el centro oeste de la Argentina, desde el sur de la provincia de San Juan, a los 32º S, en el norte, a las provincias de Mendoza (Formación Tres Esquinas) y Neuquén (formaciones Los Molles y Lotena), en el sur. Las biozonas de amonoideos, previamente sintetizadas por Riccardi (in Riccardi et al. 1994) son actualizadas a continuación.
Aaleniano
Las asociaciones y zonas de amonites del Aaleniano fueron definidas por Westermann y Riccardi (1979) e Hillebrandt y Westermann (1985) (véase también Riccardi 1984, Hillebrandt et al. 1992).
23) Zona Estándar de Bredyia manflasensis. Introducida por Hillebrandt y Westermann (1985), y pobremente representada en las provincias de San Juan y Mendoza (Riccardi 1984, Riccardi et al. 1990b, 2000, Riccardi 2000). Representada por B. manflasensis West. (Fig. 7.1), B. delicata West., Westermanniceras groeberi (West. y Ricc.), ?Leioceras sp. Aaleniano Temprano a Medio, zonas de Opalinum y Murchisonae temprano. La base de esta zona en sierra de Reyes podría servir de base para establecer el estratotipo regional del Aaleniano (cf. Westermann y Riccardi 1989).
24) Zona de Asociación de Westermanniceras groeberi. Introducida por Westermann y Riccardi (1979, Riccardi 1984, Riccardi et al. 1990b, 2000, Riccardi 2000), y representada en las provincias de San Juan, Mendoza y Neuquén con W. groeberi (West. y Ricc.) (Fig. 7.2), Planammatoceras cf. planiforme Buck., Podagrosiceras athleticum Maub. y Lamb., Tmetoceras spp. Aaleniano Medio, Zona de Murchisonae.
25) Zona Estándar de Malarguensis. Introducida por Westermann y Riccardi (1979, Riccardi 1984, Riccardi et al. 1990 b, 2000, Hillebrandt et al. 1992), incluye desde abajo (véase Westermann y Riccardi 1982, Hillebrandt y Westermann 1985), las subzonas de Puchenquia compressa y P. mendozana y el Horizonte con Podagrosiceras maubeugei. Ampliamente distribuida en el centro-oeste de la Argentina, desde el sur de San Juan al sur del Neuquén, con Puchenquia (P.) malarguensis (Burck.) (Fig. 7.3), P. (Gerthiceras) compressa West. y Ricc., P. (G.) mendozana West. y Ricc., Planammatoceras (Pseudaptetoceras) klimakomphalum (Vacek), P. (P.) tricolore West. y Ricc., P. gerthi (Jaw.), Eudmetoceras amplectens (Buckman), Csernyeiceras moerickei (Jaw.), Euhoploceras amosi West. y Ricc., Podagrosiceras athleticum Maub. y Lamb., Praeleptosphinctes jaworskii West. Aaleniano Tardío a Bajociano inicial, zonas de Concavum a Discites temprano.
Bajociano
Asociaciones de amonites del Bajociano fueron definidas por Westermann y Riccardi (1979), Riccardi (1984) y Riccardi et al. (1988b, 1990b, 1991a, véase también Hillebrandt et al. 1992).
26) Zona Estándar de Singularis. Introducida por Westermann y Riccardi (1979) está ampliamente distribuida en el centro-oeste de la Argentina, desde el sur de San Juan al sur del Neuquén. Con Pseudotoites singularis (Gottsche) (Fig. 7.4), P. sphaeroceroides (Tornq.), P. crassus West. y Ricc., P. transatlanticus (Tornq.), P. argentinus Arkell, Sonninia (Euhoploceras) amosi West. y Ricc., S. (Fissilobiceras) zitteli (Gott.), S. (Papilliceras) espinazitensis altecostata Tornq. Bajociano Temprano, zonas de Discites tardío y Laeviuscula temprano. A la base de esta zona en cerro Puchenque, Mendoza, corresponde el estratotipo regional del Bajociano (cf. Westermann y Riccardi 1989).
27) Zona Estándar de Giebeli. Introducida por Westermann y Riccardi (1979) se halla ampliamente documentada en el centro-oeste de la Argentina, con una distribución similar a la zona infrayacente. Incluye desde abajo (véase Westermann y Riccardi 1979, Hillebrandt et al. 1992) las subzonas de E. g. submicrostoma y E. multiformis y el Horizonte con Dorsetensia blancoensis. Con E. (E.) multiformis (Gott.), E. (E.) aff. brochii (Sow.), E. (E.) vagabunda Buck., E. (Chondromileia) giebeli (Gott.) (Figs. 7.5-6), Chondroceras recticostatum West. y Ricc., Stephanoceras (Skirroceras) cf. macrum (Quenst.), Sonninia (Papilliceras) espinazitensis Tornq., Witchellia sp., Dorsetensia mendozai West. y Ricc., D. blancoensis West. y Ricc. Bajociano Temprano, zonas de Laeviuscula tardío y de Propinquans.
28) Cronozona Estándar de Humphrie-sianum. Esta zona estándar europea fue extendida al norte de Chile (Westermann y Riccardi 1979, Riccardi et al. 1990b, 2000), donde ha sido dividida, de abajo hacia arriba, en las subzonas de Dorsetensia romani, de Stephanoceras chilense y de Duashnoceras chilense, está escasamente representada en el centro-oeste de la Argentina (Paso del Espinacito, arroyo Blanco, Chacay Melehue, Picún Leufú), con Stephanoceras (S.) pyritosum (Qu.) (Fig. 7.7), S. (Stemmatoceras) aff. frechi (Renz), Duashnoceras paucicostatum chilense (Hill.), Teloceras crikmayi chacayi West. y Ricc., Dorsetensia romani (Oppel), D. liostraca (Buckman).
29) Cronozona Estándar de Rotundum. Introducida para América del Norte (Hall y Westermann 1980), fue reconocida en los Andes por Westermann y Riccardi (1979, véase también Riccardi 1984, Riccardi y Westermann 1991a, Riccardi et al. 1989, 1990b, 1991a, 2000), pero es rara en el centro-oeste de la Argentina, donde se halla en el sur de San Juan (Paso del Espinacito) y en Chacay Melehue (Neuquén) con Megaspaheroceras magnum Ricc. y West. (Fig. 7.8), Cadomites cf. daubenyi (Gemm.), C. aff. deslongchampsi (d'Orb.), y Leptosphinctes sp. Bajociano Tardío, zonas de Niortense a Garantiana. Capas equivalentes a la Zona de Parkinsoni probablemente se hallan presentes en Chacay Melehue, tal como lo indica la presencia de Lobosphinctes intersertus Buck., una especie típica de esa zona en el Bajociano Superior de Inglaterra, aunque se extiende al Bathoniano Inferior.
Bathoniano
30) Zona de Asociación de Morphoceras gulisanoi. Esta asociación, definida por Riccardi y Westermann (1999) y Riccardi (en Riccardi et al. 2000), esta típicamente presente en sierra de Reyes, provincia de Mendoza, y probablemente se halla presente en Chacay Melehue (Neuquén), por arriba de la Zona de Rotundum. La asociación de M. gulisanoi está caracterizada por M. gulisanoi Ricc. y West. (Figs. 7.9-10), Procerites cf. schloenbachi (de Grossouvre), Oxycerites cf. aspidoides (Rollier). Bathoniano Temprano, Zona de Zigzag. En la base de esta zona en sierra de Reyes o en Chacay Melehue correspondería definir el estratotipo regional del Bathoniano (cf. Westermann y Riccardi 1989).
31) Asociación mixta de Cadomites-Tulitidae. Definida por Riccardi et al. (1988b, 1989, 1990b, 1991a, 1994) para una fauna parcialmente retrabajada hallada en Chacay Melehue, provincia del Neuquén. Incluye Cadomites ex gr. orbignyi (De Gross.), C. bremeri Tsereteli y C. ex gr. rectelobatus (Hauer) (Fig. 7.11), que se presentan con Tulites (Rugiferites?) cf. davaiacensis (Liss.), una especie del Bathoniano Medio, y Bullatimorphites (Kheraiceras) cf. bullatus (Orb.), y Choffatia (Homoeoplanulites) ex gr. aequalis (Roem.), que indican el Bathoniano Superior. Amonites similares fueron documentados en el norte de Chile por Gröschke y Hillebrandt (1994) y ubicados en la Zona de Asociación de Rugiferites sofanus del Bathoniano Medio. Bathoniano Medio a Superior.
32) Zona Estándar de Steinmanni. Definida por Riccardi et al. (1988b, 1989, 1990b, 1991b, 1994) para Chacay Melehue, provincia del Neuquén. La parte inferior está caracterizada por la especie nominal, Lilloettia steinmanni (Spath) (Fig. 7.12) y por Choffatia (Homeoplanulites) ex gr. aequalis (Roem.), Iniskinites crassus, I. gulisanoi, Ricc. y West. spp., y la parte superior, i.e. el Horizonte de Stehnocephalites gerthi, por Stehnocephalites gerthi (Spath), L. australis Ricc. y West. Xenocephalites cf. araucanus (Burck.), Neuqueniceras (N.) steinmanni Stehn, N. (N.) biscissum (Stehn), Oxycerites obsoletoides Ricc. et al., Choffatia jupiter (Stein.), Ch. gottschei (Steinmann) (Figs. 7.13-14). Bathoniano Tardío, zonas de Retrocostatum y Discus. La posible diferenciación (Parent 1998, p. 72) de dos horizontes adicionales, sobre la base de I. gulisanoi e I. crassus, en la parte inferior y media de esta zona, no ha sido adecuadamente fundamentada, especialmente considerando que los rangos estratigráficos de varias especies han sido incorrectamente citados en la propuesta realizada.
Calloviano
Las asociaciones de amonites del Calloviano de los Andes fueron definidas, descriptas e ilustradas por Riccardi et al. (1988b, 1989, 1990b, 1991a, 1994) y Riccardi y Westermann (1991a, b). La edad de la base del Calloviano fue datada en 160,5 ± 0,3 (Odin et al. 1992), a partir de zircones de un nivel de tobas que en Chacay Melehue se halla en la base de la Zona de Vergarensis.
33) Zona Estándar de Vergarensis. Típicamente representada en el noroeste de la provincia del Neuquén, con Eurycephalites vergarensis (Burck.) (Figs. 7.15-16), Neuqueniceras (N.) steinmanni Stehn, Xenocephalites gottschei (Tornq.). Calloviano inicial. La posible inclusión de E. vergarensis y X. gottschei en un par dimórfico (Parent 1997, 1998), y el cambio consecuente en la denominación de esta zona, resulta cuestionable, considerando, que no se han provisto los datos básicos necesarios, que las relaciones entre macro y microconchas no resultan claras, que los rangos de ambas especies son diferentes y que no se ha estudiado material topotípico. A la base de esta zona en Chacay Melehue (nivel 267) corresponde el estratotipo regional del Calloviano (cf. Westermann y Riccardi 1989).
34) Zona Estándar de Bodenbenderi. Definida para Chacay Melehue, provincia del Neuquén, se conoce desde el sur de San Juan a Chacay Melehue, con Neuqueniceras (Frickites) bodenbenderi (Tornq.) (Fig. 7.17), Eurycephalites rotundus (Tornq.) (Figs. 7.18-19), E. extremus (Tornq.), Xenocephalites stipanicici Ricc. et al., X. involutus Ricc. y West. Calloviano Temprano, zonas de Bullatus tardío y Gracilis.
35) Zona Estándar de Proximum. Definida para Chacay Melehue, provincia del Neuquén, con Hecticoceras (H.) proximum Elmi (Figs. 7.20-21), H. (H.) cf. hecticum (Rein.), H. (H.) boginense (Pet.), H. (Chanasia) navense Román, H. (Ch.) ardescicum Elmi, Xenocephalites stipanicici Ricc. et al., X. involutus Ricc. y West., Neuqueniceras (Frickites) cf. antipodum (Gott.), Rehmannia (R.) cf. paucicostata (Tornq.), R. (R.) brancoi (Stein.) (Figs. 7.22-23), R. (R.) stehni (Zeiss), Oxycerites (Paroxycerites) oxynotus (Leanza). Está también representada en el sur de San Juan. Calloviano Temprano, Zona de Gracilis tardío.
36) Horizonte de Rehmannia patagoniensis. Horizonte con abundante R. (Loczyceras) patagoniensis (Weaver) (Figs. 7.24-25), típicamente representado en facies costeras a lo largo del margen este de la cuenca Neuquina, desde el sur del Neuquén al sur de la provincia de San Juan. La presencia de esta especie (Parent et al. 2006b, p. 309, Parent 2006) en Chacay Melehue, supuestamente asociada con otros taxa, no ha sido adecuadamente documentada y se halla en contradicción con detallados estudios bioestratigráficos (Riccardi et al. 1989, Riccardi y Westermann 1991a, b, Riccardi et al. 1994) y probablemente se debe a una errónea interpretación de la especie. Calloviano Medio, zonas de Jason y Coronatum.

Figura 8: Zonación de amonites del Jurásico Superior del centrooeste de la Argentina (duración de los pisos tomada de Remane 2000).
Figura 9:
Amonites del Jurásico Superior. Oxfordiano: 1, Peltoceratoides pressulus (Leanza) (SEGEMAR s/n), Chacay Melehue, 2-3, Araucanites stipanicici West. y Ricc. (MLP 12240), Aguada de Campos, 4, Lithacosphinctes aff. janus (Choffat) (LPB 426), Chacay Melehue, 5, Perisphinctes (Arisphinctes) sp. (YPF 16718), arroyo de la Manga, Tithoniano: 6-7, Pseudinvoluticeras douvillei Spath (MLP 8210), cerro Lotena, 8-9, Choicensisphinctes choicensis (Burck.) (MLP 6342), cerro Lotena, 10-11, Pseudolissoceras zitteli (Burck.) (SEGEMAR 7333/1), 12-13, Pseudhimalayites steinmanni (Haupt), (MLP 900), cerro Lotena, 14-15, Aulacosphinctes proximus (Steuer) (MLP 14629), Mina la Eloisa, 16-17, Aspidoceras andinum Steuer (MLP 3568), río Diamante, 18-19, Corongoceras lotenoense Spath (MLP 3355), río Diamante, 20-21, Windhauseniceras internispinosum (Krantz) (SEGEMAR 8941/1), 22-23, Parodontoceras calistoides (Behrendsen), 24-25, Substeueroceras koeneni (Steuer) (MLP 6768). Repositorios: MLP, Museo de La Plata, SEGEMAR, Servicio Geológico Minero Argentino, YPF, Yacimientos Petrolíferos Argentinos, GPIT, Universidad de Bonn, LPB, Universidad Nacional de Rosario. 1, modificado de Leanza 1947, 4, modificado de Parent et al., 2006b, 5, modificado de Stipanicic, 1951, 10-11, 20-21, modificados de Leanza, 1980. Todas las figuras X 0,5.
Oxfordiano
El Oxfordiano marino se halla bien representado en el centro-oeste de la Argentina, aunque con una fauna relativamente mal conservada (Leanza 1947, Stipanicic 1951, Stipanicic et al. 1976, Riccardi, en Riccardi et al. 1990c, Parent 2006, Parent et al. 2006b), mayormente restringida a niveles del Oxfordiano Medio de la Formación La Manga. Faunas del Calloviano Tardío y Oxfordiano Temprano no se encuentran usualmente representadas debido a la existencia de un hiato regional, o se hallan geográficamente restringidas y están pobremente conservadas. Estas circunstancias han sido claramente puestas en evidencia en un trabajo reciente (Parent 2006), en el que se ha intentado refinar la bioestratigrafía del Calloviano Tardío-Oxfordiano de la región andina, donde la mayor parte de los taxones mencionados están basados en material escaso, fragmentario y pobremente preservado y cuya procedencia exacta, especialmente estratigráfica, no está claramente establecida o se desconoce.
37) Asociación de Peltoceratoides - Parawedekindia. Definida en Riccardi et al. (1990c), en el centro-oeste de la Argentina se halla presente en la cuenca Neuquina, con Peltoceratoides pressulus (Leanza) (Fig. 9.1) y Parawedekindia sp. Esta asociación ha sido denominada "Biozona de P. pressulus" por Parent (2006). Calloviano Tardío - Oxfordiano Temprano, zonas de Lamberti final a Cordatum temprano.
38) Zona de Asociación de Perisphinctes-Araucanites. Esta asociación (Riccardi, en Riccardi et al. 1990c) está ampliamente distribuida a través de la cuenca Neuquina. Incluye Araucanites mulai, A. reyesi, A. stipanicici (Figs. 9.2-3) West. y Ricc. spp., P. (Kranaosphinctes) spp., P. (Arisphinctes) spp. (Fig. 9.5), "Perisphinctes" spp., Euaspidoceras ajax Leanza, Euaspidoceras aff. waageni Spath. Esta asociación es equivalente a las "Biozonas de Subvinalesphinctes pseudokranaus, Passendorferia y Euaspidoceras tarapacaense", mayormente a la primera, propuestas por Parent (2006). Oxfordiano Temprano final a Oxfordiano Medio, zonas de Cordatum tardío a Transversarium (? y Bifurcatum).
39) Horizonte con Lithacosphinctes. Representantes de Lithacosphinctes Oloriz, descriptos por primera vez en la región andina del norte de Chile (Gygi y Hillebrandt 1991), parecen hallarse presentes en la cuenca Neuquina, de acuerdo con la presencia de un ejemplar en Chacay Melehue (Parent et al. 2006b) (Fig. 9.4), que ha sido usado (Parent 2006) para definir una "Biozona de L. desertorum" (= L. desertorus). Oxfordiano Tardío, Zona(s) de Bimammatum (a Planula).
Kimmeridgiano
En el centro-oeste de la Argentina la parte más alta del Oxfordiano y el Kimmeridgiano consisten mayormente de facies continentales. Solo en la cuenca Austral del sur de las Argentina algunos amonites pobremente preservados han sido asignados tentativamente al Kimmeridgiano Tardío (cf. Feruglio 1936-1937, Leanza 1968).
Tithoniano
En el centro-oeste de la Argentina, la base del Grupo Mendoza contiene una rica fauna de amonites del Tithoniano (cf. Leanza, en Riccardi et al. 1990c). Una fauna pobremente conservada pero de la misma edad se halla en Patagonia austral (Feruglio 1936-1937, Leanza 1968, Blasco et al. 1979, Kraemer y Riccardi 1997). Sobre esta base se desarrolló, para el centro- oeste de la Argentina, un detallado esquema de biozonas (véase más abajo). Propuestas realizadas en años recientes (véase Parent y Capello 1999, Parent 2001, 2003, Parent et al. 2006a), en las cuales se han modificando reiteradamente la sistemática y bioestratigrafía, necesitan ser fundamentadas adecuadamente con un estudio taxonómico de alcance regional basado en nuevas colecciones.
40) Zona de Asociación de Virgatosphinctes mendozanus. Zona originalmente propuesta por Burckhardt (1900, 1903, cf. Leanza 1980), caracteriza la base de la Formación Vaca Muerta, inmediatamente por encima de las facies continentales del Kimmeridgiano correspondientes a la Formación Tordillo. Se conoce en una faja que se extiende inmediatamente al sur del cerro Aconcagua, Mendoza, hasta el sur del Neuquén. Con: V. mendozanus (Burck.), V. andesensis (Douv.), V. denseplicatus rotundus Spath, V. evolutus Leanza, V. mexicanus (Burck.), V. burckhardti (Douv.), Pseudinvoluticeras douvillei Spath (Figs. 9.6-7), P. windhauseni (Weav.), P. (?) wilfridi (Douv.), Choicensisphinctes choicensis (Burck.) (Figs. 9.8-9), Ch. erinoides (Burck.), Subplanites malarguensis Spath. La transferencia (Parent 2003) de la última especie citada al género Euvirgalithacoceras Zeiss et al., es cuestionable dado que la especie tipo, E. supremum (Schneid), tal como ha sido ilustrada por Schneid (1915, lám. 4, fig. 4) difiere claramente en tamaño, involución y costulación del lectotipo de la especie andina (cf. Parent et al. 2006a). Tithoniano Inferior tardío a Tithoniano Medio bajo, zonas de Darwini a Semiforme.
41) Zona de Asociación de Pseudolissoceras zitteli. Zona introducida por Burckhardt (1900, 1903, véase Leanza 1980). Parte inferior de la Formación Vaca Muerta, representada desde el sur del río Diamante (Mendoza) al sur del Neuquén. Con: P. zitteli (Burck.) (Figs. 9.10-11), P. pseudoolithicum (Haupt), Glochiceras steueri Leanza, Hildoglochiceras wiedmanni Leanza, Parastreblites comahuensis Leanza, Aspidoceras cieneguitense (Steuer), Pseudhimalayites steinmanni (Steuer), Simocosmoceras adversum andinum Leanza y Oloriz. Tithoniano Medio temprano, zonas de Semiforme a Fallauxi.
42) Zona de Asociación de Aulacosphinctes proximus. Zona propuesta por Burckhardt (1900, 1903, véase Leanza 1980). Representada en la Formación Vaca Muerta, con una extensión geográfica similar a la de la zona subyacente. Caracterizada por: A. proximus (Steuer) (Figs. 9.14-15), Subdichotomoceras sp., Pseudhimalayites steinmanni (Steuer) (Figs. 9.12-13), Aspidoceras andinum (Steuer) (Figs. 9.16-17), A. neuquensis Weaver. La propuesta (Parent 2003) de incluir A. proximus (Steuer) en Torquatisphinctes Spath carece de fundamentación, al igual que otras suposiciones relacionadas, e.g. la supuesta presencia de tubérculos en las vueltas interiores de Aulacosphinctes Uhlig, la semejanza de A. proximus a material de México que Verma y Westermann (1973) compararan con Subdichtomoceras inversum (Spath) y Aulacosphinctoides lauri (Aguilera) y A. potosinus (Aguilera), y que fueran incluidas en Torquatisphinctes por Parent (2003). Tithoniano Medio, zonas de Fallauxi a Ponti.
43) Zona de Asociación de Windhauseniceras internispinosum. Zona introducida por Weaver (1931, véase Leanza 1980). Representada en la Formación Vaca Muerta, desde la región de El Sosneado, Mendoza, hasta el sur de Zapala, Neuquén. Caracterizada por W. internispinosum (Krantz) (Figs. 9.20-21), Wichmanniceras mirum Leanza, Pachysphinctes americanensis Leanza, Hemispiticeras aff. steinmanni (Steuer), Subdichotomoceras araucanense Leanza, S. windhauseni (Weaver), Parapallasiceras aff. pseudocolubrinoides Oloriz, P. aff. recticosta Oloriz, Aulacosphinctoides aff. hundesianus (Uhlig), Aspidoceras euomphalum Steuer, Corongoceras lotenoense Spath (Figs. 9.18-19). En el Miembro Los Catutos, Los Catutos, provincia del Neuquén, Leanza y Zeiss (1990, 1992, 1994) reconocieron, desde abajo, las subzonas de Catutosphinctes rafaeli Leanza y Zeiss y Zapalia fascipartita Leanza y Zeiss, la última de las cuales incluye los Horizontes de Aspidoceras aff. altum Biro y Djurjuriceras catutosense Leanza y Zeiss. Tithoniano Medio tardío, Zona de Ponti.
44) Zona de Asociación de Corongoceras alternans. Zona propuesta por Leanza (1945, véase Leanza 1980). Es conocida en la Formación Vaca Muerta y equivalentes, desde el río Diamante, Mendoza, a la región de Picún Leufú, sur del Neuquén. Caracterizada por C. alternans (Gerth), C. mendozanum (Behr.), C. rigali Leanza, Aulacosphinctes mangaensis (Steuer), Lytohoplites burckhardti Spath, Micracanthoceras tapiai Leanza, M. lamberti Leanza, Berriasella australis Leanza, B. pastorei Leanza, B. krantzi Leanza, B. bardensis Krantz, Chigaroceras gerthi (Krantz). Tithoniano Tardío, zonas de Microcanthum y Durangites spp.
45) Zona de Asociación de Substeueroceras koeneni. Propuesta por Gerth (1925, véase Leanza 1981), tiene la misma extensión geográfica de la zona subyacente y está caracterizada por S. koeneni (Steuer) (Figs. 9.24-25), S. exstans Leanza, Aulacosphinctes azulensis Leanza, A. mangaensis (Steuer), Pectinatites (?) striolatus (Steuer), Berriasella fraudans inflata Leanza, B. inaequicostata Gerth, Parodontoceras calistoides (Behr.) (Figs. 9.22-23), Aspidoceras longaevum Leanza, Blanfordiceras vetustum (Steuer), Himalayites andinus H. Leanza, Spiticeras acutum Gerth. Tradicionalmente ubicada en el Tithoniano Superior (véase Leanza 1981), probablemente llega al Berriasiano Inferior, tal como lo sugiere la presencia en su parte mas alta de Schaireria longaeva Leanza y "Spiticeras" acutum Gerth, un probable antecesor de Groebericeras (cf. Zeiss 1986, Leanza 1996). Tithoniano Tardío - Berriasiano Medio temprano, zonas de Durangites spp. - Jacobi/Grandis y Occitanica inicial.
CUENCA NEUQUINA: FAUNA DE AMONITES, CICLOS SEDIMENTARIOS Y PALEOGEOGRAFÍA
El origen de las rocas mesozoicas de América del Sur está relacionado con la tectónica extensional del Triásico Tardío - Jurásico Temprano que afectó al margen occidental del Gondwana y conformó una serie de fosas separadas, orientadas en sentido NNO, limitadas por fallas (cf. Vergani et al. 1995). En estas depresiones se acumularon rocas volcánicas y volcanoclásticas, conjuntamente con sedimentos continentales y, localmente, marinos. En el mismo lapso en la región del río Atuel se abrió, desde el Pacífico, un corredor marino, a partir del cual en el Sinemuriano - Pliensbachiano, mediante una subsidencia generalizada, se desarrolló una cuenca de retroarco, i.e. la cuenca Neuquina.
Cambios eustáticos del nivel del mar y aporte sedimentario determinaron la formación de secuencias depositacionales (véase Legarreta y Gulisano 1989), limitadas por discontinuidades, evidenciadas en paraconcordancias, asociadas a modificaciones paleogeográficas importantes. Estas secuencias fueron agrupadas (Legarreta y Gulisano 1989) en varias "mesosecuencias", i.e. Cuyo (espesor máximo c. 3000 m, incluyendo las secuencias C1 a C7), Lotena-Chacay (espesor máx. c. 800 m, incluyendo las secuencias L1 a L5 y Mendoza inferior (espesor máx. c. 3800 m, incluyendo las secuencias M1-M6). El conjunto de secuencias comprendido entre el Triásico Superior y el Oxfordiano, delimitado por dos discordancias de primer orden, también fue diferenciado (Riccardi y Gulisano 1992) en varias unidades limitadas por discordancias o discontinuidades.
Existe una clara vinculación entre los pulsos transgresivo-regresivos que determinan las secuencias y la diversidad, densidad y distribución de la fauna y como consecuencia una relación inversa entre éstos y la presencia e importancia de discontinuidades (véase Riccardi 1984, 1991 b, Riccardi y Gulisano 1992, Riccardi et al. 1994). Así, las faunas de amonites se encuentran más ampliamente distribuidas y son más diversas en la parte media y media-superior de las secuencias, mientras que en la proximidad a sus límites presentan restricción geográfica y baja diversidad (Fig. 10).
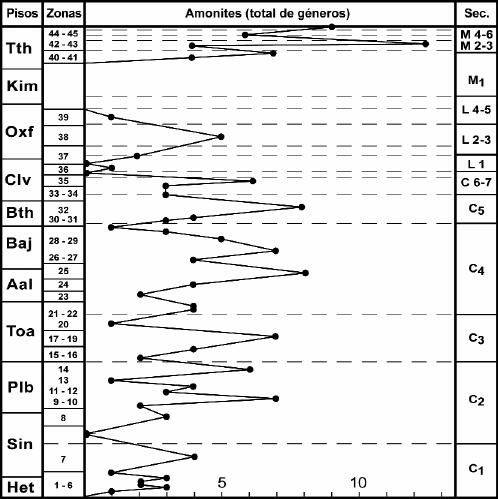
Figura 10: Diversidad de amonites en el Jurásico del centro-oeste de la Argentina.
La primera entrada del mar Jurásico se produjo al norte de 40° S, donde el mar Retiano-Sinemuriano cubrió las áreas costeras de Chile llegando, hacia el este, a la zona de río Atuel (Riccardi et al. 1988a, 2004, Lanés 2005). A consecuencia de este avance del mar se depositaron pelitas y areniscas de plataforma, las cuales se hallan ausentes hacia el sur, donde la depositación marina comenzó en el Pliensbachiano. Una expansión importante del mar del Jurásico Temprano se produjo al final del Pliensbachiano Temprano. Niveles bajos del mar produjeron depósitos turbidíticos (Formación Los Molles) en zonas más profundas y, niveles altos, pelitas de plataforma externa (Formación Los Molles), areniscas litorales (Formación Puesto Araya), y facies continentales (Formación El Freno). Una caída del nivel del mar durante el lapso Pliensbachiano Tardío - Toarciano fue interrumpida por una elevación en el Toarciano Temprano tardío.
Estos cambios concuerdan con los registrados en la diversidad de los amonites (considerada como una medida del número de géneros) (Fig. 10). La diversidad es relativamente baja en el Hettangiano y muestra un incremento en el Sinemuriano Temprano - aunque esta elevación podría ser un artefacto debido a una bioestratigrafía poco definida -. Posteriormente se registra un incremento progresivo de la diversidad desde el Sinemuriano Tardío al Pliensbachiano Temprano (Zona de M. externum, ≈ zonas de Jamesoni tardío a Ibex temprano), alcanzando un máximo, caracterizado por elevado endemismo (40 % o 80 %, según se lo calcule incluyendo o no los taxa pandémicos), en coincidencia con la parte media de la secuencia C2, sigue una pronunciada caída hacia el límite entre el Pliensbachiano Temprano y Tardío (Zona de F. fannini, zonas de Davoei tardío a Margaritatus temprano), interrumpida por una ligera elevación en la Zona de A. behrendseni ≈ Zona de Davoei medio). Posteriormente hay un aumento en la diversidad al final del Pliensbachiano (Zona de F. disciforme, ≈ Zona de Spinatum), seguido por una pronunciada disminución al comienzo del Toarciano (Zona de Tenuicostatum), y un incremento, hasta un máximo, al final del Toarciano Temprano (zonas de P. largaense-C. chilense, ≈ zonas de Serpentinum a Bifrons), en coincidencia con la parte media de la secuencia C3. Posteriormente hay una pronunciada caída en la diversidad - salvo por un breve incremento intermedio -, a través del límite Toarciano-Aaleniano, con reducida diversidad de amonites en el Aaleniano Temprano.
Durante el Jurásico Medio, partes del arco volcánico se volvieron continentales, con desarrollo de volcanismo subaéreo. Hacia el este, la sedimentación marina fue continua en la cuenca de retroarco. Un máximo transgresivo se produjo durante el Bajociano entre 40° y 31° S, cuando una elevación del nivel del mar amplió el área de la plataforma y un alto influjo clástico desde el sur y este produjeron progradación deltaica (Formación Lajas, secuencia C4). La diversidad de los amonites (Fig. 10) muestra un marcado incremento a través del Aaleniano, para alcanzar un máximo en el límite Aaleniano- Bajociano (Zona de Malarguensis, ≈ zonas de Concavum-Discites), seguido por una ligera caída en el límite entre las zonas de Malarguensis y Singularis (≈ Zona de Laeviuscula) y un nuevo incremento en la Zona de Giebeli (≈ Zona de Propinquans). Ambos máximos coinciden con la parte media de la secuencia C4.
Durante el Bathoniano, debido a una importante caída del nivel del mar, se produjo una importante regresión del mar en el centro-oeste de la Argentina, de forma tal que en áreas marginales entre 26° S y 37° S el Calloviano descansa directamente sobre el Bajociano o rocas más antiguas. La diversidad de los amonites muestra una fuerte caída a través del límite Bajociano - Bathoniano, y se incrementa fuertemente hasta alcanzar un máximo en la Zona de Steinmanni del Bathoniano Tardío, en coincidencia con la parte media de la secuencia C5.
La fase regresiva del Bathoniano se extendió al Calloviano Temprano, cuando la cuenca se redujo fuertemente, con depositación de evaporitas (secuencias C 6- 7). La diversidad de los amonites (Fig. 10) muestra una caída a través del límite Bathoniano- Calloviano, seguida por una ligera elevación al final del Calloviano Temprano. Durante el Calloviano Tardío hubo una pronunciada caída del nivel del mar, áreas de la plataforma fueron parcialmente erosionadas y en las zonas más profundas se depositaron areniscas y conglomerados continentales (parte inferior de la Formación Lotena, secuencia L1), aunque una breve elevación del nivel del mar durante el Calloviano Medio-Tardío produjo una renovada transgresión con una marcada expansión del área de sedimentación marina que, sin embargo, no alcanzó la extensión del mar del Bajociano. Así el Horizonte de Rehmannia patagoniensis se dispone transgresivamente sobre facies evaporíticas, deltaicas e incluso continentales de diferentes edades del Jurasico Medio.
Durante el Oxfordiano Temprano, debido a una elevación del nivel del mar, se produjo una sucesión turbidítica en la región andina, seguida por pelitas dispuestas transgresivamente sobre el área de plataforma (secuencia L2). Hacia el final de esta fase se desarrolló una progradación de carbonatos (Formación La Manga, secuencia L3), seguida en el Oxfordiano Tardío por sedimentación evaporítica en las partes centrales de la cuenca (Formación Auquilco, secuencias L 4-5). La diversidad de los amonites muestra un incremento aproximadamente uniforme desde el comienzo del Oxfordiano hasta alcanzar un máximo en la Zona de Perisphinctes-Araucanites (≈ zonas de Plicatilis y Tansversarium), seguido por una pronunciada disminución de los valores.
La somerización progresiva de la cuenca del centro-oeste de la Argentina durante el Calloviano-Oxfordiano se originó en una caída general del nivel del mar relacionado a la pronunciada elevación del arco volcánico de la Cordillera de la Costa de Chile, que actuó como fuente de material clástico y volcánico. La depositación resultante desde el oeste y el desplazamiento del eje de la cuenca hacia el este llegó a su clímax durante el Kimmeridgiano, cuando el volcanismo alcanzó la Cordillera Principal. Las zonas marginales estuvieron sujetas a fuerte erosión, al tiempo que en otras partes de la cuenca se producía depositación. Así se desarrolló un complejo depósito de naturaleza continental, i.e. Formación Tordillo y equivalentes.
El área fué cubierta nuevamente por el mar durante el Tithoniano-Cretácico Temprano por una extensa transgresión. En el Tithoniano Temprano medio, una elevación del nivel del mar causó la rápida inmersión de la cuenca y una fuerte expansión del área marina, con depositación de margas y calizas bituminosas (Formación Vaca Muerta). La depositación de areniscas litorales estuvo restringida a algunas áreas marginales. Durante el Tithoniano Tardío, como resultado de una fuerte progradación de carbonatos clásticos desde el sur y este, el área de sedimentación marina se redujo y las facies marginales y continentales se distribuyeron ampliamente. Coincidentemente la diversidad de los amonites (Fig. 10) muestra un fuerte incremento entre el Tithoniano Temprano (Zona de Virgatosphinctes mendozanus, ≈ Zona de Darwini tardío) y el Medio (Zona de Windhauseniceras internispinosum, ≈ Zona de Ponti), con una ligera disminución en la Zona de Aulacosphintes proximus (≈ límite Fallauxi- Ponti), seguido por una fuerte caída al comienzo del Tithoniano Tardío (Zona de Corongoceras alternans, zonas de Microcanthum- Durangites) y otra elevación a través del límite Tithoniano-Berriasiano (Zona de Substeueroceras koeneni ≈ zonas de Durangites spp. - Jaco-bi/Grandis y Occitanica inicial).
AGRADECIMIENTOS
Se agradece a los árbitros, S. Fernández López, Madrid y H.A. Leanza, Buenos Aires, y a las editoras A.M. Zavattieri y L. Giambiagi, Mendoza, los comentarios y sugerencias, que ayudaron a mejorar la versión original. Los errores que puedan encontrarse son responsabilidad exclusiva del autor. Este trabajo fue financiado con fondos provistos por el Consejo Nacional de Investigaciones Científicas y Técnicas (PIP 5635) y la Agencia Nacional de Promoción Científica y Tecnológica (PICT 07-26236) y es una contribución al IGCP 506 "Jurassic Boundary Events".
TRABAJOS CITADOS EN EL TEXTO
1. Álvarez, P. 1996. Los depósitos triásicos y jurásicos de la Alta Cordillera de San Juan. Dirección Nacional del Servicio Geológico, Anales 24: 59-137. [ Links ]
2. Blasco, G., Nullo, F. y Proserpio, C. 1979. Aspidoceras en Cuenca Austral, Prov. de Santa Cruz. Revista de la Asociación Geológica Argentina 34: 282-293. [ Links ]
3. Burckhardt, C. 1900. Profils géologiques transversaux de la Cordillère Argentino-Chilienne. Stratigraphie et tectonique. Anales del Museo de La Plata, Sec. Geol. Min. 2: 1-136. [ Links ]
4. Burckhardt, C. 1903. Beiträge zur Kenntniss der Jura- und Kreide-formation der Cordillera. Palaeontographica 50(1-3): 1-144. [ Links ]
5. Feruglio, E. 1936-37. Palaeontographia Patagonica. Istituto Geologico della Universita di Padova, Memorie 11-12: 1-381. [ Links ]
6. Gerth, E. 1925. La fauna Neocomiana de la Cordillera Argentina en la parte meridional de la provincia de Mendoza. Academia Nacional de Ciencias, Actas 9: 57-132, Córdoba. [ Links ]
7. Gröschke, M. y Hillebrandt, A. v. 1994. The Bathonian in northern Chile. Geobios, M.S. 17: 255-264. [ Links ]
8. Gygi, R.A. y Hillebrandt, A. v. 1991. Ammonites (mainly Gregoryceras) of the Oxfordian (Late Jurassic) in northern Chile and time-correlation with Europe. Schweizerische Paläontologische Abhandlungen 111: 137-185. [ Links ]
9. Hall, R.L. y Westermann, G.E.G 1980. Lower Bajocian (Jurassic) cephalopod fauna from western Canada and proposed assemblage zones for the Lower Bajocian of North America. Palaeontographia Americana 9(52): 5-93. [ Links ]
10. Hillebrandt, A. v. 1987. Liassic ammonite zones of South America and correlations with other provinces. With description of new genera and species of ammonites. En Volkheimer,W. (ed.) Bioestratigrafía de los Sistemas regionales del Jurásico y Cretácico de América del Sur: 111-157, Mendoza. [ Links ]
11. Hillebrandt, A. v. 1988. Ammonite biostratigraphy of the South American Hettangian. En: Rocha, R.B. y Soares, A.F. (eds.) 2nd International Symposium on Jurassic Stratigraphy, 1: 55-70. [ Links ]
12. Hillebrandt, A.v. 1990. Der Untere Jura im Gebiet des Rio Atuel (Provinz Mendoza, Argentinien). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 181(1-3): 143-157. [ Links ]
13. Hillebrandt, A. v. 2000. Die Ammoniten-Fauna des sudamerikanischen Hettangium (basaler Jura), Teil I, II, III, Palaeontographica A257: 85-189, A258: 1-64, 65-116. [ Links ]
14. Hillebrandt, A.v. 2002. Ammoniten aus dem oberen Sinemurium von Sudamerika. Revue de Paléobiologie 21(1): 35-147. [ Links ]
15. Hillebrandt, A.v. 2006. Ammoniten aus dem Pliensbachium (Carixium und Domerium) von Sudamerika. Revue de Paléobiologie 25(1): 1-403. [ Links ]
16. Hillebrandt, A. von y Schmidt-Effing, R. 1981. Ammoniten aus dem Toarcium (Jura) von Chile (Sudamerika). Zitteliana 6: 3-74. [ Links ]
17. Hillebrandt, A. v. y Westermann, G.E.G. 1985. Aalenian (Jurassic) Ammonite Faunas and Zones of the Southern Andes. Zitteliana 12: 3-55. [ Links ]
18. Hillebrandt, A.v., Smith, P., Westermann, G.E.G. y Callomon, J.H. 1992. Ammonite zones of the Circum-Pacific region. En: Westermann, G.E.G. (Ed.): The Jurassic of the Circum-Pacific, Cambridge University Press, 247-272, Cambridge. [ Links ]
19. Iglesia Llanos, M.P. y Riccardi, A.C. 2000. The Neuquén composite section: magnetostratigraphy and biostratigraphy of the marine lower Jurassic from the Neuquén basin (Argentina). Earth and Planetary Science Letters 181: 443-457. [ Links ]
20. Kraemer, P.E. y Riccardi, A.C. 1997. Estratigrafía de la región comprendida entre los lagos Argentino y Viedma (49° 40' - 50°10' lat. S), Provincia de Santa Cruz. Revista de la Asociación Geológica Argentina 52(3): 333-360. [ Links ]
21. Lanés, S. 2005. Late Triassic to Early Jurassic sedimentation in northern Neuquén Basin, Argentina: Tectosedimentary Evolution of the First Transgression. Geologica Acta 3(2): 81- 103. [ Links ]
22. Leanza, A.F. 1945. Ammonites del Jurásico superior y del Cretácico inferior de la sierra Azul, en la parte meridional de la provincia de Mendoza. Anales del Museo de La Plata (N.S.) 1: 1-99. [ Links ]
23. Leanza, A.F. 1947. Descripción de la Fáunula Kimmeridgiana de Neuquén. Dirección de Nacional de Geología y Minería, Informes Preliminares y Comunicaciones 1: 1-24. [ Links ]
24. Leanza, A.F. 1968. Anotaciones sobre los fósiles Jurásico-Cretácicos de Patagonia Austral (Colección Feruglio) conservados en la Universidad de Bologna. Acta Geológica Lilloana 9: 121-187. [ Links ]
25. Leanza, H.A. 1980. The Lower and Middle Tithonian Ammonite Fauna from Cerro Lotena, Province of Neuquén, Argentina. Zitteliana 5: 3-49. [ Links ]
26. Leanza, H.A. 1981. The Jurassic-Cretaceous boundary beds in West Central Argentina and their ammonite zones. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 161(1): 62-92. [ Links ]
27. Leanza, H.A. 1996. Advances in the ammonite zonation around the Jurassic/Cretaceous boundary in the Andean Realm and correlation with Tethys. Jost Wiedmann Symposium, Abstracts: 215-219, Tübingen. [ Links ]
28. Leanza, H.A. y Blasco, G. 1990. Estratigrafía y ammonites pliensbachianos del area del arroyo Ñireco, Neuquen, Argentina, con la descripcion de Austromorphites gen. nov. Revista de la Asociación Geológica Argentina 45(1-2): 159- 174. [ Links ]
29. Leanza, H.A. y Zeiss, A. 1990. Upper Jurassic Lithographic Limestones from Argentina (Neuquén Basin): Stratigraphy and Fossils. Facies 22: 169-186. [ Links ]
30. Leanza, H.A. y Zeiss, A. 1992. On the ammonite fauna of the Lithographic Limestones from the Zapala region (Neuquén province, Argentina), with the description of a new genus. Zentralblatt für Geologie und Paläontologie Teil I, 1991, H.6: 1841-1850. [ Links ]
31. Leanza, H.A. y Zeiss, A. 1994. The "Lithographic limestones" of Zapala (Central Argentina) and their Ammonite fauna. Geobios M.S.16: 245-250. [ Links ]
32. Legarreta, L. y Gulisano, C.A. 1989. Análisis estratigráfico secuencial de la cuenca Neuquina. Serie Correlación Geológica 6: 221-243. [ Links ]
33. Legarreta, L. y Uliana, M.A. 1996. The Jurassic succession in west-central Argentina: stratal patterns, sequences and paleogeographic evolution. Palaeogeography, Palaeoclimatology, Palaeoecology 120: 303-330. [ Links ]
34. Legarreta, L. y Uliana, M.A. 1999. El Jurásico y Cretácico de la Cordillera Principal y la Cuenca Neuquina. 1, Facies sedimentarias. Instituto de Geología y Recursos Minerales, Anales 29 (16): 399-432. [ Links ]
35. Odin, G.S., Baadsgaard, H., Hurford, A.J. y Riccardi, A.C. 1992. U-Pb & fission track geochronology of Bathonian-Callovian "tuffs" from Argentina. En Odin, G.S. (ed.) Phanerozoic time scale. Bulletin de Liaison et Information 11: 11-17. [ Links ]
36. Ogg, J.G. 2004. Status of Divisions of the International Geologic Time Scale. Lethaia 37: 183-199. [ Links ]
37. Parent, H. 1997. Ontogeny and sexual dimorphism of Eurycephalites gottschei (Tornquist) (Ammonoidea) of the Andean Lower Callovian (Argentine-Chile). Geobios 30(3): 407- 419. [ Links ]
38. Parent, H. 1998. Upper Bathonian and lower Callovian ammonites from Chacay Melehué, (Argentina). Acta Palaeontologica Polonica 43(1): 69-130. [ Links ]
39. Parent, H. 2001. The Middle Tithonian (Upper Jurassic) ammonoid fauna of Cañadón de los Alazanes, southern Neuquén-Mendoza Basin, Argentina. Boletín del Instituto de Fisiografía y Geología 71(1-2): 19-38. [ Links ]
40. Parent, H. 2003. The ataxioceratid ammonite fauna of the Tithonian (Upper Jurassic) of Casa Pincheira, Mendoza (Argentina). Journal of South American Earth Sciences 16: 143-165. [ Links ]
41. Parent, H. 2006. Oxfordian and Late Callovian ammonite faunas and biostratigraphy of the Neuquén-Mendoza and Tarapacá basins (Jurassic, Ammonoidea, western South America). Boletín del Instituto de Fisiografía y Geología 76(1-2): 1-70. [ Links ]
42. Parent, H. y Capello, O.D. 1999. Amonites del Tithoniano de Casa Pincheira. Revue de Paléobiologie 18(1): 347-353. [ Links ]
43. Parent, H., Scherzinger, A. y Schweigert, G. 2006 a. The earliest ammonite faunas from the Andean Tithonian of the Neuquén-Mendoza Basin, Argentina. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 241 (2): 253-268. [ Links ]
44. Parent, H., Schweigert, G. y Meléndez, G. 2006b. Oxfordian perisphinctid ammonites from Chacay Melehué, Argentina. Palaontologische Zeitschrift 80(4): 307-324. [ Links ]
45. Remane, J. 2000. International Stratigraphic Chart. International Union of Geological Sciences. [ Links ]
46. Riccardi, A.C. 1983. The Jurassic of Argentina and Chile. En Moullade, M. y Nairn, A.E.M. (eds.) The Phanerozoic Geology of the World II, Elsevier, The Mesozoic B: 201-263. [ Links ]
47. Riccardi, A.C. 1984. Las asociaciones de amonitas del Jurásico y Cretácico de la Argentina. 9 Congreso Geológico Argentino, Actas 4: 559- 595. [ Links ]
48. Riccardi, A.C. 1985. Los Eurycephalitinae Andinos (Ammonitina, Jurásico Medio): Modelos evolutivos y Resolución Paleontológica. Instituto de Fitotecnia, Boletín Genético 13: 1- 27. [ Links ]
49. Riccardi, A.C. 1988. The Cretaceous System of southern South America. Geological Society of America, Memoir 168: 1-161. [ Links ]
50. Riccardi, A.C. 1991a. Jurassic and Cretaceous marine connections between the southeast Pacific and Tehys. Palaeogeography, Palaeoclimatology, Palaeoecology 87: 155-189. [ Links ]
51. Riccardi, A.C. 1991b. Significación estratigráfica de la diversidad de los amonites en el Jurásico de la Argentina. 6° Congreso Geológico Chileno, Resúmenes Ampliados: 149-152. [ Links ]
52. Riccardi, A.C. 1993. El Jurásico en la Argentina. En: Riccardi, A.C. y Damborenea, S.E. (Eds), Léxico Estratigráfico de la Argentina, Volumen 9, Jurásico. Asociación Geológica Argentina, Serie B (Didáctica y Complementaria) 21: 9-27. [ Links ]
53. Riccardi, A.C. 1996. Heterochronic changes in the Andean Neuqueniceratinae (Ammonoidea, Middle Jurassic). 4º International Symposium Cephalopods, Present and Past (Granada, 1996), Abstracts Volume: 146. [ Links ]
54. Riccardi, A.C. 2000. The Podagrosiceratinae (Ammonitida) in the Upper Toarcian-Lower Bajocian (Jurassic) of Argentina. Revue de Paleobiologie, vol. spec. 8: 13-28. [ Links ]
55. Riccardi, A.C. 2005. First teuthid cephalopod from the Lower Jurassic of South America (Neuquén Basin, Argentina). En: Riccardi, A.C. (Ed.): New insigths into the Juurassic record of South America (Andes and Patagonia). Geologica Acta 3(2): 179-184. [ Links ]
56. Riccardi, A.C. y Gulisano, C.A. 1992. Unidades Limitadas por Discontinuidades. Su aplicación al Jurásico andino. Revista de la Asociación Geológica Argentina 45(3-4): 346-364. [ Links ]
57. Riccardi, A.C. e Iglesia Llanos, M.P. 1999. Primer hallazgo de amonites en el Triásico de la Argentina. Revista de la Asociación Geológica Argentina 54(3): 298-300. [ Links ]
58. Riccardi, A.C. y Westermann, G.E.G. 1991a. Middle Jurassic Ammonoid Fauna and Biochronology of the Argentine-Chilean Andes. Part III: Bajocian-Callovian Eurycephalitinae, Stephanocerataceae, Palaeontographica A 216: 1-110. [ Links ]
59. Riccardi, A.C. y Westermann, G.E.G. 1991b. Middle Jurassic Ammonoid Fauna and Biochronology of the Argentine-Chilean Andes. Part IV: Bathonian-Callovian Reineckeiidae. Palaeontographica A216: 111-145. [ Links ]
60. Riccardi, A.C. y Westermann, G.E.G. 1999. An early Bathonian Tethyan ammonite fauna from Argentina. Palaeontology 42(2): 193-209. [ Links ]
61. Riccardi, A.C., Damborenea, S.E., Manceñido, M.O. y Ballent, S.C. 1988a. Hettangiano y Sinemuriano marinos en Argentina. Actas V Congreso Geológico Chileno 2: C359-C373. [ Links ]
62. Riccardi, A.C., Westermann, G.E.G. y Elmi, S. 1988b. Zonas de amonites del Bathoniano - Caloviano inferior de los Andes Argentino- Chilenos. 5° Congreso Geológico Chileno, Actas 2: C415-C426. [ Links ]
63. Riccardi, A.C., Westermann, G.E.G. y Elmi, S. 1989. The Bathonian Callovian Ammonite Zones of the Argentine Chilean Andes. Geobios 22(5): 553-597. [ Links ]
64. Riccardi, A.C., Damborenea, S.E. y Mancenido, M.O. 1990a. The Lower Jurassic of South America and Antarctic Peninsula. Newsletter on Stratigraphy 21(2): 75-103. [ Links ]
65. Riccardi, A.C., Westermann, G.E.G. y Damborenea, S.E. 1990b. Middle Jurassic of South America and Antarctic Peninsula. Newsletter on Stratigraphy 21(2): 105-128. [ Links ]
66. Riccardi, A.C., Leanza, H.A. y Volkheimer, W. 1990c. Upper Jurassic of South America and Antarctic Peninsula. Newsletter on Stratigraphy 21(2): 129-147. [ Links ]
67. Riccardi, A.C., Westermann, G.E.G. y Elmi, S. 1991a. Biostratigraphy of the upper Bajocianmiddle Callovian (Middle Jurassic), South America. Journal of South American Earth Sciences 4(3): 149-157. [ Links ]
68. Riccardi, A.C., Damborenea, S.E., Manceñido, M.O. y Ballent, S.C. 1991b. Hettangian and Sinemurian (Lower Jurassic) biostratigraphy of Argentina. Journal of South American Earth Sciences 4(3): 159-170. [ Links ]
69. Riccardi, A.C., Gulisano, C.A., Mojica, J., Palacios, O., Schubert, C. y Thomson, M.R.A. 1992. Western South America and Antarctica. En Westermann, G.E.G. (ed.) The Jurassic of the Circum-Pacific, Cambridge University Press, 122-161, Cambridge. [ Links ]
70. Riccardi, A.C., Damborenea, S.E., Manceñido, M.O. y Ballent, S.C. 1994. Middle Jurassic biostratigraphy of Argentina. Geobios M.S. 17: 423-430. [ Links ]
71. Riccardi, A.C., Leanza, H.A., Damborenea, S.E., Manceñido, M.O., Ballent, S.C. y Zeiss, A. 2000. Marine Mesozoic Biostratigraphy of the Neuquén Basin. Zeitschrift für Angewandte Geologie, SH 1(2000): 103-108. [ Links ]
72. Riccardi, A.C., Damborenea, S.E., Manceñido, M.O. e Iglesia Llanos, M.P. 2004. The Triassic/ Jurassic boundary in the Andes of Argentina. Rivista Italiana di Paleontologia e Stratigrafia 110(1): 69-76. [ Links ]
73. Schneid, T. 1915. Die Geologie der frankischen Alb zwischen Eichstatt und Neuburg a.D. [ Links ]
74. Stipanicic, P.N. 1951. Sobre la presencia del Oxfordense Superior en el Arroyo de la Manga (Provincia de Mendoza). Revista de la Asociación Geológica Argentina 6: 213-239. [ Links ]
75. Stipanicic, P.N. 1996. Milestones in the Study of the Jurassic of Argentina. En Riccardi, A.C. (ed.) Advances in Jurassic Research, Transtec Publications, 23-42, Zurich. [ Links ]
76. Stipanicic, P.N., Westermann, G.E.G. y Riccardi, A.C. 1976. The Indo-Pacific Ammonite Mayaites in the Oxfordian of the Southern Andes. Ameghiniana 12: 281-305. [ Links ]
77. Valencio, S.A., Cagnoni, M.C., Ramos, A.M., Riccardi, A.C. y Panarello, H.O. 2005. Chemostratigraphy of the Pliensbachian, Puesto Araya Formation (Neuquén Basin, Argentina). En Riccardi, A.C. (ed.) New insigths into the Jurassic record of South America (Andes and Patagonia). Geologica Acta 3(2): 147-154. [ Links ]
78. Vergani, G.D., Tankard, A.J., Belotti, H.J. y Welsink, H.J. 1995. Tectonic evolution and paleogeography of the Neuquén basin, Argentina. En Tankard, A.J., Suárez S.R. y Welsink, J. (eds.) Petroleum basins of South America, American Association of Petroleum Geologists, Memoir 62: 383-402. [ Links ]
79. Verma, H.M. y Westermann, G.E.G. 1973. The Tithonian (Jurassic) Ammonite fauna and stratigraphy of Sierra Catorce, San Luis Potosi, Mexico. Bulletins of American Paleontology 63(277): 1-320. [ Links ]
80. Vicente, J.C. 2005. Dynamic paleogeography of the Jurassic Andean Basin: pattern of transgression and localisation of main straits through the magmatic arc. Revista de la Asociación Geológica Argentina 60(1): 221-250. [ Links ]
81. Vicente, J.C. 2006. Dynamic paleogeography of the Jurassic Andean Basin: pattern of regression and general considerations on main features. Revista de la Asociación Geológica Argentina 61(3): 408-437. [ Links ]
82. Weaver, C. 1931. Paleontology of the Jurassic and Cretaceous of West Central Argentina. Memoir University of Washington 1: 1-496, Seattle. [ Links ]
83. Westermann, G.E.G. y Riccardi, A.C. 1979. Middle Jurassic Ammonoid Fauna and Biochronology of the Argentine-Chilean Andes. Part II: Bajocian Stephanocerataceae. Palaeontographica A 164(4-6): 85-188. [ Links ]
84. Westermann, G.E.G. y Riccardi, A.C. 1982. Ammonoid fauna from the early Middle Jurassic of Mendoza Province, Argentina. Journal of Paleontology 56(1): 11-41. [ Links ]
85. Westermann, G.E.G. y Riccardi, A.C. 1985. Middle Jurassic ammonite evolution in the Andean Province and emigration to Tethys. En: Bayer, U. y Seilacher, A. (eds.), Sedimentary and Evolutionary Cycles. Lecture Notes in Earth Sciences 1: 6-34. [ Links ]
86. Westermann, G.E.G. y Riccardi, A.C. 1989. Jurassic Stage boundaries in South America. International Subcommission on Jurassic Stratigraphy, Newsletter 19:16-22. [ Links ]
87. Zeiss, A. 1986. Comments on a tentative correlation chart for the most important marine provinces at the Jurassic/Cretaceous boundary. Acta Geologica Hungarica 29(1-2): 27-30. [ Links ]
Recibido: 15 de febrero, 2008.
Aceptado: 23 de junio, 2008.


{kind=link}