










Services on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista de la Asociación Geológica Argentina
Print version ISSN 0004-4822On-line version ISSN 1851-8249
Rev. Asoc. Geol. Argent. vol.64 no.1 Buenos Aires Apr. 2009
Darwin's great patagonian tertiary formation at the mouth of the río Santa Cruz: a reappraisal
Ana Parras and Miguel Griffin
Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa y CONICET, Santa Rosa, La Pampa. Emails: aparras@exactas.unlpam.edu.ar, miguelgriffin@aol.com
ABSTRACT
The classical section described by Darwin at the mouth of the Río Santa Cruz was a landmark in the later stratigraphy of the marine Cenozoic of Patagonia. Sedimentological and paleontological studies of rocks from the Monte León Formation at the Darwin and Mount Entrance sections - mouth of the Río Santa Cruz - suggest that they were deposited in an early Miocene inner shelf to subtidal - or even intertidal at the top - environment. The base of the sections is covered and the top is unconformably overlain by Quaternary deposits. The invertebrate fauna (mainly mollusks) mentioned herein - the earliest of which were collected by Darwin and described by Sowerby - were collected from the two sections with a detailed stratigraphic control. It includes brachiopods, echinoderms, scaphopods, bivalves, and gastropods. The similarity of this fauna with that one from the sections at Restinga Norte, Cabeza de León, Las Cuevas, Yegua Quemada and Monte Observación, all south of the study area, allow correlation with them. In addition to the equivalence of the faunas, sedimentological features at all these localities are also the same. Therefore, all the shell beds involved should be referred to the Punta Entrada Member of the Monte León Formation. The Monte Observación Member should be restricted to its original use and included as a member at the base of the overlying Santa Cruz Formation, outcrops at Cerro Monte León and Cerro Observación.
Keywords: Darwin Section; Mount Entrance Section; Mouth of Santa Cruz River; Paleoenvironments; Invertebrates; Patagonia.
RESUMEN: La Gran Formación Terciaria Patagónica de Darwin en la desembocadura del río Santa Cruz: una revaluación. La sección clásica descripta por Darwin en la desembocadura del río Santa Cruz fue un hito en el posterior desarrollo de la estratigrafía del Cenozoico marino de Patagonia. Los estudios sedimentológicos y paleontológicos realizados en sedimentitas de la Formación Monte León, en las secciones Darwin y Monte Entrada, en el área de la desembocadura del río Santa Cruz, sugieren que las mismas fueron depositadas en un ambiente de plataforma interna a submareal en la base, hasta intermareal en el techo, durante el Mioceno temprano. La base de las secciones se encuentra cubierta y su techo es cortado en forma discordante por depósitos del Cuaternario. La fauna de invertebrados (mayormente moluscos) mencionada en este trabajo, cuyos primeros representantes fueran coleccionados por Darwin y descriptos por Sowerby, fue colectada con un detallado control estratigráfico a lo largo de las dos secciones. La misma incluye braquiópodos, equinodermos, escafópodos, bivalvos y gasterópodos. La similitud en la composición taxonómica, como así también en las características sedimentológicas, entre estas secciones y aquellas ubicadas más al sur como son Restinga Norte, Cabeza de León, Las Cuevas, Yegua Quemada y Monte Observación, sugiere que las mismas se correlacionan lateralmente. Por lo tanto estos niveles deben ser referidos al Miembro Punta Entrada de la formación Monte León, restringiéndose el Miembro Monte Observación a los niveles basales de la suprayacente Formación Santa Cruz, aflorantes en Cerro Monte León y Cerro Observación.
Palabras clave: Sección Darwin; Sección Monte Entrada; Desembocadura del río Santa Cruz; Paleoambientes; Invertebrados; Patagonia.
INTRODUCTION
On April 13th 1834, HMS Beagle anchored within the mouth of the Río Santa Cruz, a few miles upstream from the Atlantic Ocean. During his numerous excursions ashore, Darwin made geological observations on the cliffs that rise along the southern bank of the river. He also assembled a collection of invertebrate marine fossils later described by Sowerby (1846). Thus, the locality from where this material was collected became the type locality of most of the taxa described by Sowerby. The main purposes of this work are to pinpoint and re-describe as accurately as possible the locality at the mouth of the Río Santa Cruz where Charles Darwin (1846, p. 112-113) described the geological section of marine Tertiary rocks, and discuss the possible paleoenvironmental conditions in which it was deposited. This should aid in the formulation of more precise correlations of the beds mentioned by Darwin with others exposed in the area, and thus clarify the meaning of the formal names proposed by later authors for the rocks involved. Darwin (1839, 1846) himself included these rocks in his "great Patagonian tertiary formation" or "Patagonian Tertiary Formation", a unit subject to very different interpretations ever since. According to his description of the beds exposed and after a survey of the area to try to identify the exact location of his section, we were able to identify it on the southern bank of the Río Santa Cruz, a short distance away from its mouth in the Atlantic Ocean.
The rocks exposed in this area are now included in the lithostratigraphic unit known as Monte León Formation (Bertels 1970) and deemed to be of late Oligocene-early Miocene age (Malumián 1999, 2002, Barreda and Palalmarczuk 2000). This unit is exposed along the coast of Patagonia between Puerto San Julián and Coy Inlet, at the mouth of the Río Coyle (or Coig) (Fig. 1). It comprises mainly sandstones and siltstones with a high content of pyroclastic material, and carries numerous shell beds.
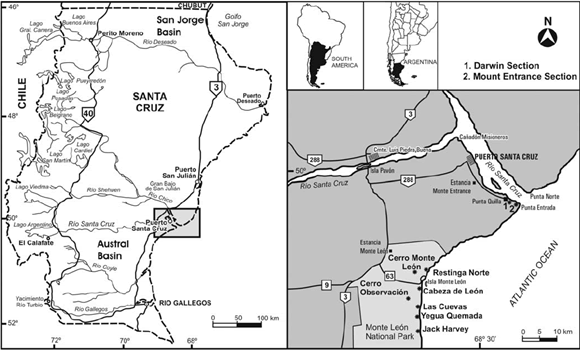
Figure 1: Location map of the Darwin Section and the Mount Entrance Section at the mouth of the Río Santa Cruz.
In order to clarify the exact location of Darwin's section, we surveyed the coast along the southern bank of the river. Based on his writings (1846, p. 112-113) it became clear the section is the one lying just east of the present port of Punta Quilla (S 50° 07' 39.9"; W 68° 24' 10.04"), a point where the marine Tertiary rocks are best exposed (Fig. 2), and where they best meet the description and measurements he offers in his "Geological Observations…".

Figure 2: Monte León Formation at the mouth of the Río Santa Cruz, Darwin Section, showing Darwin's strata and facies associations described herein.
As there has been some confusion as to the exact location of Darwin's locality, we also measured and described the section lying further east, approximately one kilometer away from Mount Entrance, at the very mouth of the river (S 50° 08' 32.2"; W 68° 22' 23.6"). The two localities lie within the present Department of Corpen Aike, in the province of Santa Cruz (Fig. 1). They both carry a rich fauna of exceptionally well preserved mollusks that can be counted among the best in southern South America. They have been mentioned in the literature (Becker 1964, Bertels 1970, 1980, Di Paola and Marchese 1973, Olivera 1990, del Río and Camacho 1998, Barreda and Palamarczuk 2000), but detailed descriptions that could enable correlation and paleoenvironmental reconstructions are still lacking.
Study of these two stratigraphic sections allowed recognition of sedimentary facies, and hence the dominant deposition mechanisms could be interpreted. These facies were grouped into three facies associations and a depositional model thus emerged. Detailed analysis of the taxonomic composition of the fossil assemblages revealed the presence of at least 38 species of bivalves and 70 species of gastropods, in addition to scaphopods, echinoderms, and brachiopods. Diffe-rences among the different shell beds suggest a complex history. The different fossil concentrations were originated in different settings, ranging from inner shelf to nearshore subtidal environments - and even intertidal, representing deposition during the early Miocene transgression in Patagonia.
GEOLOGICAL FRAMEWORK
The surface of Patagonia is covered mainly by marine rocks deposited during the Atlantic transgressions that occurred during the late Cretaceous, Paleogene and Neogene. These are intercalated with non marine rocks and non deposition and erosion intervals (Malumián 1999). One of these transgressions is commonly referred to as "Patagoniano" and occurred during the high sea level period that began in the latest Oligocene and ended in the early middle Miocene. The final phases of this transgression are coincident with the Neogene climatic optimum. It covered the largest extent of land among the various Cenozoic transgressions that occurred in Patagonia (Malumián 2002). This transgression deposited rocks containing the Paleogene/Neogene boundary in Patagonia and in the Cordillera sector of southern Chile. They are presently included in the San Julián and Monte León formations along the coast of Santa Cruz and in the Centinela Formation in the Andean region of this province. Exposures of these units lie within the Austral Basin. Further north, the rocks deposited during the Patagonian transgression are included in the Chenque and Gaiman formations, in the San Jorge Basin. In southern Chile, these rocks are known as Guadal Formation, and are exposed south of Lago General Carrera.
The basement rocks underlying the Cenozoic deposits are a Jurassic lava-pyroclastic complex known as Bahía Laura Group, including the Chon Aike Formation (ignimbrites and scarce rhyolitic lavas and tuffs) and the La Matilde Formation (tuffs, claystones, siltstones and coal beds). These two units are interdigitated and unconformably overlain, in the San Julián area, by the San Julián Formation. This late Oligocene unit includes predominantly marine clastic -sandstone to siltstone - rocks (Parras and Casadío 2005, Parras et al. 2008).
Deposition of the probably unconformably overlying Monte León Formation occurred during the late Oligocene and early Miocene. This unit includes siltstones, sandstones and tuffs (Bertels 1970, 1980), and is overlain by the early to middle Miocene continental rocks of the Santa Cruz Formation. Unconformably overlying these are variably thick Quaternary deposits of the Shingle Formation described by Darwin too (Darwin 1839, 1846).
The earliest record of marine rocks in Patagonia was by d'Orbigny (1842 a, b), who explored this part of South America during the years 1826-1833. D'Orbigny himself never went ashore further south than the coast of northern Patagonia. However, he named all the deposits that constitute the cliffs along the coast between Carmen de Patagones (Río Negro) and the Strait of Magellan as "Tertiaire Patagonien". Although he did not inspect the cliffs further south, he did have available the large oysters from San Julián, which he believed were the same ones as he had collected in Río Negro and further north in Entre Ríos. Darwin was aware of this, and therefore used an equivalent name for these rocks. However, he also noticed that there were differences between the faunas from the southern and northern areas in which his "Patagonian Tertiary Formation" was exposed.
After these initial surveys the area remained largely unexplored from a geological point of view. A Chilean settlement near the present town of Puerto Santa Cruz was headed by Vidal Gormaz, who collected a few fossils from the cliffs surrounding the settlement and sent them to R.A. Philippi, then at the Museo Nacional de Historia Natural in Santiago de Chile. These fossils were described by Philippi (1887), but hardly any comments were made on the geology of the area, except to note that the specimens came from the Tertiary marine beds there exposed. It was not until the final years of the 19th Century that this region was again surveyed and its geology described in more detail. This was due mainly to three early explorers who visited the mouth of the Río Santa Cruz and assembled the data that were then - and for many years - used to formulate the first stratigraphic divisions and the earliest paleogeographic and paleobiogeographic interpretations about the rocks in the area.
In the first place should be mentioned Carlos Ameghino, brother of Florentino Ameghino-the famed Argentine paleontologist. C. Ameghino surveyed the area and collected fossils that were subsequently studied by his brother. Although he did not publish his results extensively, his keen and detailed geological observations were later used by Florentino to formulate the earliest biostratigraphic scheme of the Tertiaire Patagonien or the Great Patagonian Formation. Thus, based on the fossil content (mainly mollusks) of these marine beds at the mouth of the Río Santa Cruz and in the surroundings of Puerto San Julián, F. Ameghino (1898) subdivided the marine beds he had named (Ameghino 1896) "Formación Patagónica" into two chronostratigraphic units he called "Juliense" and "Leonense". Overlying these (Ameghino 1900-1902) he recognized the continental "Formación Santacruceña", at the base of which he could distinguish a marine stage he called "Superpatagónico". The faunas of marine invertebrates that were the basis of Ameghino's subdivisions were collected by Carlos and described by Maurice Cossmann (1899) and Hermann von Ihering (1897, 1899, 1902, 1907, 1914).
During the same years, the area was also visited by J. B. Hatcher, geologist in charge of the Princeton Expedition to Southern Patagonia. Hatcher's observations were published in the Reports and Geology of this expedition (Hatcher 1903) and in a series of papers in the American Journal of Science (Hatcher 1897, 1900). The large collection of Tertiary marine fossils that he assembled was described by Ortmann (1902). Hatcher's conclusions on the geology of the area were different from those of Ameghino. Hatcher did not recognize the existence of the "Superpatagónico", as the fauna described by Ortmann was essentially the same one as the one in the underlying "Formación Patagónica". This initial controversy pervaded all further work on these units and has never been satisfactorily resolved. The reasons for this are varied and include imperfect knowledge on the sedimentology and stratigraphy at the localities involved, inaccurate location of some of these localities, outdated taxonomic placement of some of the fossil material on which the biostratigraphy was based, among other.
At about the same time, C. Bicego was entrusted by H. von Ihering with the task of assembling further collections at the localities initially visited by C. Ameghino. His large collection of mollusks was described by von Ihering, but Bicego himself did not provide further geological data beyond that mentioned by Ameghino.
Wichmann (1922), Windhausen (1931) and Feruglio (1949) published detailed accounts on the geological features of Patagonia, including the marine Cenozoic rocks. Their detailed sections and geological observations are generally easy to corroborate in the field. However, interpretations largely reflect the ideas of Ameghino and von Ihering.
After these initial studies, the use of "Patagonian Formation" for the Cenozoic marine rocks in southern South America was widespread, although the exact chronostratigraphic and lithostratigraphic meaning of this formational name remained somewhat unstable. Accounts on the uses and controversies attached to such usage can be found in Camacho (1974, 1979), Zinsmeister (1981), Legarreta and Uliana (1994), and Malumián (1999, 2002).
At the mouth of the Río Santa Cruz, these rocks were formally named Monte León Formation by Bertels (1970). The unit includes mainly yellowish-grey siltstones and fine sandstones with a high pyroclastic content and intercalated shell beds. These rocks record a relative sea level increase characterized by the greatest extent of the Patagonian Cenozoic sea, thus allowing input of corrosive Antarctic water onto the Argentine continental shelf (Malumián 2002). Most authors agree that in the study area, i.e. the mouth of the Río Santa Cruz, these rocks were deposited in a shallow marine environment, under littoral and neritic conditions (Panza et al. 1995, Barreda and Palamarczuk 2000). Bertels (1980) suggested outer shelf conditions for the lower part of this formation. Panza et al. (1995) stated that towards the top of the unit conditions became progressively shallower, ending in a marshy environment just before the onset of the continental environment in which the overlying Santa Cruz Formation was deposited. The abundant tuff beds and cineritic material in these rocks led Bertels (1970) and Di Paola and Marchese (1973) to suggest they originated in coetaneous volcanic events in the Cordillera. The ash was probably transported towards the coast by the western winds and deposited in an environment that was protected from the action of marine currents.
Although discrepancies still exist concerning the age of the Monte León Formation, most authors refer it to the late Oligocene (Bertels 1970) or late Oligocene - early Miocene (Malumián 1999, 2002). Yet, Náñez (1988) restricted it to the Oligocene (middle section of P19 Zone and top of P21 Zone) while suggesting that the top may reach into the early Miocene. This was later confirmed by 40Ar/39Ar ages published by Fleagle et al. (1995). Their sample came from the top of the unit at Cerro Monte León and it yielded an age of 19.33 Ma (Burdigalian, early Miocene). More recently Barreda and Palamarczuk (2000) considered that in the study area the Monte León Formation should be restricted to the early Miocene on the basis of its palynomorph content, an age we agree with herein.
METHODOLOGY
Just east of Punta Quilla (Darwin section; S50°07'39.9"; W68°24'10.04") and at Mount Entrance (S50°08'32.2"; W68° 22'23.6") we measured detailed (1:10) stratigraphic-sedimentologic sections (Fig. 3). We recorded bed geometry, lithology, grain size, sedimentary structures, and color. Each bed was sampled and fossils and trace fossils were photographed and collected. Taphonomic observations were recorded too. Close-packing and size-sorting were estimated following the semi-quantitative categories for coarse bioclastic fabrics of Kidwell and Holland (1991). Terminology for geometry of concentrations and orientation of bioclasts follows Kidwell et al. (1986). Sedimentary facies were distinguished and grouped in facies associations for paleoenvironmental analysis. The invertebrate fauna collected was identified taxonomically. The different beds mentioned by Darwin (1846, p. 112-113) were identified as was the exact provenance of the type material of the species described by Sowerby (1846).


Figure 3: Studied stratigraphic sections showing identified facies associations.
SEDIMENTOLOGY
Sections
The type section of the Monte León Formation lies on the right bank of the Río Santa Cruz, one kilometer upstream of its mouth into the Atlantic Ocean (Bertels 1970, p. 499). However, Bertels (1970) did not describe or illustrate her section until a decade later (Bertels 1980, p. 214-217), when she also defined the Punta Entrada Member of the Monte León Formation. This locality is precisely where our Mount Entrance section lies. According to her description (Bertels 1980) the section measures there 103.8 m, but this could not be confirmed in the field by us, as the bottom of the section is completely covered by modern foothill deposits and only 60 m are exposed. Therefore, the base of Bertels' section - as inferred from her description - must have been measured at Darwin's section, where the exposure is complete, although the top part is here extremely difficult to reach. Riggi (1978, p. 166) measured 89 m but it remains unclear exactly where his section is (his Mount Entrance section lies "on the right bank of the Río Santa Cruz and its mouth in the ocean"). Barreda and Palamarczuk (2000) measured 80 m south of the mouth of the river, but most other published sections are compounds based on partial sections measured at various points ranging between south of the mouth of the river to 30 km upstream along the right bank (Olivera 1990, del Río and Camacho 1998). Other authors based their stratigraphic framework on unpublished data (Becker 1964) or previous work by others (Di Paola and Marchese 1973).
According to Darwin's description, the section mentioned by him is the one just east of Punta Quilla (our Darwin Section, Figs. 2 and 3). This is where the cliff is best exposed and where the description and measurements provided by Darwin himself best agree with field observations. In order to refer the beds described by Darwin to the type section, we provide herein a description of both, even if lithology is very similar - as is fossil content. The Darwin Section is more complete because the lower beds lie exposed at low tide. However, access to the topmost beds is extremely difficult. This is probably the reason why the section at Mount Entrance (and nearby) has been better studied.
a) Darwin Section (S50º07'39.9"; W68º 24'10.04")
0-1.2 m: 1.2 m Grey tuffaceous clayish siltstones with abundant fossils and numerous Ophiomorpha isp. Dominant species is "Turritella" patagonica. It appears in clump-geometry packets. Also recorded are Nucula sp., Limopsis insolita, Cucullaea alta, Crassostrea? hatcheri, Fasciculicardia patagonica, Pleuromeris cruzensis, Dosinia laeviuscula, Ameghinomya darwini. Close-packing within these clumps is dense/loose (25%), size-sorting is poor, orientation chaotic, disarticulation is total, fragmentation low in some clumps and high in others, abrasion is nil. Encrusting and bioerosion is observed on the valves of C.? hatcheri. The taxa recorded were en-crusting bryozoans, sponge borings (En-tobia isp.), polychaetes (Maeandropolydora isp.), and bivalve borings such as Gastro-chaenolites isp.
1.2-6 m: 4.8 m Grey tuffaceous clayish siltstones similar to the previous ones. This bed carries three lenses, each ca. 0.2 m thick and dominated by "Turritella" patagonica. These lenses are separated by siltstones with very few clumps that carry almost exclusively "T". patagonica. Also present are, among many other taxa, Nucula (Lamellinucula) reticularis, Iheringinucula crassirugata, Scaeoleda? ortmanni, Neilo ornata, Arca patagonica, Cucullaea alta, Limopsis insolita, Glycymeris cuevensis, Atrina magellanica, Neopanis quadrisulcata, Swiftopecten nodosoplicatus, Zygochlamys geminata, Zygochlamys quemadensis, Reticulochlamys proxima, Jorgechlamys centralis, Crassostrea? hatcheri, Pteromyrtea crucialis, Pleuromeris cruzensis, Fasciculicardia patagonica, Spissatella lyelli, Hedecardium? ameghinoi, Hedecardium? puelchum, Trachycardium pisum, Cardium patagonicum, Lahillia patagonica, Maorimactra indistincta, Macoma? santacruzensis, Retrotapes striatolamellata, Dosinia laeviuscula, Ameghinomya darwini, Ameghinomya meridionalis, Caryocorbula hatcheri, Panopea quemadensis, Liotia scotti, Gibbula (Phorcus) margaritoides, Valdesia dalli, Solariella dautzenbergi, Calliostoma santacruzense, Calliostoma peraratum, Calliostoma cossmanni, Calliostoma garretti, "Turritella" ambulacrum, Struthiochenopus santacruzensis, Perissodonta ornata, Perissodonta patagoniensis, Crepidula gregaria, Cerithioderma patagonica, Polinices santacruzensis, Polinices ortmanni, Glossaulax vidali, Lunatia consimilis, Sassia bicegoi, Cerithiopsis juliana, Cirsotrema rugulosum, Cirsotrema quemandense, Trophon santacruzensis, Xymenella dautzenbergi, Xymene cossmanni, Enthacanthus monoceros, Crassilabrum hatcheri, cf. Ocenebra iheringi, Urosalpinx archipatagonica, Peonza torquata, "Cominella" annae, Penion subrectus, Miomelon petersoni, Miomelon gracilior, Adelomelon pilsbryi, Neoimbricaria patagonica, Dentimargo deuterolivella, Austroimbricaria quemadensis, Zeadmete ameghinoi, Zeadmete cruzialis, Antimelatoma quemadensis, Splendrillia santacruzensis, Austrotoma cuevensis, Fusiguraleus iheringi, Terebra quemadensis, Terebra santacruzensis, Odostomia synarthrota, Turbonilla cuevensis, Turbonilla observatonis, Semiacteon argentinus, Cylichna juliana, Kaitoa patagonica, "Odontostomia" euryope, Dentalium sp. Within the lens close-packing is dense (55%), size-sorting poor - with juvenile and adult specimens alike - , orientation is chaotic, disarticulation is total, fragmentation very low, and abrasion nil. Encrusting and bioerosion occur on the oysters.
6-27 m: 21 m Yellowish-brown tuffaceous siltstones with at least two beds dominated by "Turritella" patagonica. They measure 1 and 2.5 m thick respectively. The top bed is the most conspicuous and shows sectors in which it is hardened into concretions that outstand in the section. Faunal composition almost identical to that of the underlying bed. Contacts are wavy, close-packing is dense/loose (25%) at some places and dense (55%) at others, size-sorting is poor (full size range of "T." patagonica), orientation is chaotic, disarticulation high with very few specimens keeping conjoined valves, fragmentation is high in some sectors and low in others, abrasion nil. Between 15 and 18 m above the base of this bed there are thin intercalated lenses of 0.05-0.3 m thickness dominated also by "T." patagonica. At the very top there is Dentalium in life position and Limopsis insolita.
27-30 m: 3 m Grey tuffaceous siltstones. Lithology similar to previous ones, but the color change from yellowish to grey is notable.
30-34 m: 4 m Grey tuffs. The first 0.4 m carry large Ophiomorpha isp. The rest of the bed is massive, with only a few sparse fragments of shells.
34-56 m: 22 m Yellow tuffaceous siltstone highly bioturbated, with numerous very large Thalassinoides isp. (up to 0.08 m diameter). Sparse body fossils include crabs, very small Lucinoidea in life position and larger fragments of maybe the same taxon. Preservation is gypsum replacement. The top 8 m carry very sparse valves of Cucullaea alta and Dosinia laeviuscula, in all cases disarticulate and convex-up, some of them broken. Numerous concretions (as recorded by Darwin) of 0.8 to 1.4 m diameter and conjoined in a "necklace" manner. An especially conspicuous bed of concretions is at 6 m from the bottom of this bed.
56-56.5 m: 0.2-0.5 m Very fine sandstone with abundant specimens of Dosinia laeviuscula. This is a monospecific bed with a wavy bioturbated contact at the bottom and a wavy one at the top. Close-packing is dense (55%), size-sorting is well sorted, orientation chaotic, disarticulation high, fragmentation is moderate to low, abrasion nil. At the base there are numerous tubes that reach down up to 0.7 m into the underlying bed. This shell bed is replaced laterally by light grey to white tuff of approximately the same thickness.
56.5-75.5 m: 19 m Interstratified siltstone, claystone and fine sandstone. At the bottom it carries right valves of Crassostrea? hatcheri, in all cases convex-down. At 11 m from the bottom the color changes from brown to yellowish-green.
b) Mount Entrance Section (S50º08'32.2"; W68º22'23.6")
0-17 m: 17 m Grey and greenish tuffaceous clayish siltstones with a few specimens of "Turritella" patagonica that may be isolated or forming clumps. At 7 m from the base there is a sandy lens dominated by this species but also containing a few brachiopods and bivalves. At 14 m there are large "necklace" concretions of about 0.3 m diameter. Between 16 and 17 m the bed is strongly bioturbated (Ophiomorpha isp. and other small tubes in all directions).
17-27 m: 10 m Very bioturbated tuffaceous sandstones.
27-27.8m: 0.8 m Fine tuffaceous sandstones, dark grey and very hard, with crossbedding. At the base it carries load structures. These sandstones are very bioturbated, with many tubes of about 0.01 m diameter placed obliquely to the stratification planes.
27.8-38.8 m: 11 m Tuffaceous bioturbated sandstones similar to the underlying bed.
38.8-39.4 m: 0.6 m Very fine sandstones with abundant fossils. This is a shell bed with a wavy contact at the base and a gradual transition at the top. Close-packing is dense (55%) at the base and loose (15%) at the top, size-sorting is well sorted, as complete specimens and fragments show approximately the same size, a few large whole bivalves are also observed, orientation is chaotic, disarticulation complete, fragmentation high, and many naticid and muricid boreholes are evident on the shells. This bed carries a very rich fauna of well preserved mollusks, including among others Nucula (Lamellinucula) reticularis, Iheringinucula crassirugata, Scaeoleda? ortmanni, Neilo ornata, Arca patagonica, Cucullaria darwini, Cucullaea alta, Limopsis insolita, Glycymeris cuevensis, Atrina magellanica, Neopanis quadrisulcata, Swiftopecten nodosoplicatus, Zygochlamys geminata, Zygochlamys quemadensis, Reticulochlamys proxima, Jorgechlamys centralis, Crassostrea? hatcheri, Pteromyrtea crucialis, Pleuromeris cruzensis, Fasciculicardia patagonica, Spissatella lyelli, Spissatella kokeni, Hedecardium? ameghinoi, Hedecardium? pauciradiatum, Hedecardium? puelchum, Trachycardium pisum, Cardium patagonicum, Lahillia patagonica, Maorimactra indistincta, Serratina jeguaenis, Macoma? santacruzensis, Retrotapes ortmanni, Retrotapes striatolamellata, Dosinia laeviuscula, Ameghinomya darwini, Ameghinomya meridionalis, Caryocorbula hatcheri, Panopea quemadensis, Liotia scotti, Homalpoma philippii, Gibbula (Phorcus) margaritoides, Valdesia dalli, Solariella dautzenbergi, Calliostoma santacruzense, Calliostoma peraratum, Calliostoma cossmanni, Calliostoma garretti, "Turritella" ambulacrum, "Turritella" patagonica, Struthiochenopus santacruzensis, Perissodonta ornata, Perissodonta ameghinoi, Perissodonta patagoniensis, Calyptraea eleata, Sigapatella americana, Crepidula gregaria, Cerithioderma patagonica, Vermetus incertus, Polinices santacruzensis, Polinices ortmanni, Glossaulax vidali, Lunatia consimilis, Sconsia ovulum, Sassia bicegoi, Sassia morgani, Cerithiopsis juliana, Cirsotrema rugulosum, Cirsotrema qemandense, Trophon santacruzensis, Xymenella dautzenbergi, Xymene cossmanni, Enthacanthus monoceros, Crassilabrum hatcheri, cf. Ocenebra iheringi, Urosalpinx archipatagonica, Peonza torquata, "Cominella" annae, Penion subrectus, Austrocominella cosmanni, Retizafra improvisa, Miomelon petersoni, Miomelon gracilior, Adelome lon pilsbryi, Neoimbricaria patagonica, Dentimargo deuterolivella, Austroimbricaria quemadensis, Zeadmete ameghinoi, Zeadmete cruzialis, Sveltia maior, Scalptia vidali, Antimelatoma quemadensis, Splendrillia santacruzensis, Austrotoma cuevensis, Fusiguraleus iheringi, Eoturris santacruzensis, Borsonia patagonica, Terebra quemadensis, Terebra santacruzensis, Odostomia synarthrota, Odostomia suturalis, Turbonilla cuevensis, Turbonilla observatonis, Turbonilla iheringi, Semiacteon argentinus, Cylichna juliana, Kaitoa patagonica, "Odontostomia" euryope, Dentalium sp.
39.4-59.4 m: 20 m Interstratified fine sandstone, siltstone and claystone. It carries a few interstratified concretions with bivalves and echinoderms. At 12 m from the base of this bed there is a color change from brown to yellowish green.
Facies Analysis
Vertical and lateral facies distribution at the two studied sections allows recognition of three facies associations (Figs. 2 and 3). Facies Association 1 (FA1): Siltstones and fine sandstones rich in pyroclastic material and clumps, lens and beds of skeletal fossils concentrations. Facies Association 2 (FA2): Highly bioturbated siltstones and tuffs with concretions. Facies Association 3 (FA3): Interstratified siltstones, fine sandstones and claystones, thinly laminated in some sectors.
Facies Association 1 (FA1)
This facies association is roughly equivalent to Darwin's (Darwin 1846, p. 112) lowermost stratum (i.e., "… to a thickness of fifty or sixty feet, consists of a more or less hardened, darkish, muddy, or argillaceous sandstone…"). It includes yellowish grey siltstones and fine muddy sandstones with abundant pyroclastic material and skeletal concentrations forming clumps, lens or beds, dominated by "Turritella" patagonica (Fig. 4a). Also intercalated are specimens of Crassostrea? hatcheri in life position or slightly rotated but still with both valves articulated. This facies association also carries most of the rich and diverse fauna of the Monte León Formation (see description of Sections for a list of the most conspicuous taxa and their stratigraphic distribution). Also present are abundant bioturbations as small tubes running in all directions, and galleries assigned to Ophiomorpha isp. Concretions of variable size and shape are abundant, sometimes joined together forming "necklaces". This facies association measures 30 m thick at Darwin's Section and 17 m thick at Mount Entrance Section.

Figure 4: a) Clumps of "Turritella" patagonica (FA1); b) Ophiomorpha isp. (FA2); c) Concretions (FA2); d) Skeletal concentration dominated by Dosinia laeviscula at the top of FA2.
Some of the skeletal concentrations occur in laterally continuous beds, with wavy contacts and hardened concretionary sectors that form ledges in the section. There are a least three beds 1 - 2.5 m thick. The shell beds are densely to loosely packed (25%) in some of the sectors, and densely packed (55%) in others. Size-sorting is poor; although by far the numerically most abundant taxon is "T." patagonica; it is represented by specimens of all sizes. Orientation is chaotic, disarticulation is very high and only a few shells are conjoined. Fragmentation is low in some sectors and high in others, while abrasion is nil. Encrusting and bioerosion are evident in valves of C.? hatcheri, which carry encrusting bryozoans, sponge borings (Entobia isp.), polychaetes (Maeandropolydora isp.) and bivalve borings (Gastrochaenolites isp.).
In addition to the aforementioned shell beds also common are lenses of between 0.05 and 0.3 m thickness. The fossil content of these lenses is the same as the beds, as are all taphonomic attributes too. The uppermost part of this facies association shows a sharp decrease in fossil content, as the beds and lenses become much less frequent. However, a few disperse fossils such as a Dentalium in life position and Limopsis insolita can be observed.
Facies Association 1 represents a transition environment between the inner shelf and the subtidal part of a tidal plain, below fair weather wave base but above storm wave base. The mollusks contained in these beds are commonly considered to be indicative of shallow, fully marine conditions with normal salinity. Well over half the taxa present in this fauna are either very shallow infaunal (Turritella, Cucullaea, Cardium, etc) or epifaunal (Zygochlamys, Reticulochlamys, Swiftopecten, Trophon, Austrotoma, etc.). Most trophic levels are represented, including suspension feeders, deposit feeders and carnivores. Taxonomic composition of the assemblage and the beds with articulate and highly bored oyster clusters suggest that the background sedimentation rate was low.
The presence of thin consolidated lenses with fragments of shells or specimens lying chaotically is interpreted as of deposits formed by sporadic events recording a sharp increase in current velocity. The lenticular structure of some of these skeletal concentrations and their taphonomic features suggests they belong to the infilling of small scale tidal channels in a subtidal to inner shelf environment.
Facies Association 2 (FA2)
This facies association overlies FA1 and is equivalent to Darwin's lower two strata of the upper part of the cliff. It comprises massive and bioturbated tuffaceous siltstones and very fine sandstones, and tuffs. These are grey at the base and light yellow towards the top. The thickness is approximately 27 m at Darwin's section and 22 m at the Mount Entrance Section. The base carries large galleries assigned to Ophiomorpha isp. (Fig. 4b) and also few isolated shell fragments. Towards the top there are frequent galleries assigned to Thalassinoides isp. (up to 8 cm diameter). It also carries very sparse remains of crabs and small monospecific clumps of a small bivalve possibly referable to Pteromyrtea, and very few fragments of larger bivalve shells. The uppermost 8 m carry a few isolated valves of Dosinia laeviuscula. These are disarticulate, convex- up, and mostly unbroken. Throughout this facies association are common concretions of 0.8 - 1.4 m diameter that may be isolated or joined forming "necklaces" (Fig. 4c). Darwin had already noticed them as a conspicuous feature in this part of the section: "…Of the three beds, the central one is the most compact, and more like ordinary sandstone: it includes numerous flattened spherical concretions, often united like a necklace, composed of hard calcareous sandstone, containing a few shells: some of these concretions were four feet in diameter, and in a horizontal line nine feet apart…"(Darwin 1846, p. 113).
At the Mount Entrance Section there is a hard ledge of about 0.8 m thickness with cross-bedding. It is highly bioturbated with numerous tubes of about one cm diameter placed obliquely to stratification.
At the Darwin Section this facies association ends with a monospecific shell concentration of 0.2 - 0.5 m thickness, with a very fine sandstone matrix, and containing adult specimens of Dosinia laeviscula (Fig. 4d). Contacts of this bed are wavy and bioturbated, with many tubes reaching up to 0.7 m down into the underlying bed. Laterally it gives way to a white tuff of equal thickness. Close-packing is dense (55%), size-sorting good, orientation chaotic, disarticulation high, fragmentation moderate to low, abrasion nil. At the Mount Entrance Section this facies association ends with a skeletal concentration of about 0.6 m thickness, also with a fine sandstone matrix, but with a polyspecific taxonomic composition, carrying an abundant fauna of gastropods and bivalves (see description of Sections for a list of the most conspicuous taxa and their stratigraphic distribution). The lower contact is wavy and the upper one transitional, as close-packing is dense at the base and dispersed at the top. Size-sorting is good, as the whole specimens and the shell fragments are of the same size with only a few interspersed large bivalves and gastropods. Orientation is chaotic, disarticulation is complete, and fragmentation of large specimens is high but with a few whole specimens, while small specimens are largely unbroken. Many specimens show muricid and naticid boreholes.
Facies association 2 represents a subtidal environment at the lowest part of a tidal plain. The low diversity in the fauna throughout the facies association (Pteromyrtea and Dosinia), suggests a more marginal shallow environment than FA1. The high degree of bioturbation and dominance of trace fossils assigned to Thalassinoides isp. and Ophiomorpha isp. suggests low sedimentation rate and an environment with moderate to relatively low energy conditions. Taphonomic features of the specimens in the skeletal concentrations at the top, together with the presence of apparently unselected material suggests that these concentrations were formed during a high energy event, perhaps representing a storm event.
Facies Association 3 (FA3)
This facies association comprises the uppermost bed of Darwin's "upper part of the cliff" (Darwin 1846, p. 113). It includes 19 - 20 m of interstratified siltstones, fine sandstones and claystones, sometimes with heterolithic lamination. At the base there are convex-down right valves of Crassostrea? hatcheri. It carries a few concretions with bivalves and echinoderms. At 11 m above the base color changes abruptly from brown to yellowish green. Fine grain-size and sedimentary structures produced by variations between calm and turbulent water indicates deposition by combined traction and decantation processes. Such a process suggests that this facies association developed in a subtidal to intertidal environment.
Paleonvironments
Facies associations together with the invertebrate fauna recorded in each bed allow a detailed paleoenvironmental reconstruction of the rocks of the Monte León Formation in the Río Santa Cruz area. These rocks can be included in the regressive stage of the high sea-level period that began at the end of the Oligocene. The proposed deposition model suggests that it took place in a shallow sea, with variations that range from the inner shelf to subtidal or even intertidal environments.
At the base of the section (FA1) - i.e., the inner shelf to subtidal environment - there was a continuous deposition of siltstone with volcanic material (bioturbated) as result of decantation, with a few intercalated episodes recording higher energy events. These were recorded as beds and lenses of skeletal concentrations interpreted as having been deposited by channeled high energy unidirectional currents, perhaps tidal channels.
Towards the top of the sections (FA2) these lenses become sparser, while the bioturbation and pyroclastic content of the deposit increase. The top carries finely laminated siltstones and sandstones (FA3) that suggest a subtidal - or even intertidal - environment.
PALEONTOLOGY
One of the most conspicuous features of the Monte León Formation is the rich and excellently preserved mollusk fauna contained in the beds and lenses within the Punta Entrada Member; it occurs mainly in FA1 and the most conspicuous and common elements are mentioned above. This fauna has been known ever since Darwin collected the first specimens at the mouth of the Río Santa Cruz, and has been used over the years in the formulation of diverse hypothesis on the biostratigraphic and paleobiogeographic distributions of the taxa involved (del Río 2004 and references therein). Darwin's initial collection of 16 species of mollusks has been increased to over 110 recognized from that locality nowadays. Of these, virtually all are also known from the shell beds that were described later by Ameghino and Hatcher. The type locality of the species described by Sowerby (1846) is the section described here as Darwin Section. Thus, all his taxa come from beds lying within the Punta Entrada Member. It is important to note that taxa later described by von Ihering (1897, 1907, 1914) and Ortmann (1902) came from further south. The beds bearing these taxa were then included in the Superpatagonian, although Hatcher and Ortmann could see no difference between the Patagonian and Superpatagonian fauna. This is not surprising, as the Superpatagonian included two suites of taxa from entirely different sources. In the first place it included the species that is unique to the lowermost beds of the Santa Cruz Formation, i.e., Crasssotrea orbignyi. This oyster is quite different from Crassostrea? hatcheri, and is widespread in Patagonia and always appears associated to the marginal marine rocks at the base of the Santa Cruz Formation or its equivalent units. Secondly, the Superpatagonian included many taxa common to the "Leonense" chronostratigrapic unit of Ameghino, together with a suite of exclusive taxa not known from the underlying strata. However, these were based on material coming from two different sources. In the first place, it included material collected from the shell beds at the top of the Punta Entrada member, at localities where this unit is exposed along the coast between the mouth of the Río Santa Cruz and Monte Observación, just off limits to the south of Monte León National Park. It also includes material collected from beds overlying the former and containing Crassostrea orbignyi, exposed a few kilometers further inland and clearly at the base of the Santa Cruz Formation. The taphonomic attributes of the material other than the autochtonous accumulations of C. orbignyi suggest that it is in all cases reworked material from the underlying shell beds. In most cases, taxonomic differences can be attributed to the differential preservation of the two suites of material.
MEANING OF THE GREAT PATAGONIAN FORMATION AT THE MOUTH OF THE RÍO SANTA CRUZ
Upstream the Río Santa Cruz, the rocks described by Darwin at the mouth of the river are gradually replaced by continental beds nowadays included in the Santa Cruz Formation (Darwin 1846, p. 113-115). Although he described a section at about 160 km west of Santa Cruz, Darwin was doubtful that the rocks could be included in "the great Patagonian tertiary formation", as the only fossils he could find were very worn pieces of Ostrea patagonica (i.e. Crassostrea? hatcheri or more likely C. orbignyi). As Darwin never went ashore further south than Mount Entrance, he was unable to observe neither the beds at Las Cuevas nor those within the Monte Observación Member. Therefore, his concept of "great Patagonian tertiary formation" or "Patagonian Tertiary Formation" only included those rocks exposed at Santa Cruz (and other places further north).
At Mount Entrance Bertels (1980, p. 214) defined a lower member of the Monte León Formation that she named Punta Entrada Member and included in it all the marine beds exposed there. Darwin's Section is only 2.6 km away and no major structural features alter the stratigraphic relations between the beds exposed at the two localities. Consequently, we can confidently assume that (as confirmed by descriptions above) the two sections are equivalent. About 40 km further south along the coast and within the present Monte León National Park, these rocks continue to form part of the coastal cliff and lithological features remain overall constant. However, a short distance inland a second higher ledge is composed mainly by the continental Santa Cruz Formation. The contact between the two units has been subject of heated debates over the years, ever since the first subdivisions of the Patagonian Tertiary were made by Ameghino. The Patagonian Tertiary Formation in Darwin´s sense was restricted by Ameghino (1894) to the marine beds exposed along the coast between Chubut and Monte Observación in Santa Cruz. His subdivision of this unit (Ameghino 1898) into "Juliense" and "Leonense" was followed shortly afterwards (Ameghino 1900-1902) by his recognition that the overlying "Formación Santacruceña" included a lower marine bed he called "Superpatagoniano". In this he included the shell beds at the top of the cliff exposed at Las Cuevas, Yegua Quemada, Jack Harvey and Monte Observación, and also the oyster beds at the base of the Santa Cruz Formation exposed further inland at Cerro Monte León and Cerro Observación. He based such a subdivision on the fossil content, a fact that was later disputed by other authors (Wilckens 1905, Hatcher 1900, Ortmann 1902) who could see no difference in taxonomic composition between the Superpatagonian beds and those appearing in the typical Patagonian Formation.
Di Paola and Marchese (1973) did not recognize Bertels' introduction of a formational status for Ameghino's Juliense and Leonense (i.e., her San Julián and Monte León Formations). Instead they subdivided the Patagonia Formation (sensu Ameghino 1894) into three members based on lithological differences among them. The uppermost one, overlying the Monte León Member, they named Monte Observación Member. They defined this unit as the rocks appearing below the Santa Cruz Formation at Cerro Observación and that showed features common to the Monte León Member at the base and to the Santa Cruz Formation at the top. The bottom of this member is - according to them - an oyster bed of Crassostrea orbignyi, and the top is the last bed of this same oyster before giving way completely to the continental mammal-bearing rocks of the Santa Cruz Formation (the contact here is transitional). The total thickness measured by them between the two oyster beds was 27 m. When introducing her Punta Entrada Member, Bertels (1980, p. 214) also acknowledged the difference between this unit and the topmost beds of the Monte León Formation. Therefore she admitted the formal status given by Di Paola and Marchese (1973, p. 214) to the Monte Observación Member. However, she stated that a) the Monte Observación Member included all the shell beds at the top of the sections at Monte León (Cabeza de León), Las Cuevas, and Monte Observación (i.e., Cerro Observación, just west of Las Cuevas), and b) it replaced Ameghino's chronostratigraphic term "Piso Superpatagónico". However, these assumptions appear to be unwarranted. The lithological features described by Di Paola and Marchese (1973) for the Monte Observación Member are quite clearly distinct from those observed (as noted by Bertels herself; Bertels 1980, p. 216) in the shell beds at the top of the sections at Monte León (Cabeza de León) and Las Cuevas. The lithology in these shell beds is clearly the same as that in the underlying beds attributed by her to her Punta Entrada Member. The Superpatagonian of Ameghino originally included not only the shell beds at Monte León and Las Cuevas, but also those at Yegua Quemada, Jack Harvey, and Monte Observación all of them south of Las Cuevas. He also included the shell beds with Crassostrea orbignyi at Cañadón de los Artilleros, inland from San Julián, also underlying the continental Santa Cruz Formation. Neither Ameghino nor any author after him considered the fact that the shell beds at these places are thicker, with a far more abundant and better preserved fauna than those at the mouth of the Río Santa Cruz, the most likely reason for them supporting the difference between the Patagonian and Superpatagonian beds. Such a difference was minimized by Ortmann (1902) and Hatcher (1900) based on the very abundant fauna from Santa Cruz (possibly near Mount Entrance) that they had available. In fact, these authors were right in stating that the different composition of the fauna was due to local facies variations. Moreover, the far more abundant fauna at Las Cuevas is also surely the cause of bias in this sense, as the chances of collecting uncommon taxa are naturally much higher there.
Di Paola and Marchese (1973), while correctly defining their Monte Observación Member, never mentioned the fact that between the base of this unit (i.e., the lowest oyster bed at Cerro Observación) and the top of the shell beds at Las Cuevas there are ca. 20 m of rocks that are not exposed anywhere between Santa Cruz and the last exposures with marine shells at Puerto Coyle. As the lithology (and sedimentary structures as well) of the Monte Observación Member (other than the in situ beds with oysters in life position) is identical with that of the Santa Cruz Formation and the beds with other marine shells included in this unit are clearly reworked material with taxa that appear at Las Cuevas too, we suggest that the Monte Observación Member should be restricted to the rocks described by Di Paola and Marchese (1973, p. 214) and included in the Santa Cruz Formation. Likewise, the shell beds at the top of the sections at Monte León (Cabeza de León) and Las Cueva, Yegua Quemada, Jack Harvey and Monte Observación should be definitely referred to the Monte León Formation as they are equivalent to the shell beds lying within the Punta Entrada Member at Mount Entrance and Darwin's Section. Therefore, the use of the term Superpatagonian becomes superfluos.
CONCLUSIONS
As a result of this study we were able to pinpoint the most probable location where Charles Darwin made his observations in the Río Santa Cruz mouth. At this locality, i.e., our Darwin Section, we also identified the beds mentioned by him, including the location of the beds from where he collected the fossil mollusks that were described by Sowerby. The strata recognized by him are equivalent to those described by us and of which a detailed lithological description is provided.
Rocks belonging to the Monte León Formation exposed at the mouth of the Río Santa Cruz can be included in the regressive stage of the Atlantic transgressive-regressive cycle that flooded Patagonia during the late Oligocene-early Miocene. The deposition model suggests that it took place in a shallow sea, with variations that range from the inner shelf to subtidal or even intertidal environment. Analysis of sedimentological features of these rocks, together with the mollusk fauna contained in them, and comparison with those from nearby exposures further south along the coast and traditionally referred to the "Superpatagonian", suggest that the Monte Observación Member of the Monte León Formation should be restricted to its original concept. As a consequence, the term "Superpatagonian" becomes meaningless, at least as a chronostratigraphic unit in the area studied.
ACKNOWLEDGEMENTS
We would like to thank Sergio Martínez (Montevideo, Uruguay) and Jeff Stilwell (Melbourne, Australia) for their thoughtful comments that improved the original version of this manuscript. Help in the field by Juan Griffin is duly acknowledged too. The Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa supported part of the fieldwork (Project PI188). CONICET financed field and lab work (Project PIP1442 /05).
WORKS CITED IN THE TEXT
1. Ameghino, F. 1894. Ennumération synoptique des espèces des mammifères fossiles des formations eocenes de Patagonie. Boletín de la Academia Nacional de Ciencias 13: 259-445, Córdoba. [ Links ]
2. Ameghino, F. 1896. Notas sobre cuestiones de geología y paleontología argentina. Boletín del Instituto Geográfico Argentino 17: 87-119. [ Links ]
3. Ameghino, F. 1898. Sinopsis Geológico-Paleontológica. Segundo Censo de la República Argentina 1: 111-225. [ Links ]
4. Ameghino, F. 1900-1902. L'age des formations sédimentaires de Patagonie. Anales de la Sociedad Científica Argentina 50: 109-130, 145-165, 209-229 (1900); 51: 20-39, 65-91 (1901); 52: 189-197, 244-250 (1901); 54: 161-180, 220-249, 283-342 (1902). [ Links ]
5. Barreda, V. and Palamarczuk, S. 2000. Palinomorfos continentales y marinos de la Formación Monte León en su área tipo, provincia de Santa Cruz, Argentina. Ameghiniana 37(1): 3-12. [ Links ]
6. Becker, D. 1964. Micropaleontología del Superpatagoniense de las localidades Las Cuevas y Monte Entrance (provincia de Santa Cruz). Ameghiniana 3(10): 319-351. [ Links ]
7. Bertels, A. 1970. Sobre el "Piso Patagoniano" y la representación de la época del Oligoceno en Patagonia austral, República Argentina. Revista de la Asociación Geológica Argentina 25(4): 495-501. [ Links ]
8. Bertels, A. 1980. Estratigrafía y foraminíferos (Protozoa) bentónicos de la Formación Monte León (Oligoceno) en su área tipo, provincia de Santa Cruz, República Argentina. Actas 2º Congreso Argentino de Paleontología y Bioestratigrafía y 1º Congreso Latinoamericano de Paleontología (Buenos Aires, 1978) 2: 213- 273. [ Links ]
9. Camacho, H. H. 1974. Bioestratigrafía de las formaciones marinas del Eoceno y Oligoceno de la Patagonia. Anales de la Academia de Ciencias Exactas, Físicas y Naturales 26: 39-57. [ Links ]
10. Camacho, H. H. 1979. Significados y usos de "Patagoniano", "Patagoniense", "Formación Patagonica", "Formación Patagonia" y otros términos de la estratigrafía del Terciario marino argentino. Revista de la Asociación Geológica Argentina 34(3): 235-242. [ Links ]
11. Cossmann, M. 1899. Description des quelques coquilles de la formation Santacruzienne en Patagonie. Journal de Conchiliologie 47(3): 223-241. [ Links ]
12. Darwin, Ch. 1839. Narrative of the surveying voyages of His Majesty's Ships Adventure and Beagle, between the years 1826 and 1836, describing their examination of the southern shores of South America, and the Beagle's circumnavigation of the globe. Colburn, H. Ed., London, vol. 3, 615 p. [ Links ]
13. Darwin, Ch. 1846. Geological observations on South America. Being the third part of the geology of the voyage of the Beagle, under the command of Capt. Fitzroy, R. N. during the years 1832 to 1836. Smith Elder and co Ed., London, 279 p. [ Links ]
14. del Río, C. J. 2004. Tertiary marine Molluscan Assemblages of Eastern Patagonia (Argentina): a biostratigraphic analysis. Journal of Paleontology 78(6):1097-1122. [ Links ]
15. del Río, C. J. and Camacho, H. H. 1998. Tertiary nuculoids and arcoids of eastern Patagonia (Argentina). Palaeontographica Abt. A, 250: 47-88. [ Links ]
16. Di Paola, E. C. and Marchese, H. G. 1973. Litoestratigrafía de la Formación Patagonia en el área tipo (Bajo de San Julián-desembocadura del río Santa Cruz). Provincia de Santa Cruz. República Argentina. 5º Congreso Geológico Argentino, Actas 3: 207-222. [ Links ]
17. d'Orbigny, A. 1842a. Voyage dans l'Amérique Méridionale (le Brésil, la République Orientale de l'Uruguay, la République Argentine, la Patagonie, la République du Chili, la République du Bolivia, la République du Perou), exécuté pendant les années 1826,1827,1828,1829, 1830,1831, 1832 et 1833. P. Bertrand. Paris, V. Levrault, Strasbourg, vol. 3, part. 3, 290 p. [ Links ]
18. d'Orbigny, A. 1842b. Voyage dans l'Amérique Méridionale (le Brésil, la République Orientale de l'Uruguay, la République Argentine, la Patagonie, la République du Chili, la République du Bolivia, la République du Perou), exécuté pendant les années 1826,1827,1828,1829, 1830,1831, 1832 et 1833. P. Bertrand. Paris, V. Levrault, Strasbourg, vol. 3, part. 4, 96 p. [ Links ]
19. Feruglio, E. 1949. Descripción Geológica de La Patagonia. Ministerio de Industria y Comercio de La Nación, Dirección General de Yacimientos Petrolíferos Fiscales 2: 1-349. [ Links ]
20. Fleagle, J.G., Bown, T.M., Swisher, C. and Buckley, G. 1995. Age of the Pinturas and Santa Cruz Formations. 6º Congreso Argentino de Paleontología y Bioestratigrafía (Trelew), Actas: 129-135. [ Links ]
21. Hatcher, J.B. 1897. On the geology of Southern Patagonia. American Journal of Science 4(23): 327-354 [ Links ]
22. Hatcher, J. B. 1900. Sedimentary rocks of Southern Patagonia. American Journal of Sciences (4) 9(50): 85-107. [ Links ]
23. Hatcher, J.B. 1903. Geography of Southern Patagonia. In Scott, W.B. (ed.) Reports of the Princeton University Expedition to Patagonia 1896-1899, J. Pierpoint Morgan Publishing Foundation, Narrative of the expeditions 1: 45-332, Princeton. [ Links ]
24. Ihering von, H. 1897. Os Molluscos dos terrenos terciarios da Patagonia. Revista do Museo Paulista 2: 217-382. [ Links ]
25. Ihering von, H. 1899. Die Conchylien der patagonischen Formation. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie 1899: 1-46. [ Links ]
26. Ihering von, H. 1902. Historia de las ostras argentinas. Anales del Museo Nacional de Buenos Aires 7: 109-123. [ Links ]
27. Ihering von, H. 1907. Les mollusques fossiles du Tertiaire et du Crétacé superieur de l'Argentine. Anales del Museo Nacional de Buenos Aires, serie 3(7): 1-611. [ Links ]
28. Ihering von, H. 1914. Catálogo de molluscos Cretáceos e Terciarios da Argentina da collecçao do auctor [with appendix in German]. Notas Preliminares da Revista do Museu Paulista 1(3): 1-148. [ Links ]
29. Kidwell, S. M. and Holland, S. M. 1991. Field description of coarse bioclastic fabrics. Palaios 6: 426-434. [ Links ]
30. Kidwell, S. M., Fürsich, F. T. and Aigner, T. 1986. Conceptual Framework for the Analysis and Classification of Fossil Concentrations. Palaios 1: 228-238. [ Links ]
31. Legarreta, L. and Uliana, M. A. 1994. Asociaciones de fósiles y hiatos en el Supracretácico-Neógeno de Patagonia: una perspectiva estratigráfico-secuencial. Ameghiniana 31(3): 257-281. [ Links ]
32. Malumián, N. 1999. La sedimentación y el volcanismo terciarios en la Patagonia extraandina. In Caminos, R. (ed.) Geología Argentina, Anales del Instituto de Geología y Recursos Minerales 29(18): 557-612. [ Links ]
33. Malumián, N. 2002. El Terciario marino. Sus relaciones con el eustatismo. In Haller, M.J. (ed.) Geología y Recursos Naturales de Santa Cruz, 15º Congreso Geológico Argentino, Relatorio I-15: 237-244. [ Links ]
34. Náñez, C. 1988. Foraminíferos y bioestratigrafía del Terciario Medio de Santa Cruz oriental. Revista de la Asociación Geológica Argentina 43(4): 493-517. [ Links ]
35. Olivera, A. M. 1990. Aspectos paleoecológicos de asociaciones de moluscos fósiles de la Formación Monte León (Oligoceno). Puerto Santa Cruz (Santa Cruz). 5º Congreso Argentino de Paleontología y Biostratigrafía, Actas 1: 155-160. [ Links ]
36. Ortmann, A. E. 1902. Tertiary Invertebrates. In Scott, W. B. (ed.) Reports of the Princeton University Expedition to Patagonia 1896-1899, J. Pierpoint Morgan Publishing Foundation, Paleontology volume 4(I), Part 2: 45- 332, Princeton. [ Links ]
37. Panza, J. L., Irigoyen, M. V. and Genini, A. 1995. Hoja Geológica 4969-IV, Puerto San Julián, provincia de Santa Cruz, República Argentina. Boletín de la Secretaría de Minería de la Nación, Dirección Nacional del Servicio Geológico 211: 1-77. [ Links ]
38. Parras, A. and Casadío, S. 2005. Taphonomy and sequence stratigraphic significance of oysterdominated concentrations from the San Julián Formation, Oligocene of Patagonia, Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology 217: 47-66. [ Links ]
39. Parras, A., Griffin, M., Feldmann, R., Casadío, S., Schweitzer, C. and Marenssi, S. 2008. Correlation of marine beds based on Sr- and Ar- date determinations and faunal affinities across the Paleogene/Neogene boundary in southern Patagonia, Argentina. Journal of South American Earth Sciences 26: 204-216. [ Links ]
40. Philippi, R. A. 1887. Los fósiles terciarios i cuartarios de Chile. F. A. Brockhaus, Leipzig and Museo Nacional de Historia Natural, 256 p., Santiago. [ Links ]
41. Riggi, J. C. 1978. La importancia de los sedimentos piroclásticos y de la sílice biogenética en la estratigrafía de la Formación Patagonia. Revista de la Asociación Geológica Argentina 33(2): 158-171. [ Links ]
42. Sowerby, G. B. 1846. Descriptions of the Tertiary fossil shells from South America. In Darwin, Ch. Geological observations on South America. Being the third part of the geology of the voyage of the Beagle, under the command of Capt. Fitzroy, R. N. during the years 1832 to 1836. Smith Elder and Co. Ed., Appendix, p. 249-264, London. [ Links ]
43. Wichmann, R. 1922. Observaciones geológicas en el Gran Bajo de San Julián y sus alrededores (Territorio de Santa Cruz). Boletín Dirección General de Minas 30B: 1-34. [ Links ]
44. Wilckens, O. 1905. Die Meeresablagerugen der Kreide und Tertiärformation in Patagonien. Neues Jahrbuch für Geologie, Mineralogie und Paläontologie 21: 98-195. [ Links ]
45. Windhausen, A. 1931. Geología Argentina. Segunda Parte: Geología Histórica y Regional del territorio Argentino. Editorial Peuser, 676 p., Buenos Aires. [ Links ]
46. Zinsmeister, W. J. 1981. Middle to late Eocene invertebrate fauna from the San Julián Formation at Punta Casamayor, Santa Cruz province, Southern Argentina. Journal of Paleontology 55: 1083-1102. [ Links ]
Recibido: 1 de octubre de 2008
Aceptado: 20 de noviembre de 2008

