










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkDarwiniana, nueva serie
versão impressa ISSN 0011-6793versão On-line ISSN 1850-1699
Darwiniana v.44 n.1 San Isidro jan./jul. 2006
Primeros aislamientos de Thraustochytriales (Labyrinthulomycetes, Heterokonta) de ambientes estuariales y salinos de la Argentina
Silvina M. Rosa1,2, Miguel A. Galvagno1,3 & Carlos G. Vélez2
1 Instituto de Investigaciones Biotecnológicas, Universidad Nacional de San Martín, Av. Colectora General Paz 5445, 1650 San Martín, Buenos Aires, Argentina; silvinarosa@bg.fcen.uba.ar (autor corresponsal).
2 Departamento de Biodiversidad y Biología Experimental, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, 1428 Buenos Aires, Argentina.
3 Departamento de Ingeniería Química, Facultad de Ingeniería, Universidad de Buenos Aires, 1428 Buenos Aires, Argentina.
Resumen. Rosa, S. M., M. A. Galvagno & C. G. Vélez. 2006. Primeros aislamientos de Thraustochytriales (Labyrinthulomycetes, Heterokonta) de ambientes estuariales y salinos de la Argentina. Darwiniana 44(1): 81-88.
Se presentan y describen cinco cepas de Thraustochytriales aisladas de dos ambientes: uno salino continental y otro estuarial. Debido a que las observaciones no coincidieron exactamente con las descripciones originales, tanto en lo morfológico como en lo ecológico, las cepas se determinaron como Thraustochytrium aff. antarcticum, Ulkenia aff. radiata y U. aff. visurgensis. Estos registros constituyen las primeras citas de Labyrinthulomycetes para Argetnina.
Palabras clave. Thraustochytriales; Thraustochytrium;Ulkenia; Taxonomía; Argentina.
Abstract. Rosa, S. M., M. A. Galvagno & C. G. Vélez. 2006. First isolations of Thraustochytriales (Labyrinthulomycetes, Heterokonta) from estuarial and saline environments of Argentina. Darwiniana 44(1): 81-88.
This is the first report of Labyrinthulomycetes for Argentina. Five strains of Thraustochytriales were isolated from both, a saline continental environment and an estuarial one from Argentina. The strains were identified as Thraustochytrium aff. antarcticum, Ulkenia aff. radiata and U. aff. visurgensis because the observations did not exactly fit the original descriptions in the morphological and ecological aspects.
Keywords. Thraustochytriales; Thraustochytrium;Ulkenia; Taxonomy; Argentina.
Original recibido el 11 de noviembre de 2005;
aceptado el 9 de marzo de 2006
INTRODUCCIÓN
Los Labirintulomicetes son microorganismos heterotróficos, de nutrición absorbente, comunes en la microflora marina y estuarial, que también pueden encontrarse en ambientes salinos continentales (Johnson & Sparrow, 1961; Amon, 1978). Considerados tradicionalmente hongos primitivos (Sparrow, 1960), han mostrado estar emparentados con las algas heterocontas (Diatomeas, Feofíceas, etc.) y los Oomicetes (Cavalier-Smith et al., 1994; Dick, 2001; Kirk et al., 2001). Se caracterizan por sus paredes celulares constituidas por escamas delgadas derivadas del apara to de Golgi y por la presencia de una red ectoplasmática producida a partir de organelas especializadasdenominadas "sagenetosomas" (Porter, 1990; Dick, 2001).
Estudios recientes de filogenia molecular sugieren que los Labirintulomicetes comprenden tres clados genéticamente distintos, correspondientes a tres grupos morfológicos: labirintúlidos, aplanoquitridios y traustoquitridios (Leander & Porter, 2000, 2001; Leander et al., 2004). Los labirintúlidos presentan células con forma de huso, que se deslizan dentro de la red ectoplasmática que las envuelve completamente (Porter, 1990). Por otra parte, los aplanoquitridios y traustoquitridios poseen esporangios globosos unidos al sustrato mediante un sistema rizoidal formado por los delicados filamentos ramificados de la red ectoplasmática que no los rodea completamente (Porter, 1990), y se diferencian entre sí por la presencia en los primeros de esporas no flageladas que se desplazan arrastrándose sobre el sustrato ("crawling spores") (Leander et al., 2004). Para una información detallada de la morfología, taxonomía, ecología, ultraestructura y métodos de cultivo de los Labirintulomicetes pueden consultarse los trabajos de Pokorny (1967), Olive (1975), Moss (1986), Fuller & Jaworski (1987), Porter (1990), Ulken (1990), Raghukumar (1996, 2002) y Dick (2001). A pesar de su ubicuidad y de su importante papel ecológico como degradadores (Raghukumar, 2002), no existen registros de estos organismos para la Argentina.
Los traustoquitridios (Thraustochytriales) han despertado recientemente un particular interés biotecnológico debido a su capacidad para producir ácidos grasos poliinsuturados (PUFAs) omega 3 con altos rendimientos, especialmente ácido docosahexaenoico (DHA) (Lewis et al., 1999). De acuerdo con los criterios taxonómicos tradicionales, el orden comprende cinco géneros que se diferencian principalmente por las características de los esporangios y el modo de producción de zoosporas (Dick, 2001; Raghukumar, 2002). En el género Thraustochytrium el protoplasto del esporangio se divide directamente en zoosporas, mientras que en Ulkenia es liberado en estado ameboidal antes de la división en zoosporas. Althornia sólo se diferencia de Thraustochytrium por la ausencia de red ectoplasmática. Schizochytrium se caracteriza por la división binaria de las células vegetativas, y Japonochytrium posee un esporangio con apófisis no presente en los otros géneros. Sin embargo, los estudios en cultivo han mostrado que la caracterización de los aislamientos no es una tarea sencilla debido a las variaciones morfológicas que se observan, incluso entre individuos de un mismo cultivo (Kazama et al., 1974; Bongiorni et al., 2005). Adicionalmente, el análisis de las secuencias de los genes de ARN ribosomal 18S muestran que no hay correlación entre los caracteres morfológicos diagnósticos y los linajes filogenéticos, sugiriendo que los criterios taxonómicos actuales deberían ser reevaluados (Honda et al., 1999).
En el marco de una investigación dirigida a estudiar la flora de traustoquitridios de la Argentina y evaluar su potencial biotecnológico, presentamos en este trabajo los primeros aislamientos de Thraustochytriales (y de Labyrinthulomycetes) para este país. Debido a lo problemático de la identificación taxonómica de estos organismos, las identificaciones se presentan con el estatus de "affinis" y se discuten las discrepancias con las características morfológicas y hábitats referidos en la literatura.
MATERIALES Y MÉTODOS
Las muestras de agua y materia orgánica en descomposición se tomaron en recipientes plásticos estériles de 50 ml, en tres ambientes de dos localidades de la provincia de Buenos Aires: 1. Punta Rasa, Bahía Escondida, marisma salina en el extremo sureste del Río de la Plata (salinidad en el momento de la colección 25); sedimentos, detritus y restos vegetales en descomposición en pozas de marea; 2. Pequeña laguna de origen pluvial aledaña al sitio anterior (salinidad no detectable); sedimentos y trozos de tallos de Schoenoplectus californicus (C. A. Mey.) Sojak var. californicus; 3. Pedro Luro, laguna salina continental La Salada, al norte del río Colorado, a unos 30 km de la costa marítima más cercana; restos orgánicos dominados por algas carofíceas. La salinidad de esta última no fue registrada, pero de acuerdo con García (1993) se trata de un cuerpo de agua hiposalino a mesosalino. Dicho autor registró una salinidad de 23.
El material se dispuso en cajas de Petri, a las que se les agregó granos de polen (Pinus sp.) como cebo (Porter, 1990) y antibióticos (penicilina 4 x 105 U · l-1 y estreptomicina 250 mg · l-1). Luego de una semana, se detectó una masiva colonización de los cebos, que fueron transferidos a medio PYGS (Fuller & Jaworski, 1987) diluido (glucosa 0,2%, peptona 0,1%, extracto de levadura 0,5%, sal de mar artificial 1,75%), solidificado con agar 1,5 % y adicionado con antibióticos en la concentración ya referida. Cuatro días después se obtuvieron colonias aisladas, que se transfirieron al mismo medio de cultivo, procedimiento que se repitió tantas veces como fue necesario hasta lograr cultivos puros. Para observar el proceso de formación de zoosporas, cuyos detalles son el principal criterio taxonómico utilizado para la identificación de estos organismos (Raghukumar, 2002), las cepas aisladas se sembraron en agua de mar artificial (salinidad 17, 5) esterilizada en autoclave, con granos de polen (Porter, 1990). En todos los casos se incubó a 25 °C. Para la clasificación se siguió el criterio de Dick (2001). Las dimensiones indicadas en las diagnosis originales se señalan entre paréntesis. Las salinidades se estimaron con un refractómetro Aquafauna (Bio-Marine Inc., USA). Las fotografías se tomaron en un microscopio Karl Zeiss provisto de óptica DIC y los dibujos se realizaron con la ayuda de un tubo de dibujo. Salvo el aislamiento de Thraustochytrium aff. antarcticum, que no pudo ser conservado en sucesivos repiques, el resto de las cepas se encuentran depositadas en la Colección de Cultivos de la Facultad de Ciencias Exactas y Naturales (BAFCcult.), Universidad de Buenos Aires.
RESULTADOS
1. THRAUSTOCHYTRIUM Sparrow emend. Johnson, in Johnson & Sparrow, Fungi in oceans and estuaries: 331. 1961.
Esporangio usualmente globoso, epibiótico, de paredes delgadas o gruesas, lisas, con o sin fundamento basal proliferativo (porción no esporógena que regenera el esporangio). Sistema rizoidal (o red ectoplasmática) endobiótico, simple o ramificado, con o sin apófisis. Zoosporas con dos flagelos de inserción lateral, móviles o inmóviles al momento de ser liberadas a través de una fisura irregular de la pared del esporangio parcialmente delicuescente. Esporas de resistencia, cuando conocidas, epibióticas, esféricas, lisas y con una única gútula de aceite, central o excéntrica.
1. Thraustochytrium aff. antarcticum Bahnweg & Sparrow, Amer. J. Bot. 61: 760. 1974. Figs. 1 A-C y 2 A-D.
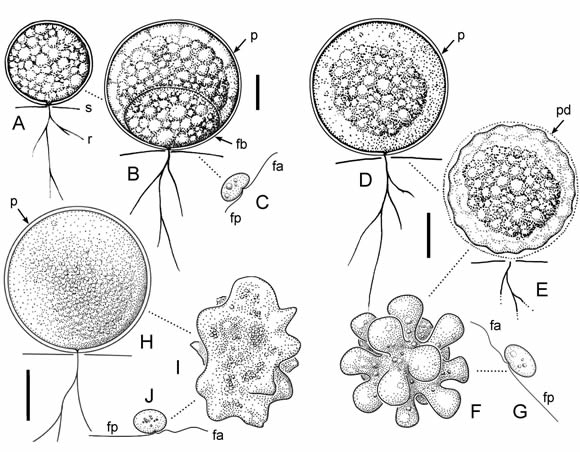
Fig. 1. A-C, Thraustochytrium aff. antarcticum. A, célula vegetativa joven. B, esporangio maduro. C, zoospora. DG, Ulkenia aff. radiata. D, esporangio maduro. E, liberación del protoplasto ameboidal por disolución de la pared celular. F, protoplasto con forma estrellada. G, zoospora. H-J, U. aff. visurgensis. H, esporangio maduro. I, protoplasto ameboidal. J, zoospora. Abreviaturas: fa, flagelo anterior; fb, fundamento basal; fp, flagelo posterior; p, pared celular; pd, pared celular en disolución; r, sistema rizoidal; s, sustrato. Las escalas indican 5 µm.
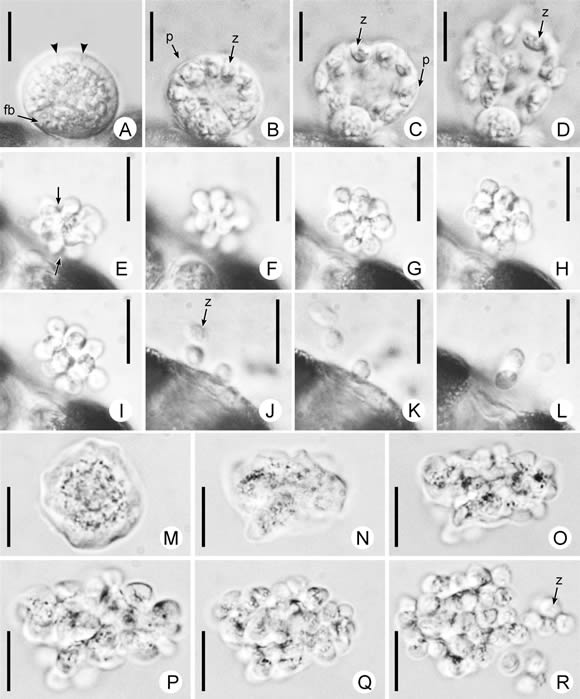
Fig. 2. A-D, Thraustochytrium aff. antarcticum. A, esporangio en maduración mostrando el fundamento basal (fb) y la periferia hialina de la zona esporógena con signos de clivaje (puntas de flecha). B-D, secuencia fotográfica (aproximadamente 5 s) de la rápida liberación de las zoosporas (z), acompañada por la disolución de la pared del esporangio (p). E-L, Ulkenia aff. radiata: zoosporogénesis (lapso aproximado:10 min). E, el protoplasto ameboidal ya liberado del esporangio, comienza a clivarse centrípetamente (flechas) originando la característica forma estrellada. F, las incisiones se hacen más profundas. G, el cuerpo de las futuras esporas ya se ha definido, permaneciendo unidas por delicadas bridas citoplasmáticas, para luego, H, ir separándose lentamente. I-L, secuencia fotográfica de la disgregación del grupo de zoosporas (z.), seguida de la adquisición de movilidad de las mismas. M-R, U. aff. visurgensis: zoosporogénesis (lapso aproximado: 10 min). M, el protoplasto acaba de emerger del esporangio y N, cambia su forma de modo irregular como una ameba. O-Q, se evidencian lóbulos, que luego de clivarse definen el cuerpo de las futuras zoosporas. R, las jóvenes zoosporas (z.) adquieren movilidad sucesivamente y se alejan nadando. Las escalas indican 10 µm.
Esporangio globoso, 19-22 (18-25) µm diám., de paredes delgadas, incoloras, con un único fundamento basal. Sistema rizoidal monoaxial, ramificado. Zoosporas elipsoidales a reniformes, 3-4 (3) µm diám. x 4-5 (5) µm long., con el flageloanterior de mayor longitud. Descarga de las zoosporas en un grupo compacto, simultánea a la desintegración completa de la pared esporangial. Luego de liberadas, las zoosporas adquieren movilidad en forma prácticamente instantánea y nadan en busca de un sustrato.
Distribución geográfica y hábitat. Océano Índico Sudoriental, en aguas superficiales oceánicas con influencia antártica (Bahnweg & Sparrow, 1974); Argentina, laguna salina continental.
Material examinado
ARGENTINA. Buenos Aires: Pdo. de Villarino: Laguna La Salada, VI-2004, Font s. n. La cepa aislada no pudo ser conservada en sucesivos repiques.
2. ULKENIA A. Gaertner, Veröff. Inst. Meeresforsch. Bremerh. 16: 141. 1977.
Talo epi- y endobiótico, interbiótico, de pared delgada durante el crecimiento activo, con un sistema rizoidal endo- o interbiótico. Talo globoso, subgloboso o piriforme, muy variable en diámetro y contenido celular durante el desarrollo. En la madurez la pared celular puede ser persistente o desaparecer parcial o totalmente, liberándose del esporangio el protoplasto desnudo, ameboidal. Luego de la liberación el comportamiento ameboidal del protoplasto se incrementa, cambiando perceptiblemente de forma. Seguidamente se divide en zoosporas, que luego de un período de inactividad se vuelven móviles en forma sucesiva debido a la formación de dos flagelos de diferente longitud.
2. Ulkenia aff. radiata A. Gaertner, Veröff. Inst. Meeresforsch. Bremerh. 16: 149. 1977. Figs. 1 D-G y 2 E-L.
Esporangios globosos, 10-20 (8-18) µm diám., de paredes delgadas. En la madurez la pared desaparece y el protoplasto ameboidal liberado adquiere forma de estrella. Seguidamente, las zoosporas se forman por división centrípeta del protoplasto en porciones aproximadamente cónicas, quedando adheridas entre sí por su extremo delgado durante un lapso variable. A continuación se forman los flagelos y el contacto en el centro se pierde. Zoosporas inicialmente cónicas, luego reniformes, 2-3 (3-4) µm diám. x 3-4 (5-8) µm long.
Distribución geográfica y hábitat. Alemania, en pozas de marea (Gaertner, 1977); Argentina, en laguna salina continental, en marisma salina y en laguna de origen pluvial aledaña a la marisma.
Material examinado
ARGENTINA. Buenos Aires: Pdo. de Villarino: Laguna La Salada, VI-2004, Font s. n. (BAFCcult. 3113); Pdo. de la Costa: Punta Rasa, IV-2004, Rosa s. n. [BAFCcult. 3109, 3110 (marisma) y 3111 (laguna aledaña)].
3. Ulkenia aff. visurgensis (A. Ulken) A. Gaertner, Veröff. Inst. Meeresforsch. Bremerh. 16: 141. 1977. Thraustochytrium visurgense A. Ulken, Veröff. Inst. Meeresforsch. Bremerh. 9: 290. 1965. Figs. 1 H-J y 2 M-R.
Esporangios globosos, 13-20 (15-20) µm diám. En la madurez la pared desaparece y el protoplasto liberado adquiere comportamiento ameboidal, de forma cambiante, emitiendo pseudopodios lobulados. Seguidamente se produce el clivaje irregular del protoplasto y se individualizan las zoosporas, las cuales yacen juntas por un tiempo, para luego alejarse nadando solas o en grupos. Zoosporas reniformes, 2-4 (2,5-3,5) µm diám. x 2,5- 4,5 (4-5,5) µm long.
Distribución geográfica y hábitat. Cosmopolita, habita en distintos ambientes marinos y estuariales (Artemchuk, 1972; Gaertner, 1974; Gaertner & Raghukumar, 1980; Ulken, 1965, 1966, 1990). En Argentina, se desarrolla en lagunas salinas continentales.
Material examinado
ARGENTINA. Buenos Aires: Pdo. de Villarino: Laguna La Salada, VI-2004, Font s. n. (BAFCcult. 3112).
DISCUSIÓN
Como referimos en la introducción, el valor taxonómico de los caracteres morfológicos de los traustoquitridios, según su uso tradicional, es dudoso. Nuestras observaciones parecen confirmar dicha presunción, como se discute a continuación. En el caso de Thraustochytrium antarticum, es llamativa la disparidad de los hábitats en los que fue encontrada, siendo la cita original para aguas superficiales antárticas (ca. 50 m de profundidad). En el caso analizado aquí, fue hallada en una laguna salada, sin conexión directa con el mar y varios kilómetros tierra adentro. Aún así, las características morfológicas de nuestros materiales se ajustan bien a la descripción original, que señala la presencia de un único fundamento basal y la delicuescencia completa de la pared esporangial en el momento de la zoosporogénesis, como rasgos diagnósticos centrales de esta especie. Sin embargo, otras características observadas tales como la zona clara periférica del citoplasma del esporangio joven (Fig. 2 A) y la extremadamente rápida liberación de las zoosporas (Fig. 2 B-D), no han sido puntualizadas por los autores de la especie y no aparecen claras en sus ilustraciones y fotografías. Si sumamos a esto la diferencia con el hábitat original, cabe preguntarse si no se trataría de un nuevo taxón. Lamentablemente la cepa aislada se perdió, por lo que deberán realizarse nuevos aislamientos para estudiar y precisar la identidad de este organismo.
El género Ulkenia parece ser el más frecuente en los ambientes estudiados. De los cuatro aislamientos de este género presentados en este trabajo, tres corresponden a U. radiata y uno a U. visurgensis. Esto es llamativo, dado que de U. radiata ha sido raramente citada (Gaertner, 1977; Huang et al., 2003), mientras que U. visurgensis parece poseer una amplia distribución, siendo la especie que se halla con más frecuencia (Dick, 2001) y con registros incluso para Brasil (Ulken, 1966). Ambas comparten algunos rasgos diagnósticos como la disolución total de la pared esporangial cuando se libera el protoplasto ameboidal, y la ausencia tanto de cuerpos proliferativos como de fisión binaria, lo que las distingue del resto de las especies del género (Gaertner, 1977). La diferencia morfológica más conspicua entre ambas es el patrón de clivaje del estado ameboidal, irregular en U. visurgensis y marcadamente radial en U. radiata. Aunque nosotros observamos estas características en nuestros materiales, y nos basamos en ellas para establecer las afinidades propuestas, también encontramos algunas discordancias con las descripciones originales. En el caso de U. aff. radiata, las zoosporas resultaron ser más pequeñas y redondeadas, y no exhibían el cuerpo protoplasmático denso posterior descripto por Gaertner (1977). U. aff. visurgensis, por su parte, presentó paredes esporangiales más delgadas que lo indicado en la diagnosis de la especie.
De acuerdo con lo expuesto hasta aquí y tal como se señalara en la introducción, hacer coincidir las observaciones con las diagnosis de los taxones no es una tarea sencilla, tanto desde el punto de vista morfológico como ecológico. Por esta razón, coincidimos con lo señalado por la mayoría de los autores que actualmente trabajan en el grupo: resulta imprescindible la realización de otro tipo de estudios (de filogenia molecular, de ultraestructura, bioquímicos, etc.) que complementen la información actualmente disponible para la identificación de estos organismos (Honda et al., 1999; Leander et al., 2004; Huang et al., 2003) y que permitan definir criterios taxonómicos más seguros.
AGRADECIMIENTOS
Queremos agradecer a Fabián Font por el legado de muestras, al personal de los Laboratorios de Micología de la Facultad de Ciencias Exactas y Naturales de la Universidad de Buenos Aires por el generoso apoyo prestado durante la realización de este trabajo, y a los árbitros por sus constructivas sugerencias.
BIBLIOGRAFÍA
1. Amon, J. P. 1978. Thraustochytrids and labyrinthulids of terrestrial, aquatic and hypersaline environments of the Great Salt Lake, USA. Mycologia 70: 1299-1301. [ Links ]
2. Artemchuk, N. J. 1972. The fungi of White Sea. III. New Phycomycetes discovered in the Great Salma Strait of the Kanadalakshial Bay. Veröff. Inst. Meeresf. Bremerhaven 13: 231-237. [ Links ]
3. Bahnweg, G. & F. K. Sparrow. 1974. Four new species of Thraustochytrium from Antartic regions, with notes on the distribution of zoosporic fungi in the Antartic marine ecosystems. Am. J. Bot. 61: 754-766. [ Links ]
4. Bongiorni, L., R. Jain, S. Raghukumar & R. Aggarwal, 2005. Thraustochytrium gaertnerium sp. nov.: a new Thraustochytrid Stramenopilan Protist from Mangroves of Goa, India. Protistología 156: 303-315. [ Links ]
5. Cavalier-Smith, T., M. T. E. P. Allsopp & E. E. Chao. 1994. Thraustochytrids are chromist, not Fungi: 18s rRNA signatures of Heterokonta. Philos. Trans., Ser. B 346: 387-397. [ Links ]
6. Dick, M. W. 2001. Straminipilous Fungi. Kluwer Academic Publishers, Dordrecht, Holanda. [ Links ]
7. Fuller, M. S. & A. Jaworski (eds.). 1987. Zoosporic fungi in teaching and research. Southeastern Publishing Corporation, Athens, Georgia. [ Links ]
8. Gaertner, A. 1974. Vorkommen und Verteilung mariner niederer Pilze in der Nordsee und im Nordmeerbecken. Veröff. Inst. Meeresforsch. Bremerh. Supp. 5: 123-147. [ Links ]
9. Gaertner, A. 1977. Revision of the Thraustochytriaceae (Lower Marine Fungi) I. Ulkenia nov. gen., with description of three new species. Vroff. Inst. Meeresf. Bremerhaven Suppl. 16: 139-157. [ Links ]
10. Gaertner, A. & S. Raghukumar. 1980. Ecology of thraustochytrids (lower marine fungi) in the Fladen Ground and other parts of the North Sea. Meteor Forschungsergebn., D 22: 165-185. [ Links ]
11. García, A. 1993. Quaternary and recent Lamprothamniun groves (Charophyta) from Argentina. Hydrobiologia 267: 143-154. [ Links ]
12. Honda, D., T. Yocochi, T. Nakahara, S. Raghukumar, A. Nakagiri, K. Schauman & T. Higashihara. 1999. Molecular phylogeny of labyrinthulids and thraustochytrids based on the sequencing of 18S ribosomal RNA gene. J. Eukar. Microbiol. 46: 637-647. [ Links ]
13. Huang, J., T. Aki, T. Yocochi, T. Nakahara, D. Honda, S. Kawamoto, S. Shigeta, K. Ono & O. Suzuki. 2003. Grouping newly isolated docosahexaenoic acid-producing thraustochytrids based on their polyunsaturated fatty acid profiles and comparative analysis of 18S rRNA genes. Mar. Biotechnol. Abstr. 5: 450-457. [ Links ]
14. Johnson, T. W. & F. K.Sparrow. 1961. Fungi in Oceans and Estuaries. Cramer, Weinheim, Alemania. [ Links ]
15. Kazama, F., A. Zachary & Schornstein, K. 1974. Observations on Thraustochytrium sp.: development and behaviour in culture. Canad. J. Bot. 53: 360-374. [ Links ]
16. Kirk, P. M., P. F. Cannon, J. C. David & J. A. Stalpers (eds). 2001. Ainsworth and Bisbys Dictionary of the Fungi. 9na ed. CABI Publishing, Wallingford, UK. [ Links ]
17. Leander, C. & D. Porter. 2000. Redefining the genus Aplanochytrium (phylum Labyrinthulomycota). Mycotaxon 76: 439-444. [ Links ]
18. Leander, C. 2001. The Labyrinthulomycota is comprised of three distinct linages. Mycologia 93: 459-464. [ Links ]
19. Leander, C., D. Porter & B. Leander. 2004. Comparative morphology and molecular phylogeny of aplanochytrids (Labyrinthulomycota). Eur. J. Protistol. 40: 317-328. [ Links ]
20. Lewis, T.E., P. D. Nicholson & T. A. McMeekin. 1999. The biological potential of thrausthochytrids. Mar. Biotechnol. Abstr. 1: 580-587. [ Links ]
21. Moss, S. T. 1986. Biology and phylogeny of the Labyrinthulales and Thraustochytriales, pp 105-129, en S.T. Moss (ed.), The Biology of Marine Fungi. Cambridge University Press, Cambridge. [ Links ]
22. Olive, L. S. 1975. The Mycetozoan. Academic Press, New York. [ Links ]
23. Pokorny, K. S. 1967. Labyrinthula. J. Protozool. 14: 697:708. [ Links ]
24. Porter, D. 1990. Phylum Labyrinthulomycota, pp. 338-398, en L. Margulis, J.O. Corliss, M. Melkonian & J. Chapman (eds.), Handbook of Protoctista. Jones and Barlett Publishers, Boston. [ Links ]
25. Raghukumar, S. 1996. Morphology, taxonomy and ecology of thraustochytrids and labyrinthulids, the marine counterparts of zoosporic fungi, pp 35-60, en R. Dayal, (ed.), Advances in zoosporic fungi. M.D. Publications Pvt. Ltd., New Delhi. [ Links ]
26. Raghukumar, S. 2002. Ecology of the marine protists, the Labyrinthulomycetes (Thraustochytrids and Labyrinthulids). Eur. J. Protistol. 38: 127-145. [ Links ]
27. Sparrow, F.K. 1960. Aquatic Phycomycetes. 2da. ed. Ann Arbor. Univ. Michigan Press, Michigan. [ Links ]
28. Ulken, A. 1965. Zwein neue Thraustochytrien aus der AuBen-weser. Veröff. Inst. Meeresf. Bremerhaven. 9: 289-295. [ Links ]
29. Ulken, A. 1966. Untersuchugen über marine Pilze im äquatorialen Atlantik vor der Küste Brasiliens. Veröff. Inst. Meeresf. Bremerhaven 10: 107-116. [ Links ]
30. Ulken, A. 1990. Marine thraustochytrids and chytridiomycetes in the North Sea area and in selected other regions. J. Cramer, Berlin. [ Links ]

