










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkDarwiniana, nueva serie
versión impresa ISSN 0011-6793
Darwiniana vol.46 no.2 San Isidro ago./dic. 2008
ARQUEOBOTÁNICA Y ETNOBOTÁNICA
Caracterización cuantitativa de productos intermedios y residuos derivados de alimentos del Algarrobo (Prosopis flexuosa y P. chilensis, Fabaceae): aproximación experimental aplicada a restos arqueobotánicos desecados
Aylen Capparelli
Departamento Científico de Arqueología, Museo de Ciencias Naturales de La Plata. Paseo del Bosque s/n, 1900 La Plata, Argentina; aylencapparelli@fcnym.unlp.edu.ar
Original recibido el 24 de junio de 2008;
aceptado el 13 de diciembre de 2008.
Resumen. Mediante una aproximación experimental se caracterizan los atributos macromorfológicos cuantitativos de productos intermedios y residuales derivados de Prosopis chilensis y P. flexuosa (Algarrobo blanco y Algarrobo negro respectivamente) que potencialmente podrían llegar a formar parte del registro arqueobotánico. Se provee descripción morfoanatómica de la vaina y la semilla de las especies tratadas. Se elaboró harina no refinada y refinada, añapa, aloja y arrope, siguiendo técnicas tradicionalmente utilizadas en el Valle de Hualfín, Catamarca, Argentina, las cuales fueron registradas por la autora en trabajos previos. Se concluye que el análisis cuantitativo de restos macrobotánicos de Prosopis, en conjunto con el cualitativo, permite la identificación de diferentes etapas de procesamiento del Algarrobo. Para ello resulta esencial la distinción entre las dos especies. La proporción de diferentes categorías de semillas y endocarpos es útil para distinguir la harina refinada de la no refinada. Esta última podría indicar la manufactura de patay, ulpo o aloja. Los residuos de la añapa y aloja se caracterizan por presentar semillas con testa plegada, enrollada o levantada, o carecer de ella, y sus cantidades se encuentran disminuidas o aumentadas con respecto a la cantidad inicial de harina utilizada dependiendo de si las semillas que se recuperan son enteras o fragmentadas. Los residuos del arrope se identifican por poseer grandes piezas de epicarpo y porque todos los endocarpos correspondientes a la cantidad de artejos utilizados inicialmente en su preparación se encuentran presentes. Dichos endocarpos se encuentran cerrados, y excepto en el caso de los residuos de arrope, se considera que la mayoría de las asociaciones arqueológicas de restos de Prosopis representa una proporción muy baja del volumen de materia que le dio origen en su contexto dinámico del pasado.
Palabras clave. Argentina; Arqueobotánica; Prosopis; Valle de Hualfín.
Abstract. Quantitative features of intermediate products and residues derived from the Algarrobo as a food (Prosopis flexuosa and P. chilensis, Fabaceae): experimental approach applied to dehydrated macroremains.
Experimental approach was used to characterize quantitative macromorphological attributes of intermediate and final food products and residues that might have reached the archaeological record as macrobotanical remains of Prosopis chilensis and P. flexuosa (white and black "Algarrobo" respectively). General morphology of beans and seeds of these species are provided. Unrefined and refined flour, añapa, aloja and arrope were elaborated following traditional techniques registered by the author along the Hualfín Valley, Catamarca Province, Argentina, during previous works. It was concluded that quantitative analysis of macrobotanical remains of Prosopis, together with qualitative features, allowed the identification of different stages in the preparation and processing of Algarrobo. Distinguishing between black and white algarroba is essential to interpret Prosopis macroremains. The proportion of different categories of seeds and endocarps may help to distinguish the production of unrefined and refined flour. The last one may indicate the elaboration of patay, ulpo or aloja. Añapa and aloja residues are characterized mainly by rolled, folded or loosening of testa seed and their quantities are diminished or increased with respect to the initial amount of flour used depending on entire or fragmented seeds are being considered. The arrope residues are identified by the presence of closed endocarps, which are fully recovered after processing in their totality, and large pieces of epicarp. Except for the arrope residues, the majority of the Prosopis archaeological assemblage is considered to represent a very low proportion of the ancient processed volume of each dynamic context.
Keywords. Archaeobotany; Argentina; Prosopis; Hualfín Valley.
INTRODUCCIÓN
El género Prosopis pertenece a la subfamilia Mimosoidea y posee 44 especies reconocidas. Cuatro de ellas se distribuyen en el viejo mundo y África, mientras que las cuarenta restantes a lo largo del continente americano, desde el SO de Norte América hasta la Patagonia. Argentina parece ser el centro de origen primario de este género mientras que México un centro de origen secundario. En los Estados Unidos y México se distribuyen 8 especies, mientras que en América del Sur 32 especies, 28 de las cuales se hallan en Argentina. De éstas últimas, 13 especies son endémicas de dicho país (Burkart, 1940, 1952, 1976a, b). El fruto de este género consiste en una legumbre drupácea indehiscente clasificada por Burkart (1952: 126, 1976a, b) como lomento drupáceo. El fruto de varias especies de Prosopis es empleado en la elaboración de diversos alimentos en distintas partes del mundo, tales como África: P. africana (Guill., Perr. & Rich.) Taubert in Engler & Prantl. (Sanni et al. 1993), India (Arya, en Harris et al., 1998), México y SO de U.S.A.: P. glandulosa Torrey, P. velutina Wooton, P. laevigata (Humboldt & Bonpland ex Willdenow) M.C. Johnston, P. juliflora (Swartz) DC (Burkart, 1976b; Felger, 1977), Chile, Perú, Bolivia y Paraguay: P. chilensis (Molina) Stuntz emend. Burkart, P. pallida (Humboldt & Bonpland ex Willdenow) H.B.K. (Latcham, 1936; Felker, 1979; Beresford-Jones, 2004; Horkheimer, 2004), y Argentina: P. alba Grisebach, P. nigra (Grisebach) Hieronymus, P. flexuosa DC, P. chilensis, P. ruscifolia Grisebach, P. vinalillo Stuckert, P. elata (Burkart) Burkart, P . torquata (Cavanilles. ex Lagasca) DC, P. hassleri Harms., P. ferox Gris. (Outes, 1917; Sánchez de Oviedo, 1936; Di Lullio, 1943, 1944; Métraux, 1944; Canals Frau, 1946; Rusconi, 1958; Cáceres Freyre, 1961, 1962; Martínez Crovetto, 1964, 1968, 1987; Ruiz Leal, 1970; D'Antoni, 1975; Burkart 1976b, D'Antoni & Solbrig, 1977; Villafuerte, 1984; Roig, 1993; Filipov, 1996; Scarpa, 1999; Arenas 2003). Entre los productos elaborados a partir de los mismos se incluyen: panes; jaleas, bebidas con y sin alcohol, vainas simplemente hervidas y harina tostada o desleída en agua. El empleo de este fruto en la elaboración de alimentos posee una profunda raíz temporal. En el Viejo Mundo, de donde se han recuperado semillas datadas en 11500 años AP (sitio Tell Abu Hureyra; Moore et al., 2000), su uso prehistórico como alimento ha sido recientemente confirmado a través del análisis químico de residuos de una mano de moler proveniente de la cueva Hayonim, en el sur del Levante, en la que se identificaron restos de azúcares característicos del género mencionado (McLaren & Capparelli, 2004). En el Nuevo Mundo, mientras que un sólo registro arqueológico confirma con certeza su ingesta (Holden & Núñez, 1993), existen muchas evidencias tanto ethnohistóricas que avalan la importancia de la utilización del género en momentos de contacto (Bell & Castetter en Felger, 1977; Noli, 1998, 1999; Arana, 1999, entre otras), como arqueológicas (Towle, 1961; Felger 1977; Beresford Jones, 2004), a las que se les puede atribuir un probable uso alimenticio. Estas evidencias arqueológicas son abundantes en Argentina (para una recopilación detallada véase Giovannetti et al., 2008).
En el Viejo Mundo, algunos arqueobotánicos han desarrollado exitosos modelos interpretativos sobre el procesamiento de cereales basados en la observación etnográfica y etnobotánica (e.g. Hillman, 1984; Jones, 1984) quienes siguiendo los preceptos de la teoría de rango medio (Schiffer, 1972, 1976; Binford, 1981) ayudaron a interpretar contextos dinámicos del pasado a partir de la distribución estática de los restos arqueológicos recuperados. El presente trabajo contribuye a dichos estudios en lo que se refiere a la interpretación del procesamiento de Prosopis.
Los macrorrestos de Prosopis son particularmente difíciles de interpretar más allá del obvio reconocimiento de su uso o no uso, dado que su procesamiento es complejo y puede dar lugar a numerosos productos intermedios, finales y residuos (véase Capparelli, 2007). En el Valle de Hualfín, por ejemplo, su harina es o era usada para hacer una especie de pan -patay-, una bebida no alcohólica -añapa-, otra bebida alcohólica -aloja- y una especie de lejía llamada ulpo que los pastores tomaban mientras alimentaban a su ganado. Las vainas son también empleadas para hacer el arrope, similar a una jalea. Estos productos finales, excepto el patay como se explicará en los resultados, comparten la particularidad de que, por ser bebidas, lejías o jaleas, no poseen visibilidad arqueológica como macrorrestos, por lo cual su manufactura debe ser evaluada en función de los productos o residuos generados por la secuencia de las varias etapas involucradas en su procesamiento. Es a lo largo de dichas etapas que se producir diferentes complejos de macrorrestos (Fig. 1) capaces de brindar información sobre el producto final elaborado y posibles de ser diferenciados arqueológicamente por sus cualidades (Capparelli, sine data).

Fig. 1. Esquema que muestra los distintos productos intermedios y finales derivados del procesamiento del Algarrobo, sus principales etapas de elaboración y los diferentes tipos de residuos que generan (modificada de Capparelli, sine data).
La antigüedad del uso prehispánico de Prosopis, la complejidad de su procesamiento y las particularidades de su visibilidad arqueológica, hacen necesario el desarrollo de una aproximación arqueobotánica que permita interpretar adecuadamente sus macrorrestos. Sólo de esta manera se podrá comprender y explicar la importancia de este género para las sociedades desde una perspectiva diacrónica.
En un trabajo previo donde se combinaron las aproximaciones etnobotánica y arqueobotánica experimental (Capparelli, sine data) se analizó el manejo espacial durante las actividades relacionadas con la producción de alimentos de Prosopis, como por ejemplo áreas de procesamiento y descarte en unidades domésticas tradicionales del Valle de Hualfín, Catamarca. Se registraron los artefactos utilizados durante dichas actividades y los posibles puntos de contacto de los especímenes de Prosopis con el fuego, con el objeto de establecer los rasgos de origen de potenciales restos carbonizados. En el mismo trabajo se caracterizaron cualitativamente los atributos macromorfológicos de los productos intermedios, finales y residuales que pueden haber formado parte del registro arqueológico como restos macrobotánicos y se establecieron las categorías morfológicas cualitativas de los órganos o partes de órganos más diagnósticos y posibles de ser cuantificados. Se concluyó que la distinción entre Prosopis flexuosa (Algarrobo negro) y P. chilensis (Algarrobo blanco), las dos especies utilizadas en el valle de Hualfín, es esencial para interpretar los macrorrestos de este género en dicha área. Cabe mencionar que los nombres vulgares de Algarrobo negro (AN) y Algarrobo blanco (AB) se utilizan aquí arbitrariamente ya que son los más difundidos en la literatura de habla hispana. Otras denominaciones vernáculas de estas especies y una mayor discusión sobre su terminología se encuentran en Capparelli (2007). La distinción entre Prosopis flexuosa y P. chilensis es importante no sólo porque puede brindar información acerca de la preferencia de los habitantes en el pasado (el Algarrobo blanco en dicho valle es más preciado que el negro por ser el primero más suave y dulce y el segundo más áspero), sino porque bajo el mismo procesamiento cada una de estas especies produce productos intermedios, finales y residuos de diferentes calidades, muchos de los cuales pueden llegar a formar parte del registro arqueológico y por lo tanto, necesariamente deben ser analizados si se pretende distinguir claramente entre la elaboración de un producto y de otro. De los órganos vegetales representados en los productos intermedios y residuales, la morfología de las semillas resultó ser uno de los caracteres más diagnósticos. En este sentido, el Algarrobo blanco brinda mayor información que el negro ya que libera mayor cantidad de semillas al ser procesado. La presencia/ausencia tanto de semillas fragmentadas como con testa fisurada son caracteres que, para el caso del Algarrobo blanco, permiten diagnosticar la manufactura de productos intermedios, e.g. harinas, destinados a la elaboración de añapa, aloja, patay o ulpo. No obstante, es aconsejable que no sólo las semillas sino también los endocarpos sean tenidos en cuenta para este tipo de análisis. Por ejemplo, en el caso del Algarrobo negro los endocarpos fisurados o fragmentados son indicadores de la manufactura de dichas harinas mientras que la mayoría de las semillas quedan en el interior de los mismos. Por otra parte, y a partir del análisis de residuos, la presencia de semillas con la testa enrollada, plegada, o directamente sin testa indicaría la elaboración de añapa o aloja. Sin embargo, la recuperación de semillas con estas características no permite rechazar la hipótesis de que se haya preparado patay o ulpo, ya que la fracción gruesa residual de harina derivada de estos dos últimos productos puede haber sido utilizada para elaborar las bebidas antes mencionadas, privando al patay o al ulpo de dejar residuos propios de su elaboración y disminuyendo así su visibilidad arqueológica. Contrariamente a lo que sucede con el patay y el ulpo, la manufactura de arrope puede distinguirse por la presencia de residuos como largas piezas de epicarpo y endocarpos cerrados con parches de mesocarpo hervido en su superficie.
Tal como se mencionó anteriormente, la presencia/ausencia tanto de semillas con testa entera, fisurada, plegada o enrollada y de semillas fragmentadas, como de endocarpos enteros, fisurados o fragmentados son caracteres que, entre otros, permiten diagnosticar la manufactura de productos finales e intermedios, tales como harinas y otros destinados a un proceso de elaboración posterior. Se piensa que la proporción numérica de dichos órganos o partes de órganos puede constituir un carácter diagnóstico importante, que complemente al cualitativo, para identificar la manufactura de dichos productos y evaluar groseramente qué cantidad de la utilizada en el contexto dinámico representan las recuperadas en el contexto arqueológico. Por tal motivo, el presente trabajo tiene por objetivo principal analizar cuantitativamente la presencia de órganos o partes de órganos en harinas derivadas de Prosopis chilensis y P. flexuosa, y en residuos derivados de la preparación de patay, ulpo, añapa, aloja y arrope, obtenidos experimentalmente. Al respecto, se pone especial énfasis en registrar los cambios dimensionales acontecidos luego de la elaboración de aquellos productos que requieran remojo o cocción (añapa, aloja y arrope). También se pretende analizar los tiempos de molienda necesarios para obtener harina apta para la elaboración de patay o ulpo y su potencial rendimiento con respecto a la fracción gruesa residual después de realizada la molienda. Para llevar a cabo los objetivos mencionados es necesario comprender cabalmente la morfología interna de la vaina y el fruto, por lo cual primeramente se describen las estructuras que conforman ambos órganos.
MATERIALES Y MÉTODOS
Para realizar la descripción morfoanatómica interna de la vaina y la semilla de Prosopis chilensis y P. flexuosa se realizaron cortes transversales (CT) de las partes medias de ambos órganos, que fueron observados a través de microscopio óptico de transparencia marca Leica.
Luego se llevó a cabo el procesamiento experimental de las dos especies mencionadas, ambas nativas del Valle de Hualfín (Fig. 2), empleadas actualmente para la elaboración de productos alimenticios. Este conocimiento proviene de trabajos etnobotánicos previos realizados en las localidades de Cerro negro, El Shincal, Puesto Zapata, Los Colorados, La Puntilla, Londres de Quimivil, Quillay (La Ciénaga), Hualfín, Jacipunco y Corral Quemado (Capparelli, 2007).

Fig. 2. Localización de los sitios en los que se obtuvo la información etnobotánica utilizada y se coleccionaron las distintas vainas de Algarrobo blanco y Algarrobo negro (tomado de Capparelli, 2007).
El macroclima actual del valle es cálido, continental, árido (Morello, 1958:32). Las precipitaciones anuales, que son principalmente estivales, aumentan de S a N hasta la Sierra de Belén (de 100 mm en Cerro Negro a 374-411 mm en Belén); mientras que disminuyen nuevamente al N de esta sierra a menos de 200 mm (Ardissone, 1961).
Los especímenes de referencia de los taxones tratados fueron depositados en el Herbario del Departamento de Plantas Vasculares del Museo de Ciencias Naturales de La Plata (LP) como Capparelli 7 [LP] y Capparelli 6 [LP] respectivamente.
Se elaboró añapa, patay, aloja, ulpo y arrope utilizando los métodos tradicionales empleados actualmente en el área mencionada, con la excepción del tipo de tamizado, ya que en lugar de una tela de arpillera se empleó una zaranda metálica de 1 mm de abertura con el objeto de hacer más sencillo el procesamiento. Aunque es común que la gente mezcle diferentes proporciones de una y otra especie de Algarrobo, en el presente trabajo ambas especies se molieron por separado con el fin de distinguir claramente el efecto de la molienda en una u otra.
El número de artejos contenido en las vainas y el peso de cada muestra fueron registrados previamente a la molienda. Se procesaron diferentes muestras según el caso. En primer lugar se obtuvieron dos tipos de harinas: no refinada y refinada. Se consideró harina no refinada a aquella en la cual los artejos estuvieron casi totalmente desarticulados y comenzó a aparecer la fracción de harina <1mm, aunque aún se observó mesocarpo adherido al epicarpo y a los endocarpos. Se consideró harina refinada a aquella en la cual no quedan remanentes de mesocarpo adherido al epicarpo o endocarpo. Esta categorización fue propuesta a partir de la observación por parte del autor de estos dos tipos de harina en el valle, en donde aún son elaboradas por los pobladores actuales. La primera generalmente es almacenada y utilizada posteriormente para la elaboración de harina refinada o de añapa. La harina no refinada se obtuvo luego de 90 segundos de molienda en un mortero de piedra con una mano de piedra de 770 g, en muestras de vainas que oscilaron entre 5,9 y 6,8 g; mientras que la harina refinada se obtuvo luego de 50 segundos adicionales de molienda.
La fracción fina (<1mm) tanto de la harina no refinada como de la refinada fue pesada a los 90 y a los 140 segundos de la molienda. Su peso fue utilizado para graficar el pasaje de harina no refinada a refinada a lo largo del tiempo de molienda y el rendimiento de las vainas con respecto a la producción de dicha fracción <1mm. Con el fin de testear si este rendimiento puede ser homogéneo para todo el valle, se seleccionaron fragmentos de vainas de ambas especies con cantidades extremas de mesocarpo apreciables a simple vista. De los mismos se calculó la proporción de este tejido en relación con el peso total del fragmento. Para ello se pesaron fragmentos de vaina en un total de tres para P. chilensis y dos para P. flexuosa. Luego se extrajeron de cada uno de ellos los tejidos epicárpico, mesocárpico y el complejo endocarpo-semilla, mediante el pelado de los frutos y el raspado del mesocarpo con una trincheta. Cada parte del fruto fue pesada individualmente. Al mismo tiempo, se calculó el peso y el número de artejos promedio de 6 vainas de P. chilensis y 9 vainas de P. flexuosa provenientes de distintas localidades a lo largo del valle.
De la fracción gruesa (>1 mm) de la harina no refinada y de la refinada se analizaron los fragmentos de tamaño comprendidos entre 1 y 2 mm y aquellos mayores a 2 mm. Los primeros, identificables a microscopio estereoscópico, fueron evaluados sólo cualitativamente. Mientras que los segundos, distinguibles a ojo desnudo caracterizados por su morfología cualitativa externa (Capparelli, sine data), fueron contados antes y después de cada estadio de molienda. A partir de estos datos se calcularon los porcentajes relativos de cada tipo de órgano en el total de la muestra y se calculó el modo en que los mismos disminuyeron o aumentaron a medida que avanzó la molienda. Este procedimiento se realizó también en el caso de los residuos de añapa-aloja y de arrope. Se consideró que esta categoría de restos (> 2 mm) conformada mayoritariamente por vainas en distintos estados de fragmentación, endocarpos y semillas es la más conveniente para definir el tipo de residuo o el grado de refinamiento de la harina por dos razones. En primer lugar, porque debido a su tamaño son fácilmente distinguibles a ojo desnudo, por lo que el balance del costo-beneficio en cuanto al tiempo consumido para su identificación es positivo. En segundo lugar, debido a que este tipo de restos es el que se ha recuperado previamente de varios sitios a lo largo de la República Argentina (véase Giovannetti et al., 2008), antes de que el interés estuviera puesto en la arqueobotánica en particular y de que la misma se consolidara como disciplina en este país. Debido a esto último, los restos mencionados seguramente no constituyen la totalidad de los que se encuentraban presentes en los sitios en donde se recuperaron, sino sólo aquéllos más conspicuos de cada contexto. Partiendo de esta suposición, el análisis de la categoría de restos > 2mm haría posible las comparaciones entre los resultados experimentales y los restos de Algarrobo registrados para otros sitios arqueológicos. No obstante, para realizar dichas comparaciones de una manera más acertada aún restaría salvar las distancias correspondientes a los métodos de recuperación empleados y a la falta de sistematización de los muestreos.
Para preparar la añapa, la fracción gruesa (>1mm) de una de las muestras de harina refinada fue sumergida en agua a una proporción de 1:4 durante 20 horas. Tiempos menores al mencionado no produjeron modificación alguna en los diferentes órganos vegetales. La misma muestra se dejó en remojo luego por 10 días para preparar aloja. Como en el caso de los endocarpos no se evidenció cambio alguno en sus dimensiones ni en sus características morfológicas antes y después de los períodos de remojo, se puso mayor énfasis en evaluar el efecto de este procesamiento en las semillas. Para ello, un conjunto adicional de semillas de AB y AN se dejó en remojo durante 21 días, a lo largo de los cuales se registraron sus dimensiones (largo, ancho y espesor) en tres ocasiones: a las 20 horas, a los 10 días y a los 21 días. El período de 21 días se seleccionó para evaluar las condiciones de las semillas con posterioridad al período comprendido en la elaboración de aloja. Una última medición fue realizada posteriormente al secado de distintos conjuntos de semillas, luego de cada período de remojo, con el fin de evaluar posibles diferencias morfológicas entre semillas secadas inmediatamente después de cada período de remojo mencionado. El secado posterior a los 10 días de remojo se realizó sólo con semillas de AB, ya que los análisis previos demostraron que las diferencias entre AB y AN sometido a un período de 10 días de remojo son mínimas.
Todas las medidas se realizaron mediante calibre manual.
Para realizar el arrope se partieron groseramente a mano 9,2 g de P. flexuosa y 9,2 g de P. chilensis y se hirvieron separadamente durante 10 minutos en el doble de su volumen de agua. Luego, las vainas fueron presionadas con fuerza contra la malla de metal mediante movimientos de vaivén con el fin de extraer su jugo.
RESULTADOS
Morfología interna de la vaina y la semilla de Prosopis chilensis y P. flexuosa
P. flexuosa posee la legumbre globosa, generalmente recta y más o menos arrosariada debido a constricciones producidas entre un artejo y otro (Fig. 3A), considerando artejo a cada segmento de la vaina que contiene un receptáculo de endocarpo, mientras que Prosopis chilensis, posee la legumbre aplanada y más o menos falcada (Fig. 3A). No se observaron diferencias notorias con respecto a la estructura interna de las vainas de ambas especies. La vaina consta internamente de epicarpo, mesocarpo (parte dulce y comestible) y endocarpo (Figs. 3C-E). El endocarpo es duro y se encuentra segmentado en receptáculos cerrados uniseminados (Fig. 3F). El epicarpo consta de cuatro capas de células. La primera vista en corte transversal de vaina corresponde a una capa de células epidérmicas cuadrangulares de paredes engrosadas (Fig. 4A), con cutícula gruesa, depósitos de ceras en forma de pequeñas hojuelas en su superficie (Fig. 4B-C) y estructuras anexas como pelos tectores unicelulares, la mayoría cortos, de forma cónica, verrucosos y en ocasiones con la punta levemente ganchuda (Figs. 4D-G). La forma de las células epidérmicas, los depósitos de ceras, los estomas y los pelos observados fueron similares a los descritos para los folíolos de ambas especies por Money et al. (1977).

Fig. 3. A-B, morfología externa de las vainas. A, Prosopis flexuosa. B, Prosopis chilensis. C-D, corte transversal de vaina donde se observa la proporción de mesocarpo con respecto a los otros tejidos. C, Prosopis chilensis. D, Prosopis flexuosa. E, morfoanatomía de la vaina de P. flexuosa en corte transversal, a,h,i, hacen referencia a la ubicación topográfica de los detalles presentados en las letras respectivas de la Fig. 4. F, segmentación de los endocarpos en P. flexuosa. Abreviaturas: endo, endocarpo; epi, epicarpo; escl tr, esclereidas transversales; ext, externo; meso, meso-carpo; int, interno; Hfv, haces fibrovasculares. La escala de A vale para B y F; la de C vale para D.

Fig. 4. A, morfología del epicarpo de Prosopis chilensis. B-C, disposición de las ceras. B, en corte transversal; C, en vista superficial. D-G, pelos tectores. H, mesocarpo externo. I, mesocarpo interno y endocarpo. Abreviaturas: cel meso, célula de mesocarpo; cer, ceras; endo, endocarpo; ep, epidermis; est, estoma; hip, hipodermis. La escala de A vale para toda la figura.
La hipodermis está constituida por tres capas alternadas de células alargadas y cuadrangulares, colenquimatosas (Fig. 4A). El mesocarpo (Fig. 4H) está conformado por una ancha franja de células parenquimáticas de paredes muy delgadas, más o menos isodiamétricas, que se agrandan hacia el interior. Poseen contenidos parduzcos que pueden ocupar toda la célula o estar colapsados. También se ha observado la presencia de gránulos de almidón (véase Giovannetti et al., 2008). En la porción exterior del mesocarpo se encuentran haces fibrovasculares unidos por células esclerenquimáticas alargadas transversalmente (Fig. 3E). El endocarpo (Fig. 4I) está formado por células esclerenquimáticas que se reducen en tamaño hacia la línea de sutura de las dos valvas que conforman el mismo.
La semilla (Fig. 5A) de Prosopis chilensis y P. flexuosa en CT consta de epidermis externa, subepidermis, parénquina esponjoso, endosperma o albumen y embrión. Entre el parénquima esponjoso y el endosperma debiera existir una epidermis interna, que según Winton & Winton (1935: 296) no es visible en la mayoría de las especies de la familia Fabaceae. Tampoco lo fue en las especies de Prosopis aquí tratadas. La epidermis externa está formada por células en empalizada o Malpighianas (cuya altura excede varias veces su ancho). En el margen se observa una capa de dichas células, mientras que en la parte media de las caras de la semilla se observan dos superior de dichas células termina en forma achacapas en P. chilensis (Fig. 5B) y una tercera capa tada, posee lumen delgado y engrosamientos que incipiente en P. flexuosa (Fig. 5C). El extremo penetran en el lumen a modo de gajos de una naranja, característicos de la familia Fabaceae en general (véase por ejemplo otras especies en Winton & Winton 1935: 295-6); mientras que su extremo inferior presenta lumen más ancho y paredes más delgadas. Se pueden producir una o más líneas lúcidas en estas capas. La subepidermis está formada por aproximadamente 5 capas de células engrosadas, de las cuales la primera y la última presentan forma de reloj de arena (Figs. 5B-C) y en las que no se ha observado la presencia de cristales. La epidermis externa y la subepidermis conforman lo que Boelcke (1946: 248) denomina testa, para las semillas de Mimosoideas y Cesalpinioideas en general. El parénquima esponjoso (Fig. 5C-D), al cual Boelcke (1946) llama tegmen, consiste en tres o cuatro capas de células de paredes muy delgadas, comprimidas. Según Boelcke (1946) la testa más el tegmen conforman el tegumento seminal que ontogenéticamente deriva del tegumento del óvulo y al cual Winton & Winton (1935) denominan espermodermo. El endosperma o albumen se halla solo en las semillas que se denominan albuminadas (Boelcke 1946: 252) y es un buen carácter para distinguir en CT semillas del género Prosopis con respecto a aquellas del género Acacia, ya que este último posee semillas exalbuminadas, es decir carentes de albumen. El albumen consiste en varias capas de células de paredes extremadamente delgadas cuyo tamaño y largo aumenta hacia el interior de la semilla (Fig. 5D); poseen núcleos muy evidentes. El albumen se interrumpe en el margen de la semilla y aumenta de espesor en la parte media de las caras (Fig. 5A). El embrión consiste en dos cotiledones y una radícula. Los dos cotiledones se pueden observar en el CT de la parte media de la semilla (Fig. 5A) y consisten en una capa de células epidérmicas isodiamétricas que los recubre y en un mesófilo, de células de paredes delgadas (Fig. 5E), más o menos isodiamétricas hacia el exterior y en empalizada hacia el interior (Fig. 5F). En el embrión se observan gránulos de proteína.

Fig 5. Anatomía de la semilla de Prosopis chilensis y P. flexuosa. A, Estructura general; b, c, d, e, f, hacen referencia a la ubicación topográfica de los detalles que se presentan en las letras respectivas de esta misma figura. B-C, testa y tegmen. B, Prosopis chilensis. C, Prosopis flexuosa. D, tegmen y albumen de P. chilensis. E-F, cotiledón de P. chilensis. E, epidermis externa y células isodiamétricas del mesofilo. F , epidermis interna y células en empalizada del mesofilo. Abreviaturas: ab, albumen; ep, epidermis; cel emp, células en empalizada; cotil, cotiledón; ext, externa; int, interna; pa es, parénquima esponjoso (tegmen); subep, subepidermis. La escala de B vale para C, D y E.
Proporción diferencial de fracciones fina (< 1mm) y gruesa (>1mm) en las harinas no refinada y refinada
Algarrobo blanco. La harina no refinada estuvo formada por un 59,3 % de partículas mayores a 1 mm y por un 40,7 % de partículas menores a esta cifra. Esta última fracción estuvo constituida principalmente por mesocarpo. Con la obtención de la harina refinada las proporciones se inviertieron, la fracción > 1 mm constituyó el 45,8 % de la muestra mientras que la fracción < 1 mm representa el 54,2 % (Fig. 6A).

Fig. 6. A-B. Pasaje de harina no refinada a refinada en función del tiempo de molienda. A, Algarrobo blanco. B, Algarrobo negro.
Algarrobo negro. La harina no refinada estuvo formada por un 85,3 % de partículas mayores a 1 mm y por un 14,7 % de partículas menores a 1 mm. Esta última fracción estuvo constituida principalmente por mesocarpo. En la harina refinada la fracción > 1 mm constituyó el 82,4 % de la muestra mientras que aquélla < 1 mm, el 17,6 % (Fig. 6B).
Productos intermedios, productos finales y residuos. Harina no refinada
Algarrobo blanco. Las partículas mayores a 2 mm que conformaron esta harina pueden observarse en la Tabla 1. Las mismas constituyeron principalmente fragmentos de epicarpo con mesocarpo adherido (Fig. 7A), semillas (23 %) y medios endocarpos (Fig. 7B). También se observaron endocarpos cuyas dos mitades estuvieron abiertas por tres lados, denominados aquí endocarpos abiertos (1 %), los cuales han perdido su semilla (Fig. 7D). En una menor proporción se observaron endocarpos cerrados conservando su respectiva semilla, algunos de los cuales presentaron restos de mesocarpo aún adherido (Fig. 7C). Dos pares de estos endocarpos enteros permanecieron apareados. Las semillas se presentaron enteras en un 5 % con respecto al total de estructuras de la muestra (Fig. 7E), con la testa fisurada en un 9 % (Fig. 7F) y fragmentadas en un 9 % (Fig. 7G). Las partículas entre 1 y 2 mm estuvieron constituidas por fragmentos pequeños de epicarpo, epicarpo-mesocarpo y tejidos seminales. La porción <1mm estuvo constituida por mesocarpo.
Tabla 1. Número absoluto (nro. abs.) y cantidad relativa (%) de estructuras o partes de estructuras >2mm presentes en la harina no refinada y refinada, derivadas de los Algarrobos blanco y negro.

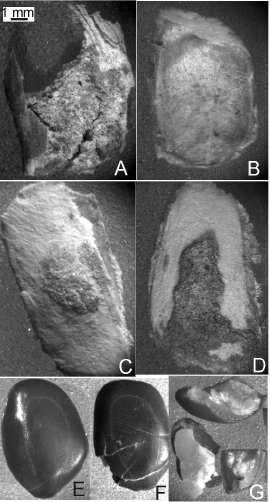
Fig. 7. Estructuras o sus partes >2mm más abundantes de la harina no refinada derivada del Algarrobo blanco. A, epicarpo-mesocarpo. B, medio endocarpo. C, endocarpo cerrado con mesocarpo. D, endocarpo abierto. E, semilla entera con testa sana. F, semilla entera con testa fisurada. G, semillas fragmentadas.
Se calcularon los porcentajes de cada tipo de órgano presente luego de la manufactura de la harina no refinada, con respecto a la cantidad de ese mismo órgano antes de comenzar la molienda (Tabla 2). Los endocarpos cerrados disminuyeron al 28 % del 100 % original, mientras que un 68 % se dividió en mitades. Dentro de las semillas predominaron aquéllas enteras (44 %) por sobre las fragmentadas (28 %). Algunas permanecieron dentro de los endocarpos cerrados (28 %). Los fragmentos de epicarpo-mesocarpo se estimaron en un 40 % de la cantidad presente originalmente
Tabla 2. Porcentaje de estructuras presentes con respecto al porcentaje inicial luego del procesamiento de vainas de Algarrobo blanco en harina no refinada (90 segundos de molienda) y harina refinada (140 segundos de molienda) calculados sobre la base del número absoluto de fragmentos.

Algarrobo negro. Las partículas mayores a 2 mm que conformaron esta harina pueden observarse en la Tabla 1. Las mismas constituyeron principalmente endocarpos cerrados, tanto sanos (35 %) (Fig. 8A), como con una fisura alargada (21 %) (Fig. 8B). En una menor proporción se observaron endocarpos fragmentados (32 %) (Fig 8C). A diferencia del Algarrobo blanco cuyos endocarpos se abren, por lo general sin dañarse, por la línea media de sutura, en el Algarrobo negro las roturas aparecen en forma irregular generalmente abarcando ambas valvas del endocarpo. Se observó también, aunque en muy baja cantidad, la presencia de semillas enteras con testa sana y de fragmentos de vaina (6 % cada uno) (Fig. 8D). Las partículas entre 1 y 2 mm estuvieron constituidas por fragmentos pequeños de epicarpo y endocarpo y la porción <1mm por mesocarpo.

Fig. 8. Estructuras o sus partes >2mm más abundantes de la harina no refinada derivada del Algarrobo negro. A, endocarpo cerrado. B, endocarpo figurado. C, endo-carpo fragmentado. D, fragmento de vaina. La escala de A vale para B y C.
Los porcentajes de cada tipo de órgano presente luego de la manufactura de la harina no refinada con respecto a la cantidad de ese mismo órgano antes de comenzar la molienda se muestran en la Tabla 3. Predominaron los endocarpos cerrados (76 %) por sobre los fragmentados (24 %). Las semillas liberadas constituyeron solo el 8 % de su cantidad inicial.
Tabla 3. Porcentaje de estructuras presentes luego del procesamiento de vainas de algarrobo negro en harina no refinada (90 segundos de molienda) y harina refinada (140 segundos de molienda) calculados sobre la base del número absoluto de fragmentos.

Productos intermedios, productos finales y residuos. Harina refinada
Algarrobo blanco. Las partículas mayores a 2 mm que conformaron esta harina pueden observarse en la Tabla 1. Las mismas estuvieron constituidas principalmente por medios endocarpos (54 %), muchos de ellos con una abertura en forma de V (Fig. 9A). Los endocarpos cerrados fueron nulos, mientras que una pequeña proporción de endocarpos permanecieron abiertos (5 %). Las semillas fragmentadas (35 %) aumentaron en relación a la harina no refinada, mientras que disminuyeron las semillas con testa fisurada (6 %). Las fisuras en estas últimas fueron más pronunciadas (Fig. 9B). Las partículas entre 1 y 2 mm estuvieron constituidas por fragmentos pequeños de epicarpo y tejidos seminales. La porción <1mm estuvo constituida por todo el mesocarpo y parte del epicarpo.
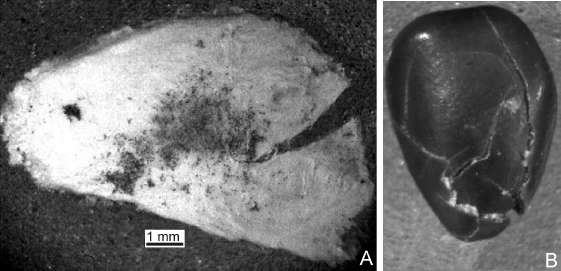
Fig. 9. Estructuras o sus partes >2mm presentes en la harina refinada derivada del Algarrobo blanco. A, medio endocarpo con fisura en V. B, semilla entera testa muy fisurada.
Los porcentajes de cada tipo de órgano posible de ser recuperado luego de la manufactura de la harina refinada con respecto a la cantidad de órganos presentes antes de comenzar la molienda se muestran en la Tabla 2. Se observó que la mayoría de los endocarpos se partió al medio (84 %) y que la mayoría de las semillas se presentaron fragmentadas (80 %) por sobre las enteras con testa fisurada (20 %); se destaca también la carencia de fragmentos de epicarpo-mesocarpo.
Algarrobo negro. Las partículas mayores a 2 mm que conformaron esta harina pueden observarse en la Tabla 1. Las mismas estuvieron constituidas principalmente por endocarpos (85 %), tanto cerrados sanos (28 %), así como con distintos grados de fisuras (10 %) (Figs. 10A-B) y fragmentados (46 %) (Fig. 10C), algunos de los cuales conservaron su semilla (Fig. 10D). Aparecieron semillas con testa fisurada (8 %) y fragmentadas (8 %), que no estaban presentes en la harina no refinada. Las partículas entre 1 y 2 mm estuvieron constituidas por fragmentos pequeños de epicarpo, endocarpo y tejidos seminales. La porción <1mm estuvo constituida, en su mayor medida, por mesocarpo, aunque también por escasas cantidades de epicarpo y endocarpo.

Fig. 10. Estructuras o sus partes >2mm presentes en la harina refinada derivada del Algarrobo negro. A-B, endocarpos fisurados. C, endocarpo fragmentado. D, endocarpo fragmentado con semilla.
Los porcentajes de cada tipo de órgano posible de ser recuperado luego de la manufactura de la harina refinada con respecto a la cantidad de órganos presente antes de comenzar la molienda se muestran en la Tabla 3. Los endocarpos fragmentados (40 %), si bien no se equipararon a los cerrados (60 %), aumentaron mucho su valor. Pocas semillas fueron liberadas (24 %), mientras que el resto permaneció en los endocarpos cerrados (76 %). Patay y ulpo. Aunque el patay podría llegar a formar parte del registro arqueológico como pan debido a que sus partículas de harina fina provenientes principalmente del mesocarpo, se aglutinan gracias a sus azúcares, este pan tiene pocas probabilidades de preservarse en estado seco debido a que se convierte fácilmente en migajas y a que se presta para ser ingerido por animales silvestres durante el período posterior a su depósito en los sitios. El ulpo, por ser una dilución, directamente no se puede confirmar arqueológicamente a través de macrorrestos. (véase Capparelli, sine data). En estos casos, una manera más certera de evaluar la posible elaboración de patay y/o ulpo es a través de la presencia de sus residuos (elementos característicos de la fracción gruesa de la harina refinada o residuo tipo 5 en la Fig. 1). La cantidad de residuo producido varía según la cantidad de vainas procesadas y el rendimiento que las mismas poseen, es decir, la proporción de mesocarpo -principal tejido formador de harina fina- con respecto al epicarpo y al endocarpo y semilla. Debido a que dicha proporción es variable entre las distintas vainas, tanto de manera intraespecífica como interespecífica (Fig. 3C, D; Tabla 4), se calculó la proporción promedio de mesocarpo aportada por vainas de cada una de las especies tratadas en el valle de Hualfín. Dicha proporción varió entre 44,6 y 53,1 % en el AB y entre 20,8 y 40,8 % en el AN, siendo el promedio del peso de las vainas completas de AB de 4 g y de AN de 2,3 g y el número de artejos promedio de 20 y 15 respectivamente (Tabla 5). En el caso del AB las vainas variaron entre 1,7 y 5,7 g de peso y tuvieron entre 13 y 26 artejos. En el caso del AN las vainas variaron entre 0,6 y 4,1 g de peso y tuvieron entre 9 y 26 artejos. En suma, el Algarrobo negro presentó valores menores de rendimiento, peso y número de artejos con respecto al Algarrobo blanco.
Tabla 4. Proporción del peso en porcentaje de cada uno de los tejidos que componen la vaina de Algarrobo blanco y Algarrobo negro. Abreviaturas: epi, epicarpo; meso, mesocarpo; end+sem, complejo endocarpo-semilla; fragm, fragmento; DEM, desvío estándar de la media.
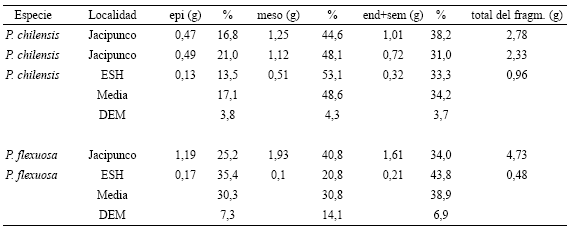
Tabla 5. Peso y cantidad de artejos de vainas individuales de Algarrobo blanco y Algarrobo negro provenientes de distintas localidades a lo largo del Valle de Hualfín. Abreviatura: DEM, desvío estándar de la media.
Si bien la presencia de la fracción gruesa de harina refinada es útil para determinar la manufactura de patay o ulpo, ésta posee dos limitantes. Por un lado, no es posible discernir si el residuo corresponde a una u otra preparación. Por otro lado, puede darse el caso de que esta fracción gruesa haya sido utilizada para la elaboración de añapa, por lo cual los residuos de la preparación de patay-ulpo pierden su visibilidad arqueológica como tales y pasarían a verse como residuos de la elaboración de añapa.
Añapa y aloja. Luego de remojar las distintas partes de órganos producidas a partir de las harinas de Algarrobo durante 20 horas (añapa), el 98 % de los órganos pudo ser recuperado (Tabla 6). Los endocarpos se recuperaron en un 100 % sin observar cambio morfológico alguno en sus dimensiones ni en el Algarrobo blanco ni en el negro. En el caso del AB sus medidas de largo y ancho promedio fueron de 10 x 6,9 mm respectivamente y en el caso de AN de 11 y 9,6 mm respectivamente. El caso de las semillas fue distinto. El 33 % de las semillas enteras sanas de Algarrobo blanco pasó a tener la testa fisurada por efecto del aumento de volumen durante la hidratación (posiblemente tendrían ya fisuras imperceptibles antes del remojo, por donde pudo haber penetrado más rápidamente el agua). El 20 % de las semillas con testa fisurada de dicha especie se fragmentó en varias partes. Esta última disminución es imperceptible dado que esta categoría percibe el aporte de semillas que antes del remojo tenían su testa sana. Si bien en el AN se recuperaron el 100 % de las semillas enteras, éstas, al igual que en el Algarrobo blanco, se encontraron aumentadas de volumen.
Tabla 6. Número absoluto y porcentaje de las estructuras recuperadas luego de un remojo de 20 horas y 10 días correspondiente al procesamiento de añapa y aloja respectivamente. Abreviatura: Nro. abs, número absoluto; AB, Algarrobo blanco. AN, Algarrobo negro.

Luego del remojo durante 10 días (aloja), volvieron a recuperarse el 100 % de los endocarpos sin alteración morfológica alguna. No obstante, se obtuvo sólo el 33 % de las semillas enteras sanas tanto en el caso del AB como del AN. El 67 % de las semillas enteras sanas pasó a tener la testa fisurada por lo que esta última categoría aumentó al 120 % en el caso del AB y al 200 % en al caso del AN. En aquellas semillas que ya inicialmente poseían fisuras notorias en su testa, las mismas se pronuncian (Figs. 11A-B) y la testa se levanta o enrolla (Figs. 11C-D), dejando en ocasiones el albumen y/o los cotiledones parcial (Figs. 11E-F) o totalmente (Fig. 11G) a la vista. Otras semillas se fragmentaron y algunas se separaron en los distintos tejidos seminales (Fig. 11H). Es importante mencionar aquí que cuanto mayor fue la proporción de mesocarpo de la harina utilizada para realizar la bebida (por ejemplo cuando se la realizó a partir de la totalidad de las fracciones de las harinas, ya sean éstas refinadas o no, y no a partir del residuo de la preparación de patay/ulpo), más se aglutinaron los residuos de añapa-aloja al secarse (véase Capparelli, sine data) (Figs. 11I-J).

Fig. 11. Estructuras o sus partes >2mm presentes en el residuo de añapa-aloja. A-C, semillas enteras con testa fisurada de Algarrobo blanco (A) y Algarrobo negro (B,C). D, semilla entera testa levantada, Algarrobo blanco. E-F, semillas enteras con tejidos internos visibles de Algarrobo blanco (E) y Algarrobo negro (F). G, semilla sin testa. H, semillas fragmentadas. I-J, endocarpos con aglutinamiento de fragmentos pequeños de vaina de Algarrobo blanco (I) y Algarrobo negro (J).
Con respecto a la cuantificaci ón del cambio de volumen de semillas antes y después del sumergido en agua y luego de su secado posterior, se observó en primer lugar que a las 20 horas de remojo las semillas de ambas especies aumentaron su largo entre 26 y 41 %, su ancho entre 19 y 35 % y su espesor entre 21 y 100 % (Tablas 7 - 8). Esto se debe a que el albumen se hidrató adquiriendo un aspecto gelatinoso. En segundo lugar, a los 10 días de remojo, el largo aumentó entre 43 y 51 %, el ancho entre 37 y 50% y el espesor entre 52 y 90 % (Tablas 7 y 9), con respecto a su estado inicial. Por último, a los 21 días de remojo las semillas mantuvieron similares proporciones de aumento en largo (de 39 a 48 %) y ancho (de 33 a 38 %) con respecto al período de 10 días, no obstante, es llamativo el hecho de que su espesor disminuyó. Esto último podría deberse a un daño en los tejidos internos producido por la proliferación de bacterias y/u hongos, tal como puede ocurrir en ocasiones durante los procesos de fermentación (Ferrarese, 1984:130-131). Una vez que las semillas se secaron nuevamente, recuperaron casi sus largos y anchos originales ya sea que provengan de 20 horas, 10 ó 21 días de remojo (el largo sólo aumentó de 0 a 11 % y el ancho de 0 a 18 % con respecto al tamaño original de la semilla), en tanto que el espesor en algunas se conservó, aumentado entre 14 y 15 %, mientras que en otras disminuyó drásticamente desde 17 a 42 % de su espesor inicial (Tablas 7 - 9).
Tabla 7. Promedio (n=3) del largo (L), ancho (A) y espesor (E) (en milímetros) y porcentaje de su aumento o disminución con respecto a la medida en el estado inicial de semillas enteras de testa sana de Algarrobo blanco y Algarrobo negro sometidas a diferentes períodos de remojo (20 horas y 10 días y 21 días) y a un secado posterior. Abreviaturas: sp., especie; AB, Algarrobo blanco; AN, Algarrobo negro.
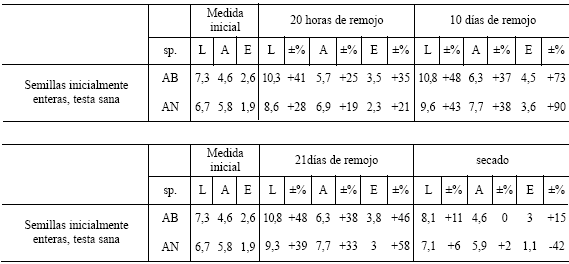
Tabla 8. Promedio del largo (L), ancho (A) y espesor (E) (en milimetros) y porcentaje de su aumento o disminución con respecto a la medida en el estado inicial de semillas enteras de testa sana de Algarrobo blanco y Algarrobo negro sometidas a un período de remojo de 20 horas y a un secado posterior. Abreviaturas: sp., especie; AB, Algarrobo blanco; AN, Algarrobo negro.
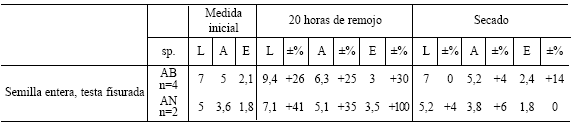
Tabla 9. Promedio del largo (L), ancho (A) y espesor (E) (en milímetros) y porcentaje de su aumento o disminución con respecto a la medida en el estado inicial de semillas enteras de testa sana de Algarrobo blanco sometidas a un período de remojo de 10 días y a un secado posterior.

Arrope. Los residuos obtenidos en húmedo estuvieron constituidos por los endocarpos cerrados (Figs. 12A-B) y por largas piezas de epicarpo. Ambas estructuras presentan áreas donde quedó aún adherido el pastoso mesocarpo hervido, que se evidenció como una pátina oscura al secarse, más evidente en el AB. Esta pátina es producto del amasado o rallado de las vainas contra la malla para separar el jarabe de su residuo y se puede observar con microscopio estereoscópico. El 100 % de los endocarpos utilizados inicialmente se recuperaron como residuo y conservaron las mismas dimensiones que en su estado inicial. La mayoría se encontraron libres, aunque unos pocos estuvieron soldados por tejido epicárpico o mesocárpico (Figs. 12C-E) o por mesocarpo solamente (Fig. 12F). Al secarse, algunos de los endocarpos libres se aglutinaron también a través del mesocarpo (Fig. 12A). Ambas especies observaron el mismo comportamiento, aunque en el AN las piezas de epicarpo fueron más finas y largas (Fig. 12G) que en el AB (Fig. 12H). Al deshidratarse, las piezas más finas de epicarpo se enrollaron.
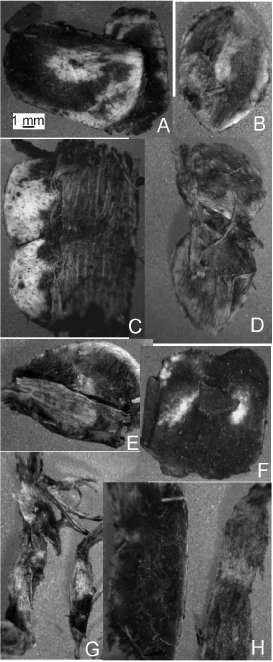
Fig. 12. Estructuras o sus partes >2mm presentes en el residuo de arrope. A-B, endocarpos simples en Algarrobo blanco (A) y Algarrobo negro (B). C-E, endocarpos con epi y mesocarpo en Algarrobo blanco (C) y Algarrobo negro (D-E). F, endocarpos de Algarrobo blanco aún apareados por mesocarpo. G-H, fragmentos de epicarpo en Algarrobo negro (G) y Algarrobo blanco (H).
DISCUSIÓN Y CONCLUSIONES
La proporción de la fracción < 1mm de la harina refinada extraída de las vainas determina el rendimiento de las mismas para la manufactura de patay o ulpo. Esta harina es la que Cáceres Freyre (1962:13) en un trabajo donde relata la manufactura del patay en el Bolsón de Pipanaco, llama "ñuta" que según él significa "echa polvo muy tenue". Una parte de ella, la porción más fina en extremo, se denomina "lustre" y es la que se coloca en las capas más externas del patay para darle mejor vista. A la fracción > a 1mm de la harina refinada, que conformaría el residuo del patay, Cáceres Freyre (1962) la clasifica en dos categorías: el "aunchi" o "amchi" (fracción > a 2mm del presente trabajo) y la "sépuca" o "seuca" (fracción entre 1 y 2 mm del presente trabajo). El "aunchi" dice que puede ser utilizada para realizar arrope. No obstante en el área tratada en el presente trabajo sólo se mencionó, a partir de la misma, la elaboración de añapa. La "sépuca" dice que se usa para alimentar a las gallinas. En el Valle de Hualfín es común que los siete distintos tipos de residuos derivados del Prosopis se usen con este fin (Capparelli, sine data).
En el caso del AB del Valle de Hualfín, el rendimiento en harina fina fue alto, llegando casi al 54 % del peso inicial. El porcentaje promedio de la proporción de mesocarpo estimado para vainas de AB del Valle de Hualfín es de 49 %, por lo cual, teniendo en cuenta los tejidos que componen esta harina, se deduce que el 5 % restante de la fracción fina obtenida deriva del epicarpo. Cáceres Freyre (1962) registró rendimientos similares para el Algarrobo negro. Este autor dice que 10 kg de algarroba dan 5 kg de patay. Sin embargo, se contradice al afirmar que en un día se producen de 140 a 160 kg de algarroba que darán lugar a 130 patayes, los cuales por datos que se brindan más adelante en el trabajo, se sabe que pesan 250 g cada uno. Según este último cálculo, el rendimiento se reduciría a un 20-23 % del peso inicial de las vainas. Figueroa & Dantas (2006), en un trabajo etnográfico realizado desde una perspectiva arqueológica a través de entrevistas a pobladores locales, abordaron la producción actual de patay en las proximidades de la ciudad de Añatuya, Santiago del Estero. Según estos autores, en esa localidad se utiliza en la confección de patay tanto Prosopis nigra como P. alba, de las cuales los pobladores remitieron un rendimiento generalizado cercano al 50 %. Este rendimiento generalizado no coincide con los datos aportados en el acápite resultados del presente trabajo, donde se registra una alta variabilidad en la producción de mesocarpo entre P. chilensis (47%) y P. flexuosa, e incluso en forma intraespecífica. Una similar registran Solbrig et al. (1977) cuando hacen referencia a las variaciones en la producción de vainas de Prosopis gordas o flacas, cuyo mayor espesor está generalmente asociado a sitios de mayor humedad. Es conocida la alta variabilidad poblacional de este género (Brizuela et al., 2000) y la elevada hibridación interespecífica dentro de distintas Series de la Sect. Algarobia (véase Palacios & Bravo, 1974, 1981; Trobok, 1984; Verga, 2000, entre otros). Esta variabiliadad está asociada al hecho de que muchas de las especies de Prosopis son genéticamente polimórficas, aunque se relaciona también con factores de selección tanto ambientales como bióticos (Solbrig et al., 1977) y probablemente también culturales (Capparelli, sine data). Volviendo al caso de la producción de mesocarpo, un menor rendimiento del AN con respecto al AB implicaría a nivel arqueológico por ejemplo, el procesamiento de mayor cantidad de vainas del primero para obtener la misma cantidad de harina de ambos y consecuentemente un mayor número de horas por persona de trabajo y una mayor producción de residuos en relación con la harina extraída. Por lo tanto, al momento de realizar generalizaciones al nivel de rendimiento, es importante conocer las características específicas del ambiente donde cada una de las especies tratadas crecen, establecer claramente cuál es el rendimiento de sus vainas y determinar la proporción en que las distintas especies pueden haber sido mezcladas, si es que lo fueron.
Es llamativo que aunque la proporción de mesocarpo estimada para el AN en el valle de Hualfín fue del 31%, la fracción fina de la harina refinada producida experimentalmente en este trabajo apenas llegó al 14%. Se piensa que esta baja proporción de harina fina no es el resultado de una baja cantidad de mesocarpo en la vaina, sino de las condiciones particulares de almacenamiento en que las vainas utilizadas se encontraban. Por ejemplo, una deficiencia en su empaquetamiento y una consecuente absorción de humedad, pudo provocar que durante la molienda gran parte del mesocarpo quedara adherido al endocarpo y a los fragmentos de epicarpo. Se deduce entonces la gran importancia que el efecto del secado posee para obtener buenos rendimientos de harina destinada a la elaboración de patay o ulpo, lo cual fue remarcado anteriormente por varios autores (Di Lullo, 1944: 269; Cáceres Freyre, 1962:11; Capparelli, 2007). Según Figueroa & Dantas (2006: 43), la inversión en tiempo que los pobladores deber realizar para esta tarea es elevada.
Por otro lado, Figueroa & Dantas (2006) proponen que en Santiago del Estero, y con morteros de madera o piedra, se muelen de 3 a 6 kg de algarroba por hora. Estos valores en eficiencia son mucho mayores que los empleados en el presente trabajo (0,2 kg/hora). Otro ejemplo plantea que mediante la utilización de la cimbra, un mortero a modo de balancín, cuya mano denominada "maray" pesa 40 kg, se muelen de 35 a 40 kg de algarroba por hora (Cáceres Freyre, 1962). El autor aclara, además, que dicho trabajo es realizado por un hombre que mueve el balancín y por dos mujeres que empujan las vainas al sitio donde el maray golpea. Estas diferencias en cuanto a la eficiencia de la molienda probablemente se deban a variaciones en la duración de la acción, en el tipo de dispositivo empleado para moler, en la intensidad del golpe al moler, en el peso de la mano empleada, en la cantidad de personas involucradas en la tarea y, finalmente, en la destreza del que realiza la acción.
Tanto el análisis como la correcta interpretación de las proporciones relativas de cada categoría de órgano vegetal son un complemento necesario de los caracteres morfológicos cualitativos a fin de identificar la elaboración de productos intermedios y residuos derivados del Algarrobo. Los estudios llevados a cabo en el Valle de Hualfín han puesto nuevamente de manifiesto que la distinción entre Algarrobo blanco y Algarrobo negro es esencial para interpretar los macrorrestos de Prosopis (Capparelli, 2007), ya que un procesamiento idéntico de ambas especies produce diferentes calidades y cantidades de órganos o partes de órganos que deben ser evaluados con parámetros propios de cada tipo de Algarrobo como para poder ser interpretados fehacientemente. Dadas las similitudes en cuanto a las características de los frutos y sus usos entre P. chilensis y P. flexuosa por un lado y P. alba y P. nigra, Algarrobo blanco y Algarrobo negro respectivamente, ambos chaqueños por otro, se piensa que los resultados de este trabajo podrían aplicarse potencialmente también a estos últimos. Sin embargo, se requiere una mayor cantidad de evidencias como para confirmar dicha suposición, las cuales serán abordadas en trabajos futuros.
Los órganos o partes de órganos más diagnósticos para determinar la harina no refinada de la refinada y por ende la posible manufactura de las distintas bebidas y/o alimentos consideradas en el presente trabajo, son los endocarpos, las semillas y en menor medida, los restos de vaina o de fragmentos de epicarpo-mesocarpo. La presencia de harina no refinada podría indicar la futura preparación de añapa, aloja, patay o ulpo. En el caso del AB la harina no refinada se caracteriza por tener muy altos porcentajes de medios endocarpos y de fragmentos de epicarpomesocarpo, bajos porcentajes de endocarpos cerrados, de semillas con testa fisurada y de semillas fragmentadas y muy bajos porcentajes de endocarpos abiertos y de semillas de testa sana; mientras que en el caso del AN la misma posee muy altos porcentajes de endocarpos cerrados (tanto sanos como fisurados) y fragmentados, así como muy bajos porcentajes de vaina fragmentadas y de semillas enteras de testa sana. Por otro lado, la identificación de harina refinada podría indicar que ha sido preparada para elaborar aloja, patay o ulpo. La harina refinada de Algarrobo blanco se caracteriza por tener muy altos porcentajes de medios endocarpos y de semillas fragmentadas y muy bajos porcentajes de endocarpos abiertos y de semillas enteras de testa fisurada, así como por carecer de semillas enteras de testa sana. La harina refinada de Algarrobo negro posee muy altos porcentajes de endocarpos cerrados sanos y de endocarpos fragmentados y bajos porcentajes de endocarpos fisurados, así como de semillas enteras de testa fisurada y de semillas fragmentadas. Como se ha mencionado anteriormente, individualizar los restos provenientes de cada una de las especies utilizadas es fundamental para poder evaluar cuantitativamente los mismos y poder contribuir a la determinación de la manufactura de harina refinada y/o no refinada. En el caso del AB, un índice que refleje el cociente entre la cantidad de semillas enteras (tanto sanas como con testa fisurada) y de semillas fragmentadas, podría darnos mayor confiabilidad para la identificación de harina no refinada (si el valor del mismo es mayor a 1) y de harina refinada (si dicho valor es inferior a 1). Resulta importante tener en cuenta que la cantidad de semillas fragmentadas de dicho cociente representa el número de semillas que antes de la molienda estaban enteras y luego de ésta se fragmentaron. Este número puede calcularse a partir del peso de los fragmentos de semilla recuperados con respecto al peso de una sola semilla, y tiene la ventaja de ser independiente del tamaño de los fragmentos. La diferencia en la cantidad de capas de células en empalizada en la parte media de las caras de las semillas de las especies tratadas (véase resultados) sería un carácter muy útil en la determinación de semillas fragmentadas a escala específica, que por su condición no permitirían registrar los caracteres diagnósticos tradicionales como tamaño y forma de la semilla, ubicación y características de la línea fisural, entre otros caracteres. En el caso de que la identificación a escala específica de las semillas fragmentadas no sea posible, se debe recurrir sólo a los caracteres cualitativos (en particular a la presencia de semillas de testa sana, de restos de epicarpo-mesocarpo y de endocarpos cerrados) para distinguir la harina no refinada de la refinada de AB. En el caso del AN, el cociente entre cantidad de endocarpos cerrados sanos y la cantidad de endocarpos fragmentados, resulta ser un carácter útil para determinar la harina no refinada de la refinada. Al igual que en el caso de las semillas, se debe tener en cuenta que la cantidad de endocarpos fragmentados no estaría representada por el número de fragmentos sino por el número de endocarpos que antes de la molienda estaban enteros y luego de ésta se fragmentaron. Este número debe calcularse a partir del peso de los fragmentos de endocarpo recuperados con respecto al peso de una solo endocarpo sin su respectiva semilla. Un cociente mucho mayor que 1 (en el caso del presente trabajo: 2) indicaría la presencia de harina no refinada y un cociente cercano a 1 (en este caso: 1,1) indicaría la presencia de harina refinada. En el caso de la harina refinada podría darse un posible sesgo tendiente a subrepresentar los endocarpos fragmentados dado por el hecho de que parte de éstos se pierden entre la fracción < a 2 mm de la harina, pero se considera que en función del peso dicha proporción es despreciable.
Para distinguir la elaboración de añapa/aloja con respecto al patay/ulpo se debe recurrir al análisis morfológico cualitativo de las semillas y no de los endocarpos, ya que el 100 % de los endocarpos no evidenciaron cambios notorios en sus cualidades después del remojo. Por lo tanto, existe una mayor probabilidad de poder distinguir el complejo añapa-aloja cuando éste deriva del Algarrobo blanco, ya que en este caso la mayoría de los endocarpos se abre con la primer molienda, liberando las semillas, más que cuando deriva de AN donde un 88 % de los residuos de la muestra se compone de endocarpos. Se debe tener en cuenta que la proporción de semillas inicialmente remojadas, se distorsiona después del período de remojo (aquellas de testa sana se encuentran subestimadas y las de testa fisurada sobrestimadas). Desafortunadamente no se evidenciaron caracteres conspicuos que permitan diferenciar claramente la manufactura de añapa con respecto a la de aloja (véase también Capparelli, sine data). Incluso, en el caso de que se evidencien residuos de añapa-aloja, no es posible dejar de lado la elaboración de patayulpo, ya que el primer complejo pudo haberse realizado a partir de los residuos del segundo.
La presencia de restos de epicarpo y de un 100 % de endocarpos cerrados es diagnóstica para identificar la manufactura de arrope. Sin embargo, se requiere especial atención para diferenciar los endocarpos derivados del procesamiento para la elaboración de harina de aquellos obtenidos como residuo final del arrope. En restos desecados esta discriminación es más sencilla ya que los segundos presentan una pátina pastosa marrón rojiza que los recubre aleatoriamente en algunas áreas, que proviene del mesocarpo hervido y es producto del amasado o rallado de las vainas contra la malla que separa el jarabe de su residuo. Esta pátina se puede observar utilizando microscopio estereoscópico. No obstante, en restos carbonizados su identificación puede ser más dificultosa. Los estudios sobre este último tipo de restos serán abordados en trabajos futuros.
En el caso del arrope, aunque se ha propuesto que su introducción como técnica en el Noroeste de Argentina se habría dado a partir de los conquistadores españoles (Scarpa, 1999), se considera interesante que dicha propuesta pueda ser contrastada en el futuro por la arqueoetnobotánica. El término arrope habría sido dado a estos productos por su similitud con aquellos del Viejo Mundo. La voz arrope proviene del árabe "arrubb" y significa "el jugo de frutos cocidos" (Scarpa, 1999: 96). Scarpa (1999: 94) propone que su origen en América, "...podría rastrearse tanto en las mieles elaboradas por los aborígenes americanos a partir de zumos vegetales, como en los jarabes de la antigua farmacopea árabe introducida por los musulmanes en el sur de España...". Ante estas dos posibilidades, y luego de analizar varias fuentes bibliográficas, este autor reconoce que la técnica en sí misma era ya conocida en Perú y Nueva España donde antes de llegar los españoles se realizaban jugos cocidos de molle (según Scarpa, 1999: 98, este nombre podría corresponder a Schinus molle L. o a otras entidades afines como Sch. areira L.), Caña de maíz (Zea mays L.) o Maguey (Agave atrovirens Karw. y A. americana L.). Sin embargo, y debido, entre otras cosas, a que no hay registros etnohistóricos ni etnográficos de elaboración de estos jugos en el Chaco, el autor concluye que sería a partir de los conquistadores españoles que esta técnica se habría introducido en el Noroeste argentino como derivación subsidiaria de la producción vitivinícola y utilizando materias primas locales (Scarpa, 1999: 94). Los datos aportados por el presente trabajo permiten suponer que la manufactura de arrope, en el caso de que se haya llevado a cabo en tiempos precolombinos, pudo haber liberado al sedimento residuos potencialmente reconocibles y con gran probabilidad de preservación. Se considera necesario, entonces, analizar cuidadosamente los restos arqueológicos de Prosopis que se recuperen en el futuro, como para confirmar o rechazar la elaboración de arrope en el pasado.
A partir de este trabajo, se corrobora que las evidencias provenientes de la elaboración de añapa, aloja, patay o ulpo, se encontrarán altamente fragmentadas antes de llegar a formar parte de algún depósito arqueológico. Por lo tanto, será necesario poder distinguir en el futuro si la fragmentación de una asociación de restos de Algarrobo se pudo haber originado por la preparación de harinas, si es el resultado de la acción de factores que actúan posteriormente al depósito de un ítem en el sitio o bien del efecto de la carbonización, en el caso de restos carbonizados. Se requiere aún una mayor cantidad de información para responder a estos interrogantes. En relación con los restos carbonizados, es llamativo advertir que son escasas las etapas de procesamiento de Algarrobo donde sus fragmentos pueden estar en contacto con el fuego (para más detalles, véase Capparelli, sine data). Por lo que se deduce que, en el caso de estos restos, además de todos los factores relativos al procesamiento considerados en el presente trabajo, se debe tener en cuenta que la porción de residuos recuperada constituirá sólo una pequeña fracción de la totalidad de los residuos desechados.
En síntesis, a partir de los datos previamente presentados es evidente que no es sencillo evaluar los restos de Prosopis recuperados hasta el momento desde una perspectiva diacrónica. En el caso de los caracteres cuantitativos, toda estimación de cantidad de Algarrobo procesado en un espacio y tiempo determinado a partir del análisis de restos arqueobotánicos asociados con productos intermedios o residuos de este vegetal, debe partir tanto de la identificación de las especies involucradas, cuyos restos aportados al registro son diferentes, como de un reconocimiento de las condiciones de procesamiento del que fue objeto este recurso, así como también del tipo de producto final que se esperaba obtener a partir del mismo. La cuantificación de órganos derivados de distintos procesamientos del Algarrobo es compleja pero posible y debe ser realizada en forma criteriosa.
Es probable que en muchos casos la cantidad de restos de Prosopis recuperados hasta el momento en distintos sitios arqueológicos se encuentre severamente subestimada con respecto a las cantidades utilizadas durante el contexto dinámico que le dio origen y será un desafío comprobarlo en el futuro. Para finalizar y a modo de ejemplo, según los cálculos aquí presentados, del 100 % de los artejos iniciales de vaina de Algarrobo blanco procesados para distintos fines, se recuperará el 28 % de semillas con testa sana luego de la elaboración de harina no refinada, el 20 % de semillas con testa sana luego de la elaboración de harina refinada y sólo el 7 % (33 % de la categoría anterior) de semillas con testa sana luego de la elaboración de aloja. Si a esto se le suma el efecto de los procesos post-depósito en los sitios arqueológicos, ese 28 %, 20 % ó 7 % se verá aún más reducido. Y aún mayor será la reducción si se considera la posibilidad de recuperar sólo restos carbonizados en el caso de un sitio en cuestión, dada la escasa probabilidad de carbonización que poseen los restos provenientes del procesamiento del Algarrobo según analogías etnobotánicas provenientes del valle de Hualfín.
AGRADECIMIENTOS
A los pobladores del Valle de Hualfín por su disposición y colaboración de siempre. Al CONICET (PEI 6281, PIP 5148) y a la Agencia de Promoción Científica (PICT 10987) por el soporte económico brindado a quien suscribe.
BIBLIOGRAFÍA
1 Arana, M. M. 1999. El tiempo de la algarroba, en C. Aschero, A. Korstanje & P. Vuoto (eds.), En los tres reinos: prácticas de recolección en el cono sur de América, pp. 205-216. Tucumán: Instituto de Arqueología y Museo, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán. [ Links ]
2 Ardissone, R. 1961. Lineamientos de la instalación Humana en el Bolsón de Pipanaco. Anales de la Sociedad Argentina de Estudios Geográficos 11:171-244. [ Links ]
3 Arenas, P. 2003. Etnografía y alimentación entre los Toba-Nachilamole#ek y Wichí-Lhuku'tas del Chaco Central (Argentina). Buenos Aires: Latín Gráfica. [ Links ]
4 Beresford-Jones, D. G. 2004. Prehispanic Prosopis human relationships on the South Coast of Peru: riparian forest in the context of environmental and cultural trajectories of the Lower Ica Valley. Ph.D. diss., Magdalene Colledge, University of Cambridge. [ Links ]
5 Binford, L. 1981. Bones: Ancient men and modern myths. New York: Academic Press. [ Links ]
6 Boelcke, O. 1946. Estudio morfológico de las semillas de leguminosas Mimosoideas y Cesalpinioideas de interés agronómico en Argentina. Darwiniana 7: 240-321 [ Links ]
7 Brizuela, M. M.; A.D. Burghardt, D. Tanoni & R. A. Palacios. 2000. Estudio de la variación morfológica en tres procedencias de P. flexuosa y su manifestación en cultivo bajo condiciones uniformes. Multequina 9: 7-15 [ Links ]
8 Burkart, A. 1940. Materiales para una monografía del género Prosopis (Leguminosae). Darwiniana 4: 57-128 [ Links ]
9 Burkart, A. 1952. Las Leguminosas argentinas silvestres y cultivadas, 2ª. Edición. Buenos Aires: Acme. [ Links ]
10 Burkart, A. 1973a. A monograph of the genus Prosopis (Leguminosae subfam. Mimosoideae). J. Arnold Arb. 57 (4): 219-249 [ Links ]
11 Burkart 1973.b. A monograph of the genus Prosopis (Leguminosae subfam. Mimosoidea). Catalogue of the recognized species of Prosopis. J. Arnold Arb. 57 (4):450-525 [ Links ]
12 Cáceres Freyre, J. 1961. Diccionario de regionalismos de la provincia de La Rioja. Instituto Nacional de Investigaciones Folklóricas, Ministerio de Educación y Justicia de la Nación Argentina, Buenos Aires. [ Links ]
13 Cáceres Freyre, J. 1962. Fabricación del "patay" en los algarrobales del Campo de Palcipas (La Rioja y Catamarca). Instituto de la Producción, Facultad de Ciencias Económicas, Univ. Nac. La Plata, Serie Contribuciones 76:1-22. [ Links ]
14 Canals Frau, S. 1946. Etnología de los Huarpes. Una síntesis. Anales del Instituto de Etnología Americana 7: 9-147. [ Links ]
15 Capparelli, A. 2007. El Algarrobo blanco y negro -Prosopis chilensis (Mol.) Stuntz y P. flexuosa DC, Fabaceae, en la vida cotidiana de los habitantes del NOA: subproductos alimenticios. Kurtziana 33:103-119 [ Links ]
16 Capparelli, A. (Sine data). Archaeobotanical recognition of food products derived from the Algarrobo [Prosopis flexuosa DC and P. chilensis (Mol.) Stuntz] in Hualfín Valley, Argentina: An ethnobotanical and experimental approach for non-carbonised remains. Vegetation, History and Archaeobotany, 18; de próxima aparición. [ Links ]
17 D'Antoni, H. L. 1975. Introducción al estudio etnobotánico del Algarrobo. Actas del Primer Congreso de Arqueología Argentina, 23-28 de mayo de 1970, Rosario (Argentina), Tomo I, pp. 423-442. [ Links ]
18 D'Antoni, H. L. & O. T. Solbrig. 1977. Algarrobos in South American cultures, past and present, en B. B. Simpson (ed.), Mesquite: its biology in two desert scrub ecosystems, pp.189-199, Pennsylvania: Dowden, Hutchinson & Ross. [ Links ]
19 Di Lullo, O. 1943. El folklore de Santiago del Estero. Material para su estudio y ensayos de interpretación. Instituto de Historia, Lingüística y Folklore, Sec. Folk. X, Publicación Especial III: 1-446, Universidad Nacional de Tucumán, Departamento de Investigaciones Regionales, Tucumán. [ Links ]
20 Di Lullo, O. 1944. El folklore de Santiago del Estero. Medicina y alimentación. Provincia de Santiago del Estero. Publicación oficial. Santiago del Estero. [ Links ]
21 Felger, R. S. 1977. Mesquite in Indian cultures of southwestern North America, en B. B. Simpson (ed.), Mesquite: its biology in two desert scrub ecosystems, pp.150-176, Pennsylvania: Dowden, Hutchinson & Ross. [ Links ]
22 Felker, P. 1979. Mesquite, an all purpose leguminous arid land tree, en G. A. Ritchie (ed.), New Agricultural Crops, pp 89-132. Colorado: Westview Press, Boulder. [ Links ]
23 Ferrarese, M. 1984. Il vino fatto in casa. Bologne: Edagricole [ Links ]
24 Figueroa, G.G. & M. Dantas. 2006. Recolección, procesamiento y consumo de frutos silvestres en el noroeste semiárido argentino. Casos actuales con implicancias arqueológicas. La Zaranda de Ideas 2:35-50 [ Links ]
25 Filipov, A. 1996. Estudio Etnobotánico de la recolección entre los Pilagá. Tesis Doctoral. Universidad Nacional de La Plata. Facultad de Ciencias Naturales y Museo. [ Links ]
26 Giovannetti M. A.; V. Lema, C. G. Bartoli & A. Capparelli. 2008. Starch grains characterization of Prosopis chilensis (Mol.) Stuntz and P. flexuosa DC, and their implication in the analysis of archaeological remains in Andean South America. Journal of Archeological Science 35: 2973-2985 [ Links ]
27 Harris, P. J.C.; N. M. Pasiecznik, M. Bradbury & L. Ramirez. 1998. Problems and potential of Prosopis, en H. D. V. Prendergast, N. L. Etkin, D. R. Harris & P. J. Houghton (eds.), Plants for food and medicine, pp. 277-293. London: Royal Botanical Gardens, Kew. [ Links ]
28 Hillman, G. 1984. Interpretation of archaeological plant remains: The application of ethnographic models from Turkey, en W. Van Zeist & W. A. Caspary (eds.), Plants and Ancient Man. Studies in palaeoethnobotany, pp.1-41. Biologisch-Archaeologisch Institute, State University, Groningen. Rotterdam: AA Balkema. [ Links ]
29 Holden, T.G. & L. Núñez. 1993. An analysis of the gut contents of five well-preserved human bodies from Tarapacá, Northern Chile. Journal of Archaeological Science 20: 595-611. [ Links ]
30 Horkheimer, H. 2004. Alimentación y obtención de alimentos en el Perú prehispánico, 2ª. Edición. Perú: Instituto Nacional de Cultura. [ Links ]
31 Jones, G. 1984. Interpretation of archaeological plant remains: Ethnographic models from Greece, en W. Van Zeist & W. A. Caspary (eds.), Plants and Ancient Man. Studies in Palaeoethnobotany, pp. 43-59. Biologisch-Archaeologisch Institute, State University, Groningen. Rotterdam: AA Balkema. [ Links ]
32 Latcham, R. 1936. La agricultura precolombina en Chile y los países vecinos. Santiago de Chile: Universidad de Chile [ Links ]
33 Martínez Crovetto, R. 1964. Estudios Etnobotánicos I. Nombres de plantas y su utilidad según los indios Tobas del E. del Chaco. Bonpl. 1: 279-333. Corrientes. [ Links ]
34 Martínez Crovetto, R. 1968. Estudios Etnobotánicos III: Nombres de plantas y su utilidad según los indios araucanos-pampas del W de Buenos Aires. Rep. Argentina. Etnobiológica 12:1-24. [ Links ]
35 Martínez Crovetto, R. 1987. El Algarrobo sagrado de los Pampas. Buenos Aires. [ Links ]
36 Mc. Laren, F. & A. Capparelli. 2004. The early use of Prosopis L. Species as food: Old World and New World evidence [on line]. Abstracts of The International Society of Ethnobiology- Ninth International Congress, Panel 27: Ethnobota-nist of distant pasts: honour of Gordon Hillman. <http://www.kent.ac.uk/anthropology/ice2004/panels/panel 27.htlm>. [Accessed June 2004] [ Links ].
37 Métreaux, A. 1944. Estudios de Etnografía Chaquense. Anales del Instituto de Etnografía Americana 5: 263-314. [ Links ]
38 Mooney, H. A.; B. B. Simpson & O. T. Solbrig. 1977. Phenology, morphology and physiology, en B. B. Simpson (ed.), Mesquite: its biology in two desert scrub ecosystems, pp.26-43. Pennsylvania: Dowden, Hutchinson & Ross, Inc. [ Links ]
39 Moore, A. M. T.; G. C. Hillman & A. J. Legge. 2000. Village on the Euphrates from Foraging to Farming at Abu Hureyra. Oxford: OUP. [ Links ]
40 Morello, J. 1958. La provincia fitogeográfica del Monte. Opera Lilloana 2. Tucumán (Argentina): Universidad Nacional de Tucumán. [ Links ]
41 Noli, E. 1998. Algarrobo, maíz y vacas. Los pueblos indios de San Miguel del Tucumán y la introducción de ganados europeos (1600-1630). Mundo de Antes 1: 31-67. [ Links ]
42 Noli, E. 1999. La recolección en la economía de subsistencia de las poblaciones indígenas: una aproximación a través de fuentes coloniales (piedemonte y llanura tucumano-santiagueña, Gobernación del Tucumán), en C. Aschero, A. Korstanje & P. Vuoto (eds.), En los tres reinos: prácticas de recolección en el cono sur de América, pp. 205-216. Instituto de Arqueología y Museo. Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán, Tucumán. [ Links ]
43 Outes, F. 1917. Observaciones etnográficas de Francisco Javier Muñiz. Physis 3: 197-215. [ Links ]
44 Palacios, R. & L. Bravo 1974. Estudio morfológico de las semillas de algunos Prosopis del Nordeste argentino. Darwiniana 18: 437-452 [ Links ]
45 Palacios, R. & L. Bravo 1981. Hibridización natural en Prosopis: evidencias morfológicas y cromatográficas. Darwiniana 23: 3-35 [ Links ]
46 Roig, F. A. 1993. Aportes a la etnobotánica del género Prosopis, en Unidades de Botánica y Fisiología Vegetal (IADIZA)-CRICYT-CIID (eds.), Contribuciones Mendocinas a la Quinta Reunión Regional para América Latina y El Caribe de la Red de Forestación del CID: Conservación y mejoramiento de especies del género Prosopis, pp. 99-119. Mendoza. [ Links ]
47 Ruiz Leal, A. 1970. Flora Popular Mendocina. Deserta 3:1-296. [ Links ]
48 Rusconi, C. 1958. Acerca del pan de "patay". Rev. Mus. Hist. Nat. Mendoza 11: 193-208. [ Links ]
49 Sánchez de Oviedo, C. 1936. La vieja terapéutica vegetal en el noroeste argentino, en Almanaque del Ministerio de Agricultura de la Nación, sección publicaciones e informes, Año XI, pp 289-297. Buenos Aires [ Links ]
50 Sanni, A.I.; E. Lie & A. M. Lindberg. 1993. Fatty acid composition of Prosopis africana and its fermented product okpehe. Chem. Mikrobiol. Techno. Lebensm. 15 (3/4): 89-90 [ Links ]
51 Scarpa, G. F. 1999. El arrope en el Noroeste Argentino. Ayer una fiesta, hoy un capital, en A. Garrido Aranda (ed.), Los sabores de España y América, pp. 93-139. Huesca, España: Ed. La Val de Onsera. [ Links ]
52 Schiffer, M. B. 1972. Archaeological context and systemic context. American Antiquity 37: 156-185 [ Links ]
53 Schiffer, M. B. 1976. Behavioral Archaeology. New York: Academic Press. [ Links ]
54 Solbrig, O.T.; A. Bawa, N. J. Carman, J. L. Hunziker, C. A. Naranjo, R. A. Palacios, L. Poggio & B. B. Simpson. 1977. Patterns of variation, en B. B. Simpson (ed.), Mesquite: its biology in two desert scrub ecosystems, pp. 44-60, Pennsylvania: Dowden, Hutchinson & Ross. [ Links ]
55 Towle, M. 1961. The Ethnobotany of pre-Columbian Perú, Viking Fund Publications in Anthropology, Nº 30, New York. [ Links ]
56 Trobok, S. 1985. Morfología de frutos y semillas de Prosopis (Fabaceae-Mimosoideae) chilenos, en M. Habit (ed.), Estado actual del conocimiento sobre Prosopis tamarugo, pp 239-253, Organización de las Naciones Unidas para la Agricultura y Alimentación, Oficina Regional de la FAO para América Latina y El Caribe, Dirección de Producción y Protección Vegetal. [ Links ]
57 Verga, A. 2000. Algarrobos como especies para forestación: una estrategia de mejoramiento. Publ. trim. Secret. Agricult., Ganad. y Pesca y Alimen. 16: 12-19 [ Links ]
58 Villafuerte, C. 1984. Diccionario de árboles, arbustos y yuyos en el folklore argentino. Buenos Aires: Ed. Plus Ultra. [ Links ]
59 Winton, A. L. & K. B. Winton. 1935. The structure and composition of foods, Vol. II. New York: John Willey & Sons, Inc. [ Links ]

