










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de la Asociación Geológica Argentina
versión impresa ISSN 0004-4822versión On-line ISSN 1851-8249
Rev. Asoc. Geol. Argent. v.64 n.1 Buenos Aires abr. 2009
Darwin forest at agua de la zorra: the first in situ forest discovered in South America by Darwin in 1835
Mariana Brea1,4, Analía E. Artabe2,4 and Luis A. Spalletti3,4
1Laboratorio de Paleobotánica, Centro de Investigaciones Científicas, Diamante. CICYTTP-Diamante, Consejo Nacional de Investigaciones Científicas y Técnicas, Diamante, Entre Ríos, Email: cidmbrea@infoaire.com.ar
2División Paleobotánica, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata. Email: aeartabe@museo.fcnym.unlp.edu.ar
3Centro de Investigaciones Geológicas (CONICET-UNLP), La Plata. Email: spalle@cig.museo.unlp.edu.ar
4Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET).
ABSTRACT
The Agua de la Zorra area (near Uspallata, Mendoza, Argentina) is one of the best renowned fossil localities of the country because of its spectacular in situ fossil forest. This forest was discovered by Charles Darwin in 1835, who described this forest as monotypic and assigned it a Tertiary age. Nowadays, this fossil locality is known as the Darwin Forest. Over a century and a half later it was reinterpreted as a mixed Middle Triassic forest and a new fossil monotypic palaeocommunity of horsetails was discovered. This palaeovegetation is included in the Paramillo Formation (i.e., lower section the Potrerillos Formation) of northwestern Cuyo Basin, Mendoza province (69°12' W and 32°30' S). The sediments were deposited in a sinuous fluvial system, in which channel-filling sand bodies were associated with mud-dominated floodplain deposits. The palaeoforest grew on an andisol soil that developed on volcaniclastic floodplain deposits. It had a density of 427 -759 trees per hectare, and was constituted by conifers and corystosperms distributed in two arboreal strata. The highest reached 20-26 m tall, and was dominated by corystosperms, but it also included the tallest conifers. The second stratum, mainly composed of conifers, ranged between 16-20 m tall. The forest has also emergent corystosperms, which reached 30 m tall. The understorey was composed of ferns. Growth ring anatomy suggests that conifers could have had an evergreen habit. Structure of vegetation, growth ring analyses and sedimentation suggest that the forest developed under dry, subtropical, and strongly seasonal conditions.
Keywords: Palaeovegetation; Palaeoecology; Palaeoenvironments; Middle Triassic; Southwestern Gondwana.
RESUMEN: El Bosque Darwin en Agua de la Zorra: El primer bosque in situ descubierto en América del Sur por Darwin en 1835. El área de Agua de la Zorra (cerca de Uspallata, Mendoza, Argentina) es uno de los sitios fósiles más espectaculares y renombrados del país porque aflora un bosque in situ. Este bosque descripto como monotípico y asignado al Terciario fue descubierto por Charles Darwin en 1835. Un siglo y medio más tarde, se reinterpretó como un bosque mixto del Triásico Medio y se reconoció una nueva paleocomunidad monotípica de esfenófitas. La paleovegetación proviene de la Formación Paramillo (= sección inferior de la Formación Potrerillos) de la región noroeste de la Cuenca Cuyana, provincia de Mendoza (69°12'O y 32°30'S). Los sedimentos fueron depositados por un sistema fluvial de alta sinuosidad. El bosque fósil creció sobre suelos del tipo andisol que se desarrollaron sobre depósitos de planicies volcaniclásticas. El bosque Darwin tiene una densidad de 427-759 árboles por hectárea. Este está constituido por coníferas y corystospermas y tiene dos estratos arbóreos. El más alto se desarrolla entre los 20-26 m, y es dominado por corystospermas, pero también por las coníferas más altas. El segundo estrato arbóreo, principalmente compuesto de coníferas, con un rango entre 16-20 m. El bosque tiene también algunos emergentes de corystospermas las cuales alcanzan hasta 30 m de altura. El sotobosque estaba compuesto de helechos. La anatomía de los anillos de crecimiento sugiere que las coníferas podrían haber tenido hábito siempreverde. La estructura de la vegetación, el análisis de los anillos de crecimiento y la sedimentología sugieren que este bosque se desarrollo bajo condiciones climáticas subtropicales, secas y estacionales.
Palabras clave: Paleovegetación; Paleoecología; Paleoambientes; Triásico Medio; Sudoeste de Gondwana.
INTRODUCTION
The first fossil plants in South America were recorded by European naturalists during the 18th and 19th centuries (Ottone 2005). The British naturalist Charles Robert Darwin (1809- 1882) was the first who provided palaeontological and geological observations of the Agua de la Zorra area, Uspallata, near Villavicencio, Mendoza province in Argentine territory (Darwin 1839a, 1839b, 1845, 1846). Later, this famous fossiliferous locality was visited by other European naturalistexplorers such as Burmeister 1861, Doering 1882, Stelzner 1885, Avé-Lallemant 1891 (see Ottone 2005).
The expeditions of Charles Darwin -on board HMS Beagle- during his historic journery around the world between 1831 and 1836, contributed valuable information and observations on the geology, plant and animal fossils, and extant organisms of South America. Darwin also collected a huge number of specimens and samples, many of them new to science.
Darwin arrived in Argentina in 1833 and visited the Río de La Plata area, La Bajada (now Paraná city, Entre Ríos), the Santa Cruz River and many other sites in Patagonia and Tierra del Fuego. Then, after crossing the Cordillera de los Andes from Chile, he arrived in Mendoza province and discovered the first in situ forest in South America, at Agua de la Zorra, located about 25 km from the town of Uspallata (Conwentz 1885, Rusconi 1941, Harrington 1971, Brea 1995, Ottone 2005). He recorded the presence of 52 fossil tree trunks measuring 90-152 cm in diameter, buried in sandstones or volcanic sandstones and standing out as columns several meters high (Darwin 1846). This discovery by Darwin is remembered by a monolith (Fig. 1).


Figure 1: a. Charles Darwin's tribute plaque at Agua de la Zorra, near Uspallata, Mendoza, Argentina. This photo was taken in 1993. b. Panoramic view of Triassic sequences, the arrow shows the plaque.
On 30 March 1835 he collected the first specimens of fossil wood from Agua de la Zorra. The samples were sent to London and were referred by Robert Brown to the genus Araucarites (Darwin 1846). Recently, one of the authors (M. B.) carried out three research field trips as part of work towards her PhD dissertation. Brea (1995) characterized and studied the Triassic units at Agua de la Zorra from a palaeobotanical and sedimentological viewpoint. She defined two palaeocommunities in the Paramillo Formation, together with their associated palaeoenviroments (Fig. 2). The first paleocommunity, the in situ Darwin Forest (Fig. 2), appears at four fossiliferous sites (Brea 1995), while the second was found at only one locality and included only fossil horsetails (Brea and Artabe 1999, Brea et al. 2008).

Figure 2: The Darwin Forest at Agua de la Zorra area (near Uspallata, Mendoza). a-b-c-e. Large fossil tree stumps in growth position in Triassic sequences; d-f-h. Petrified standing trees at fossiliferous sites C; g. Large fossil log exposed in Triassic sediments at fossiliferous site B.
The Darwin Forest was first referred to the Tertiary by Darwin (1839a, 1839b), but new data now available suggest a late Middle Triassic age (Spalletti et al. 1999, Brea et al. 2008). The most important feature of the Darwin Forest is that the trees are still preserved at the sites on which they grew (Fig. 2); it is the best renowned of its age in southwest Gondwana. Although petrified forests in growth position are very scarce in the geological record, the Triassic fossil record of SW Argentina shows four in situ fossil forests (Paramillo, Cortaderita, Ischigualasto, and Río Blanco Formations) and their occurrences have contributed significantly to our understanding of Triassic Gondwana ecosystems (Zamuner 1992, Artabe et al. 2001, 2007a, Brea et al. 2008).
The Darwin Forest was reinterpreted as a subtropical dry seasonal forest (Brea et al. 2005, 2008). It grew on an andisol soil that developed on volcaniclastic floodplain deposits. The volcanic detritus and the rhythmic amalgamation of upper flow-regime tractional deposits overlying the andisol indicate that the forest was buried rapidly by a subaerial, cool and wet pyroclastic base surge flow (Poma et al. 2004, Brea et al. 2008).
The continental Triassic succession of southwestern Gondwana occurs in a series of narrow rifts produced as a result of Triassic continental extension. These rift basins are composed of a continental clastic infilling, and record complex interactions between alluvial, fluvial, deltaic and lacustrine depositional systems with intercalations of volcaniclastic sequences in most of these basins. The rich floristic record allowed recognition of several assemblages, biozones and stages characterized by floristic events (Spalletti et al. 1999, 2003, Artabe et al. 2001, 2003, 2007b, Morel et al. 2003).
Over the last two decades, most investigations on the Argentinian Triassic have focused on the gross stratigraphy and taxonomy of fossil plants (Stipanicic 2001, Stipanicic and Marsicano 2002, Zamuner et al. 2001, and references therein). However, recently published studies (Spalletti et al. 1999, 2003, Artabe et al. 2001, 2003, 2007b, Morel et al. 2003) provided an increasing understanding of how the Triassic palaeofloras developed and changed over time as a response to variations in depositional systems, tectonism and climate. The most spectacular fossil sites of the Paramillo Formation- in the Agua de la Zorra area -, are located in the northernwestern sector of the Cuyo Basin (between 32º and 36º SL). The Paramillo Formation is composed of a 140 m thick succession of clast-supported conglomerates, pebbly sandstones, tuffaceous sandstones and mudstones (Figs. 3 and 4). These deposits have been previously described by Harrington (1971), Strelkov and Álvarez (1984), Kokogian and Mancilla (1989), Ramos and Kay (1991), Massabie (1985), Massabie et al. (1985), Linares and González (1990), Ramos (1993), Kokogian et al. (1993). These authors all agreed in interpreting the Paramillo Formation as deposited in highly sinuous fluvial systems.
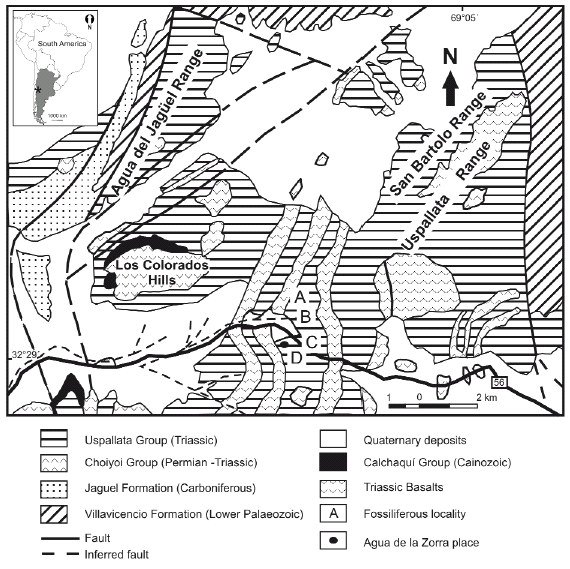
Figure 3: Location map showing Agua de la Zorra fossiliferous locality, Mendoza province,
Argentina indicating the four fossiliferous sites: A, B, C, D respectively.
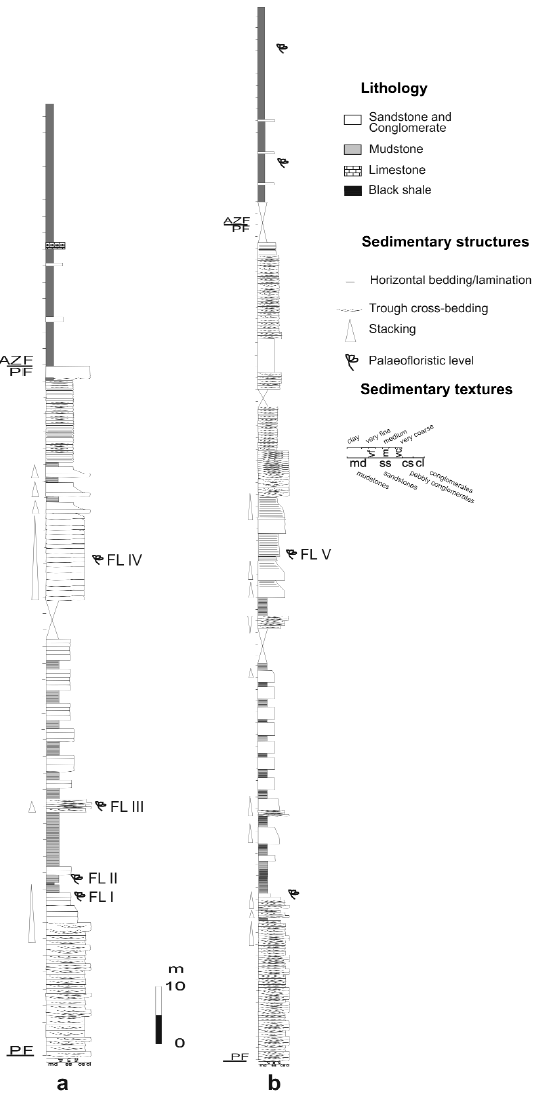
Figure 4: a. Lithostratigraphic section of the Paramillo and Agua de la Zorra Formations at locality A, showing the main lithofacies and the position of the fossil forest level. b. Lithostratigraphic section of the Paramillo and Agua de la Zorra Formations at locality C. PF: Paramillo Formation, AZF: Agua de la Zorra Formation. FL I - V: fossiliferous levels.
Previous palaeobotanical studies, comprising mostly lists of fossil plants, were published by Darwin (1846), Conwentz (1885), Stappenbeck (1910), Kurtz (1921), Du Toit (1927), Groeber (1939), Windhausen (1941), Harrington (1971) and Stipanicic et al. (1996), while modern systematic contributions were offered by Brea (1995, 1997, 2000), Brea and Artabe (1999), Artabe and Brea (2003) and Brea et al. (2005, 2008).
THE PARAMILLO FORMATION
The Paramillo Formation is a volcaniclastic unit composed of yellowish lithic sandstones, brownish and yellowish tuffaceous sandstones, dark gray and green shales and mudstones, and pink to reddish ash fall tuffs (Fig. 4). The sedimentary record of the overlying Agua de la Zorra Formation is dominated by bituminous shales and marls with subordinate intercalations of yellowish fine-grained sandstones and mudstones (Fig. 4). Both units are intruded by several sills of olivine diabase dated 235 + 5 Ma (K/Ar whole age) by Ramos and Kay (1991). At that time Mendoza was a part of the vast supercontinent called Gondwana and it was placed approximately at the same geographic latitude as it is nowadays.
Brea (1995) studied in more detail the upper part of the Paramillo Formation and the lowermost Agua de la Zorra Formation and defined several lithofacies and facies associations. The Paramillo Formation consists of cross-bedded conglomerates, cross-bedded, plane-bedded and massive pebbly sandstones, cross-bedded, massive, plane bedded and ripple-laminated sandstones and intercalations of laminated mudstones and shales. Brea (1995) interpreted that these sediments were deposited in a highly sinuous (meandering) fluvial system, in which channel-filling sand bodies are associated with mud-dominated floodplain deposits. The cyclic arrangement of well preserved soil horizons and sedimentary deposits lacking evidence of soil formation indicates alternation of periods with strong sediment aggradation produced by non-channelized high-regime flows with periods characterized by very low accommodation rates that favoured the development of immature soil profiles (Brea et al. 2008). The volcanic nature of detrital components suggests that the highly aggrading non-channelized flows that produced the burial of the Darwin Forest could be related to pyroclastic events (Poma et al. 2004). The forest might have died as a result of a diluted, subaerial, cool and wet base surge pyroclastic flow (Poma et al. 2004, Brea et al. 2008).
The Paramillo Formation, where the Darwin Forest emerges, has been correlated with the lower section of the Potrerillos Formation (Spalletti et al. 1999, Morel et al. 2003) on the basis of Assemblage Biozone chronology, analysis of main stratigraphic unconformities, and evolution of basin infill.
ECOLOGICAL RECONSTRUCTION OF THE DARWIN FOREST
The Triassic landscape at the Agua de la Zorra area was very different in comparison with the modern-day scenery. Sphenopsids dominated the flooded areas and conifers and corystosperms were the most important components of the arboreal vegetation. Four exposures of the Darwin Forest were found in the Paramillo Formation during fieldwork carried out in 1993-1994 (Figs. 2 and 3). This unit was thoroughly examined at two localities: Darwin and El Sauce (Figs. 3 and 4), where the lithology was logged bed-by-bed in vertical sections at a 1:100 scale (see Brea et al. 2008).
Imperfect carbonization processes (Poma et al. 2004) preserved the stumps and in just over 2 km2 one hundred and twenty stumps in growth position and fallen logs were counted (Fig. 2). The fossiliferous levels (FL) with stumps were found at four localities belonging to the same in situ forest. Thus, FL IV (at A locality), FL V (at B and C localities) and FL VI (at D locality) belong to a single stratigraphic level (Figs. 2 and 4). Fossiliferous levels FL I and FL III found at locality A (Fig. 4) have ferns preserved as impressions-compressions (Brea 2000).
The sedimentary sequences in which this forest is preserved consist of continental volcaniclastic units, resulting from deposition on a highly sinuous fluvial system associated with river flood-plains (Fig. 4). The palaeoecological reconstruction of the Darwin Forest was based on the quantitative data of the mapped forest (mean separation of trees, basal area per ha, species distribution) integrated with the taxonomic and sedimentological information (Brea et al. 2008). The mixed forest was composed by corystosperms (30-40%) and conifers (60-70%). The corystosperms are a group of extinct plants with mostly fern-like foliage with ovules borne on modified leaves or cupules.
This group was the dominant seed plant in Gondwana during the Triassic (Stewart 1983). All conifers are a diverse group of trees and shrubs that underwent a major radiation during the Triassic period, when the first occurrence and radiation of the eight conifer families occurred (Willis and McElwain 2002). The conifers have a pyramidal arborescent growth form, with cone-bearing seed plants; the foliage is either needle-like or scale-like. The plants are mainly small, long, and thin arranged spirally and bearing the male and female reproductive organs in separate cones on different trees, or indifferent parts of the same.
The average density of the tree stumps in this forest is 556 trees/ha. The spatial autocorrelation for the species variable indicates that in some places the corystosperms and conifers are intermingled while in others the corystosperms and conifers seem to be aggregated into cohesive social groups (Fig. 5). The Darwin Forest reveals two strata: the highest, developed between 20-24 m, has a preponderance of corystosperms but also the tallest conifers; the second stratum, mainly composed of conifers, ranges between 16-20 m. The forest also has emergent corystosperms which reach 30 m tall (Brea et al. 2008). The corystosperms were assigned by Artabe and Brea (2003) to Cuneumxylon spallettii and constitute the dominant species in the Darwin Foest.

Figure 5: Reconstruction of the Triassic Darwin Forest landscape in a high sinuosity fluvial system, in which channel-filling sand bodies are associated with mud-dominated floodplain deposits. The canopy is integrated by two arboreal strata and emergent trees with conifers and corystosperms. The understorey is formed by ferns. Cinodonts are characteristic tetrapods during the Triassic of the Cuyo Basin (drawing by Jorge Gonzalez).
This fossil forest presents values of biomass and stand basal area comparable to those of the current subtropical seasonal forests (Brea et al. 2008). These forests develop under a climatic regime that includes an annual cycle with one season in which water is unavailable to plants because of lack of precipitation. The dry season alternates with another in which there is abundant water. In addition to structural data of the Darwin Forest that match those of an extant monsoon forest, the polyxyly found in Cuneumxylon has been considered an important adaptive wood character to avoid water stress. As suggested by the functional anatomy and distribution of living groups (Fahn 1990), the included phloem associated with great amounts of parenchyma could be a strategy of subtropical plants to fight against water stress in arid regions during drought seasons.
During Permian-Triassic biosphere reorganization, aridity of the Earth increased and in the Triassic the Pangea was characterized by strongly seasonal climates in a warm-house period (Parrish et al. 1982, Dubiel et al. 1991, Parrish 1993, Scotese et al. 1999). The spreading of continental climates caused the extension of semiarid belts into middle latitudes and, partly, into high latitudes too (Chumakov and Zharkov 2003). Recently, general circulation models (GCMs) were developed by Sellowood and Valdes (2006) to simulate Mesozoic climatic patterns. In this scheme the Triassic Southwestern Gondwana is modelled as seasonal and winter-wet against the opinion of Robinson (1973) who stated that it was seasonal and summer-wet.
Growth-ring analysis of coniferous wood - assigned by Brea (1997) to Araucarioxylon protoaraucana - was used to evaluate climatic conditions (Fritts 1976, Holmes 1985). Conifer growth rings are narrow and subtly demarcated; they are characterized by a relatively wide zone of large, thin-walled early-wood cells terminated by ten to seventeen thick-walled late-wood cells. The presence of narrow growth rings indicates poor conditions for cell division and expansion during growth season (Creber and Chaloner 1984). In addition, a thin late-wood zone within a ring can be the result of water shortage at the end of the growing season, a sharp photoperiod indicating end of growth season or abrupt leaf shedding.
Therefore, A. protoaraucana growth rings suggest strongly seasonal conditions. The average ring width in these fossil woods is 1.49 mm (0.80-2.53 mm), the narrowest ring is 0.12 mm and the widest ring is 4.44 mm. Mean sensitivity (MS) values range between 0.14 and 0.38. Average MS values (0.30) indicate that the growing environment was stressed and not uniform. Although available data are limited, the complicated growth trends suggest that competition, disturbance events, or climatic stress influenced the growth of trees in the Darwin Forest. Furthermore, Falcon Lang's method (Falcon-Lang 2000a, 2000b) was used to distinguish between evergreen and deciduous species (see Brea et al. 2008). The quantitative growth ring anatomy analysis method (Falcon-Lang 2000a, 200b) indicates that the conifer fossil woods of the Darwin Forest probably belong to an evergreen gymnosperm.
The understorey of the Darwin Forest includes Cladophlebis mesozoica Frenguelli 1947 (Fig. 6, b), Cladophlebis mendozaensis (Geinitz) Frenguelli 1947 (Fig. 6, c), and Cladophlebis kurtzi Frenguelli 1947 (Fig. 6 a) (Brea 2000). Cladophlebis is an extinct genus of ferns characterized by the presence of large sterile bipinnate foliage and it was widely distributed in southern Gondwana during the Mesozoic.

Figure 6: a. Cladophlebis kurtzii Frenguelli (LPPB 12618); b. Cladophlebis mesozoica Frenguelli (LPPB 12614); c. Cladophlebis mendozaensis (Geinitz) Frenguelli (LPPB 12621); d-e. Neocalamites carrerei (Zeiller) Halle (LPPB 12584 and LPPB 12581); f. aff. Nododendron suberosum Artabe et Zamuner (LPPB 12591); g-h. Neocalamostachys arrondoi Brea et Artabe (LPPB 12593 and 12565). Scale bars, e =2 cm, f, g and h= 1cm.
Fossiliferous level FL II found at locality A (Figs. 3 and 4) records another monotypic palaeocommunity dominated by sphenopsids (Brea and Artabe 1999).The sedimentary sequences in which this community is preserved consist of continental clastic units composed of black shales and mudstones resulting from deposition on the flood plains of a fluvial system. This autochtonous taphocenosis is constituted by stems, nodal diaphragms and reproductive structures assigned to Neocalamites carrerei (Zeiller) Halle 1908 (Fig. 6 d-e), aff. Nododendron suberosum Artabe and Zamuner 1991 (Fig. 6 f), and Neocalamostachys arrondoi Brea and Artabe 1999 (Fig. 6 g-h). The stem of Neocalamites and nodal diaphragm of Nododendron found in close association with reproductive Neocalamostachys were probably based on parts of individuals of a single taxon (Brea and Artabe 1999). Because plants growing in wet habitats are hygrophytes, this monotypic horsetail palaeocommunity conformed a waterdependent, bamboo-like thicket with hygrophytic adaptations associated to the flood plains of a fluvial environment (Fig. 7).

Figure 7: Reconstruction of Triassic horsetail landscape in the flood-plain of a fluvial environment. In the background are groups of conifers (drawing by Jorge Gonzalez).
COMPARISON WITH OTHERS TRIASSIC PETRIFIED FORESTS
Three other in situ Triassic petrified forests dominated by corystosperms have been discovered in Argentina, i.e., in the Cortaderita (late Middle Triassic), Ischigualasto (early Late Triassic) and Río Blanco (Late Triassic) formations (Zamuner 1992, Spalletti et al. 1999, Artabe et al. 2001, Artabe et al. 2007b). Presently, the only available structural data are from the Late Triassic corystosperm forest found at La Elcha Mine (Río Blanco Formation, Cuyo Basin). This is an evergreen and monotypic community composed of 150 stumps in life position, and the trees colonized well-drained proximal flood-plain areas, close to channel belts (Artabe et al. 2007b). The permineralized stumps were described and assigned to Elchaxylon zavattieriae (Corystospermaceae) by Artabe and Zamuner (2007).
Vegetation analysis of the La Elcha forest shows that it has a clustered distribution pattern, with a forest density of 727-1504 tree /ha. The deduced height of Elchaxylon and the distribution of class diameters suggest that the canopy in the forest community would have had the majority of specimens ranging 13-21 m. Growth ring analysis indicates that the forest community colonized stressed ecosystems (Artabe et al. 2007b). At very high latitudes, in the central Transantarctic Mountains, another in situ corystosperm riparian forest was found and assigned to the Middle Triassic (Cúneo et al. 2003). The permineralized stumps were described as Jeffersonioxylon Del Fueyo et al. (1995) and later assigned to the Corystospermaceae by Cúneo et al. (2003). This monotypic forest is interpreted as deciduous and the tree density and the basal area are around 274 tree/ha and 20.83 m2/ha respectively. The Antarctic plant has been reconstructed as a tree of ~20-30 m with coniferous-like habit (Cúneo et al. 2003).
Petrified remains of a Late Triassic forest were preserved in the Paraná Basin, Brazil (Pires et al. 2005). This forest was dominated by Sommerxylon spiralosus, a morphotaxon with taxacean affinities (Pires and Guerra Sommer 2004). Growth ring analysis indicates that the climate was not equable but rather with marked seasonal variations. The fossil wood shows a distinctive seasonal pattern related to growth cycles, with extensive favourable and restricted unfavourable growth periods. The external factors that affected the cycles were mainly related to cyclic restrictions of water supply and irregular changes in environmental conditions, probably linked to occasional droughts during the growing season (Pires et al. 2005).
The Petrified Forest National Park in Arizona is a better known Late Triassic fossil forest in the southwestern United States (Ash and Creber 1992). This fossil forest lies within the Chinle Formation and is mainly dominated by Woodworthia arizonica and Schilderia adamanica (Creber and Ash 2004). The fossil woods of this forest do not show annual growth rings but contain irregular growth interruptions similar to those found in trees nowadays growing in humid tropics. These interruptions could be due to endogenous hormonal effects or to occasional local variations in water supply (Ash and Creber 1992).
SUMMARY
Two late Middle Triassic palaeocommunities were preserved in the Paramillo Formation, at Agua de la Zorra, northwestern Cuyo Basin, in southwestern Gondwana, i.e., the Darwin Forest associated with banks of highly sinuous fluvial systems, and second including fossil horsetails that grew on flood plains of fluvial systems. The Darwin Forest was a mixed forest, integrated by corystosperms (Cuneumxylon spallettii) and conifers (Araucarioxylon protoaraucana). Species distribution shows that in some places corystosperms and conifers are intermingled, whereas in others corystosperms and conifers appear to be aggregated into cohesive monotypic groups. The canopy is formed by two arboreal strata with a small number of emergent; the understorey with ferns.
The included phloem and associated parenchyma present in Cuneumxylon spallettii may be an important adaptative strategy to avoid water stress. Thus, polyxyly as a functional anatomy characters correlates with a seasonal climatic regime. Morpho-functional analysis, structural parameters, biomass and basal stand area of the Darwin Forest allows classifying it as a mainly dry evergreen subtropical seasonal forest. The anatomy of the growth rings of Araucarioxylon protoaraucana suggests strongly seasonal conditions. Moreover, growth ring analysis indicates that this species was an evergreen gymnosperm. The second palaeocommunity integrated by sphenopsids shows herbaceous-arbustive vegetation with hygrophytic adaptations. A fluvial flood-plain in an open landscape was the environment in which this fossil vegetation developed.
ACKNOWLEDGEMENTS
The authors would like to express their thanks to Guillermo Ottone, Greg Retallack and one anonymous reviewer for their critical and constructive comments on an earlier version of this paper. We would like to thank the financial support provided by the Consejo Nacional de Investigaciones Científicas y Técnicas (Project PIP 5430) and Universidad Nacional de La Plata (11/N535). The charts of the Triassic vegetation reconstruction were made by Jorge Gonzalez.
WORKS CITED IN THE TEXT
1. Artabe, A.E., and Brea, M. 2003. A New Approach to Corystospermales based on petrified stems from the Triassic of Argentina. Alcheringa 27: 209-229. [ Links ]
2. Artabe, A.E., and Zamuner, A.E. 1991. Una nueva equisetal del Triásico de Cacheuta, Argentina, con estructura interna preservada. Ameghiniana 28: 287-294. [ Links ]
3. Artabe, A.E., and Zamuner, A.E. 2007. Elchaxylon, a new corystosperm based on permineralized stems from the late Triassic of Argentina. Alcheringa 31: 85-96. [ Links ]
4. Artabe, A.E., Morel, E.M. and Spalletti, L.A. 2001. Paleoecología de las floras triásicas argentinas. In Artabe, A.E., Morel, E.M. and Zamuner, A.E. (eds.) El Sistema Triásico en la Argentina. Fundación Museo de La Plata Fracisco P. Moreno: 199-225, La Plata. [ Links ]
5. Artabe, A.E., Morel, E.M. and Spalletti, L.A. 2003. Caracterización de las provincias fitogeográficas triásicas del Gondwana extratropical. Ameghiniana 40: 387-405. [ Links ]
6. Artabe, A.E., Morel, E.M and Ganuza, D.G. 2007 a. Las floras trásicas de La Argentina. In Archangelsky, S., Sánchez, T. and Tonni, E.P. (eds.) Asociación Paleontológica Argentina, Publicación Especial 11, Ameghiniana 50° aniversario: 75-86. [ Links ]
7. Artabe, A.E., Spalletti, L.A., Brea, M., Iglesias, A., Morel, E.M. and Ganuza, D.G. 2007 b. Structure of a corystosperm fossil forest from the Late Triassic of Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology 243: 451-470. [ Links ]
8. Ash, S.R. and Creber, G.T. 1992. Palaeoclimatic interpretation of the wood structures of the trees in the Chinle Formation (Upper Triassic), petrified forest National Park, Arizona, USA. Palaeeogeography, Palaeoclimatology, Palaeecology 96: 299-317. [ Links ]
9. Avé-Lallemant, H. 1891. Estudios mineros en la provincia de Mendoza. La parte septemtrional de la Sierra de Uspallata. Academia Nacional de Ciencias, Boletín 12: 131-176, Córdoba. [ Links ]
10. Brea, M., 1995. Estudio de la paleoflora de la secuencia triásica de Agua de la Zorra, provincia de Mendoza. Ph. D. Thesis N° 642, Facultad de Ciencias. Naturales y Museo, Universidad Nacional de La Plata, Argentina (unpublished), 202 p. [ Links ]
11. Brea, M. 1997. Una nueva especie fósil del género Araucarioxylon Kraus 1870 emend. Maheshwari 1972 del Triásico de Agua de la Zorra, Uspallata, Mendoza, Argentina. Ameghiniana 34: 485-496. [ Links ]
12. Brea, M. 2000. Paleoflora triásica de Agua de la Zorra, Uspallata, provincia de Mendoza, Argentina: Lycophyta y Filicophyta. Ameghiniana37: 199-204. [ Links ]
13. Brea, M., and Artabe, A.E. 1999. Apocalamitaceae (Sphenophyta) triásicas de la Formación Paramillo, Agua de la Zorra, provincia de Mendoza, Argentina. Ameghiniana 36: 389-400. [ Links ]
14. Brea, M., Artabe, A.E. and Spalletti, L.A. 2005. Paleovegetation studies and growth-ring analysis of a mixed Middle Triassic forest from Argentina. Abstracts Gondwana 12: 77. [ Links ]
15. Brea, M., Artabe, A.E. and Spalletti, L.A. 2008. Ecological reconstruction of a mixed Middle Triassic forest from Argentina. Alcheringa 32: 365-393. [ Links ]
16. Brown, S. 1997. Estimating biomass and biomass change in tropical forests. FAO Forestry Paper 134, Food and Agriculture Organization of the United Nations. 49 p., Rome. [ Links ]
17. Burmeister, H. 1861. Reise durch die La Plata-Staaten, mit besonderer Rücksicht auf die physische Beschaffenheit und den Culturzustand der Argentinischen Republik. Ausgeführt in den Jahren 1857, 1858, 1859 und 1860. Erster Band. H.W. Schimdt, v. 1, 503 p., v. 2, 538 p., Halle. [ Links ]
18. Chumakov, N.M. and Zharkov, M.A. 2003. Climate during the Permian-Triassic biosphere reorganizations. Article 2. Climate of thr Late Permian and Early Triassic: General inferences. Stratigraphy and Geological Correlation 11(4): 361-375. [ Links ]
19. Conwentz, H. 1885. Sobre algunos árboles fósiles del Río Negro. Academia Nacional de Ciencias Boletín , 7: 435-456, Córdoba. [ Links ]
20. Creber, G.T. and Ash, R. 2004. The Late Triassic Schilderia adamanica and Woodworthia arizonica trees of the petrified forest National Park, Arizona, USA. Palaeontology 47: 21-38. [ Links ]
21. Creber, G.T. and Chaloner, W.G. 1984. Influence of environmental factors on the wood structure of living and fossil trees. Botanical Review 50: 357-448. [ Links ]
22. Cúneo, N.R., Taylor, E.L., Taylor, T.N. and Kring, M. 2003. In situ fossil forest from the upper Fremouw Formation (Triassic) of Antarctica: paleoenvironmental setting and paleoclimate analysis. Palaeogeography, Palaeoclimatololgy, Palaeoecology 197: 239-261. [ Links ]
23. Darwin, C. 1839a. Narrative of the surveying voyages of His Majesty's Ships Adventure and Beagle between the years 1826 and 1836, describing their examination of the southern shores of South America, and the Beagle's circumnavigation of the globe. Third Volume: Journal and remarks, 1832-1836, Henry Colburn, 615 p. London. [ Links ]
24. Darwin, C. 1839b. Journal of researches into the geology by HMS Beagle under the command of captain FitzRoy, R.N., from 1832 to 1836, Henry Colburn, 615 p., London. [ Links ]
25. Darwin, C. 1845. Journal of researches into the natural history and geology of countries visited during the voyage of HMS Beagle round the world, under the command of Capt. FitzRoy, R.N. Second Edition, corrected, with additions. John Murray, 519 p., London. [ Links ]
26. Darwin, Ch. 1846. Geological observations on South America. Being the third part of the Geology of the voyage of the Beagle, during the years 1832 to 1836. Smith, Elder and Co. 280 p., London. [ Links ]
27. Del Fueyo, G., Taylor, E.L., Taylor, T.N. and Cúneo, N.R. 1995. Triassic Wood from the Gordon Valley, central transantarctic Mountains, Antarctica. IAWA Journal 16: 111-126. [ Links ]
28. Doering, A. 1882. Informe official de la Comisión Científica agregada al Estado Mayor General de la Expedición al Río negro (Patagonia) realizadas en los meses de Abril, Mayo y Junio de 1879, bajo las órdenes del General D. Julio A. Roca. Tercera Parte. Geología. Ostwald & Martinez, 299-530, Buenos Aires. [ Links ]
29. Dubiel, R.F., Parrish, J.T., Parrish, J.M. and Good, S.C. 1991. The Pagaean megamonsoon - evidence from the Upper Triassic Chinle Formation, Colorado Plateau. Palaios 6: 347-370. [ Links ]
30. Du Toit, A.L. 1927. A geological comparison of South America with South Africa. Carnegie Institution Publication 381, 158 p., Washington. [ Links ]
31. Falcon-Lang, H.J. 2000a. A method to distinguish between woods produced by evergreen and deciduous coniferopsids on the basis of growth ring anatomy: a new palaeoecological tool. Palaeontology 43: 785-793. [ Links ]
32. Falcon-Lang, H.J. 2000b. The relationship between leaf longevity and growth ring markedness in modern conifer woods and its implications for palaeoclimatic studies. Palaeogeogrphy, Palaeoclimatololgy, Palaeoecololy 160: 317- 328. [ Links ]
33. Fahn, A. 1990. Plant anatomy. Pergamon Press. 588 p., Oxford. [ Links ]
34. Frenguelli, J. 1947. El género Cladophlebis y sus representantes en Argentina. Gaea 7: 159-369. [ Links ]
35. Fritts, H.C. 1976. Tree rings and climate. Academic Press. 567 p., London. [ Links ]
36. Groeber, P. 1939. Mapa geológico de Mendoza. Physis 14: 171-220. [ Links ]
37. Halle, T.G. 1908. Zur Kenntnis der Mesozoischen equisetales Schwedens. Kungliga. Svenska vetenskaps akademie Handlingar 43: 3-42. [ Links ]
38. Harrington, H.J. 1971. Descripción geológica de la hoja 22c, "Ramplón" provincias de Mendoza y San Juan. Carta Geológica-Económica de la República Argentina Escala 1:200.000. Ministerio de Industria y Minería, Subsecretaria de Minería, Dirección de Geología y Minería, Buenos Aires, Boletín 114: 87 p. [ Links ]
39. Holmes, R.L. 1985. Computer-assisted quality control in tree-ring dating and measurement. A users manual for program COFECHA. Laboratory of tree-ring Research: 2-23. [ Links ]
40. Kokogian, D., and Mancilla, O. 1989. Análisis estratigráfico y secuencial de la Cuenca Cuyana. In Chebli, G. and Spalletti, L. (eds.) Cuencas Sedimentarias Argentinas. Universidad Nacional de Tucumán, Serie Correlación Geológica 6: 169-201. [ Links ]
41. Kokogián, D., Fernández Seveso, F., and Mosquera, A. 1993. Las secuencias sedimentarias triásicas. In Ramos, V.A. (ed.), Relatorio Geología y Recursos Naturales de Mendoza. 12° Congreso Geológico Argentino y 2° Congreso de Exploración de Hidrocarburos 1(7): 65-78. [ Links ]
42. Kurtz, F. 1921. Atlas de plantas fósiles de la República Argentina. Academia Nacional de Ciencias, Actas 7 : 129-153, Córdoba. [ Links ]
43. Linares, E., and González, R. 1990. Catálogo de edades radimétricas de la República Argentina 1957-1987. Asociación Geológica Argentina, Serie B (Didáctica y Complementaria) 19: 628 p. [ Links ]
44. Massabie, A.C. 1985. Filón capa Paramillo de Uspallata, su caracterización geológica y edad, Paramillo de Uspallata, Mendoza. Primeras Jornadas sobre Geología de Precordillera, Actas 1: 325-330. [ Links ]
45. Massabie, A.C., Rapalini, A.E. and Soto, J.L. 1985. Estratigrafía del Cerro Los Colorados, Paramillo de Uspallata, Mendoza. Primeras Jornadas sobre Geología de Precordillera, Actas 1: 71-76. [ Links ]
46. Morel, E.M., Artabe, A.E., and Spalletti, L.A. 2003. The Triassic floras of Argentina: Biostratigraphy, Floristic events and comparison with other areas of Gondwana and Laurasia. Alcheringa 27: 231-243. [ Links ]
47. Murphy, P.G. and Lugo, A.E. 1986. Structure and biomass of a subtropical dry forest in Puerto Rico. Biotropica 18: 89-96. [ Links ]
48. Ottone, E.G. 2005. The history of palaeobotany in Argentina during the 19th century. In Bowden, A.J., Burek, C.V. and Wilding, R. (eds.) History of Paleobotany: Selected Essays. Geological Society, Special Publications 241: 281-294. [ Links ]
49. Parrish, J.T. 1993. Climate of the supercontinent Pangea. Journal of Geology 101: 215-233. [ Links ]
50. Parrish, J.T., Ziegler, A.M., and Scotese, C.R. 1982. Rainfall patterns and the distribution of coals and evaporates in the Mesozoic and Cenozoic. Palaeogeography, Palaeoclimatology, Palaeoecology 40: 67-101. [ Links ]
51. Pires, E.T. and Guerra Sommer, M. 2004. Sommerexylon spiralosus from Upper Triassic in southernmost Paraná Basin (Brazil): a new taxon with taxacean affinity. Anais da Academia Brasileira de Ciências 76: 595-609. [ Links ]
52. Pires, E.T., Guerra Sommer, M. and Dos Santos Scherer, C.M. 2005. Late Triassic climate in southernmost Parana Basin (Brazil): evidence from dendrochronological data. Journal of South American Earth Sciences 18: 213-221. [ Links ]
53. Poma, S., Quenardelle, S. and Koukharsky, M. 2004. Un flujo piroclástico del Triásico en la Formación Paramillo, provincia de Mendoza. 7º Congreso Argentino de Mineralogía y Metalogenia: 385-390. [ Links ]
54. Ramos, V.A. 1993. El magmatismo Triásico-Jurásico de intraplaca. In Ramos, V.A. (ed.) Relatorio Geología y Recursos Naturales de Mendoza. 12 Congreso Geológico Argentino y 2 Congreso de Exploración de Hidrocarburos 1(8): 79-86. [ Links ]
55. Ramos, V.A., and Kay, S.M. 1991. Triassic rifting and associated basalts in the Cuyo Basin, central Argentina. In Harmon, R.S. and Rapela, C.W. (eds.) Andean magmatism and its tectonic setting. Geological Society of America, Boulder, Special Paper 265: 79-91. [ Links ]
56. Robinson, P.L. 1973. Palaeoclimatology and continental Drift. Implications of Continental Drift in the Earth Sciences 46: 451-476. [ Links ]
57. Rusconi, C. 1941. "Bosques petrificados" de Mendoza. Anales de la Sociedad Científica Argentina 132: 80-95. [ Links ]
58. Sellwood, B.W., and Valdes, P.J. Mesozoic climates: General circulation models and the rock record. Sedimentary Geology 190: 269-287. [ Links ]
59. Scotese, C.R., Boucot, A.J. and McKerrow, W.S. 1999. Gondwanan palaeogeography and palaeoclimatology. Journal of African Earth Sciences 28: 99-114. [ Links ]
60. Spalletti, L.A., Artabe, A.E. and Morel, E.M. 2003. Geological factors and evolution of Southwestern Gondwana Triassic Plants. Gondwana Research 6: 119-134. [ Links ]
61. Spalletti, L.A., Artabe, A.E., Morel, E.M. and Brea, M. 1999. Biozonación paleoflorística y cronoestratigrafía del Triásico Argentino. Ameghiniana 36: 419 -451. [ Links ]
62. Stappenbeck, R. 1910. La Precordillera de San Juan y Mendoza. Anales del Ministerio de Agricultura, sección Geología, Mineralogía y Minería, 4 (3): 3-183, Buenos Aires. [ Links ]
63. Stelzner, A. 1885. Beiträge zur Geologie der Argentinischen Republik und des Angrenzender, zwischen dem 32. und 33°. S. gelegenen Theiles der chilenischen Cordillere. In Stelzner, A. (ed.) Beiträge zur Geologie und Palaeontologie der Argentinschen Republik, 1. Geologischer Theil. Palaeontograhica, Supplementbände 3, 29: 1-329. [ Links ]
64. Stewart, W.N. 1983. Paleobotany and the evolution of plants. Cambridge University Press. 405 p. [ Links ]
65. Stipanicic, P.N. 2001. Antecedentes Geológicos y Paleontológicos. In Artabe, A.E., Morel, E.M. and Zamuner, A.B. (eds.) El Sistema Triásico en la Argentina. Fundación Museo de La Plata Francisco Pascasio Moreno: 1-21, La Plata. [ Links ]
66. Stipanicic, P.N. and Marsicano, C.A. 2002. Léxico estratigráfico de la Argentina, Volumen VIII: Triásico. Asociación Geológica Argentina, Serie B (Didáctica y Complementaria) 26, 370 p. [ Links ]
67. Stipanicic, P.N., Herbst, R. and Bonetti, M.I.R. 1996. Floras Triásicas. Academia Nacional de Ciencias, Actas 11: 127-184, Córdoba. [ Links ]
68. Strelkov, E.E., and Álvarez, L.A. 1984. Análisis estratigráfico y evolutivo de la Cuenca Triásica Mendocina-Sanjuanina. 9° Congreso Geológico Argentino (Bariloche), Actas 3: 115-130. [ Links ]
69. Willis, K.J. and McElwain, J.C. 2002. The evolution of plants. Oxford University Press, 378 p. [ Links ]
70. Windhausen, H. 1941. El "Rético" del Paramillo de Uspallata (Mendoza), PhD Thesis, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, (unpublished), 28, 90 p., La Plata. [ Links ]
71. Zamuner, A.B. 1992. Estudio de una tafoflora de la localidad tipo, de la Formación Ischigualasto (Neotrias), provincia de San Juan. Ph.D. Thesis, Facultad de Ciencias Naurales y Museo, Universidad Nacional de La Plata, (unpublished), 583, 97 p., La Plata [ Links ]
72. Zamuner, A.B., Zavattieri, A.M., Artabe, A.E. and Morel, E.M. 2001. Paleobotánica. In Artabe, A.E., Morel, E.M. and Zamuner, A.B. (eds.) El Sistema Triásico en la Argentina. Fundación Museo de La Plata Francisco P. Moreno : 143-184. La Plata. [ Links ]
Recibido: 8 de agosto de 2008
Aceptado: 28 de octubre de 2008

