










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkDarwiniana, nueva serie
versión impresa ISSN 0011-6793
Darwiniana vol.47 no.1 San Isidro ene./jun. 2009
ARQUEOBOTÁNICA Y ETNOBOTÁNICA
Criterios de selección en los procesos de manipulación vegetal: el aporte de la etnobotánica a la interpretación de restos arqueobotánicos de Cucurbita sp.
Verónica S. Lema
Departamento Científico de Arqueología, Laboratorio de Etnobotánica y Botánica Aplicada, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata; vslema@hotmail.com
Original recibido el 20 de mayo de 2008;
aceptado el 18 de marzo de 2009.
Resumen. En este trabajo se realiza el estudio etnobotánico-etnoarqueológico de diversos cultivos locales de Cucurbita maxima subsp. maxima, Cucurbitaceae. El objetivo es comprobar si los caracteres cuantitativos y cualitativos de las semillas procedentes de los mencionados cultivos, así como también de plantas resultantes de prácticas de cultivo o cuidado, se agrupan o no en morfotipos diferenciales que reflejen materialmente la diversidad localmente reconocida y los distintos comportamientos involucrados. La generación de dichos morfotipos se propone como una vía para explorar si es factible reconocer cultivares locales y distintas formas de relación hombre - planta en muestras arqueobotánicas. Las plantas domesticadas son el producto de la puesta en práctica de criterios de selección socialmente construidos y por lo tanto susceptibles de constituir morfotipos diferenciales que operen como portadores de significados sociales. De allí que podamos hablar de una etnobotánica etnoarqueológica generadora de modelos a contrastar en el trabajo paleoetnobotánico. Se aplicaron los descriptores morfológicos macroscópicos comúnmente empleados en paleoetnobotánica y arqueobotánica para el estudio de semillas de Cucurbita sp., así como también un conjunto nuevo de descriptores micro morfológicos. Se constató que los caracteres cuantitativos micro morfológicos son los que mejor sirven para crear morfotipos que se corresponden con los cultivares localmente reconocidos y con las prácticas de cultivo o cuidado ejercidas sobre las poblaciones vegetales.
Palabras clave. Cultivo; Etnoarqueología; Etnobotánica; Morfotipo.
Abstract. Selection criteria in processes of plant manipulation: ethnobotanical contribution to the interpretation of Cucurbita sp. archaeobotanical remains.
In this paper, an ethnobotanical-ethnoarchaeological study of different local landraces of Cucurbita maxima subsp. maxima, Cucurbitaceae, was carried on. The aim was to evaluate if different qualitative and quantitative seed characters of these local landraces, as well as those which came from plants resulting from cultivation or husbandry, get together or not in different morphotypes which materially reflect the diversity locally recognized, as well as local practices. The construction of these morphotypes is proposed as a way to recognize local landraces and different forms of Man - plant interactions in archaeobotanical samples. Domesticated plants are the result of actions derived from selection criteria socially built and transformed, therefore they can constitute different morphotypes which act as carriers of social meanings. For this reason we believe that an ethnoarcheological ethnobotany which generates models to contrast in paleoethnobotanical work will be a good option. Macroscopical morphological descriptors commonly used in archaeobotany were employed together with new micromorphological ones. It was concluded that quantitative micromorphological characters are the most adequate ones to create morphotypes which correspond with locally recognized landraces and with husbandry practices locally developed upon vegetable populations.
Keywords. Crop; Ethnoarchaeology; Ethnobotany; Morphotype.
INTRODUCCIÓN
La problemática de la manipulación humana de poblaciones vegetales y las distintas formas de relación que esto produce ha sido ampliamente abordada en estudios etnobotánicos (Ford, 1985; Harlan, 1992; Casas, 2001; Hildebrand, 2003). Si bien investigadores tales como G. Hillman han hecho importantes aportes en la generación de modelos etnobotánicos sobre prácticas de procesamiento y manipulación de vegetales que fueron luego aplicados a estudios paleoetnobotánicos (Hillman, 1984; Hillman & Davies, 1990), siguen siendo escasos los intentos por identificar categorías "emic" o locales en estudios arqueológicos de restos vegetales (Farrington & Urry, 1985; Lema, 2008).
Es importante aclarar que en este trabajo se asume que los estudios de restos arqueológicos vegetales (restos arqueobotánicos) pueden ser de dos clases, siguiendo la propuesta que surgió de la IV Reunión Internacional de Teoría Arqueológica en América del Sur, Inter.-Congreso WAC (Archila et al., 2008). El primer tipo de estudio es el arqueobotánico que comparte su objetivo con la arqueología y consiste en la reconstrucción de sociedades y comportamientos sociales en el pasado. El segundo es el paleoetnobotánico, con metas cercanas a la etnobotánica, que consisten en el análisis de las interrelaciones entre comunidades humanas y poblaciones vegetales, difiriendo de esta última sólo en el aspecto cronológico.
En este trabajo se realiza un estudio etnobotánico de aplicación etnoarqueológica, en tanto se procura identificar el reflejo material de comportamientos vinculados con el manejo de diversos cultivares locales de Cucurbita maxima Duchesne subsp. maxima, Cucurbitaceae. El objetivo es verificar si los caracteres cuantitativos y cualitativos de las semillas procedentes de los mencionados cultivares, así como también de plantas resultado de distintas prácticas de cultivo o cuidado, se agrupan o no en morfotipos diferenciales que reflejen materialmente la diversidad localmente reconocida y los distintos comportamientos involucrados. La generación de dichos morfotipos se propone como una vía para explorar si es factible reconocer cultivares locales y distintas formas de relación hombre- planta en muestras arqueobotánicas.
Algunas definiciones
Los estudios etnobotánicos han demostrado que las relaciones entre sociedades humanas y comunidades vegetales son muy diversas. Entre estas distintas formas de relación puede haber únicamente aprovechamiento de plantas o partes útiles de plantas de poblaciones silvestres (recolección), pueden encontrarse plantas útiles protegidas contra factores adversos (protección), otras que no son protegidas pero tampoco eliminadas (tolerancia) o bien estrategias para aumentar el número de ciertas poblaciones naturales (fomento o inducción) (Harlan, 1992; Casas et al., 1991; Casas, 2001). Para ciertos autores la protección y el fomento serían ya formas de cultivo, en tanto son actos intencionales de cuidar u ocuparse de una planta (Farrington & Urry, 1985; Harlan, 1992; Gremillion, 1997); para otros, en cambio, el cultivo sólo estará presente cuando se plante una semilla o propágulo en un área especialmente preparada para tal fin (Ford, 1979; Piperno & Pearsall, 1998; Casas, 2001; Hather & Mason, 2002). Como puede verse, estos comportamientos ligados con la acción de cultivar son muy difíciles de detectar arqueológicamente, salvo que causen modificaciones en el fenotipo de las plantas cultivadas, lo cual se encuentra aun bajo discusión (Ford, 1979; Harris, 1989; Gremillion, 1997; Gepts, 2005).
El cultivo es un prerrequisito para la domesticación, un tipo de relación donde la mayoría de los autores coinciden que existen cambios genotípicos y fenotípicos en las poblaciones vegetales manipuladas (Higgs, 1972; Ford, 1979, 1985; Farrington & Urry, 1985; Hillman & Davies, 1990; Harlan, 1992; Gremillion, 1997; Casas, 2001; Hather & Mason, 2002; Geps, 2005).
Se entiende que las poblaciones domesticadas serán menos diversas que las silvestres ya que reunirán el mismo conjunto de caracteres deseados y seleccionados por la comunidad humana con la cual se relacionan. Sin embargo, al generarse cultivares -entendidos como poblaciones de plantas de una misma especie domesticada con características distintivas que se originaron y persisten bajo cultivo- dentro de una misma especie domesticada, la variabilidad a nivel específico aumentará, si bien los cultivares tenderán a ser muy uniformes en sus características, por lo cual la variabilidad intrínseca de los mismos será baja (Gremillion,1993).
La aplicación de la taxonomía linneana a las plantas domesticadas es problemática, debiendo pensarse en vías alternativas para su agrupamiento y ordenamiento taxonómico, tanto en estudios etnobotánicos (Harlan, 1992) como arqueobotánicos (Marconetto, 2008). Esta complejidad taxonómica aumenta al considerarse los múltiples criterios de evaluación (Nazarea, 1998) empleados por los grupos humanos que crean, mediante selección cultural dirigida, diversos cultivares. Estas percepciones locales que generan una gran diversidad de criterios de evaluación sin límites claros se contraponen con los principios ordenadores reduccionistas de la ciencia occidental que entiende al mundo en blancos y negros, en tanto la gente lo ve en una amplia gama de grises (Nazarea, 1998).
Asumiendo esta dificultad, exploraremos vías que permitan reconocer estos criterios múltiples de evaluación y selección en el pasado. La vía que hemos elegido es la realización de trabajos etnobotánicos desde una perspectiva etnoarqueológica.
La posibilidad de una etnobotánica etnoarqueológica
A pesar de que la etnoarqueología ha recibido distintas denominaciones (disciplina, subdisciplina, abordaje o perspectiva) puede aproximarse una primer idea de la misma diciendo que es el estudio de los correlatos o reflejos materiales de las conductas sociales actuales con el objetivo de lograr un mejor entendimiento acerca de las conductas sociales pasadas estudiadas por medio del registro arqueológico. El mecanismo lógico por el cual se vincula lo observado en la sociedad actual (fuente) con la interpretación de las sociedades pretéritas (sujeto o meta) es la analogía. Los arqueólogos procesuales fueron los primeros en realizar estudios etnoarqueológicos buscando generar leyes universales a-históricas y transculturales (Politis, 2004) bajo una perspectiva comparativa general que buscaba regularidades en la conducta humana (Fournier & Freeman, 1991). Esto se lograba fragmentando aspectos culturales que, por lo general, eran conjuntos de acciones concretas y observables (Fournier & Freeman, 1991; Frére et al. 2004) -como el faenamiento de presas o la confección de artefactos- que se vinculaban entre la fuente y el sujeto totalmente despojados de su contexto cultural general. Al incluirse la totalidad cultural, tal como ocurrió a partir de los trabajos de arqueólogos post procesuales (Politis, 2004), la posibilidad de generar esta clase de asunciones generales alcanzó su fin, ya que cuando se consideraban los aspectos superestructurales, las acciones concretas eran más difíciles de universalizar. En esta nueva línea teórica ya no se buscaba generar modelos como en el procesualismo positivista, sino marcos interpretativos desde una postura hermeneútica, no se construían relaciones causales entre comportamiento y consecuencias materiales, ni se hacía de la correlación entre variables una explicación (Politis, 2004).
De acuerdo con Politis (2004) toda conducta social posee un correlato material, pero sólo si la misma es recurrente generará un patrón estructurado de dicha materialidad que tendrá posibilidades de ser arqueológicamente detectado. Manuel Gándara (2006) establece con mucha claridad lo que debe cumplirse en la analogía como inferencia inductiva: "la probabilidad de que la(s) propiedad(es) presentes en el conjunto base estén también presentes en el conjunto meta es una función de la probabilidad de que el conjunto base ocurra con una cierta regularidad con todas sus propiedades. De ser cierta esta premisa, entonces se puede postular que las propiedades no observadas en el conjunto meta probablemente estén/estuvieran contenidas en él a partir de que las propiedades generalmente ocurren juntas" (Gándara, 2006:16) "determinar cuáles y cuántas de estas propiedades deben de co-ocurrir como para ser una base sólida al momento de formular la hipótesis, o deben de añadirse a ambos conjuntos, en el momento de corroborarla, es un asunto que requiere de una argumentación más profunda, teórica, de qué propiedades son las más relevantes" (Gándara, 2006: 19). De acuerdo con este autor, la proyección inductiva requiere de la asunción de un principio general, por lo cual no se puede tener una ontología particularista. Esto no sería lo mismo que aceptar la existencia de leyes a-históricas, ya que el materialismo histórico asume que hay leyes generales para lo social y otras que se aplican sólo a ciertas formaciones socio-económicas en un momento histórico de su desarrollo. Este problema de historicidad no lo tienen las analogías cuyo principio general involucrado no es social, sino físico o químico: "hay analogías de diferentes órdenes de generalidad, que combinadas con otras que dependen de principios físicos, químicos o biológicos, son menos susceptibles a problemas de distorsión histórica" (Gándara, 2006: 22).
Entendemos en este trabajo que en toda sociedad las prácticas o acciones sociales conjugan conocimientos, conceptos y categorías socialmente compartidos e individualmente aprehendidos y transformados en la acción misma (Toledo, 2002). Por lo tanto, si nos ocupamos del cultivo, debemos estar atentos a que el mismo es la puesta en práctica de criterios de selección y toma de decisiones locales que guían la acción social o individual de los cultivadores (Nazarea, 1998). La domesticación y generación de cultivares es un fenómeno contemporáneo susceptible de proyectarse al pasado asumiendo que los criterios de selección y la manipulación de poblaciones vegetales son operaciones cognitivas y prácticas, respectivamente, que toda sociedad de cultivadores realiza sobre el entorno natural con el que se relaciona. Asimismo, se asume que si actualmente se observa que las poblaciones vegetales cultivadas responden a la manipulación humana generando cambios morfológicos, lo mismo debió de ocurrir en las poblaciones vegetales del pasado.
Ahora bien, como paleoetnobotánicos que trabajamos con restos arqueológicos nos interesa la posibilidad de que estas prácticas de selección cultural se plasmen materialmente en los restos arqueobotánicos, entendidos como artefactos (Ford, 1985; Lema, 2008). Por ello, a semejanza de lo que ocurre en paleobotánica, en esta disciplina debemos manejarnos con la creación de morfotipos, los cuales son codificaciones de fenotipos particulares por medio del empleo de descriptores morfológicos estandarizados (Lobot et al., 1999). Es en este punto donde es esencial investigar si la acción resultante de la aplicación de criterios de selección y toma de decisiones se materializa en cambios morfo-anatómicos en los órganos vegetales. De ser así, un morfotipo diferencial en el registro arqueobotánico podría hablarnos de prácticas locales respaldadas en criterios que identifican socialmente a un grupo.
Alejandro Casas y colaboradores han llevado a cabo gran número de estudios etnobotánicos donde constatan que formas de cultivo sin domesticación modifican rasgos morfológicos de las plantas involucradas en este tipo de relación, hallándose la diferencia más significativa en los caracteres que son objeto de la selección artificial (i.e. tamaño y forma de frutos de Cactaceae) (Casas et al, 1991; Casas et al, 1997; Casas, 2001; Arellano & Casas, 2003).
En una aplicación etnoarqueológica de esta clase de estudios morfológicos, además de interesarnos si existe o no una modificación en la anatomía de las plantas, nos interesa evaluar las posibilidades de que dichas modificaciones se ubiquen en órganos o tejidos que sobrevivan al paso del tiempo y a los procesos de formación del registro arqueológico: "Even if selection does produce directional changes in fruit morphology, these changes may not be detectable in the archaeobotanical record. This is often the case for drupes, where selection is directed towards changes in the quality of the flesh. For instance, domesticated olives (Olea europaea L.) and dates (Phoenix dactylifera L.) have larger and better quality flesh than the wild counterparts, but this is not reflected in the archaeologically preservable endocarps" ("Incluso si la selección produce cambios direccionales en la morfología del fruto, estos cambios pueden no ser detectables en el registro arqueológico. Este es a menudo el caso de las drupas, donde la selección está dirigida hacia cambios en la calidad de la porción carnosa del fruto. Por ejemplo, los olivos domesticados (Olea europaea L.) y los dátiles (Phoenix dactylifera L.) tienen una porción carnosa más desarrollada y de mejor calidad que sus contrapartes silvestres, pero esto no se refleja en los endocarpos arqueológicamente preservables) (Zohary & Hopf, 1994)" (Lepofsky et al., 1998: 1002). En el caso de C. maxima subsp. maxima también existen órganos o tejidos de escasa o nula conservación (i.e. hojas y mesocarpo), en tanto otros se recuperan con frecuencia en los sitios arqueológicos (i.e. semillas, fragmentos de pericarpio y pedúnculos). Por ende, nos interesa explorar la posibilidad de que los rasgos de estas últimas estructuras sean susceptibles de reflejar la selección humana sobre poblaciones de la mencionada especie.
Antecedentes de estudios arqueobotánicos del género Cucurbita
El genero Cucurbita se caracteriza por ser morfológicamente muy variable, un rasgo que incluso se cree compartieron gran parte de las especies primeramente domesticadas por el hombre, ya que ofrecían una mayor variabilidad desde la cual seleccionar caracteres deseables. Esto se constituyó en un obstáculo difícil de sortear por quienes emprendieron el estudio de este género, incluso Tomas Whitaker, experto reconocido mundialmente en el estudio de cucurbitáceas -tanto en sus manifestaciones actuales como arqueológicas-declaró que existe una enorme variabilidad en la morfología de semillas dentro de una única especie y es por ello que no son del todo confiables las claves de identificación que se basan sólo en los caracteres de las semillas (Whitaker, 1980).
La identificación arqueológica de las formas de relación entre sociedades humanas pretéritas y poblaciones de Cucurbita sp. se ha realizado por medio del análisis de caracteres métricos en restos de frutos y semillas (Decker & Wilson, 1986; Newsom et. al., 1993; Smith, 2000, 2006). La mayoría de los estudios se ha centrado en estas últimas ya que se consideran las menos afectadas por factores ambientales en relación al fruto y otros órganos vegetativos (Decker & Wilson, 1986) y además porque son abundantes en el registro arqueológico (Cutler & Whitaker, 1961). Estos motivos nos llevaron a seleccionar a las semillas para efectuar el presente estudio.
Para poder contar con parámetros de referencia que permitan evaluar los caracteres presentes en los restos arqueobotánicos, se han analizado hasta el presente los rasgos exhibidos por poblaciones silvestres de Cucurbita sp. Cowan & Smith (1993) elaboraron su colección de referencia con quince plantas de una población de morfotipo silvestre de Cucurbita pepo L., tomando de cada planta un fruto y de cada fruto veinte semillas. De cada semilla midieron el largo y el ancho observando que ambas medidas poseían un coeficiente de variación (CV) bajo (con valores de 5,4 y 6,6 respectivamente), atribuyendo dichos resultados al carácter silvestre de la población estudiada (Cowan & Smith, 1993). Para estos y otros autores (Gremillion, 1993, Newsom et al., 1993) los valores de CV bajos indican homogeneidad morfológica, existiendo uno o pocos morfotipos en las poblaciones silvestres. Estos valores aumentarán con los procesos de domesticación por la presencia de más de un tipo de fruto y/o los efectos iniciales de la hibridación, cultivo y cría selectiva: "... the coefficients of variation may be used indirectly to verify the advent of gardening or the intensification of gardening practices and perhaps human-directed selection for fruit characters..." ("...el coeficiente de variación puede ser usado [en arqueología] indirectamente para verificar el advenimiento de la horticultura o la intensificación de prácticas hortícolas y quizá la selección humana dirigida de caracteres del fruto...") (Newsom 1993: 87). Cowan & Smith (1993) verifican que el CV aumenta cuando se consideran diversos cultivares actuales de morfología muy diferente, desprendiéndose de ello que un aumento en el CV o un CV alto en conjuntos arqueológicos puede indicar la presencia de variedades con frutos de morfología muy diferente. La persistencia de una alta variabilidad a lo largo de una secuencia arqueológica señala además que ciertas prácticas tales como el aislamiento reproductivo y la cría selectiva estaban probablemente limitadas en frecuencia o grado (Gremillion, 1993).
Por lo tanto, la etapa inicial de domesticación estaría caracterizada por una tendencia a la disminución en el CV respecto de las poblaciones silvestres, si éste permanece alto en la primera etapa puede ser a causa de la persistencia de formas con caracteres silvestres o malezoides (Gremillion, 1993), las cuales irán disminuyendo en frecuencia si asumimos una selección dirigida de un conjunto reducido de caracteres. La segunda etapa se caracterizará por un aumento en el CV debido a la generación de diversos cultivares dentro de una única especie domesticada. Es esta segunda etapa la que analizaremos en este trabajo teniendo en cuenta que para diferenciar cultivares en conjuntos de semillas arqueológicas se han empleado medidas lineales tales como largo, ancho y alto de las mismas (Whitaker, 1983).
Por último, mencionaremos que en las poblaciones biológicas silvestres la forma es menos variable y está menos sujeta a factores ambientales que el tamaño, como la primera se representa a través de índices y el segundo a través de valores absolutos, los índices serían más adecuados para identificar distintos taxones en ejemplares arqueo-botánicos (Mangafa & Kotsakis, 1996). La forma es un rasgo genético más conservador que el tamaño en las semillas (Kay et al., 1980), por lo tanto es aún más aconsejable el empleo de índices para identificar taxones a partir de las mismas.
Tomando en consideración lo dicho hasta aquí, podemos ver que los estudios generales referidos a morfología de órganos vegetales y otros realizados exclusivamente sobre semillas de Cucurbita sp. han demostrado que el empleo de valores métricos lineales, el CV de dichos valores y las relaciones entre ellos como representativas de la forma, pueden ser indicadores adecuados para identificar cultivares en los restos arqueobotánicos.
METODOLOGÍA DEL TRABAJO DE CAMPO ETNOBOTÁNICO Y ORGANIZACIÓN DE LOS DATOS
El estudio etnobotánico se llevó a cabo en dos comunidades: Santa Victoria Oeste (SVO) ubicada en el centro de la provincia de Salta, en su extremo norte y El Shincal (ESH) en el valle de Hualfin, Catamarca (Fig. 1). Estas comunidades fueron seleccionadas por estar emplazadas en dos ambientes fitogeográficos distintos: la primera en la provincia de las Yungas, distrito de los Bosques Montanos (Hurrell, 1989; Hilgert, 2007a) y la segunda en la del Monte (Capparelli, 2007).
Fig. 1. Localidades en donde se efectuaron las investigaciones etnobotánicas.
La localidad de Santa Victoria Oeste, constituye la cabecera del departamento homónimo y se ubica a 2560 m s. m. en la quebrada del río que lleva también su mismo nombre dentro de la Alta Cuenca del Río Bermejo (Hurrell, 1989; Hilgert, 2007a). El acceso a esta comunidad se realiza por la ruta nacional Nº 5 que parte de La Quiaca (Jujuy), pasa por la localidad de Yavi, cruza la Sierra de Santa Victoria por el Abra de Lizoite a 4500 m s. m. y en territorio salteño recorre las quebradas de los Ríos Lizoite y La Huerta hasta llegar a SVO. Tras su trabajo en la segunda mitad de la década de 1980, Hurrell (1989) destaca el aislamiento de esta comunidad, siendo la mencionada ruta una vía para el intercambio comercial, principalmente de productos alimenticios. A raíz de esto, en SVO se concentran todas las mercancías provenientes de La Quiaca (LQ) para luego distribuirse a las pequeñas comunidades de la zona. Esto mismo fue constatado en nuestro trabajo de campo observándose que periódicamente ciertos pobladores de SVO concurren al mercado municipal de La Quiaca para aprovisionarse de diversas mercancías que ofrecen luego a la venta en su comunidad de origen. Es por ello que nuestro trabajo etnobotánico incluyó también el relevamiento y la colecta de ejemplares de C. maxima subsp. maxima vendidos en el mercado municipal de La Quiaca.
Los pobladores de SVO ubican sus viviendas en los fondos de valles o laderas bajas, poseyendo en la cima de los cerros una vivienda menor o puesto para el cultivo y cría de ganado al cual concurren algunos miembros de la familia periódicamente para el mantenimiento de la producción. El cultivo se realiza en terrazas fluviales aprovechando la capa de suelo negro de unos 40 cm de espesor (Zardini & Pochettino, 1983). Los estudios etnobotánicos llevados a cabo en la década de 1980 evidencian que, si bien los pobladores locales siembran una gran diversidad de plantas nativas americanas, incluso en pequeñas parcelas, desde 1983 existe un aumento considerable en la introducción de cultivos foráneos (principalmente hortalizas) que desplazan a los autóctonos (Zardini & Pochettino, 1983; Hurrell 1989). Esto también fue constatado por nosotros al relevar la composición de los huertos locales de SVO donde el número de especies nativas americanas fue siempre menor al de las originarias del Viejo Mundo.
Estos huertos y las parcelas de cultivos ubicadas en los puestos son para autoconsumo y trueque local, siendo la principal fuente de semillas o propágulos vegetativos la propia producción. Estas mismas características de las áreas agrícolas se ha visto en diversas comunidades de la Alta Cuenca del Río Bermejo a través de estudios etnobotánicos recientes (Hilgert & Gil, 2005; Hilgert, 2007a-b).
La comunidad de El Shincal se ubica a siete kilómetros del poblado de Londres de Quimivil en el Departamento de Belén. La misma está compuesta por familias cuya economía es principalmente de base agrícola-ganadera, poseyendo también empleos estatales (Capparelli, 2007). La mayor parte de las familias poseen puestos en los cerros que rodean a El Shincal en donde crían animales y cultivan parcelas. Dentro de las actividades agrícolas debemos distinguir el cultivo orientado para el autoconsumo que se realiza en huertas o rastrojos, de aquel orientado a la venta en el mercado local, practicado en fincas de mayor extensión. En el caso de ESH, no existen comercios que expendan verduras en el ámbito de la comunidad, sí los hay en Londres, pero los habitantes que cultivan sus propios vegetales en ESH para autoconsumo no los adquieren comercialmente. Las semillas para sembrar en los huertos o rastrojos se guardan de un año para otro o bien son intercambiadas dentro de la comunidad.
En ambas comunidades se realizaron entrevistas abiertas y semiestructuradas (Granai, 1962; Martin, 1995; Cotton, 1998) a individuos adultos de ambos sexos (19 individuos en ESH y 10 en SVO) pertenecientes cada uno a una unidad doméstica distinta. El muestreo representó entre un 50 y un 75% del total de unidades domésticas de cada una de las comunidades. Las entrevistas y la observación directa libre y metódica (Granai, 1962) se llevaron a cabo en las unidades domésticas y en las áreas de cultivo, fundamentalmente en las orientadas al autoconsumo. Las entrevistas semiestructuradas estuvieron orientadas al relevamiento de prácticas agrícolas, criterios de selección de especies y variedades a sembrar, caracteres deseados y alentados en las distintas especies sembradas y formas de cultivo, manipulación y procesamiento de las mismas. En el caso del mercado municipal de La Quiaca se efectuaron entrevistas semiestructuradas con las personas que atendían los puestos de venta y observación metódica en los mismos. Ambas técnicas estuvieron dirigidas exclusivamente al relevamiento de información sobre la venta de distintos tipos de zapallos: nombre asignado, procedencia y compradores más frecuentes.
Las entrevistas fueron registradas en libreta y soporte digital de audio y las observaciones fueron complementadas con registro gráfico y fotográfico de las áreas de cultivo (huertas, fincas, etc.) y de los artefactos vinculados con la obtención y el procesamiento de recursos vegetales (morteros, harneros, palas, etc). Se colectó material etnobotánico con el previo consentimiento de los pobladores locales (tanto plantas enteras como órganos útiles: frutos y semillas) a fin de llevar a cabo identificaciones taxonómicas y análisis morfo-anatómicos.
Las muestras coleccionadas fueron depositadas en el Herbario de Plantas Útiles del Laboratorio de Etnobotánica y Botánica Aplicada (Facultad de Ciencias Naturales y Museo, U.N.L.P.) asignándoles sigla y número de registro personal. Las entrevistas fueron desgravadas y la información procesada en una base de datos considerando: área de estudio, fecha en que se realizo el trabajo de campo, características de los informantes (nombre, sexo y edad), especies mencionadas, nombres vulgares asignados a las mismas, sigla y número correspondiente de las muestras coleccionadas con el que se ingresaron al mencionado herbario, usos reconocidos localmente, modos de empleo y prácticas de manipulación (recolección, protección, cultivo, entre otras posibilidades).
El reconocimiento taxonómico de los ejemplares del género Cucurbita coleccionados se realizó empleando tanto caracteres del fruto y pedúnculo -cuando fue posible tener acceso a los mismos- así como también por medio de características macroscópicas y microscópicas de las semillas (Cutler & Whitaker, 1961; Lira Saade, 1995; Teppner, 2004; Lema et al., 2008).
Selección de la muestra
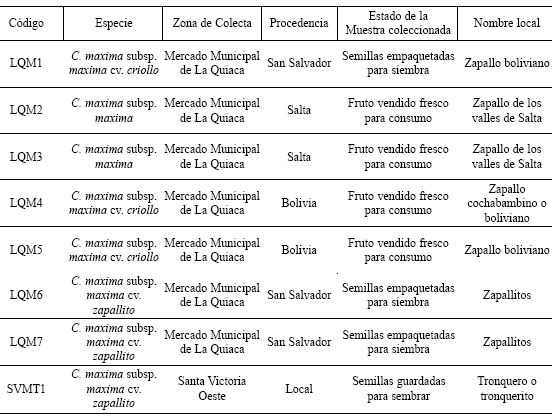
De la totalidad de especies registradas se seleccionaron aquellas mencionadas por los entrevistados como Zapallos. De los siete cultivares mencionados en ESH, tres fueron taxonómicamente identificados como C. pepo y cuatro como C. maxima subsp. maxima. Del análisis de las entrevistas referidas a esta última especie se comprobó que sólo un individuo (D.S.R., masc., 77 años) menciona modos distintos de manipulación de poblaciones de la misma. El entrevistado señaló que cultiva Zapallo criollo (muestra de semillas aportada por el entrevistado registradas bajo el código DSR1) y guarda la semilla de otro Zapallo que también consume, "el cual crece sólo con la lluvia" sin ningún cuidado de su parte, en un campo abandonado ubicado frente a su vivienda (muestra DSR2). Señaló también que el segundo no es tan rico como el criollo, pero su beneficio es que no necesita riego extra para crecer. A pesar de que estas prácticas fueron registradas en un único caso del total analizado, el mismo es significativo ya que en el marco del trabajo etnobotánico que se viene realizando en El Shincal y en otras comunidades del Noroeste argentino como SVO desde hace cuatro años, nunca se mencionaron cuidados diferenciales para los tipos de Zapallos reconocidos localmente. Consideramos por lo tanto que el caso de este poblador de ESH es de suma importancia ya que puede ser representativo de formas de manipulación que se han ido perdiendo con el paso del tiempo.
En el caso de LQ todas las muestras fueron identificadas como C. maxima subsp. maxima (LQM1 a LQM7). Sólo dos muestras de zapallos fueron coleccionadas en SVO, una (localmente denominada Guineo) fue identificada como C. pepo y otra (Tronquero o Tronquerito) como C. maxima subsp. maxima cv. zapallito (SVMT1). Estas semillas fueron aportadas por una informante (D.L.M., fem., 56 años) y corresponden a los zapallos que cultiva en su huerto. Según nos informó, las semillas son guardadas tras la cosecha y son plantadas cada año.
Finalmente, para el análisis efectuado en este trabajo se seleccionaron, en primer lugar, muestras que representaran modos distintos de relación hombre- planta en una comunidad (DSR1 y DSR2) y en segundo lugar muestras que representaran cultivares que son reconocidos y circulan en una misma comunidad, distinguiéndose entre los que son manipulados localmente en todo su ciclo productivo (SVMT1) de los que ingresan a la comunidad para ser consumidos, pero que no son producidos localmente y por lo tanto no reflejan criterios locales de manejo (LQM1 a LQM7) (Tabla 1)
Tabla 1. Datos correspondientes a las muestras obtenidas en el trabajo de campo etnobotánico en la localidad de Santa Victoria Oeste (Salta, Argentina).
METODOLOGÍA DE TRABAJO EN GABINETE Y SELECCIÓN DE LOS DESCRIPTORES MORFOLÓGICOS
Como descriptores morfológicos macroscópicos se seleccionaron el largo, ancho y alto de las semillas. Se midieron la totalidad de las semillas obtenidas en el trabajo de campo por medio del empleo de calibre digital. De cada uno de estos descriptores se calculó la media, el desvío estándar, el coeficiente de variación (CV) y los valores máximos y mínimos. A su vez, el CV fue usado específicamente como descriptor en sí mismo. Este descriptor es especialmente valioso ya que permite la comparación de muestras de distinto tamaño (N). Agrupamos los valores de CV en alto, medio y bajo siguiendo los criterios de Newsom (1993): los valores de entre 1 y 6 se consideran bajos, entre 6 y 10 medios y mayores a 10 altos.
En segunda instancia se calcularon tres índices (largo/ancho, largo/alto y ancho/alto) empleados también como descriptores morfológicos.
Para el análisis de caracteres microscópicos se siguió la metodología propuesta por Lema et al. (2008) para el reconocimiento de especies sudamericanas de Cucurbita sp. a partir de rasgos cualitativos y cuantitativos de la cubierta seminal. Se seleccionó un subconjunto de muestras (LQM3, LQM5, SVMT1, DSR1 y DSR2) y se realizaron cortes delgados de las cubiertas seminales de una semilla por muestra, los cuales fueron observados por transparencia bajo microscopio óptico. Entre los rasgos microscópicos propuestos por Lema et al. (2008) para identificar especies de Cucurbita sp. se seleccionaron cinco como descriptores morfológicos: largo y ancho de las células epidérmicas, alto y ancho de las esclereidas y alto de la capa hipodérmica.
Se obtuvieron veinte medidas por descriptor mediante el empleo de una cámara digital unida al microscopio junto al "software" acompañante (Motic Image Plus 2.0).
Los caracteres cuantitativos fueron organizados en una base de datos con el fin de poder realizar un análisis jerárquico de conglomerados con empleo de representación gráfica de diagrama de árbol o dendograma. Se utilizó el método de Ward´s y la distancia euclidiana como medida de similitud. El análisis se realizó empleando el "Software Statistica" 6.0. Se seleccionó este tipo de análisis ya que se reconoce su utilidad para evaluar grados de similitud/diferencia en caracteres de semillas de diferentes plantas alimenticias que poseen un valor diferencial asignado por los pobladores entrevistados (Höft et al., 1999).
RESULTADOS DEL ANÁLISIS DE LOS DESCRIPTORES MORFOLÓGICOS: LA BÚSQUEDA DE MORFOTIPOS
En el caso de las semillas, el carácter macroscópico cualitativo más destacado es el color de las mismas, distinguiéndose aquellas de superficie opaca y color castaño (DSR1, LQM1, LQM4 y LQM5, correspondientes al cultivar criollo) de las que poseen superficie brillante y color blanco (DSR2, SVMT1, LQM2, LQM3, LQM6 y LQM7). La utilidad de este carácter para distinguir cultivares disminuye al trabajar con restos arqueobotánicos, ya que los mismos se encuentran por lo general desecados y descoloridos o bien carbonizados. Sin embargo, estos colores de la superficie se correlacionan con tipos de células epidérmicas (Teppner, 2004; Lema et al., 2008) tal como veremos al analizar caracteres cuantitativos microscópicos.
Santa Victoria Oeste. Descriptores en morfología macroscópica
Largo, ancho y alto de las semillas (Tabla 2, Figs. 2-4). Para el caso de SVO, los tres descriptores indican la presencia de dos morfotipos. El morfotipo 1 está compuesto por semillas de zapallos llamados Bolivianos o Cochabambinos, vendidos en el mercado municipal de La Quiaca (LQM1, LQM4 y LQM5). A simple vista se aprecia que estos zapallos de gran tamaño poseen semillas también de grandes dimensiones. Esta diferencia queda plasmada en un morfotipo caracterizado por un largo de 25 - 30 mm, un ancho de 13 - 15 mm. (pudiendo llegar hasta 17 mm) y un alto de 5 - 7, 5 mm.
Tabla 2. Valores de los descriptores en morfología macroscópica cuantitativa correspondientes a las muestras obtenidas en la comunidad de Santa Victoria Oeste (Salta) y El Shincal (Catamarca). Abreviaturas: N, cantidad de semillas analizadas; DS, desvío estándar; CV, coeficiente de variación; Min, valor mínimo registrado; Max, valor máximo registrado.

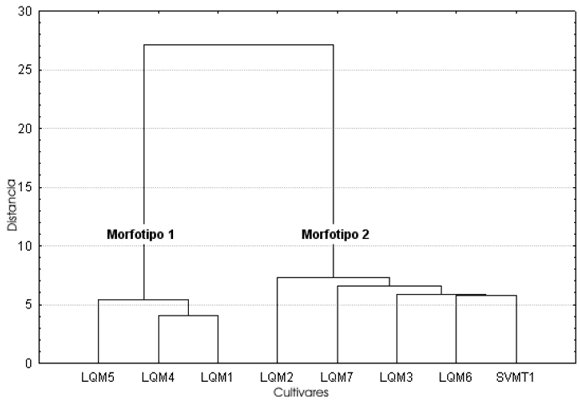
Fig. 2. Análisis de conglomerados del largo de las semillas correspondientes a las muestras de cultivos locales procedentes de la comunidad de SVO (Salta).
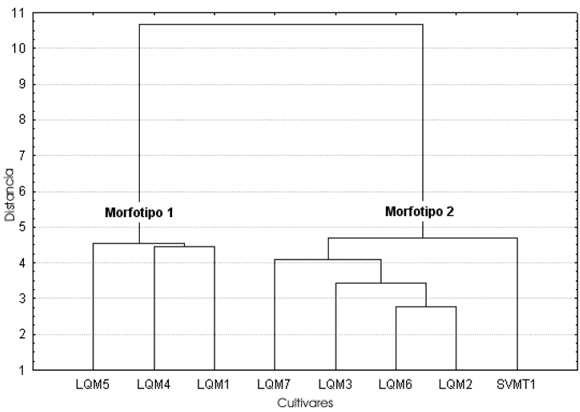
Fig. 3. Análisis de conglomerados del ancho de las semillas correspondientes a las muestras de cultivos locales procedentes de la comunidad de SVO (Salta).
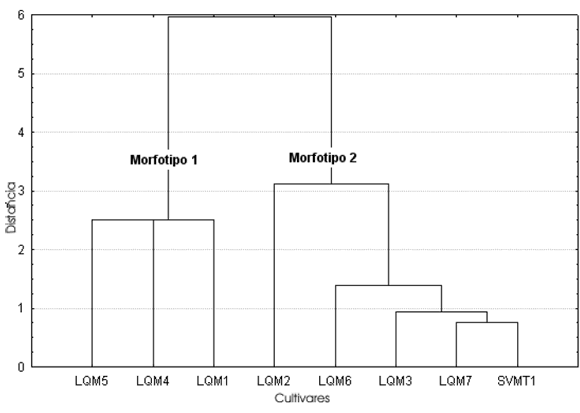
Fig. 4. Análisis de conglomerados del alto de las semillas correspondientes a las muestras de cultivos locales procedentes de la comunidad de SVO (Salta)
El morfotipo 2 es mucho más variable que el anterior, agrupando ejemplares de C. maxima subsp. maxima y de C. maxima subsp. maxima cv. zapallito (LQM2, LQM3, LQM6, LQM7 y SVMT1). En ninguno de los tres descriptores los conglomerados menores dentro del morfotipo 2 lograron reflejar las clasificaciones locales: los valores obtenidos para el largo y ancho distinguen aquellos ejemplares que los habitantes de la localidad agrupan bajo un mismo nombre -Zapallos de los valles de Salta- y otros que se clasifican localmente por separado y poseen trayectorias distintas. Para este último caso, pueden citares los Zapallitos que proceden de San Salvador y se venden en el mercado de La Quiaca junto con Zapallos que responden a un cuidado y cultivo local llamados Tronquero o Tronquerito, o bien con zapallos procedentes de Salta.
La distancia de ligamiento entre las muestras del morfotipo 1 es menor en el alto, siendo las del largo y ancho similares entre sí. La distancia de amalgamiento entre ambos morfotipos es menor también en el caso del alto, siguiéndoles en orden creciente el largo y el ancho. Coeficiente de variación (Tablas 2-3, Fig. 5). En general el CV no se comporta de la misma manera en cada muestra para los tres descriptores considerados (largo, ancho y alto). Si bien en las muestras LQM3, LQM4 y LQM6 los valores de CV se comportan de igual manera como bajos, medios o altos para las tres medidas, debemos considerar que cada una de estas muestras representa una parte de una categoría mayor reconocida por los pobladores locales. Por ejemplo LQM3 corresponde a Zapallos de los valles de Salta. No obstante, dentro de esta categoría está también la muestra LQM2 cuyos valores de CV son totalmente distintos, lo mismo ocurre con LQM4 que forma parte del grupo de Zapallos bolivianos o cochabambinos y LQM6, que junto con LQM7, proveen las semillas de Zapallitos que se venden en el mercado de La Quiaca.
Tabla 3. Agrupamiento de los valores de coeficiente de variación (CV) para las tres medidas lineales de las muestras obtenidas en la comunidad de Santa Victoria Oeste (Salta) siguiendo a Newsom (1993). Abreviaturas: B, valores bajos; M, valores medios; A, valores altos.
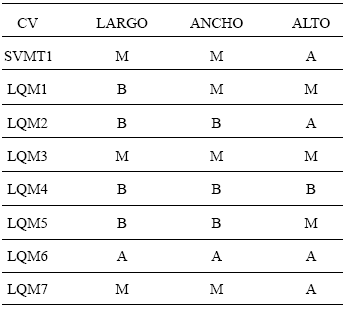
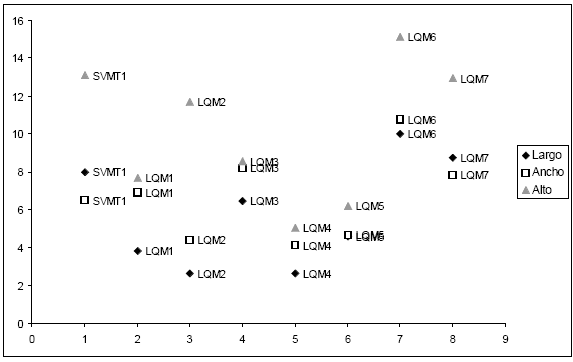
Fig. 5. Gráfico que representa la distribución de los valores de CV para las tres medidas lineales de las muestras obtenidas en la comunidad de Santa Victoria Oeste (Salta).
Si quitamos el alto de la Tabla 3 veremos que en todos los casos los valores de CV de largo y ancho coinciden en su carácter de bajo, medio o alto, salvo en LQM1. Teniendo en cuenta este hecho tendríamos entonces tres conjuntos representados por estos tres rangos de medidas, que están agrupando y a la vez separando distintos cultivares locales. El grupo de CV bajo agrupa una muestra de los valles de Salta junto con dos muestras de Zapallos bolivianos; el grupo de CV medio reúne un cultivar local, Zapallos de los valles de Salta y Zapallitos de San Salvador y el CV alto está representado solamente por una muestra de Zapallitos del circuito comercial.
El segundo aspecto de interés es el análisis de la variabilidad presente en un cultivar. Si éste se encuentra representado por semillas procedentes de diversos frutos, puede ser que el CV aumente, ya que las mismas pueden ser el producto de distintas condiciones a las que estuvieron sometidas las plantas y que a la vez pudieron causar cambios morfológicos. Pero si consideramos la variabilidad representada por semillas de un mismo fruto, tendremos una visión más ajustada de cuán variable es el cultivar. Cada una de las muestras de los Zapallos bolivianos y las de los valles de Salta proceden de frutos distintos, sin embargo los CV de ambos cultivares son muy discrepantes. Las muestras LQM4 y LQM5 presentan valores bajos de CV, por lo que este sería un cultivar bastante homogéneo. Los Zapallos de Salta, en cambio, presentan valores muy dispares entre uno y otro fruto (LQM2 y LQM3), motivo por el cual se trataría de un cultivar heterogéneo. Por lo tanto, el mantenimiento de CV altos puede estar presente en un cultivar.
Índices (Tabla 4, Figs. 6-8). El análisis de conglomerados de los tres índices indica que, si bien ninguno refleja la diversidad localmente reconocida, el índice largo/ancho (Fig. 6) correctamente agrupa a los zapallitos comerciales (LQM6 y LQM7) y separa al único cultivar no comercial (SVMT1) del resto de los ejemplares que se comercializan en La Quiaca. En los casos restantes volvemos a encontrar agrupamientos de cultivares diferenciados a nivel local y separación de muestras clasificadas como un único cultivar.
Tabla 4. Índices correspondientes a los descriptores en morfología macroscópica cuantitativa de las muestras obtenidas en la comunidad de Santa Victoria Oeste (Salta) y El Shincal (Catamarca)
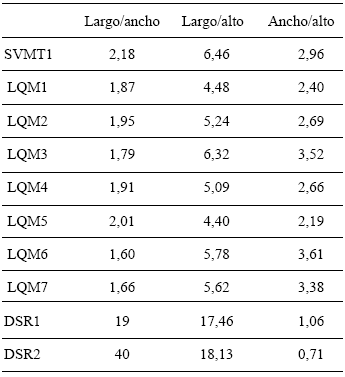
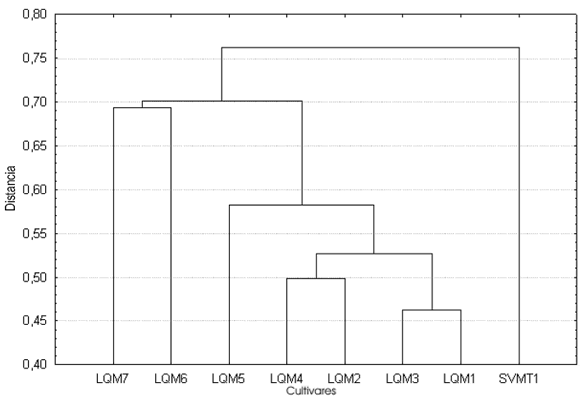
Fig. 6. Análisis de conglomerados del índice largo/ancho de las semillas correspondientes a las muestras de cultivos locales procedentes de la comunidad de SVO (Salta).
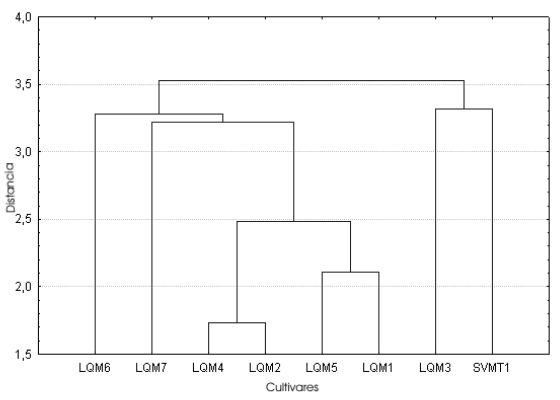
Fig. 7. Análisis de conglomerados del índice largo/alto de las semillas correspondientes a las muestras de cultivos locales procedentes de la comunidad de SVO (Salta).
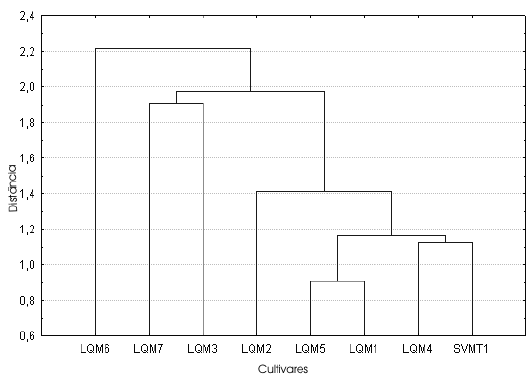
Fig. 8. Análisis de conglomerados del índice ancho/alto de las semillas correspondientes a las muestras de cultivos locales procedentes de la comunidad de SVO (Salta)
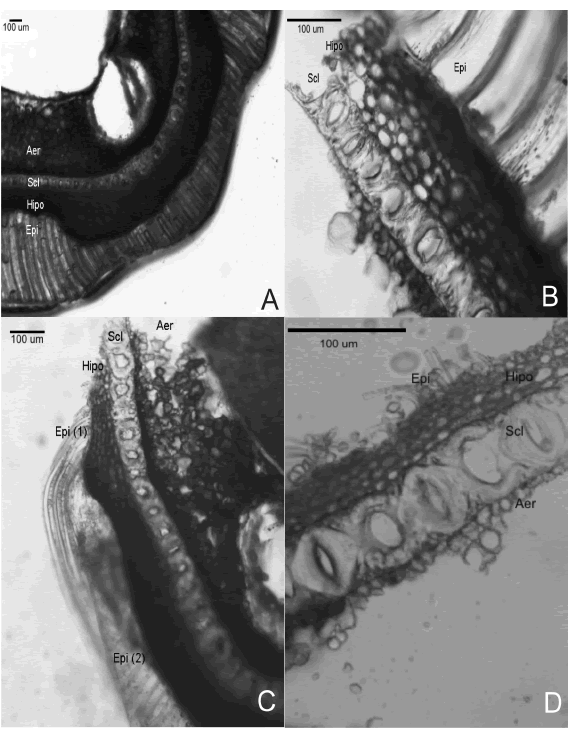
Fig. 9. Cortes delgados de cubiertas seminales de ejemplares coleccionados en el trabajo de campo etnobotánico. A-B. Muestra LQM5: A, margen; B, cara mayor. C-D. Muestra DSR1: C, margen; D, cara mayor. Abreviaturas: Epi, epidermis (1, células largas; 2, células cortas); Hipo, hipodermis; Scl, esclereidas; Aer, aerénquima
Descriptores en morfología microscópica Entre los caracteres cualitativos, la presencia de células epidérmicas largas de paredes engrosadas en las caras mayores de la semilla que no llegan a rodear a las células epidérmicas cortas en el margen de la misma, distinguen a LQM5 como C. maxima subsp. maxima cv. criollo (Lema et al., 2008) (Fig. 9A-B). Por el contrario, la presencia de células epidérmicas largas de paredes delgadas en las caras mayores que rodean a las células cortas de los márgenes es compartido por LQM3 y SVMT1.
Tras el análisis de los datos cuantitativos (Tabla 5) puede verse que sólo el alto de la capa hipodérmica en caras mayores (Fig. 9B) permite diferenciar a los tres cultivares como tres morfotipos distintos dado que las medidas no se solapan entre sí.
Tabla 5. Resultados de los descriptores en morfología cuantitativa microscópica para las muestras de Santa Victoria Oeste (Salta) y El Shincal (Catamarca). Abreviaturas: N, cantidad de semillas analizadas; DS, desvío estándar; CV, coeficiente de variación; Min, valor mínimo registrado; Max, valor máximo registrado.
Los valores de CV para los distintos descriptores son muy discrepantes al interior de cada muestra, incluso al considerar dos caracteres de una misma célula (Tabla 5), por lo cual no se considera al CV como un descriptor válido por sí mismo para caracterizar a un cultivar.
El Shincal. Descriptores en morfología macroscópica
Largo, ancho y alto de las semillas (Tabla 2). Los resultados muestran que ambas muestras poseen valores que se solapan entre sí, por lo cual no se pueden distinguir morfotipos diferentes. Coeficiente de variación (Tabla 2). Los valores de CV de la muestra DSR2 son más bajos en los tres casos.
Índices (Tabla 4). Los valores muy similares en los índices (con alto grado de solapamiento) impiden distinguir a estas dos muestras como unidades distintas.
Descriptores en morfología microscópica
Mediante el empleo de caracteres cualitativos se constató que, al igual que en el caso de las muestras de Santa Victoria Oeste, hay dos cultivares: la muestra DSR2 corresponde a C. maxima subsp. maxima cv. criollo y la muestra DSR1 a un cultivar de epidermis de paredes delgadas (Fig. 9 C-D).
Al igual que en el caso de SVO el CV no resultó útil para distinguir cultivares (Tabla 5).
En este caso el largo de las células epidérmicas fue el parámetro que mejor diferenció a ambos grupos al no haber solapamiento de medidas.
DISCUSIÓN
En el caso de los cultivares el empleo de descriptores macroscópicos puede tanto reducir como aumentar artificialmente la variabilidad reconocida por una comunidad al agrupar cultivares que los entrevistados consideran distintos y a la vez separar los que son tomados como uno , tal es el caso de SVO. Asimismo, los valores de CV obtenidos para este último conjunto de cultivares difiere en gran medida de acuerdo al descriptor elegido, por lo tanto no es indistinto tomar el CV del largo, ancho o alto de una muestra de semillas. En los estudios paleoetnobotánicos y arqueobotánicos, por lo general, se toma como indicador de cambio el CV de una medida lineal, frecuentemente del largo (véase Newsom, 1993) sin justificar esta elección. En el caso del alto -a pesar de que en los análisis de conglomerados evidenciaron las menores distancias de ligamiento y de amalgamiento-recomendamos que no sea considerado en estudios de restos arqueobotánicos ya que el mismo se ve muy afectado por procesos de desecación, degradación y carbonización. El ancho, en cambio, puede ser empleado sin inconvenientes, incluso es mencionado por Cowan & Smith (1993) como un carácter más conservador en las semillas del género Cucurbita. Pero incluso al no considerar los CV del alto en los cultivares de SVO, los CV de largo y ancho desagrupan muestras de cultivares que los pobladores locales reúnen. Asimismo, la consideración de que sólo valores diferentes de CV obtenidos en una muestra de una localidad o sitio arqueológico indicarán la presencia de distintos cultivares debe también reconsiderarse, ya que los resultados aquí obtenidos señalan que un mismo rango de valores de CV puede estar agrupando cultivares distintos.
En el caso de grupos de plantas sujetas a distintos tipos de manipulación (DSR1 y DSR2), los descriptores macroscópicos tampoco resultaron de utilidad para diferenciar dichos tipos.
Finalmente, los resultados obtenidos tanto para las muestras de SVO como para las de ESH indican que el uso exclusivo de valores bajos de CV como indicadores de cultivos en muestras arqueobotánicas puede llegar a subestimar la presencia de cultivares en donde el mantenimiento de una alta diversidad es un rasgo seleccionado por los cultivadores. En este sentido es contundente el hecho de que muestras procedentes de poblaciones con escaso cuidado humano (DSR2) obtengan CV más bajos que las que sí son cuidadas y regadas (DSR1). Esto estaría en contraposición con la idea de que los cuidados humanos tienden a reducir la variabilidad presente en las poblaciones naturales, generando poblaciones homogéneas de acuerdo con los caracteres seleccionados. Este es un punto importante ya que de acuerdo con Cowan & Smith (1993) las medidas actuales de semillas del género Cucurbita muy variables no son útiles para ser empleadas en la identificación de material arqueológico con escasas semillas.
El empleo de índices para evaluar si la forma refleja mejor las categorías locales que el tamaño no arrojó resultados favorables para el caso de ESH y sólo resultó útil el índice largo/ancho para dos cultivares de SVO.
En cambio, los descriptores empleados en morfología cuantitativa microscópica generaron morfotipos que reflejan de manera más ajustada los cultivares localmente reconocidos. Sin embargo, no todos fueron igualmente útiles; por lo tanto, es indispensable realizar el análisis de un número elevado de descriptores micro morfológicos con el fin de hallar diferencias significativas.
Siguiendo a Gándara (2006) podemos decir que hemos generado a partir de ciertos rasgos cuantitativos macroscópicos y microscópicos de semillas de C. maxima subsp. maxima una base inductiva cuyos elementos deben ir en aumento a fin de elaborar nuevas inferencias analógicas. Por lo tanto, si bien descartamos el empleo de ciertos indicadores, hemos corroborado la utilidad de otros. Esta corroboración irá tomando mayor fuerza al contrastarse con nuevos conjuntos de referencia. En este trabajo sólo hemos dado un primer paso en la elaboración de un argumento de relevancia, tal como lo entiende Gándara (2006), en el sentido de que la composición y frecuencia del conjunto base sean fundamento suficiente para la inferencia.
CONCLUSIONES
El desarrollo de estudios etnobotánicos como herramienta etnoarqueológica puede ser de gran ayuda para evaluar la posibilidad de identificar comportamientos sociales pasados a través del estudio de restos arqueobotánicos. El traspaso de lo comportamental a lo material es siempre dificultoso y requiere una constante revisión de los medios que empleamos para hacer esta transformación, puesto que con frecuencia los indicadores que estamos seleccionando para hacerlo no son los más adecuados. En este trabajo, vimos que ciertos atributos empleados en paleoetnobotánica y arqueobotánica para hablar de comportamientos pasados (vg. caracteres macroscópicos de las semillas) pueden ser engañosos y llevarnos a conclusiones erradas. El estudio de nuevos atributos tales como caracteres microscópicos de la cubierta seminal resultó ser un camino promisorio a explorar arqueológicamente y a seguir contrastando con más estudios etnobotánicos de carácter o aplicación etnoarqueológica. Si bien en este trabajo hemos indagado la utilidad de ciertos indicadores propios de los restos macrobotánicos, somos conscientes de que la reconstrucción de comportamientos pasados requiere el análisis de la totalidad del registro arqueológico en el cual se encuentran dicha clase de restos. Por lo tanto, creemos que el estudio etnobotánico de situaciones de manipulación de comunidades vegetales y su correlato material puede ser una vía fructífera de información a ser aplicada en análisis contextuales en arqueología, la cual no ha sido aún explorada.
Para finalizar, consideramos que las vías de análisis planteadas en este trabajo son válidas para investigar arqueológicamente el cultivo en tanto comportamiento humano en el pasado. Intentamos a la vez desarrollar nuevas alternativas de interpretación que nos ayuden a imponer cada vez menos nuestra propia visión del mundo a las comunidades que nos antecedieron.
AGRADECIMIENTOS
A los pobladores de El Shincal y Santa Victoria Oeste por cederme su tiempo, sus plantas y su amabilidad. A Tany Pochettino y Aylen Capparelli por la posibilidad que me dieron de realizar ambos estudios de campo y por la lectura crítica de las distintas versiones manuscritas de este trabajo. A los evaluadores anónimos y a los editores de la revista Darwiniana por sus correcciones, comentarios y apoyo para la realización de este trabajo. A Diego Gobbo por su asistencia en el procesamiento de imágenes. Todo lo expresado en esta publicación es responsabilidad exclusiva de la autora.
BIBLIOGRAFÍA
1. Archila S.; M. Giovannetti M & V. Lema V. (eds.). 2008. Arqueobotánica y teoría arqueológica. Discusiones desde Suramérica. Bogotá: UNIANDES-CESO [ Links ]
2. Arellano E. & A. Casas. 2003. Morphological variation and domestication of Escontria chiotilla (Cactaceae) under silvicultural management in the Tehuacán Valley, Central Mexico. Genet. Resources Crop Evol. 50: 439-453 [ Links ]
3. Capparelli, A. 2007. Los productos alimenticios derivados de Prosopis chilensis (Mol.) Stuntz y P. flexuosa DC., Fabaceae, en la vida cotidiana de los habitantes del NOA y su paralelismo con el algarrobo europeo. Kurtziana 33(1) 103-120 [ Links ]
4. Casas, A. 2001 Silvicultura y domesticación de plantas en Mesoamérica, en B. Aguilar, S. Domínguez, J. Caballero Nieto & M. Martínez Alfaro (eds.), Plantas, cultura y sociedad. Estudio sobre la relación entre seres humanos y plantas en los albores del siglo XXI, pp. 123-158. México: UAM-S.M.A.R.N.y P. [ Links ]
5. Casas, A; A. Otero-Arnaiz, E. Pérez-Negrón & A. Valiente Banuet. 1991-2003. Manejo y domesticación de cactáceas en Mesoamérica. Zonas Áridas 7: 75-104 [ Links ]
6. Casas, A; B. Pickersgill, J. Caballero & A. Valiente Banuet. 1997. Ethnobotany and domestication in Xoconochtli, Stenocereus stellatus (Cactaceae), in the Tehuacán Valley and La Mixteca Baja, México. Econ. Bot. 51:279-292 [ Links ]
7. Cotton, C. M. 1998. Ethnobotany. Principles and applications. England: Wiley & Sons. [ Links ]
8. Cowan, C.W. & B. D. Smith. 1993. New perspectives on a wild gourd in eastern North America. J. Ethnobiol. 13(1): 17-54. [ Links ]
9. Cutler, H. & T. Whitaker. 1961 History and distribution of the cultivated cucurbits in the Americas. Amer. Antiquity 26(4): 469-485. [ Links ]
10. Decker, D. & H. Wilson. 1986. Numerical analysis of seed morphology in Cucurbita pepo. Syst. Bot. 11(4): 595-607 [ Links ]
11. Farrington, I. & J. Urry. 1985. Food and the early history of cultivation. J. Ethnobiol. 5(2): 143-157. [ Links ]
12. Ford, R. 1985. The processes of plant food production in prehistoric north America, en R. Ford (ed.), Anthropological papers 75, Vol: Prehistoric food production in North America, pp: 1-19. Michigan: University of Michigan Press. [ Links ]
13. Frére, M.; M. González, A. Guráieb & S. Muñoz. 2004. Etnoarqueología, arqueología experimental y tafonomía, en A. Aguerre & J. Lanata (eds.), Explorando algunos temas en arqueología, pp. 97-120. Buenos Aires: Ed. Gedisa. [ Links ]
14. Fournier, P. & A. Freeman. 1991. El razonamieno analógico en etnoarqueología, el caso de la tradición alfarera de Mata Ortiz, Chihuahua, México. Boletín de Antropología Americana 23:109-114 [ Links ]
15. Gándara, M. 2006. La inferencia por analogía: más allá de la analogía etnográfica. Treballs D´Etnoarqueología 6: 13-23. [ Links ]
16. Granai, G. 1962. Técnicas de investigación sociológica, en G. Gurvitch (ed.), Tratado de sociología, pp. 153-171. Buenos Aires: Editorial Kapeluz. [ Links ]
17. Gremillion, K. 1993. The evolution of seed morphology in domesticated Chenopodium: an archaeological case study. J. Ethnobiol. 13(21): 149-169. [ Links ]
18. Harlan, J. R. 1992. Crops and man. Wisconsin: American Society of Agronomy. [ Links ]
19. Hildebrand, E. 2003. Motives and opportunities for domestication: an ethnoarchaeological study in southwest Ethiopia. Journal of anthropological archaeology 22: 358-375. [ Links ]
20. Hilgert, N. I. 2007a. Plantas silvestres, ámbito doméstico y subsistencia, en A.D. Brown, M. García Moritán, B. N. Ventura, N. I. Hilgert, L. R. Malizia (eds.), Finca San Andrés. Un espacio de cambios ambientales y sociales en el Alto Bermejo, pp.187-228. Tucumán: Ediciones del Subtrópico. [ Links ]
21. Hilgert, N. I. 2007b. La vinculación del hombre actual con los recursos naturales y el uso de la tierra, en A. D. Brown, M. García Moritán, B. N. Ventura, N. I. Hilgert, L. R. Malizia (eds.), Finca San Andrés. Un espacio de cambios ambientales y sociales en el Alto Bermejo, pp. 159-186. Tucumán: Ediciones del Subtrópico. [ Links ]
22. Hilgert, N. I. & G. E. Gil. 2005. Traditional Andean agriculture and changing processes in the Zenta River Basin, Salta, Northwestern Argentina. Darwiniana 43(1-4): 30-43 [ Links ]
23. Hillman, G. 1984. Interpretation of archaeological plant remains: the application of ethnographic models from Turkey, en W. Van Zeist & W. Casparie (eds.), Plant and ancient man. Studies in palaeoethnobotany, pp. 1-41. Rotterdam-Boston: A. Balkema. [ Links ]
24. Hillman, G. & M. S. Davies. 1990. Measured domestication rates in wild wheats and barley under primitive cultivation, and their archaeological implications. Journal of World Prehistory 4(2): 157-222. [ Links ]
25. Höft, M.; S. Barik & A. Lykke. 1999. Quantitative ethnobotany. Applications of multivariate and statistical analyses in ethnobotany. People and plants, working paper 6. Paris: UNESCO [ Links ]
26. Hurrell, J. A. 1989. Interpretación de relaciones en ecología a partir de la noción de sistema. Para el referente empírico Santa Victoria e Iruya (Salta, Argentina). Tesis Doctoral. Universidad Nacional de La Plata, Facultad de Ciencias Naturales y Museo. [ Links ]
27. Kay, S.; A. King & M. Robinson. 1980. Study of archaeobotanical seeds. Amer. Antiquity 35: 26-39. [ Links ]
28. Lema, V. 2008. ¿De qué hablamos cuando hablamos de domesticación vegetal en el NOA? Revisión de antiguas propuestas bajo un nuevo abordaje, en S. Archila, M. Giovannetti & V. Lema (eds.)), Arqueobotánica y teoría arqueológica. Discusiones desde Sudamérica, pp. 97-126. Bogotá: UNIANDES. [ Links ]
29. Lema V.; A. Capparelli & M. L. Pochettino. 2008 Taxonomic identification of dry and carbonized archaeobotanical remains of Cucurbita species through seed coat micromorphology. Veg. Hist. & Archaeobot. 17 (Suppl. 1): 277-286. [ Links ]
30. Lepofsky, D.; P. Kirch & K. Lertzman.1998. Metric analysis of prehistoric morphological change in cultivated fruits and nuts: an example from Island Melanesia. J.Archaeol. Sci. 25: 1001-1014. [ Links ]
31. Lira Saade, R. 1995. Estudios taxonómicos y ecogeográficos de las Cucurbitaceas latinoamericanas de importancia económica. México: Instituto de Biología. U.N.A.M. [ Links ]
32. Lobot, V; E. Johnston, Q. E. Zheng, D. McKern & D. Mc Kenna. 1999. Morphological, phytochemical and genetic variation in Hawaiian Cultivars of ´Awa (Kava, Piper methysticum, Piperaceae). Econ. Bot. 53(4): 407-418. [ Links ]
33. Mangafa, M & K. Kotsakis. 1996. A new method for the identification of wild and cultivated charred grape seeds. J. Archaeol. Sci. 23: 409-418. [ Links ]
34. Marconetto, M. B. 2008. Linnaeus en el Ambato. El uso de la clasificación taxonómica en arqueobotánica, en S. Archila, M. Giovannetti & V. Lema (eds.), Arqueobotánica y teoría arqueológica. Discusiones desde Sudamérica, pp. 143-166. Bogotá: UNIANDES. [ Links ]
35. Martin, G. 1995. Ethnobotany. A methods manual. London: Chapmann & Hall, World Wide Fund for Nature. [ Links ]
36. Nazarea, V. D. 1998. Cultural memory and biodiversity. Tucson: The University of Arizona Press. [ Links ]
37. Newsom, L. A.; S. D. Webb & J. S. Dunbar.1993. History and geographic distribution of Cucurbita pepo gourds in Florida. J. Ethnobiol. 13(1): 75-98. [ Links ]
38. Politis, G. 2004. Tendencias de la etnoarqueología en América Latina, en G. Politis & D. Peretti (eds.), Teoría arqueológica en América del Sur, Serie Teórica Nº 3, pp. 85-118. Buenos Aires: INCUAPA- UNICEN. [ Links ]
39. Smith, B. 2000. Guilá Naquitz Revisited: Agricultural origins in Oaxaca, Mexico, en G. Feinman & L. Manzanilla (eds.), Cultural evolution, contemporary viewpoint, pp. 15-60. New York: Kluwer. [ Links ]
40. Smith, B. 2006. Seed size increase as a marker of plant domestication, en M. Zeder, E. Emshwiller, D. Bradley & B. Smith (eds.), Documenting domestication: new genetic and archaeological paradigms, pp.25-31. Berkeley: University of California Press. [ Links ]
41. Teppner, H. 2004 Notes on Lagenaria and Cucurbita (Cucurbitaceae). Review and new contributions. Phyt. 44 (2): 245-308. [ Links ]
42. Toledo, V.M. 2002. Ethnoecology. A conceptual framework for the study of indigenous knowledge of nature, en J. R. Stepp, F. S. Wyndham & R. K. Zarger (eds.), Ethnobiology and biocultural diversity, pp. 3-10. Athens (Georgia, USA): University of Georgia Press. [ Links ]
43. Whitaker, T. 1983 Cucurbits in Andean prehistory. Amer. Antiquity 48(3): 576-585. [ Links ]
44. Whitaker, T. 1980. Cucurbitáceas americanas útiles al hombre. Conferencia dictada en la ciudad de La Plata en octubre de 1980. Publicaciones de la Comisión de Investigaciones Científicas de la provincia de Buenos Aires (CIC). [ Links ]
45. Zardini, E. & M. L. Pochettino. 1983 Resultados de un viaje etnobotánico al norte de Salta (Argentina). IDIA: 108-121. [ Links ]


{kind=link}