










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de la Asociación Geológica Argentina
versión impresa ISSN 0004-4822versión On-line ISSN 1851-8249
Rev. Asoc. Geol. Argent. v.64 n.1 Buenos Aires abr. 2009
Young Darwin and the ecology and extinction of pleistocene south american fossil mammals
Sergio F. Vizcaíno1, Richard A. Fariña2 and Juan Carlos Fernicola3
1 División Paleontología Vertebrados, Museo de La Plata, La Plata, Argentina. CONICET. Email: vizcaino@fcnym.unlp.edu.ar
2 Departamento de Paleontología, Facultad de Ciencias, Universidad de la República, Montevideo. Uruguay. Email: fari~a@fcien.edu.uy
3 Sección Paleontología de Vertebrados, Museo Argentino de Ciencias Naturales "Bernardino Rivadavia", Buenos Aires. Argentina. Email: jctano@macn.gov.ar
ABSTRACT
During his two years in South America Charles Darwin became fascinated not only with the lush vegetation of Brazil, but also with the gigantic Pleistocene mammals that he found in the drier areas of Uruguay, and in the pampas and Patagonian coast of Argentina. These findings included various ground sloths and glyptodonts among xenarthrans, and hoofed herbivores like Toxodon and Macrauchenia, in addition to horses and small rodents. He concluded that the general assumption that large animals require luxuriant vegetation was false and that vitiated the reasoning of geologists on some aspects of Earth's history. He also reflected on the evident changes that occurred in the continent, the extinct fauna of which suggested to him an analogy to southern parts of Africa. He wondered about our ignorance of biological traits in extinct creatures and the reasons for their extinction. Thus, not only did Darwin inspire phylogenetic studies on fossil mammal lineages, he also opened a gate to the research on their behaviour, physiology and extinction; i.e., their palaeobiology. Whereas the first approach was largely developed in South America beginning about the second half of the 19th century due to the intellectual influence of Florentino Ameghino, palaeobiology became a much more recent line of work, in apparent relation to innovations in methodology and technology. This contribution provides an overview of recent contributions on the palaeobiology of Pleistocene fossil mammals of South America as attempts to provide answers for Darwin's questions.
Keywords: Darwin; Ecology; Extinction; South America; Mammals.
RESUMEN: El joven Darwin y la ecología y extinción de los mamíferos fósiles sudamericanos. Durante los dos años que Charles Darwin estuvo en América del Sur no sólo se deslumbró con la profusa vegetación de Brasil, si no también con los gigantescos mamíferos pleistocenos que colectó en áreas más secas de Uruguay y en la pampa y la costa patagónica de Argentina. Sus hallazgos incluyeron distintos perezosos y gliptodontes, ungulados herbívoros como Toxodon y Macrauchenia, además de caballos y pequeños roedores. Darwin, desechó la presunción general de que los grandes animales requieren una exuberante vegetación, reconociendo que la misma condicionó las interpretaciones que hicieran los geólogos sobre algunos aspectos de la historia de la Tierra. También reflexionó sobre los cambios acaecidos en el continente, cuya fauna le sugirió una clara analogía con lo observado en el sur de África. Darwin se preguntó acerca de nuestro desconocimiento de las características biológicas y de las causas que llevaron a su extinción. Así, no sólo inspiró el estudio filogenético de distintos linajes de mamíferos fósiles, si no que abrió la puerta a las investigaciones sobre su comportamiento, fisiología y extinción; i.e. su paleobiología. En América del Sur, la influencia intelectual de Florentino Ameghino permitió apuntalar el estudio filogenético de los mamíferos durante la segunda mitad del Siglo XIX, mientras que el fortalecimiento de la paleobiología se dio en tiempos recientes en relación con innovaciones metodológicas y tecnológicas. Esta contribución provee una visión global de las contribuciones realizadas en paleobiología de mamíferos pleistocenos de América del Sur como un intento de responder los cuestionamientos que se hiciera Darwin.
Palabras clave: Darwin; Ecología; Extinción; América del Sur; Mamíferos.
INTRODUCTION
When HMS Beagle reached the coast of Brazil in February, 1832, Charles Darwin was a 22-year old theology student at Cambridge University and his ambition was to become a rural pastor. He had started to develop an interest in natural history some time before, while studying medicine at Edinburgh University. The overwhelming power of South American biodiversity that greeted the young Charles Darwin led him to declare that "The elegance of the grasses, the novelty of the parasitical plants, the beauty of the flowers, the glossy green of the foliage, but above all the general luxuriance of the vegetation, filled me with admiration." (Voyage of the Beagle, Chapter I, Feb. 1832)
However, it was not only the luxuriance that influenced his mind but also the lesser plains of the south and their fossils. Actually, he became fascinated by the discovery of gigantic Pleistocene mammals during his journeys in Uruguay, and the Pampean region and Patagonian coast of Argentina, before leaving the continent in May 1834 (see letters sent by Charles Darwin to Caroline Darwin and John Stevens Henslow in Burkhardt and Smith 1985, p. 276 and 280 respectively). As developed more fully in another contribution (see Fernicola et al., 2009) the extraordinary zoological collection of fossil and extant specimens made by Charles Darwin during his voyage to South America was studied by an important group of naturalists who published their conclusions between February 1838 and October 1843 in the "The Zoology of the Voyage of H.M.S. Beagle". All fossils mammals included in this work were studied by Richard Owen, who recognized the South American ungulates Toxodon platensis and Macrauchenia patachonica, a fossil horse identified as Equus sp., the mastodont Mastodon angustidens, and the ground sloths Glossotherium sp., Mylodon darwinii, Scelidotherium leptocephalum, Mega-lonyx jeffersonii, Megatherium cuvierii, and the glyptodonts Glyptodon clavipes, Hoplophorus euphractus among xenarthrans, as well as some smaller forms like rodents. The taxonomic history of several of these taxa is complex, including issues as varied as nomenclatural problems (e.g. Megatherium cuvierii nomen illegit.) to the mixing of specimens of different species or genera assigned to the same species (e.g. Megatherium americanum), an issue that is treated by Fernicola et al. (2009).
It is widely accepted that by the time Darwin boarded the Beagle he had been influenced by Lamarck's ideas on evolution (see Woodward 1987). Also, that the giant fossil quadrupeds he found were significant toward the development of his evolutionary theory, as very early they suggested to him that the similarities between extinct and living forms should be explained by the existence of common ancestors, and that the transformation of species to a large degree was not a vertical sequence, as Lamarck had proposed, but a tree with asymmetric branches (see Huxley and Kettlewell 1965). In his own words: "to my view, in S. America parent of all armadilloes might be brother to Megatherium - uncle now dead" (Darwin, 1837-1838 in Barret 1960). Woodward (1987) stated that by that time Darwin was also familiar with adaptation (The red notebook of Charles Darwin in Herbert 1980, p. 67), another important biological issue related to evolution, but an idea then associated with Natural Theology (Paley 1802), which argued that every organism was intentionally perfectly designed to its particular life conditions by God.
But it was not only the idea, means and processes of evolution that impressed Darwin. During his journey between Buenos Aires and Santa Fe he wrote "We may therefore conclude that the whole area of the Pampas is one wide sepulchre for these extinct quadrupeds" (Voyage of the Beagle, Chapter VII, Oct. 1833). By that time his sharp mind had already noted "That large animals require luxuriant vegetation has been a general assumption, which has passed from one work to another. I do not hesitate, however, to say that is completely false; and that it has vitiated the reasoning of geologists, on some points of great interest in the ancient history of the world" (Chapter V, of the Beagle, Aug. 1833, The red notebook of Charles Darwin in Herbert 1980, p. 54).
In January 1834, Darwin collected remains of Macrauchenia in Patagonia near San Julián, in what today is Santa Cruz province. He made inferences about the environment in which this beast lived and reflected on its extinction: "Mr. Owen... considers that they form part of an animal allied to the guanaco or llama, but fully as large as the true camel. As all the existing members of the family of Camelidae are inhabitants of the most sterile countries, so we may suppose was this extinct kind… It is impossible to reflect without the deepest astonishment, on the changed state of this continent. Formerly it must have swarmed with great monsters, like the southern parts of Africa, but now we find only the tapir, guanaco, armadillo, capybara; mere pigmies compared to antecedents races... Since their loss, no very great physical changes can have taken place in the nature of the Country. What then has exterminated so many living creatures?…We are so profoundly ignorant concerning the physiological relations, on which the life, and even health (as shown by epidemics) of any existing species depends, that we argue with still less safety about either the life or death of any extinct kind" (Voyage of the Beagle, Chapter IX, Jan. 1834).
In this way, Darwin not only triggered the studies on the genealogical interpretation of the fossil mammal lineages, but also opened a gate to the research on their behaviour, physiology and extinction; in others words, on their palaeobiology. Phylogenetic studies, combined with morphological and taxonomic analyses, flourished in South America, particularly beginning during the latter half of the 19th century due to the intellectual influence of Florentino Ameghino, but palaeobiology became a much more recent line of work, in apparent relation to innovations in methodology and technology. Indeed, Pleistocene South American fossil mammals show a greater morphological diversity than their living counterparts, as they include representatives of great body size and very peculiar features. Their peculiarity and general lack of modern analogues have encouraged creative palaeobiological approaches that will be outlined below. The aim of this contribution is to overview the recent contributions on the palaeobiology and palaeoecology of the Pleistocene fossil mammals of South America, in order to investigate if we can provide answers for the questions that the young Darwin made himself when he first collected them.
RECONSTRUCTING PALAEOBIOLOGY OF SOUTH AMERICAN PLEISTOCENE MAMMALS
Palaeobiologists are interested in reconstructing the form of the fossils as living animals, their habitat, ecological role, behaviour, and basic biology. Vizcaíno et al. (2004) and Vizcaíno et al. (2008) describe a basic protocol for palaeobiological studies that identifies three biological attributes that are essential for each taxon: size, diet and usage of substratum or type of locomotion. Such principles have been used for the last three decades (e.g., Andrews et al. 1979, Van Couvering 1980, Reed 1998, etc.) though not fully applied to South Aerican mammals. Morphological study of the masticatory and locomotor apparatuses allows predictions on the movements for which the apparatuses are optimized. In addition, analyses of mastication are useful for formulating hypotheses about the diet of the organism, while analyses of the locomotor apparatus allow inferences about the type of locomotion or preferences in the usage of substratum: runner, hopper, digger, burrower, etc. Obviously, these two aspects, added to body size, yield relevant data for the interpretation of an organism in a palaeobiological context. Palaeomammalogists have largely applied actualism, according to which past events are surmised by analogy with currently observable processes assuming that fossil species had similar habits to their current relatives. However, when phylogenetic affinity is not very close or fossil lineages possess morphologies not represented in extant species (Vizcaíno et al. 2004), this methodology does not provide reliable results, a circumstance that is particularly applicable to the mammalian faunas that evolved in relative isolation in South America during a good part of the Tertiary. Vizcaíno et al. (2004) and Vizcaíno et al. (2008) provide accounts on the reconstruction of palaeobiology through the application of the "form-function correlation approach" (Radinsky 1987), according to which function can be inferred from form, to make good use of the main sources of information like fossilized bones and teeth (though indirect evidence can also be used). Form-function relationships can be studied through different approaches, like functional morphology, biomechanics and ecomorphology (see definitions in Vizcaíno et al. 2008, and references therein)
It was not until the second part of the 1990s that authors began to apply biomechanic, morpho-geometrical, and ecomorphological methods to the study of morphology as part of a major project aimed at understanding the great palaeobiological diversity of the South American extinct forms. These results allowed the development of novel interpretations of their modes of life that, coupled with palaeoenvironmental data (geology, palaeoclimatology and reconstruction of palaeovegetation), provide insightful information on the paleoecological context in which these animals existed.
Body size
Body size has a remarkable influence on an animal's life because it can be correlated, among other features, with metabolism, limb bone dimensions and biomechanics of locomotion, or particular solutions for food intake. Body mass in Pleistocene xenarthrans was estimated using scale and computer generated (geometric) models, and allometric equations (see Vizcaíno et al. 2008 and references therein). Using these approaches, Fariña et al. (1998), Bargo et al. (2000) and Christiansen and Fariña (2003) estimated the masses of most Lujanian megamammals. Among the late Quaternary mammals of South America, three species rival for the title of the largest of them: the giant ground sloths Megatherium americanum and Eremotherium laurillardi whose body masses must have reached between three and six tonnes, depending on the approach used (Casinos 1996, Fariña et al. 1998), and the mastodont Stegomastodon superbus, estimated at four or five tonnes. The mylodontid ground sloth Lestodon armatus (Fig. 1) follows closely, with an estimated mass of three to four tonnes, while the other my-lodontids are smaller: between one and two tonnes for Glossotherium robustum and Mylodon darwini, and with Scelidotherium leptocephalum, at 900 kg, falling just short of the megamammal category (Bargo et al. 2000). Among glyptodonts the largest was Glyptodon clavipes, individuals which may have reached nearly two tonnes. Others, though smaller, were also very large mammals: Doedicurus clavicaudatus at 1400 kg, Panochthus tuberculatus at (1100 kg), and Glyptodon reticulatus at 850 kg.

Figure 1: Life reconstruction of Lestodon sp. The different size of the tusks suggest sexual dimorphism. Drawing by Néstor Toledo. Scale=100 cm.
There were also non-xenarthran giants, such as the camel-like Macrauchenia and the rhinoceros-like Toxodon (Fig. 2), whose body masses must have surpassed the one tonne limit (Fariña and Álvarez 1994, Fariña et al. 2005). The Carnivora, in turn, reached impressive sizes, although well below one tonne, with the sabre-tooth Smilodon populator and the short faced bear Arctotherium spp. attaining sizes of three or four hundred kilograms (Fariña et al. 1998).
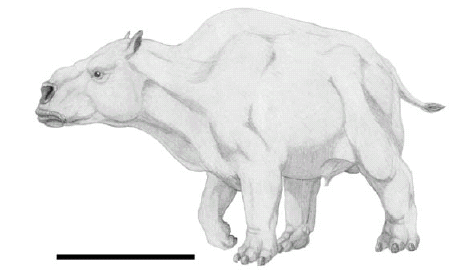
Figure 2: Life reconstruction of Toxodon sp. Drawing by Néstor Toledo. Scale=100 cm.
Limbs, locomotion and habits
Biomechanical studies performed in the last decade on large glyptodonts and ground sloths provided insight into the capacity of the limb bones to withstand bending forces, forearm extension and velocity, bipedalism or digging abilities. Within cingulates, Fariña's (1995) analyses of limb bones and locomotory habits in some glyptodonts indicated that femur strength indicators of large Pleistocene forms were equivalent to those of large living mammals capable of galloping (i.e. buffalos and rhinos), but values of the humerus were similar to those of elephants, which cannot gallop. The muscular insertions suggest that glyptodonts were able to adopt bipedal postures to perform strenuous activities, such as the intraspecific fighting proposed by Fariña (1995).
For the ground sloths, different specializations may have been derived from a primitive quadrupedal way of locomotion in both main lineages of Pleistocene forms, megatheriids and mylodontids. The giant sloth Megatherium americanum has been formally proposed as bipedal based on ichnologic and biomechanical evidence (Aramayo and Manera de Bianco 1996, Blanco and Czerwonogora 2003). The latter includes analyses of body size, speed, Froude number, indicator of athletic ability, bending and resistance moments of the vertebral column, as well as a complete geometric and biomechanical analysis of the footprints assigned to this species found in Pehuén-Có, Buenos Aires Province, Argentina. Bipedalism also implies that the forelimb could have been free to perform activities other than locomotion. Fariña and Blanco (1996) tested the possibility that the forearms of Megatherium americanum were designed for optimizing speed rather than strength of extension, and concluded that such a trait may have been associated with a potentially aggressive use of the animal's large claws. Bargo et al. (2000) analysed limb proportions and resistance to bending forces in mylodontids to infer their locomotor adaptations. The analysis indicates that some of them were well adapted for strenuous activities in which force was enhanced over velocity, such as digging. Based on this work, Vizcaíno et al. (2001) considered these taxa as possible builders of large Pleistocene burrows reported in the Pampean region (Zárate et al. 1998).
As for the strange hoofed mammals collected by Darwin, their athleticism seems to have been impressive in the case of Toxodon, which must have been capable of fast locomotion, and even more impressive in Macrauchenia, albeit for different reasons. Indeed, limb bone strength of the latter was larger if measured transversely rather than anteroposteriorly, which has been interpreted as the capability of suddenly turning while being pursued by a predator (Fariña et al. 2005).
Feeding apparatus
Functional morphology and biomechanics have been applied to the study of feeding in a wide range of xenarthrans (Vizcaíno et al. 2008, and references therein). Within the Pleistocene armoured forms, plant-eating was determined in eutatines (Vizcaíno and Bargo 1998), pampatheres (Vizcaíno et al. 1998, De Iuliis et al. 2000) and glyptodonts (Fariña and Vizcaíno 2001), although different kinds of herbivory may have occurred in each group. The studies also revealed that some cingulates evolved mechanical solutions not present in any related taxa, and do not have current analogues that can be used as models to investigate and interpret adaptations of lineages without living representatives. For instance, the masticatory apparatus in glyptodonts underwent a telescoping process that placed it well below the cranium (Fariña 1985, 1988), creating problems in the way that stresses produced by mastication were absorbed by the mandible and implying unusual jaw mechanics (Fariña and Vizcaíno 2001).
Bargo (2001), Bargo et al. (2006a, b), and Bargo and Vizcaíno (2008) studied the masticatory apparatus of the large South American Pleistocene ground sloths. Jaw mechanics, morphogeometric analyses, and the correlation between cranio-dental variables (hypsodonty, dental occlusal surface area and relative width of the muzzle) and diet, all suggested probable niche differentiation among ground sloths based on dietary categories. While the masticatory pattern of mylodontids is rather generalized with a clear anteromedial powerstroke, as previously proposed by Naples (1989), Megatherium americanum was well adapted for strong, mainly vertical biting. This information, in addition to tooth shape, suggests that teeth were mainly used for cutting, rather than grinding, and that fibrous food was not the main dietary component.
Hypsodonty is the relative increase in crown height of a tooth. It has been traditionally viewed as a response to dietary shifts toward abrasive vegetation, although recent work indicates that evolution of hypsodonty is also due to the higher prevalence of grit and dust in more open environments (Bargo et al. 2006a and references therein). Bargo et al.'s (2006a) comparative analyses of eleven species of Pleistocene sloths suggest that differences in hypsodonty may be explained by diet, habitat and behaviour. Among mylodontids, hypsodonty was unlikely due solely to dietary preferences, such as grazing. As mentioned above, some mylodontids were capable diggers that likely dug for food, and ingestion of abrasive soil particles probably played a considerable role in shaping their dental characteristics. Geographical distributions of the megatheriids Eremotherium and Megatherium indicate differing habitats as possible factors in hypsodonty differences. Vizcaíno et al. (2006) investigated the relationship between dental occlusal surface area (OSA) and diet, and other biological factors in fossil xenarthrans. They found that for most fossil xenarthrans OSA is smaller than expected compared to extant herbivorous mammals of equivalent body size. Within xenarthrans, cingulates show the highest OSA values, suggesting more extensive oral food processing than in tardigrades. Among ground sloths, mylodontids have extremely low OSA values, suggesting low efficiency in oral food processing that was probably compensated for by high fermentation in the digestive tract, and/or lower metabolic requirements. On the other hand, Megatherium americanum has an OSA as high as, or even higher than, that expected for a mammal of its size, which indicates higher oral food processing, lower fermentation capacity, and/or higher metabolic requirements.
Other features besides teeth are also important. For instance, Bargo et al. (2006b) used muzzle shape and facial musculature reconstructions to develop models of food intake in five species of South American Pleistocene giant ground sloths. Ground sloths with wide muzzles (Glossotherium robustum and Lestodon armatus) had a square, non-prehensile upper lip that, coupled with the tongue, were used to pull out grass and herbaceous plants (mostly bulk-feeders). Sloths with narrow muzzles (Mylodon darwini, Scelidotherium leptocephalum and Megatherium americanum) had a cone-shaped and prehensile upper lip (Fig. 3) that was used to select particular plants or plant parts (mixed or selective feeders).
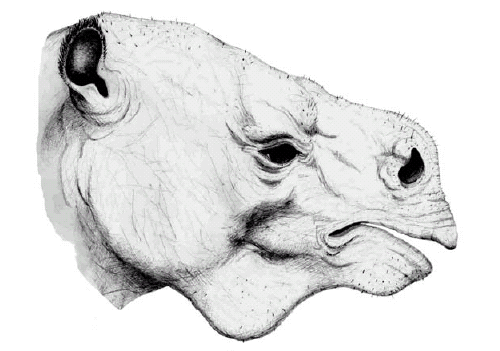
Figure 3: Life reconstruction of the head of Megatherium sp. Drawing by Néstor Toledo. Scale=50 cm.
A morphofunctional approach has been less often applied to other Lujanian mammals because they are more readily comparable to living analogues. The upper incisors of Toxodon were strongly arched, whereas the lower ones were horizontally arranged. Their great lateral expansion gave the lower jaw a giant, spade-like appearance. In Macrauchenia the retracted position of the large, elliptical nostrils suggests the presence of a trunk. Considering morphological and isotopic evidence, they have been mainly considered as grazers and mixed-feeders respectively (MacFadden and Shockey 1997). Among horses, Hippidion shows a narrower nasomaxillar region, with well developed preorbital fossae and a retracted nasal notch, a combination of features that has been interpreted as an adaptation to more closed habitats, such as a savanah. Also, Hippidion shows teeth with relatively lower degree of folding in the enamel and less hypsodont than Equus (Amerhippus), which suggest a diet less rich in silica. Congruently, biogeochemical data suggest that the species of Equus (Amerhippus) had a more grazing diet than Hippidion (MacFadden et al. 1996, Mac Fadden and Shockey 1997, MacFadden et al. 1999).
PALAEOECOLOGY
As for palaeoecological interpretations, there have been several approaches to the study of Cenozoic South American faunas, including feeding habits, locomotion, and trophic relationships. Fariña (1996) analysed the trophic relationships of the South American Lujanian (late Pleistocene-early Holocene) megamammals, from the perspective of the ecological implications of their body sizes. Based on an estimation of population density derived from body sizes Fariña (1996), as Darwin realized 160 years before, emphasized that the fauna contained a significant diversity of large herbivores. Conversely, according to Fariña, it did not contain a proportionally diverse suite of large carnivores. Assuming a basal metabolism in agreement with the body size of the beasts, the food energy required to sustain the fauna was calculated. The results suggest that based on the requirements of the species under consideration (i.e., mammals over 10 kg), they alone must have needed about 1.8 megajoules per square metre per year (hereafter, MJ m-2 year-1) of the vegetation which sustained them. Since a primary productivity of 7.3 MJ m-2 year-1 is considered excellent for modern open field ecosystems, it is difficult to explain how the smaller species of mammals could have survived, let alone reptiles, birds, insects and other consumers. If the Lujanian plains had been as productive as the African savannah is today, about 38 MJ m-2 year-1, the consumption efficiency would fall to little more than 3%, which is a typical value for modern grassland systems.
However, available evidence points to a different scenario. The fossiliferous Guerrero Member of the Luján Formation was deposited between about 20 to 10 thousand years before present, when the Last Glacial Maximum (LGM) was established and, due to the extensive glaciation in the Andes, the climate was much dryer and decidedly cooler than present conditions in that region (Clapperton 1983). Different sources of evidence are congruent with this paleoclimatic interpretation (Cantú and Becker 1988, Tonni 1990). As noted above, this was precisely Darwin's (1839) intuition about the landscape being less luxuriant than today. However, it seems that it was even more arid than surmised by the great naturalist. Current biogeographic reconstructions for the period of the LGM show that the Pampean plains underwent intense aeolian activity that redeposited large masses of silt and fine sand of periglacial origin. Southwest from the rivers where the sand was trapped in, a sand-sea in the southwestern half of the Pampas was formed, as well as a broad loessic belt over the remainder of the area. Also, the remains of the still-extant mammals, i.e., those whose habitat preference can be safely assigned, belong to species confined to Central and Patagonian faunistic provinces (Tonni 1985, Prado et al. 1987, Alberdi et al. 1989). The same can be said about the birds of this age (Tonni and Laza 1980), and analyses of pollen and ostracods have yielded congruent results (Quattrocchio et al. 1988, Markgraf 1989, Prieto 1996).
According to Iriondo and García (1993), the shift was about 750 km south west relative to present conditions. Hence, the place where the current city of Luján lies would have had climatic conditions similar to the climate that exists currently in the northern Patagonian locality of Choele-Choel (39ºS, Río Negro province), whose climogram indicates a lower mean annual temperature (2.5-3ºC less than at the present), with more marked seasonality (summers only about 1ºC colder but winters up to 4ºC colder). More importantly, the aridity must have been higher, with rainfall considerably lower, about 350 mm per year as compared with the nearly 900 mm current for that area. These figures are considered as approximations, given the likely influence of other factors, such as the well-known high edaphic quality of the Pampean region, and perhaps due to the impact of the local water bodies, a topic discussed below. Thus, primary productivity in the mid-latitude Lujanian might have been higher on average than that in today's Choele-Choel area but it does not seem likely that it could have been higher than the most productive present-day cattle field of Uruguay, and, hence, it must have been dramatically lower than the African savannah. As a consequence of this reasoning, Fariña (1996) suggested that the coexistence of so many large herbivores in a poor environment led to strong competition for resources. It is worth to mention here that by the Pleistocene large carnivorous birds, like the phorusrhacids and theratorns had long before become extinct in the Pampean region. He concluded that some of the mammals previously considered strict herbivores might have been flesh-eaters to some degree. After eliminating large ungulates for varied morphological reasons, Fariña (1996) proposed that ground sloths were opportunistic carrion eaters.
This challenging view, in turn, renewed interest in other ecological topics, such as niche partitioning in the Pleistocene and reinterpretation of the systematics of some South American Carnivora, among others. For instance, Vizcaíno's (2000) brief analysis of plant resource exploitation among sympatric Lujanian herbivorous armoured xenarthrans suggested that the main dietary difference among these cingulates was in the coarseness of the vegetation they were capable of processing. Bargo (2001) and Bargo et al. (2006b) proposed a niche differentiation among Lujanian ground sloths based on the different degrees of ability for plant selection due to muzzle morphology. Moreover, Vizcaíno et al. (2006) proposed that, like living sloths (see Gilmore et al. 2008 and references therein), mylodontids had very low metabolism, which suggests they were probably neither particularly abundant nor did they require as much food as originally calculated.
A recent revision of the bears from South America proposed that during the Late Pleistocene there were three species of bears (Soibelzon 2004), instead of one as considered by Fariña (1996). Bears may have acted as large scavengers, which was probably true for other carnivores such as felids and canids as well, forcing reexamination of Fariña's (1996) estimates of trophic diversity, at least to a certain extent. Prevosti and Vizcaíno (2006) reviewed carnivore richness in the Lujanian of the Pampean Region, describing the palaeoecology of these species (including their probable prey choices) and assessing the available information on taphonomy, carnivore ecology, and macroecology to test the hypothesis of "imbalance" of the Río Luján fauna. They found that the carnivore richness of the Río Luján fauna comprises five species: Smilodon populator, Panthera onca, Puma concolor, Arctotherium tarijense, and Dusicyon avus, plus two other species that may be added when the Lujanian of Buenos Aires province is included: Arctotherium bonariense and Canis nehringi. With the exception of D. avus and A. tarijense, these are hypercarnivores that could prey on large mammals (100-500 kg) and juveniles of megamammals (>1000 kg). Smilodon populator could also hunt larger prey with body mass between 1000 and 2000 kg. The review of the "imbalance" hypothesis reveals contrary evidence and allows the proposal of alternative hypotheses. If high herbivore biomass occurred during the Lujanian, a higher density of carnivores could be supported.
EXTINCTION
The study of extinctions has become a particularly relevant issue in palaeontology. Over the last few decades, analyses of biodiversity during the history of life and its ups and downs, both gradual and sudden (Sepkoski 1978, 1979, Raup and Sepkoski 1984) have become the cornerstone of the way we look into the deep past. Moreover, the spectacularity of mass extinction and of the non-actualistic proposals of extraterrestrial causes (Álvarez et al. 1980) attracts the attention of academics as well as of the general public.
The extinction of the giant mammals has long been attributed to the purported competition that followed the interchange of mammalian contingents across the Panama isthmus when it emerged some three million years ago. Known as the Great American Biotic Interchange, this asymmetrically reciprocal invasion brought carnivores (cats, dogs, mustelids, and bears), insectivores, rodents (in addition to the already present caviomorphs), rabbits, mastodons, tapirs, horses, camels, deer and peccaries to South America, where many of those groups still thrive. On the other hand, some South American lineages trekked north across the bridge: opossums, caviomorph rodents, toxodonts, glyptodonts, armadillos (including pampatheres), anteaters and ground sloths, and platyrhine primates, although not all were equally successful or ventured equally far in their new land. According to a proposal long held for the first half of the 20th century (Matthew 1930, Webb 1976, Simpson 1950), the evolutionarily advantaged northern mammalian lineages, on their triumphal path from their alleged evolutionary cradle in central Asia to all the corners of the globe, outcompeted their South American counterparts, driving them to extinction or forcing them to take refuge in marginal habitats. In the last part of that century, some authors critically revised the success of the North American immigrants in South America (Marshall et al. 1982, Marshall 1988, Webb 1985), and Webb (1991) provided an ecogeographic model that explains the assymmetrical results of the land-mammal interchange between both land masses. An analysis of the pattern of Pleistocene extinction (Lessa and Fariña 1996) revealed that large body size, rather than continent of origin, was the leading factor in determining which mammals went extinct.
Another interesting issue is that of human impact, as it relates both to the timing of the peopling of the Americas and to ethical, environmentally oriented issues. Up to present, evidence of this human impact was scarce, although some remains do show interesting connections. Particularly, a clavicle of a mylodontid ground sloth found in Arroyo del Vizcaíno, Uruguay, shows marks assigned to human activities. This specimen, as well as others, have been dated at about 29,000 years before present, a much older age than the 12,000 or 13,000 ybp currently accepted for human presence in the Americas (Arribas et al. 2001, Fariña and Castilla 2007).
ANSWERS TO DARWIN'S QUESTIONS
As noted above, we intend here to overview the palaeobiological and palaeoecological contributions on the Pleistocene mammals of South America and assess their measure of progress in answering young Darwin's questions and comments on such matters. As conclusions of this article, his assertions will be considered one by one and our current point of view will be added.
"We may therefore conclude that the whole area of the Pampas is one wide sepulchre for these extinct quadrupeds", said Darwin, and time has corroborated this statement in its fullest sense. Beginning with the collections made by Francisco Muñiz and Da´maso Larrañaga, as well as the subsequent work of the Ameghino brothers, the Museo de La Plata, the Museo Argentino de Ciencias Naturales "Bernardino Rivadavia", the Museo Nacional de Historia Natural de Montevideo and a multitude of smaller museums exhibit exquisite specimens of those amazing fossil beasts, as well as of many other remains that are housed in collection rooms.
"That large animals require luxuriant vegetation has been a general assumption, which has passed from one work to another. I do not hesitate, however, to say that is completely false; and that it has vitiated the reasoning of geologists, on some points of great interest in the ancient history of the world", claimed the young travelling naturalist. It seems he was absolutely right: as far as modern evidence is concerned, climates in middle latitudes seem to have shifted some 750 km towards the South-West since pleniglacial times, and hence the places where this impressive fauna must have lived then in a rather arid environment, as suggested by studies on small mammals, invertebrates, pollen and sediments, although more precise biogeographic and ecological hypotheses are yet in great need.
"Mr. Owen... considers that [the remains of what today is called Macrauchenia] form part of an animal allied to the guanaco or llama..." This statement would have been right if referred to the fossil camelid Palaeolama. However, Macrauchenia is not a camelid but a Litoptern, a group not related to any living order of mammals and its phylogenetic relationships are yet to be established. Nevertheless it is worth to consider his reflection on the palaeoenvironments: "As all the existing members of the family Camelidae are inhabitants of the most sterile countries, so may we suppose was this kind…It is impossible to reflect without the deepest astonishment, on the changed state of this continent. Formerly it must have swarmed with great monsters, like the southern parts of Africa, but now we find only the tapir, guanaco, armadillo, capybara; mere pigmies compared to antecedents races... Since their loss, no very great physical changes can have taken place in the nature of the Country". Darwin's astonishment is to be shared. As is evident from the preceding sections, not so long ago, in geological terms, the mid latitude plains of south eastern South America were home to perhaps the most spectacular mammalian fauna of all times. How an environment of apparently such a low productivity could support so many herbivores remains a mystery. On the other hand, advancements in studies in astronomical forcing and palaeoclimatology partially refutes Darwin's impression, as the nature of the country has changed since its coldest period during the LGM (approximately 18 kybp) and sea level has
risen more than 100 m, causing vast areas to become submerged. Ironically, the large beasts went extinct when the climate became less arid and the vegetation more luxuriant. That leads us to the following question: What then has exterminated so many living creatures?
Extinction, a major topic in current evolutionary thinking was surprisingly not a major issue for Darwin. References to the subject are scarce, although important, in his writings and we are left to wonder whether he accepted the ideas common in his time of catastrophes (Cuvier's global revolutions) having wiped species from the Earth. On the other hand, his proposal, many years later as a settled gentleman living in the quiet comfort of his Kent home, that species were originated by means of natural selection has been interpreted as the foundation of modern views about extinction. In other words, species were easily viewed as prone to extinction if their individuals were not fit, if their members lost in great numbers the struggle for life. As for the giant mammals that inspired his thoughts as a young traveller, he was more than eager to accept they no longer lived, although the question why it may have happened did not surface frequently enough, probably accepting Cuvier's point of view on the subject.
Our knowledge has increased considerably since then. Today, we pay enormous attention to this subject, due to our growing concern about present, dwindling biodiversity, our fascination with the spectacular extraterrestrial causes that have been proposed, and the ongoing
human impact on megamammals. However, we end this article with one last reflection.
"We are so profoundly ignorant concerning the physiological relations, on which the life, and even health (as shown by epidemics) of any existing species depends, that we argue with still less safety about either the life or death of any extinct kind" (Voyage of the Beagle, Chapter IX, Jan. 1834). Science has made considerable progress since those words were written by the person who changed the way humankind views itself and the rest of nature. However, we should be persuaded that his claim, which emphasized ignorance, be taken as a call to continue his approach. In this context, modern research on the marvellous beasts that roamed South America during the Pleistocene has indeed continued forward on the path set out by Darwin, contributing to the understanding of evolution intermingled with ecology and phylogeny. Thus, in the process of expanding our knowledge, as often happens in science, further ignorance has been revealed. However, we hope that this will only whet our appetite for the acquisition of more knowledge.
ACKNOWLEDGEMENTS
We want to acknowledge the editors for inviting us to participate in this volume. Gerry De Iuliis read critically an early version of the manuscript. Susana Bargo, Teresa Manera, Christine Janis and Darin Croft reviewed the manuscript. Néstor Toledo made the life reconstructions.
WORKS CITED IN THE TEXT
1. Alberdi, M.T., Menegaz, J.L., Prado, J.L. and Tonni, E.P. 1989. La fauna local de Quequén Salado - Indio Rico (Pleistoceno tardío) de la provincia de Buenos Aires, Argentina. Aspectos paleoambientales y bioestratigráficos. Ameghiniana 25: 225-236. [ Links ]
2. Alvarez, L.W., Alvarez, W., Asaro, F. and Michel, H.V. 1980. Extraterrestrial cause for the Cretaceous-Tertiary extinction: Experimental results and theoretical interpretation. Science 208: 1095-1108. [ Links ]
3. Andrews, P., Lord, J.M. and Evans, E.M.N. 1979. Patterns of ecological diversity in fossil and modern mammalian faunas. Biological Journal of the Linnean Society 11: 177-205. [ Links ]
4. Aramayo, S.A. and Manera de Bianco, T. 1996. Edad y nuevos hallazgos de icnitas de mamíferos y aves en el yacimiento paleo iconológico de Pehuen-Co (Pleistoceno Tardío), Provincia de Buenos Aires, Argentina. Asociación Paleontológica Argentina, Publicación Especial 4: 47-57. [ Links ]
5. Arribas, A., Palmqvist, P., Pérez-Claros, J.A., Castilla, R., Vizcaíno, S.F. and Fariña, R.A. 2001. New evidence on the interaction between humans and megafauna in South American. Publicaciones del Seminario de Paleontología de Zaragoza 5: 228-238. [ Links ]
6. Bargo, M.S. 2001. The ground sloth Megatherium americanum: skull shape, bite forces, and diet. In Vizcaíno, S.F., Fariña, R.A. and Janis, C. (eds.) Biomechanics and Paleobiology of Vertebrates. Acta Paleontologica Polonica (Special Issue) 46: 41-60. [ Links ]
7. Bargo, M.S. and Vizcaíno, S.F. 2008. Paleobiology of Pleistocene ground sloths (Xenarthra, Tardigrada): biomechanics, morphogeometry and ecomorphology applied to the masticatory apparatus. Ameghiniana 45:175-196. [ Links ]
8. Bargo, M.S., Vizcaíno, S.F., Archuby, F.M. and Blanco, R.E. 2000. Limb bone proportions, strength and digging in some Lujanian (Late Pleistocene-Early Holocene) mylodontid ground sloths (Mammalia, Xenarthra). Journal of Vertebrate Paleontology 20: 601-610. [ Links ]
9. Bargo, M.S., Vizcaíno, S.F. and Kay, R.F. 2004. Evidence for Predominance of Orthal Masticatory Movements in Early Sloths. 7° International Congress of Vertebrate Morphology, Journal of Morphology 260: 276. [ Links ]
10. Bargo, M.S., De Iuliis, G. and Vizcaíno, S.F. 2006a. Hypsodonty in Pleistocene ground sloths. Acta Paleontologica Polonica 51: 53-61. [ Links ]
11. Bargo, M.S., Toledo, N. and Vizcaíno, S.F. 2006b. Muzzle of South American ground sloths (Xenarthra, Tardigrada). Journal of Morphology 267: 248-263. [ Links ]
12. Barret, P. 1960. Transcription of Darwin`s first notebook on "Transmutation of species". Bulletin of the Museum of Comparative Zoology at Harvard College 122(6A): 248-296. [ Links ]
13. Blanco, R.E. and Czerwonogora, A. 2003. The gait of Megatherium Cuvier 1796 (Mammalia, Xenarthra, Megatheriidae). Senckenbergiana Biologica 83: 61-68. [ Links ]
14. Burkhardt, F. and Smith, S. 1985. The Correspondence of Charles Darwin Volume 1 1821-1836. Cambridge University Press, 752 p., Cambridge. [ Links ]
15. Cantú, M. and Becker, A. 1988. Holoceno del arroyo Spernanzoni, Dpto. Río Cuarto, Prov. Córdoba, Argentina. International Symposium Holocene in South America, Abstracts: 24. [ Links ]
16. Casinos, A. 1996. Bipedalism and quadrupedalism in Megatherium: an attempt at biomechanical reconstruction. Lethaia 29: 87-96. [ Links ]
17. Christiansen, P. and Fariña, R.A. 2003. Mass estimation of two fossil ground sloths (Xenarthra; Mylodontidae). In Fariña, R.A., Vizcaíno, S.F. and Storch, G. (eds.) Morphological studies in fossil and extant Xenarthra (Mammalia). Senckenbergiana Biologica 83: 95-101. [ Links ]
18. Clapperton, C. 1993. Quaternary geology and geomorphology of South America. Elsevier, 779 p., Amsterdam. [ Links ]
19. Darwin, C.R. 1839. Narrative of the surveying voyages of His Majesty's Ships Adventure and Beagle between the years 1826 and 1836, describing their examination of the southern shores of South America, and the Beagle's circumnavigation of the globe. Volume III journal and remarks 1832-1836. Henry Colburn Press, 615 p., London. [ Links ]
20. De Iuliis, G., Bargo, M.S. and Vizcaíno, S.F. 2000.Variation in skull morphology and mastication in the fossil giant armadillos Pampatherium spp. and allied genera (Mammalia: Xenarthra: Pampatheriidae), with comments on their systematics and distribution. Journal of Vertebrate Paleontology 20: 743-754. [ Links ]
21. Fariña, R.A. 1985. Some functional aspects of mastication in Glyptodontidae (Mammalia). Fortschritte der Zoologie 30: 277-80. [ Links ]
22. Fariña, R.A. 1988. Observaciones adicionales sobre la biomecánica masticatoria en Glyptodontidae (Mammalia; Edentata). Boletín de la Sociedad Zoológica (2a. época) 4: 5-9. [ Links ]
23. Fariña, R.A. 1995. Limb bone strength and habits in large glyptodonts. Lethaia 28: 189-96. [ Links ]
24. Fariña, R.A.1996. Trophic relationships among Lujanian mammals. Evolutionary Theory 11: 125-34. [ Links ]
25. Fariña, R.A. and Álvarez, F. 1994. La postura de Toxodon: una nueva reconstrucción. Acta Geologica Leopoldensia 39: 565-571. [ Links ]
26. Fariña, R.A. and Blanco, R.E. 1996. Megatherium, the stabber. Proceedings of the Royal Society London, Series B 263: 1725-1729. [ Links ]
27. Fariña, R.A. and Castilla, R. 2007. Earliest evidence for human-megafauna interaction in the Americas. In Corona, M.E. and Arroyo-Cabrales, J. (eds.) Human and Faunal Relationships Reviewed: An Archaeozoological Approach, British Archaeological Reports, International Series 1627: 31-34, Oxford. [ Links ]
28. Fariña, R.A. and Vizcaíno, S.F. 2001. Carved Teeth And Strange Jaws: How Glyptodonts Masticated. In Vizcaíno, S.F., Fariña, R.A. and Janis, C. (eds.) Biomechanics and Paleobiology of Vertebrates. Acta Paleontologica Polonica (Special Issue) 46: 87-102. [ Links ]
29. Fariña, R.A., Vizcaíno, S.F. and Bargo, M.S. 1998. Body mass estimations in Lujanian (Late Pleistocene-Early Holocene of South America) mammal megafauna. Mastozoología Neotropical 5: 87-108. [ Links ]
30. Fariña, R.A., Blanco R.E. and Christiansen, P. 2005. Swerving as the escape strategy of Macrauchenia patachonica (Mammalia; Litopterna). Ameghiniana 42: 751-760. [ Links ]
31. Fernicola, J.C., Vizcaíno, S.F. and De Iuliis, G. 2009. The fossil mammals collected by Charles Darwin in South America during his travels on board the HMS Beagle. Revista de la Asociación Geológica Argentina 64(1): 147-159. [ Links ]
32. Gilmore, D., Fittipaldi Duarte, D. and Peres da Costa, C. 2008. The physiology of two- and three-toed sloths. In Vizcaíno, S.F. and Loughry, W.J. (eds.) The Biology of the Xenarthra, p. 130-142. University Press of Florida. Gainesville. [ Links ]
33. Herbert, S. 1980. The red notebook of Charles Darwin. Bulletin of the British Museum (Natural History) Historical Series 7: 1-164. [ Links ]
34. Huxley, J. and Kettlewell, H.B.D. 1965. Charles Darwin and his world. Viking Press, 144 p., New York. [ Links ]
35. Iriondo, M. and García, N.O. 1993. Climatic variations in the Argentine plains during the last 18.000 years. Palaeogeography, Palaeoclimatology, Palaeoecology 10: 209-220. [ Links ]
36. Lessa, E.P. and Fariña, R.A. 1996. Reassessment of extinction patterns among the late Pleistocene mammals of South America. Palaeontology 39(3): 651-662. [ Links ]
37. MacFadden, B.J. and Shockey, B.J. 1997. Ancient feeding ecology and niche differentiation of Pleistocene mammalian herbivores from Tarija, Bolivia: morphological and isotopic evidence. Paleobiology 23: 77-100. [ Links ]
38. MacFadden, B.J., Cerling, T.E. and Prado, J.L. 1996. Cenozoic Terrestrial Ecosystem in Argentina Evidence from Carbon isotopes of Fossil Mammal Teeth. Palaios 11: 319-327. [ Links ]
39. MacFadden, B.J., Cerling, T.E., Harris, J.M. and Prado, J.L. 1999. Ancient latitudinal gradients of C3/C4 grasses interpreted from stable isotopes of New World Pleistocene horses. Global Ecology and Biogeography 8: 137-149. [ Links ]
40. Markgraf, V. 1989. Palaeoclimates in Central and South America since 18,000 BP based on pollen and lake-level records. Quaternary Science Review 8: 1-24. [ Links ]
41. Marshall, L.G. 1988. Land Mammals and the Great American Interchange. American Scientist 76(4): 380-388. [ Links ]
42. Marshall, L.G., Webb, S.D., Sepkoski, J.J. Jr. and Raup, D.M. 1982. Mammalian Evolution and the Great American Interchange. Science 215(4538): 1351-1357. [ Links ]
43. Matthew, W.D. 1930. Range and limitations of species as seen in fossil mammal faunas. Bulletin of the Geological Society of America 41: 271-274. [ Links ]
44. Naples, V.L. 1989. The feeding mechanism in the Pleistocene ground sloth, Glossotherium. Contributions in Science, Los Angeles County Museum of Natural History 415: 1-23. [ Links ]
45. Paley, W. 1802. Natural Theology. Oxford University Press, 384 p., Oxford. [ Links ]
46. Prado, J.L., Menegaz, A.N., Tonni, E.P. and Salamme, M.C. 1987. Los mamíferos de la fauna local Paso Otero (Pleistoceno tardío), provincia de Buenos Aires. Aspectos paleoambientales y bioestratigráficos. Ameghiniana 24: 217-233. [ Links ]
47. Prevosti, F. and Vizcaíno, S.F. 2006. Paleoecology of the large carnivore guild from the late Pleistocene of Argentina. Acta Palaeontologica Polonica 51: 407-422. [ Links ]
48. Prieto, A.R. 1996. Late Quaternary Vegetational and Climatic Changes in the Pampa Grassland of Argentina. Quaternary Research 45: 73-88. [ Links ]
49. Raup, D.M. and Sepkoski, J.J. Jr. 1984. Periodicity of extinctions in the geologic past. Proceedings National Academy of Sciences 81: 801-805. [ Links ]
50. Quattrocchio, M., Deschamps, C., Martínez, D., Grill, S. and Zavala, C. 1988. Caracterización paleontológica y paleoambiental de sedimentos cuaternarios, Arroyo Napostá Grande, Provincia de Buenos Aires. 2º Jornadas Geológicas Bonaerenses, Actas: 37-46, Buenos Aires. [ Links ]
51. Radinsky, L.B. 1987. The Evolution of Vertebrate Design. The University of Chicago Press, 188 p., Chicago. [ Links ]
52. Reed, K.E. 1998. Using large mammal communities to examine ecological and taxonomic structure and predict vegetation in extant and extinct assemblages. Paleobiology 24: 384-408. [ Links ]
53. Sepkoski, J.J. Jr. 1978. Kinematic model of Phanerozoic taxonomic diversity 1: analysis of marine orders. Paleobiology 4: 223-251. [ Links ]
54. Sepkoski, J.J. Jr. 1979. Kinematic model of Phanerozoic taxonomic diversity 2: early Phanerozoic families and multiple equilibria. Paleobiology 5: 222-251. [ Links ]
55. Simpson, G.G. 1950. History of the fauna of Latin America. American Journal of Science 38: 361-389. [ Links ]
56. Soibelzon, L. 2004. Revisión sistemática de los Tremarctinae (Carnivora, Ursidae) fósiles de América del Sur. Revista del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" 6: 107-133. [ Links ]
57. Tonni, E.P. 1985. Mamíferos del Holoceno del Partido de Lobería, Provincia de Buenos Aires. Aspectos paleoambientales y bioestratigráficos del Holoceno del sector oriental de Tandilia y Área Interserrana. Ameghiniana 22: 283-288. [ Links ]
58. Tonni, E.P. 1990. Mamíferos del Holoceno en la Provincia de Buenos Aires. Paula-Coutiana 4: 3-21. [ Links ]
59. Tonni, E.P. and Laza, J.H. 1980. Las aves de la Fauna local Paso de Otero (Pleistoceno tardío) de la provincia de Buenos Aires. Su significación ecológica, climática y zoogeográfica. Ameghiniana 17: 313-322. [ Links ]
60. Van Couvering, J.A.H. 1980. Community evolution in Africa during the Cenozoic. In Berensmeyer, A.K. and Hill, A. (eds.) Fossils in the Making, University of Chicago Press, p. 272-298, Chicago. [ Links ]
61. Vizcaíno, S.F. 2000. Vegetation partitioning among Lujanian (Late Pleistocene-Early Holocene) armored herbivores in the pampean region. Current Research in the Pleistocene 17: 135-137. [ Links ]
62. Vizcaíno, S.F. and Bargo, M.S. 1998. The masticatory apparatus of Eutatus (Mammalia, Cingulata) and some allied genera. Evolution and paleobiology. Paleobiology 24: 371-383. [ Links ]
63. Vizcaíno, S.F., Zárate, M., Bargo, M.S. and Dondas, A. 2001. Pleistocene burrows in the Mar del Plata area (Buenos Aires Province, Argentina) and their probable builders. In Vizcaíno, S.F., Fariña, R.A. and Janis, C. (eds.) Biomechanics and Paleobiology of Vertebrates. Acta Paleontologica Polonica, Special Issue 46(2): 157-169. [ Links ]
64. Vizcaíno, S.F., De Iuliis, G. and Bargo, M.S. 1998. Skull shape, masticatory apparatus, and diet of Vassallia and Holmesina (Mammalia: Xenarthra: Pampatheriidae). When anatomy constrains destiny. Journal of Mammalian Evolution 5: 293-321. [ Links ]
65. Vizcaíno, S.F., Fariña, R.A., Bargo, M.S. and De Iuliis, G. 2004. Phylogenetical assessment of the masticatory adaptations in Cingulata (Mammalia, Xenarthra). Ameghiniana 41: 651-664. [ Links ]
66. Vizcaíno, S.F., Bargo, M.S. and Cassini, G.H. 2006. Dental occlusal surface area in relation to food habits and other biologic features in fossil Xenarthrans. Ameghiniana 43: 11-26. [ Links ]
67. Vizcaíno, S.F., Bargo, M.S. and Fariña, R.A. 2008. Form, Function and Paleobiology in Xenarthrans. In Vizcaíno, S.F. and Loughry, W.J. (eds.) The Biology of the Xenarthra, University Press of Florida, p. 86-99, Gainesville. [ Links ]
68. Webb, S.D. 1976. Mammalian faunal dynamics of the Great American interchange. Paleobiology 2: 216-234. [ Links ]
69. Webb, S.D. 1985. Late Cenozoic mammal dispersal between the Americas. In Stehli, F.G. and Webb, S.D. (eds.) The Great American Biotic Interchange, Plenum Press, p. 201-217, New York. [ Links ]
70. Webb, S.D. 1991. Ecogeography and the great American interchange. Paleobiology 17(3): 266-280. [ Links ]
71. Woodward, J. 1987. Darwin. Editorial Alianza, 141 p., Madrid. [ Links ]
72. Zárate, M.A., Bargo, M.S., Vizcaíno, S.F., Dondas, A. and Scaglia, O. 1998. Estructuras biogénicas en el Cenozoico tardío de Mar del Plata (Argentina) atribuibles a grandes mamíferos. Revista de la Asociación Argentina de Sedimentología 5: 95-103. [ Links ]
Recibido: 5 de agosto de 2008
Aceptado: 5 de octubre de 2008

