










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista de la Asociación Geológica Argentina
Print version ISSN 0004-4822On-line version ISSN 1851-8249
Rev. Asoc. Geol. Argent. vol.65 no.4 Buenos Aires Dec. 2009
Paleoambientes litorales durante el inicio de la trasgresión marina holocena en bahía Lapataia, canal Beagle, parque nacional Tierra del Fuego
Jorge Rabassa1,2, Andrea Coronato1,2, Sandra Gordillo3 , María S. Candel4 y Marcelo A. Martínez4
1 CONICET-CADIC, Ushuaia, Tierra del Fuego, Emails jrabassa@gmail.com, acoro@cadic.gov.ar
2 Universidad Nacional de la Patagonia San Juan Bosco, Sede Ushuaia, Tierra del Fuego.
3 CONICET, CICTERRA, Universidad Nacional de Córdoba, Córdoba. Email gordillosan@yahoo.es
4 CONICET-INGEOSUR, Departamento de Geología, Universidad Nacional del Sur, Bahía Blanca. Emails scandel@uns.edu.ar, martinez@criba.edu.ar
RESUMEN
Los sitios Aserradero Lapataia 1 y 2 presentan depósitos limo-arcillosos portadores de valvas de moluscos: Mulinia edulis, Mytilus chilensis, Aulacomya atra y Yoldia sp. Además predominan los quistes de dinoflagelados, revestimientos de foraminíferos bentónicos y huevos de copépodos. Valvas de Mulinia edulis fueron datadas radiocarbónicamente en 8.094 ± 43 (AA74047) y 8.167 ± 43 (AA74048) a A.P. En Arroyo Baliza depósitos limo-arcillosos son portadores de Venus antiqua, Aulacomya atra, Mytilus chilensis, entre otros. La microflora está dominada por dinoquistes protoperidinioideos, acritarcos y restos de copépodos. Se sugiere ambientes marino-marginales, con baja a moderada salinidad, y altas concentraciones de nutrientes en las aguas superficiales. La datación radiocarbónica sobre Venus antiqua en Arroyo Baliza indica 2.844 ± 34 a A.P. (AA74046). Aserradero Lapataia confirma las edades más antiguas conocidas para la fase estuárica de la transgresión holocena en el canal Beagle. Arroyo Baliza aporta nueva evidencia de la fase regresiva en este canal durante el Holoceno tardío.
Palabras clave: Transgresión holocena; Canal Beagle; Paleoambientes.
ABSTRACT: Littoral paleoenvironments during the beginning of the Holocene marine transgression in Lapataia Bay, Beagle Channel, National Park of Tierra del Fuego. The Aserradero Lapataia 1 and 2 sites present clayey silty sediments including mollusk shell; Mulinia edulis, Mytilus chilensis, Aulacomya atra and Yoldia sp. Besides, dinoflagelate cysts, covers of benthonic foraminifera and copepod eggs are dominant. The microflora is represented by sporomorphs and fresh to salty water microplancton. Radiocarbon dates on Mulinia edulis shells yielded ages of 8094 ± 43 (AA74047) and 8167 ± 43 (AA74048) yr B.P. At Arroyo Baliza, clayey silts were found including Venus antiqua, Aulacomya atra, Mytilus chilensis, among others, accompanied by acritarchs and remnants of copepods. This association suggests marginal marine environments, with low to moderate salinity, and high nutrient concentration in the surface waters. Radiocarbon dating on Venus antiqua shells yielded 2844 ± 34 yr B.P. (AA74046). Marine deposits of the Aserradero Lapataia sites confirm the oldest ages known for the estuarine phase of the Holocene transgression in the Beagle Channel. Arroyo Baliza provide new evidence for the Late Holocene regressive phase in the Beagle Channel.
Keywords: Holocene transgression; Beagle Channel; Paleoenvironments.
INTRODUCCIÓN
Este trabajo presenta los resultados del estudio geomorfológico y paleontológico de dos nuevas localidades que revelan las condiciones paleoambientales durante el inicio de la transgresión holocena en el Canal Beagle, Tierra del Fuego. En la costa sur de la Bahía Lapataia se estudiaron los sitios Aserradero Lapataia 1 y 2 (54o51´S, 68o34´O) y Arroyo Baliza (54o51´S, 68o33´O; Fig. 1a). Los dos primeros forman la ribera aluvial del arroyo Los Castores, mientras que el sitio Arroyo Baliza forma parte del cauce actual, en su desembocadura en el sector costero intermareal. Ambas localidades están bajo influencia de las mareas.
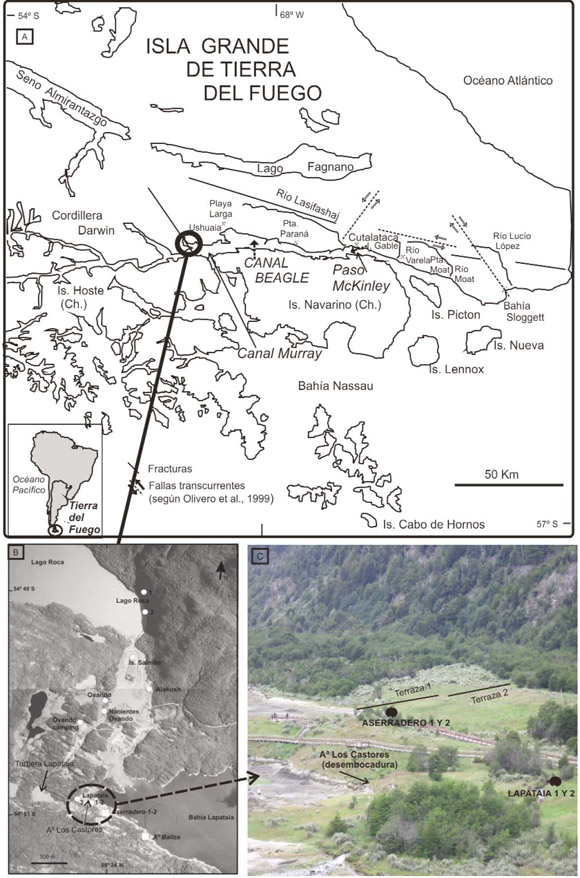
Figura 1: Ubicación del área de trabajo: a) Sitios de interés a lo largo del canal Beagle y ubicación de la bahía Lapataia; b) Emplazamiento de los sitios Aserradero 1 y 2 y Arroyo Baliza en Bahía Lapataia y Lago Roca; con círculos se indican otros sitios estudiados previamente; c) vista hacia el sector de Bahía Lapataia donde se ubican el sitio Aserradero Lapataia y los sitios Lapataia 1-2.
Antecedentes
El ingreso del agua marina a la artesa glacial en la que hoy se emplaza el canal Beagle ha sido objeto de interés científico desde las primeras investigaciones en temas del Cuaternario del extremo sur de Sudamérica. La región fue cubierta por glaciares de descarga durante la Última Glaciación Pleistocena (ca. 25 ka A.P.), los cuales exhondaron los valles pre-existentes. Se postula que a medida que los glaciares retrocedieron, varios cuerpos de agua de fusión se formaron en las zonas más deprimidas de la artesa (Caldenius 1932, Isla et al. 1999, Bujalesky et al. 2004, Bujalesky et al. 2008) dando lugar a lagos glaciarios hasta tanto se produjo el avance definitivo del mar. El ascenso del nivel del mar en tiempos postglaciales formó la intrincada red de fiordos y canales marinos de la costa sudoccidental de Sudamérica, desarrollando en algunos sectores secuencias de playas marinas elevadas. Las primeras observaciones en la región estudiada corresponden a Halle (1910) quien reconoció la existencia de costas aterrazadas a lo largo del Canal Beagle atribuyéndole un origen marino. Urien (1968) reafirmó la existencia de playas elevadas mediante el descubrimiento de tres niveles marinos en la bahía Golondrina, ubicada 20 km al este del área de estudio. En el extremo sur de esta bahía se ubica la Punta Pingüinos, límite sudoccidental de la Península Ushuaia, donde Auer (1959, 1974) identificó un estrato de gravas, presumiblemente de origen marino, dada la presencia de huesos de ballena y restos de Mytilus sp., a 5-6 m s.n.m. Estos depósitos suprayacen a niveles de limos arcillosos con desarrollo de varves y a niveles de turba, dispuestos sobre el till basal que conforma una morena. La madera contenida en los niveles limo-arcillosos fue posteriormente fechada por radiocarbono, arrojando edades de ca. 7,0-7,6 ka A.P (Auer 1974, Rabassa et al. 1986). El fechado de los niveles subyacentes a las gravas predatan el desarrollo de la supuesta playa marina. Posteriormente, Porter et al. (1984) postularon un probable origen antropogénico al nivel portador de huesos y valvas, sin confirmar el origen marino de las gravas. A lo largo de la costa norte del canal Beagle, entre bahía Lapataia (68º34´O) e isla Gable (67º 23´ O, Fig. 1a), se han determinado tres niveles de playas holocenas ubicados a 8-10, 4-6 y 1.5-3 m s.n.m. actual, respectivamente (Rabassa et al. 1986, Porter et al. 1984, Gordillo et al. 1993, Gordillo 1993, Rabassa et al. 2000). Hacia el este, en la zona de Río Varela (Fig. 1a), los depósitos marinos holocenos se hallan sumergidos bajo la línea de mareas (Rabassa et al. 2004) o sepultados por secuencias fluviales (Grill et al. 2002) en las cercanías a las desembocaduras de los ríos. En base a estas evidencias, Bujalesky et al. (2004) postularon un comportamiento tectónico diferencial a lo largo de la costa norte del Canal Beagle, indicando al sector occidental del sector argentino del canal Beagle como el de mayor actividad, debido a la presencia de playas ascendidas hasta ca. 10 m s.n.m. (Rabassa et al. 1986, Gordillo et al. 1993). El descubrimiento de depósitos marinos holocenos por debajo del nivel del mar actual, ubicados al este de la falla regional Lasifashaj, de rumbo NO, debilita la propuesta de Mörner (1991) sobre la inexistencia de niveles marinos continuos a lo largo del canal Beagle. No obstante, su teoría sobre la fuerte incidencia de la componente geotectónica sobre el ascenso glacioeustático es aplicable al sector occidental, donde se ubica el área de estudio. La curva de variación del nivel del mar para el Canal Beagle (Porter et al. 1984, Rabassa et al. 1986, Gordillo et al. 1993, Fig. 2) muestra que los niveles más altos se alcanzaron hacia 5.000-6.000 a 14C A.P., interpretándose que los sitios de mayor antigüedad podrían representar condiciones de incremento del nivel relativo del mar con predominio de la recuperación glacioisostática, mientras que los depósitos más elevados y más jóvenes corresponderían a procesos de alzamiento tectónico dominante. Debe puntualizarse que a lo largo del canal Beagle, no se han encontrado depósitos cuyos fechados radiocarbónicos indiquen edades mayores a los de la bahía Lapataia. El área de ingreso del agua de mar a la artesa glacial y cuencas lacustres del paleocanal Beagle es aún incierta. El proceso de inundación podría haber ocurrido a través del canal Murray (Fig. 1a), ubicado inmediatamente al sur de bahía Lapataia comunicando la zona con el Pasaje Drake (Rabassa et al. 1986, 2000). Por otra parte, una vez que la recuperación glacioisostática del mar alcanzó la isobata de 30 m, las aguas marinas pudieron haber ingresado desde el Océano Atlántico, superando el umbral del Paso McKinley (Bujalesky 2007) (Fig. 1A) y ocupando las cuencas lacustres ubicadas entre esta localidad y bahía Ushuaia (Isla et al. 1999).
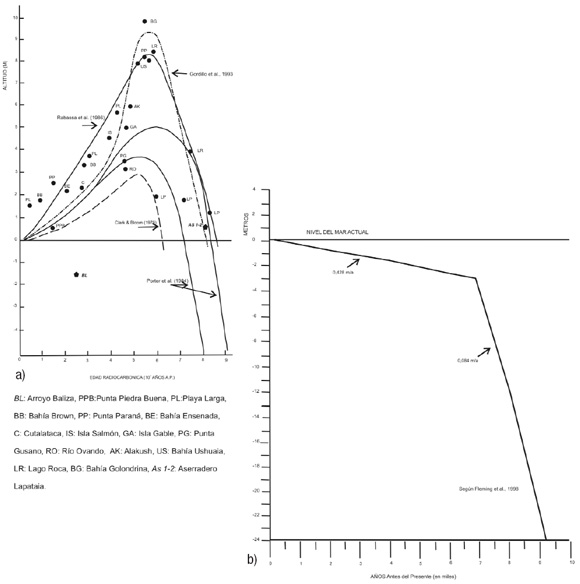
Figura 2: Curvas del nivel relativo del mar durante el Holoceno: a) Curva para el Canal Beagle, según Porter et al. (1984), Rabassa et al. (1986) y Gordillo et al. (1993). Para su interpretación, véase este último trabajo. En ella se agregan los sitios estudiados en este trabajo; b) Curva de ascenso eustático global durante el Holoceno, modificada de Flemming et al. (1998).
Hasta el presente, los estudios sobre paleomicroplancton marino de pared orgánica en la isla Grande de Tierra del Fuego, para el área de bahía Lapataia, corresponden a Borromei et al. (1997), Borromei y Quattrocchio (2001, 2007), en los cuales se registraron quistes de dinoflagelados como Spiniferites sp., Operculodinium centrocarpum, Chystroiesphaeridia sp., Cleistosphaeridium sp., Hystrichokolpoma sp. y Pixidinopsis sp., indicando condiciones inestables, con salinidades variables y temperaturas frías del agua de mar. En el área de Río Ovando, Candel et al. (2009) mencionan la presencia del "complejo Islandinium-Echinidinium", Selenopemphix quanta, Seleno-pemphix spp., Brigantedinium spp., Polykrikos schwartzii, P. kofoidii, Operculodinium cf. centrocarpum, cf. Pentapharsodinium dalei. Esta asociación de dinoquistes presente en esta secuencia marina refleja condiciones ambientales de fiordos cercanos al campo de hielo de la Cordillera Darwin, sector occidental de la isla Grande de Tierra del Fuego, los cuales fueron afectados por descarga de derretimiento glacial con baja salinidad y alto aporte de nutrientes, debido a la descarga de agua dulce. En depósitos aflorantes en la localidad de Río Varela (Grill et al. 2002) se registraron Brigantedinium spp., Selenopemphix nephroides, Operculodinium centrocarpum y Spiniferites sp., sugiriendo ambientes neríticos con alto aporte de nutrientes. Resultados preliminares de este trabajo han sido presentados en Rabassa et al. (2008).
Las primeras observaciones de moluscos fósiles cuaternarios en playas elevadas del extremo austral de América fueron hechas por Darwin (1846). Posteriormente, fue Hägg (en Halle 1910) quien proporcionó el primer listado de moluscos holocenos de una terraza elevada en isla Gable sobre el canal Beagle. Más recientemente, nuevos listados para el canal Beagle fueron proporcionados por Gordillo (1993, 1999). Los estudios sobre estos moluscos han permitido abordar aspectos tafonómicos (Gordillo 1992, 2007, Cárdenas y Gordillo 2009), paleoecológicos (Gordillo 1991, 1999), y realizar reconstrucciones paleoambientales (Gordillo et al. 1993, 2005).
MATERIALES Y MÉTODOS
Se estudió el área en base a fotografías aéreas escala 1:20.000 del Servicio de Hidrografía Naval Argentina, Año 1970 (C1, ffaa 30-31-32). Se realizaron perfiles topográficos mediante transectas con GPS tipo Garmin Vista con altímetro barométrico, ajustado al nivel medio de mareas. Se describió la composición sedimentaria de los perfiles. Se tomaron muestras de sedimentos portadores de microplancton y valvas de moluscos en los perfiles expuestos denominados Aserradero Lapataia 1 y 2, ubicados en la desembocadura del Arroyo Los Castores (Fig. 1 b y c), y en el área intermareal de la desembocadura del Arroyo Baliza, donde se realizó una calicata de 1,5 m (Fig. 1 b). Los muestreos se realizaron durante bajamar, momento durante el que quedan expuestos los perfiles o se puede realizar muestreo en calicatas.
Las muestras micropaleontológicas obtenidas fueron procesadas para su análisis palinológico, según las técnicas de Heusser y Stock (1984), con modificaciones de Dale (1976), para la preservación de los quistes de dinoflagelados de pared orgánica. Las muestras fueron tratadas con ácidos clorhídrico y fluorhídrico y no se realizó oxidación ni acetólisis para evitar la pérdida de aquellos quistes de dinoflagelados de pared orgánica (dinoquistes) de protoperidiniáceos más frágiles. Esporas exóticas de Lycopodium fueron incorporadas para permitir el cálculo de la concentración de palinomorfos por gramo de sedimento seco (Stockmarr 1971). El residuo fue filtrado a través de mallas 150 µm y 10 µm y montado en glicerinagelatina sobre portaobjetos. Los preparados palinológicos están archivados en el Laboratorio de Palinología, Universidad Nacional del Sur, Bahía Blanca, Argentina, bajo la denominación UNSP seguido por la sigla respectiva de la sección estudiada: As1 (Aserradero Lapataia 1), As2 (Aserradero Lapataia 2) y Bz (Arroyo Baliza). Las muestras fueron estudiadas utilizando un microscopio de luz transmitida Olympus BH2 con magnificación de x 400 a x 1000. Las microfotografías fueron tomadas con una cámara digital Olympus y los palinomorfos seleccionados están localizados por coordenadas de la reglilla England Finder. El análisis palinofacial fue llevado a cabo siguiendo a Tyson (1995) y Batten (1996), reconociéndose cinco tipos de materia orgánica palinológica (MOP): fitoclastos translúcidos, fitoclastos opacos, materia orgánica amorfa (AOM), palinomorfos y zooclastos. Para el análisis palinofacial se realizaron recuentos de 500 partículas mayores a 10 µm con magnificación x 400 y los distintos tipos de materia orgánica palinológica fueron expresados como frecuencias relativas. Los recuentos palinológicos se realizaron sobre un mínimo de 200 palinomorfos. Además, para garantizar la presencia de los taxa menos representados, los preparados fueron recorridos en su totalidad.
Los palinomorfos fueron asignados a distintas familias y/o grupos supragenéricos de esporomorfos y palinomorfos acuáticos. Dentro de los primeros se encuentran esporas (hongos, Lycopodiaceae, Polypodiaceae); granos de polen de árboles (Nothofagus tipo dombeyi) y granos de polen de hierbas y arbustos (Poaceae, Misodendrum, Empetrum, Gunnera, Asteroideae, Caryophyllaceae, Apiaceae, Cyperaceae). Los palinomorfos acuáticos están representados por algas dulceacuícolas a marino-salobres, dinoquistes, acritarcos, huevos de copépodos, foraminíferos. Las frecuencias relativas de las asociaciones microflorísticas se realizaron sobre la base de los recuentos palinológicos y se obtuvieron porcentajes para cada grupo de palinomorfos. El diagrama de porcentajes de palinofacies y palinomorfos se realizó con el programa Tilia 2.0.b.4 (Grimm 1991) y TGview 2.0.2 (Grimm 2004). Los ejemplares de malacofauna obtenidos con su sedimento portador fueron lavados utilizando tamices de abertura de malla fina para retener ejemplares pequeños y/o juveniles. Luego del lavado y cepillado, los moluscos fueron acondicionados, numerados y depositados en el Repositorio del Centro de Investigaciones Paleobiológicas de la Universidad Nacional de Córdoba bajo la sigla CEGH-UNC. Sobre la base del material colectado se analizó la composición taxonómica y los principales atributos tafonómicos (fragmentación, abrasión, disolución, bioerosión e incrustación) para caracterizar y evaluar el tipo de asociación.
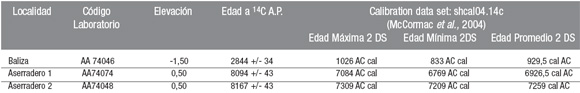
Las dataciones radiocarbónicas sobre valvas de moluscos fueron realizadas en el AMS Facility de la University of Arizona, Tucson, U.S.A. La calibración se realizó con el programa CalPal (Stuiver y Reimer 1993), set de calibración para sedimentos marinos del Hemisferio Sur (shcal04.14c; McCormac et al. 2004). A las edades radiocarbónicas presentadas en el Cuadro 1 se les debe considerar un efecto reservorio de ca. 556 ± 60 años, según lo analizado por Albero et al. (1988).
CUADRO 1: Fechados radiocarbónicos en las localidades estudiadas. En las muestras Aserradero 1 y 2 las dataciones radiocarbónicas se realizaron sobre valvas de Mulinia edulis, mientras que en las muestras de Baliza fueron realizadas sobre valvas de Venus antiqua. A las edades radiocarbónicas se les debe considerar un efecto reservorio de ca. 556 ± 60 años (Albero et al. 1988).
RESULTADOS
Los depósitos marinos del sitio Aserradero Lapataia se hallan adosados al nivel inferior de un sistema de terrazas de 11 m s.n.m. (Terraza 1) y 6 m s.n.m. (Terraza 2)( Fig. 1 c), que se apoyan al pie de la ladera del cerro Mesa Real (700 m s.n.m.). Si bien los depósitos que las conforman no son visibles, en sectores costeros se observan afloramientos de esquistos cuarzosos de la Formación Lapataia (Borrello 1969), mientras que en cercanías a los depósitos estudiados se disponen bloques erráticos resultantes de la acción glacial previa a la trasgresión marina. La relación topográfica con la morfología glacial de la zona permite interpretar a estas terrazas como parte del sistema de drenaje de los glaciares en retroceso, el cual fluía en dirección O-E al pie del Cerro Mesa Real (Gordillo et al. 1993) hasta desembocar en la actual bahía Lapataia. El alineamiento de turberas formadas a lo largo de este sistema de drenaje evidencia la presencia de lagos someros luego del retiro de los hielos. La turbera Lapataia (Fig. 1 b) presenta una edad basal de 10080 años 14C A.P. (Heusser 2003) indicando no sólo la edad mínima del retiro del hielo en la zona, sino también el inicio de la colmatación de los lagos someros formados durante la fase de deglaciación (Gordillo et al. 1993). El sitio Arroyo Baliza forma parte del cauce en cercanías a la desembocadura.
Los depósitos marinos del sitio Aserradero Lapataia 1 y 2, datados en ca. 8.100 años 14C A.P., presentan similares edades radiocarbónicas y elevación sobre el nivel del mar actual (+0,50 m; 8.094 ± 43 y 8.167 ± 43 años 14C A.P., respectivamente; Cuadro 1), y confirman el ingreso del mar holoceno en bahía Lapataia, demostrando que el agua de mar afectó un área mayor a la ya conocida en dicha bahía (Rabassa et al. 1986). La relación altitud/ edad radiocarbónica de los sitios Aserradero es semejante a los sitios Lapataia 1, 2 y 3 establecidos en la curva de variación del nivel del mar holoceno para el canal Beagle (Fig. 2) aunque con un defasaje de casi 15 m con respecto a la curva de nivel del mar por ascenso eustático global planteada por Flemming et al. (1998). El sitio Arroyo Baliza (- 1,50 m; 2844 ± 34 a 14C A.P.), en cambio, es el primer sitio que muestra valores por debajo del nivel promedio actual del mar para el Holoceno tardío; sin embargo, está ubicado apenas 0,50 m por debajo del valor establecido por la curva eustática global (Flemming et al. 1998).
MICROPALEONTOLOGÍA
Los palinomorfos predominantes en el sitio Aserradero Lapataia 1 son los esporomorfos, principalmente granos de polen de Nothofagus tipo dombeyi (42%), Poaceae (4,3%) y esporas de hongos (15%) (Fig. 3). Otros grupos polínicos muestran una representación inferior al 2%: Asteroideae (1,8%), Polypodiaceae (1,8%), Misodendrum y Empetrum (1,2%) y Caryophyllaceae (0,6%). Los palinomorfos acuáticos están representados por dinoquistes como Brigantedinium spp. (9,3%), "round brown" indet. (3,7%), Selenopemphix quanta (1,8%) y en menor proporción Echinidinium spp. (1,2%), Echinidinium granulatum (0,6%), Brigantedinium cariacoense (0,6%) y Lejeunecysta sp. (0,6%). También se encuentran presentes acritarcos como Halodinium sp. (3,7%) y Micrhystridium sp. (0,6%), huevos de copépodos (4,9%) y foraminíferos (4,3%). Respecto a la palinofacies, se observó alto predominio de fitoclastos translúcidos (86,3%) principalmente nobioestructurados pseudoamorfos y gelificados, de colores hialinos y muy degradados, algunos de los cuales muestran una transición a materia orgánica amorfa (Fig. 3). Los fitoclastos opacos se encuentran en muy baja proporción (0,8%) y predominan aquellos con forma de tablillas en tamaños variables. La materia orgánica amorfa está presente en un 10%, representada por amorfo de tipo esponjoso a una transición con amorfo granular. Se observa pirita framboidal en forma diseminada y formando agregados (Fig.4). En el sitio Aserradero Lapataia 2, la asociación microflorística está representada principalmente por Nothofagus tipo dombeyi (55,8%), seguido también en menores proporciones por esporas de hongos (16,1%), Poaceae (5%), Polypodiaceae (4,7%), Lycopodiaceae y Asteroideae (2,6%), Empetrum (2,3%), Cyperaceae (0,9%) y Apiaceae (0,3%) (Fig. 3). El microplancton se registra en muy bajas proporciones reconociéndose dinoquistes, Brigantedinium spp. (2%), "round brown" indet. (2%) y Echinidinium granulatum (0,3%), acritarcos, Halodinium sp. (1,8%) y algas de agua dulce a marino-salobre como Botryococcus spp. y Tasmanites sp., con el 0,6% en ambos casos. Restos de microorganismos bentónicos, como huevos de copépodos y revestimientos de caparazones de foraminíferos, están presentes con 1,8 y 0,6%, respectivamente (Fig. 4). La palinofacies está dominada por fitoclastos translúcidos (79%) fundamentalmente no-bioestructurados degradados de colores ámbar a castaño oscuro, mientras que los bioestructurados están representados por tejidos, traqueidas e hifas de hongos. La materia orgánica amorfa se presenta con 11%, de tipo esponjoso y granular con inclusiones de pirita. La pirita de tipo framboidal observada se encuentra concentrada sobre los fitoclastos y está presente en menor proporción que en el nivel anterior.
En el sitio Arroyo Baliza, la palinoflora se caracteriza por altos valores de Nothofagus tipo dombeyi (88,6%), acompañado también por Polypodiaceae (9,5%), esporas de hongos (8,3%), Poaceae (5,5%), Asteroideae (3,6%) y Empetrum (1,8%). Con bajas proporciones se registraron Lycopodiaceae (0,9%), Misodendrum y Gunnera (0,6%) y Apiaceae (0,3%) (Fig. 3). El microplancton está dominado por dinoquistes, los cuales evidencian los valores más altos de diversidad registrados (seis especies). Se identificó Brigantedinium spp. (3%), "castaño redondeado indet." (1,8%), Echinidinium granulatum (1,2%) y en proporciones inferiores al 1%: Selenopemphix quanta (0,6%), Echinidinium spp. (0,3%), y Quinquecuspis concreta (0,3%). Completan la asociación, acritarcos, Halodinium sp. (1,8%) y algas dulceacuícolas a marino-salobres como Botryococcus spp., Zygnema sp. y Tasmanites sp. con 0,92%, 0,30% y 0,30%, respectivamente. Se registraron además restos de microorganismos bentónicos, tales como huevos de copépodos (1,8%) y foraminíferos (0,3%). La palinofacies muestra una dominancia de fitoclastos translúcidos (88,3%) principalmente no-bioestructurados degradados y de colores pálidos, muchos de ellos en transición a amorfo; dentro de los bioestructurados predominan las hifas de hongos y tejidos de colores castaños. La materia orgánica amorfa está presente en un 6,4%, representado por amorfo de tipo granular acompañado en menor proporción por amorfo de tipo esponjoso. La pirita framboidal se observa en forma diseminada o como inclusiones sobre los fitoclastos (Fig. 4).
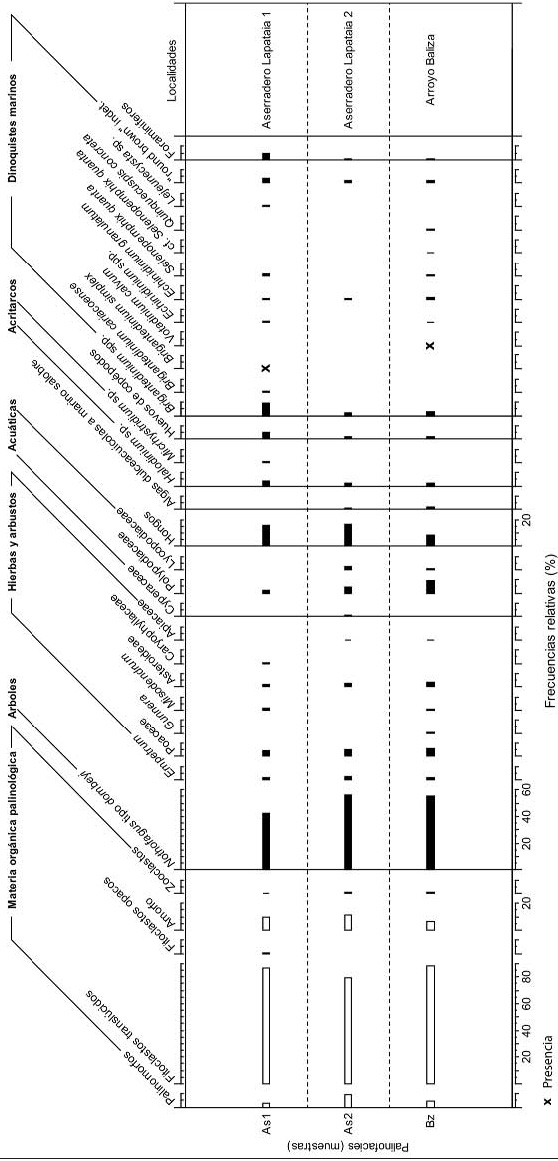
Figura 3: Diagrama de frecuencias relativas (%) de las asociaciones microflorísticas y palinofacies de Aserradero 1, Aserradero 2 y Arroyo Baliza.
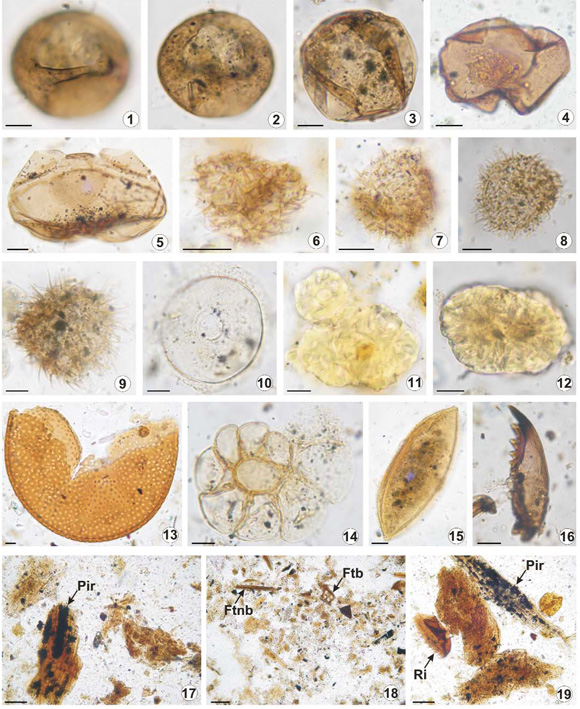
Figura 4: Palinomorfos acuáticos y Palinofacies de Aserradero 1 (AS1), Aserradero 2 (AS2) y Arroyo Baliza (BZ). Escala 1 a 16: 10 µm; 17-19: 30 µm y 18: 50 µm. Número de muestra seguido por coordenadas de England Finder. 1-2) Brigantedinium simplex. UNSP AS1 3053, P46/2; 3) Brigantedinium spp. UNSP BZ 3055, O49; 4) Quinquecuspis concreta. UNSP AS1 3053; 5) Votadinium calvum. UNSP BZ 3055, G49/3; 6) Echinidinium granulatum. UNSP AS2 3054, J37/3; 7) Echinidinium granulatum. UNSP BZ 3055, Z23/3; 8) Echinidinium granulavtum. UNSP BZ 3055, S33; 9) Selenopemphix quanta. UNSP AS13053, M52/2; 10) Halodinium sp. UNSP BZ 3055, M38/4; 11) Botryococcus spp. UNSP AS2 3054, T41/3; 12) Botryococcus spp. UNSP AS2 3054, C10/4; 13) Tasmanites sp. UNSP AS2 3054, X22; 14) Foraminífero. UNSP AS1 3053, C18/4; 15) Huevo de copépodo. UNSP AS1 3053, A16; 16) Pieza de copépodo. UNSP BZ 3055, Z33/3; 17-19) Palinofacies de AS1 (x400) con detalle de fitoclastos no-bioestructurados (Ftnb) con inclusiones de pirita (Pir) y restos de invertebrados (Ri). UNSP AS1 3053, T20; J27/1; 18) Palinofacies de BZ (x200) con detalle de fitoclastos bioestructurados (Ftb) y no-bioestructurados (Ftnb). UNSP BZ 3055, B52.
MACROPALEONTOLOGÍA
Los invertebrados hallados en los sitios estudiados son principalmente moluscos bivalvos y gastrópodos, seguidos de crustáceos cirrípedos. Entre los moluscos, y según sus requerimientos ecológicos (i.e. sustrato, modo de vida, locomoción, tipo de alimentación), hay una gran variedad de formas (Cuadro 2), aunque se destacan en su abundancia numérica los bivalvos suspensívoros y los gastrópodos depredadores.
La asociación faunística del sitio Aserradero 2 está compuesta solamente por dos especies: la almeja Mulinia edulis y el mejillón Mytilus chilensis. Ambos bivalvos cohabitan en la zona intermareal, aunque uno (M. edulis) es excavador y se entierra y el otro (M. chilensis) se fija como epifauna sobre sustratos duros. Estas especies son capaces de vivir en ambientes estuáricos y/o en planicies de mareas, caracterizados por amplias fluctuaciones en la cantidad de materia orgánica en suspensión (Navarro et al. 2003), y su presencia en el canal Beagle indicaría condiciones estuáricas para el Holoceno temprano.
CUADRO 2: Caracterización ecológica de las principales especies de moluscos
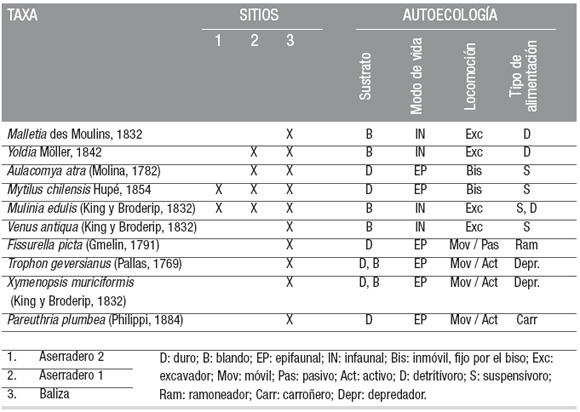
En el sitio Aserradero 1, la asociación faunística está compuesta por las dos especies anteriores (Mulinia edulis, Mytilus chilensis), a las que se le suman otras dos: Aulacomya atra y Yoldia sp. Estos últimos bivalvos también tienen plasticidad ecológica y toleran ambientes variables en salinidad y materia orgánica. En el sitio Arroyo Baliza, del Holoceno tardío, la asociación de moluscos se compone de un mayor número de especies y entre las más comunes figuran Trophon geversianus, Xymenopsis muriciformis y Pareutrhia plumbea, entre los gastrópodos, y Venus antiqua, Aulacomya atra, Mytilus chilensis, Mulinia edulis, Yoldia sp. y Malletia sp., entre los bivalvos (Cuadro 2). Todas éstas son especies bentónicas típicas de aguas someras de la Región Magallánica. Desde el punto de vista tafonómico estos ensambles de conchillas representan, en su conjunto, asociaciones para-autóctonas o autóctonas (ver terminología en Kidwell et al. 1986) y sus integrantes, en general, se hallan bien preservados, con leves alteraciones tafonómicas (Fig. 5, a-j). Los bivalvos presentan sus valvas articuladas, o desarticuladas pero en asociación con la valva opuesta. La fragmentación es baja, salvo en los mitílidos (Fig. 5, c-d). En el conjunto faunístico las conchillas no evidencian signos de disolución ni de abrasión. La bioerosión también es baja, pudiéndose mencionar la presencia de una valva perforada (Fig. 5, f), lo que se atribuye a la actividad de depredación de un gastrópodo perforador (Gordillo 1994). Finalmente, y en relación a la presencia de organismos incrustantes y/o epibiontes, algunas conchillas presentan remanentes de crustáceos cirrípedos (Fig. 5, g-h).

Figura 5: Principales moluscos hallados en los perfiles holocenos de Bahía Lapataia: a) Yoldia sp. (vista exterior, valva derecha, 17 mm largo), Baliza, CEGH-UNC 23363. b) Malletia sp. (vista exterior, valva izquierda, 30 mm largo), Baliza, CEGH-UNC 23364. c) Mytilus chilensis (vista interior, fragmento, valva izquierda, 20 mm largo), Baliza, CEGH-UNC 23370. d) Aulacomya atra (vista interior, fragmento, valva izquierda, 20 mm largo), Baliza, CEGH-UNC 23369. e) Mulinia edulis (vista exterior, valva derecha, 54 mm largo), Aserradero 1, CEGH-UNC 23386. f) Venus antiqua con perforación cilíndrica (vista exterior, valva derecha, 59 mm largo), Baliza, CEGH-UNC 23374. g) Fisurella picta con marcas de cirrípedos (vista dorsal, 38 mm largo), Baliza, CEGH-UNC 23373. h) Xymenopsis muriciformis con cirrípedos (vista lateral, 36 mm alto), Baliza, CEGH-UNC 23371. i) Trophon geversianus (vista lateral, 44 mm alto), Baliza, CEGH-UNC 23377. j) Pareuthria plumbea (vista lateral, 28 mm alto), Baliza, CEGH-UNC 23380.
Las asociaciones microflorísticas de los tres sitios estudiados se caracterizan por un predominio en la concentración de Nothofagus tipo dombeyi acompañada de Poaceae, Asteroideae y Cyperaceae, Empetrum, Polypodiaceae, Misodendrum y Lycopodiaceae entre otros. Esto indica el desarrollo de un ambiente con bosque abierto y sectores con herbáceas, bajo condiciones de clima frío, húmedo y variable durante el Holoceno.
Cabe destacar el registro de dos especies de dinoquistes (Quinquecuspis concreta y Lejeunecysta sp.) en las muestras de Aserradero Lapataia 1 y 2 y Arroyo Baliza, no halladas previamente en las secciones fósiles de la zona.
Los taxones dominantes de dinoquistes marinos sugieren la ocurrencia de ambientes marino-marginales. La asociación de dinoquistes presentes en estos niveles refleja condiciones ambientales de fiordos cercanos a campos de hielo terrestres, afectados por descarga de agua de fusión glacial, con salinidad variable. La descarga de agua dulce al medio marino también es confirmada por la presencia de algas dulceacuícolas a salobres como Zygnematáceas y Botryococcus. Esta descarga y condiciones de circulación restringida propia de ambientes de fiordos favorecen el desarrollo de salinidad del agua de mar sub-normal, lo cual es avalado por el registro de algas de Prasinophytas (Tasmanites). Los fitoclastos no-bioestructurados, mayormente degradados constituyen la materia orgánica palinológica predominante en las tres palinofacies, junto con materia orgánica amorfa de tipo granular y esponjoso. También se encuentran presentes fitoclastos bioestructurados tales como cutículas, traqueidas e hifas de hongos. Esto sugiere el gran aporte de origen continental al medio marino, evidenciando la materia orgánica palinológica distancias y/o tiempos de transporte relativamente cortos con respecto a la fuente de origen continental, dado el amplio predominio de fitoclastos translúcidos por sobre los opacos. La pirita se encuentra presente en todas las palinofacies indicando condiciones ambientales reductoras asociada probablemente a la escasa circulación; sin embargo, el predominio de fitoclastos degradados sugiere que no siempre prevalecieron estas condiciones. La dinámica misma de los ambientes marino-marginales como fiordos o estuarios produce una preservación selectiva concentrando aquellos constituyentes más resistentes.
Luego del retroceso general del hielo, que en el canal Beagle tuvo lugar hacia los 10,8 ka 14C A.P. (Rabassa et al. 1986), la invasión marina fue acompañada hacia los 8,2 ka 14C A.P. por la primera aparición de dos especies euritópicas: Mulinia edulis y Mytilus chilensis, seguida de la adición temprana de otras dos especies: Aulacomya atra y Yoldia sp., que hacen su aparición hacia los 8,1 ka 14C A.P. Todas estas especies conforman un grupo pionero (Cuadro 3) caracterizado por su plasticidad ecológica, ya que vivieron en un período con gran aporte de agua dulce por fusión glacial, es decir condiciones de baja salinidad y fluctuaciones en la depositación y redepositación de materia orgánica. Posteriormente, y por la incorporación progresiva de agua marina, aparecen otras especies, dando lugar a un grupo variado, conformado por asociaciones faunísticas más diversificadas. En un trabajo previo realizado en esta región (Gordillo 1999) se describen diferentes asociaciones de moluscos holocenos desarrollados en épocas en que las condiciones marinas ya estaban plenamente establecidas, siendo la más antigua la asociación diversificada hallada en el lago Roca, que se corresponde con una edad cercana a los 7,5 ka 14C A.P. Además del grupo pionero, en el área estudiada también se ha preservado un grupo variado (Cuadro 3) compuesto por diversas especies, presente hacia los 2,8 ka 14C A.P. en un ambiente con condiciones similares a las actuales. La diversidad de asociaciones faunísticas del canal Beagle durante todo el Holoceno ya fue previamente considerada por Gordillo et al. (2005) y se atribuyó a la combinación de dos razones: la sucesión faunística gradual que tiene lugar en un ambiente marino previamente englazado, sumada a la heterogeneidad de hábitats que alberga el canal Beagle, lo que posibilita el desarrollo de una gran variedad de ensambles o asociaciones de moluscos que varían de una localidad a otra. Las condiciones paleoambientales indican que este sector del canal Beagle ya presentaba condiciones de ambiente estuárico de baja energía hacia inicios del Holoceno, en sincronía con el desarrollo de la transgresión marina a nivel planetario. No obstante, la altura actual de los depósitos marinos holocenos presenta condiciones específicas a lo largo del canal Beagle. La figura 2 muestra el defasaje entre la curva de ascenso eustático planetario y las evidencias para esta región, indicando una fuerte condicionante tectónica y/o recuperación glacioeustática a lo largo de las costas del canal Beagle. Según Flemming et al. (1998), durante el período 9,5-7,0 ka A.P., el nivel del mar post-glacial sufrió un ascenso sostenido desde los -24 m hasta los -3 m de profundidad, lo cual indica una tasa de ascenso del orden de 0,084 m/a. En el área de estudio, los depósitos marinos del Holoceno temprano se hallan entre 0,50-1,5 m por encima del nivel del mar actual.
CUADRO 3: Sucesión faunística de moluscos durante el Postglacial en Bahía Lapataia, Canal Beagle.

La tasa de ascenso del nivel del mar a escala global habría acelerado su ritmo a 0,428 m/a a partir de los 7,0 ka A.P. hasta alcanzar el nivel actual.
Los fechados radiocarbónicos presentados en este trabajo confirman plenamente las edades mínimas de penetración de las aguas marinas en la paleoartesa del canal Beagle, y posterior ascenso hasta alcanzar posiciones del nivel relativo del mar equivalentes a los actuales, que fueran informadas en trabajos anteriores (Rabassa et al. 1986, Gordillo et al. 1993). Estas edades, cercanas a 8,2 ka 14C A.P. (Cuadro 1), corresponden a posiciones del nivel del mar global cercanas a -20 m por debajo del nivel del mar actual (Fig. 2B). Sin embargo, estos depósitos se encuentran hoy a cotas cercanas al nivel del mar actual, demostrando que la región ha sufrido un claro ascenso de, por lo me-nos, unos 20-25 m en los últimos 9000 años, a una tasa de ascenso continental de alrededor a 2,0-2,5 mm/año. Esta tasa de elevación es muy fuerte y se asemeja más a tasas de elevación tectónica que a recuperación glacioisostática, como fuera sugerido por Rabassa et al. (2004), de acuerdo al comportamiento esperado del sistema de fallas transcurrentes del canal Beagle, con marcada componente vertical en el sector occidental. Sin embargo, si existe una componente de recuperación glacioisostática regional, es aún imposible discernir su magnitud en relación al ascenso regional total.
Los resultados micro- y macropaleontológicos presentados aquí demuestran además que los ambientes dominantes en el canal Beagle en el Holoceno temprano eran de tipo estuárico, de baja salinidad. Estos ambientes habrían sido el resultado de la mezcla de las aguas marinas en ascenso con lagos de origen glacial que se habrían desarrollado en la depresión del Beagle luego de la recesión definitiva de los glaciares (ca. 11 ka A.P.; Rabassa et al. 2000). Estos ambientes de baja salinidad habrían sobrevivido por al menos varios milenios, a medida que la influencia de aguas marinas fue expandiéndose, por el ascenso eustático global. Se trataba seguramente de cuerpos de agua salobre, profundos, con escasa biodiversidad. La información disponible hasta el momento no permite establecer si existía estratificación de las aguas en bahía Lapataia. El aumento de la salinidad fue acompañado por un crecimiento en la biodiversidad, hasta que hacia 7,0 ka A.P. se habría instalado definitivamente el ambiente marino como el que existe en la actualidad. La evidencia de campo disponible sugiere que el ascenso tectónico en este sector del canal Beagle ha continuado ininterrumpidamente hasta el presente. En caso que hubiera existido recuperación glacioisostática en esta región, ésta habría tenido lugar exclusivamente en el Tardiglacial y el Holoceno temprano, si bien es imposible por el momento discernir la magnitud de su contribución al ascenso regional total.
AGRADECIMIENTOS
Este trabajo fue llevado a cabo en el marco del PICTR67/02 FONCyT-ANPCyT. Los autores agradecen al Lic. E. L. Piana (CADIC-CONICET) y a autoridades y guardaparques del Parque Nacional Tierra del Fuego por el aviso de descubrimiento de los depósitos en estudio en Arroyo Baliza, en ocasión de bajamares extremas, y a la Delegación Técnica Patagonia de la Administración Nacional de Parques por el apoyo permanente a nuestros trabajos de investigación. F. I. Isla y M. Aguirre realizaron valiosas sugerencias y comentarios que contribuyeron a mejorar el trabajo. J. F. Ponce, M. Fernández (CADIC) y A.M. Borromei (UNS) colaboraron en los trabajos de campo.
TRABAJOS CITADOS EN EL TEXTO
1. Albero, M., Angiolini, F. y Piana, E. 1988. Holocene 14C reservoir effect at Beagle Channel (Tierra del Fuego, Argentine Republic). A.A. Balkema Publishers, Quaternary of South America and Antarctic Peninsula (1987) 5: 59-73, Rotterdam. [ Links ]
2. Auer, V. 1959. The Pleistocene of Fuego-Patagonia: Part III: Shorelines Displacements. Annales Academiae Scientiarum Fennicae Series A 3. Geologiska-Geographica, 247 p., Helsinki. [ Links ]
3. Auer, V. 1974. The isorythmicity subsequent to the Fuego-Patagonian and Fennoscandian ocean level transgressions and recessions of the Last Glaciation. The significance of tephrochronology, C-14 dating and micropaleontology for Quaternary research. Annales Academiae Scientiarum Fennicae, Series A 3. Geologiska-Geographica 115:1-98, Helsinki. [ Links ]
4. Batten, D.J. 1996. Palynofacies and palaeoenvironmental interpretation. En Jansonius, J. y McGregor, D.C. (eds.) Palynology: Principles and applications. American Association of Stratigraphic Palynologists Foundation 3: 1011-1064, Salt Lake City. [ Links ]
5. Borrello, A. 1969. Los Geosinclinales de la Argentina. Dirección General de Geología y Minería, Anales 14, 188 p. Buenos Aires. [ Links ]
6. Borromei, A.M. y Quattrocchio, M. 2001. Palynological study of Holocene marine sediments from Bahía Lapataia, Beagle Channel, Tierra del Fuego, Argentina. Revista Española de Micropaleontología 33: 61-70. Logroño. [ Links ]
7. Borromei, A.M. y Quattrocchio, M. 2007. Holocene sea-level change and marine palynology of the Beagle Channel, southern Tierra del Fuego, Argentina. Ameghiniana 44: 161-171, Buenos Aires. [ Links ]
8. Borromei, A., Quattrocchio, M. y Rabassa, J. 1997. Estudio palinológico de sedimentos marinos holocénicos en bahía Lapataia, Tierra del Fuego, Argentina. 6º Congreso de la Asociación Brasilera para Estudios Cuaternarios (ABEQUA), Abstracts 317-321, Curitiba. [ Links ]
9. Bujalesky, G. 2007. Coastal geomorphology and evolution of Tierra del Fuego (Southern Argentina). Geologica Acta 5(4): 337-362. [ Links ]
10. Bujalesky, G., Coronato, A., Roig, C. y Rabassa, J. 2004. Holocene differential tectonic movements along the Argentine sector of the Beagle Channel (Tierra del Fuego) inferred from marine palaeoenvironments. Geosur 2004 International Symposium on the Geology and Geophysics of the Southernmost Andes, the Scotia Arc and the Antarctic Peninsula. Bollettino de Geofisica Teorica ed applicata 45(2): 235-238. [ Links ]
11. Bujalesky, G., Coronato, A., Acevedo, R. y Rabassa, J. 2008. El Canal Beagle, un ambiente esculpido por el hielo. En Sitios de Interés Geológico de la República Argentina. Los geólogos nos cuentan, Servicio Geológico Minero Argentino, Anales 46(2): 849-864., Buenos Aires. [ Links ]
12. Caldenius, C. 1932. Las Glaciaciones Cuaternarias en Patagonia y Tierra del Fuego. Ministerio de Agricultura de la Nación. Dirección General de Minas y Geología, Boletín 95: 1-148 Buenos Aires. [ Links ]
13. Candel, M.S., Borromei, A.M., Martínez, M.A., Gordillo, S., Quattrocchio, M., y Rabassa, J., 2009. Middle-Late Holocene palynology and marine mollusks from Archipiélago Cormoranes area, Beagle Channel, southern Tierra del Fuego, Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology 273: 111-122. [ Links ]
14. Cárdenas J. y Gordillo, S. 2009. Paleoenvironmental interpretation of late Quaternary molluskan assemblages from southern South America: A taphonomic comparison between the Strait of Magellan and the Beagle Channel. Andean Geology 36 (1): 89-93. [ Links ]
15. Dale, B., 1976. Cyst formation, sedimentation, and preservation: factors affecting dinoflagellate assemblages in Recent sediments from Trondheimsfjord, Norway. Review of Palaeobotany and Palynology 22: 39-60. [ Links ]
16. Darwin, C. 1846. Geological observations on South America being the third part of the geology of the voyage of the Beagle, under the command of Capt. Fitzroy, R.N. during the years 1832 to 1836. Smith, Elder and Co 286 p. London. [ Links ]
17. Fleming, K., Johnston, P., Zwartz, D.,Yokoyama,Y., Lambeck, K. y Chappell, J. 1998. Refining the eustatic sea level curve since the Last Glacial Maximum using far- and intermediate field sites. Earth and Planetary Science Letters 163: 327-342. [ Links ]
18. Gordillo, S. 1991. Paleoecología de moluscos marinos del Holoceno Medio en Isla Gable, Canal Beagle (Tierra del Fuego, Argentina). Ameghiniana 28(1 2): 127 133, Buenos Aires. [ Links ]
19. Gordillo, S. 1992. Tafonomía y paleoecología de moluscos bivalvos del Holoceno del Canal Beagle, Tierra del Fuego. Tesis doctoral, Universidad Nacional de Córdoba, (inédita), 286 p., Córdoba. [ Links ]
20. Gordillo, S. 1993. Las terrazas marinas holocenas de la región del Beagle (Tierra del Fuego) y su fauna asociada. 12º Congreso Geológico Argentino, Actas 6: 34-39, Mendoza. [ Links ]
21. Gordillo, S. 1994. Perforaciones en bivalvos subfósiles y actuales del Canal Beagle, Tierra del Fuego. Ameghiniana 31(2): 177-185, Buenos Aires. [ Links ]
22. Gordillo, S. 1999. Holocene molluscan assemblages in the Magellan Region. Scientia Marina, 63(Supl. 1): 15-22, Barcelona. [ Links ]
23. Gordillo, S. 2007. Análisis tafonómico de quitones (Polyplacophora: Mollusca) holocenos de Tierra del Fuego, Argentina. Ameghiniana 44: 407-416. Buenos Aires [ Links ]
24. Gordillo, S., Coronato, A. y Rabassa, J. 1993. Late Quaternary evolution of a subantarctic paleofjord, Tierra del Fuego. Quaternary Science Reviews 12(10): 889-897. [ Links ]
25. Gordillo, S., Coronato, A. y Rabassa, J. 2005. Quaternary molluscan faunas from the island of Tierra del Fuego after the Last Glacial Maximum. Scientia Marina 69(Suppl. 2): 337-348. [ Links ]
26. Grill, S., Borromei, A., Quattrocchio, M., Coronato, A., Bujalesky, G. y Rabassa, J. 2002. Palynological and sedimentological analyses of Holocene sediments from Río Varela, Beagle Channel, Tierra del Fuego, Argentina. Revista Española de Micropaleontología 34(2): 145-161, Madrid. [ Links ]
27. Grimm, E. 1991. Tilia software. Illinois State Museum. Research & Collection Center. Springfield, Illinois. [ Links ]
28. Grimm, E. 2004. Tilia y TGView 2.0.2 Illinois State Museum. Research and Collection Center. Springfield, Illinois. [ Links ]
29. Halle, T. 1910. On Quaternary Deposits and Changes of Level in Patagonia and Tierra del Fuego. Bulletin of the Geological Institute of Uppsala 9: 93-117. Uppsala. [ Links ]
30. Heusser, C. 2003. Ice Age of Southern Andes. A chronicle of palaeocological events. En Rose, J. (ed) Elsevier, Developments in Quaternary Science 3, 240 p., London. [ Links ]
31. Heusser, L. y Stock, C. 1984. Preparation techniques for concentrating pollen from marine sediments and other sediments with low pollen density. Palynology 8: 225-227, Nottingham. [ Links ]
32. Isla, F., Bujalesky, G. y Coronato, A. 1999. Procesos estuarinos en el Canal Beagle, Tierra del Fuego. Revista de la Asociación Geológica Argentina 54(4): 307-318. Buenos Aires. [ Links ]
33. Kidwell, S.M., Fürsich, F.T. y Aigner, T. 1986. Conceptual framework for the analysis and classification of fossil concentrations. Palaios 1: 228-238. [ Links ]
34. McCormac, F.G., Hogg, A.G., Blackwell, P.G., Buck, C.E., Higham, T.F.G. y Reimer, P.J. 2004. SHCal04 Southern Hemisphere Calibration 0 - 11.0 cal kyr BP. Radiocarbon 46: 1087-1092. [ Links ]
35. Mörner, N. 1991. Holocene sea level changes in the Tierra del Fuego Region. Boletin IG-USP, Publicación Especial 8:133-151, São Paulo. [ Links ]
36. Navarro, J.M., Labarta, U., Fernández-Reiriz, M.J. y Velasco, A. 2003. Feeding behavior and differential absorption of biochemical components by the infaunal bivalve Mulinia edulis and the epibenthic Mytilus chilensis in response to changes in food regimes. Journal of Experimental Marine Biology and Ecology 287: 13-35. [ Links ]
37. Porter, S., Stuiver, M. y Heusser, C. 1984. Holocene Sea Level Changes along the Strait of Magellan and Beagle Channel, Southernmost South America. Quaternary Research 22: 59-67. [ Links ]
38. Rabassa, J. Heusser, C. y Stuckenrath, R. 1986. New Data on Holocene Sea Transgression in the Beagle Channel: Tierra del Fuego, Argentina. En Rabassa, J. (ed.) Balkema, Quaternary of South America and Antarctic Peninsula, 4: 291:309, Rotterdam. [ Links ]
39. Rabassa, J., Coronato, A., Bujalesky, G., Roig, C., Salemme, M., Meglioli, A., Heusser, C., Gordillo, S., Borromei, A. y Quattrocchio, M. 2000. Quaternary of Tierra del Fuego, Southernmost South America: an updated review. Quaternary International 68-71: 217-240. [ Links ]
40. Rabassa, J., Coronato, A., Roig, C., Martínez, O. y Serrat, D. 2004. Un bosque sumergido en Bahía Sloggett, Tierra del Fuego, Argentina: evidencias de comportamiento tectónico diferencial en el Holoceno tardío. Procesos geomorfológicos y evolución costera. En Blanco Chao, R., López Bedoya, J y Pérez Alberti, A. (eds.) Universidad de Santiago de Compostela, 2º Reunión de Geomorfología Litoral, Actas 333-346, Santiago de Compostela. [ Links ]
41. Rabassa, J., Coronato, A., Gordillo, S., Candel, M.S. y Martínez, M.A. 2008. El inicio de la trasgresión marina holocena en el canal Beagle, Parque Nacional Tierra del Fuego. 12º Congreso Geológico Argentino, Actas: 889890, San Salvador de Jujuy. [ Links ]
42. Stockmarr, J. 1971. Tablets with spores used in absolute pollen analysis. Pollen et Spores 13: 615-621. [ Links ]
43. Stuiver, M. y Reimer, P.J. 1993. Extended 14C data base and revised CALIB 3.0 14C age calibrating program. Radiocarbon 35: 215-230, Tucson. [ Links ]
44. Tyson, R. 1995. Sedimentary organic matter. Chapman & Hall Eds., 615 p., London. [ Links ]
45. Urien, C. 1968. Edad de algunas playas elevadas en la Península Ushuaia y su relación con el ascenso costero. 3º Jornadas Geológicas Argentinas, Actas 2: 35-42, Buenos Aires. [ Links ]
Recibido: 15 de abril, 2009
Aceptado: 29 de octubre, 2009

