










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkDarwiniana, nueva serie
Print version ISSN 0011-6793On-line version ISSN 1850-1699
Darwiniana vol.44 no.2 San Isidro Aug./Dec. 2006
Estudios en las Apocynaceae Neotropicales XXVIII: la familia Apocynaceae (Apocynoideae, Rauvolfioideae) de El Salvador, Centroamérica
J. Francisco Morales
Instituto Nacional de Biodiversidad (INBio), Apto. 23-3100, Santo Domingo de Heredia, Costa Rica;
E-mail: fmorales@inbio.ac.cr
Resumen. Morales, J. F. 2006. Estudios en las Apocynaceae neotropicales XXVIII: la familia Apocynaceae (Rauvolfioideae, Apocynoideae) de El Salvador, Centroamérica. Darwiniana 44(2): 453-489.
Se presenta un tratamiento florístico de la familia Apocynaceae, subfamilias Rauvolfioideae y Apocynoideae, para la República de El Salvador, Centroamérica. Como resultado, se informan 17 géneros nativos (Allotoonia, Alstonia, Aspidosperma, Cascabela, Echites, Fernaldia, Forsteronia, Mandevilla, Mesechites, Pentalinon, Plumeria, Prestonia, Rauvolfia, Stemmadenia, Tabernaemontana,Thevetia, Vallesia) y cinco introducidos (Allamanda, Beaumontia, Carissa, Catharanthus,Nerium). Se trata un total de 34 especies, siendo Mandevilla y Prestonia los géneros con mayor número de especies (4), ninguna de ellas endémica. Se incluyen descripciones de la familia, los géneros y las especies, además de claves y especímenes representativos examinados. Asimismo se incluyen referencias bibliográficas, nombres comunes, usos (cuando disponibles) y fotografías.
Palabras clave. Apocynaceae; Apocynoideae; El Salvador; Flora; Rauvolfioideae; Sistemática.
Abstract. Morales, J. F. 2006. Studies on neotropical Apocynaceae XXVIII: the family Apocynaceae (Rauvolfioideae,
Apocynoideae) from El Salvador, Central America. Darwiniana 44(2): 453-489.
The family Apocynaceae s. str. (Apocynaceae s. l. subfamilies Apocynoideae and Rauvolfioideae) is reviewed for the Republic of El Salvador, Central America. Seventeen native genera (Allotoonia,Alstonia, Aspidosperma, Cascabela, Echites, Fernaldia, Forsteronia, Mandevilla, Mesechites, Pentalinon,Plumeria, Prestonia, Rauvolfia, Stemmadenia, Tabernaemontana, Thevetia, Vallesia) are accounted for, as well as five introduced genera (Allamanda, Beaumontia, Carissa, Catharanthus,Nerium). The most species-rich genera are Mandevilla and Prestonia, with four species each. A total of 34 species are reported. Family, genus, and species descriptions are provided, together with keys and representative specimen citations. Bibliographic references, common names, uses (when known), and photographs are also included.
Keywords. Apocynaceae; Apocynoideae; El Salvador; Floristics; Rauvolfioideae; Systematics.
Original recibido el 11 de agosto de 2006;
aceptado el 9 de octubre de 2006
INTRODUCCIÓN
La familia Apocynaceae, tomando en cuenta la inclusión de las Asclepiadaceae como lo han sugerido estudios moleculares y filogenéticos recientes (e.g., Endress & Albert, 1995; Sennblad & Bremer, 1996; Potgieter, 1999), es una de las más grandes de las Angiospermas, con un total de alrededor de 355 géneros y 3700 especies, distribuidas principalmente en las regiones tropicales (Endress & Bruyns, 2000). Bajo este concepto de unificación, cinco subfamilias fueron propuestas por Endress y Bruyns (2000), quedando los géneros tradicionalmente tratados como Apocynaceae a las subfamilias Apocynoideae y Rauvolfioideae. En este artículo, este grupo es denominado "Apocynaceae s. str."
A pesar de su tamaño geográfico relativamente pequeño, El Salvador carece de un tratamiento integral de su flora que permita la identificación de sus diferentes especies. La familia Apocynaceae s. str. no es la excepción, y el único estudio disponible que trató en parte las especies de El Salvador (aunque sin un detalle de localidades o especímenes examinados) es el de la Flora de América del Norte (Woodson, 1938a). Este trabajo comprende un total de 13 géneros y 18 especies [Allamanda cathartica L, Fernaldia pandurata (A. DC.) Woodson, Forsteronia spicata (Jacq.) G. Mey., Lochnera rosea (L.) Rchb. ex Endl., Mandevilla subsagittata (Ruiz & Pav.) Woodson, M. villosa (Miers) Woodson, Nerium oleander L., Plumeria rubra L., Prestonia portobellensis (Beurl.) Woodson, P. speciosa Donn. Sm., Rauvolfia hirsuta Jacq., Stemmadenia glabra Benth., S. obovata Benth., Tabernaemontana amygdalifolia Jacq., Thevetia peruviana (Pers.) K. Schum., T. plumeriifolia Benth., Tonduzia longifolia (A. DC.) Woodson)]. Continuando con el proceso de tratamientos de la familia Apocynaceae s. str. en el Nuevo Mundo, se presenta una revisión para la República de El Salvador, Centroamérica, siguiendo el mismo esquema propuesto por Morales (2005c) para Costa Rica.
MATERIALES Y MÉTODOS
La República de El Salvador, con una extensión aproximada de 21000 kilómetros cuadrados, se encuentra ubicada en el N de América Central, entre los 13 y 15 grados latitud Norte y entre los 87 y 90 grados longitud Oeste. Limita al NO con la República de Guatemala, al N y E con la República de Honduras, al SE con la República de Nicaragua (dividido por el Golfo de Fonseca) y al S y O con el océano Pacífico (Fig. 1). El territorio se encuentra dividido en 14 departamentos, donde las elevaciones oscilan de 0 a 2385 m s.m. (volcán Santa Ana). El Salvador es un país predominantemente volcánico, ya que se conocen alrededor de 25 volcanes hoy extintos, por lo que el terreno es mayormente montañoso, aunque existe una meseta central y áreas relativamente planas distribuidas en una estrecha franja costera.
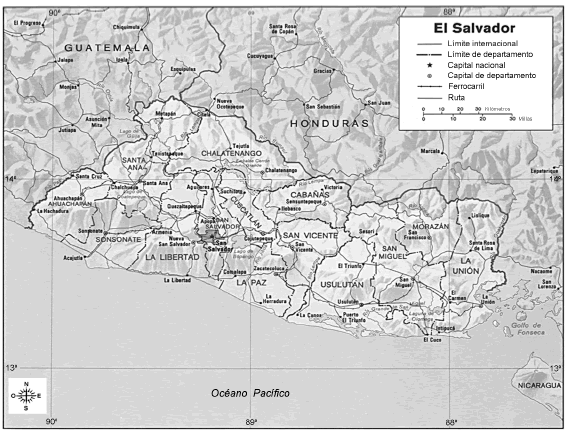
Fig. 1. Mapa de la República de El Salvador (cortesía de la Librería de la Universidad de Texas, Austin, Estados Unidos, http://www.lib.utexas.edu/maps/el_salvador.html)
El clima es tropical con dos estaciones: una lluviosa, que se extiende de mayo a noviembre, y una seca, de diciembre a abril, con una temperatura promedio que ronda los 24 °C. Las precipitaciones promedio alcanzan los 1830 mm anuales, aunque este valor varía según los diferentes ecosistemas presentes en el país. La vegetación original de El Salvador es hoy escasa, debido principalmente a la alta densidad poblacional que ha provocado la desaparición de la mayoría de los bosques, y su reemplazo por cultivos agrícolas o sistemas de producción ganadera.
Dada la ausencia total de un tratamiento o de un listado reciente de las Apocynaceae s. str. para El Salvador, se visitaron los herbarios de mayor importancia en Estados Unidos de América (DUKE, F, MO, NY, US), Europa (BM, BR, CGE, G, K, MA, P, U, W, WAG, WU) y Mesoamérica (BIGU, CR, EAP, INB, LAGU, TEFH, UVAL) en los últimos 9 años (1997-2006), con el objeto de elaborar un listado preliminar de las especies y géneros presentes en El Salvador. En forma paralela, se elaboró una base de datos para catalogar todos los especímenes de Apocynaceae examinados, de manera que todos sus datos de localidad y estado fenológico pudieran ser documentados, así como los usos y nombres comunes registrados en las etiquetas de herbario. En forma general, se examinaron más de 1000 especímenes de herbario, incluyendo la mayoría de los tipos nomenclaturales. Las siglas de los herbarios citados siguen a Holmgren et al. (1990).
Las descripciones de los géneros y especies fueron tomadas de diversas monografías o revisiones previas, que son citadas para cada género, incluyéndose a su vez, bibliografía de importancia particular en algún punto específico (por ejemplo tipificación). Morales (2005c) proveyó una descripción detallada de los géneros y especies de las Apocynaceae de Costa Rica, así como fotografías para la mayoría de ellos. Por lo tanto, dichas descripciones e ilustraciones no serán repetidas en este tratamiento y se brindará únicamente la referencia bibliográfica específica cuando corresponda. Para cada especie, se brinda la sinonimia más relevante, sobre todo de taxones descriptos originalmente para El Salvador o países circundantes. Las descripciones de las especies se complementaron con el análisis de las flores y los frutos coleccionados en Costa Rica y Colombia, y preservados en alcohol (una mezcla de 80% de alcohol etílico al 99% de pureza, con un adicional de 20% de glicerol), así como el estudio de especímenes secos depositados en el Herbario INB. Para cada uno de los géneros, se trató de incluir al menos una fotografía representativa, tomadas en su mayoría por el autor en Honduras, Costa Rica, Colombia, Perú y Bolivia en los últimos 14 años. En algunos casos se utilizaron fotografías con el permiso de otras personas e instituciones, especialmente citadas en los agradecimientos. Por otro lado, algunas descripciones fueron tomadas de diversas monografías en desarrollo (e.g., Mandevilla, Prestonia), por lo que los datos de distribución o morfología pueden diferir de otros anteriormente publicados (e.g., Woodson, 1933, 1936). Los datos de distribución altitudinal, fenología, nombres comunes y usos fueron tomados directamente de las etiquetas de herbario, por lo que la veracidad de estos últimos no pudo ser comprobada. Nombres comunes adicionales de otros países de la misma región, pueden ser consultados en Morales (2005c). Únicamente se cita una colección representativa para cada Departamento, pero se brinda una lista completa de los especímenes estudiados al final de esta revisión.
TERMINOLOGÍA Y CARACTERES TAXONÓMICOS SINGULARES
En el tratamiento de las Apocynaceae de Costa Rica, Morales (2005c) proveyó una explicación detallada de algunas estructuras morfológicas características de las Apocynaceae, muchas de ellas acompañadas de fotografías ilustrativas. Por lo tanto, solo se repetirá aquella información que es de relevancia para la adecuada interpretación de las claves y los límites genéricos. Más información e ilustraciones adicionales sobre los diversos tipos de coléteres, pueden ser encontradas en Morales (2005c), así como en los trabajos de Ezcurra (1981) y de Simões & Kinoshita (2002). Se discuten los caracteres de los géneros del Nuevo Mundo de las subfamilias Apocynoideae y Rauvolfioideae únicamente, según Endress & Bruyns (2000).
Algunas estructuras adicionales en las hojas, que merecen una atención importante, son los domacios que algunas veces están presentes en la superficie abaxial de la lámina foliar, a lo largo del nervio central (Fig. 2 B). En las Apocynaceae s. str. presentes en El Salvador dichas estructuras pueden ser observadas con facilidad en Forsteronia. Otros géneros presentes en países adyacentes y que tienen domacios son Tintinnabularia (Guatemala y Honduras) y Malouetia (Guatemala, Honduras y Nicaragua).
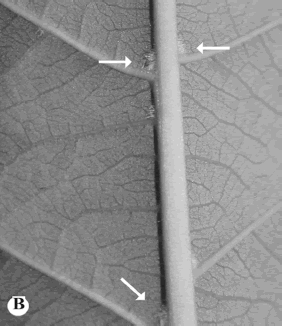
Fig. 2. Coléteres y domacios.B, domacios (Forsteronia spicata).
En relación a las corolas y sus partes, se sigue a Ezcurra (1981), Simões & Kinoshita (2002) y Morales & Fuentes (2004). La estivación de los lóbulos de la corola es un carácter clave para poder distinguir los diferentes miembros de las subfamilias Apocynoideae y Rauvolfioideae, ya que en la primera predominan las corolas con estivación dextrorsa (Fig. 3A), mientras que en las Rauvolfioideae predomina la estivación sinitrorsa (Fig. 3B). Aunque si bien, las estructuras coronales presentes en Nerium y Prestonia fueron explicadas por Morales (2005c), se hace de nuevo ilustrarlas con el objeto de facilitar el uso de las claves en este tratamiento. La corona anular (Fig. 3C) se encuentra ubicada alrededor de la fauce del tubo, mientras que los lóbulos coronales (Fig. 3D), se encuentran en forma interna y opuesta a cada antera, por esta razón es que pueden ser confundidos comúnmente con ellas. La primera estructura está limitada a Prestonia en El Salvador, mientras que la segunda se puede encontrar aparte de ese género en Nerium.
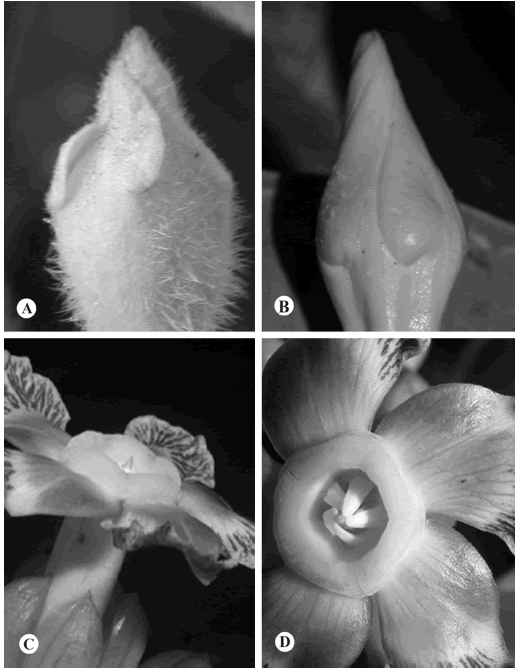
Fig. 3. Estructuras morfológicas de corolas. A, estivación dextrorsa (Mandevilla hirsuta). B, estivación sinitrorsa (Cascabela ovata). C, corona anular (Prestonia trifida). D, lóbulos coronales libres (Prestonia portobellensis).
Al menos en los géneros presentes en El Salvador, es común que el ovario esté rodeado por un nectario, que puede ser anular o estar limitado a dos o tres nectarios individuales (Fig. 4A, B, C). Sin embargo, el nectario está ausente en unos pocos géneros, en donde el ovario se halla en contacto directo con el tubo de la corola (Fig. 4D).
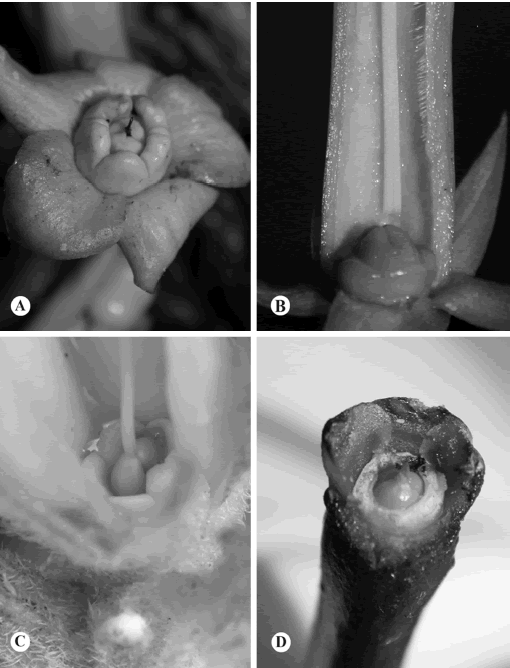
Fig. 4. Nectarios y ovario en Apocynaceae. A, Cascabela ovata. B, Cascabela thevetia. C, Mandevilla hirsuta. D,Plumeria rubra (nectario ausente).
TRATAMIENTO TAXONÓMICO
APOCYNACEAE Juss. Gen. Pl. 143. 1789.
Familia con ca. 355 géneros y 3700 especies, mayormente en los trópicos (Endress & Bruyns, 2000). En El Salvador 22 géneros y 34 especies. Véase la descripción de la familia en Morales (2005c).
Funtumia elastica (Preuss) Stapf (Apocynoideae, Malouetieae) es una liana que crece en el Jardín Botánico La Laguna (La Libertad) y aparentemente se encuentra colonizando zonas aledañas como una especie invasora; se puede reconocer por sus hojas opuestas, láminas eglandulares, pecíolos con hasta 3 hileras de coléteres en la superficie adaxial, inflorescencias paniculadas, termina les y usualmente con numerosas flores, así como folículos apocárpicos, de más de 20 cm long. y semillas con coma en ambos extremos. Esta especie no se incluye en el tratamiento, dado que solo se conoce en cultivo de un área muy reducida y no existen evidencias de que se cultive en otras áreas.
Clave para la identificación de los géneros que crecen en El Salvador
1. Hojas alternas o subalternas, algunas veces agrupadas al final de las ramas o nudos pareciendo subopuestas, pero nunca consistentemente opuestas . . . . . . . . . . . . . . . . . . .2
1. Hojas estrictamente opuestas o verticiladas, nunca alternas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6
2(1). Sépalos con numerosos coléteres en la base de la cara adaxial; corola infundibuliforme o hipocraterimorfa; frutos drupáceos o abayados, rojos, morados o morado-rojizos al madurar . . . . . . . . . . . . . . 3
2. Sépalos sin coléteres en la base de la cara adaxial; corola estrictamente hipocraterimorfa; frutos foliculares o si drupáceos (Vallesia), entonces blancos o blanco-verdosos al madurar . . . . . . . . . . . . 4
3(2). Corola hipocraterimorfa, crema a verde-crema; frutos abayados, rojos al madurar. . . . . . . . . . . . . . 21. Thevetia
3. Corola infundibuliforme, amarillo intenso; frutos drupáceos, morados a morado-rojizos al madurar . . . . . . 7. Cascabela
4(2). Frutos drupáceos, carnosos al madurar; semillas sin alas marginales; hojas por lo común dísticas . . . . . . . . . . . . . 22. Vallesia
4. Frutos foliculares, secos, leñosos o firmemente membranáceos al madurar; semillas usualmente con alas marginales; hojas nunca dísticas . . . . . . . . . . . . . 5
5(4). Lóbulos de la corola usualmente de más de 2 cm long.; folículos falcados y cilíndricos, usualmente de más de 1,5 cm de diámetro . . . . . . . . . . . . . . . . . . 16. Plumeria
5. Lóbulos de la corola de menos de 1,5 cm long.; folículos cilíndricos y continuos, menores de 1 cm de diámetro, o leñosos y aplanados de forma lateral . . . . . . 4. Aspidosperma
6(1). Hojas mayormente verticiladas, raramente algún nudo con hojas opuestas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7
6. Hojas mayormente opuestas, rara vez algún nudo con hojas verticiladas. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
7(6). Tubo de la corola con cinco lóbulos coronales petaloides, opuestos a cada antera; semillas con coma apical; plantas estrictamente cultivadas, raramente subespontánea cerca de las ciudades . . . . . . . . . . . . . . . . . . . . . 14. Nerium
7. Tubo de la corola sin lóbulos coronales; semillas aladas, ciliadas marginalmente o desnudas, sin coma apical; plantas silvestres o cultivadas . . . . . 8
8(7). Corola infundibuliforme, amarilla; frutos capsulares, espinosos en la superficie exterior; semillas con alas marginales; plantas cultivadas . . . . . . . . . . .1. Allamanda
8. Corola hipocraterimorfa a urceolada, blanca o crema; frutos drupáceos o foliculares, pero entonces lisos en la superficie exterior, sin espinas; plantas silvestres . . . . . . . . . 9
9(8). Folículos; semillas con los márgenes ciliados; pecíolos sin coléteres en la superficie adaxial; lóbulos de la corola de 3-6 mm long.; hojas siempre isofilas . . . . . . . . . . .3. Alstonia
9. Drupas; semillas no ciliadas; pecíolos con coléteres en la superficie adaxial; lóbulos de la corola ca. 1 mm long.; hojas isofilas o anisofilas . . . . . . . . . 8. Rauvolfia
10(6). Corola mayor de 10 cm long.; filamentos largos y conspicuos, evidentes, de varios cm de longitud; plantas siempre cultivadas . . . . . . . . . 5. Beaumontia
10. Corola menor de 8 cm long.; filamentos inconspicuos, menores de 1 cm de longitud; plantas silvestres o cultivadas . . . . . . . . . . . 11
11(10). Tallos con espinas bifurcadas en los nudos; plantas conocidas solo en cultivo . . . . . . . . . . . . . . . . . 6. Carissa
11. Tallos siempre sin espinas; plantas silvestres o cultivadas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12.
12(11). Árboles o arbustos erectos de varios metros de altura, usualmente con un tronco principal o varios tallos múltiples definidos, nunca escandentes. . . . . . . . . . . .13
12. Lianas, arbustos escandentes o hierbas erectas, algunas veces arbustivas (Catharanthus), pero entonces inferiores a 1,5 m de altura y nunca con un tallo leñoso principal y erecto de varios metros de longitud. . . . . . . . . . . . . . . 14
13(12). Tubo de la corola con cinco proyecciones o crestas bajo las anteras; lóbulos de la corola amarillo intenso . . . . . 19. Stemmadenia
13. Tubo de la corola sin alas bajo las anteras; lóbulos de la corola blancos. . . . . . . . . . . . . . . . 20. Tabernaemontana
14(12). Láminas foliares con coléteres diminutos en la superficie abaxial del nervio central, distribuidos a lo largo de su entera extensión o agrupados en la base. . . . . . . . . . . . . . .15
14. Láminas foliares sin coléteres en el nervio central . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 17
15(14). Anteras exsertas, con al menos 1/3 de su longitud exserto; tubo de la corola inferior a 3,5 mm long.; láminas foliares algunas veces con domacios . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11. Forsteronia
15. Anteras siempre inclusas; tubo de la corola superior a (4-)10 mm de longitud; láminas foliares usualmente sin domacios . . . . . 16
16(15). Inflorescencias racemosas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12. Mandevilla
16. Inflorescencias cimosas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13. Mesechites
17 (14). Lóbulos de la corola con estivación sinistrorsa; anteras libres de la cabeza estigmática; semillas sin comas presentes; sépalos de forma usual sin coléteres en la base de la cara adaxial, plantas cultivadas y ocasionalmente subespontáneas . . . . . . . . 8. Catharanthus
17. Lóbulos de la corola con estivación dextrorsa o valvar; anteras aglutinadas a la cabeza estigmática; semillas de forma usual con algún tipo de coma presente en uno solo de los extremos; sépalos en forma usual con coléteres en la base de la cara adaxial, de forma más rara sin coléteres . . . . . . . . . . . . . . . . .18
18(17). Corola con una corona anular alrededor de la fauce, de forma usual con cinco lóbulos coronales libres en las superficie interna (corona epiestaminal), algunas veces estos ausentes y reducidos a callos o crestas callosas longitudinales . . . . . . . . . . . .17. Prestonia
18. Corola sin una corona anular alrededor de la fauce; lóbulos coronales ausentes . . . . . . . . . . . . . . . . . . . . . . . . . 19
19(18). Lóbulos de la corola muy angostamente elípticos a casi filiformes, acuminados a largamente acuminados y filiformes en el ápice, de manera usual enroscados en espiral distalmente. . . . . . . . . . . . . . . . . . . . . . 2. Allotoonia
19. Lóbulos de la corola obovados, redondeados a obtusos en el ápice, no enroscados en espiral distalmente. . . . . 20
20(19). Lóbulos de la corola densa a moderadamente vilosos en la superficie adxial, los pelos blancos, conspicuos y largos, de manera usual de varios mm de longitud; inflorescencias estrictamente racemosas . . . . . . 10. Fernaldia
20. Lóbulos de la corola glabros, glabrescentes a diminutamente puberulentos en la superficie adaxial, los pelos diminutos e inconspicuos, ca. 1 mm long. o menos, no blancos; inflorescencias de varios tipos, a veces racemosas, pero los lóbulos sin pelos largos y conspicuos . . . . . . . . . . .21
21(20). Corola hipocraterimorfa; anteras con apéndices apicales inconspicuos, triangulares, pero nunca filiformes o de varios mm de longitud . . . . . . . .9. Echites
21. Corola infundibuliforme; anteras con apéndices apicales filiformes, usualmente de varios mm de longitud y entrelazados entre sí . . . . . . . . . . . 15. Pentalinon
Descripción de los géneros y las especies
1. ALLAMANDA L., Mant. Pl. 2: 146, 214 (576). 1771.
Género con 14 especies restringidas principalmente a Brasil. Dos de ellas se conocen solo en cultivo en el El Salvador: Allamanda blanchetii A. DC. (sinónimo: A. violacea Gardner & Fielding) y A. cathartica L. Una tercer especie, A. schottii Pohl (conocida en Centroamérica) podría encontrarse en cultivo en El Salvador, aunque no se pudo encontrar un ejemplar de herbario que constate esta posibilidad. Allamanda blanchetii se ha cultivado en forma muy esporádica en los Departamentos de La Libertad y San Salvador; y se puede reconocer de A. cathartica por sus corolas lilacinas o moradas (Sakane & Sheperd, 1987). A. schottii se reconoce de A. cathartica por sus flores, sépalos y frutos más pequeños. Para la descripción del género y mayores detalles véase Morales (2005 c).
1. Allamanda cathartica L., Mant. Pl. 2: 214- 215. 1771. TIPO. " habitat per totam Guianam, juxta fluvios ", sin fecha (fl), Herb. Linn. 298.1 (neotipo S-LINN! designado por Leeuwenberg, Regnum. Veg. 127: 16. 1993).
Referencia. Morales (2005c: 101).
Distribución y hábitat. Originaria de la cuenca baja amazónica, pero cultivada en todo el mundo. Cultivada ocasionalmente en El Salvador, sobre todo en elevaciones de 500-1400 m. Las flores y los frutos se producen durante todo el año.
Nombres vulgares. "San José" (Ahuachapán).
Observaciones. En El Salvador se conoce solo en cultivo, pero esta especie crece en forma natural en otras zonas de la región del Caribe de Mesoamérica.
Material representativo examinado
EL SALVADOR. Ahuachapán. Ahuachapán, 22-V- 1994, R. López s.n. (LAGU, MO). Chalatenango. La Palma, El Gramal, M. Smeets 63 (ITIC, LAGU, U).
2. ALLOTOONIA J. F. Morales & J. K. Williams, Sida 21(1): 135-137. 2004.
Cinco especies distribuidas desde México hasta Panamá y las Antillas, dos registradas en El Salvador. (Morales & Williams, 2004, 2005). Para la descripción del género y mayores detalles véase Morales (2005c: 105).
1. Lóbulos de la corola más pequeños que el tubo; ápice del botón floral agudo o cortamente acuminado; tubo 8-10 mm long. . . . . . . . . . . . . . . . . . 3. A. tuxtlensis
1. Lóbulos de la corola más largos que el tubo; ápice del botón floral obtuso a redondeado; tubo 4-7 mm long.
2.. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A. caudata
2. Allotoonia caudata (Woodson) J. F. Morales, Sida 21(1): 145. 2004. Prestonia caudata Woodson, Ann. Missouri Bot. Gard. 47: 79. 1960, non E. caudata Blanco, 1837. Echites puntarenensis J. F. Morales, Brittonia 49: 332. 1997. TIPO: Costa Rica. Puntarenas: cercanías de Cascajal, 25 km ESE de Puntarenas, 3-VII- 1949 (fl), R. Holm & H. Iltis 243 (holotipo, MO!; isotipos, A!, CR!, G!, GH!, P!).
Referencia. Morales (2005c: 105).
Distribución y hábitat. Guatemala, El Salvador, Nicaragua y Costa Rica. Bosque secos o bosques estacionalmente secos, en elevaciones de 0-500 m. Floración de septiembre a enero.
Observaciones. En forma general, los lóbulos de este taxón son muchos más largos que el tubo y extendidos, mientras que en Allotoonia tuxtlensis, la otra especie presente en El Salvador, son cortos y de manera usual enrollados.
Material representativo examinado
EL SALVADOR. Santa Ana. Coatepeque, 3-IX- 1994, R. Villacorta & M. Méndez 2181 (LAGU, MO).
3. Allotoonia tuxtlensis (Standl.) J.F. Morales & J.K. Williams, Sida 21(1): 153. 2004. Echites tuxtlensis Standl., Contr. U.S. Natl. Herb. 23: 1164. 1924. TIPO: México. Chiapas: cerca de Tuxtla, 1-IX-1895 (fl), E. Nelson 3080 (holotipo, US!). Fig. 5A.
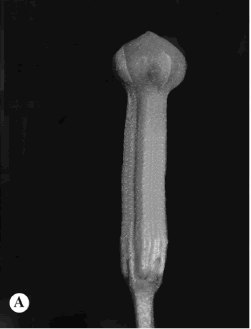
Fig. 5. Allotoonia, Alstonia y Aspidosperma. A, Allotoonia tuxtlensis.
Lianas. Hojas de 4,5-11 x 1,4-3,7(-4,8) cm, angostamente elípticas a angostamente ovadas, acuminadas apicalmente, obtusas a cuneadas en la base, pecíolo de 3-11 mm long. Inflorescencias con numerosas flores, pedúnculo de 2,6-8 cm long., pedicelos de 0,8-1,9 cm long., brácteas de 0,8-1,5 mm long., escariosas; sépalos de 1,5-1,8 mm long., ovadas, acuminados; corola amarillocrema, verde-crema a verde, tubo de 8-10 mm long., lóbulos de 6-8 mm long., glabros; anteras de 4 mm long., nectarios ligeramente más cortos que el ovario. Folículos de 13-15 cm x 1-3 mm, continuos, glabros; semillas de 11-13 mm long., coma de (2,3)2,5-3,2 cm long.
Distribución y hábitat. S de México (Chiapas, Campeche, Oaxaca, Quintana Roo y Yucatán), Guatemala, Belice, Honduras y El Salvador. Bosques secos en elevaciones de 150-700 m. Especímenes con flores han sido coleccionados entre abril y junio, septiembre y en diciembre. Los frutos se producen entre febrero y mayo. Nombres vulgares. "Bejuco lengua de culebra" (Ahuachapán, San Benito); "Loroco de Culebra" (El Salvador, Cuscatlán).
Material representativo examinado
EL SALVADOR. Ahuachapán. San Francisco Menéndez, El Corozo, 12-V-2000, J. Rosales 702 (LAGU, MO). Cuscatlán. cerca de Cerrón Grande, 11- XII-1997, M. Renderos 378 (B, BM, BONN, INB, LAGU, MO). Santa Ana. Metapán, 28-IV-1995, J. Linares & C. Martínez 2728 (EAP, MO).
3. ALSTONIA R. Br., Mem. Wern. Nat. Hist. Soc. 1: 75. 1811.
Género con 45 especies distribuidas en el trópico, principalmente en el Paleotrópico, dos conocidas de México a Costa Rica, con solo una en El Salvador. (Morales, 1995, 2005c; Sidiyasa, 1998). Para la descripción del género y mayores detalles véase Morales (2005c: 108).
4. Alstonia pittieri (Donn. Sm.) A. Gentry, Missouri Bot. Gard. 70: 206. 1983. Tonduzia pittieri Donn. Sm., Bot. Gaz. 49(6): 456. 1910. TIPO: Guatemala. Fiscal, 1-VI-1909 (fl), C. Deam 6098 (lectotipo, US! designado por Sidiyasa, Blumea, Suppl. 11: 145, 1998; isolectotipo, F!, foto F neg. 56525 INB!). Fig. 5 B, C.


Fig. 5. Allotoonia, Alstonia y Aspidosperma.B-C, Alstonia pittieri.
Rauvolfia stenophylla Donn. Sm., Bot. Gaz. 44: 115. 1907. Tonduzia stenophylla (Donn. Sm.) Pittier, Contr. U.S. Natl. Herb. 12(2): 104, t. 9. 1908. Alstonia stenophylla (Donn. Sm) J. F. Morales, Darwiniana 43: 108. 2005, nom. inval., non A. stenophylla Guillaumin, 1941. TIPO: El Salvador. San Salvador. cercanías de San Salvador, 1905 (fl), C. Renson 289 (holotipo, US! [2 cartulinas]; isotipo, F!)
Tonduzia parvifolia Pittier, Contr. U.S. Natl. Herb. 12(2): 103, f. 5-6. 1908, non A. parvifolia Merr., 1905. TIPO: Costa Rica. Cartago:Angostura, cerca de Turrialba, 11-IV-1903 (fl), C. Doyle 33 (holotipo, US!; isotipo, MO!).
Tonduzia longipedunculata Woodson, Publ. Field Mus. Nat. Hist., Bot. Ser. 23(2): 78. 1944. TIPO: Guatemala. Sololá: sendero entre San Pedro, vía San Juan, San Cristobal, Buena Vista y falda NO del Volcán Santa Clara, 8-VI-1942 (fl), J. Steyermark 47313 (holotipo, MO!; isotipo, F!, foto F neg. 56526 INB!).
Referencia. Morales (2005c: 108, bajo A. stenophylla).
Distribución y hábitat. México a Costa Rica. Bosques húmedos y bosques estacionalmente secos, en elevaciones de 200-1100 m s.m. Florece en enero y de junio a septiembre. Los frutos se producen entre julio y enero.
Nombres vulgares. "Chilindrón" (Ahuachapán); "Chilindrón de montana" (La Libertad); "Sulfatillo" (Ahuachapán).
Observaciones. Alstonia pittieri se puede distinguir con facilidad por la siguiente combinación de caracteres: hábito arbóreo, con las hojas dispuestas en verticilos, flores blancas con los sépalos sin coléteres en la base de la cara adaxial; así como por sus frutos foliculares, con las semillas sin una coma en su extremo micropilar, aplanadas y con una hilera de cilios en los márgenes.
Material representativo examinado
EL SALVADOR. Ahuachapán. San Benito, Cerro León, V-1993, E. Sandoval 1254 (LAGU, MO). La Libertad. Teotepeque, 22-IV-1995, J. González 315 (LAGU, MO). San Miguel. Volcán Conchagua, 30-I- 1998, A. Monro et al. 2111 (BM, MO). San Vicente, volcán de San Vicente, III-1922 (fl), S. Calderón 339 (MO). Santa Ana. Santa Ana, 6-V-1959, E. Montalvo 3385 (MO). Sonsonate. Cerro Verde, 21-IX-1958, P. Allen & M. van Severen 6924 (EAP).
4. ASPIDOSPERMA Mart. & Zucc., Flora 7(1) (beil. 4): 135. 1824, nom. cons.
Género con ca. 45 especies distribuidas desde México y las Antillas al N de la Argentina (excepto Chile); solamente 1 reportada en el El Salvador. (Marcondes-Ferreira, 1988). Para la descripción del género y mayores detalles véase Morales (2005 c: 111).
5. Aspidosperma megalocarpon Müll. Arg., Linnaea 30: 400. 1860. Macaglia megalocarpa (Müll. Arg.) Kuntze, Revis. Gen. Pl. 2: 416. 1891. TIPO: México. Colipa, 1841-1842 (fr), W. Karwinsky 1348 (holotipo, LE!). Fig. 5 D.

Fig. 5. Allotoonia, Alstonia y Aspidosperma.D, Aspidosperma megalocarpon.
Cufodontia lundelliana Woodson, Arch. Bot. Sist. 10(1): 40. 1934. TIPO: Guatemala. Petén: La Libertad, 26- V-1933 (fl), C. Lundell 3408 (holotipo, MO!; isotipos, F!, foto F neg. 56459 INB!, MICH!, US!).
Aspidosperma lundellianum Woodson, Amer. J. Bot. 22(7): 684. 1935. México. Campeche: Tuxpeña, 4-II- 1932 (fl, fr), C. Lundell 1284 (holotipo, MO!; isotipos, F!, foto F neg. 56433 INB!, GH!).
Cufodontia arborea Woodson, Ann. Missouri Bot. Gard. 21(4): 617. 1934. TIPO: México. Oaxaca: Cafetal, Concordia, 1-15-IV-1933 (fl), C. Morton & E. Makrinius 2692 (holotipo, US!; isotipos, CAS!, F!, foto F neg. 56458 INB!).
Cufodontia escuintlensis Matuda, Madroño 10(6): 174. 1950. TIPO: México. Chiapas: Calcuta, Acacovagua, 17-VIII-1947 (fl), E. Matuda 16978 (holotipo, MEXU!; isotipos, CAS! [2 cartulinas], F!, foto F neg 51100 INB!).
Referencia. Morales (2005c: 112).
Distribución y hábitat. México a Panamá y en el N de Colombia. Crece en bosques húmedos y formaciones asociadas a afloramientos de piedras calizas, en elevaciones de 200-650 m s.m. Las flores se producen entre enero y mayo. Los frutos son registrados entre diciembre y marzo y en agosto.
Nombres vulgares. "Molleja de pato" (Ahuachapán).
Observaciones. Aspidosperma megalocarpon se puede reconocer con facilidad por su hábito arbóreo, hojas alternas, inflorescencias caulifloras, las corolas con el tubo inferior a 6 mm long. y folículos lateralmente aplanados, leñosos y con las semillas aladas
Material representativo examinado
EL SALVADOR. Ahuachapán. San Francisco Menéndez, San Benito, 20-II-1992, M. Sandoval & Chinchilla 272 (LAGU, MO). Cabañas. Cinquera, quebrada la Creciente, 22-IV-2001, R. Carballo et al. 301 (INB, LAGU).
5. BEAUMONTIA Wall., Tent. Fl. Napal. 14. 1824.
Género con 9 especies en Asia tropical, India e Indonesia (Bali); en El Salvador 1 especie cultivada. (Rudjiman, 1986). Para la descripción del género y mayores detalles véase Morales (2005c: 114).
6. Beaumontia grandiflora Wall., Tent. Fl. Napal. 1: 15, pl. 7. 1824. TIPO: Nepal. Noakote, Nawakot, Wallich 1629,1 (holotipo, KWALL no visto)
Referencia. Morales (2005c: 115).
Distribución y hábitat. Nativa del Este de la India. Cultivada ocasionalmente en parques y jardines, en elevaciones de 700-1200 m s.m.
Nombres vulgares. "Campana blanca" (La Libertad).
Observaciones. Es una de las especies de Apocynaceae más fáciles de distinguir, principalmente por el gran tamaño de sus flores. Esta especie es cultivada en casi toda América tropical, pero en forma esporádica. Casi no se conocen frutos, por lo que su descripción fue tomada de Rudjiman (1986).
Material representativo examinado
EL SALVADOR. La Libertad. Jardín Botánico La Laguna, 17-III-1984, W. Berendsohn & B. Berendsohn 69 (LAGU, MO).
6. CARISSA L., Syst. Nat. (ed. 12) 2: 135, 189. 1767.
Arbustos o árboles pequeños, tallos aplanados cuando jóvenes, cilíndricos a subcilíndricos con la edad, con secreción lechosa, glabros, glabrescentes o inconspicuamente puberulentos, coléteres intrapeciolares diminutos, pero evidentes, usualmente con presencia de espinas, algunas veces bifurcadas. Hojas opuestas, sin coléteres en el nervio central, la lámina glabra (en especies comúnmente cultivadas), coriáceas, corto-pecioladas. Inflorescencias cimas axilares o reducidas a 1 ó 2 flores fasciculadas, usualmente con pocas flores, brácteas escariosas, deciduas o persistentes; cáliz con 5 sépalos, iguales o subiguales, escariosos a subfoliáceos, usualmente glabros (en especies comúnmente cultivadas), usualmente sin coléteres en la base de la cara adaxial, de manera inusual unos pocos coléteres presentes; corola hipocraterimorfa, sin corona anular o lóbulos coronales libres, blanca o crema, limbo con 5 lóbulos, con estivación sinistrorsa o dextrorsa; estambres totalmente inclusos, anteras no conniventes ni aglutinadas a la cabeza estigmática, cabeza estigmática anchamente ovada, apenas diferenciada del estilo, sin collar basal; gineceo sincárpico, con pocos o relativamente numerosos óvulos, nectario usualmente ausente.
Frutos sincárpicos, bayas indehiscentes, globosos o subglobosos, lisos, carnosos; semillas desnudas, usualmente pocas, raramente hasta 15 por fruto. Género paleotropical con ca. 20 especies; una sola especie ocasionalmente cultivada en El Salvador. (Gentry, 2001; Leeuwenberg & Van Dilst, 2001).
7. Carissa macrocarpa (Eckl.) A. DC., Prodr. 8: 336. 1844. Arduina macrocarpa Eckl., S. African Quart. J. 1: 372. 1830. TIPO: no localizado. Fig. 6A.

Fig. 6. Carissa y Cascabela. A, Carissa macrocarpa.
Arduina grandiflora E. Mey., Comm. Pl. Afr. Austr. 190. &1835. Carissa grandiflora (E. Mey.) A. DC., Prodr. 8: 335. 1844. TIPO: Sur África: "capem zwischen Omtendo und Amsamculo", II-(fl), J. Drege 88 (holotipo, S!; isotipo, MO!).
Arbustos de 0,5-1,5(-2) m de altura, tallos glabros o glabrescentes, con escasas espinas furcadas en los nudos. Hojas de (1,7-)3-7,5 x (1,5-)2-4,5 cm, elípticas, angostamente elípticas a angostamente ovadas, el ápice apiculado, mucronado o mucronulado, la base obtusa o muy levemente cordada, glabras, pecíolo de 1-4 mm long. Inflorescencia sésiles o subsésiles, pedicelos de 2,5-4 mm long., brácteas de c. 1 mm long.; sépalos de 3-5 x 2-4 mm, angostamente ovados a angostamente ovado-elípticos, el ápice acuminado con coléteres; corola blanca, tubo de 0,9-1,7 cm long., con un diámetro de 2- 2,5 mm en la fauce, lóbulos de 17-22 x 4-7 mm, obovados a angostamente obovados, extendidos; anteras de 3-4 mm long., ovario de 1,5-2 mm long., glabro. Frutos de 1,8-2,5(-3) x 1,2-3 cm, rojos al madurar; semillas de 6-8 mm long.
Distribución y hábitat. Originaria del S de África, pero cultivada ocasionalmente en El Salvador, en elevaciones de (400-)700-1000 m s.m. Las flores y los frutos se producen de manera intermitente durante todo el año.
Observaciones. Carissa macrocarpa es la única especie con espinas bifurcadas presente en el neotrópico. Se cultiva principalmente por su porte ornamental, aunque sus frutos son comestibles.
Material representativo examinado
EL SALVADOR. La Libertad. La Libertad, 31-X- 1989, R. Cruz 222 (LAGU, MO). San Salvador. San Salvador, 19-III-1992, P. Quintana s.n. (LAGU, MO).
7. CASCABELA Raf., Sylva Tellur. 162. 1838.
Arbustos o árboles, tallos subcilíndricos, con secreción lechosa, glabros a puberulentos, los pecíolos con coléteres axilares pequeños e inconspicuos. Hojas alternas, usualmente dispuestas en espiral, lámina eglandular, sin coléteres, glabras, glabrescentes o puberulentas en la superficie abaxial, firmemente membranáceas a subcoriáceas, sin domacios, márgenes no revolutos, sésiles, subsésiles a pecioladas. Inflorescencias cimosas, axilares a subterminales, con pocas o muchas flores, glabrescentes o puberulentas, brácteas escariosas; cáliz con 5 sépalos, iguales, escariosos a subfoliáceos, con numerosos coléteres en la base; corola infundibuliforme, amarilla, sin corona anular, de manera usual glabra, el interior con cinco lóbulos coronales en forma opuesta sobre cada antera, limbo con 5 lóbulos, con estivación sinistrorsa; estambres inclusos, anteras no conniventes ni aglutinadas a la cabeza estigmática, cabeza estigmática con lóbulos basales; gineceo bicarpelar, apocárpico, con 2-4 óvulos, nectario anular y a veces lobulado. Frutos sincárpicos, drupas, morados o rojo morados al madurar, el exocarpo carnoso o esponjoso, el mesocarpo leñoso; semillas 2-4, desnudas, no comosas.
Género con cerca de 5 especies distribuidas desde México hasta Suramérica, dos presentes en el Salvador.
1. Láminas foliares de 2-6 cm de ancho; venación secundaria prominente y siempre conspicua en el envés . . . . . . . . . . . . . . . .8. C. ovata
1. Láminas foliares de 0,4-1,1 cm de ancho; venación secundaria apenas impresa y usualmente inconspicua por el envés . . . . . . . . . . . . . . . . . . .9. C. peruviana
8. Cascabela ovata (Cav.) Lippold, Feddes Repert. 91(1-2): 53. 1980. Cerbera ovata Cav., Icon. 3: 35, t. 270. 1796. Thevetia ovata (Cav.) A. DC., Prodr. 8: 344. 1844. TIPO: "Nueva España", sin fecha (fl), Cavanilles s.n. (holotipo, MA!). Fig. 3 B, 4 A, 6 B.

Fig. 6. Carissa y Cascabela.B, Cascabela ovata.
Thevetia plumeriifolia Benth, Bot. Voy. Sulphur. 124, t. 43. 1845. Cascabela plumeriifolia (Benth.) Lippold, Feddes Repert. 91(1-2): 53. 1980. TIPO: Honduras. Golfo de Fonseca, sin fecha (fl), Sinclair s.n. (lectotipo K! designado por Nelson, Fontqueria 44: 60. 1996).
Referencia. Morales (2005c: 177).
Distribución y hábitat. México a Costa Rica. Crece en bosques secos, acantilados costeros, en elevaciones de 0-800 m s.m. Las flores y los frutos se producen intermitentemente durante todo el año.
Nombres vulgares. "Cojón de costa hojas largas" (Ahuachapán); "Chilindrón" (La Libertad). Observaciones. Esta especie puede llegar a formar poblaciones bastante densas en zonas cercanas a la costa, donde se puede reconocer con facilidad por sus hojas con la venación secundaria conspicuamente impresa en el envés, flores amarillas y frutos sincárpicos y drupáceos. La variación de la pubescencia de la superficie abaxial de las casos encontrarse individuos casi glabros.
Material representativo examinado
EL SALVADOR. Ahuachapán. El Refugio, San Benito, 15-II-1997, J. González 507 (LAGU, MO). Cabañas. Cinquera, 2-II-2002, R. Carballo et al. 189 (LAGU, MO). Chalatenango. Nueva Concepción, Pañanalapa, 17-V-2000, K. King & J. Chávez 39 (INB, LAGU, MO). La Libertad. Deininger, IV-1976, D. Current 44 (MO). La Unión. El Carmén, 1928, S. Calderón 2375 (MO). Morazán. Perquín, río Sapo, 25-I- 2000, J. Monterrosa et al. 4. (LAGU, MO). San Miguel. volcán Conchagua, 31-I-1998, A. Monro et al. 2157 (BM, MO). San Vicente. Cercanías de San Vicente, 7-14-II-1947, P. Standley & E. Padilla 3440 (EAP). Santa Ana. Santa Ana, 23-IV-1995, L. Linares & C. Martínez 2549 (MO).
9. Cascabela thevetia (L.) Lippold, Feddes Repert. 91 (1-2): 52. 1980. Cerbera thevetia L., Sp. Pl. 1: 209. 1753. Cerbera peruviana Pers., Syn. Pl. 1: 267. 1805, basado en C. thevetia. Cascabela peruviana (Pers.) Raf., Sylva Tellur. 162. 1838. Thevetia peruviana (Pers.) K. Schum., Nat. Pflanzenfam. 4(2): 159. 1895. Thevetia thevetia (L.) Millsp., Field Mus. Nat. Hist., Bot. Ser. 2(1): 83. 1900, nom. inval. Thevetia peruviana (Pers.) Merr., Philipp. J. Sci. 9(2): 130. 1914, nom. illeg. TIPO: " habitat in America calidiore, Cerbera foliis linearibus, longissimus" in Plumier, Codex Boerhaavianus (lectotipo, designado por Lippold, Feddes Repert. 91: 52. 1980). Figs. 4 B, 6 C-D.

Fig. 6. Carissa y Cascabela.C-D, Cascabela thevetia.
Referencia. Morales (2005c: 177, bajo Thevetia).
Distribución y hábitat. Originaria del NO de Sudamérica, se conoce en El Salvador únicamente cultivada, en elevaciones de 400-1100 m s.m. La floración y la fructificación ocurre durante todo el año.
Observaciones. Cascabela peruviana se ha usado con regularidad como planta ornamental y en El Salvador se encuentra plantada principalmente en parques. Aunque en forma general las semillas y el látex de esta planta son tóxicos, se ha informado el consumo ocasional de los frutos maduros y casi secos, en distintas áreas de Mesoamérica.
Material representativo examinado
EL SALVADOR. San Salvador. San Salvador, 18- IX-1987, J. Flores s.n. (LAGU, MO). Santa Ana. Santa Ana, 2-X-2001, V. Martínez 39 (INB, LAGU, MO).
8. CATHARANTHUS G. Don, Gen. Hist. 4: 71, 95. 1837.
Género con 8 especies, en su mayoría endémicas de Madagascar, una de ellas cultivada en zonas tropicales y templadas de todo el mundo (Bergen, 1996). Para la descripción del género y mayores detalles véase Morales (2005c: 115).
10. Catharanthus roseus (L.) G. Don., Gen. Hist. 4: 95. 1837. Vinca rosea L., Syst. Nat. (ed. 10) 2: 944. 1759. Lochnera rosea (L.) Rchb. ex Endl., Gen. Pl. 583. 1838. TIPO: Madagascar, Java, sin fecha (fl), Miller, Fig. Pl. Gard. Dict., 2: t. 186, 1757 (lectotipo designado por Codd, Fl. S. Africa 26: 268. 1963).
Referencia. Morales (2005c: 115). Distribución y hábitat. Originaria de Madagascar, pero cultivada en todo el mundo, en elevaciones entre 0-1500 m s.m. Las flores y los frutos se producen durante todo el año.
Nombres vulgares. "Loroco de zope" (Ahuachapán); "Chula" (La Libertad).
Material representativo examinado
EL SALVADOR. Ahuachapán. San Alfonso, parque nacional El Imposible, 17-VI-1990, A. Sermeño 205 (LAGU, MO). La Libertad. hacienda Cuyagualo, al S del Lago de Zapotitlán, río Gualo, 27-I-1946, M. Carlson 314 (EAP). San Salvador. San Salvador, 10-VIII- 1987, J. Flores s.n. (LAGU, MO).
9. ECHITES P. Browne, Civ. Nat. Hist. Jamaica: 182. 1756.
Lianas, tallos algo aplanados cuando jóvenes, cilíndricos a subcilíndricos con la edad, con secreción lechosa, glabros a inconspicuamente puberulentos, coléteres interpeciolares inconspicuos. Hojas opuestas, pecioladas, sin coléteres en el nervio central, glabras o puberulentas, membranáceas, sin domacios, los márgenes de manera usual no revolutos. Inflorescencias cimosas, axilares y con numerosas flores, algunas veces reducidas a 1 ó 2 flores, glabras a inconspicuamente puberulentas, brácteas escariosas e inconspicuas; cáliz con 5 sépalos, iguales o subiguales, escariosos, glabros o puberulentos, con 1 coléter en la base de la cara adaxial, a veces profundamente lacerado y dando la impresión de muchos coléteres individuales; corola hipocraterimorfa, sin corona anular o lóbulos coronales, blanca, blanco-crema a crema, limbo con 5 lóbulos, con estivación dextrorsa; estambres inclusos, anteras conniventes y aglutinadas a la cabeza estigmática, filamentos inconspicuos, cabeza estigmática fusiforme, con un anillo membranoso basal; gineceo bicarpelar, apocárpico, con numerosos óvulos, nectario anular irregularmente pentalobulado o separado en cinco nectarios individuales. Frutos apocárpicos, compuestos por dos folículos, continuos, levemente leñosos; semillas numerosas, truncadas o levemente rostradas, desnudas y secas, comosas en el ápice micropilar.
&Género con 3 especies distribuidas desde el S de Estados Unidos de América (Florida) y las Antillas hasta Panamá; una sola especie presente en el Salvador (Morales, 1997 b).
11. Echites yucatanensis Millsp. ex Standl., Publ. Field Columbian Mus., Bot. Ser. 8(1): 35. 1930. TIPO: México.Yucatán: Chichankanab, fecha perdida (fl), G. Gaumer 1979 (holotipo, F!; isotipos, C!, MO!, NY!, S!, TEX!).
Echites turriger Woodson, Ann. Missouri Bot. Gard. 19(4): 381. 1932. TIPO: Guatemala. Zacapa: Gualan, 20-VI-1909 (fl), C. Deam 6376 (holotipo, MO!; isotipos, F!, foto F neg. 51101 INB!, GH!, NY!, US!).
Echites circinnalis Woodson, Ann. Missouri Bot. Gard. 21: 616. 1934. TIPO: Nicaragua. Granada: cercanías de Granada, 1896 (fl), P. Lévy 1074 (holotipo, C!, foto INB!). Echites elegantulus Woodson, Amer. J. Bot. 22(7): 686. 1935. TIPO: México. Yucatán: Chichén Itzá, 23-VI- 1932 (fl), W. Steere 1477 (holotipo, MO!; isotipos, F!, MO!, NY!).
Liana, tallos cilíndricos, glabros a esparcidamente puberulentos. Hojas con pecíolo de 0,9-3,7 cm long., láminas de 4-16 x 2-10,5 cm, ovadas, elípticas, angostamente elípticas o panduradas, el ápice acuminado, la base obtusa o redondeada, membranáceas, glabras o glabrescentes, algunas veces con pubescencia muy esparcida en el envés, nervaduras impresas y conspicuas en ambas caras. Inflorescencia con (3-)6-25 flores, algo aglomerada, pedúnculo de 1,6-17 cm long., pedicelos de 0,8-3 cm long., brácteas de 1,5-4,5 mm long.; sépalos de 2-5 mm long., ovados, el ápice largoacuminado; corola blanca o crema, glabra o puberulenta en la superficie externa, tubo de 2,5-5,3 cm long., recto, sin torciones, con un diámetro de 3-4 mm en la fauce, lóbulos de 1,7-3,5 x 9-2,3 cm, obovados; anteras de 7-8 mm long., ovario de ca. 2 mm long., glabro, disco anular, entero o irregularmente pentalobulado. Folículos de 18-29 cm x 5-8 mm, glabros y lisos, continuos; semillas de 1,7-2 cm long., coma 2,5-3,5 cm long.
Distribución y hábitat. México a Nicaragua. Crece en bosques secos y estacionalmente secos en elevaciones de 20-700 m s.m. Florece entre abril y julio. Los frutos se han coleccionado en julio. Nombres vulgares. "Flor blanca" (La Libertad); "Loroco de Venado" (Ahuachapán); "Loroco Montes" (San Salvador)".
Observaciones. En forma general, las especies de Echites actualmente reconocidas; y de géneros relacionados (Fernaldia), son morfológicamente muy variables. Aunque se ha propuesto reconocer E. yucatanensis y E. turriger como dos especies distintas (Williams, 2002), varios de las caracteres utilizados para distinguirlas se superponen. En todo caso, la misma variabilidad está presente en E. umbellatus, por lo que no tiene sentido reconocer una serie de taxones basados en caracteres no excluyentes, considerando la alta variabilidad del género.
Material representativo examinado
EL SALVADOR. Ahuachapán. Parque Nacional El Imposible, Los Najarro, 1997, E. Sandoval 1647 (INB, LAGU). La Libertad. Zaragoza, Guadalupe, 10-VI- 2001, M. Hernández PBT-561 (INB, LAGU). San Salvador. Aguijares, sitio Arqueológico Sihuatán, 9-VII- 1997, M. Renderos 186 (INB, LAGU). Santa Ana. Entre El Pedregal San Isidro y Hacienda La Presa, 1- VII-1995, C. Linares & C. Martínez 2845 (EAP). Sonsonate. Entre El Pedregal San Isidro y la Chacarita, 1- VII-1995, C. Linares & C. Martínez 2866 (EAP).
10. FERNALDIA Woodson, Ann. Misouri Bot. Gard. 19(1): 48. 1932.
Género con 3 especies distribuidas desde México al NE de Panamá, con una sola presente en El Salvador (Morales, 2002). Para la descripción del género y mayores detalles véase Morales (2005c: 119).
12. Fernaldia pandurata (A. DC.) Woodson, Ann. Missouri Bot. Gard. 19: 48. 1932. Echites panduratus A. DC., Prodr. 8: 458. 1844. Amblyanthera pandurata (A. DC.) Müll. Arg., Linnaea 30: 448. 1860. Angadenia pandurata (A. DC.) Miers, Apocyn. S. Am.: 182. 1878. TIPO: México. Oaxaca: San Dionicio, VIII-1832 (fl), G. Andrieux 245 (lectotipo, G-DC! designado por Morales, Rhodora 104: 192. 2002, foto F neg. 7559, F!, INB!, MO!, NY!, US!; isolectotipo, K!). Fig. 7 A.

Fig. 7. Fernaldia y Forsteronia. A, Fernaldia pandurata.
Fernaldia brachypharynx Woodson, Ann. Missouri Bot. Gard. 19: 380. 1932. TIPO: Guatemala: carretera entre Escuintla y el puerto de San José de Guatemala, 23-VIII-1860 (fl), S. Hayes s.n. (holotipo, GH!). Fernaldia pandurata var. glabra A. Molina, Ceiba 3: 95. 1952. Fernaldia glabra (Molina) Lundell, Wrightia 5: 256. 1976. TIPO: Honduras. Cortés: faldas de la Montaña Santa Ana, río Santa Ana, 6-XII- 1950 (fl), A. Molina 3640 (holotipo, EAP!; isotipo, F!, foto F neg. 56469 INB!).
Referencia. Morales (2005c: 119).
Distribución y hábitat. México a Costa Rica. Crece en bosque secos o estacionalmente secos, en elevaciones de 0-850 m s.m. La floración y la fructificación ocurren principalmente de mayo a diciembre.
Nombres vulgares. "Bejuco quilite" (Ahuachapán); "Loroco" (Ahuachapán)" Observaciones. Para una información más detallada sobre los usos culinarios de las flores, véase el trabajo de Morton et al. (1990).
Material representativo examinado
EL SALVADOR. Ahuachapán. parque nacional El Imposible, San Alfonso, 10-VII-1990, A. Sermeño 232 (LAGU, MO). San Salvador. San Salvador, L. Químico 2 (MO). Santa Ana. Coatepeque, 3-IX-1994, R. Villacorta & M. Méndez 2196 (LAGU, MO).
11. FORSTERONIA G. Mey., Prim. Fl. Esseq. 133. 1818.
Género con ca. 46 especies distribuidas desde México hasta la Argentina y las Antillas; dos taxones conocidos en El Salvador (Hansen, 1985). Para la descripción del género y mayores detalles véase Morales (2005c: 121).
1. Inflorescencia de apariencia espiciforme, ocasionalmente con 1 a 3 ramificaciones cortas en la base; lámina foliar moderada- a esparcidamente puberulenta en el envés, con mechones de pelos en las axilas de los nervios; sépalos de 2,5-4,5 mm long.; semillas de 8,5-10 mm de long. . . . . . . . . . . . . . . . . . .14. F. spicata
1. Inflorescencia subcorimbosa a tirseiforme, con varias ramificaciones, pero no pareciendo una espiga; lámina foliar glabra, glabrescente o esparcida e inconspicuamente puberulenta en el envés, sin mechones de pelos; sépalos de ca. 1 mm long.; semillas de 12-16 mm long. . . . . . . . . . 13. F. myriantha
13. Forsteronia myriantha Donn. Sm., Bot. Gaz. 27 (6): 435. 1899. TIPO: Guatemala. Zacatepequez: Embaulada, XII-1889, E. Heyde & E. Lux 4534 (lectotipo US! designado por Hansen, 1985, US! [3 cartulinas]; isolectotipos, BM!, G!, GH!, K!, M!, US!). Figs. 7 B, C.


Fig. 7. Fernaldia y Forsteronia.B-C, Forsteronia myriantha.
Referencia. Morales (2005c: 123).
Distribución y hábitat. México a Panamá, Colombia, Ecuador, Perú y Bolivia. Crece en bosques húmedos y bosques estacionalmente secos, en elevaciones de 100-500 m s.m. la floración ocurre entre marzo y agosto.
Nombres vulgares. "Bejuco de Tacuazín" (Ahuachapán).
Observaciones. Forsteronia myriantha se puede reconocer de F. spicata por sus hojas glabras, glabrescentes o inconspicuamente puberulentas abaxialmente, inflorescencias mucho más abiertas, nunca de aspecto espiciforme, flores con sépalos más pequeños y folículos mucho más largos y desarrollados. Adicionalmente, esta especie crece en bosques más húmedos y no se encuentra en bosques secos.
Material representativo examinado
EL SALVADOR. Ahuachapán. San Francisco Menéndez, El Corozo, 31-III-2000, J. Rosales 455 (LAGU, MO).
14. Forsteronia spicata (Jacq.) G. Mey., Prim. Fl. Esseq. 135. 1818. Echites spicatus Jacq., Enum. Syst. Pl. 13. 1760. TIPO: Colombia. Lectotipo, designado por Hansen (1985), Jacquin, Sel. Stirp. Amer. Hist. 2: pl. 29. 1763. Fig. 2 B, 7 D.

Fig. 7. Fernaldia y Forsteronia.D, Forsteronia spicata.
Referencia. Morales (2005 c: 124). Distribución y hábitat. México a Colombia y Cuba. Crece en bosques secos, en elevaciones de 0-550 m s.m. Las flores se han observado de mayo a septiembre; los frutos se producen entre agosto y octubre.
Nombres vulgares. "Bejuco Tacuazín blanco" (Ahuachapán).
Observaciones. Esta especie aparentemente no es común en El Salvador, y se encuentra restringida principalmente en zonas de bosques secos.
Material representativo examinado
EL SALVADOR. Ahuachapán. San Francisco Menéndez, El Corozo, 7-VIII-2000, J. Rosales 1261 (LAGU, MO).
12. MANDEVILLA Lindl., Edwards´s Bot. Reg. 26: t. 7. 1840, nom. cons.
Género con ca. 145 especies distribuidas desde México y las Antillas hasta la Argentina (excepto Chile); 4 presentes en El Salvador. (Morales, 1998 a). Mandevilla hirsuta (Rich) K. Schum. (S de México a Brasil, Paraguay y Bolivia) no ha sido coleccionada en el El Salvador, pero es de esperarse. Puede reconocerse del resto de las especies por sus hojas con los coléteres agrupados en la base del nervio central, así como sus inflorescencias con brácteas foliáceas y corolas infundibuliformes con el tubo mayor de 33 mm long. Para la descripción del género y mayores detalles véase Morales (2005c: 131).
1. Coléteres distribuidos a lo largo del nervio central; tubo de la corola basalmente giboso . . . . . . . . . . . .2
1. Coléteres agrupados en la base del nervio central; tubo de la corola recto, no giboso . . . . . . . . . . . . . .3
2(1). Brácteas 2-5 mm long., escariosas. . . . . . . . . . . . . . . . . . . . . . . . . . .15. M. subsagittata
2. Brácteas 12-37(-42) mm long., foliáceas. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .18. M. villosa
3(1). Brácteas de 3-7(-11) mm long.; sépalos de 5-9,5(- 11) x 2,5-6,5(-7,5) mm, obovados a angostamente obovados; anteras de 4,9-5,3 mm long.; semillas de (10-)12-16 mm long . . . . . . . . . . .16. M. subsessilis
3. Brácteas 1,3-4 x 0,8-1,1 mm long., sépalos 2,1-3,3 x 1-1,3 mm, ovados; anteras 2,9-3,5 mm long.; semillas 8-9 mm long. . . . . . . . . . . . . . .17. M. tubiflora
15. Mandevilla subsagittata (Ruiz & Pav.) Woodson, Ann. Missouri Bot. Gard. 19(1): 69. 1932. Echites subsagittatus Ruiz & Pav., Fl. Peruv. Prodr. 2: 19. 1799. TIPO: Perú: sin datos de localidad y fecha (fl), H. Ruiz & H. Pavón 11/56 (holotipo, MA!, foto F neg. 29216 INB!; isotipo, F!). Figs. 8 A-B.

Fig. 8. Mandevilla y Pentalinon. A-B, M. subsagittata.
Echites jasminiflorus M. Martens & Galeotti, Bull. Acad. Roy. Sci. Bruxelles 11: 357. 1844. Mesechites jasminiflorus (M. Martens & Galeotti) Miers, Apocyn. S. Amer.: 235. 1871. TIPO: México. Oaxaca, sine locus, sine data (fl), H. Galeotti 1602 (holotipo BR!; isotipo P!, foto INB!).
Echites cuspidifer S. F. Blake, Contr. Gray Herb. 52: 79. 1917. TIPO: Belice. Pine ridge cerca del Lago Manatee, 11-VI-1905 (fl, fr), M. Peck 35 (holotipo GH!; isotipo K!).
Referencia. Morales (2005c: 134).
Distribución y hábitat. México hasta Colombia, Venezuela, Ecuador y Perú. Crece en bosques húmedos, bosques estacionalmente secos o secos, en elevaciones de 100-1200 m s.m. Las flores y los frutos se producen de forma intermitente durante todo el año.
Nombres vulgares. "Flor de Veleta" (Ahuachapán).
Material representativo examinado
EL SALVADOR. Ahuachapán. San Francisco Menéndez, Hacienda San Benito, 16-XII-1991, E. Sandoval & Chinchilla 125 (B, LAGU, MO). Chalatenango. La Palma, 6-VII-1976, E. Montalvo 4690 (MO). La Libertad. Pelón de Comasagua, 7-16-II-1963, D. Winkler & J. Lagos 13 (EAP). Morazán. Arambala, Área Protegida Cerro Sapo, Cumaro, 14-V-2003, J. Monterrosa & M. Rivera 582 (INB, LAGU). Santa Ana. Entre Metapán y Cerro Monte Cristo, 30-VII-1977, T. Croat 42279 (MO). San Salvador. San Salvador, 1922, S. Calderón 815 (US).
16. Mandevilla subsessilis (A. DC.) Woodson, Ann. Missouri Bot. Gard. 19(1): 59. 1932. Echites subsessilis A. DC., Prodr. 8: 451. 1844. TIPO: México, sine locus (fl), H. Pavón s.n. (holotipo, G-DC!, foto F neg 34141 INB!; isotipo, MA!).
Mandevilla platydactyla Woodson, Ann. Missouri Bot. Gard. 19(1): 55. 1932. TIPO: México. Oaxaca: Tolaga, VI-1842 (fl), C. Liebmann 11986 (holotipo, C!). Mandevilla mollis Lundell, Contr. Univ. Michigan Herb. 7: 46. 1942. TIPO: México. Chiapas: Santa Rosa, Escuintla, 20-VI-1941 (fl), E. Matuda 4240 (holotipo, MICH!; isotipos, CAS!, F!, LL! [2 cartulinas], MO!, NY!).
Liana; ramitas densamente hirsútulo-tomentulosas o tomentulosas cuando jóvenes, volviéndose puberulentas a glabrescentes con la edad; coléteres interpeciolares inconspicuos. Hojas de 5-18,5 x 2,4-10,5 cm, angostamente elípticas, obovadoelípticas, angostamente obovadas a obovadas, el ápice caudado-acuminado, la base cordada, más raramente obtusa, los coléteres agrupados en la base del nervio medio, y distribuidos en grupos de 6 a 9 a lo largo del nervio medio hasta en 9 mm, láminas esparcidamente puberulentas a glabrescentes en la superficie adaxial, densa a moderadamente tomentosas a tomentulosas por el envés, de manera inusual puberulentas o glabrescentes, pecíolos de 2-7(-10) mm long. Inflorescencia con 5 a 15 (o más) flores, densamente a esparcidamente hirsútulas o puberulentas, pedúnculo de 10-38(- 120) mm long., pedicelos de 6-14 mm long., brácteas de 3-7(-11) mm, escariosas a foliáceas o subfoliáceas; sépalos de 5-9,5(-11) x 2,5-6,5(-7,5) mm, obovados a angostamente obovados, el ápice obtuso o redondeado-mucronulado, foliáceos, los coléteres distribuidos en hileras irregulares; corola hipocraterimorfa, amarilla, glabra en la superficie externa, tubo de 13-19 x 2,5-4 mm, recto, lóbulos de 4,5-10 x 3-6,5 mm, angostamente obovados; estambres insertos en el tercio distal del tubo, justo por debajo de la fauce, anteras de 4,9-5,3 mm long.; nectario anular, profunda e irregularmente pentalobulado, algunas veces algunos lóbulos totalmente divididos y formando nectarios independientes. Folículos de 20,5-32 cm x 2-3,5 mm, continuos, glabros; semillas de (10-)12-16 mm long., coma de 1,1-2,6 cm long.
Distribución y hábitat. SO de México, Guatemala y el N de El Salvador. Habita bosques subcaducifolios, en elevaciones de 800-1600 m s.m. La floración ocurre entre mayo y septiembre. El material con frutos es de julio y diciembre
Observaciones. En El Salvador, esta especie se puede reconocer fácilmente por sus sépalos foliáceos, de más de 5 mm long. En la última monografía del género, Woodson (1933) informó que esta especie era de Perú, pero más recientemente Morales (1998a) aclaró que en realidad este taxón está restringido al N de Centro América y el SO de México.
Material representativo examinado
EL SALVADOR. Ahuachapán. parque nacional El Imposible, 1-VIII-1991, R. Toledo 18 (B, LAGU, MO). La Libertad. Antiguo Cuscatlán, faldas de La Laguna, 10-VI-1991, R. Villacorta & R. Toledo 830 (B, INB, LAGU, MO). Sonsonate. Pedregal de San Isidro, S del lago Coatepeque, 29-VII-1977, T. Croat 42243 (INB, MO 2 láminas).
17. Mandevilla tubiflora (M. Martens & Galeotti) Woodson, Ann. Missouri Bot. Gard. 19(1): 52. 1932. Echites tubiflorus M. Martens & Galeotti, Bull. Acad. Roy. Sci. Bruxelles 11(1): 358. 1844. Amblyanthera tubiflora (M. Martens & Galeotti) Müll. Arg., Linnaea 30: 423. 1860. TIPO: México. Oaxaca: cerca de Xalapa, X.1841 (fl), H. Galeotti 1579 (holotipo, K!, foto INB!; isotipo, P!).
Echites cobanensis Donn. Sm., Bot. Gaz. 40(1): 6. 1905. TIPO: Guatemala. Alta Verapaz: Cobán, VIII.1904 (fl), H. Tuerckheim 8709 (holotipo, US!; isotipo, NY!). Mandevilla donnell-smithii Woodson, Ann. Missouri Bot. Gard. 19(1): 54. 1932. TIPO: Guatemala. Santa Rosa: Cerro Gordo, VIII.1892 (fl), E. Heyde & E. Lux 3993 (holotipo, GH!; isotipo, K!, foto INB!, NY!, US!).
Liana; tallos jóvenes hirsútulos a hirsútulopuberulentos, raramente glabrescentes, puberulentos o glabrescentes con la edad, coléteres interpeciolares inconspicuos. Hojas de 2-11 x (0,5-)0,7- 4,5(-6,5) cm, elípticas, muy angostamente elípticas, obovado-elípticas, ovado-elípticas, ovadas a angostamente ovadas, el ápice cortamente acuminado, cortamente cuspidado o más raramente agudo-mucronado, la base cordada, obtusa, redondeada a truncada, los coléteres agrupados en la base del nervio central, esparcidamente hirsútulas, puberulentas a glabrescentes o raramente glabras en la superficie abaxial, densamente tomentosas, tomentulosas o raramente glabrescentes abaxialmente, pecíolos de 4-27 mm long. Inflorescencia pluriflora, hirsútula o puberulenta, pedúnculo de (6-)10-70(-85) mm long., pedicelos de 5-12 mm long., brácteas de 1,3-4 x 0,8-1,1 mm, escariosas; sépalos de 2,1-3,3 x 1-1,3 mm, ovados, el ápice agudo a cortamente acuminado, escariosos, los coléteres en grupos alternos de 3-5; corola hipocrateriforme, amarilla o crema, glabra en la superficie externa, tubo de 12-17 x 1,7-3,3 mm, no giboso; de lóbulos 2,5-5(-6) x 2-3,5 mm, angostamente ovados a ovado-elípticos; estambres insertos cerca de la fauce, justo debajo de la boca, anteras de 2,9-3,5 mm long.; nectarios cinco, a veces dos o más connados entre sí, igualando al ovario o ligeramente más cortos. Folículos de 15-18 cm x 1,3-2,8 mm, continuos o torulosos, glabros; semillas de 8-9 mm long., coma de 1,3-2,7(-3,3) cm long.
Distribución y hábitat. México, Guatemala, Honduras y el N de Nicaragua. Habita bosques secos y formaciones de pino-encino en elevaciones de 1100-1500 m s.m. Especímenes con flores han sido coleccionados en febrero y agosto.
Observaciones. Mandevilla tubiflora se puede confundir con M. acutiloba, pero se separa por sus estambres insertos en la mitad distal del tubo de la corola por debajo de la boca del tubo y botones florales con el ápice agudo. Esta especie no había sido registrada previamente para El Salvador (Morales, 1998a).
Material representativo examinado
EL SALVADOR. Chalatenango. al NO del cerro El Pital, cerca de San Ignacio, 7-16-II-1963 (fl), S. Winkler & E. Montalvo 10 (EAP). Santa Ana. entre la Peña del Cuervo y el cerro El Yupe, 18-VIII-1994 (fl), J. Linares & C. Martínez 1614 (EAP)
18. Mandevilla villosa (Miers) Woodson, Ann. Missouri Bot. Gard. 19(1): 70. 1932. Laseguea villosa Miers, Apocyn. S. Amer.: 250. 1878. TIPO: Nicaragua, sine locus, 1848 (fl), C. Seemann 95 (holotipo, BM!; isotipo, K!). Fig. 8 C.

Fig. 8. Mandevilla y Pentalinon. C, M. villosa.
Echites comosus O. Kuntze, Revis. Gen. Pl. 2: 414. 1891. TIPO: Panamá. Colón: Gale, sin fecha (fl), O. Kuntze 1891 (lectotipo NY!, designado por Morales, Anales Jard. Bot. Madrid 62(1): 67. 2005, fotocopia, INB!).
Referencia. Morales (2005c: 135).
Distribución y hábitat. México, Guatemala y de El Salvador a Colombia. Habita bosques muy húmedos, de 0-1000 m s.m. Especímenes con flores han sido coleccionados en febrero, julio y septiembre. La fructificación se ha registrado entre agosto y septiembre.
Material representativo examinado
EL SALVADOR. Ahuachapán. San Francisco Menéndez, El Corozo, 14-VII-2000, J. Rosales 1070 (B, BM, INB, LAGU, MO). Cabañas. Cinquera, calle a San Antonio, 29-VIII-2002, J. Monterrosa & R. Carballo 298 (INB, LAGU). La Libertad. San Juan Buenavista, 19-IX-2000, R. Carballo et al. s.n. (B, BM, LAGU, MO). San Salvador. San Salvador, VII-1922, S. Calderón 938 (US).
13. MESECHITES Müll. Arg., in Martius, Fl. Bras. 6(1): 150. 1860.
Género con 8 especies desde México y las Antillas a Bolivia y Argentina (excepto Uruguay y Chile); con solo una de ellas en El Salvador. (Morales, 2006; Woodson, 1933). Para la descripción del género y mayores detalles véase Morales (2005c: 137).
19. Mesechites trifidus (Jacq.) Müll. Arg., in Martius, Fl. Bras. 6(1): 151. 1860. Echites trifidus Jacq., Enum. Syst. Pl. 13. 1760. TIPO: Tab. 24 en Jacquin, Select. Stirp. Amer. Hist. 1763 (lectotipo, designado por Morales, Darwiniana 43: 137. 2005).
Referencia. Morales (2005 c: 137).
Distribución y hábitat. México a Trinidad, Brasil, Bolivia, Paraguay y el N de Argentina. Bosques secos y bosques muy húmedos, en elevaciones de 0-1000 m. Floración y fructificación se produce durante todo el año.
Nombres vulgares. "Loroquillo" (Ahuachapán).
Material representativo examinado
EL SALVADOR. Ahuachapán. San Francisco Menéndez, El Corozo, 5-IV-2000, J. Rosales 515 (B, BM, LAGU, MO). San Miguel. Laguna El Jocotal, 12- VI-1996, R. Villacorta 2385 (LAGU, MO). Santa Ana. Cabanas, E de Ilobasco, 21-I-1998, G. Davidse et al. 37123 (B, INB, ITIC, LAGU, M, MO). San Vicente. cercanías de San Vicente, 11-III-1922, P. Standley 21864 (NY, US)
14. NERIUM L., Sp. Pl. 1: 209. 1753. Género monotípico originario de Eurasia, pero ampliamente cultivado en todo el mundo. (Pagen, 1987). Para la descripción del género y mayores detalles véase Morales (2005c: 139).
20. Nerium oleander L., Sp. Pl. 1: 209. 1753. TIPO: Creta, Palaestina, Syria, India, sin fecha (fl), Herb. Clifford 76, Nerium No. 1B (lectotipo, BM!, designado por Stearn, Fl. Turkey 6: 159. 1978).
Referencia. Morales (2005c: 139).
Distribución y hábitat. Conocido únicamente en cultivo, entre los 100-1100 m s.m. Florece durante todo el año.
Observaciones. Nerium es fácil de reconocer por ser una especie cultivada, con hábito arbustivo, hojas verticiladas, corola con 5 lóbulos coronales petaloides opuestos a cada antera, así como sus frutos foliculares de tipo doble folículo, originados por un ovario sincárpico. Las plantas cultivadas no producen frutos con regularidad.
Material representativo examinado
EL SALVADOR. La Libertad. Jardín Botánico La Laguna, zona 1, 17-V-2005 (fl), J. Monterrosa 900 (LAGU).
15. PENTALINON Voigt, Host. Suburb. Calcutt.: 523. 1845.
Lianas con tallos algo aplanados cuando jóvenes, cilíndricos a subcilíndricos con la edad, con secreción lechosa, glabros, glabrescentes o pubescentes, coléteres intrapeciolares diminutos, pero evidentes. Hojas opuestas, sin coléteres en el nervio central, lámina glabra, glabrescente a puberulenta o hirsútula, membranáceas, pecioladas. Inflorescencias cimosas, axilares, subterminales o terminales, pauci- a pluriflora, brácteas escariosas a foliáceas, deciduas o algunas pocas persistentes; cáliz con 5 sépalos, iguales o subiguales, escariosos, glabros o glabrescentes, con varios coléteres en la base de la cara adaxial; corola infundibuliforme, sin corona anular o lóbulos coronales libres, amarilla o crema, limbo con 5 lóbulos, con estivación dextrorsa; estambres totalmente inclusos, anteras conniventes y aglutinadas a la cabeza estigmática, con conspicuos apéndices filiformes apicales, cabeza estigmática fusiforme, con un anillo en la base; gineceo bicarpelar, apocárpico, con numerosos óvulos, nectario anular, irregularmente lacerado en el ápice. Frutos apocárpicos, foliculares, fusiformes, lisos y continuos, subleñosos; semillas desnudas y secas, numerosas, rostradas, comosas en el ápice micropilar.
Género con 2 especies confinadas al S de Estados Unidos de América y las Antillas hasta Nicaragua, una de ellas presente en El Salvador. (Hansen & Wunderlin, 1986; Woodson, 1936).
21. Pentalinon andrieuxii (Müll. Arg.) B. F. Hansen & Wunderlin, Taxon 35(1): 168. 1986. Urechites andrieuxii Müll. Arg., Linnaea 30: 442. 1860. TIPO: México. Oaxaca: cerca de Tehuantepec, VIII-1834 (fl, fr), G. Andrieux 247 (lectotipo, G-DC! designado por Morales, Lankesteriana 5: 159. 2005, foto F neg. 26898 INB!; isolectotipo, P!). Fig. 8 D.

Fig. 8. Mandevilla y Pentalinon.D, Pentalinon andrieuxii.
Echites cupulifer L. O. Williams, Fieldiana, Bot. 32(12): 179, f. 1. 1970. TIPO: Guatemala. Petén: Tikal, Parque Nacional, 5-XI-1969 (fl), R. Ortíz 418 (holotipo, F!, foto F neg. 56461 INB!; isotipo, MO!)
Liana, tallos glabros, glabrescentes o diminutamente puberulentos. Hojas de 4,5-11,8 x 2-7,2 cm, elípticas a obovado-elípticas, el ápice acuminado o caudado-acuminado, la base obtusa o redondeada, glabras o glabrescentes en la superficie adaxial, glabras, glabrescentes o esparcidamente puberulentas por el envés, pecíolo de 0,7-2,1 cm long.. Inflorescencia glabra o glabrescente, pedúnculo de (1,5-)6- 12 cm long., pedicelos de 1,6-4,1 cm long., brácteas de 1-1,5 mm long.; sépalos de 4-6 mm long., ovados, el ápice acuminado; corola verde-amarilla o amarilla, glabra, la parte inferior del tubo de 0,9-1,6 cm long., la parte superior de 2-4,5 cm long., tubular a tubularcampanulada, con un diámetro de 0,6-1,2 cm en la garganta, lóbulos de 1,2-2,4 x 0,8-1,2 cm, obovados; anteras de 6-7 mm long., apéndices apicales de 7-10 mm long., ovario de 1,5-2 mm long., glabro, disco anular ca. 3 mm long., entero a levemente lacerado. Folículos de 14-30 cm x 5-8 mm; semillas de 1,5-2 cm long., coma de 1,2-1,6 cm long.
Distribución y hábitat. S de Estados Unidos de América a Nicaragua. Habita bosques secos, en elevaciones de 0-500 m s.m. Especímenes con flores han sido coleccionados entre abril y agosto.
Observaciones. Pentalinon andrieuxii se puede reconocer con facilidad en El Salvador, por sus anteras con conspicuos apéndices apicales, filiformes y entrelazados en espiral, usualmente de más de 7 mm long. Puede ser confundido con Nerium y Tintinnabularia (este último no registrado en El Salvador), géneros cuyas anteras también tienen apéndices, pero Pentalinon difiere de Nerium por sus hojas opuestas (vs. verticiladas), mientras que de Tintinnabularia se separa por sus hojas que carecen de coléteres en la base del nervio central en la superficie adaxial de la hoja, así como de crestas o costillas longitudinales en la cabeza estigmática.
Material representativo examinado
EL SALVADOR. Ahuachapán. San Francisco Menéndez, El Corozo, 26-V-2000, J. Rosales 843 (BM, LAGU, MO). La Libertad. Al O de la Libertad, 6-VIII- 1962, G. Webster et al. 12804 (MO, UC).
16. PLUMERIA L., Sp. Pl. 1: 209. 1753.
Género con 7 a 8 especies distribuidas desde México y las Antillas a las Guyanas, Venezuela y Perú, con una sola citada para El Salvador. (Woodson, 1938b). Para la descripción del género y mayores detalles véase Morales (2005c: 145).
22. Plumeria rubra L., Sp. Pl. 1: 209. 1753. TIPO: Habitat in Jamaica, Surinamo, sin fecha (fl), Sloane, Voy. Jamaica, 2: 61, t. 185, f. 1, 1725 (lectotipo, BM-SL!, designado por Wijnands, Bot. Commelins: 44. 1983). Fig. 4 D.
Plumeria tricolor Ruiz & Pav., Fl. Peruv. 2: 20, t. 139. 1799. TIPO: Perú. Lima: Lima, ene-feb, 1778-1788 (fl), H. Ruiz & H. Pavón 11/59 (lectotipo, MA!, designado por Morales, Darwiniana 43: 145. 2005). Plumeria purpurea Ruiz & Pav., Fl. Peruv. 2: 20, t. 187. 1799. TIPO: Perú. Lima: Lima, ene-feb, 1778-1788 (fl), H. Ruiz & H. Pavón 11/58 (lectotipo, MA! designado por Morales, Darwiniana 43: 145. 2005).
Referencia. Morales (2005c: 145).
Distribución y hábitat. México y las Antillas a Venezuela y Perú, introducida en el S de Suramérica (Brasil, Bolivia y Paraguay) y en áreas tropicales y subtropicales de todo el mundo. Habita bosques secos, bosques estacionalmente secos, más raramente en bosques húmedos, en elevaciones de 200- 1400 m s.m. La floración y la fructificación ocurren principalmente de marzo a junio, pero también de manera irregular en el resto del año.
Nombres Vulgares. "Flor Blanca" (Ahuachapán); "Flor de mayo", "Flor de Ensarta" (Ahuachapán); "Pacayita" (Santa Ana); "Pacallita macho" (Santa Ana).
Observaciones. Existen bastantes variedades cultivadas, en las cuales la coloración de las flores varía desde amarilla a rosada o rojiza. Sin embargo, en las poblaciones naturales las flores blancas son predominantes. El ovario semi-ínfero de este género es algo bastante raro en las Apocynaceae s. str.
Material representativo examinado
EL SALVADOR. Ahuachapán. El Imposible, San Benito, 8-III-1992, E. Sandoval & Chinchilla 296 (LAGU, MO). Chalatenango. La Palma, El Caballero, XII-2001, M. Smeets & F. Quiñónez 367 (ITIC, LAGU, U). La Libertad. Primavera, Quetzaltepeque, 2-II- 1998, A. Monro et al. 2888 (BM, MO). Santa Ana. San José Ingenio, Parque Nacional Montecristi, 20-IX-2001, V. Martínez 93 (INB, LAGU).
17. PRESTONIA R. Br., Asclepiadeae: 58. 1810.
Género con 55 especies distribuidas desde México y las Antillas a Argentina (excepto Chile); 4 son conocidas en el Salvador (Morales, 1997a, Woodson, 1936). Para la descripción del género y mayores detalles véase Morales (2005 c: 147).
1. Corola y hojas variadamente pubescentes . . . . . . . .2
1. Corola y hojas glabras o glabrescentes, nunca conspicuamente pubescentes . . . . . . . . . . . . . . . . . . . . . . .3
2(1). Corola infundibuliforme . . . . . . . .26. P. speciosa
2. Corola hipocraterimorfa . . . . . . . . . .24. P. mexicana
3(1). Sépalos 3-6(-8) x 2-3 mm . . . . . .23. P. longifolia
3. Sépalos (9-)10-18(-21) x 4-7 mm . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .25. P. portobellensis
23. Prestonia longifolia (Sessé & Moç.) J. F. Morales, Novon 6(3): 286. 1996. Echites longifolius Sessé & Moç., Fl. Mexic.: 45. 1893. TIPO: México. Veracruz: San Andrés Tuxtla, sin fecha (fl), J. Sessé & J. Moçiño 5007 (lectotipo, MA!, designado por Morales, Novon 6: 287. 1996, foto INB!; isolectotipo, F!). Fig. 9A.

Fig. 9. Prestonia, Rauvolfia y Stemmadenia. A, P. longifolia.
Belandra concolor S. F. Blake, Contr. Gray Herb. 52: 78. 1917. Prestonia concolor (S. F. Blake) Woodson, Field Mus. Nat. Hist., Bot. Ser. 12: 327. 1936. TIPO: Belice. Bancos del río Grande, 25-III-1907 (fl), M. Peck 953 (holotipo, GH!, fotocopia INB!).
Referencia. Morales (2005c: 148).
Distribución y hábitat. México al N de Colombia. Crece en bosques muy húmedos, en elevaciones de 300-500 m s.m. Especimenes con flores fueron coleccionados en febrero.
Observaciones. Esta especie puede ser confundida con Prestonia portobellensis, ya que las hojas y frutos de ambas especies son prácticamente idénticas, pero P. longifolia puede ser separada con facilidad por sus sépalos más pequeños y angostos. Véase la sinonimia adicional en Morales (1996, 2005c).
Material representativo examinado
EL SALVADOR. Ahuachapán. San Francisco Menéndez, río Cuayapa, I-1996, M. Sandoval & Chinchilla 283 (B, LAGU, MO).
24. Prestonia mexicana A. DC., Prodr. 8: 429. 1844. Mitozus mexicanus (A. DC.) Miers, Apocyn. S. Am.: 225. 1878. TIPO: México. Oaxaca: San Bartolo, Oaxaca, VIII-1834 (fl), G. Andrieux 251 (holotipo, G-DC!, foto F neg. 7542 INB!; isotipos, FI-W!, K! [2 cartulinas], P!).
Prestonia amanuensis Woodson, Ann. Missouri Bot. Gard. 23(2): 359. 1936. TIPO: Belice: 6 millas de línea Stan Creek, VIII-1929 (fl), W. Schipp S-7 (lectotipo, NY, designado por Morales, Novon 7: 61. 1997; isolectotipos, F!, foto F neg. 56492 INB!, GH!, Z!, foto INB!).
Referencia. Morales (2005c: 149).
Distribución y hábitat. México al NO de Colombia (Chocó). Crece en bosques muy húmedos a bosques estacionalmente secos, en elevaciones de 0-1300 m s.m. Florece en enero y de mayo a agosto. Fructifica de septiembre a febrero.
Observaciones. Esta especie se puede confundir con P. speciosa, ya que ambos taxones comparten caracteres morfológicos similares, pero P. mexicana se reconoce con facilidad por sus corolas hipocraterimorfas, en contraposición de P. speciosa, donde tenemos corolas claramente infundibuliformes. Sin embargo, especímenes con frutos pueden ser difíciles de separar. Para una sinonimia adicional véase Morales (1997a, 2005c).
Material representativo examinado
EL SALVADOR. Ahuachapán. San Francisco Menéndez, El Corozo, 19-VIII-2000, J. Rosales 765 (B, BM, LAGU, MO). Chalatenango. Ojos de Agua, Las Vueltas, 16-VI-1997, R. Villacorta 2393 (LAGU, MO). La Libertad. Carretera a La Libertad, 8-VII-1957, 23- II-1976, Lagos 836 (USF). San Miguel. NO de San Miguel, T. Croat 32798 (MO, Z).
25. Prestonia portobellensis (Beurl.) Woodson, Ann. Missouri Bot. Gard. 18(4): 553. 1931. Echites portobellensis Beurl., Kongl. Vetensk. Acad. Handl. 40: 137. 1854. [1856]. TIPO: Panamá. Colón: Portobello, IV-1826 (fl), J. Billberg s.n. (holotipo, S!, foto MO!). Fig. 3 D.
Prestonia guatemalensis Woodson, Ann. Missouri Bot. Gard. 23(2): 339. 1936. TIPO: Guatemala. Alta Verapaz, Sepacuitle, X-1901 (fl), P. Owen 1 (holotipo, US!, foto INB!). Prestonia laxa Rusby ex Woodson, Ann. Missouri Bot. Gard. 23(2): 317. 1936. TIPO: Colombia. Magdalena, Valparaiso, 26-III-1899 (fl), H. Smith 1647 (holotipo, NY no visto, fotocopia en INB; isotipos, CM, GH, K no vistos) Prestonia schippii Woodson, Ann. Missouri Bot. Gard. 23(2): 337. 1936. TIPO: Belize, El Dorado, IX-1922 (fl), W. Schipp S-388 (holotipo, F!, foto F neg. 52873 INB!).
Referencia. Morales (2005c: 150).
Distribución y hábitat. México a Colombia, el N de Venezuela y Ecuador. Habita bosques muy húmedos, en elevaciones de 0-800 m s.m. La floración y la fructificación se producen de febrero a mayo.
Nombres vulgares. "Bejuco loroco de venado" (Ahuachapán).
Observaciones. Prestonia portobellensis presenta una alta variación en los patrones de coloración de los lóbulos de la corola, de manera que en algunas poblaciones los lóbulos pueden tener motas o líneas intensamente moradas o lilacinas, mientras que en otras éstos son de un color verde-crema, con una ausencia casi total de marcas lilacinas. Material representativo examinado
EL SALVADOR. Ahuachapán. San Benito, 18-IV- 1995, M. Sandoval & E. Sandoval 894 (B, EAP, INB, LAGU, MO). San Vicente. Las Galeras, entre San Miguel y San Vicente, 9-II-1998, G. Davidse et al. 37486 (B, BM, ITIC, LAGU, MO). Sonsonate. Cerca de Sonsonate, III-1922, P. Standley 22349 (GH, NY, US).
26. Prestonia speciosa Donn. Sm., Bot. Gaz. 27(6): 435. 1899. TIPO: Guatemala. Santa Rosa: Buena Vista, IV-1893 (fl), N. Heyde & E. Lux 4497 (holotipo, US!; isotipos, BM!, foto INB!, GH!, K!, US! [3 cartulinas]).
Prestonia grandiflora L. O. Williams, Fieldiana, Bot. 31(17): 402. 1968. TIPO: México. Chiapas: La Grandeza, 19-V-1945 (fl), E. Matuda 15570 (holotipo, F!; isotipos, EAP!, F!, foto F neg. 56493 y 56494 INB!, MO! [2 cartulinas], S!, TEX-LL!).
Liana con ramitas pubescentes, algunas veces glabrescentes en los tallos viejos, con secreción acuosa, coléteres intrapeciolares conspicuos, 3,5-5 mm long. Hojas con láminas de 8-26 x 6-13,5 cm, obovadas, anchamente elípticas, o orbiculares, el ápice acuminado a caudado-acuminado, la base cuneada, obtusa a redondeada, membranáceas, esparcidamente puberulentas a glabrescentes en la superficie adaxial, tomentosas o tomentulosas por el envés, pecíolo de 0,4-1,1 cm long. Inflorescencia axilar, tomentulosa, pedúnculo de 0,4-2,2 cm long., pedicelos de 0,9-1,7 cm long., brácteas de 3- 12 mm long., foliáceas o subfoliáceas; sépalos de 9-18 x 4-8 mm, ovados, ovado-elípticos, a angostamente elípticos, el ápice agudo a acuminado o mucronulado; corola infundibuliforme, amarilla, tomentulosa en la superficie externa, parte inferior del tubo 12-19 x 3-4 mm long., la parte superior 10-20 long., cónica a cónico-campanulada, con una diámetro de 10-20 mm en la fauce, lóbulos coronales ausentes, reducidos a crestas callosas inconspicuas localizadas sobre los filamentos, corona anular de 1-1,5 mm long., lóbulos de la corola 20-25 x 11-14 mm, obovados; anteras de 8- 9 mm long., inclusas, nectario pentalobulado a irregularmente lacerado, más largo que el ovario. Folículos de 7-9 x 1,8-2,7 cm, divaricados, seríceos o tomentulosos; semillas de 11,5-14 mm long., coma de (1,8-)2-3(-3,3) cm long.
Distribución y hábitat. S de México al N de Nicaragua. Habita bosques húmedos y bosques nubosos, en elevaciones de 1300-2300 m s.m. Especímenes con flores han sido coleccionados de febrero hasta agosto. Especímenes con frutos han sido coleccionados en febrero.
Observaciones. Es una de las especies más fáciles de reconocer dentro del género, Prestonia speciosa se distingue al por sus corolas infundibuliformes. Este carácter es compartido únicamente con P. clandestina, endémica de México, que se diferencia por sus corolas con el tubo mucho más corto.
Material representativo examinado
EL SALVADOR. Ahuachapán. Parque Nacional El Imposible, San Benito, 11-VI-1997, M. Sandoval 1586 (B, EAP, INB, LAGU, MO). La Libertad. Puerta de La Laguna, 27-IV-1922, P. Standley 23673 (GH, US). Sonsonate. Laguna de Las Ninfas, 4-III-1991, R. Villacorta & Cortez 742 (LAGU, MO).
18. RAUVOLFIA L., Sp. Pl. 1: 208. 1753.
Género pantropical con ca. de 100 especies (35 en el neotrópico y 65 en el paleotrópico); únicamente dos se registran en El Salvador. (Rao, 1956). Para la descripción del género y mayores detalles véase Morales (2005c: 153).
1. Hojas mayormente en verticilos de 3 . . . . . . . . . . . . . . . .27. R. ligustrina 1. Hojas mayormente en verticilos de 4 . . . . . . . . . .. . . . . .28. R. tetraphylla
27. Rauvolfia ligustrina Willd. ex Roem. & Schult., Syst. Veg. 4: 805. 1819. TIPO: Colombia, sine locus, sine data (fl), A. Humboldt & B. Bonpland s.n. (holotipo, P-HB!). Fig. 9 B.
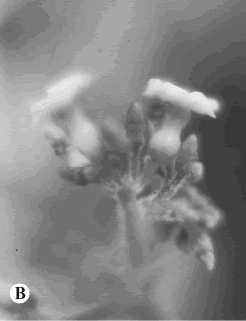
Fig. 9. Prestonia, Rauvolfia y Stemmadenia.B, R. ligustrina.
Rauvolfia ternifolia Kunth, Nov. Gen. Sp. 3: 232. 1819. TIPO: Colombia. Bolivar: río Magdalena, cerca de Mompox, IV-(año perdido), A. Humboldt 1480 (holotipo, P-HB!; isotipo B no visto, foto F neg. 4475 INB!).
Referencia. Morales (2005c: 155).
Distribución y hábitat. México al NE de Brasil y Bolivia y las Antillas. Crece en bosques secos, en elevaciones de 0-400 m s.m. Especimenes con flores se han coleccionado en junio y julio. Los frutos han sido observados en enero, febrero, mayo, junio, septiembre y noviembre.
Nombres vulgares. "Amatillo" (La Libertad); "Canforillo" (San Miguel); "Chalchupa" (Ahuachapán).
Observaciones. A pesar de su amplia distribución mundial, esta especie no es común en Mesoamérica, y usualmente se conoce solo de poblaciones aisladas en cada país. En diversas etiquetas de campo se ha informado que su raíz es medicinal, sin especificar su acción terapéutica.
Material representativo examinado
EL SALVADOR. Ahuachapán. Área Protegida Santa Rita, 2004, J. Rosales 2475 (INB, LAGU). La Libertad. San Diego, 21-IX-1997, R. Aparicio & R. Rivera 5 (INB, LAGU, MO).
28. Rauvolfia tetraphylla L., Sp. Pl. 1: 208. 1753. TIPO: Habitat in America calidiore, sin fecha (fl), Herb. Clifford 75, Rauvolfia No. 1, (lectotipo, BM, designado por Stearn, Introd. Linnaeus ´ Sp. Pl. (Ray. Soc. Ed.: 47. 1957). Fig. 2 A.

Fig. 2. Coléteres y domacios. A, coléteres (Rauvolfia tetraphylla).
Rauvolfia hirsuta Jacq., Enum. Syst. Pl. 14. 1760. TIPO: Sin datos, Jacquin s.n. (no localizado). Rauvolfia tomentosa Jacq., Enum. Syst. Pl. 14. 1760. TIPO: Colombia. Atlántico: Cartagena, sin fecha, Jacquin s.n. (no localizado).
Referencia. Morales (2005c: 157).
Distribución y hábitat. México y las Antillas hasta Venezuela y Perú; introducida en el S de Estados Unidos y Africa. Habita bosques secos o estacionalmente secos, en elevaciones de 0-850 m s.m. Las flores y los frutos se producen en forma intermitente durante casi todo el año.
Nombres vulgares. "Amatillo", "Chalchupa" (Ahuachapán); "Cojoncito" (Ahuachapán).
Observaciones. Rauvolfia tetraphylla, es una hierba común, que llega a formar rodales bastantes puros en áreas alteradas o de crecimiento secundario. Aunque se puede confundir con R. ligustrina, se distingue por sus hojas dispuestas mayormente en verticilos de a 4.
Material representativo examinado
EL SALVADOR. Ahuachapán. San Francisco Menéndez, El Corozo, Los Peralta, 2-V-2000, J. Rosales 675 (LAGU, MO). Chalatenango. Nueva Concepción, Buenos Aires, área protegida Pañanalapa, V-2000, K. King & J. Chávez 40 (INB, LAGU). Cuscatlán. Cuscatlán, 15-VIII-1998, Grupo Ecológico Cantón La Bermuda 3340 (LAGU, MO). La Libertad. La Cabaña, 20- VIII-1987, J. Flores s.n. (LAGU, MO). La Paz. La Lucha, Zacatecoluca, 10-VI-1994, J. González 162 (LAGU, MO). La Unión. Dunas costeras, 23-III-1939, A. Beetle 26272 (MO, UC). San Miguel. Laguna El Jicotal, 17-V-1996, R. Villacorta & E. Echeverría 2344 (LAGU, MO). San Salvador. Jardín Botánico La Laguna, 18-IX-1987, J. Flores 267 (LAGU, MO). Santa Ana. Coatepeque, 29-VI-1971, W. Harmon & J. Fuentes 5938 (MO). Sonsonate. Acajutla, 15-VIII-1971, D. Spellman et al. 747 (MO).
19. STEMMADENIA Benth., Bot. Voy. Sulphur: 124. 1844 [1845].
Género con 16 especies distribuidas desde México al N de Brasil, Ecuador, Perú y el N de Bolivia, pero con un marcado centro de diversidad en el S de Mesoamérica; 3 especies se conocen en El Salvador. (Leeuwenberg, 1994; Morales & Méndez, 2005). Para la descripción del género y mayores detalles véase Morales (2005c: 159).
1. Corola conspicuamente infundibuliforme, la parte superior del tubo con un diámetro de 14-17 mm en la fauce; parte superior del tubo de la corola iguala en longitud la parte inferior del tubo . . . . . . . . . . . . . . . . .31. S. pubescens
1. Corola hipocraterimorfa o angostamente infundibuliforme, entonces con el tubo expandiéndose solo levemente en el tercio distal justo antes de la fauce, el diámetro de la fauce nunca excede los 9 mm de ancho; parte inferior del tubo (en corolas angostamente infundibuliformes), más corta que la parte superior . . . . . . . . . . . . . . . .2
2(1). Sépalos marcadamente desiguales en longitud, los externos mucho más pequeñas que las internas, las diferencias en longitud más de 1/3 long. total del sépalo . . . . . . . . . . . . . . . . . . .29. S. donnell-smithii
2. Sépalos relativamente similares en longitud, algunas veces algunos levemente más pequeños, pero la diferencia menos de 1/5 de la longitud total del sépalo . . . . . . . . . . .30. S. eubracteata
29. Stemmadenia donnell-smithii (Rose) Woodson, Ann. Missouri Bot. Gard. 15(4): 369. 1928. Tabernaemontana donnell-smithii Rose, Bot. Gaz. 18(6): 206. 1893. TIPO: Guatemala: Retalhuleu, San Felipe, IV-1892 (fl), J. Donnell- Smith 2763 (lectotipo US!, designado por Woodson, Ann. Missouri Bot. Gard. 15: 369. 1928; isolectotipos, A!, F!, G!, GH!, K!, M!, MO!, NY!, US! [2 cartulinas]). Fig. 9 C.
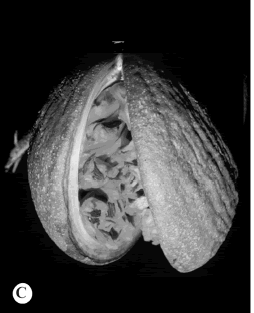
Fig. 9. Prestonia, Rauvolfia y Stemmadenia.C, S. donnell-smithii.
Referencia. Morales (2005c: 162).
Distribución y hábitat. México a Panamá. Habita bosques muy húmedos, bosques estacionalmente secos, en elevaciones de 20-900 m s.m. Las flores y los frutos se producen durante todo el año.
Nombres vulgares. "Cojón" (San Salvador); "Cojón de puerco" (San Salvador); "Cojón de tunco" (Ahuachapán).
Observaciones. Esta especie ha sido ocasionalmente confundida con S. pubescens, pero se puede separar con facilidad por sus corolas hipocraterimorfas (vs. infundibuliformes), así como por sus folículos con el ápice obtuso (vs. acuminado).
Material representativo examinado
EL SALVADOR. Ahuachapán. San Francisco Menéndez, El Corozo, Alto Los Sánchez, 12-IV-2000, J. Rosales 558 (INB, LAGU). La Libertad. Antigua Cuscatlán, 25-XI-1999, R. Guerra et al. 5 (BM, MO). San Salvador. Al S de San Salvador, IV-1958 (fl), P. Allen 6840 (EAP). Santa Ana. Santa Lucía, Santa Ana, 27-IV- 1995, J. Linares 2586 (MO).
30. Stemmadenia eubracteata Woodson, Ann. Missouri Bot. Gard. 15(4): 368, t. 49, f. 2. 1928. TIPO: Guatemala. Santa Rosa: Volcán de Tecuamburro, II-1893 (fl), E. Heyde & E. Lux 4538 (holotipo, GH!; isotipos, BM!, F!, K!, M!, US!). Fig. 9 D.

Fig. 9. Prestonia, Rauvolfia y Stemmadenia.D, S. eubracteata.
Arbusto o árboles de 1,5-4 m de altura, tallos glabros. Hojas de 3,2-13(-14,5) x 1,8-4,6(-5,5) cm, elípticas, ovado-elípticas a obovado-elípticas, ápice acuminado a caudado-acuminado, base obtusa a anchamente cuneada, glabras, raramente con unos cuantos pelos esparcidos a lo largo del nervio central por el envés, pecíolo de 2-9 mm long.. Inflorescencia con 1-4 flores, pedúnculo de 1-3 mm long., pedicelos de 4-8 mm long., brácteas de 0,5-3,5 mm long.; sépalos iguales a subiguales, 5-12(-16) x 2-5,5 mm, foliáceos, angostamente elípticos, angostamente ovado-elípticos a obovado- elípticos, el ápice agudo a acuminado; coro la angostamente infundibuliforme, el tubo expandido en el tercio distal, amarilla, tubo de 1,4-2,2 cm long., con un diámetro de 4,5-6,5(-7) mm en la fauce, lóbulos de 1,3-1,7 cm long., obovados; anteras de ca. 3,5 mm long., ovario de 1,5-2 mm long., nectario pentalobulado, ca. la mitad de la longitud del ovario. Frutos de 2-2,5 x 1,2-1,5 cm, verdes o verde-amarillentos al madurar, apiculados apicalmente; semillas no examinadas.
Distribución y hábitat. S de México, Guatemala, El Salvador y Honduras, habita en bosques húmedos entre 800-1700 m s.m. Especímenes con flores han sido coleccionadod entre enero y abril. Material con frutos se conoce de enero, junio, septiembre y octubre.
Nombres vulgares. "Cojón", "Cojón de altura", "Cojoncito verde de altura", "Cojón de tierra fría" (Ahuachapán); "Cojón de Montana" (Ahuachapán); "Cojón de Toro" (Santa Ana)".
Observaciones. Esta especie fue incorrectamente incluida en la sinonimia de S. macrophylla Greenm. por Leeuwenberg (1994). Sin embargo, recientemente Morales & Méndez (2005) realizaron una nueva sinopsis del género, estableciendo que el tipo de S. macrophylla debía ser reducido a la sinonimia de S. litoralis (Kunth) Allorge (no presente hasta el momento en El Salvador) y S. eubracteata debe ser tratado como un taxón válido. Asimismo, aunque S. eubracteata no fue citado para Honduras en esa sinopsis, el estudio de colecciones en los herbarios EAP y TEFH han confirmado su presencia en ese país. Para más detalles sobre la sinonimia y la historia taxonómica de esta especie vease el trabajo de Morales & Méndez (2005).
Material representativo examinado
EL SALVADOR. Ahuachapán. San Benito, N del Cerro León, 10-VI-1993, M. Sandoval & Fuentes 1308 (B, LAGU, MO). Santa Ana. San José Ingenio, Parque Nacional Montecristo, 17-IX-2001, V. Martínez 64 (INB, LAGU, MO).
31. Stemmadenia pubescens Benth., Bot. Voy. Sulphur: 125. 1845. Bignonia (?) obovata Hook. & Arn., Bot. Beechey Voy. 439. 1841, nom. illeg., no Spreng., (1825). Stemmadenia obovata K. Schum., Nat. Pflanzenfam. 4(2): 149. 1895, nom. illeg. TIPO: México. San Luis Potosí: Realejo, sin fecha (fl), Sinclair s.n. (holotipo K!; isotipo K!, fotos INB!). Figs. 10 A-B.

Fig. 10. Stemmadenia pubescens. A, corola. B, folículos.
Stemmadenia glabra Benth., Bot. Voy. Sulphur: 124, t. 44. 1845. TIPO: Honduras. Vallea: Golfo de Fonseca, Choluteca, sin fecha (fl), Sinclair s.n. (lectotipo, K! [2 cartulinas], designado por Nelson, Fontqueria 44: 59. 1996, fotos, INB!).
Stemmadenia mollis Benth., Bot. Voy. Sulphur: 125. 1845. Stemmadenia obovata Benth. var. mollis (Benth.) Woodson, Ann. Missouri Bot. Gard. 15(4): 358. 1928. TIPO: Ecuador. Guayaquil: Guayas, sin fecha (fl), Sinclair s.n. (holotipo, K!; foto, INB!).
Referencia. Morales (2005c: 167).
Distribución y hábitat. México a Colombia, Ecuador, Perú y Bolivia. Crece en bosques secos, en elevaciones de 0-1300 m s.m., pero es más común bajo los 800 m s.m. Las flores y los frutos se producen durante todo el año.
Nombres vulgares. "Cojón" (Cabañas); "Cojoncillo" (Ahuachapán); "Huevos de toro" (San Miguel); "Cojón verde" (Ahuachapán); "Cojón de costa hoja de hacha" (Ahuachapán); "Cojón picudo" (Ahuachapán).
Observaciones. Es el taxón más común en bosques secos, que se diferencia del resto de las especies presentes en El Salvador por el tamaño de sus flores y por la corola infundibuliforme. Para más información sobre la historia taxonómica de esta especie, véase Morales & Méndez (2005).
Material representativo examinado
EL SALVADOR. Ahuachapán. El Refugio, 17-V- 1997, J. González & M. Hernández 442 (LAGU, MO). Cabañas. Cinquera, cerro Tepeagua, 22-IV-2002, R. Carballo et al. 290 (INB, LAGU). Chalatenango. Nueva Concepción, área protegida Pañanalapa, Los Chorros, 17-V-2000, K. King & J. Chávez 30 (INB, LAGU). La Libertad. San Diego, El Amatal, 12-IV- 1997, R. Aparicio & R. Rivera 139 (LAGU, MO). La Unión. Tierra Blanca, 13-VI-1970, Davidse & Pohl 2080 (MO). San Miguel. Volcán Conchagua, 31-I- 1998, A. Monro et al. 2153 (BM, MO). San Vicente. cercanía de San Vicente, 7-14-II-1947, P. Standley & E. Padilla 3523 (EAP). Santa Ana. El Pinalito, Santa Ana, 1-IX-1994, R. Villacorta & E. Montalvo 2210 (LAGU, MO).
20. TABERNAEMONTANA L., Sp. Pl. 1: 210. 1753.
Bonafousia A. DC., Prodr. 8: 359. 1844. Peschiera A. DC., Prodr. 8: 360. 1844. Stenosolen (Müll. Arg.) Markgr., Fl. Suriname 4: 455. 1937.
Género pantropical con cerca de 100 especies, 45 de ellas restringidas al neotrópico; 1 sola conocida en El Salvador. (Leeuwenberg, 1994). Para la descripción del género y mayores detalles véase Morales (2005 c: 168).
32. Tabernaemontana amygdalifolia Jacq., Enum. Syst. Pl. 14. 1760. TIPO: Cult., Austria, Wien, Hort. Schönbrunn, Herb. Jacquin s.n. (holotipo, W!).
Distribución y hábitat. México a Colombia, Venezuela, Ecuador, Perú y Bolivia. Habita bosques muy húmedos, bosques estacionalmente secos, en elevaciones de 0-1200 m s.m. Especímenes con flores y frutos han sido colecionados principalmente entre diciembre y mayo. Para la descripción del género y mayores detalles véase Morales (2005c: 170).
Nombres vulgares. "Cojón" (Ahuachapán); "Cojón Amarillo" (Ahuachapán); "Cojón blanco" (La Libertad); "Cojón de blanco" (Ahuachapán); "Cojón de altura" (Ahuachapán); "Cojón de Toro" (Santa Ana); "Cojón verde de costa" (Ahuachapán); "Cojoncito blanco" (Ahuachapán); "Jazmín de monte" (La Libertad).
Observaciones. Es una especie bastante polimórfica en relación al tamaño de las flores y hojas. A través de Mesoamérica, los especímenes de zonas secas tienen flores y hojas mucho más pequeñas con respecto a los provenientes de bosques más humedos. Estudios monográficos posteriores, complementados con un fuerte trabajo de campo, podrían demostrar que se trata de dos entidades distintas.
Material representativo examinado
EL SALVADOR. Ahuachapán. San Francisco Menéndez, El Corozo, Los Peralta, 24-II-2000, J. Rosales 262 (LAGU, MO). Cabañas. Cinquera, 15-III-2002, A. Monro et al. 3740 (BM, LAGU, MO). La Libertad. Cordillera de Balsamó, carretera San Julián, 7-III-2002, A. Monro et al. 3677 (BM, LAGU, MO). San Salvador. Cerca del Lago Ilopango y Apulo, 19-VI-1949 (fl), L. Williams & A. Molina 16754 (EAP). Santa Ana. E de Texistepeque, Piedras Negras, 28-IV-1995, E. Linares & C. Martínez 2680 (EAP, MO). Sonsonate. Santa Emilia, 22-25-III-1922, P. Standley 2213 (MO, US).
21. THEVETIA L., Opera Var. (Syst. Nat. Ed. 8): 212. 1758.
Plumeriopsis Rusby & Woodson, Ann. Missouri Bot. Gard. 24(1): 11. 1937.
Género monotípico distribuido desde México a Colombia y Venezuela. (Alvarado-Cárdenas y Ochotorena, 2005; Gensel, 1969; Morales, 2005 c). Para la descripción del género y mayores detalles véase Morales (2005c: 174).
33. Thevetia ahouai (L.) A. DC., Prodr. 8: 345. 1844. Cerbera ahouai L., Sp. Pl. 1: 208. 1753. Plumeriopsis ahouai (L.) Rusby & Woodson, Ann. Missouri Bot. Gard. 24 (1): 11. 1937. TIPO: Herb. Clifford: 76, Plumeria No. 1, rama, fruto y flor inferior derecha (lectotipo, BM!, designado por Leeuwenberg, Regnum. Veg. 127: 94. 1993).
Cerbera nitida Kunth, Nov. Gen. Sp. 3: 225. 1818 [1819]. Thevetia nitida (Kunth) A. DC., Prodr. 8: 344. 1844, nom. Illeg. TIPO: Colombia. Atlántico: río Sinú, Turbaco, III-año perdido (fl), Bonpland 1371 (holotipo P-HB!, isotipo B no visto, foto F neg. 4482 INB!).
Referencia. Morales (2005c: 175).
Distribución y hábitat. México a Colombia y Venezuela. Habita bosques muy húmedos, en elevaciones de 250-400 m s.m. Especímenes con flores y frutos fueron coleccionados en febrero.
Nombres vulgares. "Cojón de costa" (Ahuachapán).
Observaciones. Thevetia ahouai es un taxón raro en El Salvador, que se puede reconocer con facilidad del resto de especies de Cascabela por su corola hipocraterimorfa, así como por sus frutos abayados y rojo intenso al madurar.
Material representativo examinado
EL SALVADOR. Ahuachapán. San Benito, Monte Hermoso, 21-II-1993, M. Sandoval 1030 (LAGU, MO).
22. VALLESIA Ruiz & Pav., Fl. Peruv. Prodr. 2: 26. 1799.
Género con ca. 9 especies distribuidas desde México y las Antillas a Costa Rica (excepto Nicaragua) y desde Colombia e Islas Galápagos hasta el N de Argentina; una sola especie presente en El Salvador (Meyer, 1955; Morales, 1998b). Para la descripción del género y mayores detalles véase Morales (2005c: 178).
34. Vallesia aurantiaca (M. Martens & Galeotti) J. F. Morales, Novon 8(3): 263. 1998. Neriandra aurantiaca M. Martens & Galeotti, Bull. Acad. Roy. Sci. Bruxelles 11(6): 356. 1844. TIPO: México. Veracruz: Santiago de Huatusco, IV- 1841 (fl), H. Galeotti 1591 (holotipo, BR!).
Vallesia mexicana Müll. Arg., Linnaea 30: 393. 1860. TIPO: México. Veracruz: cerca de Orizaba, II-1854 (fl), M. Botteri 473 (holotipo, G-DC!; isotipo, P!, foto, INB!).
Vallesia flexuosa Woodson, Ann. Missouri Bot. Gard. 24(1): 14. 1937. TIPO: Costa Rica. Alajuela: Zarcero, 21-XII-1924 (fl), Brenes 4156 (holotipo F!, foto F neg. 56523 INB!; isotipos, CR!, P!).
Referencia. Morales (2005c: 178).
Distribución y hábitat. México, Guatemala, Belice, Honduras, El Salvador y Costa Rica. Habita bosques muy húmedos, en elevaciones de 1700- 2100 m s.m. Especímenes con flores fueron coleccionados en abril.
Nombres vulgares. "Mango" (Santa Ana, Montecristo).
Observaciones. Una de las Apocynaceae arborescentes más fáciles de reconocer, principalmente por sus hojas alternas, usualmente dísticas, así como por sus frutos drupáceos, carnosos y blancos al madurar. En el campo, de primera impresión, puede ser confundido con un representante de la familia Annonaceae.
Material representativo examinado
EL SALVADOR. Santa Ana. San José Ingenio, parque
nacional Montecristo, 16-II-2002, V. Martínez 730
(LAGU, MO).
APÉNDICE 1
Lista completa del material examinado, ordenado alfabéticamente por coleccionista. El número entre paréntesis indica la especie a la que corresponde cada ejemplar de acuerdo a la numeración de las especies en este artículo.
Allen 1855 (24), 6840 (29), 6924 (4); Andrieux 245 (12), 247 (21), 251 (24); Aparicio 5 (27), 104 (24), 139 (31).
Beetle 26272 (28); Berendsohn 69 (6), 189 (12), 389 (4), 1502 (30); Bertero s.n. (27); Billberg s.n. (25); Bonpland 1371 (33); Botteri 473 (34); Brenes 4156 (34); Burgos 5 (4).
Calderón 32 (24), 339 (4), 815 (15), 938 (18), 2375 (8); Campos s.n. (27); Carballo s.n. (18), 189 (8), 289 (31), 301 (5); Carlson 314 (10); Case 22, 22 a, 28 (28); Castillo 551 (28), 567 (31), 605, 668 (28); Cavanilles s.n. (33); Chinchilla s.n. (4), s.n. (15), s.n. (24), s.n. (25), s.n. (26), (29), s.n. (30), s.n. (28); Croat 32798 (24), 42119 (15), 42161 (15), 42243 (16), 42279 (15); Cruz 27 (22), 86 (16), 222 (7); Cufodontis 220 (5); Current 44 (8).
Davidse 2038 (29), 2080 (31), 37123 (19), 37486 (25); Deam 6098 (4), 6376 (11); Donnell-Smith 2763, 6646 (29); Doyle 33 (Apitti.); Drege 88 (7); Dunn 36 (28); Dwyer 412 (28).
Escobar s.n., s.n. (15); Estrada 193 (12).
Flores s.n. (10), s.n. (28), 28 (29), 267 (28). Galeotti 1579 (17), 1586 (24), 1591 (34), 1602 (15); Gaumer 1979 (11); González 52 (28), 54 (27), 162 (28), 260 (31), 315 (4), 439 (8), 442 (31), 507 (8); Grupo Ecológico Cantón La Bermuda 3340 (28); Guerra 5 (29).
Harmon 5938 (28); Hayes s.n. (12); Hernández PBT- 561 (11); Heyde 3993 (17), 4497 (26), 4534 (13), 4538 (30); Holm 243 (2); Humboldt s.n., 1480 (27);
Karwinsky 1348 (5); King 30 (31), 34 (31), 39 (8), 40 (28), 80 (31).
Lagos 836 (24); Lara 2164 (28); Lévy 1074 (11); Liebmann 11986 (16); Linares 174 (28), 360 (28), 537 (8), 548 (31), 686 (28), 1321 (32), 1327 (29), 1614 (17), 1953 (3), 2125 (9), 2427 (19), 2470 (32), 2471 (31), 2549 (8), 2551 (31), 2586 (29), 2658 (31), 2680 (32), 2728 (3), 2733 (11), 2760 (8), 2785 (11), 2845 (11), 2866 (11), 2873 (4), 3315 (9), 3629 (12), 3979 (12), 3982 (3); López s.n. (1); s.n. (29); Lundell 1284 (5), 3408 (5).
Mangandí s.n. (4); Martínez s.n. (4), s.n. (25), s.n., s.n. (28), s.n. (34), 39 (9), 64 (30), 238 (4), 730 (8), 1136 (34); Matuda 4240 (16), 15570 (26), 16978 (5); Molina 3640 (12), 21551 (28); Monro 1952 (29), 2111 (4), 2153 (31), 2888 (22), 3658 (29), 3677 (32), 3686 (31), 3740 (32); Montalvo 3385 (4), 4690 (15), 6365 (32), 6370 (28), 6464 (6); Monterrosa 4 (8), 80 (28), 298 (18), 374 (31), 582 (15), 900 (20); Morales 1270 (24); Morton 2692 (5). Nelson 3080 (3).
Owen 1 (25). Pavón s.n. (16); Peck 35 (15), 953 (23); Peñate 1311 (15); Pittier 2012 (25); Purpurs 5408 (12).
Quintana s.n. (7). Renderos 186 (11), 378 (3), 722 (31); Renson 289 (4); Reyna 121 (28); Rosales 36 (29), 46 (31), 47 (32), 53 (5), 64 (28), 118 (25), 262 (32), 275 (29), 293 (22), 295 (25), 296 (31), 420 (32), 446 (28), 455 (13), 507 (22), 511, 515 (19), 544 (3), 556 (25), 558 (29), 634 (28), 635 (31), 675 (28), 696 (14), 702 (3), 765, 766 (24), 767 (31), 785 (3), 791 (29), 824 (12), 843 (21), 885 (24), 1070, 1120 (18), 1176 (28), 1212 (4), 1261 (14), 1312 (5), 1356 (28), 1770 (4), 1872 (32), 1879 (32), 1905 (31), 2083 (29), 2086 (28), 2158 (19), 2372 (29), 2464 (24), 2471 (14), 2475 (27), 2527 (11), 2547 (32), 2679 (12); Ruiz 11/55 (15).
Salazar 20 (27); Sandoval 27 (3), 47 (4), 91 (4), 107 (21), 125 (15), 158 (12), 201 (30), 272 (5), 280 (32), 283 (23), 296 (22), 309 (25), 311 (26), 341 (32), 355 (28), 378 (25), 379 (32), 502 (15), 835 (4), 894 (25), 922 (28), 957 (30), 1013 (25), 1030 (33), 1254 (4), 1259 (24), 1308 (30), 1309 (12), 1317 (31), 1440 (14), 1476 (15), 1542 (28), 1545 (31), 1586 (26), 1647 (11); Schipp S-07 (24), S-388 (25); Sessé 5007 (23); Seemann 95 (18); Sermeño s.n. (4), s.n. (29), 30 (24), 124 (22), 125 (13), 183 (30), 205 (10), 228 (15), 232 (12); Sidwell 583 (29), 713 (4); Sinclair s.n., s.n., s.n. (31), s.n. (8); Skutch 3864 (23); Smeets 63(1), 367 (22); Spellman 747 (28); Standley 3033 (28), 3440 (8), 3523 (31), 19913 (24), 20091 (26), 21864 (19), 22349 (25), 23673 (26), 41332 (24); Steere 1477 (11); Steyermark 47313 (4); Stork 2800 (27).
Thurn 198 (18); Toledo 12 (15), 18 (16); Tuerckheim 8709 (17).
Vasquez s.n. (28); Villacorta 742 (26), 804 (21), 830 (16), 837 (27), 861 (24), 1069 (32), 1176 (27), 1199 (31), 1502 (32), 2050 (31), 2181 (2), 2196 (12), 2210 (31), 2344 (28), 2355 (31), 2385 (19), 2393 (24), 2688 (31), 2891 (32).
Wallich 1629,1 (6); Weberling 843 (24); Webster 12804 (21); Wedel 1353 (24); Williams 13569 (30), 15156 (30), 16754 (32), 16760 (28); Winkler 10 (17); Witsberger 605 (26), 717 (26), 752 (30); Woodson 1180 (24).
APÉNDICE 2
Índice de nombres científicos. Los nombres válidos se indican con negritas; los sinónimos con bastardillas.
Allamanda, 457, 458
Allamanda cathartica, 458
Allotoonia, 458, 461, 462
Allotoonia caudata, 462
Allotoonia tuxtlensis, 461, 462
Alstonia, 457, 461, 462
Alstonia pittieri, 461, 462
Alstonia stenophylla, 463
Amblyanthera pandurata, 470
Amblyanthera tubiflora, 473
Angadenia pandurata, 470
Arduina grandiflora, 464
Arduina macrocarpa, 464
Aspidosperma, 457, 461, 463
Aspidosperma lundellianum, 463
Aspidosperma megalocarpon, 461, 463
Beaumontia, 457, 464
Beaumontia grandiflora, 464
Belandra concolor, 478
Bignonia obovata, 482
Carissa, 457, 464, 465
Carissa grandiflora, 464
Carissa macrocarpa, 464, 465
Cascabela, 457, 465, 466
Cascabela ovata, 459, 460, 465, 466
Cascabela peruviana, 467
Cascabela plumeriifolia, 466
Cascabela thevetia, 460, 465, 467
Catharanthus, 458, 467
Catharanthus roseus, 467
Cerbera ahouai, 484
Cerbera nitida, 484
Cerbera ovata, 466
Cerbera thevetia, 467
Cerbera peruviana, 467
Cufodontia arborea, 463
Cufodontia escuintlensis, 463
Cufodontia lundelliana, 463
Echites, 458, 467
Echites circinnalis, 468
Echites cobanensis, 473
Echites comosus, 474
Echites cupulifer, 475
Echites cuspidifer, 472
Echites elegantulus, 468
Echites jasminiflorus, 472
Echites longifolius, 478
Echites panduratus, 470
Echites portobellensis, 479
Echites puntarenensis, 462
Echites spicatus, 472
Echites subsagittatus, 472
Echites subsessilis, 473
Echites trifidus, 474
Echites tubiflorus, 473
Echites turriger, 462
Echites tuxtlensis, 462
Echites yucatanensis, 462
Fernaldia, 458, 469, 470
Fernaldia brachypharynx, 470
Fernaldia glabra, 470
Fernaldia pandurata, 469, 470
Fernaldia pandurata var. glabra, 470
Forsteronia, 458, 469, 470
Forsteronia myriantha, 469, 470
Forsteronia spicata, 456, 469, 472
Laseguea villosa, 474
Lochnera rosea, 467
Macaglia megalocarpa, 463
Mandevilla, 458, 471, 472
Mandevilla donnell-smithii, 473
Mandevilla hirsuta, 459, 460, 472
Mandevilla mollis, 473
Mandevilla platydactyla, 473
Mandevilla subsagittata, 472
Mandevilla subsessilis, 473
Mandevilla tubiflora, 473
Mandevilla villosa, 474
Mesechites, 458, 474
Mesechites jasminiflorus, 472
Mesechites trifidus, 474
Mitozus mexicanus, 478
Neriandra aurantiaca, 484
Nerium, 457, 475
Nerium oleander, 475
Pentalinon, 458, 471, 475
Pentalinon andrieuxii, 471, 475
Plumeria, 457, 477
Plumeria purpurea, 477
Plumeria rubra, 460, 477
Plumeria tricolor, 477
Plumeriopsis ahouai, 484
Prestonia, 458, 476, 478
Prestonia amanuensis, 478
Prestonia caudata, 462
Prestonia concolor, 478
Prestonia longifolia, 478
Prestonia grandiflora, 479
Prestonia guatemalensis, 479
Prestonia laxa, 479
Prestonia mexicana, 478
Prestonia portobellensis, 459, 479
Prestonia trifida, 459
Prestonia schippii, 479
Prestonia speciosa, 479
Rauvolfia, 457, 476, 480
Rauvolfia hirsuta, 480
Rauvolfia ligustrina, 480
Rauvolfia stenophylla, 463
Rauvolfia ternifolia, 480
Rauvolfia tetraphylla, 456, 480
Rauvolfia tomentosa, 480
Stemmadenia, 457, 476, 481
Stemmadenia donnell-smithii, 481
Stemmadenia eubracteata, 481
Stemmadenia glabra, 483
Stemmadenia mollis, 483
Stemmadenia obovata var. mollis, 483
Stemmadenia obovata, 482
Stemmadenia pubescens, 482
Tabernaemontana, 457, 483
Tabernaemontana amygdalifolia, 483
Tabernaemontana donnell-smithii, 481
Thevetia, 457, 484
Thevetia ahouai, 484
Thevetia nitida, 484
Thevetia ovata, 466
Thevetia plumeriifolia, 466
Thevetia peruviana, 467
Thevetia thevetia, 46
Tonduzia longipedunculata, 463
Tonduzia pittieri, 462
Tonduzia parvifolia, 463
Tonduzia stenophylla, 463
Urechites andrieuxii, 475
Vallesia, 457, 484
Vallesia aurantiaca, 484
Vallesia flexuosa, 484
Vallesia mexicana, 484
Vinca rosea, 467
APÉNDICE 3
Índice de nombres vulgares
Amatillo, 400
Bejuco de Tacuazín, 470
Bejuco lengua de culebra, 462
Bejuco loroco de venado, 479
Bejuco quilite, 470
Bejuco Tacuazín blanco, 472
Campana blanca, 464
Canforillo, 480
Chalchupa, 480
Chilindrón, 463, 466
Chilindrón de montaña, 463
Chula, 467
Cojón, 481, 483
Cojón amarillo, 483
Cojón blanco, 483
Cojón de altura, 482, 483
Cojón de blanco, 483
Cojón de costa, 484
Cojón de costa hoja de hacha, 483
Cojón de costa hojas largas, 466
Cojón de montaña, 482
Cojón de puerco, 481
Cojón de tierra fría, 482
Cojón de toro, 482, 483
Cojón de turco, 481
Cojón picudo, 483
Cojón verde, 483
Cojón verde de costa, 483
Cojoncillo, 483
Cojoncito, 480
Cojoncito blanco, 483
Cojoncito verde de altura, 482
Flor blanca, 468, 477
Flor de ensarta, 477
Flor de mayo, 477
Flor de veleta, 472
Huevos de toro, 483
Jazmín de monte, 483
Loroco, 470
Loroco de culebra, 462
Loroco de venado, 468
Loroco de zope, 467
Loroco montes, 468
Loroquillo, 475
Mango, 484
Molleja de pato, 464
Pacayita, 477
Pacallita macho, 477
Sulfatillo, 463
AGRADECIMIENTOS
Quiero agradecer a los herbarios A, BM, BR, C, CAS, CR, EAP, F, FI, FI-W, G, G-DC, GH, K, LAGU, M, MA, MEXU, MICH, MO, NY, P, P-HB, S, TEX, UC, US, W, WIS, y Z por proveer material en préstamo o por permitir el ingreso a sus colecciones. El mapa de la República de El Salvador es cortesía de la Librería de la Universidad de Texas, Austin, Estados Unidos y fue tomado de su sitio WEB (http://www.lib.utexas.edu/maps/elsalvador.html). Por otro lado, es de gran importancia el reconocer el trabajo realizado por el Herbario de la Asociación Jardín Botánico La Laguna (LAGU), en La Libertad, El Salvador, cuyas colecciones han sido fundamentales por la gran cantidad de nuevos registros para El Salvador, y por el numeroso envío de material como regalo por identificación; en especial quiero resaltar la importante colaboración de Jorge Monterrosa en diferentes aspectos de este trabajo. Se agradece a Jim Manhart (Texas A&M University, Estados Unidos), por autorizar el uso de la fotografía de Carissa macrocarpa, así como al Instituto Nacional de Biodiversidad (INBio) por permitir usar las fotografías de Alstonia pittieri, Mandevilla subsagittata, Prestonia portobellensis, Stemmadenia donnellsmithii, (tomadas por Alexander Rodríguez) y de Aspidosperma megalocarpon y Mesechites trifidus (tomadas por Reinaldo Aguilar). Asimismo, la fotografía de Mandevilla villosa es reproducida con permiso del Missouri Botanical Garden y del autor, Tom Croat. Daniel Atha (NY) autorizó el uso de la imagen de Pentalinon andrieuxii, y Ana Luc McVean (UVAL) la de Prestonia longifolia. Quiero agradecer el apoyo financiero del Missouri Botanical Garden a través del proyecto de la Flora Mesoamericana y en forma especial a Gerrit Davidse, así como a Olga Martha Montiel, quienes hicieron posible en parte la culminación de este trabajo, mediante el apoyo económico para la visita del herbario MO. Otras personas que colaboraron en Honduras para la revisión de material de El Salvador depositado en distintos herbarios fueron Frank Cardoza y José Linares (EAP) y Cirilo Nelson (TEFH). Agradezco la revisión crítica del manuscrito por parte de Mary Endress (Z), así como la colaboración de Michael Grayum (MO) por la redacción del resumen en inglés, así como por la evacuación de dudas sobre la correcta aplicación del latín en las terminaciones de los epítetos de algunas especies de Echites y finalmente, a Walter Berendsohn (B), por algunos comentarios de una versión preliminar de este manuscrito y por llamar mi atención sobre la presencia de Funtumia en algunas zonas del El Salvador.
BIBLIOGRAFÍA
1. Alvarado-Cárdenas, L. & H. Ochoterena. 2005. Phylogenetic análisis of the Cascabela-Thevetia complex (Plumerieae; Apocynaceae) based on morphology. XVII International Botanical Congress, Julio 2005, Vienna, (Austria), Abstracts: 1007. [ Links ]
2. Bergen, M. A. van. 1996. Revision of Catharanthus G. Don. Series of Revisions of Apocynaceae XLI. Agric. Univ. Wageningen Pap. 96: 9-46. [ Links ]
3. Codd, L. E. 1963. Apocynaceae, pp. 244-296, en L. E. Codd & H. B. Rycroft (eds.), Flora of Southern Africa, vol. 26. Pretoria: Dept. Agric. Techn. Serv. [ Links ]
4. Endress, M. E. & V. A. Albert. 1995. A morphological cladistic study of Apocynaceae: Trends in character evolution within a broadened familial circumscription. Amer. J. Bot. 82 (6, Abstracts): 127. [ Links ]
5. Endress, M. E. & P. Bruyns. 2000. A revised classification of the Apocynaceae s.l. Bot. review (Lancaster) 66: 1-56. [ Links ]
6. Ezcurra, C. 1981. Revisión de las Apocináceas de la Argentina. Darwiniana 23: 367-474. [ Links ]
7. Gensel, W. H. 1969. A revision of the genus Thevetia (Apocynaceae). Masters thesis. University of Connecticut. [ Links ]
8. Gentry, A. 2001. Apocynaceae, en W. Stevens, C. Ulloa, A. Pool & O. Montiel (eds.), Flora de Nicaragua. Monogr. Syst. Bot. Missouri Bot. Gard. 85: 116-132. [ Links ]
9. Hansen, B. F. 1985. A monographic revision of Forsteronia (Apocynaceae). Ph.D. diss. University of South Florida. [ Links ]
10. Endress, M. E. & R. P. Wunderlin. 1986. Pentalinon Voigt, an earlier name for Urechites Müll. Arg. (Apocynaceae). Taxon 35: 166-168. [ Links ]
11. Holmgren, P. K., N. H. Holmgren & L. C. Barnett, 1990. Index Herbariorum. Part I: The Herbaria of the World. New York: New York Botanical Garden. [ Links ]
12. Jarvis, C. E., F. R. Barrie, D. M. Allan & J. L. Reveal. 1993. A list of Linnaean generic names and their types. Regnum Veg. 127: 1-100. [ Links ]
13. Leeuwenberg, A. 1994. A revision of Tabernaemontana Two. The New World species and Stemmadenia. Kew: Royal Botanic Gardens. [ Links ]
14. Leeuwenberg, A. & F. J. H. Van Dilst. 2001. Series of revisions of Apocynaceae XLIX, Carissa L. Wageningen University Papers. Leiden: Blackhuys Publishers. [ Links ]
15. Lippold, H. 1980. Die Gattungen Thevetia L., Cerbera L. und Cascabela Rafin. (Apocynaceae). Feddes Repert. 91: 45-55. [ Links ]
16. Marcondes-Ferreira, W. 1988. Aspidosperma Mart., nom. cons. (Apocynaceae): estudos taxonômicos. Ph. D. Tésis, Universidade Estadual de Campinas. [ Links ]
17. Meyer, E. 1955. The genus Vallesia, A systematic study. Master Thesis, Washington University. [ Links ]
18. Morales, J. F. 1995. Evaluación del género Alstonia (Apocynaceae) en América Central. Phytologia 78: 192-194. [ Links ]
19. Morales, J. F. 1996. Novelties in Prestonia (Apocynaceae). Novon 6: 285-287. [ Links ]
20. Morales, J. F. 1997a. A synopsis of the genus Prestonia (Apocynaceae) section Tomentosae in Mesoamerica. Novon 7: 59-66. [ Links ]
21. Morales, J. F. 1997b. A reevaluation of Echites and Prestonia section Coalitae (Apocynaceae). Brittonia 49: 328-336. [ Links ]
22. Morales, J. F. 1998a. A synopsis of the genus Mandevilla (Apocynaceae) in Mexico and Central America. Brittonia 50: 214-233. [ Links ]
23. Morales, J. F. 1998b. Three new species and a new combination in Vallesia (Apocynaceae). Novon 8: 263-264 [ Links ]
24. Morales, J. F. 2002. Studies in Neotropical Apocynaceae II: A revision of the genus Fernaldia. Rhodora 104: 186-200. [ Links ]
25. Morales, J. F. 2005a. Estudios en las Apocynaceae Neotropicales IX: Mandevilla antioquiana sp. nov. y lectotipificación de Echites comosa. Anales Jard. Bot. Madrid 62: 65-68 [ Links ]
26. Morales, J. F. 2005b. Estudios en las Apocynaceae Neotropicales XIV: nuevas lectopificaciones en los géneros Hylaea J. F. Morales y Pentalinon Voigt (Apocynoideae, Echiteae). Lankesteriana 5: 159-160. [ Links ]
27. Morales, J. F. 2005c. Estudios en las Apocynaceae Neotropicales XIX: la familia Apocynaceae (Apocynoideae, Rauvolfioideae) en Costa Rica. Darwiniana 43: 90-191. [ Links ]
28. Morales, J. F. 2006. Estudios en las Apocynaceae Neotropicales XXVI: Una monografía del género Mesechites (Apocynoideae, Mesechiteae. Candollea 61: 215-277. [ Links ]
29. Morales, J. F. & A. Fuentes. 2004. Estudios en las Apocynaceae Neotropicales VIII: nuevas especies de Mandevilla (Apocynoideae, Mesechiteae) para Perú y Bolivia, con notas sobre la morfología floral en corolas infundibuliformes. Candollea 59: 167-174. [ Links ]
30. Morales, J. F. & M. Méndez. 2005. Estudios en las Apocynaceae Neotropicales XXII: Nuevos realineamientos taxonómicos en el género Stemmadenia (Apocynaceae, Rauvolfioideae, Tabernaemontaneae. Candollea 60: 345-371 [ Links ]
31. Morales, J. F. & J. Williams. 2004. Allotoonia, a new neotropical genus of Apocynaceae based on a subgeneric segregate of Echites. Sida 21: 133-158. [ Links ]
32. Morales, J. F. & J. Williams. 2005. Una nueva combinación en el género Allotoonia (Apocynaceae, Apocynoideae, Echiteae). Lankesteriana 5: 119-120. [ Links ]
33. Morton, J., E. Álvarez & C. Quiñónez. 1990. Loroco, Fernaldia pandurata (A. DC.) Woodson (Apocynaceae): A popular edible flower of Central America. Econ. Bot. 44: 301- 310. [ Links ]
34. Nelson, C. 1996. La flora de Honduras en la Biología Centrali- Americana, de Hemsley. Fontqueria 44:53-68. [ Links ]
35. Pagen, F. J. 1987. Oleanders. Nerium L. and the oleander cultivars, en A. J. M Leeuwenberg (ed.), Series of Revisions of Apocynaceae, part XX. Agric. Univ. Wageningen Pap. 87: 1-113. [ Links ]
36. Potgieter, K. 1999. Phylogenetic study of Apocynaceae Juss. and Aspidosperma Mart. & Zucc. Ph. D. diss., University of Illinois at Urbana-Champaign. [ Links ]
37. Rao, A S. 1956. A revision of Rauvolfia with particular reference to the American species. Ann. Missouri Bot. Gard. 43: 253-355. [ Links ]
38. Rudjiman. 1986. A revision de Beaumontia Wallich, Kibatalia G. Don y Vallariopsis Woodson (Apocynaceae). en A. J. M Leeuwenberg (ed.), Series de Revisions de Apocynaceae, Part XIX. Agric. Univ. Wageningen Pap. 85: 1-99. [ Links ]
39. Sakane, M. & G. J. Sheperd, 1987. Uma revisão do gênero Allamanda L. (Apocynaceae). Rev. Brasil. Bot. 9: 125-149. [ Links ]
40. Sennblad, B. & B. Bremer. 1996. The familial and subfamilial relationships of Apocynaceae and Asclepiadaceae evaluated with rbcL data. Pl. Syst. Evol. 202: 153-175. [ Links ]
41. Sidiyasa, K. 1998. Taxonomy, phylogeny and wood anatomy of Alstonia (Apocynaceae). Blumea, Suppl. 11: 1-230. [ Links ]
42. Simões, A. O. & L. S. Kinoshita. 2002. The Apocynaceae s. str. of the Carrancas region, Minas Gerais, Brazil. Darwiniana 40: 127-169. [ Links ]
43. Stearn, W. T. 1957. An introduction to the 'Species Plantarum' and cognate botanical works of Carl Linnaeus (Prefixed to Ray Society facsimile of Linnaeus, Species Plantarum, vol. 1.) London. [ Links ]
44. Stearn, W. T. 1978. Nerium L., en P. Davis (ed.), Flora of Turkey and the East Aegean Islands, vol. 6, p. 159. Edinburgh: Edinburgh University Press. [ Links ]
45. Wijnands, D. O. 1983. The Botany of the Commelins. Rotterdam: A.A. Balkema. [ Links ]
46. Williams, J. 2002. A further evaluation of Echites sect. Yucatanense (Apocynaceae) with additional notes on the genus. Brittonia 54: 310-317. [ Links ]
47. Woodson, R. E. 1928. Studies in the Apocynaceae II. Arevision of the genus Stemmadenia. Ann. Missouri Bot. Gard. 15: 341-379. [ Links ]
48. Woodson, R. E. 1933. Studies en the Apocynaceae IV. The American genera of Echitoideae XXVI. Ann. Missouri Bot. Gard. 20: 605-790. [ Links ]
49. Woodson, R. E.. 1936. Studies in the Apocynaceae. IV. The American genera of Echitoideae. Ann. Missouri Bot. Gard. 23: 169-438. [ Links ]
50. Woodson, R. E. 1938a. (Asclepiadales) Apocynaceae, en J. H. Barnhart (ed.), North American flora 29(2): 179-185. New York: New York Botanical Garden. [ Links ]
51. Woodson, R. E. 1938b. An evaluation of the genera Plumeria L. and Himatanthus Willd. Ann. Missouri Bot. Gard. 25: 189-224. [ Links ]

