










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkDarwiniana, nueva serie
versão impressa ISSN 0011-6793versão On-line ISSN 1850-1699
Darwiniana v.45 n.2 San Isidro ago./dez. 2007
Análisis de caractereres macroscópicos y microscópicos de Phaseolus Vulgaris (fabaceae, faboideae) silvestres y cultivados del noroeste argentino: una aplicación en arqueobotánica
María del Pilar Babot1, Nurit Oliszewski2 & Alfredo Grau3
1Instituto Superior de Estudios Sociales, Consejo Nacional de Investigaciones Científicas y Técnicas - Instituto de Arqueología y Museo, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán. San Martín 1545, 4000 San Miguel de Tucumán, Tucumán, Argentina; shypb@arnet.com.ar (autor corresponsal).
2Instituto Superior de Estudios Sociales, Consejo Nacional de Investigaciones Científicas y Técnicas - Instituto Interdisciplinario de Estudios Andinos, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán. Miguel Lillo 205, 4000 San Miguel de Tucumán, Tucumán, Argentina.
3Laboratorio de Investigaciones Ecológicas de las Yungas, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán, Centro Universitario Horco Molle, casilla de correo 34, 4107 Yerba Buena, Tucumán, Argentina.
Original recibido el 12 de enero de 2007;
aceptado el 14 de noviembre de 2007
Resumen. Babot, M. del P.; N. Oliszewski & A. Grau. 2007. Análisis de caractereres macroscópicos y microscópicos de Phaseolus vulgaris (Fabaceae, Faboideae) silvestres y cultivados del noroeste argentino. Una aplicación en arqueobotánica.
Los criterios tradicionales utilizados en las descripciones de Phaseolus vulgaris no han facilitado la identificación taxonómica de ejemplares arqueológicos de esta especie y sus variedades, debido a que se centran mayormente en aspectos de la planta difícilmente preservados. En este trabajo se establecen variaciones de utilidad arqueobotánica en un conjunto amplio de caracteres cualitativos y cuantitativos, macroscópicos (atributos de la semilla) y microscópicos (atributos de los granos de almidón) de las variedades cultivada y silvestre en el Noroeste argentino, identificando aquellos caracteres diagnósticos específicos e intraespecíficos.
Palabras clave. Arqueobotánica; Almidón; Morfología; Argentina; Phaseolus;"Poroto común".
Abstract. Babot, M. del P.; N. Oliszewski & A. Grau. 2007. Analysis of macroscopic and microscopic characters of wild and cultivated varieties of Phaseolus vulgaris (Fabaceae, Faboideae) from northwestern Argentina. An archaeobotanical application.
Current criteria used for the characterization of Phaseolus vulgaris and its varieties have not promoted the taxonomic identification of archaeological specimens, because they focused on rarely preserved characteres. This work analyzes variations within a broader set of cualitative and cuantitative characters of Phaseolus vulgaris, both macroscopic (seed) and microscopic (starch grain), with archaeobotanical relevance for northwestern Argentina. Diagnostic characters for Phaseolus vulgaris and the cultivated and wild varieties were identified.
Keywords. Archaeobotany; "Common bean"; Morphology; Argentina; Phaseolus; Starch.
INTRODUCCIÓN
Ubicación taxonómica de Phaseolus vulgaris silvestre y cultivado
Cinco de las 55 especies del género Phaseolus L. (Phaseolae, Faboideae, Fabaceae; Lackey, 1983), han sido domesticadas de manera independiente a partir de distintas especies silvestres americanas. Estas son P. vulgaris L., P. lunatus L., P. polyanthus Greenm, P. acutifolius A. Gray y P. coccineus L. Las dos primeras han sido objeto de múltiples domesticaciones ocurridas en los Andes meridionales y México-Centroamérica; la tercera y la menos influenciada por la domesticación, se halla en México, Mesoamérica, el Caribe y América del Sur (hasta el norte de Perú); y las dos últimas corresponden a México, el SO de Estados Unidos y Centroamérica (Debouck, 1994; Kaplan, 1965).
Toda la evidencia de carácter arqueológico, morfológico, químico y molecular disponible hasta el momento, sugiere que Phaseolus vulgaris var. vulgaris L., conocido vulgarmente como "Poroto común" en la región andina y sur de Sudamérica, fue seleccionada en múltiples centros de domesticación tanto en Mesoamérica como en el área andina sudamericana, a partir de diversas variedades del "Poroto común" silvestre, como lo es Phaseolus vulgaris var. aborigineus (Burkart) Baudet en el Noroeste argentino (Singh et al., 1991; Zalocchi et al., 1995). En efecto, la mayoría de los genotipos de poroto común cultivado pueden ser separados o identificados como pertenecientes a dos "pools" genéticos, uno distribuido en los Andes meridionales (sur de Perú, Bolivia y Argentina) y otro en México, América Central y Colombia, cuya diferenciación precede a la domesticación (Johnson & Gepts, 1999; Kami et al., 1995). A su vez, en Ecuador y el norte de Perú se han hallado poblaciones de poroto común silvestre que serían ancestrales e intermedias entre los dos acervos genéticos antes mencionados (Kami et al., 1995). La separación entre los "pools" principales es tal que sus cruzas generan progenies con anormalidades fenotípicas y productividad reducida, en el marco de un incipiente proceso de especiación (Johnson & Gepts, 1999).
La existencia de una variedad silvestre de Phaseolus vulgaris en el Noroeste argentino es mencionada por primera vez por Arturo Burkart en 1941, durante la I Reunión Argentina de Agronomía. Esto es ratificado en 1943, cuando el autor sostiene la presencia de porotos absolutamente espontáneos en la selva tucumano-boliviana del Noroeste argentino (Berglund-Brücher & Brücher, 1976). En 1952, Burkart publica la descripción de Phaseolus aborigineus Burkart, proponiéndolo como el ancestro silvestre del "Poroto común" cultivado y dándole el estatus de especie (Burkart, 1952). Posteriormente, el taxón es ubicando en el rango de subespecie para ser finalmente incluido como una variedad de P. vulgaris (Baudet, 1977). Actualmente, se considera que el "Poroto común" (P. vulgaris) presenta dos variedades en el Noroeste argentino: una silvestre [P. vulgaris Linnaeus var. aborigineus (Burkart) Baudet] y otra cultivada (P. vulgaris var. vulgaris) (Baudet, 1977; Zalocchi et al., 1995; Zuloaga & Morrone, 1999).
La distribución de Phaseolus vulgaris var. aborigineus en Sudamérica describe un arco que se extiende a lo largo de 5000 km entre los 1000 y 2800 m s.m., coincidiendo con la asociación Alnus-Sambucus-Juglans-Celtis de la ladera este de los Andes (Berglund-Brücher & Brücher, 1976). Esta angosta faja de selva mesotérmica constituye su hábitat natural, caracterizado por el clima montañoso de la zona subtropical, con elevadas temperaturas diurnas y noches frías. En Argentina es común en las provincias de Jujuy, Salta y Tucumán y ocurre, con menor frecuencia, en Catamarca, Córdoba y San Luis, siendo particularmente abundante en los valles intermontanos de las laderas orientales del sistema del Aconquija que alcanza alturas máximas superiores a 5000 m s.m. (Berglund-Brücher & Brücher, 1976).
Phaseolus vulgaris var. vulgaris es un cultivo mesotérmico típico de los valles y faldas templadas sudamericanas (Parodi, 1991). Crece comúnmente entre los 0 y 2000 m s.m., aunque en algunas zonas se siembra hasta los 3500 m s.m. (Tapia, 1990).
Características de las semillas de Phaseolus vulgaris var. aborigineus y P. vulgaris var. vulgaris
La variedad aborigineus puede ser fácilmente reconocida por sus numerosas semillas chatas y a veces angulares. El tamaño varía según las condiciones ecológicas pero, en promedio, sus medidas son: 5-10 mm de longitud, 3,5-7 mm de latitud y 3-4,5 mm de grosor, aunque embebidas en agua llegan a tener un tamaño considerable, de hasta 12 x 8 mm. El peso de 100 semillas es de aproximadamente 3,5 g; poseen duros tegumentos seminales que previenen un empapamiento demasiado rápido, dilatando la germinación. Las semillas son muy variables en cuanto a forma, color y patrón, siendo más comunes los patrones moteados en gris o marrón, aunque también se dan amarillos, pardos y negros (Berglund-Brücher & Brücher, 1976; Gentry, 1969; Gepts, 2006-2007).
Las semillas de Phaseolus vulgaris var. vulgaris difieren de las de P. vulgaris var. aborigineus en que son más permeables al agua, con la aparición de una gran diversidad de tamaños, formas, colores y patrones. La dormición, desfavorable para el cultivo, no se registra en P. vulgaris var. vulgaris en la cual se han eliminado los tegumentos seminales duros. Las semillas de esta variedad se encuentran en el interior de frutos grandes y carnosos, que presentan una mejora en el gusto, con respecto a los de la variedad silvestre y en los cuales se han eliminado las sustancias tóxicas (Berglund-Brücher & Brücher, 1976; Gentry, 1969).
Aunque ocurren algunas excepciones, ambas formas son autocompatibles y diploides con 2n = 22 cromosomas; hibridizan con facilidad produciendo individuos viables y fértiles (Singh et al., 1991).
La magnitud de los cambios registrados en los atributos de las semillas de las variedades cultivadas y su importancia como parte útil para consumo humano en estado maduro, permiten sostener que éstas constituyeron el objetivo de la selección antrópica a partir de las variedades silvestres (Berglund- Brücher & Brücher, 1976).
Identificación taxonómica de especímenes arqueológicos de Phaseolus vulgaris
El registro arqueológico de Phaseolus vulgaris en sitios sudamericanos es recurrente y relativamente sincrónico con los hallazgos mesoamericanos, en concordancia con la hipótesis de domesticaciones independientes. Los datos más tempranos de P. vulgaris doméstico proceden del altiplano de Perú (Cueva del Guitarrero), con cronologías ca. 10800, 6550 y 5780 años a. C., de las cuales, la más temprana ha sido cuestionada en el ámbito arqueológico (Chevalier, 1999). Los hallazgos que les siguen a éstos en antigüedad en la puna y costa peruanas, corresponden al lapso 3400-2100 años a. C. Dentro de los actuales territorios de Ecuador, Chile, Bolivia, Uruguay y Argentina las evidencias son más tardías (Pearsall, 1994; Chevalier, 1999). En Argentina, tanto Phaseolus vulgaris var. aborigineus (Burkart) Baudet como P. vulgaris var. vulgaris están registradas ya en contextos relacionados con agricultura temprana, desde ca. 2600 años a. C. en adelante (Albeck, 2000; Babot, 2004; Carrizo et al., 1999, 2003; Korstanje & Würschmidt, 1999; Lagiglia, 2001; Oliszewski, 2004a; 2005; Pochettino & Scattolin, 1991; Tarragó, 1980). Todos estos hallazgos de P. vulgaris han sido datados mediante fechados radiocarbónicos efectuados sobre materiales asociados estratigráficamente. Las únicas dataciones directas por AMS sobre P. vulgaris var. vulgaris disponibles en la actualidad proveen evidencias de su cultivo recién a partir de los ca. 2400 años a.C. para los Andes peruanos. Esta diferencia en los fechados de semillas de poroto común mediante AMS y los que se les atribuyen a otros ejemplares a partir de la cronología de sus asociaciones arqueológicas, plantea la necesidad de ampliar el conjunto de muestras arqueobotánicas datadas, a efectos de corroborar su antigüedad y establecer una fecha certera para los inicios de la agricultura precolombina del "Poroto común" (Kaplan & Lynch, 1999).
Con excepción de los macrorrestos botánicos recuperados en zonas desérticas, usualmente, la identificación taxonómica de ejemplares arqueológicos de la especie no ha resultado una tarea sencilla debido a su estado de conservación malo y fragmentario, con una alta representación de especímenes carbonizados. Por el mismo motivo, ha sido aún más difícil diferenciar entre Phaseolus vulgaris var. vulgaris y P. vulgaris var. aborigineus. Ambos constituyen aspectos que deben ser resueltos, teniendo en cuenta, por un lado, la frecuencia con que la especie es hallada y, por otro lado, su vinculación con la cuestión de la agricultura temprana en el área Centro - Sur Andina. En este sentido, Piperno & Holst (1998) han llamado la atención respecto de la necesidad de producir más información macro y micromorfológica para evaluar el grado en el que las especies pueden ser identificadas en el registro arqueológico, particularmente aquellas domesticadas que tienen parientes silvestres cercanos, como sería, en nuestra región, el caso de P. vulgaris var. vulgaris y su ancestro silvestre (Burkart, 1952).
En el caso de las dos variedades de Phaseolus vulgaris que se analizan en este trabajo, las caracterizaciones macroscópicas se han centrado, mayormente, en aspectos de la planta o la semilla difícilmente observables en los especímenes arqueológicos, a menos que éstos se encuentren deshidratados y en excelente estado de preservación. Los caracteres externos del tegumento no se conservan en ejemplares carbonizados, por lo tanto, en estos casos las únicas herramientas con las que se cuenta son la forma y las medidas de latitud, longitud y grosor de las semillas completas o sus cotiledones, las cuales no permiten ir más allá de la especie y en muchos casos sólo se puede llegar a determinar género (Oliszewski, 2005).
A nivel microscópico, varios trabajos se han referido a los atributos de los granos de almidón de Phaseolus vulgaris var. vulgaris (Korstanje & Babot 2007; Piperno & Holst, 1998; Reichert, 1913; Yánez-Farías et al., 1997, entre otros), pero no se han efectuado comparaciones con los de su ancestro silvestre. El antecedente más cercano lo constituye el trabajo de Pochettino & Scattolin (1991), en donde se realizan identificaciones a nivel intraespecífico de especimenes arqueológicos carbonizados de ambas variedades a partir del análisis morfológico general de los granos de almidón y de sus improntas en la matriz celular.
Debido a que en los contextos arqueológicos, las semillas o bien sus restos microscópicos, nos brindan las únicas evidencias sobre la utilización de Phaseolus vulgaris, se hace necesario afinar su caracterización, tanto a nivel externo como interno para aportar a la diferenciación de variedades. Teniendo en cuenta esta situación, este trabajo se propone realizar un aporte a la determinación intraespecífica de especímenes arqueológicos, a partir del análisis comparativo de un conjunto amplio de atributos cuali-cuantitativos de las semillas que han sido relevados a nivel macro- y microscópico en muestras actuales. De manera complementaria, se evalúa la incidencia de las diferencias ambientales en los caracteres diagnósticos en semillas de distinta proveniencia.
MATERIALES Y MÉTODOS
Procedencia de las muestras actuales
Se analizaron siete muestras actuales de semillas de Phaseolus vulgaris var. aborigineus y dos muestras de P. vulgaris var. vulgaris. Debe aclararse que la variación fenotípica de P. vulgaris var. vulgaris es mucho mayor a la representada por las muestras que han sido introducidas a modo de ejemplo en esta investigación a fin de compararlas con P. vulgaris var. aborigineus. Esta última variedad está mejor representada en términos de su variación fenotípica y hábitat. Las muestras proceden de diversas localidades del Noroeste argentino situadas en distintos pisos dentro del ámbito de las Yungas (Brown et al., 2001) (Tabla 1, Fig. 1A-I) y fueron coleccionadas entre abril y julio de 1990 en estado maduro; en el momento de su análisis, se encontraban naturalmente deshidratadas, aunque bien conservadas y sin germinar. El material no usado en los estudios ha sido depositado en el herbario LIL.
Tabla 1. Características de las muestras actuales de Phaseolus vulgaris analizadas. Abreviaturas: M, muestra.


Fig. 1. Ejemplares de las muestras actuales de Phaseolus vulgaris analizadas. A-G, P. vulgaris var. aborigineus. H-I, P. vulgaris var. vulgaris. A-G, Grau 911s-913s, 931s, 921s-923s, respectivamente; H-I, Grau 924s, 941s.
Análisis de caracteres
Caracteres macroscópicos. La descripción macroscópica de los especímenes se realizó a ojo desnudo y con microscopio estereoscópico Arcano (2x a 4x), teniendo en cuenta tanto caracteres cuantitativos como cualitativos, los cuales se reseñan a continuación (Burkart, 1952; Oliszewski, 2004b; Singh et al., 1991):
- Longitud, latitud y grosor (mm). Se utilizó un calibre HL (150 x 0,02 mm)
- Relación longitud/latitud
- Peso (g). Se utilizó una balanza digital SAB 61 (200 x 0,1 g). Se pesó cada muestra (10 especimenes) calculándose luego el peso para cada individuo.
- Forma. Se obtuvo a partir de la combinación de longitud, latitud y grosor. La forma puede ser esférica, ovoide, romboédrica, reniforme o cilíndrica.
- Color. Se determinó a ojo desnudo utilizando la Carta de Colores de Suelos de Munsell (Munsell Soil Color Charts, 2000)
- Textura y superficie. La textura puede ser por un lado, lisa o rugosa y por otro lado, lustrosa u opaca. La superficie puede ser mayormente uniforme o no uniforme.
- Anatomía externa del tegumento. Presencia o ausencia de una serie de características que son diagnósticas para la especie: hilo seminal, arilo, micrópila, rafe, carúncula, chalaza y radícula (Burkart, 1952)
Caracteres microscópicos. A nivel microscópico, el análisis se centró en las características de los conjuntos de granos de almidón que son apreciables mediante luz transmitida y a medianos aumentos (200x a 500x). Tomadas en conjunto, estas características poseen utilidad reconocida en Arqueología para la asignación de ciertos taxones (cf. Torrence & Barton, 2006). Para la obtención de las muestras se pelaron las semillas y se rasparon los cotiledones suavemente con el filo de un cortante. Se utilizó este procedimiento para liberar del tejido vegetal una buena cantidad de micropartículas no dañadas y en estado completo, con el fin de facilitar la observación clara de sus atributos (Cortella & Pochettino, 1994, 1995). Se efectuaron preparados semi-permanentes que permitieran la rotación de los granos en el medio de montaje para su observación completa mediante un microscopio petrográfico Zeiss-Axioskop, utilizando luz normal y polarizada. En todos los casos se examinaron atributos cualitativos y cuantitativos típicos de los granos de almidón (Babot, 2004, 2007):
- Largo y ancho (µm)
- Relación largo/ancho
- Multiplicidad: simple, múltiple (doble, triple, etc.); visibilidad y equivalencia en tamaño y forma de cada gránulo (modificado de Buttrose, 1960). Se utilizan las formas españolas gránulo/gránulos de los términos latinos "granulum"/"granula" para hacer referencia a cada constituyente individual de un grano de almidón compuesto ("sensu" Buttrose, 1960). Se emplea grano de almidón como equivalente de la forma inglesa "starch granule" para diferenciar a la estructura completa de sus partes componentes.
- Forma: se refiere a un aspecto particular de la morfología de un grano de almidón, constituido por la silueta que queda definida a la altura de su largo y ancho máximos cuando el grano es apreciado en una vista en planta. Se trata de formas planas que pueden tomar los siguientes valores: oval, plano-convexa, circular, triangular de vértices redondeados, reniforme, ovoide, elíptica, amigdaloide, piriforme, cordiforme, lobulada irregular, irregular, poligonal.
- Hilo del grano de almidón: presencia/ausencia, visibilidad y forma.
- Anillos de crecimiento: presencia/ausencia observable y grado de visibilidad.
- Cruz de extinción: forma, número de brazos o ramas visibles, apariencia y regularidad de los brazos.
- Visibilidad del contorno.
- Presencia de fisuras.
A su vez se estimaron los valores poblacionales de algunas variables:
- Largo y ancho máximo y mínimo.
- Largo/ancho máximo y mínimo.
- Media del largo, Media del ancho y Media de la relación largo/ancho.
- Moda del largo, Moda del ancho y Moda de la relación largo/ancho.
- Distribución de las formas.
- Relación número de granos simples/número de granos múltiples.
RESULTADOS
Caracteres macroscópicos de las muestras analizadas
Los caracteres macroscópicos de las muestras analizadas se resumen en la Tabla 2. En términos generales, todas las muestras de Phaseolus vulgaris analizadas comparten varios atributos: forma reniforme; color rojo oscuro; textura lisa y lustrosa; presencia de hilo, arilo y carúncula. En cuanto a la predominancia del color rojo oscuro, éste se debe al reducido tamaño de la muestra que consideramos en el trabajo, la cual se encuentra acotada geográficamente al Noroeste argentino; como se ha mencionado anteriormente, las semillas de Phaseolus vulgaris son muy variables en cuanto a forma, color y patrón (Berglund-Brücher & Brücher, 1976; Gentry, 1969; Gepts, 2006-2007). En ambas variedades, la relación longitud/latitud se presenta de manera uniforme ya que varía entre 1,3 y 1,5 no evidenciando diferencias significativas a excepción de la muestra H cuyo índice indica que el largo prácticamente duplica al ancho (1,92).
Tabla 2. Características macroscópicas de las muestras de semillas de Phaseolus vulgaris analizadas. Abreviaturas: M, muestra; hi, hilo; ar, arilo; mi, micrópila; ra, rafe; ca, carúncula; ch, chalaza; ra, radícula; +, indica presencia del carácter aludido; -, indica ausencia del carácter aludido; ±, el carácter aludido puede estar presente o ausente. a, corresponde al grupo de semillas pequeñas (< 25 g) establecido por Voysest (1983) para variedades cultivadas (Singh et al. 1991). b, corresponde al grupo de semillas medianas (25 - 40 g) establecido por Voysest (1983) para variedades cultivadas (Singh et al. 1991).

Tomando en cuenta las variables analizadas en conjunto se pueden agrupar las muestras, en dos clases, de la siguiente manera:
Variedades cultivadas. Se caracterizan por presentar grandes dimensiones: 10-13 x 7 x 5,5 mm promedio. Su peso (entre 0,2 y 0,3 g) es superior al de los especímenes silvestres. En cuanto a la forma, la muestra I se diferencia del resto ya que sus especímenes son ovoides. El color también diferencia a las variedades cultivadas de las silvestres, especialmente en el caso de la muestra I que es amarillo pálido. En cuanto a los caracteres anatómicos, la presencia clara de la micrópila permite diferenciarlas de sus parientes silvestres (Fig. 2A).

Fig. 2. Aspecto macroscópico de los ejemplares de Phaseolus vulgaris analizados. A, P. vulgaris var. vulgaris. B, P.vulgaris var. aborigineus. A, Grau 924s; B, Grau 911s.
Variedades silvestres. Se observan claramente tres grupos de muestras: uno formado por las muestras a, b y c que proceden de la zona de San Pedro de Colalao (Dpto. Trancas, Tucumán), un segundo grupo formado por las muestras e, f y g procedentes de Las Estancias (Dpto. Andalgalá,
Catamarca) y un tercer grupo formado por la muestra d, procedente de Tafí del Valle (Dpto. Tafí del Valle, Tucumán). Todos los grupos pueden adscribirse a Phaseolus vulgaris var. aborigineus pero hay ciertas variaciones en los caracteres cuantitativos que determinan su separación. En general son especímenes pequeños (6,7-8,5 mm x 4,7-6 mm x 2,2-3,1 mm). Su peso varía entre 0,04-0,05 g para las muestras de San Pedro de Colalao, pasando por 0,08-0,09 g para las muestras de Las Estancias, hasta 0,11 g para la muestra de Tafí del Valle. Las variables cualitativas no presentan diferencias importantes (Fig. 2B).
Caracteres microscópicos de las muestras analizadas
Los caracteres de los granos de almidón de las muestras analizadas se presentan en la Tabla 3.
Tabla 3. Características de los granos de almidón de las muestras de semillas de Phaseolus vulgaris analizadas. a Representadas por más de tres granos de almidón. Abreviaturas: M, muestra.

El almidón de Phaseolus vulgaris presenta un conjunto de formas simples y compuestas dominantes, oval, ovoide y circular con, al menos, un eje de simetría, las cuales aparecen asociadas en proporciones variables. A éstas se suman las formas lobulada irregular, piriforme y reniforme, típicas de algunos granos múltiples de la especie y, complementariamente y con menor representatividad, las formas poligonal, elíptica, irregular, triangular de vértices redondeados, amigdaloide y cordiforme. Entre esas formas, las ovales y reniformes son diagnósticas, como ocurre en otras especies cuyo fruto es una legumbre, mientras que los granos circulares, en el otro extremo, carecen de este atributo por su ubicuidad en diferentes taxones.
Ambas variedades se distinguen entre s í por la representación diferencial de algunas formas, y por atributos cuantitativos referidos al ancho y la relación largo/ancho de los individuos. La relación entre el número de granos simples y múltiples es variable, siendo estos últimos predominantemente dobles, aunque pueden aparecer como triples o supernumerarios. Los rangos de tamaños observados son amplios y variables; el largo máximo de cada grano individual se ubica entre los 2,5 µm y 52 µm, con una media de alrededor de 20 µm, y una distribución bi- o tri-modal, aunque se han documentado mayores dimensiones para los granos de almidón de la especie, superando los 60 µm (cf. Korstanje & Babot, 2007).
Se observan, adem ás, otros caracteres diagnósticos como la presencia de un contorno nítido y destacado en todos los casos. Como ocurre en la mayoría de los granos múltiples, una cubierta externa impide ver nítidamente los límites de los gránulos contenidos cuando éstos están bien hidratados ("sensu" French, 1984), sin embargo la presencia de cada uno de ellos se aprecia como un lóbulo dentro del grano compuesto que describe puntos de inflexión en el contorno general. Los gránulos no son equivalentes en forma y tamaño y están diferencialmente orientados, presentando un hilo y una cruz de extinción individual.
La cruz de extinci ón es centrada, posee ramas o brazos nítidamente visibles, irregulares y/o fibrosos que se cortan en un punto o línea recta, irregular o entrelazada. Los brazos observables en un grano completo pueden ser supernumerarios, esto último generalmente en el caso de los granos múltiples, siendo no todos visibles y, a veces, compartidos por más de un gránulo. Se observan algunos granos simples y múltiples con sólo dos brazos largos tangenciales que no se intersectan, formados por dos o más brazos individuales. Estos efectos se deben a que los granos de almidón compuestos pueden presentar, tanto una cruz por cada gránulo que contienen, o bien, una cruz que es propia del conjunto ("sensu" French, 1984) y que de esta manera, resulta en un patrón complejo.
El hilo del grano de almid ón, los anillos de crecimiento y una típica fisura central que se presenta como una línea irregular o ramificada, se muestran leve o nítidamente visibles, con mayor frecuencia e intensidad en la variedad cultivada que en la silvestre. Estas características han sido descritas precedentemente como típicas de varias legumbres y, entre ellas, de distintas especies domésticas de Phaseolus (Korstanje & Babot, 2007; Piperno & Holst, 1998; Reichert, 1913; Yánez-Farías et al., 1997, entre otros). Su mayor o menor observación depende en cierta medida de las técnicas de extracción, grado de hidratación, edad de la semilla, inicio de la germinación y otras variables que pueden afectar la estructura cristalina del grano de almidón. En el caso de las muestras analizadas es posible descartar que la presencia de fisura central en los granos de almidón se deba a la utilización de un método de extracción agresivo o a la germinación de las semillas de las que éstos proceden.
Los granos de almid ón simples y múltiples de tamaño pequeño o mediano, pueden aparecer conformando, en número variable, agregados arracimados empaquetados en una única cobertura exterior transparente o translúcida que no es rígida (granos semicompuestos sensu French, 1984) (Fig. 3).
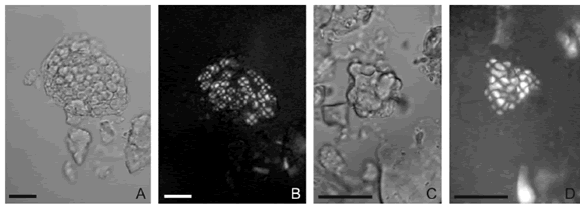
Fig. 3. Aspecto de los agregados arracimados de granos de almidón de Phaseolus vulgaris var. vulgaris. A y C, vistas con luz normal. B y D, vistas con luz polarizada. Escala: 20 µm. A-D, Grau 924s.
Ahora bien, sobre la base de las variaciones en los atributos descritos, las muestras pueden agruparse en dos clases.
Variedades cultivadas. Los conjuntos de granos de almidón se caracterizan por la apariencia general de homogeneidad, regularidad y simetría del conjunto de formas. A las morfologías dominantes se puede sumar la denominada piriforme y, a las minoritarias, la plano-convexa. Más del 75% de los granos tiene una relación largo/ancho entre 1 y 1,42-1,63, salvo en el caso de los granos múltiples de forma lobulada irregular. Tienden a ser poco alargados, poco más largos que anchos y levemente inequiaxiales, lo que está atestiguado por la Moda largo/ancho que se ubica entre 1,11 y 1,28. Este valor puede modificarse cuando abundan los granos supernumerarios lobulados irregulares que son alargados. La Media del largo/ancho no alcanza 1,30; la Moda del ancho supera los 14 µm y la Media supera los 16 µm, mientras que el ancho máximo puede superar los 30 µm. Estos atributos cuantitativos contribuyen a la apariencia general de simetría de las muestras. El hilo de los granos de almidón y los anillos de crecimiento se presentan de leve a nítidamente visibles, a veces o frecuentemente, de manera respectiva (Fig. 4A-B).
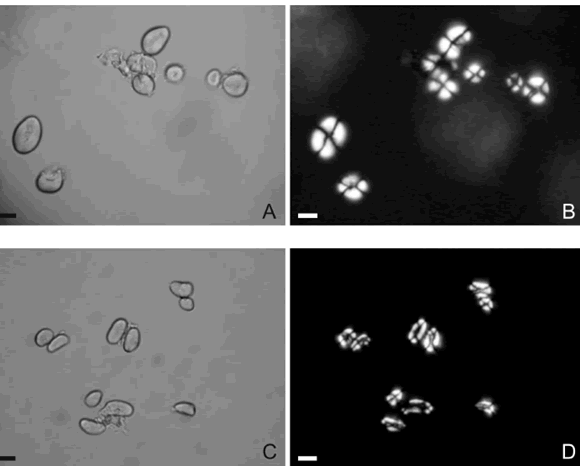
Fig. 4. Aspecto de los conjuntos de granos de almidón de poroto común. A y B, Phaseolus vulgaris var. vulgaris. C y D, P. vulgaris var. aborigineus. Vistas con luz normal a la izquierda, y polarizada a la derecha. Escala: 20 µm. A-B, Grau 924s; C-D, Grau 911s.
Variedades silvestres. Los conjuntos de granos de almidón se caracterizan por la apariencia general de cierta heterogeneidad, irregularidad y asimetría del conjunto de formas. A las morfologías dominantes se puede sumar la plano-convexa y, a las minoritarias, la piriforme; eventualmente pueden ocurrir granos amigdaloides. Más del 75 % de los granos tiene una relación largo/ancho de entre > 1 y 1,84; tienden a ser alargados, más largos que anchos y destacadamente inequiaxiales, lo que está atestiguado por la Moda largo/ancho que se ubica entre 1,15 y 1,55. La Media del largo/ancho supera 1,40; la Moda del ancho es inferior a los 14 µm y la Media del ancho no alcanza los 15 µm, mientras que el ancho máximo no supera los 28 µm. Estos atributos cuantitativos contribuyen a la apariencia general de asimetría de las muestras. El hilo de los granos de almidón y los anillos de crecimiento se presentan levemente visibles, sólo a veces (Fig. 4C-D).
DISCUSIÓN
En términos generales las semillas de Phaseolus vulgaris analizadas, incluyendo tanto a las variedades silvestre como cultivada, comparten varios atributos. A nivel macroscópico, éstos son la forma reniforme; textura lisa y lustrosa; presencia de hilo seminal, arilo y carúncula. A nivel microscópico ambas variedades presentan granos de almidón simples y múltiples (compuestos y semi-compuestos) con formas mayoritariamente simétricas, de contornos nítidos y marcados, que comparten las características de la cruz de extinción, las tendencias de valores de máxima longitud y la eventual aparición de una fisura central. Algunos rasgos microscópicos como la tendencia formal, la fisura central y la medida promedio de los granos han sido observados también en Phaseolus vulgaris cultivar "Pinto" UI-114 (Yáñez-Farias et al., 1997).
Estos atributos uniformemente compartidos pueden ser considerados como característicos de la especie y, por lo tanto, no diagnósticos de las variedades.
Otros aspectos que presentan diferentes valores para las variedades silvestre y cultivada pueden ser tomados, en cambio, como diagnósticos del estatus e indicadores a nivel intraespecífico. Entre estos últimos se destacan a nivel macroscópico las variables cuantitativas latitud, longitud, grosor, largo/ancho y peso y, ciertos caracteres cualitativos como forma, superficie y presencia o ausencia de micrópila. A nivel microscópico se observan parámetros cuantitativos relativos a la relación largo/ancho y al ancho de los granos de almidón, la tendencia formal del conjunto de los granos de almidón, su homogeneidad, regularidad y simetría, y la mayor o menor distinción del hilo y los anillos de crecimiento, principalmente, aunque estos últimos dependen del estado de conservación y de la historia de la muestra.
Como se mencionó anteriormente, es notable que la selección antrópica fue dirigida, principalmente a modificar las características de las semillas. Coincidiendo con lo relevado por otros autores para sitios arqueológicos del Noroeste argentino (Carrizo et al., 1999; Oliszewski, 2004a, 2005; Pochettino & Scattolin, 1991; entre otros) nuestros resultados muestran la búsqueda de ejemplares domésticos más grandes y voluminosos. Si bien no constituye un tema abordado en este trabajo, consideramos pertinente una futura exploración de las causas de la evidente modificación de la morfología de los granos de almidón como resultado de la domesticación tal como ocurre a raíz de variaciones en la composición del almidón en Zea mays L. (Watson, 1984).
En lo que respecta a la variación de los caracteres diagnósticos en relación con las características ambientales, a nivel macroscópico éstas no son significativas. Para el caso de la variedad silvestre cabe aclarar que, a pesar de separarse en base a sus dimensiones en tres grupos, no se observan variaciones de importancia debidas al tipo de ambiente que tengan implicancia a nivel infraespecífico. Las diferencias observadas se deberían a la variabilidad propia de Phaseolus vulgaris var. aborigineus.
Por su parte, los efectos de las condiciones de crecimiento (incluyendo el emplazamiento, estación y año de cosecha, niveles de fertilidad, edad de la plantación y prácticas de cultivo) en las características de los granos de almidón, han sido evaluados en otras especies de legumbres, tubérculos y cereales (Haase & Plate, 1996; Mitch, 1984; Shannon & Garwood, 1984). Estos estudios han demostrado que los efectos ambientales no son tan intensos como los debidos a la especie, diferentes cultivares y su grado de madurez (Shannon & Garwood, 1984). En el caso de Phaseolus vulgaris var. aborigineus, a partir de esta investigación se confirma esta tendencia, ya que no se han observado divergencias significativas entre las extracciones, a diferencia de lo que ocurre cuando se compara a esta variedad con su pariente cultivada.
CONCLUSIONES
En este trabajo se han podido diferenciar aquellos atributos macro- y microscópicos de las semillas de Phaseolus vulgaris var. vulgaris y P. vulgaris var. aborigineus debidos a la especie, de aquellos relacionados con el estatus, desestimando a las condiciones ambientales como fuente de variabilidad de dichos atributos. Los aspectos analizados en las semillas han ido más allá de aquellos tradicionalmente descritos para centrarnos en otros previamente no reconocidos, que podrían dar respuesta a cuestiones de asignación taxonómica de la especie y la variedad en ejemplares arqueológicos.
Esta línea puede ser útil tanto cuando la especie se presenta como macrorrestos arqueobotánicos en diferentes estados de conservación, así como cuando se dispone únicamente de sus microrrestos en las superficies de instrumentos y sedimentos arqueológicos. No obstante, se considera que el uso de los caracteres macroscópicos analizados se restringe a aquellos casos en que el material arqueológico presenta los rasgos diagnósticos bien preservados, ya sea en estado deshidratado o carbonizado, completo o fragmentado. Los caracteres microscópicos observables con microscopia con luz polarizada, pueden ser aplicables a conjuntos numerosos o que presenten granos con atributos diagnósticos, con la ventaja de que el almidón posee una buena resistencia a la acción de diferentes procesos tafonómicos naturales y culturales (Babot, 2003; Haslam, 2004; Pochettino & Scattolin, 1991). Aunque en el caso de Phaseolus vulgaris no es excluyente, el uso de microscopia electrónica de barrido puede aportar información complementaria para la asignación taxonómica, a partir de las características de la morfología de superficie de los granos de almidón y su observación tridimensional.
Por último, y atendiendo a la facilidad de mutua hibridación que presenta Phaseolus vulgaris (Singh et al., 1991), es importante poder distinguir con precisión en el registro arqueológico si se trata de variedades silvestres, domésticas o incluso intermedias (estas últimas, potencialmente originadas a partir de la cruza aleatoria o deliberada de ejemplares silvestres y domésticos). En efecto, la existencia de híbridos e introgresantes entre P. vulgaris var. aborigineus y P. vulgaris var. vulgaris ha sido informada en varias poblaciones bolivianas (Freyre et al., 1996). Lo mismo puede citarse para algunas poblaciones peruanas de P. lunatus que se desarrollan espontáneamente y se cruzan con las variedades silvestres circundantes (Debouck, 1994). Estas semillas intermedias, que se encuentran a medio camino entre las variedades silvestre y doméstica, podrían haber sido producidas por plantas con un grado de selección tal que no afectara totalmente sus caracteres fenotípicos. Esto es factible si consideramos a los procesos de domesticación no como acotados en un único punto en el tiempo y el espacio, sino como búsquedas constantes con objetivos variables dentro de un cierto rango, ocurriendo en múltiples lugares al mismo tiempo.
AGRADECIMIENTOS
A los evaluadores cuyas sugerencias permitieron enriquecer el trabajo; a los editores de Darwiniana, en especial a M. Fernanda Rodríguez y Raúl Pozner, por sus valiosos comentarios y a Luis Babot por su colaboración en la digitalización de las imágenes.
BIBLIOGRAFÍA
1. Albeck, M. E. 2000. La vida agraria en los Andes del Sur, en M. N. Tarragó (ed.), Nueva Historia Argentina, tomo I, pp. 187-228. Buenos Aires: Editorial Sudamericana. [ Links ]
2. Babot, M. del P. 2003. Starch grain damage as an indicator of food processing, en D. Hart & L. Wallis (eds.), Phytolith and starch research in the Australian-Pacific-Asian regions: the state of the art, pp. 69-81. Canberra: Pandamus Books for the Centre for Archaeological Research (ANU). [ Links ]
3. Babot, M. del P. 2004. Tecnología y utilización de artefactos de molienda en el Noroeste prehispánico. Tesis de Doctorado en Arqueología, Universidad Nacional de Tucumán. [ Links ]
4. Babot, M. del P. 2007. Granos de almidón en contextos arqueológicos: posibilidades y perspectivas a partir de casos del Noroeste argentino, en M. B. Marconetto, M. del P. Babot & N. Oliszewski (eds.), Paleoetnobotánica del Cono Sur: estudios de casos y propuestas metodológicas, pp. 95-125. Córdoba: Ferreyra Editor para el Museo de Antropología, Facultad de Filosofía y Humanidades, Universidad Nacional de Córdoba. [ Links ]
5. Baudet, J. C. 1977. Origine et classification des espècies cultivées du genre Phaseolus. Bull. Soc. Roy. Bot. Belg. 110: 65-76. [ Links ]
6. Berglund-Brücher, O. & H. Brücher. 1976. The Southamerican wild bean (Phaseolus aborigineus Burk.) as ancestor of the common bean. Econ. Bot. 30: 257-272. [ Links ]
7. Brown, A. D.; H. R. Grau; L. R. Malizia & A. Grau. 2001. Argentina, en M. Kappelle & A. Brown (eds.), Bosques nublados del Neotrópico, pp. 623-659. Santo Domingo de Heredia, Costa Rica: Instituto Nacional de Biodiversidad. [ Links ]
8. Burkart, A. 1952. Las leguminosas argentinas silvestres y cultivadas, 2ª. edición. Buenos Aires: Acme. [ Links ]
9. Buttrose, M. S. 1960. Submiscroscopic development and structure of starch granules in cereal endosperms. Ultraestructure Research 4: 231-257. [ Links ]
10. Carrizo, J.; S. F. Cano & M. M. Soler Nixdorff. 1999. Recursos vegetales comestibles en el Valle de Tafí durante el Período Formativo: análisis arqueobotánico del sitio Casas Viejas- El Mollar (STucTav2). Actas del XII Congreso Nacional de Arqueología Argentina, 22-26 de septiembre de 1997, La Plata (Argentina), Tomo I: 65-73. [ Links ]
11. Carrizo, J.; N. Oliszewski & J. G. Martínez. 2003. Macrorrestos vegetales del sitio arqueológico Cueva de los Corrales (El Infiernillo, Tafí del Valle, Tucumán). Revista. Mus. Argent. Cienc. Nat. 5: 253-260. [ Links ]
12. Chevalier, A. 1999. De la pomme de terre à la coca: statut de l'archéobotanique au Pérou et revue critique des données, en A. Chevalier, L. Velarde & I. Chenal-Velarde (eds.), L'Amérique du Sud. Des chasseurs-cueilleurs à l'Empire Inca, pp. 43-61. Cambridge: BAR International Series 746. [ Links ]
13. Cortella, A. & M. L. Pochettino. 1994. Starch grain analysis as a microscopic diagnostic feature in the identification of plant material. Econ. Bot. 48: 171-81. [ Links ]
14. Cortella, A. & M. L. Pochettino. 1995. Comparative morphology of starch of three Andean tubers. Stärke 47: 455-461. [ Links ]
15. Debouck, D. G. 1994. Beans (Phaseolus spp.), en J. E. Hernándo Bermejo & J. León (eds.), Neglected Crops: 1492 from a different perspective, pp. 47-62. Plant Production and Protection Series 26. Roma: Organización de las Naciones Unidas para la Agricultura y la Alimentación, FAO. [ Links ]
16. French, D. 1984. Organization of starch granules, en R. Whistler, J. Bemiller & E. Paschall (eds.), Starch: Chemistry and Technology, pp. 183-247, 2a. edición. Florida: Academic Press. [ Links ]
17. Freyre, R.; R. Ríos; L. Guzmán; D. G. Debouck & E. Gepts. 1996. Ecogeographic distribution of Phaseolus spp. (Fabaceae) in Bolivia. Econ. Bot. 50: 195-215. [ Links ]
18. Gentry, H. S. 1969. Origin of the common bean, Phaseolus vulgaris. Econ. Bot. 23: 55-69. [ Links ]
19. Gepts, P. 2006-2007. Crop evolution, domestication and biodiversity [on line]. Fezocaonline (California, USA): Department of Plant Sciences, University of California. Davis. <http://www.agronomy.ucdavis.edu/gepts>. [Consulta: diciembre 2006]. [ Links ]
20. Haase, N. U. & J. Plate. 1996. Properties of potato starch in relation to varieties and environmental factors. Starkë 48: 167-171. [ Links ]
21. Haslam, M. 2004. The decomposition of starch grains in soils: implications for archaeological residue analyses. J. Archl. Sci. 31: 1715-1734. [ Links ]
22. Johnson, W. C. & P. Gepts. 1999. Segregation for performance in recombinant inbred populations resulting from inter-gene pool crosses of common bean (Phaseolus vulgaris L.). Euphytica 106: 45-56. [ Links ]
22. Kami, J.; V. B. Velásquez; D. G. Debouck & P. Gepts. 1995. Identification of presumed ancestral DNA sequences of phaseolin in Phaseolus vulgaris. Proc. Natl. Acad. Sci. USA 92: 1101-1104. [ Links ]
23. Kaplan, L. 1965. Archaeology and domestication in American Phaseolus. Econ. Bot. 19: 358-368. [ Links ]
24. Kaplan, L. & T. F. Lynch. 1999. Phaseolus: AMS radiocarbon dates and their significance for pre-columbian agriculture. Econ. Bot. 53: 261-272. [ Links ]
25. Korstanje, M. A. & M. del P. Babot. 2007. Microfossil characterization from south Andean economic plants, en M. Madella & D. Zurro (eds.), Plants, people and places. Recent studies in phytolithic analysis. Cambridge: Oxbow Books. [ Links ]
26. Korstanje, M. A. & A. E. Würschmidt. 1999. Producir y recolectar en los valles altos del NOA: "Los Viscos" como caso de estudio, en C. A. Aschero, M. A. Korstanje & P. M. Vuoto (eds.), En los tres reinos: prácticas de recolección en el cono sur de América, pp. 151-160. San Miguel de Tucumán: Magna Ediciones. [ Links ]
27. Lackey, J. A. 1983. A review of generic concepts in American Phaseolinae (Phabaceae, Faboideae). Iselya 2: 21-64. [ Links ]
28. Lagiglia, H. A. 2001. Los orígenes de la agricultura en la Argentina, en E. E. Berberián & A. E. Nielsen (eds.), Historia argentina prehispánica, Tomo I, pp. 41-81. Córdoba: Editorial Brujas. [ Links ]
29. Mitch, E. L. 1984. Potato starch: production and uses, en R. Whistler, J. Bemiller & E. Paschall (eds.), Starch: Chemistry and Technology, pp. 479-490, 2a. edición. Florida: Academic Press. [ Links ]
30. Munsell Soil Color Charts. 2000. Revised washable edition. New York: GretagMacbeth. [ Links ]
31. Oliszewski, N. 2004a. Estado actual de las investigaciones arqueobotánicas en sociedades agroalfareras del área valliserrana del NOA (0 - 600 D.C.). Relacs Soc. Arg. Antrop. 29: 211-227. [ Links ]
32. Oliszewski, N. 2004b. Utilización de recursos vegetales en Campo del Pucará (Andalgalá, Catamarca) durante el Período Formativo (200-500 d.C.). Análisis de macrorrestos. Tesis de Doctorado en Arqueología, Universidad Nacional de Tucumán. [ Links ]
33. Oliszewski, N. 2005. Archaeobotany of archaeological sites from Northwest Argentina (1750-1450 B.P.): ceremonial use or rubbish dumps? Vegetation History and Archaeobotany 14: 465-471. [ Links ]
34. Parodi, L. 1991. Agricultura prehispánica. San Salvador de Jujuy: Editorial de la Universidad Nacional de Jujuy. [ Links ]
35. Pearsall, D. 1994. Issues in the analysis and interpretation of archaeological maize in South America, en S. Johannessen & C. A. Hastorf (eds.), Corn and Culture in the Prehistoric New World, pp. 245-272. Boulder: Westview Press. [ Links ]
36. Piperno, D. R. & I. Holst. 1998. The presence of starch grains on prehistoric stone tools from the Humid Neotropics: indications of early tuber use and agriculture in Panama. J. Archl. Sci. 25: 765-776. [ Links ]
37. Pochettino, M. L. & M. C. Scattolin. 1991. Identificación y significado de frutos y semillas carbonizados de sitios arqueológicos de la ladera occidental del Aconquija, Prov. Catamarca, Rca. Argentina. Rev. Mus. La Plata, Antropol. 9: 169-181. [ Links ]
38. Reichert, C. T. 1913. The differentiation and specificity of starches in relation to genera, species, etc. Washington D. C.: Carnegie Institution of Washington, Publication 173. [ Links ]
39. Shannon, J. C. & D. L. Garwood. 1984. Genetics and physiology of starch development, en R. Whistler, J. Bemiller & E. Paschall (eds.), Starch: chemistry and technology, pp. 25-86, 2a. edición. Florida: Academic Press. [ Links ]
40. Singh, S. P.; P. Gepts & D. G. Debouck. 1991. Races of common bean (Phaseolus vulgaris, Fabaceae). Econ. Bot. 45: 379-396. [ Links ]
41. Tapia, M. 1990. Cultivos andinos subexplotados y su aporte a la alimentación. Santiago de Chile: Organización de las Naciones Unidas para la Agricultura y la Alimentación, FAO. [ Links ]
42. Tarragó, M. N. 1980. El proceso de agriculturización en el Noroeste Argentino, zona valliserrana. Actas del V Congreso Nacional de Arqueología Argentina, 1978, San Juan (Argentina), Tomo I: 181-218. [ Links ]
43. Torrence, R. & H. Barton. 2006. Ancient starch research. Walnut Creek, California: Left Coast Press. [ Links ]
44. Watson, S. A. 1984. Corn and sorghum starches: production, en R. Whistler, J. Bemiller & E. Paschall (eds.), Starch: chemistry and technology, pp. 417-467, 2a. edición. Florida: Academic Press. [ Links ]
45. Yánez-Farias, G. A.; J. G. Moreno-Valencia; M. R. Falcón-Villa & J. M. Barrón-Hoyos. 1997. Isolation and partial characterization of starches from dry beans (Phaseolus vulgaris) and chickpeas (Cicer arietinum), grown in Sonora, Mexico. Starkë 49: 341-345. [ Links ]
46. Zallocchi, E. M.; A. B. Pomilio & R. A. Palacios. 1995. Estudio quimiotaxonómico de la subtribu Phaseolinae (Phaseoleae- Papilionoideae-Leguminosae) III: flavonoides de las especies argentinas de los géneros Phaseolus y Dolichopsis. Darwiniana 33: 135-148. [ Links ]
47. Zuloaga, F. & O. Morrone (eds.). 1999. Catálogo de las plantas vasculares de la República Argentina, Fabaceae - Zygophyllaceae (Dicotyledoneae). Monogr. Syst. Bot. Missouri Bot Gar. 74: 623-1269. [ Links ]

