










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkDarwiniana, nueva serie
Print version ISSN 0011-6793On-line version ISSN 1850-1699
Darwiniana vol.45 no.2 San Isidro Aug./Dec. 2007
Canna ascendens (Cannaceae), una nueva especie de la provincia de Buenos Aires y comentarios sobre otras especies argentinas de este género
María de las Mercedes Ciciarelli
Laboratorio de Estudios de Anatomía Vegetal Evolutiva y Sistemática (LEAVES), Facultad de Ciencias Naturales y Museo de La Plata, calle 64 Nº 3, entre calle 120 y diagonal 113, B1904 DZB La Plata, Argentina; mmciciar@yahoo.com.ar
Original recibido el 29 de noviembre de 2006;
aceptado el 8 de agosto de 2007
Resumen. Ciciarelli, M. M. 2007. Canna ascendens (Cannaceae), una nueva especie de la provincia de Buenos Aires y comentarios sobre otras especies argentinas de este género.
Canna ascendens sp. nov. se describe a partir de poblaciones situadas en localidades húmedas, costeras y centrales de la provincia de Buenos Aires, a menudo coexistiendo con C. variegatifolia, otra especie colonial y urbana representada por plantas más grandes, con hojas variegadas. Las plantas de C. ascendens son altas y grandes, aunque más delicadas y de tamaño algo menor que las de C. variegatifolia. Las hojas son concolores, verdes, ovadas a ovado lanceoladas, con frecuencia se doblan lo largo de la vena media, en forma conduplicada; en estado natural presentan una cubierta de cera epicuticular sobre la epidermis plegada y estriada y abundantes protuberancias intercelulares pécticas (verrugas) distribuidas sobre las paredes de las células de las hipodermis abaxial y adaxial. Las inflorescencias son politélicas reducidas, con 1-2 paracladios o con éstos ausentes. Las flores son pocas, a menudo una sola, de color anaranjado rosado apagado. Se efectuaron análisis de la anatomía, se da una descripción completa de la nueva especie que incluye nuevos caracteres y se la comparó con otras que crecen en la misma área y con otras especies de Argentina.
Palabras clave. Anatomía; Argentina; Buenos Aires; Canna; Cannaceae; Morfología; Taxonomía.
Abstract. Ciciarelli, M. M. 2007. Canna ascendens (Cannaceae), a new species from Buenos Aires Province, and comments on other argentine species of this genus.
Canna ascendens sp. nov. is described from several populations found in moist coastal and central locations of Buenos Aires Province, Argentina, often growing together with another colonial, urban species, C. variegatifolia, having larger plants with variegated leaves. Plants of C. ascendens are tall and large, yet smaller and more delicate than plants of C. variegatifolia; the leaves are concolorous, green, ovate to ovate-lanceolate in outline, usually conduplicate along their midrib with epicuticular wax over a striated or folded epidermis. More or less abundant intercellular pectic protuberances (warts) were found on the cell walls of both adaxial and abaxial hypodermis. The inflorescences are polytelic, reduced, with 1-2 paraclades or, eventually, no paraclades at all. Paraclade flowers are few, often only one, wtih a dull, rosy orange color. Plants were investigated anatomically, and a detailed description with diagnostic characters is included. The new species is compared with other species of Canna that grow in the area and also with other argentine species.
Keywords. Anatomy; Argentina; Buenos Aires; Canna; Cannaceae; Morphology; Taxonomy.
INTRODUCCIÓN
Canna L. es un género pantropical perteneciente a la familia Cannaceae A. L. de Jussieu. Juntamente con las Costaceae (K. Schum.) Nak., Heliconiaceae (Endl.) Nak., Marantaceae Petersen, Musaceae A. L. de Jussieu, Strelitiziaceae (K. Schum.) Hutch. y Zingiberaceae Adanson, las Cannaceae están incluidas en el orden Zingiberales. considerado monofilético (Tomlinson, 1962, 1969; Dahlgren & Rassmusen, 1983; Kress, 1990, 1995; Stevenson & Loconte, 1995; Chase et al., 1993; Smith et al., 1993; Kress et al., 2001). Cannaceae, Costaceae, Marantaceae y Zingiberaceae formarían un clado único basado en la reducción del androceo (un solo estambre funcional), los estaminodios grandes y llamativos, las semillas predominantemente perispermadas, la ausencia de ráfides en los tejidos parenquimáticos y las hojas íntegras y firmes, que no se desgarran (Judd et al., 2002).
Las especies de Canna habitan los trópicos y subtrópicos de todo el mundo. Sus flores llamativas e intensamente coloreadas han llevado al cultivo de las plantas como ornamentales y a la producción de variedades hortícolas y de híbridos muy numerosos. Mass (1988) reconoce solamente 10 especies para América y Tanaka (2001), en una revisión reciente del género, considera la existencia de 19 taxones silvestres.
En el curso de un estudio morfológico comparado de especies argentinas de Canna, se encontraron, en la provincia de Buenos Aires, numerosas comunidades de una planta que no pudo ser asignada a ninguna de las especies previamente descritas para el género, por lo que aquí se la describe como nueva para la ciencia, incluyendo numerosos rasgos diagnósticos y observaciones adicionales. También se la compara con otras especies bonaerenses y argentinas. Se incluye una clave para distinguir las especies que crecen en la provincia de Buenos Aires y una tabla en la que se resumen los caracteres más destacados de las especies argentinas.
MATERIALES Y MÉTODOS
En este estudio se empleó material fresco coleccionado por la autora. Las colecciones se realizaron desde setiembre hasta mayo, con el fin de coleccionar partes vegetativas, flores, frutos y semillas.
Los caracteres foliares externos, como la relación largo/ancho (en adelante l/a), contorno, base, ápice y venación se estudiaron en el material fresco y se confirmaron luego en material herborizado. La descripción de la base de la lámina corresponde a la porción expandida antes de escurrirse para formar la vaina foliar. El tamaño foliar promedio se obtuvo midiendo 25 hojas de posición media.
Para el estudio de la venación y la epidermis se aclararon trozos de hojas con hidróxido de sodio 3% acuoso e hipoclorito de sodio comercial (40% acuoso), sin diluir o diluido en agua en proporción 1:1 y se colorearon con azul de toluidina (en adelante TBO) 1% acuoso. Se efectuaron preparaciones transitorias. La terminología utilizada para venación es la de Hickey (1999).
Para estudiar las inflorescencias se tomaron los datos de 25 plantas y se evaluaron los siguientes caracteres: longitud total de la inflorescencia (medida desde el nudo 1 ó nudo original, del cual parte el entrenudo basal, hasta el extremo distal de la florescencia principal), número de ejes por nudo, número de brácteas por nudo; número de paracladios o brotes repetitivos y color. Se utiliza la terminología de Weberling (1965, 1985, 1997).
Para el estudio de la morfología floral se analizaron 25 flores de 25 inflorescencias, tomadas al azar de diferentes plantas. La longitud total de la flor se midió desde la base del ovario hasta el ápice del estaminodio más largo. La longitud del tubo floral se midió desde la base de los sépalos hasta el punto en el cual el estilo se separa del estambre. Las medidas de las semillas se efectuaron sobre muestras (N = 50) tomadas al azar en cada población visitada. Los términos de color se tomaron de Stern (1996).
Para el estudio de la cera epicuticular, las ornamentaciones cuticulares y el polen se utilizó microscopia electrónica de barrido. El material fresco, recientemente coleccionado, se montó sin ningún tratamiento sobre bases de vidrio de 3 x 3 mm, con cinta adhesiva de doble faz y se metalizaron con oro en un JEOL JFC-1100 (fine coat ion sputter). Las observaciones se efectuaron con un microscopio electrónico Jeol /EO JSM 6360, en el Servicio de Microscopía Electrónica de la Facultad de Ciencias Naturales y Museo de La Plata. Los datos de las restantes especies bonaerenses fueron tomados de material fresco coleccionado por la autora y de las colecciones depositadas en los herbarios BM, CTES, L, LIL, LP, SI y P (Holmgren et al., 1990).
RESULTADOS
Canna ascendens sp. nov. TIPO: Argentina. Buenos Aires. San Pedro, Panamé, 12-12-1986, Ciciarelli 5 (holotipo LP). Fig. 1 A-C

Fig. 1. Canna ascendens. A, aspecto general de las plantas. B, detalle de las inflorescencias. C, detalle de las flores.
Plantae terrestres. Rhizoma subterraneus, repente. Folia integerrima viridis, ascendens, herbacea, laevis, nitida, ovata-lanceolata, apice acutae, retortae, basi acutae decurrentes, inaequalis; venatio eucamptodroma; cuticula striata, ceracea. Inflorescentia politelicae deminuta, cymosae, 0-1(2) paracladiis. Flores rosae aurantiaci; sepala 3, imbricata, rosa virentia; petala 3, basi connata tubiformia; staminodia 3, oblanceolata, rosa aurantiacia; labellum oblanceolatum reflexum, rosa aurantiaceo, rubra maculatum adaxiales; stylum rosa aurantiaceo. Pollinis grana globosa, echinulata, innaperturata. Capsulae ellipsoideae vel ovoideae, extrapapillosae. Semina maturae fuscato-nigra, nítida, quasi sphaerica. Canna variegatifolia affinis, a qua imprimis differt statura minore et foliis minoribus semper ascendens, viridibus, non variegati, inflorescentia deminuta, flores aurantiaci, semina minore, nitida.
Plantas terrestres, de 2-2,5 m de altura. Rizomas de (2)4 (5) cm de diámetro, crasos subterráneos, monopodiales, con nudos cubiertos por catafilos castaños y entrenudos blanquecinos; abundantes raíces abaxiales. Ejes erectos adaxiales con vainas foliares grandes, adpresas, algo carinadas, glabras, verdes. Hojas concolores verdes, glabras, de 15-50 (60) cm de longitud x 7-18 (20) cm de anchura, con láminas ovado-lanceoladas (relación largo/ancho 3:1), con ápices agudos, acuminados, retorcidos, bases agudas decurrentes asimétricas, textura herbácea, superficie de diseño liso y pulidez nítida. Venación eucamptódroma, con venas primarias, secundarias, intersecundarias, terciarias y vena intramarginal. Cutícula estriada a plegada; células epidérmicas dispuestas con modelo subpoligonal; estomas paracíticos de contorno subrectangular de 30 x 19 µm, con células anexas rectangulares y células polares no envolventes. Células de las hipodermis abaxial y adaxial translúcidas, con abundantes protuberancias intercelulares pécticas (verrugas) en las paredes. Inflorescencias politélicas reducidas, de 45-50 (55) cm de longitud, con florescencia principal y 1-2 paracladios, o a veces sin paracladios. Entrenudo basal de 20-23 cm de longitud. Florescencia principal con 5-8 nudos, paracladios con 3-6. Nudos bifloros con 3 brácteas protectoras y una flor más grande que la otra; brácteas, vainas y ejes de color verde. Flores de 6-7( 9) cm de longitud, con 12 piezas, 3 estaminodios y un cuarto estaminodio rudimentario en la base del labelo. Ovario ínfero, de hasta 1,2 cm de longitud, externamente papiloso. Sépalos 3, rosado- verdosos, ovados angostos, con ápices agudos y base truncada, el externo de hasta 1,2 cm de longitud x 0,7 cm de anchura; el medio y el interno de igual tamaño, de hasta 1 cm de longitud x 0,6 cm de anchura. Pétalos 3, rosado-anaranjados, oblanceolados, con ápices agudos y connatos en la base, el externo y medio de hasta 5,2 cm de longitud x 0,8 cm de anchura y el interno algo menor, de hasta 3,4 cm de longitud x 0,5 cm de anchura. Estaminodios 3, de color amarillo-anaranjado, anaranjado claro o intenso, oblanceolados, con ápices apiculados, agudos o acuminados, el externo de hasta 6 cm de longitud x 1,5 cm de anchura, el medio y el interno algo más pequeños, de hasta 5 cm de longitud x 1 cm de anchura. Labelo reflexo, amarillo-anaranjado con manchas rojizas adaxiales, con ápice emarginado o entero. Estambre rosado-anaranjado, la parte petaloide con ápice reflexo y antera marginal retorcida de 1 cm de longitud. Estilo rosado-amarillento, erguido, con área estigmática marginal y apical. Tubo floral de hasta 2,6 cm de longitud. Granos de polen esféricos, equinados, inaperturados, de 80 (86) 90 µm de diámetro. Fruto cápsula, ovoide, de hasta 3 x 2,5 cm, externamente papilosa. Semillas casi esféricas, de 0,6 a 0,8 cm de diámetro, de color castaño oscuro a negro a la madurez, lisas.
Paratipos
ARGENTINA. Buenos Aires. La Plata: Ringuelet, Camino Centenario y 519, frente a Carrefour, 10-III-2006 Ciciarelli 7 (LP); City Bell. Camino Centenario y La Croze, 10-III-2006 Ciciarelli 8 (LP).
Distribución geográfica y hábitat
Las poblaciones de Canna ascendens se encuentran en la provincia de Buenos Aires, en localidades ribereñas de La Plata, y también en City Bell, Ringuelet y San Pedro. Crecen en suelos húmedos pero no anegados, en terrenos baldíos y en áreas urbanas abiertas, siempre en suelos ricos, a veces removidos. En Buenos Aires coexiste con C. variegatifolia, una especie cuyas plantas pueden exceder los 4 m de altura. Las comunidades de ambas especies pueden crecer próximas pero también se las encuentra independientes una de otra, en localidades distintas de la provincia. Canna variegatifolia vive también en las provincias de Santa Fe y Corrientes (Ciciarelli, 1995) pero C. ascendens no se ha encontrado aún fuera del ámbito bonaerense.
Etimología. El epíteto ascendens hace referencia al carácter saliente de las hojas, ascendentes, nunca patentes ni reflexas como en otras especies de Canna (por ejemplo C. indica L.).
Observaciones y comentarios adicionales. Además de la posición foliar, rasgo que se ha utilizado en el epíteto específico, otro aspecto destacable es la tendencia de las hojas a plegarse a lo largo de la vena primaria, desde la base, de manera más o menos conduplicada e involuta. En períodos secos este carácter se acentúa notablemente y las hojas se enroscan sobre su cara adaxial formando una prolongación de la vaina, que es fuerte y adpresa (Fig. 1A).
Canna ascendens tiene afinidades con C. variegatifolia, pero se distingue de ésta por el menor tamaño de las plantas, los rizomas más delgados, el mayor número de ejes ascendentes producidos en cada nudo y las hojas simétricas concolores, verdes en el haz y el envés, también de menor tamaño.
Los rizomas de C. ascendens y C. variegatifolia son monopodiales y estoloníferos. En C. ascendens el eje principal plagiótropo presenta 3-4 entrenudos sin raíces, y forma en el cuarto o quinto nudo hasta 4 ejes ascendentes que se transformaran en otros tantos ejes floríferos, mientras que en la zona de división del eje principal se produce un gran fascículo de raíces que están ausentes en los entrenudos previos. En C. variegatifolia el eje principal plagiótropo se divide en dos ramas cada 4-5 entrenudos sin raíces; una de las ramas continúa el crecimiento plagiótropo y la otra, ascendente, origina una sola planta. Ciciarelli (1989) reconoció dos tipos de rizomas: los monopodiales o estoloníferos, más o menos gruesos, con entrenudos cortos y los simpodiales o tuberosos, en los cuales uno de dos ejes aborta para formar un tuberibulbo, mientras el otro continúa creciendo. Ciciarelli (1995) ilustró el tipo monopodial o estolonífero para C. variegatifolia. Tanaka (2001) describió cuatro tipos de rizomas para Canna sobre la base del diámetro, la distancia entre los nudos, la cantidad de ramas producidas en el eje principal y la cantidad de raíces por nudo, sin hacer referencia a la ramificación, de manera que las clasificaciones de ambos autores no son estrictamente comparables.
Las inflorescencias de C. ascendens y C. variegatifolia son politélicas, consisten de un eje o conjunto de ejes ramificados simpodialmente de manera repetitiva, en el que todos los paracladios tienen estructura similar y son idénticos a la florescencia principal (brotes repetitivos). Canna ascendens tiene inflorescencias reducidas, con una florescencia principal con pocas flores y 1-2 paracladios, aunque éstos pueden faltar y en ese caso la florescencia principal lleva una sola flor. Las inflorescencias son más cortas que las de C. variegatifolia, que son ramificadas y tienen 2-4 paracladios. Mientras que C. ascendens presenta color verde en ejes, brácteas florales y paracladiales, en C. variegatifolia el color de los ejes, brácteas florales y paracladiales es verde, blanco y amarillo muy claro, variegado.
Las flores de C. ascendens (Figs. 1B-C y 2A) y las de C. variegatifolia (Fig. 2B) tienen 12 piezas; las de C. ascendens tienen color anaranjado-amarillento o rosado, de claro a intenso y algo opaco y las de C. variegatifolia color rosado-anaranjado a rosado salmón. El labelo es reflexo, lanceolado, con base ensanchada truncada y ápice emarginado o entero y tiene color rosado-anaranjado con manchas rojizas adaxiales en C. ascendens (Fig. 2, A), mientras que es oblanceolado, circinado o girado, con base truncada y ápice emarginado en C. variegatifolia ( Fig. 2B).

Fig. 2. Morfología vegetativa y floral de Canna. A, C. ascendens, flor. B, C. variegatifolia, flor. C-D, C. variegatifolia, epidermis. E-F, C.ascendens, epidermis. G, C.ascendens, corte tranversal de epidermis. H, C. variegatifolia, corte tranversal de epidermis. Abreviaturas: e, estilo; es, estaminodio; est, estambre; l, labelo. En las epidermis, izquierda epifilos; derecha, hipofilos. Barra = 5 cm en A-B; 70 µm en B-F; 130 µm en G-H.
La micromorfología y anatomía foliar también sugiere afinidades entre C.ascendens y C. variegatifolia. La cera epicuticular, que parece ser común en la epidermis de las especies de Canna, es particularmente abundante tanto en C. ascendens (Fig. 3E, G) como en C. variegatifolia (Fig. 3F). En ambas especies aparece en forma de escamas y placas algo irregulares, dispuestas más o menos paralelas unas con otras. La cutícula de C. ascendens (Fig. 3A-B) y C. variegatifolia (Fig. 3C-D) está ornamentada con estrías gruesas o pliegues dispuestos longitudinalmente en todas las células epidérmicas. Los modelos epidérmicos de ambas especies son poligonales a subpoligonales, de contornos más o menos rectos (Fig. 2C-F), con paredes celulares gruesas, de hasta 2,6 µm de espesor en C. ascendens y de hasta 2,2 µm en C. variegatifolia (Fig. 2G-H). Los estomas son paracíticos (Fig. 2C-F; Fig. 3A-C, E-F). El grupo de células oclusivas y células anexas más o menos reniformes o semilunares fue llamado "complejo paracítico" por Ciciarelli (1989), por su reacción conjunta ante el TBO 1% acuoso y está difundido en Canna, aunque su contorno y tamaño varian en el nivel específico. En C. ascendens y C. variegatifolia los estomas tienen contorno oblongo, con células oclusivas casi rectangulares y células anexas reniformes que contactan en los polos; tienen 30 x 19 µm en C. ascendens y 32 x 24 µm en C. variegatifolia. La cera puede ser muy abundante sobre las células anexas del complejo paracítico en C. ascendens (Fig. 3 F). El mesofilo de las hojas de C. ascendens es dorsiventral, con una hipodermis abaxial y otra adaxial cuyas células presentan protuberancias intercelulares pécticas (PIP) en forma de verrugas esféricas de diferentes tamaños (Fig. 3 H). Estas PIP son también abundantes en el tejido parenquimático del área media foliar. Las PIP se conocen desde hace más de un siglo, pero recientemente Potgieter & van Wyk (1992) las describieron con este nombre para Icacinaceae, las clasificaron en verrugas, filamentos y conexiones pécticos y efectuaron una prospección bibliográfica de las familias y géneros o especies de plantas de todos los grupos taxonómicos que las presentan. Están muy difundidas en el reino vegetal y no es posible establecer claramente relaciones de parentesco sobre la base de su presencia. Algunos autores sugieren, sin embargo, que podrían representar rasgos de afinidad en el nivel genérico o específico en ciertas familias (Potgieter & van Wyk, 1992; Rolleri et al., 1999; Prada & Rolleri, 2004). Potgieter & van Wyk (1992) y Prada & Rolleri (2004) resumieron las diversas interpretaciones sobre las funciones de estas estructuras y es posible que su amplia distribución en taxones diferentes resida en la convergencia de una o más funciones de las PIP. En Monocotiledóneas estarían presentes en familias de Arales, Arecales, Asparagales, Commelinales y Poales pero no hay antecedentes en Zingiberales, de modo que ésta es la primera cita para Zingiberales. Además de C. ascendens, las PIP se han encontrado en C. x generalis (Ciciarelli, datos no publicados), hasta el momento también en forma de verrugas. No se observaron en C. variegatifolia.
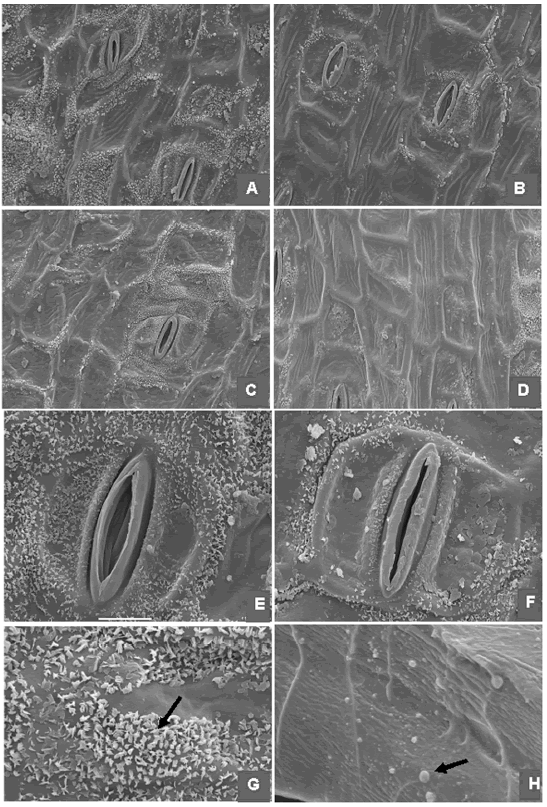
Fig. 3. Epidermis de Canna. A, C. ascendens, epifilo. B, C. ascendens, hipofilo. C, C. variegatifolia, epifilo. D, C.variegatifolia, hipofilo. E, C. ascendens, detalle estoma paracítico y cera epicuticular. F, C. variegatifolia, estoma paracítico y cera epicuticular. G, C. ascendens, detalle de cera epicuticular. H, C. ascendens, verrugas pécticas (PIP) en células de la hipodermis. Barra= 56 µm en A-B, C-D; 18 µm en E-F; 5 µm en G-H.
Los granos de polen de C. ascendens son esféricos, inaperturados, de (80) 86 (90) µm. El esporodermo está formado por una exina espinulosa muy delgada y una intina transparente de (4,5) 6 (10) µm, que representa el mayor espesor del esporodermo. La exina tiene espínulas cónicas espaciadas, tan altas como su ancho basal, de 2 x 2 µm (Fig. 4C, E). La intina tiene dos capas, una externa de (3) 4 (5) µm y una interna de (1,5) 2 (5) µm. Todas las poblaciones analizadas mostraron un porcentaje de normalidad de casi el 99 % (Fig. 4A) y muchos granos se encontraron en el proceso de emitir el tubo polínico sobre el estilo de las flores frescas. Canna variegatifolia tiene granos de (72) 80 (81) µm , es decir, de tamaño algo menor y el esporodermo, de (5) 7 (10) µm, es más grueso. Las espínulas de la exina son más cortas, de 1,5 x 1,5 µm (Fig. 4D, F) y las capas externa e interna de la intina miden (3) 4 (5) µm y (2) 3 (5) µm respectivamente. También en esta especie el porcentaje de normalidad es casi del 100 % (Fig. 4B) y se registró emisión de los tubos polínicos. Segeren & Maas (1971) estudiaron el polen de las especies de Canna del N de América del Sur y describió granos esféricos, inaperturados, con paredes gruesas y exina espinulosa, con un diámetro variable entre 55 y 77 µm. Esos autores registraron diámetros de ca. 60 µm, esporodermos de 4 µm de espesor y espínulas de 3,5 x 3 µm para granos de C. indica y diámetros de ca. 62 µm, espesor de pared de 5 µm espínulas de 3 x 2,5 µm para C. glauca, señalando que el espesor del esporodermo en Canna puede alcanzar los 8 µm. Para Segeren & Maas (1971) los caracteres del polen no aportarían información relevante para la sistemática del género, pero Nair (1960) registró variaciones que consideró de importancia en la clasificación de las especies cultivadas.

Fig. 4. Polen de Canna. A, C. ascendens, poblaciones de granos normales. B, C. variegatifolia, poblaciones de granos normales. C, C. ascendens, un grano de polen. D, C. variegatifolia, un grano de polen, con el tubo polínico emergente. E, C. ascendens, detalle de exina espinulosa. F, C. variegatifolia, detalle de exina espinulosa. Barra = 170 µm en A-B; 20 µm en C-D; 5 µm en E-F.
Las semillas se producen con regularidad, tanto en C. ascendens como en C. variegatifolia, en ésta última en mayor cantidad por la presencia de inflorescencias ramificadas y mayor número de flores: en C. variegatifolia, con inflorescencias ramificadas y hasta 16 nudos floríferos en la florescencia principal y 13 en los paracladios de mayor desarrollo, se producen, en general, hasta 6 frutos, mientras que en C. ascendens, con inflorescencias reducidas con 5-8 nudos floríferos en la florescencia principal y 3 en los paracladios, se producen de 1 a 3 frutos. Ambas especies producen semillas de tamaño similar, inicialmente blancas, negras a la madurez y con cubierta lisa. La multiplicación vegetativa en las poblaciones no parece influir en una merma en la producción de semillas, que germinan normalmente en estado natural si bien su cubierta dura debe escarificarse con el fin de lograr su germinación en laboratorio (Ciciarelli, datos no publicados). Son devoradas por pájaros de la fauna bonaerense (Colúmbidos, Pscittaciformes, Mimidiformes) lo que facilitaría la germinación natural por el paso de las semillas por el sistema digetivo de las aves y explicaría la dispersión de las poblaciones en áreas distantes de la provincia o en otras áreas, en el caso de C. variegatifolia.
Los granos de polen normales, la emisi ón de tubos polínicos observada y la producción estacional de semillas sugieren que tanto C. ascendens como C. variegatifolia son taxones sexuales normales, no híbridos.
El color reviste una importancia destacable en especies de Canna, ya que se presenta tanto en órganos vegetativos como en flores y puede variar de una especie a otra. Algunos taxones son muy característicos y se reconocen fácilmente por el color de las hojas, de las flores o de ambas. Las hojas son concolores o discolores: ejemplos de esto son las hojas concolores verdes de C. ascendens o concolores glaucas de C. glauca y las hojas discolores, con lámina verde y márgenes y venas primarias de color púrpura de C. coccinea, o las hojas verde-amarillentas, fasciadas en el haz y verde-grisáceas en el envés, de C. indica. Algunos híbridos de C. indica, C. x generalis tienen las mismas características que su presunto parental. Las hojas de C. variegatifolia podrían describirse como tricolores, con láminas de color verde a verde-grisáceo oscuro, blanco y amarillo pálido, aunque en general se las describe como variegadas. Las hojas variegadas muestran una ausencia completa de cloroplastos en los sectores foliares con parches blancos, que se debería a una mutación citológica local (quimera sectorial) que induce la formación de células protodérmicas sin cloroplastos o con plástidos que no completan la formación de su aparato pigmentario, por lo que suele interpretarse que las plantas con estas características tienen una eficiencia fotosintética menor y crecen lentamente. La ausencia de cloroplastos en las áreas del mesofilo de C. variegatifolia fueron descritas y analizadas por Ciciarelli (1989). Las poblaciones de C. variegatifolia se han visitado sistemáticamente en estudios de campo realizados en el curso de los doce años recientes y, lejos de disminuir en extensión, se ha registrado un aumento en la superficie de terreno cubierta por las mismas.
Canna ascendens y otras especies, bonaerenses y argentinas
Ciciarelli (1989) citó 6 especies de Canna para la Argentina: C. coccinea Mill., C. compacta Roscoe, C. paniculata Ruiz & Pav. (como C. confusa J.W. Richardson & L.B. Sm.), C. glauca L., C. indica L. y C. variegatifolia Ciciarelli. De acuerdo con esa autora, C. compacta crece en las selvas del NO y C. paniculata en Misiones, mientras que C. coccinea, C. glauca, C. indica y C. variegatifolia crecen silvestres desde Formosa y Misiones hasta Santa Fe y Buenos Aires, aunque algunas, como C. variegatifolia se han encontrado cultivadas como ornamentales en Corrientes. Tanaka (2001) describió 4 especies para el NE de la Argentina: C. amabilis T. Koyama & Nb.Tanaka, C. plurituberosa T. Koyama & Nb.Tanaka y C. stenantha Nb. Tanaka (las 3 para Misiones) y C. jacobiniiflora T. Koyama & Nobuyuki Tanaka, para Corrientes. Para la provincia de Buenos Aires, Fabris (1968) citó dos especies y una variedad: C. coccinea, C. glauca var. glauca y C. glauca var. rubrolutea Hook. Ciciarelli (1989) sinonimizó C. glauca var. rubrolutea con la variedad tipo y reconoció a C. coccinea y C. glauca como especies bien caracterizadas. En la provincia de Buenos Aires están presentes hasta el momento C.ascendens, C.coccinea, C. glauca, C.indica y C. variegatifolia.
Las plantas de las especies bonaerenses difieren en el tamaño: las de C. glauca son comparativamente pequeñas, de hasta 1,5 m de altura en ciertas comunidades palustres; las de C. coccinea y C. indica son siempre más altas, de hasta 3 m, similares a las de C. ascendens, y la más grandes corresponden a C. variegatifolia que puede incluso superar los 4 m de altura. Es posible que los tamaños de algunas especies varíen en relación con aspectos ecológicos locales, como es el caso de C. glauca, cuyas plantas tienen alturas variables y alcanzan los 3 m en ejemplares del S de Estados Unidos (Estado de Florida).
Los rizomas de todas las especies son monopodiales, con excepción de los de C. coccinea, que son simpodiales y presentan tuberibulbos.
En cuanto a la ecología, C. glauca se destaca porque es anfibia, crece en las orillas de pequeños lagos, lagunas y riachos, con las raíces permanentemente sumergidas y siempre relacionada con agua corriente abundante. Las restantes especies son terrestres y crecen en suelos húmicos, eventualmente removidos o perturbados. Todas, incluyendo C. ascendens, viven tanto en áreas urbanas como baldíos y bordes de terrenos libres como en sitios abiertos, menos perturbados, pero siempre en suelos ricos, formando comunidades muy abundantes. Es posible que C. ascendens se encuentre en otras zonas de la cuenca del Paraná, favorecida por la similitud de ambientes y eventualmente, por su capacidad para crecer en áreas urbanas.
Canna indica es, sin duda, la especie más difundida del género y se caracteriza por una plasticidad morfológica notable, sobretodo en lo referente a ciertos caracteres florales, como el número de estaminodios y el patrón de distribucion de los colores (colorado y amarillo) en estaminodios y labelos, algo que llevó a Kranzlin (1912), entre otros autores, a describir numerosas especies distintas. Más recientemente Tanaka (2001) reconoció solamente cuatro variedades de la especie, sobre la base del color de las flores y la distribución geografica. Maas (1988) considera a C. indica de manera muy amplia y señala la existencia de un "complejo C. indica", integrado por formas cuyas variaciones carecerían de importancia taxonómica. En ese complejo, Maas (com. pers.) incluye a C. amabilis, C. compacta, C. coccinea, C. jacobiniflora, C. plurituberosa, C. stenantha y C. variegatifolia. Aquí se participa más bien de la idea más circunscripta de la especie propuesta por Tanaka (2001), aunque las variedades aceptadas por ese autor para C. indica no se han encontrado en la Argentina.
En el caso de las especies argentinas, la combinación de ciertos caracteres exomorfológicos foliares (contorno, relación largo ancho, ápice, base e indumento de las hojas), la ecología y la distribución geográfica, sugieren grupos de especies afines. Otros rasgos foliares (color, consistencia, espesor del mesofilo) y florales (la combinación de, contorno, dirección y color, entero o maculado, del labelo) tendrían valor diagnóstico específico (Ciciarelli, datos no publicados). Los caracteres más destacables de las especies argentinas se han resumido en la Tabla 1, en la que no se ha incluido a C. plurituberosa por no haber contado con material confiable para su análisis. Canna plurituberosa tiene muchas semejanzas con C. coccinea (Tanaka, 2001) y eventualmente podrían ser coespecíficas. Maas (1988) considera a C. coccinea coespecífica con C. indica mientras que Ciciarelli (1989) y Tanaka (2001) las tratan como taxones distintos. Canna coccinea se diferencia de C. indica, entre otros rasgos, por el color púrpura de tallos, vainas, ejes, márgenes y vena primaria, debido a la presencia de antocianas, las hojas más pequeñas con ápices obtusos con acumen y las flores carmesíes con labelos concolores. Estos caracteres, además del contorno foliar, también permiten distinguir a C. coccinea de C. ascendens (Tabla 1).
Tabla 1. Caracteres foliares y florales en especies argentinas de Canna
Clave de las especies bonaerenses de Canna
1. Hojas linear lanceoladas, lanceoladas u ovado lanceoladas, plantas terrestres o anfibias, inflorescencias reducidas o ramificadas, flores grandes (hasta 15 cm) o más pequeñas (hasta 8-9 cm); estaminodios 3-4. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .2
1. Hojas ovadas angostas, elípticas u oblongas, plantas siempre terrestres, inflorescencias reducidas, 0-2 paracladios, flores siempre más bien pequeñas (hasta 6-7 cm); estaminodios 2 (3). . . . . . . . . . . . . . . . . . . . . . .4
2 (1). Plantas anfibias, hojas linear lanceoladas glaucas, inflorescencias reducidas, 0-1 paracladios, flores grandes de color amarillo limón, 10-12 (15) cm, labelo reflexo, siempre concolor, amarillo. . . . . . . . . . .C. glauca
2.Plantas terrestres, hojas ovado-lanceoladas a lanceoladas, verdes o variegadas, inflorescencias reducidas o ramificadas, flores más pequeñas (hasta de 8-9 cm), rosadas o rosado-anaranjadas, siempre con 3 estaminodios, labelos oblanceolados, reflexos o circinados a girados, concolores o maculados. . . . . . . . . . . . . . . . . . .3
3 (2).Hojas verdes ascendentes, comúnmente plegadas a lo largo de la vena media, inflorescencias reducidas, 0- 2 paracladios, flores anaranjadas, labelo reflexo, adaxialmente maculado. . . . . . . . . . . . . . . . .C. ascendens
3. Hojas variegadas de blanco y amarillo pálido, horizontales a ascendentes, planas, inflorescencias ramificadas, con hasta 4 paracladios, flores rosado anaranjadas, labelo circinado o girado, concolor. . .C. variegatifolia
4(1). Hojas ovadas angostas a casi oblongas, discolores, verdes con vena media y márgenes de color púrpura, flores concolores carmesíes, labelo concolor. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. coccinea
4. Hojas elípticas, concolores pero con epifilos verdeamarilentos e hipofilos glaucos o verde-grisáceos, flores discolores, coloradas y amarillas, labelo generalmente colorado maculado de amarillo. . . . . . . . . . . C. indica
BIBLIOGRAFÍA
1. Ciciarelli, M. M. 1989. Las Cannaceae Link argentinas. Tesis Doctoral. Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata. [ Links ]
2. Ciciarelli, M. M. 1995. Canna variegatifolia Ciciarelli sp. nov. (Cannaceae-Zingiberales). Revista Mus.La Plata, Secc. Bot. 14: 333-341. [ Links ]
3. Chase, M. W.; D. E. Soltis; R.G.Olmstead, D. Morgan, D.H. Les, B. D. Mishler, M. R. Duvall, R. Price, H. G. Hills, Y. Qui, A. Kron, J. H. Retting, E. Conti, J. D. Palmer, J. R. Manhart, K. J. Sytsma, H. Michaels, W. J. Kress, K.G. Karol, W.D. Clark, M. Hedren, B.S. Gaut, R. K. Jansen, K. Kim, C. F.Wimpee, J. F. Smith, G. R. Furnier, S. H. Strauss, Q. Xiang, G. M. Plunkett, P. S. Soltis, S. E. Swensen, S. E, Williams, P. A. Gadek, C. J. Quinn, L. E. Eguiarte, E. Golenbrg, G. H. Jr. Learn, S. Graham, C. H. Barret, S. Dayanandan & V. A. Albert.1993. Phylogenetics of the seed plants: an analysis of nucleotide sequences from the plastid gene rbcL. Ann. Missouri Bot. Gard. 80: 528-580. [ Links ]
4. Dahlgren, R. M. & F. N. Rassmusen. 1983. Monocotyledon evolution: characters and phylogenetic estimation. Evol.Biol. 16: 255-395. [ Links ]
5. Fabris, H. 1968.Cannaceae, en A. L. Cabrera (ed.), Flora de la Provincia de Buenos Aires. Colecc. Ci. Inst. Nac.Tecnol Agropecu. 4(1): 568-571. [ Links ]
6. Hickey, L. J.; A. Ash, B. Ellis, P. Wilf, K. Johnson &S. Wing 1999. Manual of leaf Architecture. Morphological description and categorization of dicotyledonous and net-veined monocotyledonous angiosperms by Leaf Architecture Working Group. Washington D.C. Smithsonian Institution Press. [ Links ]
7. Holmgren, P. K.; N. H.Holmgren & L. Barnett 1990. Index Herbariorum, Part I: Herbaria of the World. New York: New York Botanical Garden. [ Links ]
8. Judd, S.; C.S.Campbell, E. A. Kellog, P.F. Stevens & M. J. Donoghue. 2002. Plant Systematics. A phylogenetic approach. Massachusetts, Sunderland: Sinauer. [ Links ]
9. Kranzlin, F. 1912. Cannaceae. En A.Engler (ed.), Das Pflanzenreich, IV. 47 (Heft 55): 1-77. Leipzig: W. Engelmann. [ Links ]
10. Kress, J.1990.The phylogeny and classification of the Zingiberales. Ann.Missouri Bot.Gard. 77: 698-721. [ Links ]
11. Kress, W. J. 1995. Phylogeny of the Zingiberanae: Morphology and molecules, in P. J. Rudall, P. J. Cribb, D. F. Cutler & C. J. Humphries (eds.), Monocotyledons: Systematics and evolution. pp. 443-460. Kew: Royal Botanic Gardens. [ Links ]
12. Kress, W. J. & D. E. Stone.1982. Nature of the sporoderm in monocotyledons, with special reference to the pollen grains in Canna and Heliconia. Grana 21:129-148. [ Links ]
13. Kress, W. J. & L. Prince, 2000. Cannaceae, in Flora of North America Editorial Committee, Flora of North America 22: 310-314. New York. Oxford University Press [ Links ]
14. Kress, W. J.; L. M. Prince, W. J. Hahn & E. A. Zimmer. 2001. Unraveling the evolutionary radiation of the families of the Zingiberales using mophological and molecular evidence. Syst. Biol. 50: 926-944. [ Links ]
15. Maas, P. J. M. & H. Mass. 1988. Cannaceae, in G.Harling et al. (eds.), 1973 + Flora of Ecuador, 5660 + volsnos. Göteborg. Vol. 32: 1-9. [ Links ]
16. Nair, P. K. K. 1960. Pollen grains of cultivated plants, I: Canna. J. Indian Bot. Soc. 39: 373-381. [ Links ]
17. Nair, P. K. K. 1962. Pollen grains of Indian Plants. I. Natl. Bot. Gard. Bull. 53: 1-35. [ Links ]
18. Potgieter, M. J. & A. E. Van Wyk. 1962. Intercellular pectic protuberances in plants: their structure and taxonomic significance. Bot. Bull. Acad. Sin. 33: 295-316. [ Links ]
19. Prabhakar, M. 2003. Structure, Delimitation, Nomenclature and Classification of Stomata. Acta Bot .Sin. 46: 242- 252. [ Links ]
20. Prada, C. & C. H. Rolleri. 2005. A new species of Isoetes L. (Isoetaceae) with a study of microphyll intercellular pectic protuberances and their potential taxonomic value. Bot. J. Linn. Soc. 147: 213-228. [ Links ]
21. Rolleri, C. H.; M. C. Lavalle, A. Mengascini & M. Rodríguez. 1999. Protuberancias intercelulares pécticas en especies de Angiopteris Hoffm. y otras Marattiales. Physis, C. 57(132- 133): 1-11. [ Links ]
22. Segeren, W. & P. J. M. Maas. 1971. The genus Canna in Northern South America. Acta Bot. Neerl. 20: 663-680. [ Links ]
23. Smith, J.; W. J Kress & E. A. Zimmer. 1993. Phylogenetic analysis of the Zingiberales based on rbcL sequences. Ann. Missouri Bot. Gard. 80: 50-66. [ Links ]
24. Stern, W. T. 1996. Botanical Latin. Portland, Oregon: Timber Press. [ Links ]
25. Stevenson, D. W. & H. Loconte. 1995. Cladisctic analysis of the monocot families, in P. J. Rudall, P. J. Cribb, D. F. Cutler & C. J. Humphries, (eds.), Monocotyledons: Systematics and evolution. Kew: Royal Botanic Gardens. [ Links ]
26. Tanaka, N. 2001.Taxonomic revision of the family Cannaceae in the New World and Asia. Makinoa 1: 1-74. [ Links ]
27. Tomlinson, P. B. 1962. Phylogeny of the Scitaminae: Morphological and anatomical considerations. Evolution 16: 192-213. [ Links ]
28. Tomlinson, P. B. 1969. Commelinales-Zingiberales, in C. R. Metcalfe (ed.), Anatomy of the Monocotyledons, vol. 3. Oxford: Clarendon Press. [ Links ]
29. Troll, W. 1964-69. Die Infloreszenzen. Typologie und Stellung in Aufbau des vegetationsköroers. Stuttgart: G. Fisher. [ Links ]
30. Weberling, F. 1965.Tipology of the inflorescences. J. Linn.Bot. Soc. 63: 235-246. [ Links ]
31. Weberling, F. 1985. Aspectos modernos de la morfología de las inflorescencias. Bol. Soc. Argent. Bot. 24: 1-28. [ Links ]
32. Weberling, F.; U. Müller-Doblies, D. Müller-Doblies & G. Rua. 1997. Hacia una terminología descriptiva y morfológico comparativa para inflorescencias complejas. Bol. Soc. Argent. Bot. 32: 171-184. [ Links ]
33. Wilkinson, H. 1979. The plant cuticle (mainly leaf), en C. R. Metcalfe & L. Chalk (eds.). The anatomy of the Dicotyledons, pp. 97-165. Oxford: Clarendon Press. [ Links ]

